Embed Size (px)
Citation preview

Aquaculture 127 (1994) 131-144
Mitochondrial DNA variation and markers of species identity in two penaeid shrimp species:
Penaeus monodon Fabricius and P. japonicus Bate
Didier Bouchon”, Catherine Souty-Grosset, Roland Raimond Vniversife’ de Poifiers, Laboratoire de Biologic Animale, VRA CNRS 1452. G&&ique et Biologic des
Populations de Crustacb, 40 avenue du Recteur Pineau, F-86022 Poitiers cedex, France
Accepted 28 June 1994
Abstract
Mitochondrial DNA of three laboratory strains of Penaeus nwnodon from Malaysia, Australia and Fiji and one farm population of P. japonicus were analysed. The molecule size of the mtDNA of the two penaeid species was similar and close to 16 000 base pairs. The polymerase chain reaction method provided molecular markers of species identity which showed a high interspecific diversity. According to RFLP data in P. monodon, a genetic divergence of 1.68% was found between the Fiji strain and the Australia/Malaysia strains. This result yielded new genetic data about the Indo-West Pacific biogeographical region. A restriction map of the mtDNA of the Fiji strain was established and the small ribosomal subunit gene region was located. The farm population of P. japonicus was structured but, because its parental origin was unkown, it was not possible to determine intraspecific variation.
Keywords: Penaeus monodon; Penaeusjaponicus; Mitochondrial DNA; Geneticvariation; Ribosomal RNA genes; RFLP; PCR
1. Introduction
Penaeid shrimps are an important part of the aquaculture industry in many areas of the world. Although most of the annual domestic yield of penaeid shrimp comes from the harvest of wild shrimp, increasing emphasis is being placed on pond and raceway culture.
Advances in laboratory control of maturation (AQUACOP 1975, 1979) have allowed the selection of populations for commercial production.
Knowledge of the structure of the populations from which the founder individuals were
chosen is essential for maximum genetic improvement. By the 196Os, electrophoretic tech-
* Corresponding author. Tel. (33) 49 45 39 81; Fax (33) 49 45 40 15.
0044-8486/94/$07.00 0 1994 Elsevier Science B.V. All rights reserved SSDIOO44-8486(94)00185-5

132 D. Bouchon et al. /Aquaculture 127 (1994) 131-144
niques for protein separation allowed geneticists to survey natural populations in Crustacea. Data from 97 crustacean species showed low levels of polymorphism (Hedgecock et al.,
1982): the mean heterozygosity was 0.073 in the Crustacea as a whole, and it was lower (0.05 1) in Penaeoidea and Caridea. This technique was unable to detect neutral mutations, which resulted in a systematic underestimation of mean heterozygosity.
Recently, molecular genetic techniques have created new opportunities for the charac- terization of marine animal populations from the study of mitochondrial DNA (mtDNA)
(Avise, 1985). The transmission of mtDNA is usually maternal and non-recombinational
and this type of DNA has a rapid rate of evolution. Consequently, an analysis of restriction fragment length polymorphism (RPLP) provides a high-resolution technique to study the
microevolutionary processes within species. According to the review by Ovenden ( 1990), studies on mtDNA and marine stock assessment are mainly related to fish and only a few
articles are about Crustacea. In marine crustaceans, Batuecas et al. ( 1988) mapped out the
genome organization of brine shrimp (Artemiu) mtDNA in which the order of genes is
similar to the completely sequenced mtDNA of Drosophila yukubu (Clary and Wolsten-
holme, 1985). Studies on natural populations were performed on the spiny lobster, Punulirus argus (Komm et al., 1982; McLean et al., 1983)) and have provided a preliminary subdi- vision of the species. The most complete study was made on the palinurid rock lobster.
RPLP from 5 Jusus species revealed that intraspecific mtDNA nucleotide sequence diversity is generally high (Brasher et al., 1992). Recently Palumbi and Benzie (1991) reported surprisingly large mtDNA differences among morphologically and ecologically similar
species of penaeid shrimps, Penueus vunnumei, P. stylirostris, P. esculentus and Metupen- ueus enduevori, by sequencing amplified mtDNA fragments. They found about 18% dif- ference between subgenera of Penueus and 7.4% within the subgenera Litopenueus. In the
same way, Machado et al. ( 1993) have found a 11% nucleotide divergence between the studied parts of the 16s rDNA genes in P. notiuEis and P. schmitti . Benzie et al. ( 1993)
reported high variability in mtDNA relative to that of allozymes in P. monodon Australian
populations. The scarcity of papers on crustaceans is probably related to the difficulty of extracting
mtDNA. The most commonly used methods are very time-consuming with a low yield, and
they are therefore unsuitable for mtDNA studies in population genetics and evolutionary biology. To overcome this problem, the use of the polymerase chain reaction (PCR) method, which can amplify DNA sequences, could be necessary. DNA amplification and restriction enzyme digestion of portions of mitochondrial genes have provided molecular markers of
species identity for three species of the copepod genus, Culunus (Bucklin and Kann, 1991) . In the same way, Chow et al. ( 1993) have reported that PCR-RFLP analysis was a useful simple method for investigating genetic stock structure in 13 western Atlantic snappers.
