Embed Size (px)
Citation preview

J. Anat.
(2005)
207
, pp155–164
© Anatomical Society of Great Britain and Ireland 2005
Blackwell Publishing, Ltd.
Morphogenesis of the fibrous sheath in the marsupial spermatozoon
M. Ricci and W. G. Breed
Department of Anatomical Sciences, The University of Adelaide, S.A., Australia, 5005
Abstract
The spermatozoon fibrous sheath contains longitudinal columns and circumferential ribs. It surrounds the axoneme
of the principal piece of the mammalian sperm tail, and may be important in sperm stability and motility. Here we
describe its assembly during spermiogenesis in a marsupial, the brush-tail possum, and compare its structural organiza-
tion with that of eutherian mammals, birds and reptiles. Transmission electron microscopy showed that possum
fibrous sheath assembly is a multistep process extending in a distal-to-proximal direction along the axoneme from steps
4 to 14 of spermiogenesis. For the most part, assembly of the longitudinal columns occurs before that of the circum-
ferential ribs. Immunohistochemical and immunogold labelling showed that fibrous sheath proteins are first present
in the spermatid cytoplasm; at least some of the proteins of the sheath precursors differ from those in the mature
fibrous sheath. That immunoreactivity develops after initiation of chromatin condensation suggests that fibrous sheath
proteins, or their mRNAs, are stored within the spermatid cytoplasmic lobule prior to their assembly along the axoneme.
These findings are similar to those in laboratory rats, and thus suggests that the mode of fibrous sheath assembly
evolved in a common ancestor over 125 million years ago, prior to the divergence of marsupial and eutherian lineages.
Key words
spermatozoon; fibrous sheath; marsupial.
Introduction
In the flagellum of the spermatozoon of eutherian
mammals, the fibrous sheath surrounds the axoneme and
outer dense fibres of the principal piece. It is composed
of two structurally distinct segments, the dorsal and
ventral longitudinal columns
,
and a connecting array
of circumferential ribs (Fawcett, 1970, 1975). For many
years, the fibrous sheath was viewed as a passive, mechan-
ical component of the sperm flagellum (Fawcett, 1975)
that provides elastic rigidity to the sperm tail and/or
defines the shape of its beat by placing a constraint
on its plane of bending (Phillips, 1972; Fawcett, 1975;
Lindemann et al. 1992; Si & Okuno, 1993; Carrera et al. 1994;
Jassim, 1995). However, as some of its individual proteins
have become isolated and sequenced, it has become
increasingly clear that this structure also acts as a scaffold
for constituents of signalling cascade events (Carrera
et al. 1994, 1996; Fulcher et al. 1995; Vigayaraghavan
et al. 1997, 1999; Miki et al. 2002), as well as for glycolytic
enzymes (Mori et al. 1992; Welch et al. 1992, 1995).
Consequently, in addition to a structural function, the
eutherian fibrous sheath appears to have an import-
ant role in sperm motility (for a review see Eddy et al.
2003). Furthermore, comparative studies show that a
fibrous sheath-like structure is present in sperm tails of
reptiles (Harding et al. 1995; Scheltinga et al. 2001) and
non-passerine birds, with it becoming lost during the
evolution of passerines (Baccetti & Afzelius, 1976; Jones
& Lin, 1993; Lin & Jones, 1993). To date, the extent of
conservation of its molecular composition, and function,
across Amniota is unknown, although differences in ultras-
tructure suggest that its morphology and perhaps function
vary between species (Fawcett, 1970).
The formation of the fibrous sheath in eutherian
mammals has, to date, only been described in detail for
the laboratory rat (Irons & Clermont, 1982b) in spite of
the interspecific differences in its morphology. Trans-
mission electron microscopy and radioautography
Correspondence
Mario Ricci, Department of Anatomical Sciences, The University of Adelaide, S.A., 5005, Australia. T: +61 88303 6294; F: +61 88303 4398; E: [email protected]
Accepted for publication
10 May 2005

Morphogenesis of the fibrous sheath in the marsupial spermatozoon, M. Ricci and W. G. Breed
© Anatomical Society of Great Britain and Ireland 2005
156
have shown that its morphogenesis involves a lengthy,
multistep process extending from steps 2 to 17 of
spermiogenesis. Moreover, in contrast to the forma-
tion of the outer dense fibres, in the laboratory rat
fibrous sheath morphogenesis has been shown to
proceed in a distal-to-proximal direction (Irons &
Clermont, 1982a) with its proteins being evident within
the spermatid cytoplasm prior to their assembly along
the axoneme (Oko & Clermont, 1988; Clermont et al.
1990).
Little is known of fibrous sheath morphogenesis
in marsupial sperm. Sapsford et al. (1969, 1970) showed
that longitudinal column precursors occur in early sper-
matids in the long-nosed bandicoot (
Perameles nasuta
)
(Peramelidae), but their morphogenesis does not appear
to have been studied in any other marsupial. The
bandicoots diverged from the possums (Phalangeridae)
early in evolution of the Australian marsupials at least
50 million years ago (Kirsch et al. 1997; Amrine-Madsen
et al. 2003; Asher et al. 2004; Cardillo et al. 2004; Nilsson
et al. 2004). In the brush-tail possum (
Trichosurus
vulpecula
) the number and molecular weights of the
major fibrous sheath proteins have recently been found
to differ considerably from those of the laboratory rat
(Harris & Rodger, 1998; Ricci & Breed, 2001). Although
several ultrastructural studies of possum spermiogen-
esis have already been carried out (Harding et al. 1976;
Lin et al. 2004), little data are available on sperm tail
morphogenesis. In this study, we describe the assembly
of the fibrous sheath in the brush-tail possum and, using
polyclonal antibodies for immunocytochemistry, we have
determined the time and site of formation of fibrous
sheath proteins during spermiogenesis.
