Embed Size (px)
Citation preview

Netv Phytol. (1995), 131, 461-469
Mycorrhizal fungus propagules in thejarrah forest
II. Spatial variability in inoculum levels
BY M A R K C. B R U N D R E T T * AND L. K.ABBOTT
Soil Science and Plant Nutrition, The University of Western Australia, Nedlands,W.A. 6009, Australia
{Received 12 May 1994; accepted 22 December 1994)
SirMMARY
Spatial variations in the capacity of propagules of arbuscular mycorrhizal (AM) and ectomycorrhizal (ECM) fungito form associations in their natural habitats were investigated using hioassays with bait plants grown in intactcores of forest soil. These cores were collected from a sclerophyllous forest community dominated by Eucalyptusmarginata Donn ex Smith (jarrah) and E. calophylla Lindley (marri) trees with a diverse shrub understorey in themediterranean (winter rainfall) climatic zone of Western Australia. Small-scale (adjacent core) variations in thecapacity of AM fungi to form associations were found to be as substantial as differences between locations 15 mapart. Comparisons of AM fungus colonization patterns within the roots of seedlings growing in the same coreindicated that there was considerable spatial heterogeneity in the inoculum potential of ' individual' fungi withinthese 1 1 volumes of soil. A second experiment included bait plants to measure ECM formation as well as AMformation and also considered the impact of soil disturbance. The disruption of hyphal networks reducedmycorrhiza] formation somewhat, but it still remained highly variable. Some of this spatial heterogeneity couldbe attributed to differences in the organic matter content, length of fungal hyphae, or length of old mycorrhizalroots, measured withm soil cores. In jarrah forest soil, mycelial systems of AM and ECM fungi apparently werelocalized in separate domains, and there were also zones where non-mycorrhizal roots (mostly cluster rootsproduced by members of the Proteaceae) predominated. More research is required to determine the size ofdomains of m\corrhizal mycelial systems in soils, how these spatial patterns change with time, and if they areassociated with zones of resource utilization by different 'functional groups" of roots.
Key words: Mycorrhizas, eucalyptus forest soil, inoculum potential, soil spatial ^•ariability, bioassays.
networks are important drought-resistant propagulesINTRODV CTiON of AM fungi in many south-western Australian soils,
Plants with mycorrhizal associations are predomi- but are sensitive to soil disturbance (Jasper, Abbottnant in most natural ecosystems, but little is known & Robson, 1989(7, b. 1991).about the life-histories of mycorrhizal fungi in plant Inoculum potential is defined as the energy forcommunities. In particular, the capacity of propa- growth of an organism at the surface of its host, andgules of mycorrhizal fungi to persist in soil without is a consequence of the number of active propagulesroots, tolerate disturbance, resist predation by other of that organism and their nutritional status (Garrett,soil organisms and disperse to new locations are not 1956). For mycorrhizal fungi, inoculum potentialwell understood (see Brundrett, 1991). Propagules of can be defined as the capacity of their propagules toarbuscular mycorrhizal (AM) fungi are thought to form associations in a particular soil and can beinclude spores, dead root fragments and other colon- measured as the rate of colonization of host roots,ized organic material, as well as networks of hyphae lndirectestimationsof inoculum potentials of mycor-in soil. Propagules of ectomycorrhizal (ECM) fungi rhizal fungi have been obtained by using serialinclude networks ofmycehal strands, old mycorrhizal dilutions of soil or by counting propagules such asroots, sclerotia and basidiospores (Skinner & Bowen, spores, but results of these procedures do not always1974; Ba, Garbaye & Dexheimer, 1991). Hyphal correspond with mycorrhiza formation in soil
(Abbott & Robson, 1991; Brundrett, 1991). Short-• To whom correspondence- should be addressed at: CSIRO ^^^^ bioassavs that measure mycorrhiza formation
Centrt- for Mediterranean Agricultural Research, Division of , , . , ' . ., ' , u uForestry-, Private Bag PO,Wembiey, W.A. 6014, Australia. by bait plants grown m SOll samples WOuld be

462 M. C. Brundrett and L. K. Abbott
expected to provide a better estimate of inoculumpotential. The use of intact soil cores allows thecontribution of disturbance sensitive propagules -such as mycelial networks, as W"ell as more robustpropagules - such as spores or root fragments, tobe measured.
Aggregated (non-random) patterns of distributionare often observed when spores of AM fungi aresampled (Walker, Mize & McNab, 1982; Andersonet al., 1983; Sylvia, 1986; Tews & Koske, 1986;Friese & Koske, 1991), when hyphal growth in soil isobserved (St John, Coleman & Reid, 1983), or whenmycorrhizai colonization of root systems is examined(St John & Hunt, 1983; Fitter & Merryweather,1992). Ectomycorrhizal fungi often occupy patchesor 'mats' in forest soils resulting from the localizedspread of mycelial systems (Ogawa, 1985; Griffiths& Caldwell, 1992). This results in the aggregateddistribution of sporocarps (Ford, Mason & Pelham.1980; Gregory, 1982; Taylor, 1992), that might beassociated with separate clones of these fungi(Dahlberg & Stenlid, 1990). The inoculum of soil-borne plant pathogens typically also has an aggre-gated distribution pattern (Campbell & Noe, 1985).
