Embed Size (px)
Citation preview

Middle Cambrian molluscs of 'Australian' aspect from northern Siberia
ALEXANDER P. GUBANOV, ARTEM V. KOUCHINSKY, JOHN S. PEELAND STEFAN BENGTSON
GUnANOV, A.P., KOUCHINSKY, A.V., PnEL, J.S. &; BENGTSON, S., 30.4.2004. Middle Cambrian molluscs of 'Australian type' from northern Siberia. Alcheringa 28, 1-20. ISSN 0311- 5518.
An assemblage of silicified helcionelloid molluscs is described from the Kuonamka Formation (Middle Cambrian Amgan Stage) of the Bol'shaya Kuonamka and Malaya Kuonamka rivers, northern Anabar Shield, northern Siberia. In addition to the new genera Kuonamkaella and Anabaroconus, the assemblage contains species of Yochelcionella, Eotebenna, Pseudomyona, and Protowenella. These latter genera were previously unknown from the Middle Cambrian of Siberia and indicate a strong affinity with coeval molluscan faunas from the Australian Middle Cambrian. The fauna provides evidence for close palaeogeographie links to East and West Gondwana, Baltica, and Laurentia during the Middle Cambrian, and facilitates biostratigraphic correlation between these crustal blocks.
Alexander P. Gubanov [[email protected]], Artem V. Kouchinsky [[email protected]], John S. Peel [[email protected]], Department of Earth Sciences (Palaeontology), Uppsala University, Norbyviigen 22, SE-752 36 Uppsala, Sweden [present address of A. E Kouchinsky: Institute of Geophysics and Planetary Physics, University of California, Los Angeles, CA 90095-1567, USA]; Stefan Bengtson [[email protected]], Department of Palaeozoology, Swedish Museum of Natural History, Box 50007, SE- I 04 05 Stockholm, Sweden; received l 4.11.200 2, revised 17.4.2003.
Key words: Mollusca, helcionelloids, Middle Cambrian, Siberia.
VARIATION in our knowledge of Early and Middle Cambrian molluscs from different regions adversely affects the utility of molluscs for biostratigraphic and biogeographic purposes and impedes the discrimination of evolutionary lineages. Siberian helcionelloid molluscs from the Early Cambrian, especially from the Tommotian and Atdabanian, have been intensively studied (Vostokova 1962, R ozanov et al. 1969, Missarzhevsky 1989, Vassiljeva 1990, 1996), but the molluscs from overlying strata have received little attention. In contrast, molluscs from the late Early and Middle Cambrian in Australia, New Zealand and elsewhere exhibit an array of new morphologies (Runnegar & Jell 1976, MacKirmon 1985, Geyer 1986, Bengtson et al. 1990, Partdaaev 2001). However, the paucity of descriptions of the earlier Cambrian molluscs from these 0311/5518/2004/01001-20 $3.00 ©AAP
localities makes it difficult to trace the ancestry of these faunas. A silicified Middle Cambrian molluscan fauna, described herein from the Kuonamka Formation of northern Siberia, contains helcionelloids such as Eotebenna Runnegar & Jell, 1976, Protowenella Runnegar & Jell, 1976, Pseudomyona Runnegar, 1983, and Yochelcion-
ella Runnegar & Pojeta, 1974, all previously unknown from the Middle Cambrian of Siberia. The fauna provides evidence for c loser palaeogeographic links to East (Australia and New Zealand) and West (Spain and Morocco) Gondwana, Baltica and Laurentia in the Middle Cambrian, and facilitates direct biostratigraphic correlation between these crustal blocks.
