Embed Size (px)
Citation preview

www.elsevier.com/locate/brainres
Brain Research 1047
Research report
Mode of leptin action in chicken hypothalamus
Sami Dridi*, Quirine Swennen, Eddy Decuypere, Johan Buyse
Laboratory of Physiology and Immunology of Domestic Animals, KU Leuven, 3001 Heverlee, Belgium
Accepted 15 April 2005
Abstract
While there have been many studies in various species examining the mode of central leptin action on food intake, there is however a
paucity of data in birds. We have, therefore, addressed this issue in broiler chickens because this strain was selected for high growth rate, hence
high food intake. Continuous infusion of recombinant chicken leptin (8 Ag/kg/h) during 6 h at a constant rate of 3 ml/h resulted in a significant
reduction (49–57%) of food intake in 3-week-old broiler chickens (P < 0.05). The effect of leptin within the central nervous system (CNS)
was mediated via selective hypothalamic neuropeptides. Leptin significantly decreased the expression of its receptor (Ob-R), neuropeptide Y
(NPY), orexin (ORX), and orexin receptor (ORXR) (P < 0.05), but not that of agouti-related protein (AgRP) (anabolic/orexigenic effectors) in
chicken hypothalamus. However, the catabolic/anorexigenic neuropeptides namely proopiomelanocortin (POMC) and corticotropin-releasing
hormone (CRH) mRNA levels remained unchanged after leptin treatment. Despite the absence of leptin effect on AgRP (the antagonist of
melanocortin receptor MCR) and POMC (the precursor of a-melanocyte stimulating hormone which is a potent agonist for MCR), leptin
significantly decreased the expression of MCR-4/5 gene in chicken hypothalamus (P < 0.05) suggesting that leptin acts directly (as ligand) or
indirectly (via other ligands) on MCRs to regulate food intake in birds. Additionally, leptin down-regulated the expression of fatty acid
synthase (FAS) gene in chicken hypothalamus, indicating an additional pathway of leptin action on food intake such as described for FAS
inhibitors. These findings provide new insight into the mechanism of leptin control of food intake in chickens.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Chicken leptin; Food intake; Neuropeptides; FAS; Broiler; Hypothalamus
1. Introduction
Food intake is regulated via neural circuits located in the
hypothalamus. During the past decade, our knowledge on the
specific mediators and neuronal networks that regulate food
intake and body weight has increased dramatically. An
important contribution to the understanding of hypothalamic
control of food intake has been the characterization of the ob
gene product (leptin) via positional cloning [82]. Leptin, a
16-kDa peptide hormone, is mainly secreted by adipose
tissue in mammals [82] and functions as a hormonal sensing
mechanism for fat deposition and body weight homeostasis
[25,28,54]. Mammalian adipocytes produce and secrete
more leptin in the bloodstream as fat storage increases
0006-8993/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2005.04.034
* Corresponding author. Fax: +32 16321994.
E-mail address: [email protected] (S. Dridi).
[44], signaling the brain via leptin receptor [3,22,23,48] and
modulating the hypothalamic neuropeptide system to inhibit
food intake and increase energy expenditure [25,47,64,
65,81]. Among these hypothalamic neuropeptides, the
best-described anabolic effectors are neuropeptide Y
(NPY) [39,71] and agouti gene-related peptide (AgRP)
[27,60]. The catabolic counterparts of NPY/AgRP system
are proopiomelanocortin (POMC), the precursor of alpha-
melanocyte stimulating hormone (a-MSH) [11] and cocaine-
and amphetamine-regulated transcript (CART) [38]. NPY
and AgRP (NPY/AgRP) are co-localized in neurons in the
arcuate nucleus (ARC) [27], whereas POMC and CART
(POMC/CART) are expressed in distinct neurons in the ARC
[20]. NPY/AgRP and POMC/CART neurons in the ARC
project to second-order neurons, including those in the
paraventricular nucleus (PVN), perifornical area (PFA), and
lateral hypothalamus (LHA) [21]. The second-order target
neurons selectively express additional neuropeptides such as
(2005) 214 – 223

S. Dridi et al. / Brain Research 1047 (2005) 214–223 215
melanin-concentrating hormone (MCH) and orexin (hypo-
cretin) in LHA that can induce feeding [18,55,61,66] or
corticotropin-releasing hormone (CRH), thyrotropin-releas-
ing hormone (TRH), and oxytocin in the PVN that can
inhibit feeding [33,53,70]. It is the balance between these
two pathways (anabolic and catabolic) that ultimately
determines the animal’s ingestive behavior and defended
the body weight set point. More recently, it has been shown,
by using specific inhibitors, that fatty acid synthase (FAS)
regulates food intake in mammals [36,41,67] and that the
mechanism may involve hypothalamic neuropeptides as
those trigged by leptin [45]. The mechanism of central neu-
ropeptide-mediating leptin action on feeding behavior has
been the subject of many studies in mammals over the last
few years. Exogenous administration of leptin decreased the
orexigenic peptides (anabolic effectors) and increased the
anorexigenic ones (catabolic effectors) resulting in the
reduction of food intake. In chicken, leptin manifests some
particularities compared to that in mammals: (1) chicken
leptin is expressed not only in adipose tissue but also in liver
[2,78], (2) liver is the major source for leptin [58], (3) leptin
gene expression is sensitive to hormonal treatment in liver
but not in adipose tissue [2], and (4) chicken leptin protein
contains three cysteine residues as compared to two in
mammalian leptin [78]. Rock et al. [59] showed that these
two cysteines are involved in an intrachain disulfide bridge
that is critical for the structural integrity and biological acti-
vity of leptin in mammals. Despite these peculiarities, exo-
genous administration of leptin (recombinant chicken or
human leptin) to chickens resulted in reduction in food
intake as in mammals [14,17,57], whereas the mechanisms
involved are not yet known. Therefore, we undertook this
study to identify the targets of leptin action in chicken
hypothalamus.
2. Materials and methods
2.1. Recombinant chicken leptin infusion
Animal study was conducted with research protocol
approved by Ethical Commission for Experimental Use of
Animals of the Catholic University of Leuven (Belgium).
Day-old male broiler chicks (Ross) were purchased from
Avibel, Zoersel (Belgium) and reared on floor pen until 2
weeks of age, at which time the birds were transferred to
individual cages and provided with individual feeders and
drinking nipples. After 3 days of adaptation, birds were
weighed and cannulated in the brachial artery under local
anesthesia (xylocaine) [7]. The chickens were allowed to
recover and to adapt during four more days. Before the
infusion experiment, the chickens were divided into two
homogenous weight-matched groups (n = 5) and fasted for
2 h in order to increase their appetite. The mini pump
(Syringe pump series, Model 22, Harvard apparatus, USA)
infused recombinant chicken leptin prepared as previously
described [57] (8 Ag/kg/h) or saline at constant rate of 3 ml/
h during 6 h. Serial blood samples were taken at different
times (0, 1.5, 3, 4.5, and 6 h), and food intake was recorded
at 1, 2, 3, 4, 5, and 6 h after mini pump implantation. Birds
were sacrificed by cervical dislocation, and tissues (hypo-
thalamus) were removed and snap frozen in liquid nitrogen
and stored at �80 -C until use.
