Embed Size (px)
Citation preview

Molecular and cytological characterization ofgenomic variability in hexaploid wheat ‘Lindström’
Pedro Costa-Nunes, Teresa Ribeiro, Margarida Delgado, Leonor Morais-Cecílio,Neil Jones, and Wanda Viegas
Abstract: ‘Lindström’ wheat (AABBDD + rye B chromosomes) was used to study the effects of alien chromatinintrogressed into a wheat genetic background, subjecting the wheat genome to a new and transient allopolyploidisationepisode. Using this experimental material, we have previously demonstrated that no large-scale chromosomaltranslocations occurred as a result of the genomic constitution of the addition line. However, we have shown that thepresence of a number of rye B chromosomes is associated with changes in the interphase organization and expressionpatterns of wheat rDNA loci. We have now extended our studies to focus on a further characterization of ‘Lindström’5S rDNA loci and also on high molecular weight glutenin subunit (HMW-GS) patterns. In the process, we have uncov-ered an unusually large variant of the 5S rDNA locus on wheat chromosome 1B (not to be confused with rye B chro-mosomes) and 2 novel HMW glutenin y-type alleles. These changes are not directly related to variation in rye Bchromosome number in the present material, but the fact that a new, and still segregating, 1Dy HMW-GS gene wasidentified indicates a recent timescale for its origin. Strikingly, the ‘Lindström’ 5S rDNA 1B locus integrates a unitsharing 94% homology with a rye 5S rDNA sequence, suggesting the possibility that the wheat locus was colonized byhighly homologous rye sequences during the breeding of ‘Lindström’, when the rye and wheat genomes were together,albeit briefly, in the same nucleus.
Key words: Triticum aestivum ‘Lindström’, allopolyploidisation, 5S rDNA, NTS, high molecular weight glutenin(HMW-GS).
Résumé : Le blé ‘Lindström’ (AABBDD + chromosomes B du seigle) a été employé pour étudier les effets d’unechromatine étrangère introduite chez le blé, laquelle expose le génome du blé à un épisode passager d’allopolyploïdie.Chez ce matériel expérimental, les auteurs ont démontré antérieurement qu’aucune translocation chromosomique degrande taille n’était survenue malgré la composition génomique de la lignée d’addition. Cependant, il a été montré quela présence de chromosomes B du seigle était associée à des changements dans l’organisation et l’expression de gènesd’ADNr du blé au cours de l’interphase. Ces études ont maintenant été élargies pour toucher les locus d’ADNr 5S etune gluténine de masse moléculaire élevée (HMW-GS) chez le blé ‘Lindström’. Au cours de ce travail, les auteurs ontobservé un variant d’ADNr 5S d’une taille exceptionnellement grande sur le chromosome 1B du blé (à ne pasconfondre avec les chromosomes B du seigle) et 2 nouveaux allèles de gluténines HMW de type y. Ces changementsne sont pas directement liés à la variation du nombre de chromosomes B du seigle au sein du présent matériel. Cepen-dant, le fait qu’un gène 1Dy HMW-GS nouveau, toujours en ségrégation, ait été identifié implique une origine récente.Étonnamment, le locus d’ADNr 5S sur le chromosome 1B de ‘Lindström’ comprend une unité qui est à 94 % identiqueavec l’ADNr 5S du seigle. Cela suggère que le locus du blé aurait été colonisé par des séquences très homologues enprovenance du seigle au cours de la sélection de ‘Lindström’, alors que les génomes du seigle et du blé étaient pré-sents, bien que brièvement, au sein du même noyau.
Mots clés : Triticum aestivum ‘Lindström’, allopolyploïdisation, ADNr 5S, NTS, gluténine de masse moléculaire élevée(HMW-GS)
[Traduit par la Rédaction] Costa-Nunes et al. 904
Genome 48: 895–904 (2005) doi: 10.1139/G05-050 © 2005 NRC Canada
895
Received 7 January 2005. Accepted 7 May 2005. Published on the NRC Research Press Web site at http://genome.nrc.ca on20 October 2005.
Corresponding Editor: J.H. de Jong.
P. Costa-Nunes, T. Ribeiro, M. Delgado, L. Morais-Cecílio,1 and W. Viegas. Secção de Genética, Centro de Botânica Aplicadaà Agricultura, Instituto Superior de Agronomia, Tapada da Ajuda, 1349–017 Lisboa, Portugal.N. Jones. The University of Wales Aberystwyth, Institute of Biological Sciences, Cledwyn Building, Aberystwyth SY23 3DD,Wales, UK.
1Corresponding author (e-mail: [email protected]).

Introduction
The ‘Lindström’ strain of hexaploid wheat was created byLindström (1965) to make an addition line carrying the su-pernumerary B chromosomes of rye in a wheat background.In the spring of 1963, a primitive spring variety of rye fromTransbaikal, Siberia, with B chromosomes (Bs), was crossedwith the Nepal 2 line of a spring wheat from Nepal. The F1hybrids underwent spontaneous chromosome doubling togive a 2n = 56 + 4 Bs Triticale-type, which was self-pollinated. Selected plants with 2n = 56 + 2Bs were thenback-crossed to Nepal 2, and gave rise to some 49 chromo-some progeny (42 wheat + 7 rye) with Bs. On further back-crossing to Nepal 2, two progeny plants with 2n = 42 + Bswere found and formed the ‘Lindström’ strain. The rye Bs in‘Lindström’ wheat retain their autonomous properties andare transmitted in the same way as in rye (Jones and Rees1982), except that they are meiotically less stable in wheatand are maintained through selection for the +B types ineach generation. ‘Lindström’ plants are, therefore, availablewith and without the B chromosomes (Müntzing 1973) andconstitute a useful experimental resource for the study of ge-nome interactions.
Rye Bs have phenotypic effects on ‘Lindström’ wheat,which include a negative impact on fertility, grain qualityand weight (Müntzing 1973), as they do in more recentlyproduced B chromosome addition lines (Korean rye) and inother wheat strains (‘Chinese Spring’) (Niwa et al. 1997). Instudies at a cytological level, we have also demonstrated thatthe rye Bs in ‘Lindström’ cause alterations in rDNA inter-phase organization and expression patterns (Morais-Cecílioet al. 2000); and with this, and the effects on seed proteins inmind, we have used the uniqueness of this experimental ma-terial to characterize the high-molecular-weight glutenin sub-unit (HMW-GS) patterns and the 5S rDNA loci.