Material and methods
Material described herein was obtained in 1996
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

2 GUBANOV, KOUCHINSKY, pEEl.& BENGTSON ALCHER1NGA
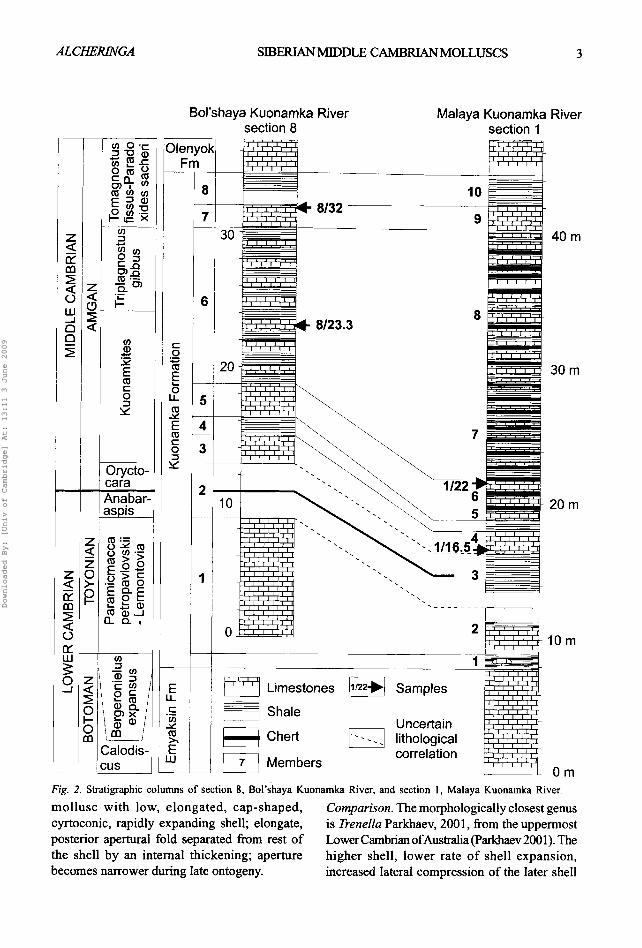
Fig. 1. Map of the Siberian Platform, showing the distribution of facies zones in the basin during the Early Cambrian and the locality for Middle Cambrian molluscs. 1, lagoonal facies zone; 2, transitional facies zone; 3, open-marine facies zone; B - Bol'shaya Kuonamka River, M - Malaya Kuonamka River.
during fieldwork by AVK and SB in northern Siberia (Fig. 1). The fossils are derived from limestone beds in the bituminous Kuonamka Formation (Fig. 2) and occur within the Amgan Stage of the Siberian Middle Cambrian. Acrolretid and lingulid brachiopods, polymerid trilobites and agnostids, simple phosphatic tooth-like and tubular structures, chancelloriids, net-l ike Microdictyon, button-shaped palaeoscolecid sclerites, hyoliths and ossicles of echinoderms co-occur with the molluscs described here.
Section 1 of the Kuo,amka Formation (Fig. 2) crops out along the left bank of the Malaya Kuonarnka River, near and downstream of the village of Zhilinda (70°02'N, 114°42'E). The lowermost part of the section belongs to the Emyaksin Formation and is composed o f bioturbated fossi l i ferous greenish-grey, glauconitic, lime mudstones with wacke- and packstones, interbedded with greenish-grey shale, especially in the upper 3 m of the formation. The Kuonamka Formation is overlain by yellowish-grey lime mudstones of the basal Olenyok Formation; sample 1/22 was collected
from the upper part of member 6 (top of bed 12 of Val'kov 1975, p. 28).
Because of its proximity to the northern part of the transitional facies zone of the Siberian Platform, the Kuonamka Formation in section 8 (Fig. 2) is considerably more calcareous than in section 1. Section 8 is exposed along the left bank of the Bol'shaya Kuonamka River, ca. 7 km downstream from the mouth of the Ulakhan- Tjulen Brook (Val'kov 1975). Sample 8/18.5 is from the upper part of member 5; sample 8/23.3 is from a lime mudstone layer near the middle of member 6; sample 8/32 was collected from the top o f member 7.