2.2. RNA extraction
Total RNA was extracted from hypothalamus using the
Trizol reagent (Invitrogen, Belgium) according to manufac-
turer’s recommendation. Pellets were suspended in 20–30 Alof DEPC-treated water. The quantity and integrity of
isolated RNA were determined for each sample by using
both UV absorbance (260/280) as well as by 1% agarose gel
electrophoresis.
2.3. Reverse transcription and polymerase chain reaction
(RT-PCR)
Total RNA from hypothalamus (1 Ag) was reverse
transcribed in a final volume of 20 Al containing 10 units
of AMV reverse transcriptase (Promega, Belgium), 1 mM
dNTP mixture (Promega, Belgium), 40 units of recombinant
RNasin ribonuclease inhibitor (Promega, Belgium), and
0.5 Ag of random primers (Promega, Belgium) in sterilized
water and buffer supplied by the manufacturer. After incu-
bation (42 -C, 45 min), the mixture was heated (80 -C,3 min). PCRwas performed in 50 Al containing 2 Al of the RTreaction, 1 unit of Taq DNA polymerase (Roche Diagnostic,
Belgium), 0.1 mM dNTP mixture, and 10 pmol of each
forward and reverse primer for FAS, hypothalamic neuro-
peptides, and ribosomal 18S (Table 1). Thermal cycling
parameters were as follows: 1 cycle of 94 -C for 2–9 min
followed by 25–30 cycles (35 cycles for characterization and
probe preparation) of 94 -C for 30–60 s, 58–60 -C for 1 min,
72 -C for 1 min with a final extension at 72 -C for 10 min.
2.4. Probe labeling
The amplified fragments (FAS, neuropeptides and 18S)
were separated on a low-melting point agarose gel (1%), and
the appropriate bands were cut out, purified by using
QIAquick Gel extraction kit protocol (Qiagen, Belgium),
and stored at �20 -C. The cDNA fragments were cloned
using the TOPO PCR cloning kit (Invitrogen, The Nether-
lands) and automatically sequenced using an ABI automated
sequencer. Then, 25–30 ng of cloned probes was labeled by
random priming with (a-32P)dCTP [24].
2.5. Southern blot analysis
The amplified PCR products were transferred to nylon
membrane by vacuum blotting apparatus (Amersham
Biosciences, the Netherlands) and cross linked by ultraviolet
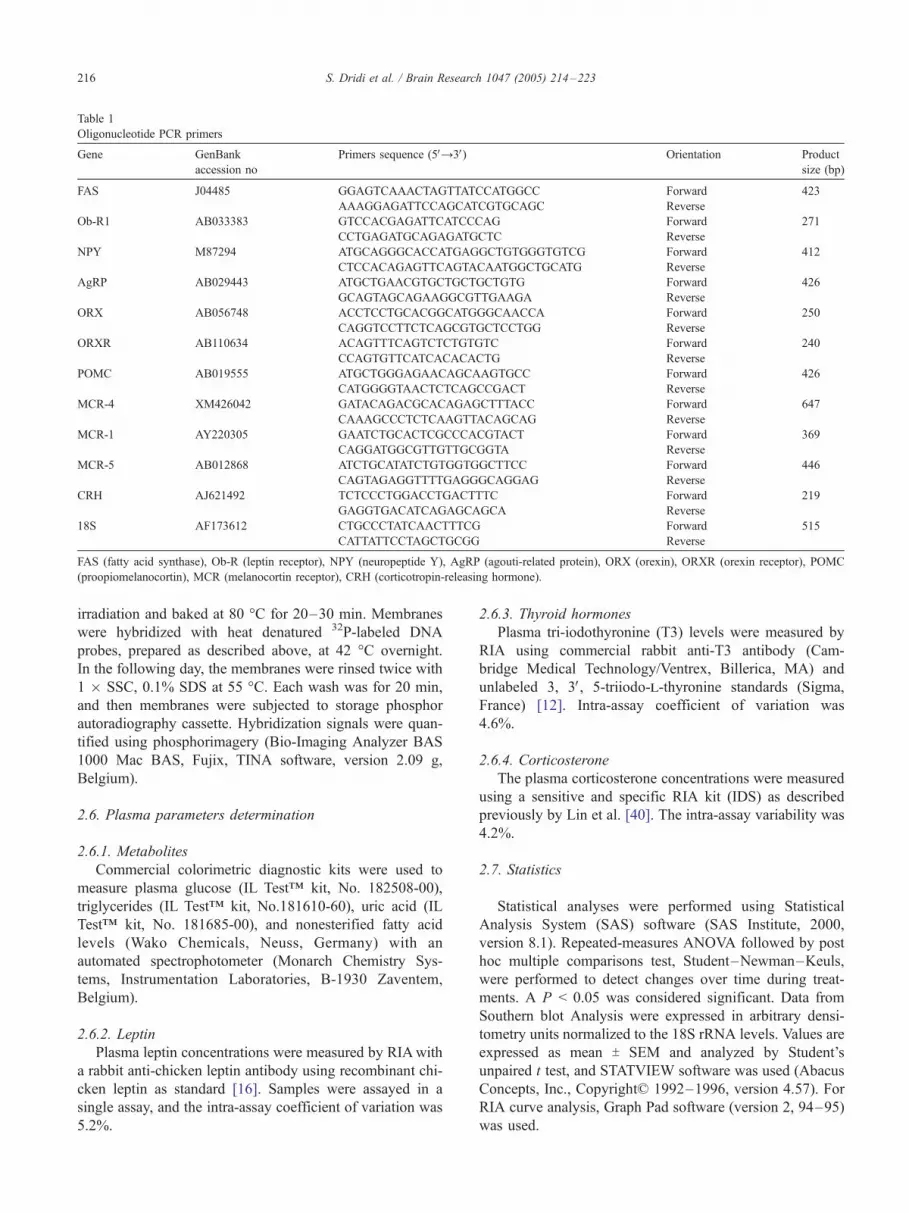
Table 1
Oligonucleotide PCR primers
Gene GenBank
accession no
Primers sequence (5VY3V) Orientation Product
size (bp)
FAS J04485 GGAGTCAAACTAGTTATCCATGGCC Forward 423
AAAGGAGATTCCAGCATCGTGCAGC Reverse
Ob-R1 AB033383 GTCCACGAGATTCATCCCAG Forward 271
CCTGAGATGCAGAGATGCTC Reverse
NPY M87294 ATGCAGGGCACCATGAGGCTGTGGGTGTCG Forward 412
CTCCACAGAGTTCAGTACAATGGCTGCATG Reverse
AgRP AB029443 ATGCTGAACGTGCTGCTGCTGTG Forward 426
GCAGTAGCAGAAGGCGTTGAAGA Reverse
ORX AB056748 ACCTCCTGCACGGCATGGGCAACCA Forward 250
CAGGTCCTTCTCAGCGTGCTCCTGG Reverse
ORXR AB110634 ACAGTTTCAGTCTCTGTGTC Forward 240
CCAGTGTTCATCACACACTG Reverse
POMC AB019555 ATGCTGGGAGAACAGCAAGTGCC Forward 426
CATGGGGTAACTCTCAGCCGACT Reverse
MCR-4 XM426042 GATACAGACGCACAGAGCTTTACC Forward 647
CAAAGCCCTCTCAAGTTACAGCAG Reverse
MCR-1 AY220305 GAATCTGCACTCGCCCACGTACT Forward 369
CAGGATGGCGTTGTTGCGGTA Reverse
MCR-5 AB012868 ATCTGCATATCTGTGGTGGCTTCC Forward 446
CAGTAGAGGTTTTGAGGGCAGGAG Reverse
CRH AJ621492 TCTCCCTGGACCTGACTTTC Forward 219
GAGGTGACATCAGAGCAGCA Reverse
18S AF173612 CTGCCCTATCAACTTTCG Forward 515
CATTATTCCTAGCTGCGG Reverse
FAS (fatty acid synthase), Ob-R (leptin receptor), NPY (neuropeptide Y), AgRP (agouti-related protein), ORX (orexin), ORXR (orexin receptor), POMC
(proopiomelanocortin), MCR (melanocortin receptor), CRH (corticotropin-releasing hormone).