Materials and methods
Experimental animals
Adult brush-tail possums housed at the Central Animal
House of the University of Adelaide were fed a diet
of mixed cereals and fruit daily with water available
ad libitum
. The University of Adelaide Ethics Committee
approved all animal experimentation.
Transmission electron microscopy
Possums were anaesthetized with isofluorane (4%, 1.5 L
min
−
1
, DBL, Mulgrave, Victoria, Australia), and the testes
fixed by vascular perfusion through the abdominal aorta.
The vascular system was first rinsed with heparinized
physiological saline containing 2.5% polyvinyl pyrolidone
and 0.5% procaine hydrochloride until blood outflow
had ceased, followed by perfusion with 3% parafor-
maldehyde/3% glutaraldehyde in 0.1
M
phosphate buffer,
pH 7.4, containing 2.5% polyvinyl pyrolidone for 5–10
min or until blood vessels had cleared. The testes were
then dissected free, cut into small cubes (
∼
1 mm
3
) and
immersed in the above fixative for 2 h. Immersion and
perfusion-fixed tissue was rinsed in two changes of
0.1
M
phosphate buffer, pH 7.4, for 10 min each, and
post-fixed in 1% osmium tetroxide in PBS, pH 7.4,
for 1 h. Tissue was dehydrated by passing through a
graded series of ethanols, cleared in two changes of
propylene oxide, and infiltrated in a 1 : 1 ratio of pro-
pylene oxide/resin overnight. It was then embedded in
pure resin (TK3, TAAB Laboratories, Berkshire, UK) and
blocks were polymerized at 60
°
C for at least 48 h. Thick
(0.5–1.0
µ
m) plastic sections were cut using a Reichert–
Jung Ultracut ultramicrotome with a glass knife and
stained with 0.25% toluidine blue in 0.5% sodium
tetraborate prior to viewing under an Olympus BH-2
light microscope. Areas of interest were selected from
thick plastic sections, the blocks trimmed and ultrathin
sections of silver/gold interference colours (0.02–0.1
µ
m)
were then cut with a diamond knife (Diatome Ltd, Bienne,
Switzerland). Sections were collected onto copper/
palladium grids (200 mesh), stained with uranyl acetate
and lead citrate, and viewed with a Phillips CM100
transmission electron microscope at 80 kV.
Polyclonal antibody preparation
The polyclonal anti-possum fibrous sheath serum was
prepared as previously described (Ricci & Breed 2001).
Briefly, the fibrous sheath was isolated by a combina-
tion of sonication, incubation in 4.5
M
urea and 25 m
M
DTT, and sucrose density gradient centrifugation. The
fibrous sheaths were then solubilized, emulsified in an
equal volume of Freund’s complete adjuvant (Sigma,
St. Louis, MO, USA), and approximately 50
µ
g of protein
was injected either intraperitoneally or subcutaneously
into Sprague–Dawley rats. The rats were boosted with
50
µ
g of solubilized protein in an equal volume of
Freund’s incomplete adjuvant at 2-week intervals for
6 weeks, and test bleeds were carried out 7 days after
each boost. After collection, the serum was stored
at
−
70
°
C. Pre-immune sera were collected from all rats
prior to immunization.

Morphogenesis of the fibrous sheath in the marsupial spermatozoon, M. Ricci and W. G. Breed
© Anatomical Society of Great Britain and Ireland 2005
157
Light microscope immunohistochemistry
Adult male possums were anaesthetized with isoflurane,
and testes and epididymides were fixed by perfusion for
20 min with Bouin’s fixative. Testes were excised, cut into
1-m
3
cubes and immersion-fixed in Bouin’s fixative for 2 h.
Then tissue was cut into smaller, 5-mm
3
cubes, re-immersed
in fixative for 2 h and dehydrated by passing through
a graded series of ethanols. Tissue was infiltrated with
paraffin wax, and 5-
µ
m sections were cut on a Leica
microtome and floated onto slides. The remaining steps
in this protocol were adapted from the protocol of Oko
& Clermont (1989). Deparaffinization of these sections
followed standard procedures except that during hydra-
tion (i) sections were immersed in 70% ethanol containing
1% (w/v) lithium carbonate to inactivate residual picric
acid, and (ii) endogenous peroxidase activity was elim-
inated by incubating sections in methanol containing
1% (v/v) hydrogen peroxide for 10 min. Hydrated sections
were subsequently washed for 5 min in distilled H
2
O
containing 300 m
M
glycine to block any free aldehyde
groups, then rinsed in 20 m
M
Tris/HCl saline (TBS), pH 7.4.
Non-specific binding of IgG was blocked by incubat-
ing sections in 10% normal goat serum (NGS) in TBS for
30 min. Sections were then incubated for 1 h with the
primary antibody diluted 1 : 100 in TBS-Tween con-
taining 1% NGS, and washed six times for 5 min
each in TBS-Tween containing 1% NGS, blocked with
10% NGS, before incubation with biotin-labelled goat
anti-rat immunoglobulin (Sigma) diluted 1 : 25 in TBS
for 1 h. The sections were washed in TBS-Tween and
incubated in streptavidin-alkaline phosphatase (SA-AP)
diluted 1 : 3000 in TBS for 1 h, washed again, and antibody
binding visualized by incubating sections with diamino-
benzidene tetrahydrochloride (Zymed, San Francisco,
CA, USA). Finally, sections were lightly counterstained
with haematoxylin, dehydrated by passing through a
graded series of ethanols and mounted under glass
coverslips with DPX mounting medium.
On all control slides pre-immune sera were used in
place of primary antibody. Immunostaining was repeated
on sections of testes from four different animals for
reproducibility.