Silicified fossils were initially extracted from the limestones using a buffered solution of 10% dilute acetic acid. Mud was removed from the resultant insoluble residues by heating to boiling point in water and immediately decanting the suspension. Sample 8/32 was additionally treated with 10% diluted hydrochloric acid to remove calcite crystals, leaving the silicified fossils intact.
Systematic palaeontology
Figured material is housed in the type collection of the Swedish Museum of Natural History in Stockholm, Sweden (SMNH Mo 160563-615). AM F refers to the collections of the Australian Museum, Sydney; and ANU refers to collections in the Department of Earth and Marine Sciences, Australian National University, Canberra.
Phylum MOLLUSCA Cuvier, 1797 Class HELCIONELLOIDA Peel, 1991 a Order HELCIONELLIDAGeyer, 1994 Family HELCIONELLIDAE Wenz, 1938
K u o n a m k a e l l a gen. nov.
Derivation of name. From the Kuonamka River.
Type and only species. Kuonamkaella oliva sp. nov.
Diagnosis. Small, bilaterally symmetrical univalve
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009
ALCI-1ERINGA SIBERIAN MIDDLE CAMBRIANMOLLUSCS 3
2
rf
Z (d <
(.9 tt :~ E: < r
Z _< 7
z 0 < >-
.in
11
~ z
I1
E c o ,-~
Ory( to cara Anat,al aspi.,
o~ := --~ t~
° - -
E ~ E
Q.. Q. ,
u)
• e£
Calodis- CUS I
Borshaya Kuonamka River section 8
Ole t
F
c O
E o
LL
E c o
E LL r-"
E LU
~ok 1
8
3 0
6
,2° i 4,
3
1 0
1
8/32
Malaya Kuonamka River section 1
8/23.3
10
I I I I I I I I I I I, , ' , ' ',',1 " - . ~ " - . \ , 4
j, ,,,, ,,,,j -. . ~ " - . .1/16.5~ , , ' , ' ' ' , 1 . . ~ . . . . . . .
4tl,i, p l,i,I - , }11111 I I i l l i ~ I F l l l l I I i I i I
. . . . , ',I . . . . . . . . . j [ 1 1 ~ l l l l l I i t l l r
OI ',.','. .',~ 2 r ~ 7 ~ , ,
• Limestones ~ Samples
Shale Uncertain
Chert ~ lithological correlation
~ Members
40 m
30 m
20 m
~ 10m
r l , , ' , ' , , , I J I I I I I I
t', ..... I I I I I [111 I I I II ! ' ' , ' , ' j I l l I I
[ I i ~ l l [1 I I II
I I I I II I I I I i i i i i i I
~ l r l r l l I Om
Fig. 2. St~atigraphic columns of section 8, Bol'shaya Kuonamka River, and section 1, Malaya Kuonamka River.