Triticum aestivum has 6 genes coding for HMW-GS or-ganised in 3 complex loci located on the long arm ofhomoeologous group 1 chromosomes (Payne 1987; Cabreraet al. 2002). The HMW-GS encoding genes lack introns(Forde et al. 1985; Thompson et al. 1985; Anderson et al.1988) and are susceptible, through duplication or deletion of1 of the repeats in the central repetitive domain, to produc-ing transcripts with different lengths (Forde et al. 1985).Therefore, size variation in HMW-GS is mainly due to thedifferent length of the central repetitive domain of variousHMW glutenin genes. The structure and organization of the5S rRNA multigene family in the Triticeae is well character-ized and is divided into 2 classes, 1 with long repeat unitsand 1 with short repeat units. These 2 size classes havehighly conserved coding regions (120 bp), but have non-transcribed spacers differing in length and nucleotide se-quence (Gerlach and Dyer 1980; Dvorák et al. 1989; Appelset al. 1992; Baum and Appels 1992). The arrays constitutedby long repeat units are assigned to the short arm ofhomoeologous group 5 chromosomes and the short units, aswell as the short arm of homoeologous group 1 chromo-somes (Appels et al. 1980; Dvorák et al. 1989; Reddy andAppels 1989; Mukai et al. 1990).
The form of the analysis we have used, involving plantswith and without B chromosomes, excludes any direct effect
of rye B chromosomes on HMW-GS patterns and 5S rDNAloci structural modifications in the present experiments. Wereport the finding of 2 novel HMW-GS alleles and an unusu-ally large variant of the 5S rDNA 1B locus that integrates arye 5S rDNA sequence, and we interpret the results in termsof modifications induced at the time of formation of the ad-dition line owing to genomic interactions between the wheatand rye chromosomes.
Materials and methods
Plant materialThe analysis was performed on Triticum aestivum (L.)
Thell ‘Lindström’ with and without B chromosomes and‘Chinese Spring’ (CS), and on Secale cereale Thell ‘JNK’and Secale cereale ‘Centeio do Alto’ (CA). Wheat kernelswere cut transversely in such a way that HMW-GS extraction,plant genomic DNA (gDNA) extraction, and cytogenetic anal-ysis of root meristematic cells could all be achieved on eachindividual kernel and (or) seedlings.
Sodium dodecyl sulfate – polyacrylamide gelelectrophoresis (SDS–PAGE) analysis
Single grain flour protein extraction was carried out using amodification of the methods described by Khan et al. (1989)and Gupta and MacRitchie (1991). Non-HMW glutenin pro-teins were sequentially extracted with 100% DMSO (50 µL/mgof flour) and twice with 70% (v/v) ethanol (50 µL/mg offlour). At each step, samples were vortexed for 10 min, cen-trifuged for 5 min at 15 900g, and the supernatant was dis-carded. Finally, HMW-GS were extracted with 1.5% (w/v)sodium dodecyl sulphate (SDS) (4 µL/mg of flour) and 2-mercaptoethanol (0.4 µL/ mg of flour). The sample wasvortexed for 60 min, centrifuged for 2 min at 15 900g (r.t.),and the supernatant was recovered. Electrophoresis was per-formed in the PhastSystem™ (Amersham Pharmacia BiotechInc., Piscataway, New Jersey, USA) in a 10%–15% gradientpolyacrilamide mini-gel.
PCR amplificationGenomic DNA was isolated from 5 g of leaves from sin-
gle plants according to the protocol described by Coen andCarpenter (1986) and was subsequently used as template forPCRs. Polymerase chain reactions for HMW-GS gene analy-sis were performed accordingly to D’Ovidio et al. (1995).The primers used were P1, 5′-ATG GCT AAG CGG TTGGTC CT-3′; P2, 5′-CTG TGT TAA CAT GCT ATG GGTTGT C-3′; P3, 5′-GGG AAC ATC TTC ACA AAA CAGTAC AA-3′; and P4, 5′-GGC TAG CCG ACA ATG CGTCG-3′ (D’Ovidio et al. 1995). Primer binding sites to the1Dy12 HMW-GS gene, determined from published sequence(Halford et al. 1987), are shown in Fig. 1. Polymerase chainreactions for 5S nontranscribed spacers (NTS) analysis wereperformed according to Van Campenhout et al. (1998).Primers PR5S5, TTT TGC GTC TCG TGA CAA G andPR5S3, GTT ATC CCC GTC TTC GCT were used to am-plify the spacer DNA region between repeated 5S rRNAgenes of chromosome 1R (5SrRNA-R1). Primers PW5S1B5,AAG TCC TCG TGT TGC ATT CC and PW5S1B3, CATTGC TTT GGC CGC TCC CT were used to amplify the
© 2005 NRC Canada
896 Genome Vol. 48, 2005

spacer between 5S rRNA genes of chromosome 1B (5SrRNA-B1) and 21 nucleotides of the gene.
Southern blot analysisAmplified products were fractioned on 1.5% agarose gel
and alkaline DNA transfer to a Hybond N+ membrane. ThepTag1290 clone, containing a 1.5-kb PstI fragment of thecentral repetitive domain of the HMW-GS genes, was usedas a probe (Thompson et al. 1983). Once isolated from theplasmid vector, the 1.5-kb fragment was purified using theUltrafree®-DA purification kit (Millipore Corp., Billerica,Mass.). Probe labelling, prehybridization, hybridization anddetection reactions were carried out as indicated by the manu-facturer of the ECL-direct nucleic acid labelling and detectionsystems (Amersham Pharmacia Biotech Inc., Piscataway, NewJersey, USA).
Cloning and sequencing reactions‘Lindström’ PCR product amplified with primer combina-
tion PR5S5/PR5S3 was cloned into the pCR2.1-TOPO® vec-tor from the TOPO TA Cloning® kit (Invitrogen Ltd.,Paisley, UK), according to the manufacturer’s instructions.Cloned plasmid DNA was isolated using a Magic MiniprepsDNA Purification System kit (Promega, Southampton, UK).The insert was subsequently sequenced using M13 primers.Polymerase chain reaction products from primer combina-tions PR5S5/PR5S3 with rye gDNA template (control) andPW5S1B5/PW5S1B3 with ‘Lindström’ gDNA template weredirectly sequenced. A computer-assisted sequence analysiswas performed using NCBI-BLAST2 (http://www.ebi.ac.uk/blastall/) and CLUSTAL W (http://www.ebi.ac.uk/clustalw/)Internet tools.
Slide preparation for in situ hybridizationSeeds of ‘Lindström’ wheat with and without B chromo-
somes, and of rye ‘JNK’ without B chromosomes, were ger-minated on moist filter paper at 24 °C until their rootsreached a length of approximately 1 cm. They were thentreated to enhance the mitotic index (Delgado et al. 1995).Root tips from each seedling were excised, immersed in ice-cold water for 24 h to induce c-metaphases, and then fixedin 3:1 (v/v) absolute alcohol : glacial acetic acid. Cells fromthe meristematic root tip apex were spread following theprocedure of Schwarzacher and Heslop-Harrison (2000).