Here we used a rapid extraction method which corresponds to a modified version of alkaline lysis commonly employed to extract plasmids (Birnboim and Doly, 1979; Ish- Horowitz and Burke, 1981). This method adapted by Souty-Grosset et al. ( 1992) in the terrestrial crustacean, Armudillidium vulgure, revealed genetic divergence between conspe- cific populations (Grandjean et al., 1993). mtDNAs of two commercially important shrimps (P. monodon and P. juponicus) were characterized by the description of the molecule (length and restriction mapping). Molecular tools were designed for analyzing the diversity between three domestic P. monodon strains and for providing markers of species identity

D. Bouchon et al. /Aquaculture 127 (1994) 131-144 133
between P. monodon and P. japonicus. The results are discussed on the basis of what is currently known on metazoan mitochondrial genomes.
2. Materials and methods
Prawn strains Two domestic populations of commercial shrimps were collected:
( 1) The giant tiger prawn, Penaeus monodon Fabricius 1798, is one of the major aquaculture species in Asia. This marine shrimp is widely distributed from East and Southeast Africa, throughout the Red Sea and Arabian Gulf, around the Indian subcontinent, and throughout the Malay Archipelago to Northern Australia and Japan. Three laboratory strains were used
from the AQUACOP, IFREMER, Tahiti (Aquacop, 1979). Six adult females originating
from Malaysia, Australia and Fiji were collected. (2) The kuruma prawn, Penaeus japonicus Bate 1888, has recently become a farm-grown
shrimp in Southern Europe. It is a marine shrimp widely ranging from South Africa to the
Red Sea throughout the entire Malay Archipelago to Korea, Japan and northern Australia. The species has migrated through the Suez Canal into the Mediterranean and is now caught off southern Turkey. Twenty juvenile females (about 6 months old) were sampled from
the CREAA French sea farm (Centre Regional d’Etudes et d’Applications Aquacoles,
OlCron Island, France). Because the broodstock was obtained from a commercial hatchery,
the origin of individuals was unknown and this sample was not genetically controlled.
DNA extraction Muscles, ovaries and egg masses of each P. monodon female sampled were collected and
frozen in liquid nitrogen. In the absence of egg masses in juvenile P. japonicus females,
muscles and ovaries only were collected in the same way. Because their smaller sizes 5
females were pooled in a sample; 1200 mg of tissues were ground in liquid nitrogen with mortar and pestle, and the powder was homogenized in 6 volumes of ice-cold NET buffer
(30 mM Tris-HCl, 30 mM EDTA, 15 mM NaCl, pH 7.8) containing proteinase K (100
pug/ml). Two volumes of freshly prepared solution of SDS l%, 0.2 N NaOH were added. The sample was then incubated at 0°C for 10 min and homogenized at intervals; 1.5 volumes
of ice-cold potassium acetate (potassium 3M, acetate 5M) were added. The sample was stored on ice for 10 min. The homogenate was then centrifuged at 12 000 X g for 10 min.
The mtDNA was extracted twice with buffer-equilibrated phenol and with chloroform/
isoamyl alcohol (24: 1). mtDNA was precipitated by the addition of 0.8 volume of isopro- panol and 0.1 volume of sodium acetate, stored overnight at -2o”C, and pelleted at
1.5 000 X g for 30 min. Washed mtDNA in 70% ethanol was vacuum dried and resuspended in 1 ~1 of TE buffer for 7 mg of starting tissue.
Amplljication of mitochondrial12S and 16s rDNA genes DNA extraction was the same as outlined above. The polymerase chain reaction was
carried out in a final volume of 50 ~1 in a reaction mixture described by Kocher et al. ( 1989). This mixture included all 4 dNTPs (each at 200 PM), 600 nM of primer concen- tration, and 2 units of Taq polymerase (Promega Corporation, Madison, USA). The poly-

134 D. Bouchon et al. /Aquaculture I27 (1994) 131-144
merase buffer, containing 1.5 mM MgCI,, is given by the manufacturer. The reaction mixture is covering by a volume of 60 ~1 of mineral oil (Sigma Chemical Company, St Louis, USA). The two pairs of primers that targeted the mitochondrial small ( 12s) and large sub-
unit ( 16s) of ribosomal DNA genes were described by Simon et al. ( 1991) and designed
to be universal in insects. The nucleotide sequence of each set of primers was as follows:
12SrDNA, 5’-AAACTAGGATTAGATACCCTATTAT-3’ and 5’-AAGAGCGA-
CGGGCGATGTGT-3’; 16SrDNA, 5’-CGCCTGTITAACAAAAACAT-3’ and 5’-
CCGGTCTGAACTCAGATCATGT-3’.
The PCR was carried out on a TRIO-Thermoblock (Biometra GmbH, Gottingen, Ger-
many) thermocycler. The thermal program was 93°C for 2 min followed by 30 cycles of
93°C for 30 s, 50°C for 30 s, 72°C for 45 s and 72°C for 5 min as a final extension after the
last cycle. Electrophoresis of 5 ~1 of the amplified mixture was performed in a 2% agarose
gel in TBE buffer (90 mM Tris-boric acid and 2 mM EDTA) and the DNA was stained
with ethidium bromide. The size of fragment amplified was estimated in comparison with
a size standard (Marker MVI, Boehringer, Mannheim, Germany).