mollusc with low, elongated, cap-shaped, Comparison. The morphologically closest genus cyrtoconic, rapidly expanding shell; elongate, is Trenella Parkhaev, 2001, from the uppermost posterior apertural fold separated from rest of Lower Cambrian ofAustralia (Parkhaev 2001). The the shell by an internal thickening; aperture higher shell, lower rate of shell expansion, becomes narrower during late ontogeny, increased lateral compression of the later shell
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009
4 GUBANOV, KOUCHINSKY, PEEI .& BENGISON ALCItF2UNGA
A B L, ~ a m k a e l l a
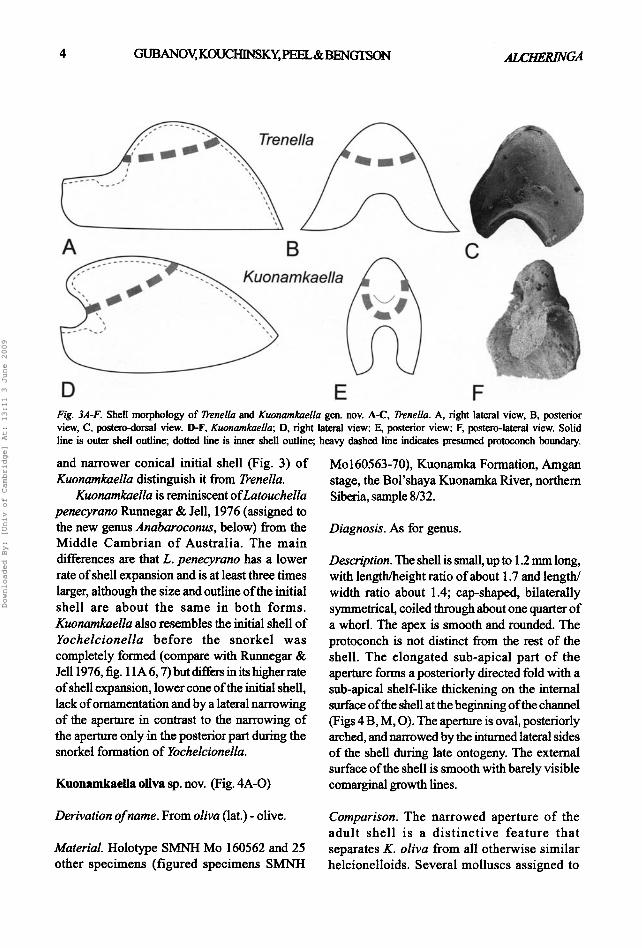
D E F Fig. 3AoF. Shell morphology of D'enella and Kuonamkaella gen. nov. A-C, D'enella. A, right lateral view, B, posterior view, C, l~stem--dorsal view. D-F, £uonamkaella; D, right lateral view; E, posterior view; F, l~stem-laterai view. Solid line is outer shell outline; dotted line is inner shell outline; heavy dashed line indicates ~esumed pmt~.~nch boundary.
and narrower conical initial shell (Fig. 3) of Kuonamkaella distinguish it from Trenella.
Kuonamkaella is reminiscent of Latouchella penecyrano Runnegar & Jell, 1976 (assigned to the new genus Anabaroconus, below) from the Middle Cambrian o f Australia. The main differences are that L. penecyrano has a lower rate of shell expansion and is at least three times larger, although the size and outline of the initial shell are about the same in both forms. Kuonamkaella also resembles the initial shell of Yochelcionella before the snorkel was completely formed (compare with Runnegar & Jell 1976, fig. l lA 6, 7) but differs in its higher rate of shell expansion, lower cone of the initial shell, lack of ornamentation and by a lateral narrowing of the aperture in contrast to the narrowing of the aperture only in the posterior part during the snorkel formation of Yochelcionella.
Kuonamkaella oliva sp. nov. (Fig. 4A-O)
Mo160563-70), Kuonamka Formation, Amgan stage, the Bol'shaya Kuonamka River, northern Siberia, sample 8/32.
Diagnosis. As for genus.