In situ hybridizationIn situ hybridization, detection procedures, and the ampli-
fication of hybridization signals were performed according
to Schwarzacher and Heslop-Harrison (2000). The DNAprobes used throughout this work were pTa794, a 410-bp 5SrDNA unit isolated from Triticum aestivum (Gerlach andDyer 1980); NTS5SR1, a 324-bp amplification product fromthe 5S rDNA short repeat unit (460 bp) of Secale cereale,isolated during this study; NTS5SB1, a 251-bp amplificationproduct from the 5S rDNA short repeat unit (420 bp) ofTriticum aestivum, also isolated during this study; andpTa71, a 9-kb EcoR1 fragment, which includes 18S–5.8S–25S rRNA genes and intergenic spacers isolated from Trit-icum aestivum (Gerlach and Bedbrook 1979). The pTa794,NTS5SR1 and NTS5SB1 probes, were PCR labelled with ei-ther digoxigenin-11-dUTP or biotin-11-dUTP. Detection re-actions of digoxigenin or biotin were performed by wereperformed by fluoresceinated (FITC) anti-digoxigenin or bystreptavidin-Cy3 conjugates, respectively. The pTa71 probewas labelled with biotin-11-dUTP by nick translation. DNAwas counterstained with 4′,6-diamidino-2-phenylindole.
Preparations were analysed with an epifluorescence mi-croscope (Zeiss Axioscop2, Zeiss, Jena, Germany) with theappropriate filters. Microphotography was performed usingthe Carl Zeiss Axiocam digital camera and Axiovision soft-ware (version 3.0.6.38). Fluorescent images were separatelycaptured and merged with Photoshop® (Adobe® SystemsInc., San Jose, California, USA).
Results
Variability in HMW-GS patternsThe analysis of HMW-GS in 39 grains of ‘Lindström’ wheat,
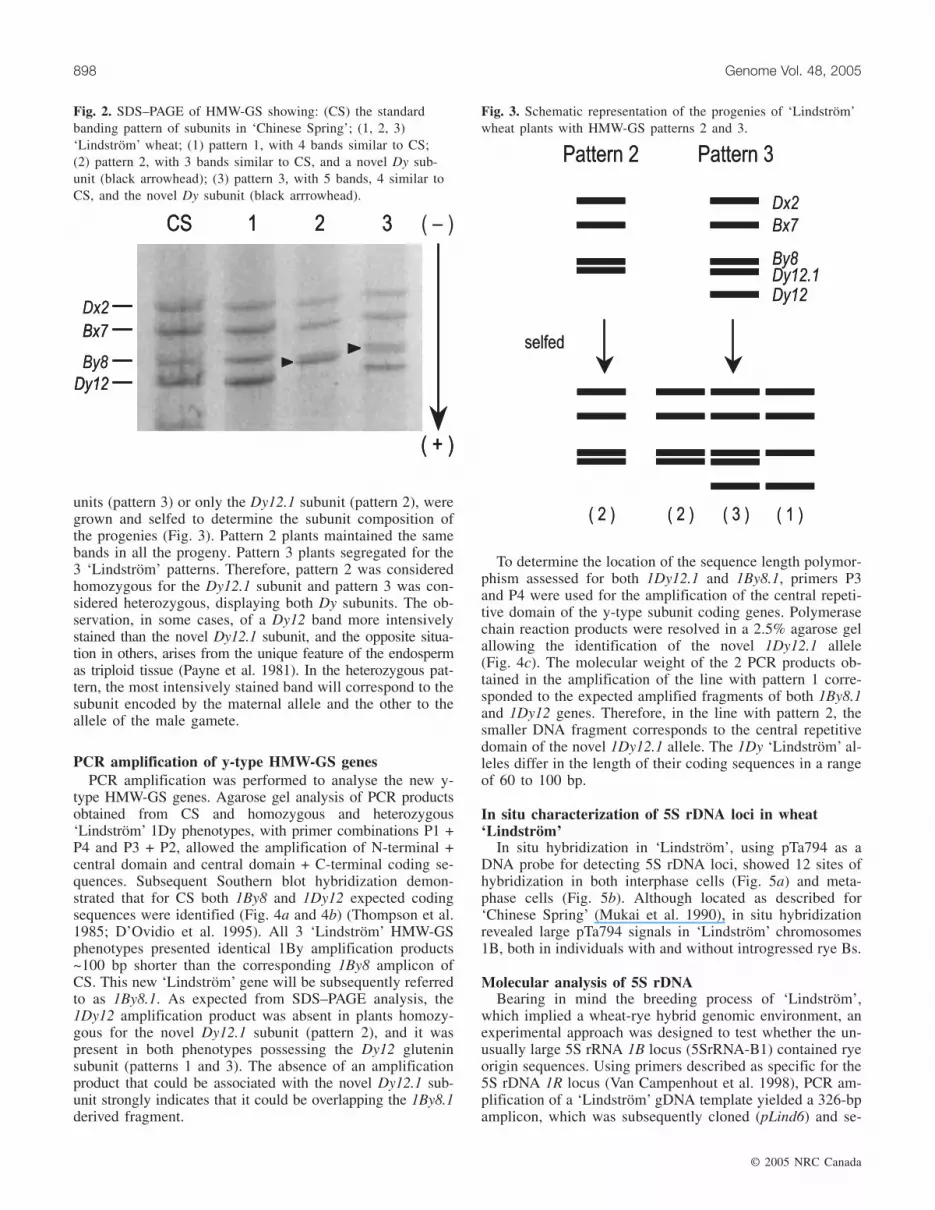
by one-dimensional sodium dodecyl sulfate – polyacrylamidegel electrophoresis (SDS–PAGE), showed 3 patterns: 2 with4 HMW-GS and 1 with 5. All 3 patterns displayed bandswith identical mobility for subunits Dx2, Bx7, and By8 ofTriticum aestivum ‘Chinese Spring’ (CS) (Fig. 2). Other thanthe common subunits, pattern 1 presents the Dy12 subunit,pattern 2 is characterized by the presence of a novel subunitdesignated as Dy12.1, and pattern 3 presents both Dy12 andDy12.1. Pattern 3 is also characterized by quantitative varia-tion of the Dy subunits, assessed by staining intensity. Insome cases, Dy12 is more intensively stained than Dy12.1(Fig. 2; lane 3) and in others the reverse situation is ob-served (not shown).