Jarrah forests are sclerophyllous plant communi-ties dominated by Eucalyptus marginata (jarrah) andE. calophylla (marri) trees with a diverse understoreycomprised mostly of shrubs (Bell & Heddle, 1989).This habitat is located in the mediterranean climaticzone of Western Australia, where cool, wet wintersare followed by 4—7 months of summer drought(Gentiili, 1989). Jarrah forest plants include manyspecies with mycorrhizal associations, but non-mycorrhizal plants are also common (Brundrett &Abbott, 1991).
The first part of this study demonstrated thatthere were no large seasonal fluctuations in numbersof mycorrhizal fungus propagules in jarrah forestsoils, but that there were consistent variations inthese numbers along a transect within a site(Brundrett & Abbott, 1994). Spatial patterns inmycorrhizal inoculum potential could not be ex-plained by variations in the cover of host and non-host plants at transect locations. Substantial varia-bility in AM fungus inoculum potential over a scaleof several metres has also been observed in a re-vegetated jarrah forest site 4 y after disturbance bymining (Jasper, Abbott & Robson, 1992). Theexperiments presented here represent furtherattempts to resolve spatial patterns in the inoculumpotential of AM and FCM fungi in jarrah forest soilusing bioassays with bait plants grown in intact soilcores.
MATERIALS AND METHODS
Relatively undisturbed, intact 1 I cores of lateriticgravel soil were collected from the jarrah forest sitedescribed in the first part of this study (Brundrett &
Abbott, 1994). Soil cores were collected fromimmediately adjacent to each other (core edges werec. 1 cm apart) and from locations 15 m apart alongtransects for two separate experiments (Fig. 1).These cores were placed in plastic-lined 1 1 buckets,which were kept in a constant-temperature watertank (20 °C) and maintained at field capacity duringbioassay experiments. The procedures used for soilcore bioassays, to clear, stain and assess root samples,and to quantify spores are described in detailelsewhere (Brundrett, Melville & Peterson, 1994).
Experiment 1
The objective of this experiment was to compare theAM fungus inoculum potential in immediatelyadjacent soil cores and in cores from other locationsalong a transect. Four adjacent intact soil cores werecollected in a 2 x 2 grid from four locations separatedby 15 m as show-n in Figure 1 a. These locations wereat the 15, 30, 45 and 60 m positions along the sametransect sampled during the first part of this study(Brundrett & Abbott, 1994). Soil cores were collectedat the end of the dry season (May 1990) and fromwithin 1 m of the same locations during the wintergrowmg season (July 1990). Estimates of the in-oculum potential of AM fungi within cores weremeasured by bioassays using clover {Trifoliumsubterraneum L. cv. Seaton Park) bait plants grownin each core for 2 wk. Ten clover seeds, which weregerminated in aerated water for 1 d, were sown (withRhizobium leguminosarum bv. trifolii [TAl] inocu-lum) into each pot and later thinned to four plantsper pot.
After the 2 wk period found optimal for AMbioassays in which the rate of mycorrhizal formationis used to estimate inoculum potential (Brundrett &Abbott, 1994), bait plant roots were washed fromsoil cores, cleared in lO" ;, KOH and stained inchlorazol black E. Mycorrhizal and non-mvcorrhizal
(a)
Transectlocation
Transectlocation 15 m 15 m
Figure 1. Sampling patterns for soil cores from the Jarrahforest site for expts 1 [a) and 2 {b). For expt 1, four cloverplants (X) were grown in each core. Experiment 2treatments: AM = 2 wk clover bioassay, ECM = 4 wkEucalyptus calophylla and Mirbelia dilatata bioassay.Hatched cores were disturbed while others remainedintact.

Mycorrhizal propagules in jarrah forest, II 463
roots were then measured by the gridline intersectionmethod (Giovannetti & Mosse, 1980). All themycorrhiza] roots from each clover plant wereseparately mounted on slides to allow total mycor-rhizal colonization within cores to be measured.Slides of roots from the first harvest were examinedwith a compound microscope and colonization bydifferent AM fungi quantified by estimating thelength (in mm) of colonies produced within roots.This method of quantification produced results thatwere in close agreement with those obtained with thegridline intersection method (i?" = 0'817), but wasmore accurate when colonization levels were low.Colonization by different AM fungi was quantifiedseparately after each had been characterized usinghyphal diameters and morphology patterns withinroots (Abbott, 1982).
Experiment 2
The objective of this experiment was to comparedifferences in the inoculum potential of both ECMand AM fungi in adjacent soil cores and in coresfrom other locations along a transect and to measurepotential propagules within these cores. Sixteenadjacent soil cores were collected from a 4 x 4 gridfrom each of three locations near the transect usedfor the previous experiment in the jarrah forest siteas shown in Figure 16. Half of the soil cores weredisturbed b\' lifting plastic bags from pots andmanually crushing any large soil aggregates (<5 tnm), before re-packing soil in pots. Alternatingcores were left undisturbed (Fig. \b). This 'mild'disturbance was expected to disrupt pre-existingnetworks of hyphae without much mixing of soillayers.