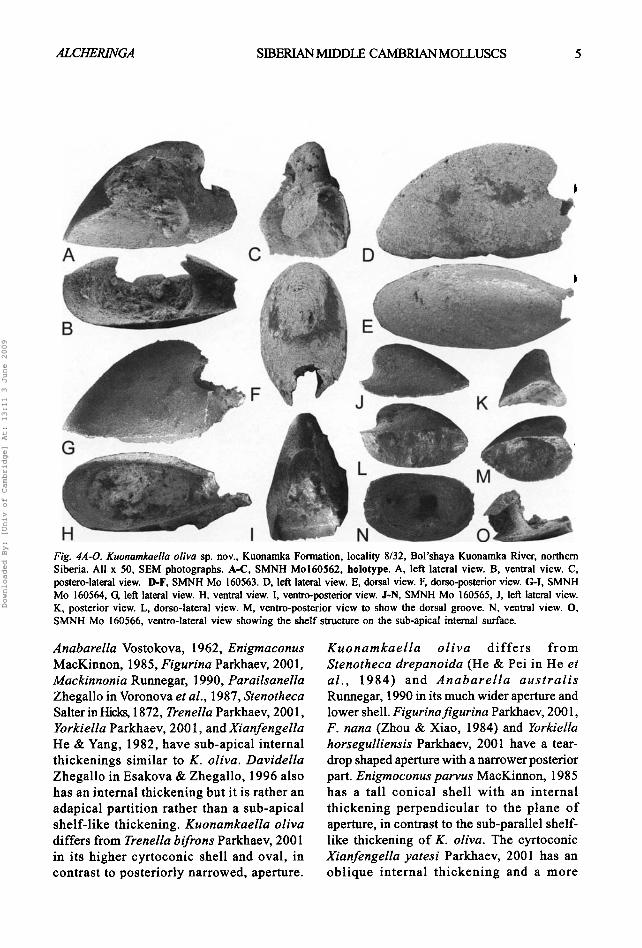
Description. The shell is small, up to 1.2 mm long, with length/height ratio of about 1.7 and length/ width ratio about 1.4; cap-shaped, bilaterally symmetrical, coiled through about one quarter of a whorl. The apex is smooth and rounded. The protoconch is not distinct from the rest of the shell. The elongated sub-apical part o f the aperture forms a posteriorly directed fold with a sub-apical shelf-like thickening on the internal surface of the shell at the beginning of the channel (Figs 4 B, M, O). The aperture is oval, posteriorly arched, and narrowed by the intumed lateral sides of the shell during late ontogeny. The external surface of the shell is smooth with barely visible comarginal growth lines.
Derivation o f name. From oliva Oat.) - olive.
Material. Holotype SMNH Mo 160562 and 25 other specimens (figured specimens SMNI-I
Comparison. The narrowed aperture of the adult shell is a d is t inc t ive fea ture that separates K. oliva from all otherwise similar helcionelloids. Several molluscs assigned to
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009
ALCHERINGA SIBERIAN MIDDLE CAMBRIAN MOLLUSCS 5
b
F
(-i
n I
Fig. 4A-O. Kuonamkaella ofiva sp. nov., Kuonamka Formation, locality 8/32, Bol'shaya Kuonamka River, northern Siberia. All x 50, SEM photographs. A-C, SMNH Mo160562, holotype. A, left lateral view. B, ventral view. C, postero-lateral view. I}-F, SMNH Mo 160563. D, left lateral view. E, dorsal view. F, dorso-posterior view. G-I, SMNH Mo 160564, G, left lateral view. H, ventral view. I, ventro-posterior view. J-N, SMNH Mo 160565, J, left lateral view. K, posterior view. L, dorso-lateral view. M, ventro-posterior view to show the dorsal groove. N, ventral view. O, SMNH Mo 160566, ventro-lateral view showing the shelf structure on the sub-apical internal surface.
Anabarella Vostokova, 1962, Enigmaconus MacKirmon, 1985, Figurina Parkhaev, 2001, Mackinnonia Runnegar, 1990, Parailsanella Zhegallo in Voronova et al., 1987, Stenotheca Salter in Hicks, 1872, Trenella Parkhaev, 2001, Yorkielta Parkhaev, 2001, and Xianfengella He & Yang, 1982, have sub-apical internal thickenings similar to K. oliva. Davidella Zhegal lo in Esakova & Zhegallo, 1996 also has an internal thickening but it is rather an adapical parti t ion rather than a sub-apical shelf- l ike thickening. Kuonamkaella oliva differs from Trenella bifrons Parkhaev, 2001 in its higher cyrtoconic shell and oval, in contrast to poster ior ly narrowed, aperture.