Restriction endonuclease digestion Total mtDNA from the three P. monodon strains were separately digested with 12
enzymes (GIBCO BRL, Life Technologies) ; 9 of them were 6-base cutters: BamHI, BgZII,
ClaI, EcoRV, PA, PvuII, SacI, S&I and StyI. Two were 4-base cutters (HhaI and h4spI)
and one a 7-base cutter (EcoO109). The mtDNA of P. japonicus population was digested
with 5 endonucleases: BamHI, BgZII, CZaI and Hi&III were 6-base cutters and HhaI was a
4-base cutter. All digests were carried out in 12 ~1 volumes in reaction conditions as specified
by the supplier. After 1 h of incubation the samples were mixed with a gel-loading buffer
(54% Ficoll, 10% electrophoresis buffer, 14% bromophenol blue, 20% EDTA 0.5 M, 2%
SDS). The digested mtDNA was electrophoresed on a 1.2% agarose gel in TEP buffer ( 80
mM Tris-phosphate, 2 mM EDTA, pH 8.0) for 15 h at 30 V. After staining with Ethidium
Bromide (0.5 mg/ml of electrophoresis buffer) the fragments were visualized with an UV
light transilluminator at 302 nm. DNA bands were photographed and negatives were ana-
lyzed with an image-processing system. The program (Lecphor, BIOCOM SA) calculates
the molecular weight of each fragment on the basis of cubic spline interpolation between
mobility and size of the chosen molecular weight standard. The standard used to estimate the fragment sizes was lambda-phage DNA (Boehringer, Mannheim) digested by Hi&III.
Amplified portions of 12s and 16s rDNA from the three strains of P. monodon and the
population of P. japonicus were digested by 29 enzymes: 2 were 7-base cutters, 21 were 6-
base cutters and 7 were 4-base cutters. The digested mtDNA was electrophoresed on a 4% NuSieve (3: 1) (FMC BioProducts, Rockland, ME) agarose gel in TAE buffer (40 mM
Tris base, 20 mM acetic acid, 2 mM EDTA, pH 8.4) for 6 h at 80 V. The size of fragment restricted was estimated in comparison with size standards (HaeIII and MspI digests of
pBR322 DNA) in the same way as for total mtDNA analysis.
Restriction site mapping mtDNA restriction sites for single and double digests of all combinations ofAcc1, BamHI,
CZaI, EcoRV, PvuII and Sac1 were mapped for the P. monodon Fiji strain.

D. Bouchon etal. /Aquaculture 127 (1994) 131-144 135
Estimation of genetic distance in P. monodon strains The degree of genetic divergence between mtDNA of P. monodon strains was estimated
with the restriction fragment method of Nei and Li (1979) and Nei and Tajima (1981). Fragments are pooled for all enzymes with the same number of bases per restriction site (i.e. 4-cutters, 6-cutters and 7-cutters) and a parametric distance (d) between mtDNA for enzyme types by the formulae given by Nei and Li ( 1979). To estimate d the parameter S is first estimated by:
sij = 2mij mi +??lj
where mi and mj are the numbers of restriction fragments in DNA sequences x and y, respectively, whereas mij is the number of fragments shared by the two sequences. The number of nucleotide substitutions per site d can be estimated by:
Jij = - Ln Sij
r
where r is the number of bases per restriction site (Nei and Li, 1979). When different kinds of enzymes with different r-values are used, the mean number of nucleotide substitutions can be estimated by the formula given by Nei and Tajima ( 198 1) :
where mk = f%(k) + mj(k)
2
and k refers to the kth class of restriction enzymes. Summation is taken over all different enzyme classes.
3. Results
mtDNA extraction from muscles was not very productive. Because of the high number of mitochondria in that tissue one might expect a good yield, but homogenization of samples was a major problem even when we increased the volume ratio of NET buffer. Preparations of mtDNA of shrimp muscles were thus highly contaminated with genomic DNA. Ovaries and eggs gave much better results (e.g. in Fig. 1). At least 0.5 pg of mtDNA must be used per digestion to ensure the detection of fragments. Our yields of mtDNA from ovaries were usually 7 pg/ g tissue. Thus 1.2 g of starting material provide enough DNA for 17 digests. Because the extraction yield method was significantly higher with egg masses, up to 26

136 D. Bouchon et al. /Aquaculture 127(1994) 131-144
M 1 2 3 4 5 6 M 7 8 9 10 11
kb
23.13
9.42
6.63
2.32
2.03
0.57
Fig. 1. Example of ethidium bromide stained agarose gel of mtDNA digestion patterns of P. monodon Fiji strain.