Mycorrhizal formation by clover bait plants wasused to estimate AM inoculum potential after 2 wkin half the cores as described above. Germinatedseeds of Eucalyptus calophylla Lindley and trans-planted seedlings of Mirbelia dilatata R. Br. wereplaced in the remaining pots and allowed to grow for4 wk to estimate ECM inoculum potential (see Fig.\b). The bait plants and bioassay procedures usedwere optimized in the earlier study (Brundrett &Abbott, 1994). Mycorrhizal formation of bait plantswas assessed using the percentage of root length withAM colonies and the number of root tips with ECMassociations relative to total root length after clearingand staining as described above.
Samples of soil from each core were used toprovide estimates of numbers of potential propa-gules, including the length of fungal hyphae withincores and the amount of mycorrhizal roots andorganic material they contained. Measurements ofsoil hyphae were made using three (14 mm di-ameter X 100 mm long) cores of soil taken from potsI wk before bioassay plants were harvested. These(15-32 g) soil samples were homogenized by blend-
ing them in 520 miof water for 10 s before membranefiltration of a 0-5 ml aliquot, trypan blue staining,and gridline intersect counting (Abbot, Robson &De Boer, 1984). Spores of AM fungi were quantifiedusing a 50-100 g subsample of soil from cores, whichwas collected when roots were extracted. Sporeswere separated by wet-sieving, followed by centH-fugation in water and then in S0% sucrose, beforefiltration. All the pre-existing roots within soil coreswere collected as the roots of bait plants wereharvested. These roots from cores were cleared andstained and the length of AM, ECM and non-mycorrhizal roots was measured as described above.Light organic material was separated from heaviersoil materials by flotation in water. This organicmaterial was collected on sieves, whereas heaviermaterial (mostly lateritic gravel) was discarded. Theorganic material was dried at 60 °C and weighed.
Correlation, multiple regression and analysis ofvariance tests were used to compare bioassay resultsbetween undisturbed and disturbed soil cores and toexamine their relationships with soil constituentsthat tnay function as propagules.
RESULTS
Experiment 1
When clover bait plants grown in adjacent soil coreswere compared, differences in mycorrhiza] coloni-zation by AM fungi were as substantial as thosebetween locations (Fig. 2). In this experiment,statistically significant differences in mycorrhiza]formation occurred within and between transectlocations but not between sampling times (Table 1).Note that the highest and lowest va]ues of mycor-rhiza] co]onization shown in Figure 2a occurredin adjacent cores.
A detailed examination of the patterns of mycor-rhiza] colonization within the roots of individualclover bait plants demonstrated substantial dif-ferences in mycorrhizai population structure be-tween cores within sites as we]] as between sites (Fig.3). For examp]e, there was more co]onization by aGlomus sp. (or spp.) with coarse hyphae in core 2than in other cores from site D, Acaulospora andScutellospora (young colonies of these fungi werehard to distinguish) were most common in core 2from site B, Glomus sp. (medium endophyte) oc-curred sporadically, while colonization by a fineendophyte was greatest in site D, core 4. There werealso differences in colonization by specific AM fungibetween the clover plants growing within each core(Fig. 3).