Kuonamkael la oliva d i f f e r s f r o m Stenotheca drepanoida (He & Pei in He et al., 1984) and Anabarel la austral is Runnegar, 1990 in its much wider aperture and lower shell. Figurinafigurina Parkhaev, 2001, F. nana (Zhou & Xiao, 1984) and Yorkiella horsegulliensis Parkhaev, 2001 have a tear- drop shaped aperture with a narrower posterior part. Enigmoconus parvus MacKirmon, 1985 has a tall conical shell wi th an internal th ickening perpendicular to the p lane o f aperture, in contrast to the sub-parallel shelf- like thickening o f K. oliva. The cyrtoconic Xianfengella yatesi Parkhaev, 2001 has an ob l ique in ternal th i cken ing and a more
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

6 GUBANOV, KOUCHINSKY, PEF~I.& BENGTSON ALCHER1NGA
C
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

ALCHERINGA SIBERIAN MIDDLE CAMBR/AN MOLLUSCS 7
strongly coiled and wider shell. Mackinnonia rostrata (Zhou & Xiao, 1984) and Parailsanella lata Parldaaev, 2001 have much taller shells with series of internal shell thickenings on the dorsum.
Anabaroeonus gen. nov.
Derivation o f name. From the Anabar River and conus (lat.) - cone.
Type species. Anabaroconus sibiricus sp. nov.
Diagnosis. Small, bilaterally symmelrical univalve mollusc with cyrtoconic moderately expanding shell, ornamented by fine radial striae and broad co-marginal plicae; apertural margin arched sub- apically.
Comparison. The new genus differs from Helcionella Grabau & Shimer, 1909 in its less strongly coiled and more laterally compressed shell, wider and less frequent and comarginal plicae that almost disappear on the anterior and posterior parts of the shell. The tear-drop shaped aperture in plan view, with the sub-apical part lifted up in lateral view, contrasts with the aperture of Helcionella, which is more oval in plan view and fiat in lateral view.
Species included. Type species and Latouchella penecyrano Rurmegar & Jell, 1976.
Anabaroconus sibiricus sp. nov. (Fig. 5 A-U)
Derivation o f name. From Siberia.
Material . Holo type SMNH Mo 160572, Kuonamka Formation, Amgan stage, northern Siberia. Three complete specimens (SMNH Mo160572-74) including the holotype; and one fragment (SMNH Mo 160575) from sample 8/32;
two fragments (SMNH Mo 160571, 160577) from sample 1/22; and one shell fragment (SMNH Mo 160576) from sample 8/18.5.
Description. The shell is small, up to 1.6 mm long and 1.3 mm high, bilaterally symmetrical , cyrtoconic (coiled about 1/8-1 / 10 of a whorl) with a slightly laterally compressed form; length/width ratio is about 1.2. The initial part of the shell is hemispherical in shape with a diameter of about 0.3-0.4 ram. Ornamentation consists of a fine radial striation and broad rounded co-marginal plicae that are most prominent and sharper on the lateral sides and almost disappear on the sub-apical part of the shell. The aperture is oval in outline with the subapical part arched to form a broad shallow channel.
Remarks. Latouchella Cobbold, 1921 is widely reported in the literature, but most o f the Cambrian molluscs placed therein do not belong to this genus (see Gubanov & Peel 1999). Latouchellapenecyrano Runnegar & Jell, 1976 is one such helcionelloid which is here referred to Anabaroconus. Anabaroconus sibiricus differs from A. penecyrano in its less strongly coiled, lower shell and less elongated subapical aper tural fold. Helc ionel la terraustral is Runnegar & Jell, 1976 has a subapically arched apertural margin similar to that ofA. sibiricus, although H. terraustralis is coiled through more than ha l f a whorl and has more frequent comarginal plicae that continue on the dorsum. It also has sub-apical ridges on the shell interior, lacking in A. sibericus, that relate H. terraustralis to Latouchella merino Runnegar & Jell, 1976, L. accordionata Runnegar & Jell, 1976, and Latouchella spp. Peel (1991 a). However, none of these molluscs with internal ridges should be attributed to Helcionella or Latouchella.