Immunogold electron microscopy
Possums were anaesthetized with isoflurane, and testes
and epididymides were fixed by perfusion for 20 min
with 4% paraformaldehyde in 0.2
M
PBS, pH 7.4. Tissue
was then dissected free and immersed in the same
fixative for 2 h, washed in two changes of PBS, and
dehydrated by passing through a graded series of
ethanols. The tissue was infiltrated in a 2 : 1 ratio of
LR White/70% ethanol for 1 h, followed by two further
incubations in pure LR White resin for 1 h and then
overnight. The resin was again replaced just prior
to polymerization in gelatin capsules at 60
°
C for 24 h.
Ultrathin sections were cut on a Reichert–Jung ultracut
microtome and mounted on nickel grids.
Sections on grids were blocked by incubating in 10%
NGS in TBS for 30 min, and then incubated with the
primary antibody diluted 1 : 100 in TBS-Tween containing
1% NGS for 1 h. They were washed six times for 5 min
each in TBS-Tween, and further blocked in 10% NGS for
15 min before being incubated in 10-nm gold-conjugated
goat anti-rat immunoglobulin (Sigma), diluted 1 : 20,
for 1 h at room temperature. They were then subject to
six 5-min washes in TBS-Tween, four washes in double
distilled water, before staining with uranyl acetate (3 min)
and examination on a Phillips CM100 transmission
electron microscope at 80 kV.
Results
Formation of possum fibrous sheath
Possum fibrous sheath morphogenesis is described using
the ten stages of the seminiferous epithelium cycle and
the 14 steps of spermatid development as described by
Lin et al. (2004).
During steps 1–3 of spermiogenesis, the axoneme
developed from the distal centriole but a precursor to
the fibrous sheath was not yet visible. Sheath assembly
first began in step 4 spermatids at the distal end of the
flagellum. Electron-dense material accumulated there
between the plasmalemma and microtubule doublets
3 and 8 of the axoneme, forming the precursors of the
longitudinal columns (Fig. 1a). In step 5 spermatids, the
precursors of the longitudinal columns had lengthened
in a proximal direction along the flagellum. In step 6
spermatids, considerable electron-dense material had
accumulated beneath the plasmalemma in the distal
segment of the flagellum attached to the margins of each
of the thickened column anlagen. In longitudinal sections,
this material presented as a series of parallel, evenly
spaced, circumferentially orientated striations and there-
fore are presumably the precursors of the circumferential
ribs (Fig. 1b,c).

Morphogenesis of the fibrous sheath in the marsupial spermatozoon, M. Ricci and W. G. Breed
© Anatomical Society of Great Britain and Ireland 2005
158
In step 8 spermatids, electron-dense material accu-
mulated between adjacent pairs of the rib precursors
(Fig. 1d), which subsequently thickened and began to
resemble the ribs in mature sperm. During step 9, these
coalescenced further to form the larger, definitive,
ribs of the fibrous sheath (Fig. 1e). The number of rib
precursors that fused at this stage ranged from 2–3 in the
most distal segment of the developing principal piece
to 6–12 in the most proximal segment where the develop-
ing fibrous sheath was therefore considerably larger.
These ribs were united at each of their ends to the
longitudinal columns, which simultaneously thickened
and grew in a distal-to-proximal direction.
During step 11, the longitudinal columns appeared
considerably larger, although the newly formed circum-
ferential ribs were conspicuously electron lucent, perhaps
reflecting their early state of development (Fig. 1f), and
projected away from the axoneme to give the flagellum
an ovoid cross-sectional shape. During steps 12 and 13,
as the ribs coalesced and thickened, small, electron-lucent
spaces appeared within the longitudinal columns of the
fibrous sheath (Fig. 1g). The final step of spermiogenesis
Fig. 1 Transverse (a,f) and longitudinal (b–e,g) sections through possum spermatids showing the development of the fibrous sheath (FS). The anlagen of the longitudinal columns (ALC) first appear in step 4 spermatids adjacent to microtubules 3 and 8 of the axoneme (a), whereas the anlagen of the circumferential ribs (RA) first appear in step 6 spermatids as a series of evenly spaced striations (b,c). The rib anlagen thicken and coalesce during steps 8 (d) and 9 (e) to form mature ribs (R). During step 11, the ribs project away from the axoneme and the longitudinal columns (LC) increase in size (f). The ribs coalesce and thicken further as spermiogenesis progresses, although electron-lucent spaces can be seen in the columns in step 13 spermatids. Scale bars: (a) 0.1 µm, (b) 0.3 µm, (c) 0.03 µm, (d–f) 0.25 µm, (g) 0.3 µm.
Morphogenesis of the fibrous sheath in the marsupial spermatozoon, M. Ricci and W. G. Breed
© Anatomical Society of Great Britain and Ireland 2005
159
was characterized by a gradual increase in size of the
fibrous sheath.