Experiment 2
This experiment aimed to measure the extent ofmycorrhiza] formation and simultaneously quantifypotentia] mycorrhiza] fungus propagu]es in the same
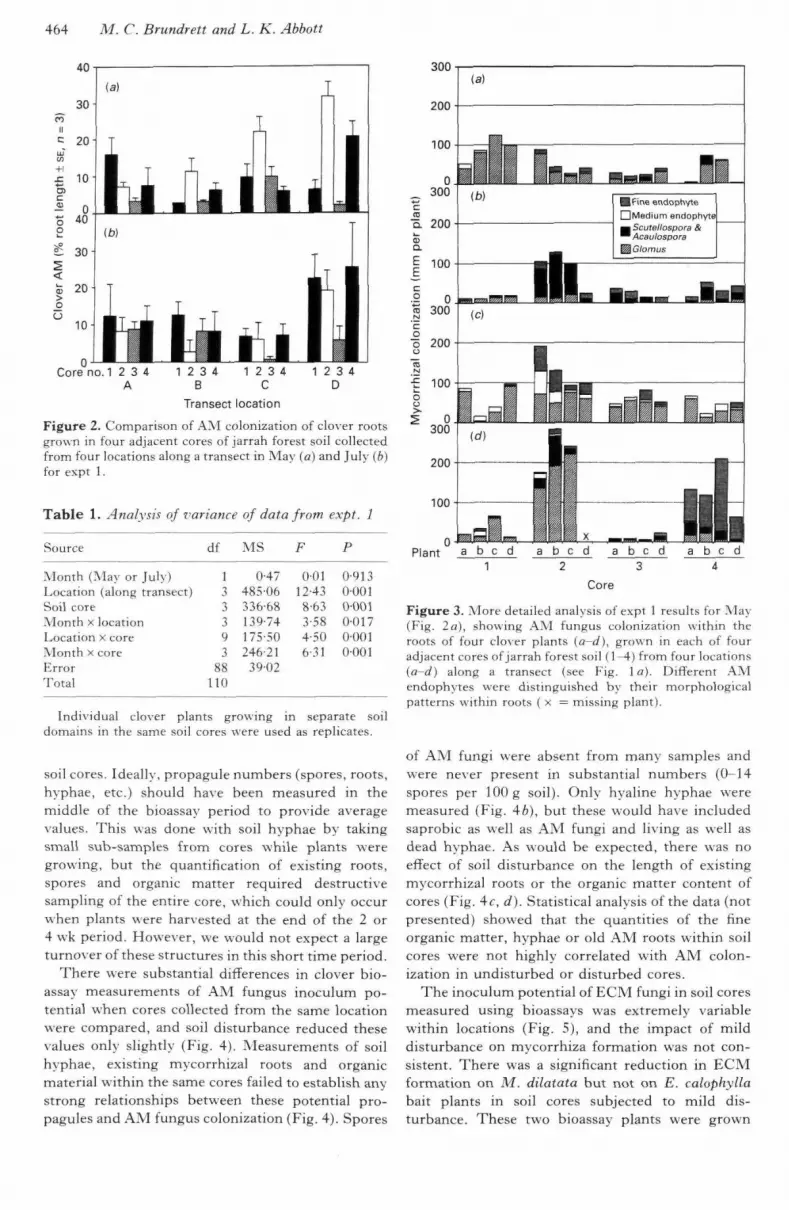
464 M. C. Brundrett and L. K. Abbott
Core no. 1 2 3 4A
1 2 3 4 1 2 3 4C D
Transect location
Figure 2. Comparison of AM colonizarion of clover rootsgrown in four adjacent cores of jarrah forest soil collectedfrom four locations along a transect in May {a) and July (b)for
Table 1. Analysis of variance of data from expt. 1
Source df MS F
Month (May or July)Location (along transect)Soil coreMonth X locationLocation X coreMonth X coreErrorTotal
0'47485-06336-68I39'74i75'5O246-21
0-0112-438'633-584-506-.31
0-9130-0010-0010'017O'OOl0-001
88 39-02UO
Individual clover plants growing in separate soildomains in the same soil cores were used as replicates.
soil cores. Ideally, propagule numbers (spores, roots,hyphae, etc.) should have been measured in themiddle of the bioassay period to provide averagevalues. This was done with soil hyphae by takingsmall sub-samples from cores while plants weregrov\ing, but the quantification of existing roots,spores and organic matter required destructivesampling of the entire core, which could only occurwhen plants were harvested at the end of the 2 or4 w-k period. However, w e would not expect a largeturnover of these structures in this short time period.
There were substantial differences in clover bio-assay measurements of AM fungus inoculum po-tential w-hen cores collected from the same locationwere compared, and soil disturbance reduced thesevalues only slightly (Fig. 4). Measurements of soilhyphae, existing mycorrhizal roots and organicmaterial within the same cores failed to establish anystrong relationships between these potential pro-pagules and AM fungus colonization (Fig. 4). Spores
300
200
edium andophyteScutellospora &AcaulosporaGlomus
P l a n t a b e d a b e d a b e d a b e d
Core
Figure 3. More detailed analysis of expt 1 results for May(Fig. 2a), showing AM fungus colonization within theroots of four clover plants {a-d), grown in each of fouradjacent cores of jarrah forest soil (1-4) from four locations(a-d) along a transect (see Fig. 1 a). Difierent AMendophytes were distinguished by their morphologicalpatterns within roots ( x = missing plant).
of AM fungi were absent from many sample.s andwere never present in substantial numbers (0-14spores per lOOg soil). Only hyaline hyphae weremeasured (Fig. 46), but these would ha\e includedsaprobic as well as AM fungi and living as well asdead hyphae. As would be expected, there was noeffect of soil disturbance on the length of existingmycorrhizal roots or the organic matter content ofcores (Fig. 4c, d). Statistical analysis of the data (notpresented) showed that the quantities of the fineorganic matter, hyphae or old AM roots within soilcores were not highly correlated with AM colon-ization in undisturbed or disturbed cores.