On the left side are given fragment sizes in kilobase pairs (kb) of Lambda-phage DNA digested by HindIII. Lanes: M = size standard, I= Std. 2 = BamHIIClaI, 3 = Sac IIBamHI, 4 = EcoRVIPuuII, 5 = Barn HIIPuuII,
6 = SacI/PuuII, 7 = Cla I/PouII, 8 = ClaIIEcoRV, 9 = Eco RVIBamHI, 10 = ClaIISacI, 11= Sac IIEcoRV.
restriction endonucleases at best could be tested in the Fiji sample. Nevertheless, a reduction
to 10 digests per sample has been detected in the Malaysia and Australia strains probably due to storage problems during transport from Tahiti to France. P. juponicus females had
the lowest extraction yield, probably because of immature ovaries.
Mitochondrial DNA restriction analysis
P. monodon samples. The fragment sizes of all digestions are given in Table 1. The size of the giant tiger prawn mtDNA is almost 16 kilobase pairs (15.83 +0.16 kb).
Of the 12 endonucleases tested in the three strains, 9 were monomorphs: EcoO109 EcoRV and Sac1 linearized the molecule while BarnHI, Clu I, PuuII, StuI gave two fragments. Sty1 and MspI produced respectively 4 and 6 fragments. Three restriction enzymes exhibited polymorphism according to the geographical origins of the strains: the Fiji strain was
D. Bouchon et al. /Aquaculture 127 (1994) 131-144 137
Table 1
Calculated size of P. monodon mtDNA fragments (kilobase pairs) obtained after digestion with restriction enzymes
Eco0109 EcoRV Sac1 BarnHI C&I PvuII StuI SI.YI MspI BgrII BgLII HhaI HhoI PSfI PstI
No difference in fragment pattern for all strains Fiji MaLAu Fiji Mal.Au Fiji Mal.Au
11.25 15.75 15.45 9.35 9.30 8.60 10.15 9.20 5.85 4.15 4.85 dnc 15.25 1 I .90 15.65
_ 7.10 6.80 7.80 6.20 3.35 2.85 3.30 3.95 2.33 -
_ _ I .95 2.50 3.20 3.20 1.65 -
_ _ _ 0.80 1.90 1.95 1.95 _ _ _ 1.60 1.55 1.15 _ -
_ _ 0.90 1.10 - _ _
Total 17.25 15.75 15.45 16.45 16.10 16.40 16.35 15.20 15.50 15.25 15.10 dnc 15.25 15.90 15.65
Fiji = Fiji strain; Ma1 = Malaysian strain; Au = Australian strain; dnc = did not cleave.
Table 2
Calculated size of P. japonicus mtDNA fragments (kilobase pairs) obtained after digestion with restriction
enzymes
BamHI B&l ClaI HhaI HindIll
9.45 17.35 9.50 7.30 17.95 4.60 _ 6.70 5.30 _
2.40 _ 2.85 _
Total 16.45 17.35 16.20 15.45 17.95
Table 3 Calculated sizes of P. monodon mtDNA fragments (kilobase pairs) of theFiji strain obtained after single-digestion
with AccI and several double-digestions
AccI AccIIClaI AccIIEcoRV AccI/SacI AccIIBamHI AccI/PvulI
5.95 3.50 3.50 3.25 2.75 2.65 2.20 2.55 0.80 2.20 0.70 0.80
0.70 _ 0.25
5.25 6.00 5.75 4.75
3.50 3.05 3.50 2.75
2.75 2.75 2.75 2.75
2.20 2.15 1.80 2.20
0.80 0.80 0.80 1.20
0.70 0.70 0.70 0.80
0.70 0.45 0.40 0.75 _ 0.20 0.70
Total 15.90 15.90 15.90 15.90 15.90 15.90
distinguished in all cases, whereas the Malaysia and Australia strains were monomorphs. The differences were related to 6 fragments for BglZI in Fiji strain against 5 in both Malaysia and Australia strains. Moreover, only two identical fragments were recognized in the two observed profiles ( 3.20 and 1.95 kb, see Table 1) . Concerning HhaI, there was no restriction site on mtDNA of Fiji shrimps while the molecule was linearized in both Malaysia and

138 D. Bouchon et al. /Aquaculture 127 (1994) 131-144
Act I Barn HI
Act I
Act I -12 4-
Cla I
8 I
Fig. 2. Restriction map of the mitochondrial genome of the Fiji strain of Penaeus monodon. The stippled area represents the presumptive localization of the amplified portion of the small ribosomal subunit. Each graduation of the circle indicates 1 kb portion. The total length of the molecule is 16 kb.
Australia shrimps. PsrI cut the mtDNA of Fiji into three fragments, whereas it linearized the molecule of Malaysia and Australia strain.
P. japonicus samples. Owing to the pooled samples of P. japonicus only 5 restriction
morphs were clearly interpretable. In all other cases mtDNA bands could not be unambig-
uously identified because two or more profiles in a sample were superimposed. The fragment sizes of all digestions of mtDNA are given in Table 2. The mean size of the mtDNA of P. japonicus is 16.68 f 0.44 kb. BamHI and HhaI produced three fragments. BgLII and Hi&III linearized the molecule. CZaI produced two fragments.
Genetic distances between the three strains of P. monodon According to the formulae of Nei and Tajima ( 1981)) the percentage of divergence
between Fiji and Malaysia-Australia strains was 1.68%.