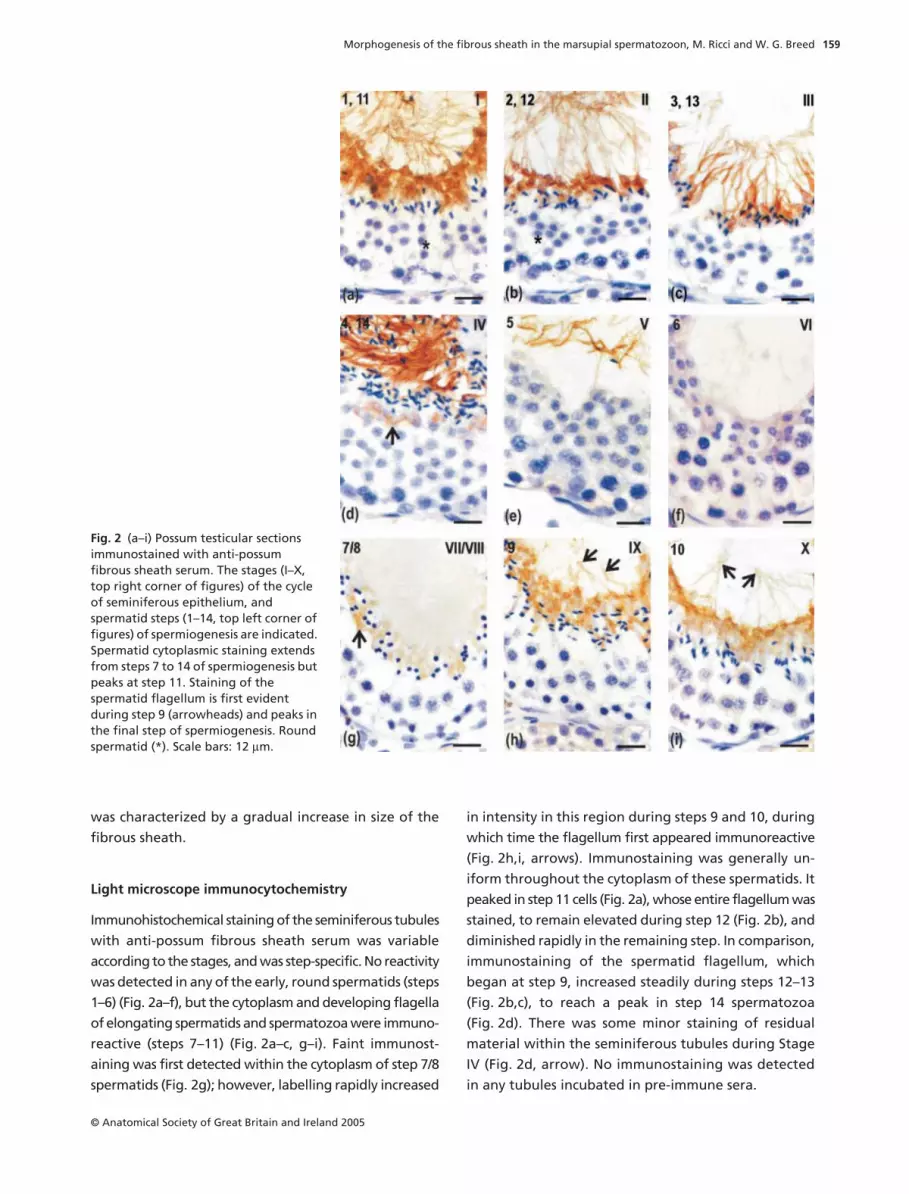
Light microscope immunocytochemistry
Immunohistochemical staining of the seminiferous tubules
with anti-possum fibrous sheath serum was variable
according to the stages, and was step-specific. No reactivity
was detected in any of the early, round spermatids (steps
1–6) (Fig. 2a–f), but the cytoplasm and developing flagella
of elongating spermatids and spermatozoa were immuno-
reactive (steps 7–11) (Fig. 2a–c, g–i). Faint immunost-
aining was first detected within the cytoplasm of step 7/8
spermatids (Fig. 2g); however, labelling rapidly increased
in intensity in this region during steps 9 and 10, during
which time the flagellum first appeared immunoreactive
(Fig. 2h,i, arrows). Immunostaining was generally un-
iform throughout the cytoplasm of these spermatids. It
peaked in step 11 cells (Fig. 2a), whose entire flagellum was
stained, to remain elevated during step 12 (Fig. 2b), and
diminished rapidly in the remaining step. In comparison,
immunostaining of the spermatid flagellum, which
began at step 9, increased steadily during steps 12–13
(Fig. 2b,c), to reach a peak in step 14 spermatozoa
(Fig. 2d). There was some minor staining of residual
material within the seminiferous tubules during Stage
IV (Fig. 2d, arrow). No immunostaining was detected
in any tubules incubated in pre-immune sera.
Fig. 2 (a–i) Possum testicular sections immunostained with anti-possum fibrous sheath serum. The stages (I–X, top right corner of figures) of the cycle of seminiferous epithelium, and spermatid steps (1–14, top left corner of figures) of spermiogenesis are indicated. Spermatid cytoplasmic staining extends from steps 7 to 14 of spermiogenesis but peaks at step 11. Staining of the spermatid flagellum is first evident during step 9 (arrowheads) and peaks in the final step of spermiogenesis. Round spermatid (*). Scale bars: 12 µm.

Morphogenesis of the fibrous sheath in the marsupial spermatozoon, M. Ricci and W. G. Breed
© Anatomical Society of Great Britain and Ireland 2005
160
Immunogold electron microscopy
No immunogold labelling was detected in step 1–5
spermatids. During steps 6–10 there was no, or only very
minor, non-specific labelling over the anlagen of the
longitudinal columns and circumferential ribs (Fig. 3a–c).
By contrast, an increased labelling over the spermatid
cytoplasm during this period reached a peak in step 11
cells (Fig. 3b,d). In step 12 spermatids, there was a marked
increase in immunogold labelling over the abaxial
cytoplasm, especially in the proximal segment of the
flagellum (Fig. 3e). From step 11, specific immunogold
labelling was seen over both the longitudinal columns
and the circumferential ribs of the fibrous sheath
itself, this labelling peaking at step 14 (Fig. 3f,g). No
immunogold labelling was evident in any sections
incubated in pre-immune sera.
Discussion
In the current study, the morphogenesis of the fibrous
sheath is described for sperm of the brush-tail possum.