Fig. 5A-U. Anabaroconus sibericus sp. nov. from the Kuonamka Formation, localities 8/18.5 and 8/32, Bol'shaya Kuonamka River and locality 1/22, Malaya Kuonamka River, northern Siberia. All except N and U x 30; SEM photographs. A, SMNH Mo 160571, left lateral view. B-D, SMNH Mo 160572, holotype. B, lateral view; C, posterior view; D, dorsal view. E-G, SMNH Mo 160575. E, left lateral view; F, ventral view; G, anterior view. H-K, SMNH Mo 160576. H, right lateral view; I, anterior view; J, dorsal view; K, postero-lateral view. L-N, SMNH Mo 160577. L, right lateral view; M, dorsal view; N, close up of doubled radial ridges on the lateral side, x 230. O-Q, SMNH Mo 160573. O, right lateral view; P, dorsal view; Q, antero-dorsal view. R-U, SMNH Mo 160574. R, left lateral view; S, posterior view; T, dorsal view; U, close up of plicae to show the radial structure on the shell surface, x 90. [see page 6 opposite].
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

8 GUBANOV, KOUCHINSKY, Pl~.h-'l & BENGTSON ALCHERINGA
Y o c h e l c i o n e l l a Runnegar & Pojeta, 1974
Type species. Yochelcionella cyrano Runnegar & Pojeta, 1974.
Diagnosis. Small, bilaterally symmelrical univalve mollusc with slightly curved, slowly expanding shell with a prominent sub-apical tube (snorkel).
Diagnosis. Small, bilaterally symmetrical univalve mollusc with tall, slowly expanding shell, S- shaped in lateral view (with snorkel to the right), with prominent sub-apical snorkel that points obliquely upwards in mature specimens; sculpture consists of sharp regular comarginal plicated ribs and fLUe radial striation.
Species included. Yochelcionella a ichal ica Fedorov 1987; Y. americana Runnegar & Pojeta, 1980; Y. angustiplicata Hinz-Schallreuter, 1997; Y. chinensis Pei, 1985; Y. cyrano Runnegar & Pojeta, 1974; Y. daleki Runnegar & Jell, 1976; Runnegar in Bengtson et aL, 1990; Y. erecta ( Stenotheca? rugosa var. erecta) (Walcott, 1891); Y. fissurata Hinz-Schallreuter, 1997; Y. ostentata Runnegar & Jell, 1976; Y. recta Missarzhevsky in Missarzhevsky & Mambetov, 1981; Y stylifera Missarzhevsky in Missarzhevsky & Mambetov, 1981; Y. trompetica Hinz-Schallreuter, 1997. Several Yochelcionella have been described under open nomenclature from Siberia (Dzik 1994), Morocco (Geyer 1986), Denmark (Berg- Madsen & Peel 1987), England (Hinz 1987) and Canada (Voronova et al. 1987, Landing & Bartowski 1996, Landing et al. 2002).
Y o c h e l e i o n e l l a o s t e n t a t a Runnegar & Jell, 1976 (Fig. 6A-S)
1976 Yochelcionella ostentata sp. nov. Runnegar & Jell, p. 130, fig. 11C5-8.
Holotype . Australian National University, Canberra, ANU 29085 from sample 10352; Coonigan Formation, Ordian - lower Temple- tonian, Middle Cambrian, western New South Wales, Australia.
Material. Four complete silicified specimens (SMNH Mo 1 6 0 5 7 8 - 8 1 ) and one shell fragment (SMNH Mo160582) from sample 8/23.3, three incomplete specimens (SMNH Mo160583-85) from sample 8/18.5, and one pyritized shell fragment (SMNH Mo 160586) from sample 1/22.