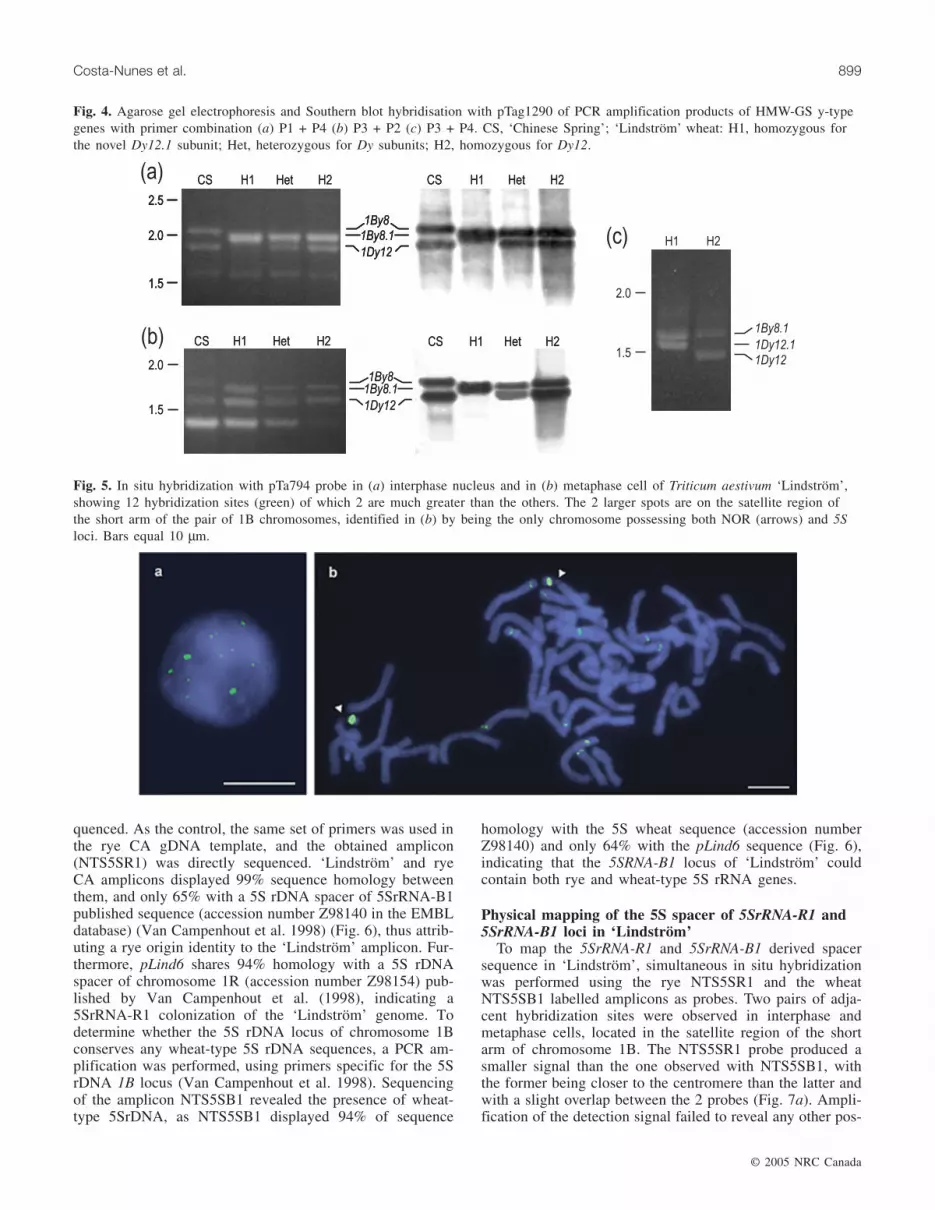
Segregation of HMW-GS patternsTo study how the transmission of the ‘Lindström’ HMW-
GS patterns differed from CS a random sample of half-grains with viable embryos, typed for carrying both Dy sub-
© 2005 NRC Canada
Costa-Nunes et al. 897
Fig. 1. Schematic representation of the 1Dy12 HMW-GS gene of ‘Chinese Spring’. P1, P2, P3, and P4 represent primer annealingsites.
units (pattern 3) or only the Dy12.1 subunit (pattern 2), weregrown and selfed to determine the subunit composition ofthe progenies (Fig. 3). Pattern 2 plants maintained the samebands in all the progeny. Pattern 3 plants segregated for the3 ‘Lindström’ patterns. Therefore, pattern 2 was consideredhomozygous for the Dy12.1 subunit and pattern 3 was con-sidered heterozygous, displaying both Dy subunits. The ob-servation, in some cases, of a Dy12 band more intensivelystained than the novel Dy12.1 subunit, and the opposite situa-tion in others, arises from the unique feature of the endospermas triploid tissue (Payne et al. 1981). In the heterozygous pat-tern, the most intensively stained band will correspond to thesubunit encoded by the maternal allele and the other to theallele of the male gamete.
PCR amplification of y-type HMW-GS genesPCR amplification was performed to analyse the new y-
type HMW-GS genes. Agarose gel analysis of PCR productsobtained from CS and homozygous and heterozygous‘Lindström’ 1Dy phenotypes, with primer combinations P1 +P4 and P3 + P2, allowed the amplification of N-terminal +central domain and central domain + C-terminal coding se-quences. Subsequent Southern blot hybridization demon-strated that for CS both 1By8 and 1Dy12 expected codingsequences were identified (Fig. 4a and 4b) (Thompson et al.1985; D’Ovidio et al. 1995). All 3 ‘Lindström’ HMW-GSphenotypes presented identical 1By amplification products~100 bp shorter than the corresponding 1By8 amplicon ofCS. This new ‘Lindström’ gene will be subsequently referredto as 1By8.1. As expected from SDS–PAGE analysis, the1Dy12 amplification product was absent in plants homozy-gous for the novel Dy12.1 subunit (pattern 2), and it waspresent in both phenotypes possessing the Dy12 gluteninsubunit (patterns 1 and 3). The absence of an amplificationproduct that could be associated with the novel Dy12.1 sub-unit strongly indicates that it could be overlapping the 1By8.1derived fragment.
To determine the location of the sequence length polymor-phism assessed for both 1Dy12.1 and 1By8.1, primers P3and P4 were used for the amplification of the central repeti-tive domain of the y-type subunit coding genes. Polymerasechain reaction products were resolved in a 2.5% agarose gelallowing the identification of the novel 1Dy12.1 allele(Fig. 4c). The molecular weight of the 2 PCR products ob-tained in the amplification of the line with pattern 1 corre-sponded to the expected amplified fragments of both 1By8.1and 1Dy12 genes. Therefore, in the line with pattern 2, thesmaller DNA fragment corresponds to the central repetitivedomain of the novel 1Dy12.1 allele. The 1Dy ‘Lindström’ al-leles differ in the length of their coding sequences in a rangeof 60 to 100 bp.