S. Dridi et al. / Brain Research 1047 (2005) 214–223216
irradiation and baked at 80 -C for 20–30 min. Membranes
were hybridized with heat denatured 32P-labeled DNA
probes, prepared as described above, at 42 -C overnight.
In the following day, the membranes were rinsed twice with
1 � SSC, 0.1% SDS at 55 -C. Each wash was for 20 min,
and then membranes were subjected to storage phosphor
autoradiography cassette. Hybridization signals were quan-
tified using phosphorimagery (Bio-Imaging Analyzer BAS
1000 Mac BAS, Fujix, TINA software, version 2.09 g,
Belgium).
2.6. Plasma parameters determination
2.6.1. Metabolites
Commercial colorimetric diagnostic kits were used to
measure plasma glucose (IL Testi kit, No. 182508-00),
triglycerides (IL Testi kit, No.181610-60), uric acid (IL
Testi kit, No. 181685-00), and nonesterified fatty acid
levels (Wako Chemicals, Neuss, Germany) with an
automated spectrophotometer (Monarch Chemistry Sys-
tems, Instrumentation Laboratories, B-1930 Zaventem,
Belgium).
2.6.2. Leptin
Plasma leptin concentrations were measured by RIAwith
a rabbit anti-chicken leptin antibody using recombinant chi-
cken leptin as standard [16]. Samples were assayed in a
single assay, and the intra-assay coefficient of variation was
5.2%.
2.6.3. Thyroid hormones
Plasma tri-iodothyronine (T3) levels were measured by
RIA using commercial rabbit anti-T3 antibody (Cam-
bridge Medical Technology/Ventrex, Billerica, MA) and
unlabeled 3, 3V, 5-triiodo-l-thyronine standards (Sigma,
France) [12]. Intra-assay coefficient of variation was
4.6%.
2.6.4. Corticosterone
The plasma corticosterone concentrations were measured
using a sensitive and specific RIA kit (IDS) as described
previously by Lin et al. [40]. The intra-assay variability was
4.2%.
2.7. Statistics
Statistical analyses were performed using Statistical
Analysis System (SAS) software (SAS Institute, 2000,
version 8.1). Repeated-measures ANOVA followed by post
hoc multiple comparisons test, Student–Newman–Keuls,
were performed to detect changes over time during treat-
ments. A P < 0.05 was considered significant. Data from
Southern blot Analysis were expressed in arbitrary densi-
tometry units normalized to the 18S rRNA levels. Values are
expressed as mean T SEM and analyzed by Student’s
unpaired t test, and STATVIEW software was used (Abacus
Concepts, Inc., Copyright* 1992–1996, version 4.57). For
RIA curve analysis, Graph Pad software (version 2, 94–95)
was used.

S. Dridi et al. / Brain Research 1047 (2005) 214–223 217
3. Results
3.1. Effect of recombinant chicken leptin infusion on food
intake in broiler chickens
Three-week-old broiler chickens were divided into two
homogeneous weight-matched groups (n = 5) and were
treated with recombinant chicken leptin or saline solution
(placebo) as described in Materials and methods. After 2 h of
food deprivation, leptin or saline solutions were continu-
ously infused, and animals were refed immediately after start
of infusion. Cumulative food intake was measured at 1, 2, 3,
4, 5, and 6 h after start of feeding. As shown in Fig. 1,
recombinant chicken leptin significantly reduced (P < 0.05
to P < 0.01) cumulative food intake during the next 2–6 h of
feeding with a mean reduction of about 49.1–57.3% as
compared to the placebo.
3.2. Effect of recombinant chicken leptin infusion on plasma
hormone levels
The administration of recombinant chicken leptin resulted
in a significant increase of plasma leptin levels during the
next 1 h 30–6 h of feeding (P < 0.05). The leptinemia in
leptin-treated group peaked 90 min postinjection at 62 ng/ml
and remained higher (17.9 to 37.3-folds) than that in vehicle-
treated group (55.7 T 4.7 to 65.03 T 2.85 vs. 2.52 T 0.25 to
3.46 T 0.23 ng/ml; mean T SEM, n = 5) (Fig. 2A). Leptin
tended to decrease plasma T3 levels, however, this reduction
(4.3 to 25.9% compared to control group) was not statisti-
cally significant at the 5% level (Fig. 2B). The effect of leptin
on plasma corticosterone levels was not consistent and not
significantly different compared to untreated group (Fig.
2C). It should be noted that a high individual variability was
observed in circulating corticosterone concentrations.
Fig. 1. Effect of recombinant chicken leptin infusion on cumulative food
intake in 3-week-old broiler chickens. Recombinant chicken leptin (8 Ag/kg/h) or saline solution was continuously administered at a constant rate of
3 ml/h during 6 h. Cumulative food intake was measured continuously and
presented graphically as mean T SEM (n = 5) and * indicates a significant
difference between leptin-treated group and the placebo at P < 0.05.
Fig. 2. Effect of recombinant chicken leptin on plasma hormone levels.
Recombinant chicken leptin (8 Ag/kg/h) or saline solution was continuouslyinfused at constant rate of 3 ml/h in 3-week-old broiler chickens during 6 h.
Circulating leptin (A), T3 (B), and corticosterone levels (C) were measured at
0, 1.5, 3, 4.5, and 6 h by RIAs, and values are mean T SEM (n = 5).