D. Bouchon et al. /Aquaculture 127 (1994) 131-144 139
Ml 1 23 4 56 M2
bp
622
527
404
309
242
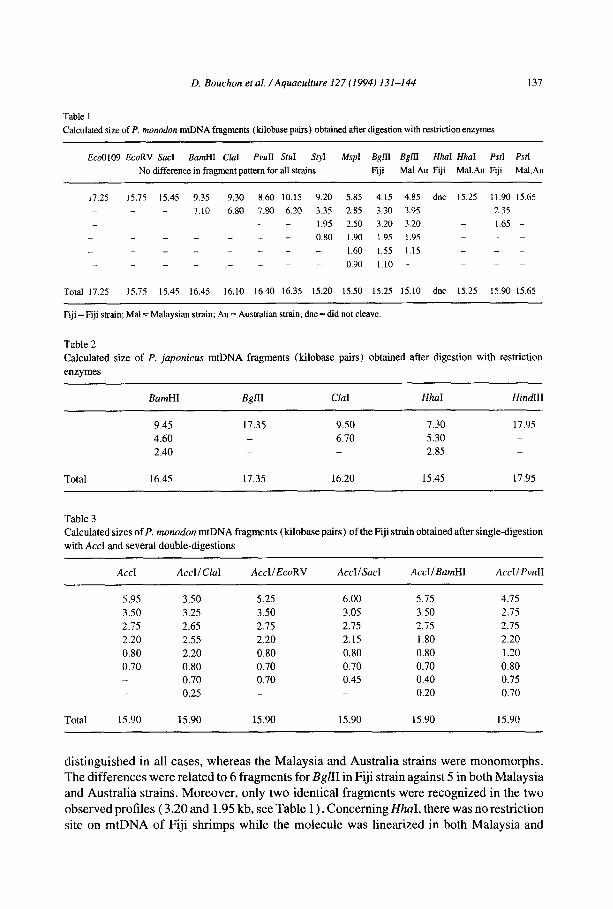
Fig. 3. Restriction profiles of 12s rDNA gene fragments of P. monodon (Pm) or P. japonicus (Pj) mtDNA
amplified by PCR. On the right side are given some fragment sizes in base pairs (bp) of pBR322 DNA digested
by HaeIII. On the left side are given some fragment sizes in base pairs (bp) of pBR322 DNA digested by Mspl. Lanes: M, = HaeIII digest of pBR322; 1 and 2 = Pm and Pj digested by DraI; 3 = Pm, no digestion by BsrEII:
4 = Pj digested by BsfEII; 5 = Pm, no digestion by SspI; 6 = Pj digested by Ssp I; MZ = MspI digest of pBR322.
Restriction site mapping We have determined the restriction sites for 5 enzymes (SacI, EcoRV, BamHI, CZaI, and
PuuII) which were monomorph in the three P. monodon strains (see Table 1 and Fig. 1).
Table 4
Calculated fragment sizes in numbers of base pairs resulting from restriction enzyme digestion of the 415-bp 12s
rDNA amplification product for samples of P. japonicus and P. monodon strains
Enzyme Recognition site P. monodon
DraI TTTlAAA 361 -t 48 BstEll GlGTnACC dnc ssp I AATtATT dnc
dnc = did not cleave; A = adenine; C = cytosine; G = guanine; T = thymine; n = A, C, C or T.
P. japonicus
235t 180
338 +77
339 + 76
140
Fig. 4 Restriction profiles of 16s rDNA gene fragments of P. monodon (Pm) or P. japonicus (Pj) mtI amplif ied by PCR. Same DNA standards as Fig. 3. Lanes: 1 and 2 = Pm and Pj digested by DraI; 3 = Pm dige by AC< :I; 4 = Pj, no digestion by Accl; 5 = Pm digested by Mbo I; 6 = Pj digested by AluI; 7 = Pm digested by J 8=Pj, no digestion by SspI; 9 = Pm, no digestion by AM; lO= Pj, no digestion by MboI.
Table Calcul rDNA
D. Bouchon et al. /Aquaculture 127 (1994) 131-144
Mt Nl2 1 23456 78 9 10
5 ated fragment sizes in numbers of base pairs resulting from restriction enzyme digestion of the 520-pb amplification product for samples of P. japonicus and P. monodon strains
rNA sted :sp1;
16s
Enzyme Recognition site P. monodon P. japonicus
DraI Accl Mb01 SspI AhI Hi&II I1
TTTIAAA GT/mkAC IGATC AATI ATT AG/CT A/ AGCTT
500+20* 429+91 358 + 162 327 + 193 dnc dnc
317+203 dnc dnc dnc 351+ 169 291+ 229
Fragments marked with asterisks were not observed but assumed under the criterion of minimum mutational steps involved in fragment changes. dnc = did not cleave; A = adenine; C = cytosine; G = guanine; T = thymine; m=AorC;k=GorT.

D. Bouchon etai. /Aqclacutture I21 (1994) 131-144 141
AccI was only tested in the Fiji strain and gave 6 fragments. The fragment sizes of some single and double digestions are given in Table 3. According to the 15 double-digestion
morphs, we had a better estimation of the molecular weight of the P. monodon mtDNA (mean value 15.90 kb). A restriction map of the Fiji mtDNA is shown in Fig. 2.
rDNA analysis: species-specific restriction site polymorphisms The size of the amplified portions of rDNA genes was 415 rf: 2 bp for the 12s and 520 + 6
bp for the 16X All the endonucleases tested in the three strains of P. monodon were monomorph.