In situ characterization of 5S rDNA loci in wheat‘Lindström’
In situ hybridization in ‘Lindström’, using pTa794 as aDNA probe for detecting 5S rDNA loci, showed 12 sites ofhybridization in both interphase cells (Fig. 5a) and meta-phase cells (Fig. 5b). Although located as described for‘Chinese Spring’ (Mukai et al. 1990), in situ hybridizationrevealed large pTa794 signals in ‘Lindström’ chromosomes1B, both in individuals with and without introgressed rye Bs.
Molecular analysis of 5S rDNABearing in mind the breeding process of ‘Lindström’,
which implied a wheat-rye hybrid genomic environment, anexperimental approach was designed to test whether the un-usually large 5S rRNA 1B locus (5SrRNA-B1) contained ryeorigin sequences. Using primers described as specific for the5S rDNA 1R locus (Van Campenhout et al. 1998), PCR am-plification of a ‘Lindström’ gDNA template yielded a 326-bpamplicon, which was subsequently cloned (pLind6) and se-
© 2005 NRC Canada
898 Genome Vol. 48, 2005
Fig. 2. SDS–PAGE of HMW-GS showing: (CS) the standardbanding pattern of subunits in ‘Chinese Spring’; (1, 2, 3)‘Lindström’ wheat; (1) pattern 1, with 4 bands similar to CS;(2) pattern 2, with 3 bands similar to CS, and a novel Dy sub-unit (black arrowhead); (3) pattern 3, with 5 bands, 4 similar toCS, and the novel Dy subunit (black arrrowhead).
Fig. 3. Schematic representation of the progenies of ‘Lindström’wheat plants with HMW-GS patterns 2 and 3.
quenced. As the control, the same set of primers was used inthe rye CA gDNA template, and the obtained amplicon(NTS5SR1) was directly sequenced. ‘Lindström’ and ryeCA amplicons displayed 99% sequence homology betweenthem, and only 65% with a 5S rDNA spacer of 5SrRNA-B1published sequence (accession number Z98140 in the EMBLdatabase) (Van Campenhout et al. 1998) (Fig. 6), thus attrib-uting a rye origin identity to the ‘Lindström’ amplicon. Fur-thermore, pLind6 shares 94% homology with a 5S rDNAspacer of chromosome 1R (accession number Z98154) pub-lished by Van Campenhout et al. (1998), indicating a5SrRNA-R1 colonization of the ‘Lindström’ genome. Todetermine whether the 5S rDNA locus of chromosome 1Bconserves any wheat-type 5S rDNA sequences, a PCR am-plification was performed, using primers specific for the 5SrDNA 1B locus (Van Campenhout et al. 1998). Sequencingof the amplicon NTS5SB1 revealed the presence of wheat-type 5SrDNA, as NTS5SB1 displayed 94% of sequence
homology with the 5S wheat sequence (accession numberZ98140) and only 64% with the pLind6 sequence (Fig. 6),indicating that the 5SRNA-B1 locus of ‘Lindström’ couldcontain both rye and wheat-type 5S rRNA genes.
Physical mapping of the 5S spacer of 5SrRNA-R1 and5SrRNA-B1 loci in ‘Lindström’
To map the 5SrRNA-R1 and 5SrRNA-B1 derived spacersequence in ‘Lindström’, simultaneous in situ hybridizationwas performed using the rye NTS5SR1 and the wheatNTS5SB1 labelled amplicons as probes. Two pairs of adja-cent hybridization sites were observed in interphase andmetaphase cells, located in the satellite region of the shortarm of chromosome 1B. The NTS5SR1 probe produced asmaller signal than the one observed with NTS5SB1, withthe former being closer to the centromere than the latter andwith a slight overlap between the 2 probes (Fig. 7a). Ampli-fication of the detection signal failed to reveal any other pos-
© 2005 NRC Canada
Costa-Nunes et al. 899
Fig. 4. Agarose gel electrophoresis and Southern blot hybridisation with pTag1290 of PCR amplification products of HMW-GS y-typegenes with primer combination (a) P1 + P4 (b) P3 + P2 (c) P3 + P4. CS, ‘Chinese Spring’; ‘Lindström’ wheat: H1, homozygous forthe novel Dy12.1 subunit; Het, heterozygous for Dy subunits; H2, homozygous for Dy12.
Fig. 5. In situ hybridization with pTa794 probe in (a) interphase nucleus and in (b) metaphase cell of Triticum aestivum ‘Lindström’,showing 12 hybridization sites (green) of which 2 are much greater than the others. The 2 larger spots are on the satellite region ofthe short arm of the pair of 1B chromosomes, identified in (b) by being the only chromosome possessing both NOR (arrows) and 5Sloci. Bars equal 10 µm.

sible sites of hybridization (not shown). The sum of signalsfrom NTS5SB1 and NTS5SR1 probes is smaller than theone from the pTa794 probe in the 1B locus. This could bedue to the length of the probes: they are smaller than thepTa794 and the amount of fluorescence is less intense. Inthe ‘JNK’ rye, the NTS5SR1 probe hybridized not only tochromosomes 1R, identified in the picture by having the sec-ondary constriction (NOR-region) labelled with pTa71, butalso to chromosomes 5R (Fig. 7b). This nonspecific hybrid-ization could be due to the high level of homology between5S rDNA sequences from chromosomes 1R and 5R. No hy-bridization signal was observed with the NTS5SB1 probe.
Discussion
In the breeding of ‘Lindström’, the wheat + rye genomesand the rye Bs shared the same nucleus in 2 Triticale-typehybrid generations and also in 2 backcrosses to wheat whenthe rye A chromosomes were being eliminated. The presentresults suggest that it was during this transient allopoly-ploidisation time frame that a genome restructuring eventtook place that produced the sequence modifications to the5SrRNA-B1 and Glu-1D loci.
The analysis of HMW-GS patterns in ‘Lindström’ re-vealed variability associated with allelic variation of the 1Dygene; but we were unable to determine whether the new al-lele we found was already present in the parental Nepalesespring wheat. However, an extensive screening of the HMW-GS content of 57 hexaploid wheat accessions from Nepal(Margiotta et al. 1988) failed to identify 1Dy12.1 among the1Dy alleles of Nepalese populations, and the fact that segre-gation at the Glu-1D locus is still occurring suggests a re-cent timescale for the event. PCR and subsequent Southernblot analysis indicates that the variability found in theHMW-GS is due to a novel 1Dy12.1 allele arising from genealteration, possibly by nucleotide sequence addition or dupli-cation of tandem repeats in the repetitive central domain ofthe coding region of the 1Dy12-like gene of ‘Lindström’.Overlapping amplicons of the 1By8.1 and 1Dy12.1 geneswere observed, and a constant difference in sequence lengthof about 100 bp was detected when compared with the CS1Dy12 fragment. Together, these results locate the polymor-phism between 1Dy12 and 1Dy12.1 to the central repetitivedomain coding sequence. When overlapped bands were re-solved in a 2.5% agarose gel, the novel 1Dy12.1 central do-main amplicon was clearly visualized. A similar case of
© 2005 NRC Canada
900 Genome Vol. 48, 2005
Fig. 6a. CLUSTAL W (1.82) multiple sequence alignment and homology scores between 5S rDNA spacer sequences of the cloned‘Lindström’ amplicon (pLind6) and 2 sequences of rye: ‘Centeio do Alto’ amplicon (NTS5SR1) and a published 5S spacer sequencefrom 1R locus (Z98154).