* Indicates a significant difference between the two groups at P < 0.05.
3.3. Effect of recombinant chicken leptin infusion on plasma
metabolite levels
Recombinant chicken leptin administration resulted in a
slight, but not significant decrease of plasma glucose levels
during the 1 h 30–6 h of feeding (Fig. 3A). The reduction
average was about 12.5–17.8% as compared to the control
group. The effect of leptin was not consistent on circulating
triglycerides, nonesterified fatty acids (NEFA), and uric acid
concentrations (Figs. 3B, C, and D, respectively).

Fig. 3. Effect of recombinant chicken leptin on plasma metabolite concentrations. Leptin administration was 8 Ag/kg/h during 6 h at constant rate of 3 ml/h.
Plasma glucose (A), triglycerides (B), nonesterified fatty acids (NEFA) (C), and uric acid (D) were continuously determined as described in Materials and
methods. Values are mean T SEM (n = 5).
S. Dridi et al. / Brain Research 1047 (2005) 214–223218
3.4. Effect of recombinant chicken leptin infusion on chicken
hypothalamic fatty acid synthase (FAS) and leptin receptor
(Ob-R) gene expression
Leptin administration during 6 h significantly down-
regulated the hypothalamic FAS and Ob-R gene expression
by 21 and 15.2% respectively (P < 0.05, Fig. 4) as compared
to the control group.
3.5. Effect of recombinant chicken leptin infusion on chicken
hypothalamic anabolic/orexigenic peptides
Recombinant chicken leptin significantly reduced both
NPY, orexin (ORX), and ORXR gene expression by 14,
13.5, and 7% respectively (P < 0.05, Fig. 5) in the
hypothalamus of 3-week-old broiler chickens. However,
AgRP mRNA levels were not significantly affected by leptin
treatment (Fig. 5).
3.6. Effect of recombinant chicken leptin infusion on chicken
hypothalamic catabolic/anorexigenic peptides
Recombinant chicken leptin administration did not signi-
ficantly affect the POMC, MCR-1, and CRH gene expres-
sion in the hypothalamus of 3-week-old broiler chickens.
However, MCR-4, which plays the most important role in
mediating catabolic effects of a-MSH, and MCR-5 were
significantly reduced in leptin-treated group compared to
untreated group (0.911 T 0.044 vs. 0.592 T 0.066 and 0.587 T0.014 vs. 0.487 T 0.031, respectively, for MCR-4 and
-5; mean T SEM; arbitrary unit; P < 0.05) (Fig. 6).
4. Discussion
The biological activity of the recombinant chicken leptin
was demonstrated previously by its ability to stimulate the
proliferation of BAF/3 1442-CL4 cells, in vitro, transfected
with the functional long form of the human leptin receptor
[2] and in vivo by its inhibitory effect on food intake after a
single peripheral (intraperitoneal or intravenous) injection
[17,57]. Furthermore, Denbow et al. [14] have shown that
intracerebroventricular (ICV) injection of recombinant
human leptin decreased food intake in both broiler and
leghorn chickens in a dose-dependent manner, with 10 Ag asthe most efficacious dose. However, Bungo et al. [5] reported
that ICVadministration of recombinant mouse leptin did not
affect food intake in chickens. In this study, we have shown
that continuous infusion of recombinant chicken leptin (8 Ag/kg/h) during 6 h resulted in a 49–57% reduction of food

Fig. 5. Effect of recombinant chicken leptin infusion on neuropeptide Y
(NPY), agouti-related protein (AgRP), orexin (ORX), and orexin receptor
(ORXR) gene expression in the hypothalamus of 3-week-old broiler
chickens. Total RNA from hypothalamus was prepared and subjected to
RT-PCR and Southern blot analysis as described previously in Materials and
methods. Levels of NPY, AgRP, ORX, and ORXR gene expression are
expressed in arbitrary units relative to 18S levels and presented graphically as
mean T SEM (n = 5) and * indicates a significant difference at P < 0.05.
Fig. 4. Effect of recombinant chicken leptin on hypothalamic fatty acid
synthase (FAS) and leptin receptor (Ob-R) gene expression in 3-week-old
broiler chickens. Recombinant chicken leptin (8 Ag/kg/h) or saline solutionwas continuously administered at a constant rate (3 ml/h) during 6 h. Total
RNA was extracted and subjected to RT-PCR coupled to Southern blot
analysis. Data are presented as ratio of specific genes (FAS and Ob-R) to
ribosomal 18S gene expression and values are mean T SEM of 5 chickens
per group. *P < 0.05 indicates a significant difference between leptin- and
vehicle-treated groups.
S. Dridi et al. / Brain Research 1047 (2005) 214–223 219
intake in 3-week-old broiler chickens and support those
found in birds and in other species [4,9,28]. The observed
numeric, but not significant, decrease of plasma T3 and
glucose levels in leptin-infused chickens may be a con-
sequence of the lowered food intake as this will affect both
circulating glucose and T3 concentrations in a complex and
interactive way as described in previous studies [8]. The new
finding in this study was the identification of the central site
of leptin action on food intake regulation in birds. We have
shown firstly that leptin administration significantly reduced
the leptin receptor gene expression in chicken hypothalamus
confirming that leptin acts within the CNS to regulate food
intake. However, the mechanism(s) involved in the transport
of leptin to across the blood–brain barrier (BBB) remained
unclear. Indeed, McMurtry et al. [46] showed that no leptin
binding protein was detected in blood of chickens, whereas,
in mammals, leptin is secreted from adipose tissue, circulates
in the blood where it binds to a family of binding proteins
including the soluble leptin receptor (Ob-Re) [15,29,69],
then crosses the BBB to enter the brain [6,31,37], and then
interacts with hypothalamic neuropeptides that regulate
ingestive behavior. In this study, we have shown also that
recombinant chicken leptin significantly reduced orexigenic
neuropeptides (NPY, ORX, and ORXR). This result corrob-
orates previous findings in mammals [42,62]. NPY stim-
ulates food intake in birds similarly to that in mammals [35]
and has also been shown to be localized within the
hypothalamus [34]. Ohkubo et al. [52] showed that ORX
was expressed within the hypothalamus of chickens and is
induced by fasting. However, ICV administration of mam-
malian ORX did not affect food intake in neonatal chicks
[26]. Interestingly, leptin administration did not affect AgRP
(orexigenic neuropeptide) and POMC and CRH (anorexi-
genic/catabolic neuropeptides). This observation is com-
pletely different from the results obtained in mammals
because leptin-treated mammals showed lower AgRP
[19,49] and higher POMC and CRH mRNA levels than that
of vehicle-treated group [50,51,63,79,80]. This discrepancy
may be related to site or species-specific effects of leptin.