The inoculum potential of FCM fungi in soil coresmeasured using bioassays was extremely variablewithin locations (Fig. 5), and the impact of milddisturbance on mycorrhiza formation was not con-sistent. There was a significant reduction in ECMfornaation on M. dilatata but not on E. calophyllabait plants in soil cores subjected to mild dis-turbance. These two bioassay plants were grown

Mycorrhizal propagules in jarrah forest, II 465
> 2 mm
2-0-75 mm
0 75-0 25 mm
Core 1 2 3 4 t
Undisturbed Disturbed •? Undisturbed
? 3
Undisturbed Disturbed
A B C
Location
Figure 4. Experiment 2 results showing spatial variability in AM colonization of clover bait plants {a) and theproportion of potential propagules measured in the same soil cores {b~d). Potential propagules are hyaline soilhyphae {b), pre-existing roots i;ontaining AM fungi (c) and organic matter fragments which may have containedfungal structijre.s [d). Results are shown for intact and disturbed cores.
together in soil cores and produced similar results insoil from two locations, but not for the third locationwhere E. calophylla remained non-mycorrhizal,whereas M. dilatata was colotiized in three out offour cores (Fig. 5 a, b). Inconsistencies in ECMformation by these two hosts were also observed inthe first part of this study. Both E. calophylla and M.
dilatata also formed AM associations to a limitedextent (data not presented).
Measurements of existing mycorrhizai roots, soilhyphae (of all types) or organic matter were takenfrom the same cores where bioassay plants weregrow'n (Fig. 5). Correlation analysis demonstratedthat ECM formation by E. calophylla was related to

466 M. C. Brundrett and L, K. Abbott
20
5O CO
Is
Core0
1IiJL ^ JLi Ji ' ^M^fei ' tsxy wXi' ( W IS^Ti-s^^t>'T»»JRa»' \my BSW MW E^'T'S
1
1 2 3 4
Undisturbed1 2 3 4
Disturbed1 2 3 4Undisturbed
1 2 3 4Disturbed
B
1 2 3 4 1 2 3 4Undisturbed Disturbed
Location A
Figure 5. Experiment 2 results showing spatial variability in ECM colonization of Eucalyptus calophylla (a) andMirhelia diiata {b) bait plant roots and the proportion of potential propagules measured in the same cores ofintact or disturbed soil (r-e). Potential propagules are pre-existing roots with ECM associations (c), total soilhyphae {d), and fine organic matter fragments (e).
the amounts of soil hyphae, fine organic matter insoil cores and M. dilatata ECM formation in theundisturbed cores (Pearson coefficients were 0764,0-819, 0-616 respectively), but not in the disturbedcores (Pearson coefficients = 0'168, -0-073, 0465).Multiple regression using data from the undisturbedcores resulted in the equation E. calophylla ECM= —5-27-1-1-63 hyphae-I-1-63 fine organic matter (r^- 7 5 7 « o ) .
There was a tendency for existing roots to bepredominantly either AM or ECM in difTerent soilcores, but there were also many cores where non-mycorrhizal roots were most abundant (Fig. 6).Most of these non-mycorrhizal roots were cluster
roots produced by members of the Proteaceae, whichwere common in the site.
DISCUSSION
Bioassays using intact soil cores demonstrated small-scale spatial heterogeneity in AM and ECM funguspropaguies in jarrah forest soil (between adjacent100 mm wide soil cores) to be as great as larger scalespatial variability (between sample sites 15m apart)observed in an earlier study (Brundrett & Abbott,1994). Differences in mycorrhiza formation byindividual bait plants within soil cores providedevidence of spatial variability in the inoculum

Mycorrhizal propagules in jarrah forest, II 467
250-
I E •S Q-100-
am ^ 50H
20 40 60 80AM colonization (% root length)
100
Figure 6. The relationship between AM and ECMcolonization of existing roots within 1 1 intact cores of soilused for bioassays in expt 2.
potential of particular fungi at an even smaller scale(less than 50 mm). Substantial differences in rootcolonization by different fungi in adjacent soil cores,would make it impossible to take uniform samples ofthis soil to use as replicates in an experiment.However, the use of mixed soil m experiments wouldalso be misleading because of the loss of activehyphae and changes in the dispersion of aggregatedpropagules. The aggregated distribution pattern ofpropagules of soil fungi must be considered duringfield sampling, experimental design and data analy-sis, as is discussed by St John & Hunt (1983),Campbell & Noe (1985), Tews & Koske (1986) andDutilleul (1993).
The slight overall reduction in mycorrhiza form-ation owing to disturbance of intact soil cores, andthe results of other bioassay experiments with jarrahforest soils (Jasper et al., 1991; Brundrett & Abbott,1994) suggest that hyphal networks were a source ofmycorrhiza] inoculum m this soil, even after longdry periods. Old mycorrhizal roots, and fine organicmaterial from these soils can also function as propa-gules and have been used to start pot cultures of AMfungi (data not presented). However, correlationsbetween these potential propaguies m soil corescould not explain all the variability in AM or ECMformation by bioassay plants. This might haveresulted because the importance of different propa-gules varied between cores. It is also likely thatthe small subsamples of soil that were used tomeasure hyphae or spores were not representativeof the entire core. Spores of AM fungi did not occurin sufficient numbers in jarrah forest soil toresult in substantial amounts of colonization, asis often the case in other natural ecosystems(Brundrett, 1991). Root colonization of bait plantswas primarily by Glomus species with coarse andmedium hyphae and by a fine endophyte. However,there was also sporadic colonization by Acaulosporaand Scutellospora in bait plant roots. The spores thatwere encountered belonged to these genera, and mayhave been important propagules for them.