length polymorphism has been reported in relation to the1Dx2 allele of ‘Chinese Spring’ and the 1Dx2.2* allele ofwheat accession MG315 (D’Ovidio et al. 1995). These al-leles differ by the insertion of 561 bp in the latter. The inser-tion is homologous to, and originates from, a 1Dx2-like genein the ancestral wheat of accession MG315. Sequence dupli-cation has been attributed to unequal crossing-over events,which is the mechanism proposed for the origin of HMW-GS variability owing to the repetitive nature of the centraldomain of these genes (Forde et al. 1985).
In relation to the By8.1 HMW-GS found in ‘Lindström’,our results strongly indicate that, although no differenceswere observed by SDS–PAGE analysis between the ByHMW-GS of ‘Chinese Spring’ and ‘Lindström’, in realitythe ‘Lindström’ subunit is encoded by an approximately100-bp smaller 1By gene. As for the novel 1Dy gene codingsequence, diversity of 1By8.1 is localized in the central re-petitive domain, as the results show. A strong relationshiphas been established between the length of the central repeti-tive domain of HMW-GS genes and the molecular weight of
the encoded subunit (D’Ovidio et al. 1995). Nevertheless, themobility of HMW-GS in SDS–PAGE is not always correlatedwith their actual molecular weight owing to protein confor-mation (Goldsbrough et al. 1989; D’Ovidio et al. 1995). Itcan be concluded that the By8.1 subunit of ‘Lindström’, al-though displaying an identical mobility to the ‘ChineseSpring’ By8 subunit, is in fact a different HMW-GS as it dif-fers in the length of its coding sequence from the 1By8 gene.
The ‘Lindström’ cultivar possesses the same number of 5SrRNA loci as described for ‘Chinese Spring’ (Mukai et al.1990), although ‘Lindström’ shows a much larger 5S rRNAcluster on chromosome 1B. This hybridization pattern wasobserved in all the analysed ‘Lindström’ individuals and indifferent generations, whether or not they carried any rye Bchromosomes. Furthermore, isolation of a rye 5SrRNA-R1nontranscribed spacer from the ‘Lindström’ genome and itsmapping to the 5SrRNA-B1 locus suggests that a rearrange-ment has occurred in ‘Lindström’ through a stable intro-gression, probably early in the development of this cultivar,and has been maintained in the following generations. (It is
© 2005 NRC Canada
Costa-Nunes et al. 901
Fig. 6b. CLUSTAL W (1.82) multiple sequence alignment and homology scores between 5S rDNA spacer sequences of the cloned‘Lindström’ amplicon (pLind6) and 2 sequences of wheat: ‘Lindström’ amplicon from 1B locus (NTS5SB1) and a published 5S spacersequence from 1B locus (Z98140). The area of homology between sequences is evidenced by (*) and highlighted in grey betweenNTS5SB1 and Z98140 sequences.

not known how many generations ‘Lindström’ has beenthrough, since it almost certainly has not been grown in ev-ery year since 1965.) Ruling out the unlikely hypothesis ofany previous wheat-rye hybrid state of the parental Nepalesewheat, the 5S rye sequence introgression could only haveoccurred in one of the generations in which wheat and ryegenomes shared the same nucleus. 5S rRNA gene familiesare compartmentalised in specific loci (Gerlach and Dyer1980; Dvorák et al. 1989; Appels et al. 1992; Baum andAppels 1992), but thus far there is no evidence for concertedevolution by interlocus interaction among 5S arrays in plants(Cox et al. 1992; Sastri et al. 1992; Kellogg and Appels1995), only of intralocus concerted evolution (Cronn et al.1996; Fulnecek et al. 2002). Therefore, a modification in 5SrRNA gene arrays in the wheat 1B locus could be main-tained and perpetuated by intralocus homogenization mecha-nisms. The unusually large size of this locus, observed inboth interphase and metaphase cells, could be interpreted asan intermediate step in intralocus homogenization both atthe level of 5S nontranscribed spacer sequence polymor-phism and gene copy number. It is known that 5S rDNA se-quences are susceptible to amplification or deletion becauseof their highly repetitive nature. In Plantago lagopus, aheterochromatic B chromosome arose essentially from mas-sive amplification of 5S rDNA sequences (Dhar et al. 2002).Furthermore, total copy number of 5S rRNA genes in a ge-nome is maintained close to a constant level through a pro-cess that compensates loss of 1 class of repeats throughincreasing the copy number in a distinct 5S rRNA locus.The mechanism of unequal crossing-over is believed to beresponsible for these large-scale locus modifications (Röderet al. 1992; Kanazin et al. 1993; Fulnecek et al. 2002) andcould have played a determinant role in the ongoing homog-enization of the acquired 5SrRNA-B1 polymorphism inwheat.
During the development of the ‘Lindström’ strain, back-crossing the spontaneous Triticale type to the wheat parentproduced progeny with an unbalanced chromosome comple-ment (42 wheat + 7 rye). The effect of such imbalance involv-ing wheat and rye genome interaction has been previously
reported. While developing disomic addition lines of ryechromosomes to wheat, Lukaszewski (1995) observed re-verse tandem duplications and chromosomal translocationsas a result of chromosome-type breakage-fusion-bridge cy-cles induced by rye monosomics. Genomic in situ hybridiza-tion (GISH) with rye gDNA as a probe never revealed anyrye segments in the wheat genome of ‘Lindström’, confirm-ing previous results (Morais-Cecílio et al. 1996) and rulingout a major translocation event between chromosomes 1Rand 1B. The rye 5S NTS insertion in 5SrDNA-B1 is un-likely to be detectable by this cytogenetical approach be-cause of high homology of wheat and rye 5S codingsequences and use of blocking gDNA of wheat in an up-to-30-fold proportion to the rye gDNA probe. Furthermore,double in situ hybridization mapped rye derived 5S se-quences closer to the centromere than for native wheat 1B5S rRNA genes, indicating a localized translocation. Al-though chromosome pairing should have been restricted tohomologous wheat chromosomes, some interactions betweenwheat and rye chromosomes could have occurred because ofthe haploid state of the latter, resulting in the insertion of ryeorigin 5S nontranscribed spacer sequences in the 5SrDNA-B1 locus via homeologous pairing of 1B and 1R chromo-somes.