Indeed, Arvaniti et al. [1] showed that leptin treatment
decreased CRH mRNA levels in the paraventricular hypo-
thalamic nucleus (PVH) and enhanced CRH gene expression
in the central nucleus of amygdala and in the bed nucleus of
the stria terminalis in mice. In our study, we sampled the
whole hypothalamus, and the effect of leptin on CRHmay be
different according to the hypothalamic area. Denbow et al.
[13] have shown that ICV administration of CRF signifi-
cantly decreased food intake in both fed and overnight-fasted

Fig. 6. Effect of recombinant chicken leptin infusion on proopiomelano-
cortin (POMC), corticotropin-releasing hormone (CRH), and melanocortin
receptors (MCR-1, -4, and -5) gene expression in 3-week-old broiler
chickens. Recombinant chicken leptin (8 Ag/kg/h) or saline solution was
administered during 6 h at a constant rate of 3 ml/h. Relative hypothalamic
anorexigenic neuropeptide (POMC, CRH, MCRs) gene expression was
determined by RT-PCR and Southern blot analysis and graphically presented
as mean T SEM (n = 5). *P < 0.05 indicates a significant difference between
leptin-treated and untreated group.
S. Dridi et al. / Brain Research 1047 (2005) 214–223220
broiler and leghorn chickens. a-MSH is derived from the
cleavage of the precursor, POMC, and significantly inhibited
fasting- and NPY-induced feeding in neonatal chicks in a
dose-dependent manner when administered by ICV injection
[32]. The anorexigenic effect of a-MSH was abolished after
the administration of AgRP in chickens [72]. It has been
shown that the effect of AgRP differed between chicken
strains; it stimulated food intake in layer but not in broiler-
type chickens [72]. These previous studies suggested that the
anorexigenic effect of endogenous a-MSH may not be
important in broiler chickens. Present results seem to support
their suggestion indicating that leptin may act selectively via
orexigenic neuropeptides (NPY and ORX, but not AgRP)
rather than anorexigenic pathways (POMC and CRH in this
study) at least in broiler chickens. and further studies are
required to examine other chicken strains. Additionally, we
used in this study male chickens, and the effect of leptin and
its interaction with hypothalamic neuropeptides may depend
on gender as previously reported for sheep [10]. It is also
known that the melanocortin system coordinates the main-
tenance of energy balance in mammals via the regulation of
both food intake and energy expenditure [30]. The adipo-
genic hormone leptin, which is involved in the regulation of
energy balance, is thought to act by stimulating the
production of a-MSH (the potent agonist of MCR-3 and
-4) [39] and inhibited the release of AgRP (the antagonist of
MCR-3 and -4) in mammals [43,56]. Five receptor genes
belonging to the MCR family have been recently cloned in
birds [73,75–77]. MCR-1 is implicated in melanogenesis
within melanocytes since it has been mapped at the genetic
locus which acts to control feather color pigmentation in the
chicken [74]. MCR-2 has been suggested to regulate
steroidogenesis in chicken [76], however, no clear function
has yet been ascribed to the other MCRs. MCR-3 is
expressed exclusively in the adrenal gland [77], while
MCR-4 and-5 showed a ubiquitous expression in peripheral
tissues as well as in the brain [73]. In this study, we have
shown that leptin significantly reduced MCR-4 and -5 gene
expression in chicken hypothalamus, whereas MCR-1
mRNA levels remained unchanged. These data suggest that
the central effects of leptin on food intake in chickens may
involve changes in signaling of MCR-4 and -5, however,
whether this interaction is direct or indirect remained unclear.
This result is intriguing and particularly of interest because it
is different from the classic model of leptin action in
mammals where leptin increased the production of a-MSH
(agonist of MCR-3 and -4) and decreased the release of
AgRP (antagonist of MCR-3 and -4) in the hypothalamus,
thereby modulating MCRs gene expression and regulating
food intake. The changes observed in MCR-4 and -5 gene
expression in this study without modification of both POMC
and AgRP gene expression suggest that leptin may act
directly as a ligand or indirectly via other ligands for MCRs.
The interaction between leptin and MRCs in the regulation
of food intake in chicken needs to be explored further. It has
been reported that naturally occurring fatty acid synthase
inhibitor cerulenin and its more potent synthetic analog C75
robustly inhibit food intake, reduce body weight, and
increase metabolic rate when injected into mice, possibly
by acting on hypothalamic glucose-sensing neurons through
a leptin-independent mechanism involving malonyl-CoA
accumulation [41]. However, other studies have reported that
these compounds modulate hypothalamic neuropeptides in a
manner similar to that of leptin [68]. In this study, we have
shown for the first time, in our knowledge, that leptin
reduces FAS gene expression, as cerulenin and C75, in the
hypothalamus of chickens suggesting a potential role of
leptin/FAS in the control of food intake in birds, while the
molecular mechanism(s) involved are unknown and beg
answers.
In conclusion, the present study is the first to report,
additionally to the single ICV or peripheral injection of

S. Dridi et al. / Brain Research 1047 (2005) 214–223 221
leptin, that continuous infusion of leptin inhibits food intake
in chickens and to identify some central targets of leptin
action in birds. It seems that leptin selectively modulates
anabolic (orexigenic) neuropeptides rather than catabolic
(anorexigenic) pathways to regulate ingestive behavior in
broiler chickens. The new finding that needs to be explored
in the future is the interaction, on the one hand, between
leptin and FAS, and on the other hand, between leptin and
MCRs in the regulation of food intake and energy balance in
chickens.
Acknowledgments
We wish to thank Gerda Nackaerts and Inge Vaesen for
their skilled technical assistance. This work was supported
by research grant (G012201) from the FWO-Flanders.
References
[1] K. Arvaniti, Q. Huang, D. Richard, Effects of leptin and corticosterone
on the expression of corticotrophin-releasing hormone, agouti-related
protein, and proopiomelanocortin in the brain of ob/ob mouse,
Neuroendocrinology 73 (2001) 227–236.
[2] C.M. Ashwell, S.M. Czerwinski, D.M. Brocht, J.P. McMurtry,
Hormonal regulation of leptin expression in broiler chickens, Am. J.
Physiol. 276 (1999) R226–R232.
[3] W.A. Banks, A.J. Kastin, W. Huang, J.B. Jaspan, L.M. Maness, Leptin
enters the brain by a saturable system independent of insulin, Peptides
17 (1996) 305–311.
[4] C.R. Barb, X. Yan, M.J. Azain, R.R. Draeling, G.B. Rampaeek, T.G.
Ramsay, Recombinant porcine leptin reduces feed intake and
stimulates growth hormone secretion in swine, Domest. Anim.
Endocrinol. 15 (1991) 77–86.
[5] T. Bungo, M. Shimojo, Y. Masuda, T. Tachibanab, S.J. Tanaka, K.
Sugahara, M. Furuse, Intracerebroventricular administration of mouse
leptin does not reduce food intake in the chicken, Brain Res. 817 (1999)
196–198.
[6] B. Burguera, M.E. Couce, Leptin access into the brain. A saturated
transport mechanism in obesity, Physiol. Behav. 74 (2001) 717–720.