Fig. 3 shows restriction-fragment patterns of the amplified portion of 12s rDNA gene.
DraI cleaved the 4 15 bp amplification product into two fragments for samples of P. monodon and P. japonicus. The sizes of the fragments (i.e. position of the restriction site) were
unique in each sample (Table 4). BstEII and SspI cleaved the P. japonicus gene portion
into two fragments, but did not cleave the gene portion of P. monodon strains. AccI, AccII,
AU, BgZII, CZaI, RsaI and TaqI profiles were monomorph in the two species and produced two fragments. The 19 remaining restriction enzymes did not cleave the 12s gene’s portion
in the two penaeid shrimps. Fig. 4 shows restriction-fragment patterns of the amplified portion of 16s rDNA gene.
DraI cleaved the 520 bp amplification product for samples of P. monodon and P. japonicus with two specific restriction sites (Table 5). The 16s gene portion was cleaved in two fragments by AccI, Mb01 and SspI for P. monodon strains whereas it was not cleaved for P. japonicus. AfuI and Hind III had no restriction site in the P. monodon amplification product
but one site in the P. japonicus product. HaeIII and SstII profiles were monomorph in the two species and produced two fragments. The 21 remaining restriction enzymes did not cleave the 16s gene’s portion in the two penaeid shrimps.
4. Discussion
By ultracentrifugation on CsCl-ethidium bromide density gradients, Benzie et al. ( 1993) obtained a yield of 10 digests per sample from unfrozen oocytes in P. monodon. In the
studies performed by Komm et al. ( 1982) and McLean et al. ( 1983) the mtDNA amounts
extracted were insufficient to survey genome variation (Ovenden, 1990). The average DNA yield (7 pg/g of eggs in the Fiji sample) of the method presented here was higher than
those using isolated mitochondria as starting material. Lansman et al. ( 1981) reported a yield of 3 pg/g tissue of rodents. Powell and Zuniga ( 1983) have obtained enough mtDNA
from 75 mg of tissues of Drosophila pseudoobscura to perform one digestion. The lowest
yield was obtained in P. japonicus samples, which suggests that there is a specificity of the extraction buffer and demonstrates the necessity of testing different tissues for optimalisa- tion. For example, when the same method is used with fresh ovaries, it gives sufficient amounts of mtDNA for at least 6-10 digests in the terrestrial isopod Armadillidium vulgare (Souty-Grosset et al., 1992); however, it does not work with egg masses.
The mtDNA of P. monodon and P. japonicus appears to be similar to that of other animal mtDNA. The molecular size of about 16 kb pairs is consistent with the size of other marine crustacean mtDNAs studied: Pant&us argus (Komm et al., 1982; McLean et al., 1983),

142 D. Bouchon et al. /Aquaculture 127 (1994) 131-144
Arfemia salina (Batuecas et al., 1988), Jasus spp. (Brasher et al., 1992), Penaeus notialis and P. schmitti (Monnerot et al., 1992). Despite an unexplained restriction morph with a
total mtDNA of 7.7 kb, Benzie et al. ( 1993) gave the same molecular weight in three P. monodon Australian populations.
Mapping of mtDNA in P. monodon can be used to determine site presence or absence in
various strains or wild populations from their restriction fragment profiles. Moreover, restriction site mapping provides information about region-specific rates of gene evolution.
We can now locate the 12s gene in the mtDNA (see Fig. 1): a 415 bp portion of this gene
is cut by either AccI or ClaI. According to the double digest fragments shown in Table 3, there is only one fragment ( = 242 bp) small enough to contain AccI and CZaI sites in a
sequence of at least 415 nucleotides long. This preliminary result provides the opportunity
to determine the gene arrangement and provides specific probes for future studies in penaeid shrimps.
Mitochondrial DNA has been widely used as a marker in population study; however, few works have been devoted to crustacean mtDNA. Previous studies which examined allozyme
variation report very low level subdivision of populations in penaeid shrimps (Mulley and
Latter 1980; Benzie et al., 1992; Sodsuk et al., 1992), and Mulley and Latter (1980) concluded that P. monodon had a homogeneous genetic structure over large geographical
areas. Moreover, Sunden and Davis (1991) have shown slightly lower levels of heterozy-
gosity and loss of rare alleles in an aquaculture population of P. uannamei . Marine species are known to have lower intraspecific mtDNA sequence diversity (i.e. lower than 1.0%)
than terrestrial or freshwater species (Ovenden, 1990). However, our survey of mitochon- drial variation in P. monodon strains has demonstrated significant genetic differentiation (divergence = 1.7%) between the Fiji population and the Malaysia and Australia popula-
tions. These two last strains were identical and if we suppose that there were sufficient endonucleases to reveal genome variation (e.g. we have found 3 informative enzymes to
separate Fiji from the two others), this result may reflect the absence of genetic subdivision. Using mtDNA genotype frequencies, Benzie et al. ( 1993) reported a significant differ-
ence between P. monodon wild populations from east and west Australian coasts but not
among the east coast populations. According to several restriction morphs (BarnHI, EcoRV
and SacI), the Australian strain studied here could be related to the east coast population. Sodsuk et al. ( 1992) found 5 differences in allele frequencies between populations of the
Gulf of Thailand (West Pacific) and of the Andaman Sea (East Indian Ocean). They concluded that this fact provides some evidence for isolation between the two seas. These two studies thus show a genetic isolation between Indian and Pacific shrimps. In addition,
our results tend to confirm a low level of subdivision among the populations in the extreme West Pacific (from Malay Archipelago to northern Australia). However, our results also show a structuration of populations within the Pacific Ocean. Therefore the Indo-West Pacific zoogeographical region could be divided into three genetically distinct P. monodon populations: Indian Ocean, extreme West Pacific and Melanesian Archipelago.