Fig. 3 Transverse (a,e,g) and longitudinal (b–d,f) sections through possum spermatids treated with anti-possum fibrous sheath serum. These show the development of the fibrous sheath. Immunogold labelling of the flagellum does not occur or occurs very sparsely in step 6 (a,b) and step 8 (c) spermatids. However, the cytoplasm lobule is reactive from step 6 (b) to step 11 (d) (arrowheads). In step 12 spermatids (e) there is intense immunogold labelling beneath the plasmalemma (arrowheads), suggesting that these proteins migrate down the flagellum before becoming incorporated into the fibrous sheath (FS). Immunogold labelling over the fibrous sheath reaches a peak in step 14 spermatids (f,g). Anlagen of the longitudinal columns (ALC), anlagen of the circumferential ribs (RA), immunogold labelling (arrowheads). Scale bars: (a,c) 0.1 µm, (b,d,f) 0.15 µm, (e) 0.06 µm, (g) 0.2 µm.

Morphogenesis of the fibrous sheath in the marsupial spermatozoon, M. Ricci and W. G. Breed
© Anatomical Society of Great Britain and Ireland 2005
161
Transmission electron microscopy has shown that this
sheath is morphologically similar to that of the long-nosed
bandicoot (Sapsford et al. 1969, 1970) and laboratory
rat (Irons & Clermont, 1982b) in spite of differences in
its final form. In these three species it has been found
that (1) the precursors of the longitudinal columns of
the fibrous sheath first assemble adjacent to axoneme
microtubules 3 and 8 in early spermatids, (2) the precursors
of the ribs appear later as circumferential striations that
gradually enlarge and coalesce, (3) growth of the columns
and ribs is largely independent of each other except for
late in spermiogenesis when their development occurs
concurrently, and (4) fibrous sheath assembly takes place
in a distal-to-proximal direction along the sperm tail.
One aim of the current study was to differentiate, by
immunohistochemistry, the time and site of formation
of the components of the fibrous sheath using a possum
fibrous sheath polyclonal antibody. This antiserum has
previously been shown, by Western blotting, to label
specifically the major possum fibrous sheath proteins,
but not any others, including those of the outer dense
fibres, thus suggesting that the sheath is composed of
a unique suite of proteins (Ricci & Breed, 2001). Further-
more, this serum also recognizes fibrous sheath proteins
from species in several other marsupial families (e.g.
tammar wallaby: Macropodidae; koala: Phascolarctidae;
and fat-tailed dunnart: Dasyuridae), which diverged from
the Phalangeridae at least 40–50 million years ago (Kirsch
et al. 1997; Asher et al. 2004; Nilsson et al. 2004), thus
suggesting an early origin of some of the fibrous sheath
proteins. In addition, the antibody also stained some
proteins in the fibrous sheath of rat sperm (Ricci &
Breed, 2001), indicating their conservation across
marsupial and eutherian lineages. The immunolabelling
data of the current study indicate that the timing of
fibrous sheath formation is, like its method of assembly,
similar in marsupials and eutherians [for data on the
laboratory rat see Oko (1988), Oko & Clermont (1989)
and Clermont et al. (1990)]. In the possum, peak cyto-
plasmic immunoreactivity occurs in step 10 spermatids,
well after chromatin condensation has become initiated
(Lin et al. 2004). Thus, as in rats (Oko, 1988; Oko &
Clermont, 1989; Clermont et al. 1990; El-Alfy et al. 1999),
some of the fibrous sheath proteins are translationally
regulated. The fact that there is marked immunolabel-
ling of the periaxonemal cytoplasm of step 11 possum
spermatids makes it likely that fibrous sheath proteins
are transferred from the spermatid cytoplasmic lobule to
the developing fibrous sheath late in spermiogenesis.
The proteins of the anlagen of the longitudinal columns
and circumferential ribs of possum spermatids are not
labelled with the anti-fibrous sheath serum and there-
fore are presumably dissimilar to those of the mature
fibrous sheath. Either the precursor structure is com-
posed of a different suite of proteins from those of the
mature fibrous sheath or these proteins are processed
during morphogenesis. Immunocytochemical analysis
of fibrous sheath formation in the laboratory rat (Oko
& Clermont, 1989; Clermont et al. 1990), mouse (Sakai
et al. 1986; Fenderson et al. 1988; Brown et al. 2003),
human (Jassim et al. 1991) and the cockerel (Bozkurt &
Holley, 1995) has yielded similar results, suggesting that
the primitive column and rib anlagen might somehow
act as organizers, or triggers, of fibrous sheath assembly
(Oko, 1988; Clermont et al. 1990) in both mammals and
birds. In the mouse, it has been suggested that a fibrous
sheath protein A-Kinase Anchoring Protein 3 (AKAP3),
which is synthesized in round spermatids and incorpo-
rated into the fibrous sheath concurrently with formation
of the rib anlagen, may be responsible for organizing
the basic structure of the fibrous sheath (Brown et al.
2003).
The current study indicates that the morphogenesis
of the fibrous sheath has been conserved across the
two major extant mammalian subclasses, despite their
divergence over 125 million years ago (Cifelli & Davis,
2003; Woodburne et al. 2003; Asher et al. 2004; Nilsson
et al. 2004). Taken together with our previous conclusion
that the 62- and 76-kDa fibrous sheath proteins have
been conserved between sperm of possum, koala, wallaby
and dunnart, as well as laboratory rat (Ricci & Breed,
2001), this suggests that an increased complexity of the
sperm flagellum evolved in a common ancestor prior to
the divergence of the eutherian and marsupial lineages.