[7] J. Buyse, A. Vanderpooten, B. Leclercq, R. Berghman, E. Decuypere,
Pulsatility of plasma growth hormone and hepatic growth hormone
receptor characteristics of broiler chickens divergently selected for
abdominal fat content, Br. Poultry Sci. 35 (1994) 145–152.
[8] J. Buyse, K. Janssens, S. Van der Geyten, P. Van As, E. Decuypere,
V.M. Darras, Pre- and postprandial changes in plasma hormone and
metabolite levels and hepatic deiodinase activities in meal-fed broiler
chickens, Br. J. Nutr. 88 (2002) 641–653.
[9] L.S. Campfield, F.J. Smith, Y. Guisez, R. Devos, P. Burn,
Recombinant mouse ob protein: evidence for a peripheral signal
linking adipocity and the central neural networks, Science 269 (1995)
546–549.
[10] I.J. Clarke, A.J. Tilbrook, A.I. Turner, B.W. Doughton, J.W. Goding,
Sex, fat and tilt of the earth: effects of sex and season on the feeding
response to centrally administered leptin in sheep, Endocrinology 142
(2001) 2725–2728.
[11] R.D. Cone, The central melanocortin system and energy homeostasis,
Trends Endocrinol. Metab. 10 (1999) 211–216.
[12] V.M. Darras, T.J. Visser, L.R. Berghman, E.R. Kuhn, Ontogeny of
type I and III deiodinase activities in embryonic and posthatch chicks:
relationship with changes in plasma triiodothyronine and growth
hormone levels, Comp. Biochem. Physiol. 103A (1992) 131–136.
[13] M.D. Denbow, N. Snapir, M. Furuse, Inhibition of food intake by CRF
in chickens, Physiol. Behav. 66 (1999) 645–649.
[14] D.M. Denbow, S. Meade, A. Robertson, J.P. McMurtry, M. Richards,
C. Ashwell, Leptin-induced decrease in food intake in chickens,
Physiol. Behav. 69 (2000) 359–362.
[15] F.B.J. Diamond, D.C. Eichler, G. Duckett, E.V. Jorgensen, D. Shul-
man, A.W. Root, Demonstration of a leptin binding factor in human
serum, Biochem. Biophys. Res. Commun. 233 (1997) 818–822.
[16] S. Dridi, J. Williams, V. Bruggeman, M. Onagbesan, N. Raver, E.
Decuypere, J. Djiane, A. Gertler, M. Taouis, A chicken leptin specific
radioimmunoassay, Domest. Anim. Endocrinol. 18 (2000) 325–335.
[17] S. Dridi, N. Raver, E.E. Gussakovsky, M. Derouet, M. Picard, A.
Gertler, M. Taouis, Biological activities of recombinant chicken leptin
C4S analog compared with unmodified leptons, Am. J. Physiol. 279
(2000) E116–123.
[18] M. Dube, S. Kalra, P. Kalra, Food intake elicited by central
administration of orexins/hypocretins: identification of hypothalamic
sites of action, Brain Res. 842 (1999) 473–477.
[19] K. Ebihara, Y. Ogawa, G. Katsuura, Y. Numata, H. Masuzaki, N.
Satoh, M. Tamaki, T. Yoshioka, M. Hayase, N. Matsuoka, M. Aizawa-
Abe, Y. Yoshimasa, K. Nakao, Involvement of agouti-related protein,
an endogenous antagonist of hypothalamic melanocortin receptor, in
leptin action, Diabetes 48 (1999) 2028–2033.
[20] C.F. Elias, C. Lee, J. Kelly, C. Aschkenasic, R.S. Ahima, P.
Couceyro, M.J. Kuhar, C.B. Saper, J.K. Elmquist, Leptin activates
hypothalamic CART neurons projecting to the spinal cord, Neuron 21
(1998) 1375–1385.
[21] C.F. Elias, C.B. Saper, E. Maratos-Flier, N.A. Tritos, C. Lee, J. Kelly,
J.B. Tatro, G.E. Hoffman, M.M. Ollmann, G.S. Barsh, T. Sakurai, M.
Yanagisawa, J.K. Elmquist, Chemically defined projections linking the
mediobasal hypothalamus and the lateral hypothalamic area, J. Comp.
Neurol. 402 (1998) 442–459.
[22] J.K. Elmquist, R.S. Ahima, C.F. Elias, J.S. Flier, C.B. Saper,
Distributions of leptin receptor mRNA isoforms in the rat brain,
J. Comp. Neurol. 395 (1998) 535–547.
[23] H. Fei, H.J. Okano, C. Li, G.H. Lee, C. Zhao, R. Darnell, J.M.
Friedman, Anatomic localization of alternatively spliced leptin
receptors (ob-R) in mouse brain and other tissues, Proc. Natl. Acad.
Sci. U. S. A. 94 (1997) 7001–7005.
[24] A.P. Feinberg, B. Vogelstein, A technique for radiolabeling DNA
restriction endonuclease fragments to high specific activity, Anal.
Biochem. 132 (1983) 6–13.
[25] J.M. Friedman, J.L. Halaas, Leptin and the regulation of body weight
in mammals, Nature 395 (1998) 763–770.
[26] M. Furuse, R. Ando, T. Bungo, R. Ao, M. Shimojo, Y. Masuda,
Intracerebroventricular injection of orexins does not stimulate food
intake in neonatal chicks, Br. Poultry Sci. 40 (1999) 698–700.
[27] T. Hahn, J. Breininger, D. Baskin, M. Schwartz, Coexpression of
AGRP and NPY in fasting-activated hypothalamic neurons, Nat.
Neurosci. 1 (1998) 271–272.
[28] J.L. Halaas, K.S. Kajiwala, M. Maffei, S.L. Cohen, B.T. Chait, D.
Rabinowitz, R.L. Lallone, S.K. Burley, J.M. Friedman, Weight-
reducing effects of the plasma protein encoded by the obese gene,
Science 269 (1995) 543–546.
[29] K.L. Houseknecht, C.S. Mantzoros, R. Kuliawat, E. Hadro, J.S. Flier,
B.B. Kahn, Evidence for leptin binding to proteins in serum of
rodents and humans: modulation with obesity, Diabetes 45 (1996)
1638–1643.
[30] D. Huszar, C.A. Lynch, V. Fairchild-Hunteress, J.H. Dunmore, Q.
Fang, L.R. Berkemeier, W. Gu, R.A. Kesterson, B.A. Boston, R.D.
Cone, F.J. Smith, L.A. Campfield, P. Burn, F. Lee, Targeted disruption
of the melanocortin-4 receptor results in obesity in mice, Cell 88
(1997) 131–141.
[31] A.J. Kastin, W. Pan, Dynamic regulation of leptin entry into brain by
the blood–brain barrier, Regul. Pept. 92 (2000) 37–43.