In retrospect, we should not be entirely surprised by thesefindings when we remind ourselves that the idea of “revolu-tionary changes in genome concepts”, by way of “genomeshock” due to hybridity, or several other classes of unexpectedevents for the genomes, was summarized by McClintockmore than 2 decades ago (McClintock 1984). In her review,McClintock pointed out a number of examples of chromo-some restructuring involving allopolyploids, including Triti-cale in which a plethora of instabilities continued to occurover several generations after the initial allopolyploids wereproduced. Although breakage of wheat chromosomes ap-pears to be well known in several situations where rye chro-mosomes are added to the wheat genome (Lukaszewski1995), there are no reports of any chromosome instabilitiesin the wheat genome because of the presence of B chromo-somes of rye in the ‘Lindström’ strain, despite the fact that
© 2005 NRC Canada
902 Genome Vol. 48, 2005
Fig. 7. Fluorescent in situ hybridization in (a) metaphase cell and 1B chromosome (inset) of Triticum aestivum ‘Lindström’ withNTS5SR1 (green) and NTS5SB1 (red) probes and in (b) metaphase cell of Secale cereale ‘JNK’ with NTS5SR1 (green) and pTa71(red) probes. The NOR is identified in (a) by an arrow in the inset and in (b) by the red signal of the pTa71 probe. Bars equal 10 µm.

several hundreds of plants of this material have been studiedcytologically (Müntzing et al. 1969). One reason for thismay be that the B chromosomes are genetically less activethan chromosomes of the normal rye genome, and, as wehave shown here, the changes maybe more cryptic and lesseasy to detect by standard cytogenetical analysis.
Although a rare event, the “gain” of novel DNA fragmentsand of different transcripts can be a consequence of allo-polyploid synthesis (Comai et al. 2000; Ozkan et al. 2001).The characterization of 5S rDNA multicopy loci and HMW-GS single copy genes in ‘Lindström’, highly homologous incoding sequence between wheat and rye genomes, unveiledmarked alterations at both loci. Our results strongly suggestthe observed polymorphisms are yet another example ofgenomic restructuring due to allopolyploidisation events, al-beit a cryptic one.
Acknowledgements
We thank Augusta Barão (Instituto Superior de Agronomia)for technical support, and Prof. Mike D. Gale (John InnesCentre) for providing the pTag1290 probe. We also thankXana Castilho for valuable and pleasant discussions. Thiswork was funded by the Portuguese Fundação para a Ciênciae Tecnologia grants PRAXIS XXI/BM/ 20747/99 and pro-ject POCTI/34000/AGR/2000.
References
Anderson, O.D., Halford, N.G., Forde, J., Yip, R.E., Shewry, P.R.,and Greene, F.C. 1988. Structure and analysis of high molecularweight glutenin genes from Triticum aestivum L. cv Cheyenne.Proceedings of the 7th International Wheat Genetics Sympo-sium, Cambridge, U.K., 13–19 July 1988. Edited by T.E. Millerand R.M.D. Koebner. Institute of Plant Science Research, Cam-bridge, U.K. pp. 669–704.
Appels, R., Gerlach, W.L., Dennis, E.S., Swift, H., and Peacock,W.J. 1980. Molecular and chromosomal organization of DNAsequences coding for the ribosomal RNAs in cereals. Chromo-soma (Berlin), 78: 293–311.
Appels, R., Baum, B.R., and Clarke, B.C. 1992. The 5S DNA unitsof bread wheat (Triticum aestivum). Plant Syst. Evol. 183: 183–194.
Baum, B.R., and Appels, R. 1992. Evolutionary change at the 5SDNA locus of species in the Triticeae. Plant Syst. Evol. 183:195–208.
Cabrera, A., Martin, A., and Barro, F. 2002. In situ comparativemapping (ISCM) of Glu-1 loci in Triticum and Hordeum.Chrom. Res. 10: 49–54.
Coen, E.S., and Carpenter, R. 1986. Transposable elements in Anthir-rinum majus: generations of genetic diversity. Trends Genet. 2:292–296.
Comai, L., Tyagi, A.P., Winter, K., Holmes-Davis, R., Reynolds,S.H., Stevens, Y., and Byers, B. 2000. Phenotypic instability andrapid gene silencing in newly formed Arabidopsis allo-tetraploids. Plant Cell, 12: 1551–1567.
Cox, A.V., Bennett, M.D., and Dyer, T.A. 1992. Use of the polymer-ase chain reaction to detect spacer size heterogeneity in plant 5S-rRNA gene clusters and to locate such clusters in wheat (Triticumaestivum L.). Theor. Appl. Genet. 83: 684–690.
Cronn, R.C., Zhao, X., Paterson, A.H., and Wendel, J.F. 1996.Polymorphism and concerted evolution in a tandemly repeated
gene family: 5S ribosomal DNA in diploid and allopolyploidcottons. J. Mol. Evol. 42: 685–705.
Delgado, M., Morais-Cecílio, L., Neves, N., Jones, R.N., andViegas, W. 1995. The influence of B chromosomes on rDNA or-ganization in rye interphase nuclei. Chrom. Res. 3: 487–491.
Dhar, M.K., Friebe, B., Koul, A.K., and Gill, B.S. 2002. Origin ofan apparent B chromosome by mutation, chromosome fragmen-tation and specific DNA sequence amplification. Chromosoma(Berlin), 111: 332–340.
D’Ovidio, R., Masci, S., and Porceddu, E. 1995. Development ofset of oligonucleotide primers specific for genes at the Glu-1complex loci of wheat. Theor. Appl. Genet. 91: 189–194.
Dvorák, J., Zhang, H.-B., Kota, R.S., and Lassner, M. 1989. Orga-nization and evolution of the 5S ribosomal RNA gene family inwheat and related species. Genome, 32: 1003–1016.
Forde, J., Malpica, J.M., Halford, N.G., Shewry, P.R., Anderson,O.D., Greene, F.C., and Miflin, B.J. 1985. The nucleotide se-quence of a HMW glutenin subunit gene located on chromo-some 1A of wheat (Triticum aestivum L.). Nucleic Acids Res.13: 6817–6832.
Fulnecek, J., Lim, K.Y., Leitch, A.R., Kovarík, A., and Matyásek,R. 2002. Evolution and structure of 5S rDNA loci in allotetra-ploid Nicotiana tabacum and its putative parental species. He-redity, 88: 19–25.