In mammals, the thickness of the sheath’s circumfer-
ential ribs and the prominence of its longitudinal
columns varies between species. For example, in a bat
species,
Myotis lucifugus
, the ribs are very broad and
flat and the longitudinal columns thin, whereas the
Chinese hamster
Cricetulus griseus
sperm tail displays
thin ribs but broad columns (Fawcett, 1970). In both the
American opossum,
Didelphis virginiana
(Fawcett, 1970),
and brush-tailed possum, the longitudinal columns are
relatively narrow, but the ribs are greatly expanded
adjacent to the columns, and there is a pyramidal electron-
lucent region adjacent to the longitudinal columns. In
the monotreme
Ornithorhynchus anatinus
, the columns
are poorly developed with the sheath being almost

Morphogenesis of the fibrous sheath in the marsupial spermatozoon, M. Ricci and W. G. Breed
© Anatomical Society of Great Britain and Ireland 2005
162
entirely a series of helically orientated dense loops
(Carrick & Hughes, 1982; Lin & Jones, 2000). An accessory
sperm tail structure, closely resembling the mammalian
fibrous sheath, also occurs in reptiles (Furieri, 1970;
Harding et al. 1995; Ishmail & Dehlawi, 1995; Scheltinga
et al. 2001) and non-passerine birds (Baccetti & Afzelius,
1976; Burgess et al. 1991; Jones & Lin, 1993; Lin & Jones,
1993). In non-passerine birds, this structure has been
described as an amorphous, irregularly beaded, fila-
mentous mesh (Baccetti & Afzelius, 1976; Thurston &
Hess, 1987; Burgess et al. 1991; Jones & Lin, 1993; Lin
& Jones, 1993; Jamieson et al. 1995; Jamieson, 1995),
whereas in squamates, a thick, rib-like coat extends into
the midpiece of the flagellum (Jamieson & Healy, 1992;
Harding et al. 1995; Ishmail & Dehlawi, 1995; Scheltinga
et al. 2001; Ferreira & Dolder, 2002). The presence in at
least some bird and reptile spermatozoa of a sheath-
like structure, or an amorphous sheath, suggests a
somewhat similar structure to that of the mammalian
fibrous sheath. However, whether it is biochemically
and/or functionally comparable is unknown. Although
there are no specific studies of bird and reptile fibrous
sheath morphogenesis, this structure appears early in
spermiogenesis in both the quail (Lin & Jones, 1993), and
a species of neotropical lizard,
Tropidurus torquantas
(Vieira et al. 2001), and it assembles in a distal-to-proximal
direction along the axoneme in the common lizard
Lacerta vivipara
(Courtens & Depeiges, 1985).
The presence of a fibrous sheath, or a fibrous sheath-
like structure, in spermatozoa from reptiles, birds and
mammals, coupled with the current finding that its
morphogenesis is similar in eutherian and marsupial
mammals, suggests an important function(s) for it in
sperm of all amniotes. In the rat and mouse, the most
abundant fibrous sheath protein is an A-kinase anchor-
ing protein, AKAP4 (Carrera et al. 1994, 1996; Fulcher
et al. 1995) which may anchor cAMP-dependent pro-
tein kinase A (Vijayaraghavan et al. 1997; Colledge &
Scott, 1999; Miki et al. 2002; for a review see Eddy et al.
2003). The molecular weight of AKAP4 is 76 kDa and,
because possum fibrous sheath extracts have a major
protein of similar molecular weight (Ricci & Breed,
2001), it too may contain much AKAP4. In addition,
using both immunofluorescence and immunogold
microscopy, we have recently found that an antibody
raised against mouse spermatogenic glycolytic path-
way enzyme glyceraldehyde 3-phosphate dehydroge-
nase (GAPDS) (Fenderson et al. 1988; Welch et al.
1992; Bunch et al. 1998) binds intensely to the possum
fibrous sheath by both immunofluorescence and
immunogold microscopy (M.R., M. Eddy and W.G.B.,
unpublished observations). Thus, in the possum GAPDS
also appears to be a major fibrous sheath protein and
may well provide its sperm with an energy source. A
more detailed investigation of the molecular biology
of the marsupial fibrous sheath may shed light on the
proteins present in this sperm tail cytoskeletal struc-
ture. Such findings will further extend our understand-
ing of the evolution of this accessory cytoskeletal
structure of the sperm tail that appears to have evolved
around the time of the evolution of internal fertiliza-
tion and viviparity in early amniotes.
Acknowledgements
We thank Chris Leigh for his technical assistance.
M.R. was in receipt of an Australian Postgraduate Award.
This work was supported, in part, by an ARC grant
References
Amrine-Madsen H, Scally M, Westerman M, Stanhope MJ,Krajewski CW, Springer MS
(2003) Nuclear gene sequencesprovide evidence for the monophyly of Australidelphianmarsupials.
Mol Phylogenetics Evol
28
, 186–196.
Asher RJ, Horovitz I, Sanchez-Villagra MR
(2004) First com-bined cladistic analysis of marsupial mammal interrelation-ships.
Mol Phylogenetics Evol
33
, 240–250.
Baccetti B, Afzelius BA
(1976) The biology of the sperm cell.
Monogr Dev Biol
10
, 1–254.
Bozkurt HH, Holley MC
(1995) Identification of a 53-kDaantigen in the fibrous sheath of avian spermatozoa.
JReprod Immunol
29
, 149–160.
Brown PR, Miki K, Harper DB, Eddy EM
(2003) A-kinaseanchoring protein 4 binding proteins in the fibrons sheathof the sperm flagellum.
Biol Reprod
68
, 2241–2248.
Bunch DO, Welch JE, Magyar PL, Eddy EM, O’Brien DA
(1998)Glyceraldehyde 3-phosphate dehydrogenase-S proteindistribution during mouse spermatogenesis.
Biol Reprod
58
, 834–841.
Burgess SA, Dover SD, Woolley DM
(1991) Architecture of theouter arm dynein ATPase in an avian sperm flagellum, withfurther evidence for the B-link.