[32] S.I. Kawakami, T. Bungo, R. Ando, A. Ohgushi, M. Shimojo, Y.
Masuda, M. Furuse, Central administration of a-melanocyte stimulat-

S. Dridi et al. / Brain Research 1047 (2005) 214–223222
ing hormone inhibits fasting- and neuropeptide Y-induced feeding in
neonatal chicks, Eur. J. Pharmacol. 398 (2000) 361–364.
[33] L. Kow, D. Pfaff, The effects of the TRH metabolite cyclo- (His-Pro-
and its analogs on feeding, Pharmacol. Biochem. Behav. 38 (1991)
359–364.
[34] W.J. Kuenzel, J. McMurtry, Neuropeptide Y: brain localization and
central effects on plasma insulin levels in chicks, Physiol. Behav. 44
(1988) 669–678.
[35] W.J. Kuenzel, L.W. Douglass, B.A. Davison, Robust feeding follow-
ing central administration of neuropeptide Y or peptide YY in chicks,
Peptides 8 (1987) 823–828.
[36] M.V. Kumar, T. Shimokawa, T.R. Nagy, M.D. Lane, Differential
effects of a centrally acting fatty acid synthase inhibitor in lean and
obese mice, Proc. Natl. Acad. Sci. U. S. A. 99 (2002) 1921–1925.
[37] D. Kurrimbux, Z. Gaffen, C.L. Farrell, D. Martin, S.A. Thomas, The
involvement of the blood–brain and the blood–cerebrospinal fluid
barriers in the distribution of leptin into and out of the rat brain,
Neuroscience 123 (2004) 527–536.
[38] P. Lambert, P. Couceyro, K. McGirr, S. Dall Vechia, Y. Smith, M.
Kuhar, CART peptides in the central control of feeding and
interactions with neuropeptide Y, Synapse 29 (1998) 293–298.
[39] B. Li, B. Xu, N. Rowland, S. Kalra, c-Fos expression in the rat brain
following central administration of neuropeptide Y and effects of food
consumption, Brain Res. 665 (1994) 277–284.
[40] H. Lin, E. Decuypere, J. Buyse, Oxidative stress induced by cortico-
sterone administration in broiler chickens (Gallus gallus domesticus),
short-term effect, Comp. Biochem. Physiol. 139 B (2004) 745–751.
[41] T.M. Loftus, D.E. Jaworsky, G.L. Frehywot, C.A. Townsend, G.V.
Ronnett, M.D. Lane, F.P. Kuhajda, Reduced food intake and body
weight in mice treated with fatty acid synthase inhibitors, Science 288
(2000) 2379–2381.
[42] M. Lopez, L. Seoane, M.C. Garcia, F. Lago, F.F. Casanueva, R.
Senaris, C. Dieguez, Leptin regulation of prepro-orexin and orexin
receptor mRNA levels in the hypothalamus, Biochem. Biophys. Res.
Commun. 269 (2000) 41–45.
[43] D. Lu, D. Willard, I.R. Patel, S. Kadwell, L. Overton, T. Kost, M.
Luther, W. Chen, R.P. Woychik, W.O. Wilkison, R.D. Cone, Agouti
protein is an antagonist of the melanocyte-stimulating hormone
receptor, Nature 371 (1994) 799–802.
[44] M. Maffei, J. Halaas, E. Ravussin, R.E. Pratley, G.H. Lee, Y. Zhang,
H. Fei, S. Kim, R. Lallone, S. Ranganathan, P.A. Kern, J.M. Friedman,
Leptin levels in human and rodent: measurement of plasma leptin and
ob RNA in obese and weight-reduced subjects, Nat. Med. 1 (1995)
1155–1161.
[45] H. Makimura, T.M. Mizuno, X.J. Yang, J. Silverstein, J. Beasley, C.V.
Mobbs, Cerulenin mimics effects of leptin on metabolic rate, food
intake, and body weight independent of the melanocortin system, but
unlike leptin, cerulenin fails to block neuroendocrine effects of fasting,
Diebetes 50 (2001) 733–739.
[46] J.P. McMurtry, C.M. Ashwell, D.M. Brocht, T.J. Caperna, Plasma
clearance and tissue distribution of radiolabeled leptin in the chicken,
Comp. Biochem. Physiol. 138 (2004) 27–32.
[47] B. Meister, Control of food intake via leptin receptors in the
hypothalamus, Vitam. Horm. 59 (2000) 265–304.
[48] J.G. Mercer, N. Hoggard, L.M.Williams, C.B. Lawrence, L.T. Hannah,
P. Trayhurn, Localization of leptin receptor mRNA and the long form
splice variant (ob-Rb) in mouse hypothalamus and adjacent brain
regions by in situ hybridization, FEBS Lett. 387 (1996) 113–116.
[49] T.M. Mizuno, C.V. Mobbs, Hypothalamic agouti-related protein
messenger ribonucleic acid is inhibited by leptin and stimulated by
fasting, Endocrinology 140 (1999) 814–817.
[50] T.M. Mizuno, S.P. Kleopoulos, H.T. Bergen, J.L. Roberts, C.A. Priest,
C.V. Mobbs, Hypothalamic proopiomelanocortin mRNA is reduced by
fasting and corrected in ob/ob and db/db mice, but is stimulated by
leptin, Diabetes 47 (1998) 294–297.
[51] H. Munzberg, L. Huo, E.A. Nillni, A.N. Hollenberg, C. Bjorbek, Role
of signal transducer and activator of transcription 3 in regulation of
hypothalamic proopiomelanocortin gene expression by leptin, Endo-
crinology 144 (2003) 2121–2131.
[52] T. Ohkubo, T. Boswell, S. Lumineau, Molecular cloning of chicken
prepro-orexin cDNA and preferential expression in the chicken
hypothalamus, Biochim. Biophys. Acta 1577 (2002) 476–480.
[53] B. Olson, M. Drutarosky, M. Chow, V. Hruby, E. Stricker, J. Verbalis,
Oxytocin and an oxytocin agonist administered centrally decrease
food intake in rats, Peptides 12 (1991) 113–118.
[54] M.A. Pelleymounter, M.J. Cullen, M.B. Baker, R. Hecht, D. Winters,
T. Boone, F. Collins, Effects of the obese gene product on body weight
regulation in ob/ob mice, Science 269 (1995) 540–543.
[55] D. Qu, D.S. Ludwig, S. Gammeltoft, M. Piper, M.A. Pelleymounter,
M.J. Cullen, W.F. Mathes, R. Przypek, R. Kanarek, E. Maratos-Flier,
A role for melanin-concentrating hormone in the central regulation of
feeding behavior, Nature 380 (1996) 243–247.
[56] K. Rahmouni, W.G. Haynes, D.A. Morgan, A.L. Mark, Role of
melanocortin-4 receptors in mediating renal sympathoactivation to
leptin and insulin, J. Neurosci. 23 (2003) 5998–6004.