Gerlach, W.L., and Bedbrook, J.R. 1979. Cloning and characteriza-tion of ribosomal RNA genes from wheat and barley. NucleicAcids Res. 7: 1869–1885.
Gerlach, W.L., and Dyer, T.A. 1980. Sequence organization of therepeating units in the nucleus of wheat which contain 5S rRNAgenes. Nucleic Acids Res. 8: 4851–4865.
Goldsbrough, A.P., Bulleid, N.J., Freedman, R.B., and Flavell, R.B.1989. Conformational differences between two wheat (Triticumaestivum) high molecular weight glutenin subunits are due to ashort region containing six amino acid differences. Biochem. J.263: 837–842.
Gupta, R.B., and MacRitchie, F. 1991. A rapid one-step one-dimensional SDS–PAGE procedure for analysis of subunit com-position of glutenin in wheat. J. Cereal Sci. 14: 105–109.
Halford, N.G., Forde, J., Anderson, O.D., Greene, F.C., andShewry, P.R. 1987. The nucleotide and deduced amino acid se-quences of an HMW glutenin subunit gene from chromosome1B of bread wheat (Triticum aestivum L.) and comparison withthose of genes from chromosomes 1A and 1D. Theor. Appl.Genet. 75: 117–126.
Jones, R.N., and Rees, H. 1982. B Chromosomes. Academic Press,London, U.K.
Kanazin, V., Ananiev, E., and Blake, T. 1993. The genetics of 5SrRNA encoding multigene families in barley. Genome, 36:1023–1028.
Kellogg, E.A., and Appels, R. 1995. Intraspecific and interspecificvariation in 5S RNA genes are decoupled in diploid wheat rela-tives. Genetics, 140: 325–343.
Khan, K., Tamminga, G., and Lukow, O. 1989. The effect of wheatflour proteins on mixing and baking — correlation with proteinfractions and high molecular weight glutenin subunit composi-tion by gel electrophoresis. Cereal Chem. 66: 391–396.
Lindström, J. 1965. Transfer to wheat of accessory chromosomesfrom rye. Hereditas, 54: 149–155.
Lukaszewski, A.J. 1995. Chromatid and chromosome type break-age-fusion-bridge cycles in wheat (Triticum aestivum L.). Ge-netics, 140: 1069–1085.
Margiotta, B., Lafiandra, D., Tomassini, C., Perrino, P., andPorceddu, E. 1988. Variation in high molecular weight glutenin
© 2005 NRC Canada
Costa-Nunes et al. 903

© 2005 NRC Canada
904 Genome Vol. 48, 2005
subunits in a hexaploid wheat collection from Nepal. Proceedingsof the 7th International Wheat Genetics Symposium, Cambridge,U.K., 13–19 July 1988. Edited by T.E. Miller and R.M.D.Koebner. Institute of Plant Science Research, Cambridge, U.K.pp. 975–980.
McClintock, B. 1984. The significance of responses of the genometo challenge. Science (Washington, D.C.), 226: 792–801.
Morais-Cecílio, L., Delgado, M., Jones, R.N., and Viegas, W. 1996.Painting rye B chromosomes in wheat: interphase chromatin or-ganization, nucleolar disposition and association in plants withtwo, three or four Bs. Chrom. Res. 4: 195–200.
Morais-Cecílio, L., Delgado, M., Jones, R.N., and Viegas, W.2000. Modifications of wheat rDNA loci by B chromosomes: achromatin organization model. Chrom. Res. 8: 341–351.
Mukai, Y., Endo, T.R., and Gill, B.S. 1990. Physical mapping ofthe 5S rRNA multigene family in common wheat. J. Heredity,81: 290–295.
Müntzing, A. 1973. Effects of accessory chromosomes of rye inthe gene environment of hexaploid wheat. Hereditas, 74: 41–56.
Müntzing, A., Jaworska, H., and Carlbom, C. 1969. Studies onmeiosis in the Lindström strain of wheat carrying accessorychromosomes of rye. Hereditas, 61: 179–207.
Niwa, K., Horiuchi, G., and Hirai, Y. 1997. Production and charac-terization of common wheat with B chromosomes of rye fromKorea. Hereditas, 126: 139–146.
Ozkan, H., Levy, A.A., and Feldman, M. 2001. Allopolyploidy-induced rapid genome evolution in the wheat (Aegilops-Triticum) group. Plant Cell, 13: 1735–1747.
Payne, P.I. 1987. Genetics of wheat storage proteins and the effect
of allelic variation on bread-making quality. Ann. Rev. PlantPhysiol. 38: 141–153.
Payne, P.I., Holt, L.M., and Law, C.N. 1981. Structural and geneticalstudies on the high-molecular-weight subunits of wheat glutenin.Theor. Appl. Genet. 60: 229–236.
Reddy, P., and Appels, R. 1989. A second locus for the 5Smultigene family in Secale L.: sequence divergence in two lin-eages of the family. Genome, 32: 456–467.
Röder, M.S., Sorrells, M.E., and Tanksley, S.D. 1992. 5S ribosomalgene clusters in wheat: pulsed field gel electrophoresis reveals ahigh degree of polymorphism. Mol. Gen. Genet. 232: 215–220.
Sastri, D.C., Hilu, K., Appels, R., Lagudah, E.S., Playford, J., andBaum, B.R. 1992. An overview of evolution in plant 5S DNA.Plant Syst. Evol. 183: 169–181.
Schwarzacher, T., and Heslop-Harrison, J.S. 2000. Practical in situhybridization. Oxford: BIOS Scientific Publishers, Oxford, U.K.
Thompson, R.D., Bartels, D., Harberd, N.P., and Flavell, R.B.1983. Characterisation of the multigene family coding for HMWglutenin subunits in wheat using cDNA clones. Theor. Appl.Genet. 67: 87–96.
Thompson, R.D., Bartels, D., and Harberd, N.P. 1985. Nucleotidesequence of a gene from chromosome 1D of wheat encoding aHMW-glutenin subunit. Nucleic Acids Res. 13: 6833–6846.Van Campenhout, S., Aert, R., and Volckaert, G. 1998.
Orthologous DNA sequence variation among 5S ribosomalRNA gene spacer sequences on homoeologous chromo-somes 1B, 1D, and 1R of wheat and rye. Genome, 41(2):244–255.
















![A genetic linkage map of sweetpotato [Ipomoea batatas (L.) Lam.] based on AFLP markers](https://img.dokumen.tips/doc/110x75/63330e96b6829c19b80c3d31/a-genetic-linkage-map-of-sweetpotato-ipomoea-batatas-l-lam-based-on-aflp-markers.jpg)