Because the shrimps sampled did not originate from natural populations but from com- mercially pooled brood stock, intraspecific variation could not be detected in P. japonicus. Nevertheless, several superimposed profiles suggest a genetic structure in this aquaculture population. Such results point out the importance of selecting broodstock from a wide genetic base using wild populations that have first been genetically determined.

D. Bouchon et al. /Aquaculture I27 (1994) 131-144 143
There is probably a high interspecific diversity in the penaeid shrimps because only CZuI
gave an identical restriction morph in the total mtDNA of P. monodon and P. japonicus. RFLP analysis of the amplified gene portions has provided molecular markers of species identity. We found 9 polymorphic and 8 monomorphic restriction sites in two mitochondrial genes (see Tables 4 and 5). By the maximum likelihood method the genetic divergence
between the two species is 8.7 +_ 3.7% for the 12s ribosomal gene and 24.0 &- 6.6% for the 16s ribosomal gene. The mean genetic divergence between P. monodon and P. juponicus is therefore 13.9 + 3.4%. These data are in the same range as those of Palumbi and Benzie
( 1991)) who have shown a divergence of 9.6% between P. vannamei and P. stylirostris by sequencing parts of the 125 ribosomal gene. Mulley and Latter ( 1980) examined 32 proteins
among 7 species of penaeid shrimps. Within the genera Penaeus, the measure of genetic similarity, based on data of 37 loci, is 0.65 f 0.08. They concluded that congeneric species
differ, on average, in at least one-third of their genes.
The present results suggest that P. monodon populations may be more structured than has been thought, in particular in the Indo-West Pacific region. Future aquaculture breeding
programs should be based on genetically differentiated populations available in the wild.
The P. japonicus case demonstrates the usefulness of the mtDNA technique in hatcheries
to determine the parental origin of larvae. Our study also shows that information could rapidly be obtained by using the polymerase chain reaction.
Acknowledgements
We thank A. Diter and all the staff of AQUACOP (IFREMER Tahiti) for obtaining P. monodon shrimps from their brood stock and Y. Zanette, manager of C.R.E.A.A. (Oltron, France), for providing P. japonicus samples. This work was supported in part by grant 89/
202 from C.O.R.D.E.T. (Commission de la Recherche dans les Departements et Territoires
d’Outre-Mer) .
References
AQUACOP, 1975. Maturation and spawning of penaeid shrimp: Penaeus merguiensis de Man, Penaeus japonicus Bate, Penaeus Qztecus Ives, Metapenaeus ensis de Haan and Penaeus semisulcatus de Haan. Proc. World
Mar&It. Sot., 6: 123-132.
AQUACOP, 1979. Penaeid reared brood stock: closing the life cycle of Penaeus monodon, Penaeus szylirostris and Penaeus oannamei. Proc. World Maricult. Sot., 10: 445452.
Avise, J.C., 1985. Identification and interpretation of mitochondrial DNA stocks in marine species. In: H.E.
Kumpf, R.N. Vaught, C.B. Grimes, A.G. Johnson and E.L. Nakamura (Editors), Proceedings of Stock
Identification Workshop, pp. 105-136.
Batuecas. B., Garesse, R., Calleja, M., Valverde, J.R. and Marco, R., 1988. Genome organization of Artemia mitochondrial DNA. Nucleic Acids Res., 16: 65 15.
Benzie, J.A.H., Frusher, S. and Ballment, E., 1992. Geographical variation in allozyme frequencies of Penaeus monodon (Crustacea: Decapoda) populations in Australia. Aust. I. Mar. Freshwater Res., 43: 715-725.
Benzie, J.A.H., Ballment,E. andFrusher, S., 1993. Geneticstructure of Penaeus monodon in Australia: concordant
results from mtDNA and allozymes. Aquaculture, 111: 89-93.
Bimboim, H.C. and Doly, J., 1979. A rapid alkaline extraction procedure for screening recombinant plasmid DNA.
Nucleic Acid Res., 7: 1513.

I44 D. Bouchon et al. /Aquaculture 127 (1994) 131-144
Brasher, D.J., Ovenden, J.R. and White, R.W.G., 1992. Mitochondrial variation and phylogeneticrelationships of Jams spp. (Decapoda: Palinuridae). J. Zool. Lond., 227: l-16.