J Cell Sci
98
, 17–26.
Cardillo M, Bininda-Emonds ORP, Boakes E, Purvis A
(2004) Aspecies-level phylogenetic supertree of marsupials.
J Zool
264
, 11–31.
Carrera A, Gerton GL, Moss SB
(1994) The major fibrous sheathpolypeptide of mouse sperm: structural and functionalsimilarities to the A-kinase anchoring protein.
Dev Biol
165
,272–284.
Carrera A, Moss J, Ning XP, Gerton GL, Kopf GS, Moss SB
(1996) Regulation of protein tyrosine phosphorylationin human sperm by a calcium/calmodium-dependentmechanism: identification of A kinase anchor proteins as

Morphogenesis of the fibrous sheath in the marsupial spermatozoon, M. Ricci and W. G. Breed
© Anatomical Society of Great Britain and Ireland 2005
163
major substrates for tyrosine phosphorylation.
Dev Biol
180
,284–296.
Carrick FN, Hughes RL
(1982) Aspects of the structure anddevelopment of monotreme spermatozoa and their relevanceto the evolution of mammalian sperm morphology.
CellTissue Res
222
, 127–141.
Cifelli RL, Davis BM
(2003) Marsupial origins.
Science
302
,1899–1900.
Clermont Y, Oko R, Hermo L
(1990) Immunocytochemicallocalization of proteins utilized in the formation of outerdense fibres and fibrous sheath in rat spermatids: an electronmicroscope study.
Anat Rec
227
, 447–457.
Colledge M, Scott JD
(1999) AKAPs: from structure to function.
Trends Cell Biol
9
, 216–221.
Courtens JC, Depeiges A
(1985) Spermiogenesis of
Lacertavivipara
.
J Ultrastruct Res
90
, 203–220.
Eddy EM, Toshimori K, O’Brien DA
(2003) Fibrous sheath ofmammalian spermatozoa.
Microsc Res Technique
61
, 103–115.
El-Alfy M, Moshonas D, Morales CR, Oko R
(1999) Molecularcloning and developmental expression of the major fibroussheath protein (FS 75) of rat sperm.
J Androl
20
, 307–318.
Fawcett DW
(1970) A comparative view of sperm ultrastruc-ture.
Biol Reprod
2
, 90–127.
Fawcett DW
(1975) The mammalian spermatozoon.
Dev Biol44, 394–436.
Fenderson BA, Toshimor K, Muller CH, Lane TF, Eddy EM(1988) Identification of a protein in the fibrous sheath of thesperm flagellum. Biol Reprod 38, 345–357.
Ferreira A, Dolder H (2002) Ultrastructural analysis of spermio-genesis in Iguana iguana (Reptilia: Sauria: Iguanidae). Eur JMorphol 40, 89–99.
Fulcher KD, Mori C, Welch JE, O’Brien DA, Klapper DG,Eddy EM (1995) Characterization of Fsc1 cDNA for a mousesperm fibrous sheath component. Biol Reprod 52, 41–49.
Furieri P (1970) Sperm morphology in some reptiles: Squamataand Chelonia. In Comparative Spermatology (ed. Baccetti B),pp. 115–131. Academia Nazionate dei Lincei. Rome, Italy.
Harding HR, Carrick FN, Shorey CD (1976) Spermiogenesis inthe brush-tailed possum, Trichosurus vulpecula (Marsupialia).The development of the acrosome. Cell Tissue Res 171, 75–90.
Harding HR, Aplin KP, Mazur M (1995) Ultrastructure of sper-matozoa of Australian blindsnakes, Ramphotyphlops spp.(Typhlopidae, Squamata): first observations on the maturespermatozoon of Scolecophidian snakes. In Advances inSpermatozoal Phylogeny and Taxonomy (eds Jamieson BG,Ausio J, Justine J-L), Mem Mus National Histoire Naturelle166, 385–396.
Harris MS, Rodger JC (1998) Characterisation of fibrous sheathand midpiece fibre network polypeptides of marsupialspermatozoa with a monoclonal antibody. Mol Reprod Dev50, 461–473.
Irons MJ, Clermont Y (1982a) Formation of the outer densefibres during spermiogenesis in the rat. Anat Rec 202,463–471.
Irons MJ, Clermont Y (1982b) Kinetics of fibrous sheath formationin the rat spermatid. Am J Anat 165, 121–130.
Ishmail MF, Dehlawi GY (1995) Ultrastructure of spermiogen-esis of Saudian reptiles. Sperm tail differentiation in Steno-dacetylus selvini. J Environ Sci 10, 97–106.
Jamieson BGM, Healy JM (1992) The phylogenetic positionof the tuatara (Sphenodontida, Amniota), as indicated bycladistic analysis of the ultrastructure of spermatozoa.Phil Trans Roy Soc Lond: Biol Sci 335, 207–219.
Jamieson BGM (1995) Evolution of tetrapod spermatozoawith particular reference to amniotes. In Advances inSpermatozoal Phylogeny and Taxonomy (eds Jamieson BG,Ausio J, Justine J-L), Mem Mus National Histoire Naturelle166, 343–358.
Jamieson BGM, Koehler L, Todd B (1995) Spermatozoalultrastructure in three species of parrots (aves, Psittaciformes)and its phylogenetic implications. Anat Rec 241, 461–468.
Jassim A, Gillot DJ, Al-Zuhdi Y (1991) Human sperm tail fibroussheath undergoes phosphorylation during its development.Human Reprod 6, 1135–1142.
Jassim A (1995) Molecular and ontogenic analysis of the humansperm tail fibrous sheath. In Advances in SpermatozoalPhylogeny and Taxonomy (eds Jamieson BG, Ausio J, JustineJ-L), Mem Mus National Histoire Naturelle 166, 431–436.