Relatively large (1 I) intact soil cores were taken tominimize disturbance impacts during sampling and
the impact of severing mycorrhizal hyphae fromfood sources. The impact of fragmenting mycelialsystems would depend on their size. Spatial varia-tions in mycorrhizal formation within individualcores of jarrah forest soil provided evidence thatthese contained a number of separate hypha! net-works belonging to different fungi within each of the1 1 volume of soil samples used in the present study.The capacity of propagules in soils to form mycor-rhizal associations perhaps could also be measuredby examining mycorrhizal formation in situ in fieldsoils. However, it would be difficult to compareresults of different in situ experiments, because of theimpact of large fiuctuations in soil temperature andmoisture conditions on root growth and mycorrhizaformation. Thus it is best to remove cores of soilfrom the field and incubate them in a controlledenvironment to obtain reproducible measurementsof mycorrhiza] inoculum potential.
The role of spatial heterogeneity in farrah forest soil
Analysis of existing roots in soil cores, which providea historical perspective on mycorrhizal fungusactivity, suggested that there was a tendency forECM, AM and non-mycorrhizal roots to occurwithin separate volumes of jarrah forest soi]. Thissegregation of root activity occurred on a smallerscale than measured variations in the above-grounddistributions of host and non-host plants at the site(Brundrett & Abbott, 1994). It seems likely thatroots of plants had a tendency to avoid patches of soilwhere roots w-ith another type of mycorrhizalassociation predominated. It is not known if rootsresponded to specific soil conditions, the activities ofparticular types of fungi, or if there w ere negativeinteractions between the roots themselves.
There are many known cases of alJelopathicinteractions, where chemicals released by plants ortheir litter adversely infiuence the roots of otherplants, and some of these interactions are thought toinvolve mycorrhizal fungi (Rice, 1984; Brundrett,1991). It is suspected that soil microbes often inhibiteach other b\' secreting toxic substances and bycreating zones where required nutrients are depleted(Alien, 1991). More information about these ty^pes ofinteractions will have to be obtained before their rolein Australian ecosystems can be determined.
Ectomycorrhizal roots often preferentially occupyorganic rich layers in forest soils (Har\'ey, Larsen &Jurgensen, 1979; Reddell & Malajczuk, 1984),Hyphae of AM fungi are more often found inmineral soil, but there have also been reports of theirproliferation in localized patches of organic material(St John et al., 1983; Warner, 1984). Cluster rootsare produced by members of the Proteaceae andsome other plants and are thought to be an adaptationto very infertile soils (Lamont, 1993). They oftenproliferate between the mineral soil and litter layer

468 M. C. Brundrett and L. K. Abbott
(Attiwill & Adams, 1993). These micro-habitatpreferences by different 'functional groups' of rootsmight partly explain their spatial heterogeneity injarrah forest soil.
The spatial segregation of different types ofmycorrhizal fungi could also result from differentsubstrate preferences. It has often been stated thathyphae of AM fungi can only utilize the samesources of nutrients that are accessible to roots, butthere is now some doubt about this (Bolan, 1991;Schweigher, 1994). Hyphae of some ECM fungi eanuse simple organic sources of nitrogen and phos-phorus, such as those JtkeJy to be found in Inter(Abuzinadah & Read. 1989; Hausling & Marschner,1989). There is also evidence of enhanced soilmineralization by organic acids within ECM fungusmats (Griffiths & Caldwell, 1992). Cluster roots maypromote nutrient uptake by their large surface areaand the production of leachates which contain organicacids and other substances (Attiwill & Adams, 1993;Lamont, 1993). It is becoming increasingly apparentthat the production of large root surface areas (fine,highly branched roots with long root hairs) alone isnot sufficient to ensure adequate nutrient uptake bynon-mycorrhizal roots, and that these roots are likelyto be more chemically active than those of mycor-rhizal species (Brundrett, 1991).
If there are differences in the forms of nutrientsand the soil micro-habitats that are accessible todifferent types of roots, it might be possible for themsequentially to occupy the same volume of soil andutiliae different resources (types of nutrients). Thiswould help to ensure that no nutrients are lost fromthe system. Efficient nutrient cycling is particularlyimportant in West Australian soils which containsome of the lowest levels of available nutrients in theworld (Hingston, O'Conneil & Grove, 1989;Lamont, 1993). The use of different forms ofnutrients by plants might also be a mechanism toreduce subterranean competition for limiting nu-trient resources within this diverse plant community.Mycorrhizas might also reduce competition betweenco-existing plant species by equalizing differences intheir capacity to obtain nutrients (Brundrett, 1991).Spatial variability in plant communities is now-known to be an important ecological factor whichallows more competing species to co-exist thanwould otherwise be theoretically possible (Tilman,1994). Further considerations of soil spatial varia-bility within ecosystems may help to explain howpopulations of mycorrhizal fungi can exist togetherin soils and how a wide diversity of plants cancompete for limiting resources such as phosphorus.