Bucklin, A. and Kann, L., 1991. Mitochondrial DNA variation of copepods: markers of species identity and population differentiation in Culunus. Biol. Bull., 181(2): 357.
Chow, S., Clarke, M.E. and Walsh P.J., 1993. PCR-RFLPanalysis on thirteen western Atlantic snappers (subfamily Lutjaninae): a simple method for species and stock identification. Fish. Bull., 91(4): 619-627
Clary, D.O. and Wolstenholme, D.R., 1985. The mitochondrial DNA molecule of Drosophihyakuba: nucleotide sequence, gene organization, and genetic code. J. Mol. Evol., 22: 252-271.
Grandjean, F., Rigaud, T., Raimond, R., Juchault, P. and Souty-Grosset, C., 1993. Mitochondrial DNA polymor- phism and feminising sex factors dynamics in a natural population of Armadillidium m&are (Crustacea, Isopoda). Genetica, 92: 55-60.
Hedgecock, D., Tracey, M.L. and Nelson, K., 1982. Genetics. In: L.G. Abele (Editor), The Biology of Crustacea. Academic Press, New York, NY, pp. 28aO3.
Ish-Horowitz, D. and Burke, J.F., 1981. Rapid and efficient cosmid cloning. Nucleic Acids Res., 9: 2989. Kocher, T.D., Thomas, W.K., Meyer, A., Edwards, S.V., P&&o, S., Villahlanca, F.X. and Wilson, AC., 1989.
Dynamics of mitochondrial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA, 86: 6196-6200.
Komm, B., Michaels, A., Tsokos, J. and Linton, J., 1982. Isolation and characterization of the mitochondrial DNA from the Florida spiny lobster, Panulirus argus. Comp. Biochem. Physiol., 738,4: 923-929.
Lansman, R.A., Shade, R.O., Shapira, J.F. and Avise, J.C., 1981. The use of restriction endonucleases to measure mitochondrial DNA sequence relatedness in natural populations. III. Techniques and potential applications. J. Mol. Evol., 17: 214-226.
Machado, E.G., Dennebouy, N., Suarez, M.O., Mounolou, J.C. and Monnerot M., 1993. Mitochondrial I6S-rRNA gene of two species of shrimps: sequence variability and secondary structure. Crustaceana, 65( 3): 279-286.
McLean, M., Okubo, C.K. and Tracey, M.L., 1983. mtDNA heterogeneity in Panulirus nrgus. Experientia, 39: 536-538.
Monnerot, M., Garcia, M., Dennebouy, N., Oliva, M. and Mounolou, J.C., 1992. Mitochondrial DNA of Penaeus notiulis and Penaeus schmitti: characterization, diversity and sequencing. In: Abstracts, Fist European Crus- tacean Conference, p. 102.
Mulley, J.C. and Latter, B.D.H., 1980. Genetic variation and evolutionary relationships within a group of thirteen species of penaeid prawns. Evolution, 904-916.
Nei, M. and Li, W.H., 1979. Mathematical model for studying genetic variation in terms of restriction endonu- cleases. Proc. Natl. Acad. Sci. USA, 76: 5269-5273.
Nei, M. and Tajima. F., 1981, DNA polymorphism detectable by restriction endonucleases. Genetics, 97: 145- 163.
Ovenden, J.R., 1990. Mitochondrial DNA and marine stock assessment: a review. Aust. J. Mar. Freshwater Res., 41: 835-853.
Palumbi, S.R. and Benzie, J., 1991. Large mitochondrial DNA differences between morphologically similar Penaeid shrimp. Mol. Mar. Biol. Biotech., 1 ( 1) : 27-34.
Powell, J.R. and Zuniga, M.C., 1983. A simplified procedure for studying mtDNA polymorphisms. Biochem. Genet., 21(9/10): 1051-1055.
Simon, C., Franke, A. and Martin, A., 1991.The polymerase chain reaction: DNA extraction and amplification. In: G.M. Hewitt, A.W.B. Johnson and J.P.W. Young (Editors), Molecular Techniques in Taxonomy. NATO ASI Series, Vol. H57, pp. 329-355.
Sodsuk, S., McAndrew, B.J. and Penman, D.J., 1992. Genetic population structure of the giant tiger prawn (Penaeus monodon Fabricius, 1798) in the gulf of Thailand and the Andaman Sea. In: D. Penman, N.
Roongratri and B. McAndrew (Editors), AADCP Workshop Proceedings, pp. 161-165.
Souty-Grosset, C., Raimond, R. and Tourte, M., 1992. D6terminismeBpigenetiquedu sexe et divergence gen&ique de I’ADN mitochondrial chez Armadillidium vu&are Latr. (Crustac, Oniscdide): variabilite inter et intrapo- pulations. CR Acad. Sci. Paris, 314(Ser. III): 119-125.
Sunden, S.L.F. and Davis, S.K., 1991. Evaluation of genetic variation in a domestic population of Penaeus uannamei (Boone): a comparison with three natural populations. Aquaculture, 97: 131-142.





