Jones RC, Lin M (1993) Spermatogenesis in birds. Oxford RevReprod Biol 15, 233–264.
Kirsch JAW, Lapointe FJ, Springer MS (1997) DNA-hybridisationstudies of marsupials and their implications for Metatherianclassification. Aust J Zool 45, 211–280.
Lin M, Jones RC (1993) Spermiogenesis and spermiation in theJapanese quail (Coturnix coturnix japonica). J Anat 183,525–535.
Lin M, Jones RC (2000) Spermiogenesis and spermiation in amonotreme mammal, the platypus, Ornithorhynchusanatimus. J Anat 196, 217–232.
Lin M, Harman A, Fletcher TP (2004) Cycle of the seminiferousepithelium in a marsupial species, the brushtail possum(Trichosurus vulpecula), an estimation of its duration. ReprodFertil Dev 16, 307–313.
Lindemann CB, Orlando A, Kanous KS (1992) The flagellarbeat of rat sperm is organized by the interaction of twofunctionally distinct populations of dynein bridges with astable central axonemal partition. J Cell Sci 102, 249–260.
Miki K, Willis WD, Brown PR, Goulding EH, Fulcher KD,Eddy EM (2002) Targeted disruption of the Akap4 genecauses defects in sperm flagellum and motility. Dev Biol 15,331–342.
Mori C, Welch JE, Sakai Y, Eddy EM (1992) In situ localizationof spermatogenic cell-specific glyceraldehyde 3-phosphatedehydrogenase (Gapd-s) messenger ribonucleic acid in mice.Biol Reprod 46, 859–868.
Nilsson MA, Arnason U, Spencer PSB, Janke A (2004) Marsu-pial relationships and a timeline for marsupial radiation inSouth Gondwana. Gene 340, 189–196.
Oko R (1988) Comparative analysis of proteins from thefibrous sheath and outer dense fibres of rat spermatozoa.Biol Reprod 39, 169–182.
Oko R, Clermont Y (1988) Isolation, structure and proteincomposition of the perforatorium of rat spermatozoa. BiolReprod 39, 673–687.
Oko R, Clermont Y (1989) Light microscopic immunocyto-chemical study of fibrous sheath and outer dense fibreformation in the rat spermatid. Anat Rec 225, 46–55.
Phillips DM (1972) Comparative analysis of mammalian spermmotility. J Cell Biol 53, 561–573.

Morphogenesis of the fibrous sheath in the marsupial spermatozoon, M. Ricci and W. G. Breed
© Anatomical Society of Great Britain and Ireland 2005
164
Ricci M, Breed WG (2001) Isolation and partial characteriza-tion of the outer dense fibres and fibrous sheath from thesperm tail of a marsupial: the brush-tail possum (Trichosurusvulpecula). Reproduction 121, 373–388.
Sakai Y, Koyama YI, Fujimoto H, Nakamoto T, Yamashina S(1986) Immunocytochemical study on fibrous sheath forma-tion in mouse spermiogenesis using a monoclonal antibody.Anat Rec 215, 119–126.
Sapsford CS, Rae CA, Cleland KW (1969) Ultrastructuralstudies on maturing spermatids and on Sertoli cells in thebandicoot Perameles nasuta Geoffroy (Marsupialia). Aust JZool 17, 195–292.
Sapsford CS, Rae CA, Cleland KW (1970) Ultrastructuralstudies on the development and form of the principal piecesheath of the bandicoot spermatozoon. Aust J Zool 18, 21–48.
Scheltinga DM, Jamieson BG, Espinoza RE, Orrell KS (2001)Descriptions of the mature spermatozoa of the lizardsCrotaphytus bicinctores, Gambelia wislizenii (Crotaphytidae),and Anolis carolinensis (Polychrotidae) (Reptilia, Squamata,Iguania). J Morphol 247, 160–171.
Si Y, Okuno M (1993) The sliding of the fibrous sheath throughthe axoneme proximally together with microtubule extru-sion. Exp Cell Res 208, 170–174.
Thurston RJ, Hess RA (1987) Ultrastructure of spermatozoafrom domesticated birds: comparative study of turkey,chicken and guinea fowl. Scanning Microsc 1, 1829–1838.
Vieira GHC, Wiederhecker HC, Colli GR, Bao SN (2001)Spermiogenesis and testicular cycle of the lizard Tropidurustorquatus (Squamata, Tropiduridae) in the Cerrado ofcentral Brazil. Amphibia–Reptilia 22, 217–233.
Vijayaraghavan S, Goueli SA, Davey MP, Carr DW (1997)Protein kinase A-anchoring inhibitor peptides arrest mam-malian sperm motility. J Biol Chem 272, 4747–4475.
Vijayaraghavan S, Liberty G, Mohan J, Winfrey V, Olson G,Carr D (1999) Isolation and molecular characterization ofAKAP110, a novel, sperm-specific protein kinase A-anchoringprotein. Mol Endocrinol 13, 705–717.
Welch JE, Schatte EC, O’Brien DA, Eddy EM (1992) Expressionof a glyceraldehyde 3-phosphate dehydrogenase gene specificto mouse spermatogenic cells. Biol Reprod 46, 869–878.
Welch JE, Brown PR, O’Brien DA, Eddy EM (1995) Genomicorganization of a mouse glyceraldehyde 3-phosphatedehydrogenase gene (Gapd-s) expressed in post-meioticspermatogenic cells. Dev Genet 16, 179–189.
Woodburne MO, Rich TH, Springer MS (2003) The evolution oftribospheny and the antiquity of mammalian clades. MolPhylogenet Evol 28, 360–385.





