The large differences in the activity of mycorrhizalfungi detected by bioassays both between and withincores demonstrate spatial heterogeneity consisting ofsoil domains w^here propagules of mycorrhizal fungimay be common or scarce. These spatial patternsprobably change with time, but will be difficult to
study in intact soil systems without the use ofdestructive sampling procedures. There is still muchto learn about spatial and chronological variation inthe activities of soil organisms, the importance ofthis heterogeneity in nutrient cyeling within soils,and its role in maintaining the diversity of mycor-rhizal fungi and plants in ecosystems.
ACKNOWLEDGEMENTS
This research was supported by a postdoctoral researchfellowship to M. Brundrett from the Natural Sfiencfs andEngineering Research Council of Canada, Karen Clarkeand Gracia Murase provided field assistance. We thankMark Adams, Peter Braunberger and Da\id Jasper fortheir comments on the manuscript,
REFERENCES
Abbott LK. 1982. Comparativt- anatomy of vesicuiar-arbuscularmycorrhiza.s formed on subterranean clover. Au.stra!ian jfournalof Botany 30: 485-499.
Abbott LK, Robson AD. 1991. Factors influencing the occurreticeof vesicular arbuscular mycorrhizas. Agriculture, Ecosystemsand Emironn/ent 35 : 121-150.
Abbott LK, Robson AD, De Boer G. 1984. The effect ofphosphorus on the formation of hyphae in soil by the\ esicular-arbuscular mycorrhiziil fungus (Jlomus fasiiculntum.New Phytologist 97: 437—1-46.
Abuzinadab RA, Read DJ. 1989. The role of proteins in thenitrogen nutrition of ectomycorrhizal plants. l\. The utilizationof peptides by birch {Betula pendula L.) infected with differentmycorrhizal fungi. NetL Phytoiogisi 112: 55-6U.
Allen MF. 1991. The ecology of mycorrhisne. Cambridge:Cambridge Uni\'ersit\- Press.
Anderson RC, Liberta AE, Dickman LA, Katz AJ. 1983.Spatial variation in vesicular-arbuscular mycorrhizal sporedensity. Bulletin of the Torrey Botanical Club 110: 511^-525.
Attiwill PM, Adams MA. 1993. Nutrient cycling in foresta. Sen:Phytoiogisi 124: 561-5S2.
Ba MA, Garbaye J, Dexbeimer J. 1991. Influence of fungalpropagules during the early stages of the time sequence ofectomycorrhiza! colonization on Afzelia africana seedlings.Canadian Journal of Botanv 69: 2442-2447,
Bell DT, Heddle EM. 1989. Floristic, morphologic and veg-etational diversity. In: Dell B, Havel JJ, .Maiajczuk X. eds. TheJarrah Forest. Dordrecht: Kluwer Academic Publishers, 53-66.
Bolan NS. 1991. A critical review on ihe role of mycorrhizal fungiin the uptake of phosphorus by plants. Plant and Soil 134:189-207.
Brundrett MC. 1991. Mycorrhizas in natural ecosystems, ln:Macfayden A, Begon M. Fitter AH, eds. Advances itt EcologicalRe.semch. vol. 21. London: Academic Press, 171-313,
Brundrett MC, Abbott LK. 1991. Roots of jarrab forest plants. I.Mycorrhizal associations of shrubs and herbaceous plants.Australian Journal of Botany 39: 445—457.
Brundrett MC, Abbott LK. 1994. Mycorrbizal fungus propa-gules in tbe jarrah forest. 1. Seasonal study of inoculum levels.A'fK Phytologist 127: 5.19 54().
Brundrett M, Melville L, Peterson L. 1994. Practical methodsin mvcorrhizal research. Waterloo: Mycologue PublitationsLtd.
Campbell CL, Noe JP. 198S. Tbe spatial analysis of soilbornepatbogens and root diseases. Annual Review of Phvtopathologv23: 120-14S.
Dahlberg A, Stenlid J. 1990. Population structure and dynamicsin Suillus bovinus as indicated by spatial distribution of fungalclones. New Phytologist 115: 487^93.
Dutilleul P. 1983. .Spatial heterogeneity and the design ofecological field experiments. Ecology 74: 1646-1658.
Fitter AH, Merry weatber JW. 1992. Why are some plants moremycorrbizal than others? An ecological inquiry. In: Read DJ,Lewis DH, Fitter AH & Alexander IJ, eds. Mycorrhizas inEcosystems. Wallingford: C.A.B. International, 26-36.

Mycorrhizal propagules in jarrak forest, II 469
Ford ED, Mason PA, Pelham J. 1980. Spatial patterns ofsporophorc distribution ar<iund a young birch tree in threesuccessive years. Tratisactwns of the British Mvcological Society75: 287 296.
Friese CF, Koske RE. 1991. The spatial dispersion of spores ofvesicular-arbusciihir niyitirrhizal funyi in a sand dune; micro-scale patterns associated with the root architecture of Americanbeachgrass. Myiologiial Rfseiirrli 95; 952-957.
Garrett SD. 1956. Bialdgy of root infecting fungi. Cambridge:Cambridge University Press.