[57] N. Raver, M. Taouis, S. Dridi, M. Derouet, J. Simon, B. Robinzon, J.
Djiane, A. Gertler, Large-scale preparation of biologically active
recombinant chicken obese protein (Leptin), Protein Expression Purif.
14 (1998) 403–408.
[58] M.P. Richards, M.C. Ashwell, J.P. McMurtry, Analysis of leptin gene
expression in chicken using reverse transcription polymerase chain
reaction and capillary electrophoresis with laser-induced fluorescence
detection, J. Chromatogr., A 853 (1999) 321–335.
[59] F.L. Rock, S.W. Altmann, M. Vanheek, R.A. Kastelein, J.F. Bazon,
The leptin haemopoietic cytokine fold is stabilized by an intrachain
disulfide bond, Horm. Metab. Res. 28 (1996) 649–652.
[60] M. Rossi, M. Kim, D. Morgan, A C-terminal fragment of agouti-
related protein increases feeding and antagonizes the effect of alpha-
melanocyte stimulating hormone in vivo, Endocrinology 139 (1998)
4428–4431.
[61] T. Sakurai, A. Amemiya, M. Ishii, I. Matsuzaki, R. Chemelli, H.
Tanaka, S. Williams, J. Richardson, G. Kozlowski, S. Wilson, J. Arch,
R. Buckingham, A. Haynes, S. Carr, R. Annan, D. McNulty, W. Liu, J.
Terrett, N. Elshourbagy, D. Bergsma, M. Yanagisawa, Orexins and
orexin receptors: a family of hypothalamic neuropeptides and G
protein-coupled receptors that regulate feeding behavior, Cell 92
(1999) 573–585.
[62] M.W. Schwartz, R.J. Seeley, L.A. Campfield, P. Burn, D.G. Baskin,
Identification of targets of leptin action in rat hypothalamus, J. Clin.
Invest. 98 (1996) 1101–1106.
[63] M.W. Schwartz, R.J. Seeley, S.C. Woods, D.S. Weigle, L.A. Camp-
field, P. Burn, D.G. Baskin, Leptin increases hypothalamic proopio-
melanocortin mRNA expression in the rostral arcuate nucleus,
Diabetes 46 (1997) 2119–2123.
[64] M.W. Schwartz, S.C. Woods, J.D. Porte, R.J. Seeley, D.G. Baskin,
Central nervous system control of food intake, Nature 404 (2000)
661–671.
[65] R.J. Seeley, S.C. Woods, Monitoring of stored and available fuel by
the CNS: implications for obesity, Nat. Rev., Neurosci. 4 (2003)
901–909.
[66] M. Shimada, N. Tritos, B.B. Lowell, J.S. Flier, E. Maratos-Flier, Mice
lacking melanin concentrating hormone are hypophagic and lean,
Nature 396 (1998) 670–674.
[67] T. Shimokawa, M.V. Kumar, M.D. Lane, Effect of a fatty acid
synthase inhibitor on food intake and expression of hypothalamic
neuropeptides, Proc. Natl. Acad. Sci. U. S. A. 99 (2002) 66–71.
[68] T. Shimokawa, M.V. Kumar, M.D. Lane, Effect of a fatty acid
synthase inhibitor on food intake and expression of hypothalamic
neuropeptides, Proc. Natl. Acad. Sci. U. S. A. 99 (2002) 66–71.
[69] M.K. Sinha, I. Opentanova, J.P. Ohannesian, J.W. Kolaczynski, M.L.
Heiman, J. Hale, G.W. Becker, R.R. Bowsher, T.W. Stephens, J.F.
Caro, Evidence of free and bound leptin in human circulation, J. Clin.
Invest. 98 (1996) 1277–1282.
[70] M. Spina, E. Merlo-pich, R. Chan, A. Basso, J. Rivier, W. Vale, G.

S. Dridi et al. / Brain Research 1047 (2005) 214–223 223
Koob, Appetite-suppressing effects of urocortin, a CRF-related
neuropeptide, Science 273 (1996) 1561–1564.
[71] B. Stanley, S. Leibowitz, Neuropeptide Y injected in the para-
ventricular hypothalamus: a powerful stimulant of feeding behavior,
Proc. Natl. Acad. Sci. U. S. A. 82 (1985) 3940–3943.
[72] T. Tachibana, K. Sugahara, A. Ohgushi, R. Ando, S. Kawakami, T.
Yoshimatsu, M. Furuse, Intracerebroventricular injection of agouti-
related protein attenuates the anorexigenic effect of alpha-melanocyte
stimulating hormone in neonatal chicks, Neurosci. Lett. 305 (2001)
131–134.
[73] S. Takeuchi, S. Takahashi, Melanocortin receptor genes in the chicken-
tissue distributions, Gen. Comp. Endocrinol. 112 (1998) 220–231.
[74] S. Takeuchi, H. Suzuki, M. Yabuuchi, S. Takahashi, A possible
involvement of melanocortin 1-receptor in regulating feather color
pigmentation in the chicken, Biochim. Biophys. Acta 1308 (1996)
164–168.
[75] S. Takeuchi, H. Suzuki, S. Hirose, M. Yabuuchi, C. Sato, H.
Yamamoto, S. Takahashi, Molecular cloning and sequence analysis
of the chick melanocortin 1-receptor gene, Biochim. Biophys. Acta
1306 (1996) 122–126.
[76] S. Takeuchi, T. Kudo, S. Takahashi, Molecular cloning of the chicken
melanocortin 2 (ACTH)-receptor gene, Biochim. Biophys. Acta 1403
(1998) 102–108.
[77] S. Takeuchi, S. Takahashi, A possible involvement of melanocortin 3
receptor in the regulation of adrenal gland function in the chicken,
Biochim. Biophys. Acta 1448 (1999) 512–518.
[78] M. Taouis, J.W. Chen, C. Daviaud, J. Dupont, M. Derouet, J. Simon,
Cloning the chicken leptin gene, Gene 208 (1998) 239–242.
[79] J.E. Thornton, C.C. Cheung, D.K. Clifton, R.A. Steiner, Regulation of
hypothalamic proopiomelanocortin mRNA by leptin in ob/ob mice,
Endocrinology 138 (1997) 5063–5066.
[80] Y. Uehara, H. Shimizu, K.I. Ohtani, N. Sato, M. Mori, Hypothalamic
corticotrophin-releasing hormone is a mediator of the anorexigenic
effect of leptin, Diabetes 47 (1998) 890–893.
[81] S.C. Woods, R.J. Seeley, J.D. Porte, M.W. Schwartz, Signals that
regulate food intake and energy homeostasis, Science 280 (1998)
1378–1383.
[82] Y. Zhang, R. Proenca, M. Maffei, M. Barone, J.M. Friedman,
Positional cloning of the mouse obese gene and its human homologue,
Nature (London) 372 (1994) 425–432.





