GentilliJ. 1989. Climate of the jiirrah forest. In : Dell B, Havel JJ& Malajczuk N, eds. The Jarrah Forest. Dordrecht: Kluwer.Academic Publishers, 23-40,
Giovannetti M, Mosse B. 1980. An evaluation of techniques formeasuring vesicular-arhuscular infection in roots. A'cre Phv-tologist 84: 4K')-500.
Gregory PH. 1982. Fairy rings; free and tethered. Bulletin of theBritish Mycolngical Society 16: I61-]f)3.
Griffiths RP, CaldweJJ BA, 1992. Mycorrhiza) mar communitiesin forest soils. In: Read DJ, Lewis DH, Fitter .AH & Alexander]J. eds. Mycorrhizas in Ecosystems. Wallingford: C.A.B.International, 98-105.
Harvey AE, Larsen MJ, Jurgensen MF. 1979. Comparativedistribution of ectomycorrhizae in soils of three westernMontana forest habitat types. Forest Science 25: 350-358.
Hausting M, Marschner H. 1989, Organic and inorganic soilphosphates and soil pho.sphatase activity in the rhizosphere of8(l-year'otd Norway spruce [Picea abies (L) Karst.] trees.Biology and Fertility of Soils 8: 128 - 133.
Hingston FJ, O'Conneil AM, Grove TS. 1989. Nutrient cyclingin jarrah forest. In : Del] B, Havel JJ, Malajczuk N , eds. TheJarrnh Forest. Dordrech t : Kluwer Academic Publishers ,"155-177.
Jasper DA, Abbott LK, Robson AD. 1989a. Soil disturbancereduces tht infi-ctivity of external hyphae of VA mycorrhizalfungi, A'ew FhytologisI 112: 93-99,
Jasper DA, Abbott LK, Robson AD. 19896. Hyphae of a \.\mycorrhizal fungus maintain infectivity in dry soiS, exceptwhen the soil is disturbed. A'w PhytologisI 112: 101- 107.
Jasper DA, Abbott LK, Robson AD. 1991. The eift-ct of soi!disturbance on vesicular-arbuscular mytorrhizal fungi m soilsfrom different vt-gt-tation types. Nejc PhyUdo^ist 118: 471—^76.
Jasper DA, Abbott LK, Robson AD. 1992. Soil disturbance in
native ecosystems - the decline and recovery of infectivity ofVA mycorrhizal fungi. In: Read DJ, Lewis DH, Fitter AH,.A.lexander IJ, eds. Mycorrhizas iv Ecosystems. Wallingford:CA.B, International, 151-155,
Lamont BB. 1993. Why are hair>' root clusters so abundant in themost nutrient-impovenshed soils of Australia? In: Barrow NJ,ed. Plant Nutrition -frum Genetic Engineering to Field Practice.Dordrecht: Kluwer .\cademjc Publishers, 309-312.
Ogawa M. 1985. Ecological characters of ectomycorrhizal fungiand their mycorrhizae. An mtroduction to the ecology of higherfunf-i. yapanese Agriculture Research Quarterly 18: 305-314.
Reddell P, Malajczuk N. 1984. Formation of mycorrhizae byjarrah {Eucalyptus marginata Donn ex Smith) in litter and soil.Australian Journal 'if Botany 32: 511-520.
Rice EL. 1984. Allelopathy. New York: Academic Press.Schweiger PFG. 1994. Factors affecting VA mycorrhizal uptake
of phosphorus. Ph.D.thesis. University of Western Australia,Perth.
Skinner MF, Bowen GD. 1974. The penetration of RO'\\ bymycelial strands of ectomycorrhizal fungi. Soil Biology andBiochemistry 6; 57 -61.
St John TV, Coieman DC, Reid CPP. 1983. Growth and spatiaidistribution of nutrient-absorhing organs: selective exploitationof .soil heterogeneity. Plant and Soil 71: 487—1-93.
St John TV, Hunt HW. 1983. Statistical treatment of VAMinfection data. Plant and Soil 73: 307-31.''.
Sylvia DM. 1986. Spatial and temporal distribution of vesicular-arhuscular mycorrhizal fungi associated with L'niola paniculatain Florida foredunes. Mvcologia 75: 728—734.
Taylor RJ. 1992. Distribution and abundance of fungal sporo-carps and diggings of the Tasmanian hettong, Bettongiagaimardi. Australian Journal of Ecology 17: 1 55-160,
Tews LL, Koske RE. 1986. Towards a sampling strategy forvesicular-arbuscular mycorrhizas. Transactions of the BritishMycological Society 87: 353-358.
Tilman D. 1994. Competition and biodiversity in spatiallystructured habitats. Ecology 75: 2- !6.
Walker C, Mize CW, McNab HS Jr. 1982. Populations ofendogonaceous fungi at two locations m central Iowa. Canadianyournal of botany 60: 25. ] 8- 2 529.
Warner A. 1984. Colonization of organic matter by vesicular-arbuscular mycorrhiza] fungi. Transactions of the BritishMycological Society 82: 352-354.






