Description. The shell is small, tall and bilaterally symmetrical, with a slightly sigmoid profile (Fig. 6F); the prominent upward-directed snorkel is located just beneath the initial part of the shell. The initial part of the shell is cap-shaped, slightly coiled, with barely visible shallow concentric folds and f'me striations which radiate from the apex (Fig. 6S). The shell is oval in cross section with antero-posterior elongation. The snorkel is perpendicular to the initial shell and may be slightly curved upward; with growth of the sigmoidal shell it becomes increasingly tilted, steeply inclined to the aperture. The snorkel is dorso-ventrally compressed at its base and is formed as an extension of the first co-marginal rib (Fig. 61, Q, K). The rest of the snorkel has a circular cross section. The later shell is slowly expanding and coiled in the opposite direction to the early stages to give the sigmoidal lateral profile; it has an oval cross section. Sculpture consists of radial striae and comarginal, evenly spaced, sharp, phcated ribs with concave interareas. The distance between ribs is about 150-200 ~tm.
Fig. 6A-S. Yochelcionella ostentata Runnegar & Jell, 1976 from the Kuonamka Formation, localities 8/18.5 and 8/ 23.3, Bol'shaya Kuonamka River and locality 1/22, Malaya Kuonamka River, northern Siberia. All x 30 except I and S, all SEM photographs. A-E, SMNH Mo 160578. A, right lateral view; B, antero-lateral view; C, postero-lateral view; D, posterior view; E, apertural view; F-I, SMNH Mo 160579. F, left lateral view; G, anterior view; H, posterior view; I, dorso-lateral view to show the initial shell and dorso-ventral compression of the snorkel, x 50. J, SMNH Mo 160582, left lateral view. K-L, SMNH Mo 160580. K, right lateral view; L dorso-lateral view showing lateral comarginal rogae. M-N. SMNH Mo 160583, adult shell with lacking apical part and snorkel. M, left lateral view; N, dorsal view. O-S, SMNH Mo 160581. O, right lateral view; P, dorsal view; R, oblique postero-lateral view to show the dorso-ventraUy compressed snorkel; Q, oblique dorso- lateral view to show transition between initial shell and snorkel; S, close up of apex with radial striation, x 180. [see page 9 opposite].
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

ALCHERINGA SIBERIAN MIDDLE CAMBRIAN MOLLUSCS 9
(
G l - ' v H
F
K "~ "e L
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

10 GUBANOV, KOUCHINSKY, PEEl.& BENGTSON ALCHERINGA
Comparison. Siberian samples differ slightly from the type series from Australia. While the size of the shell and snorkel are similar, the Australian samples show a less curved initial part of the shell and only half the distance between ribs, although this feature is rather variable within the type population. The fine radial striation is visible in samples from both areas (compare Runnegar & Jell 1976, fig. 11C and Fig. 6S herein). Dorso- ventral compression at the base of the snorkel seems to be present in the holotype (Runnegar & Jell 1976, fig. 11C 9), but is not noted in the original description.
Yochelcionella angusfiplicata Hinz-Schallreuter, 1997 (Fig. 7A-F)
1997 Yochelcionella angustiplicata sp. nov. Hinz- Schailreuter, pp. 115-7, fig. 5, pls 1-2.
Holotype. Museum fiir Naturkunde Berlin, MB Nr. Ga.448, Exsulans Limestone, Middle Cambrian, Bomholm, Denmark.
Material One specimen SMNH Mo160587 from sample 8/32.
Description. The shell is about 1 mm long, 0.8 mm high, and 0.7 mm wide, and slightly sigmoid when viewed laterally, with the snorkel to the right. The sub-apical snorkel has a circular cross section and is about 0.3 mm in diameter. The initial part of the shell is smooth, low cap-shaped and laterally compressed with a length/width ratio of about 2. Sculpture consists of transverse closely spaced sharp plicated ribs. The distance between ribs is about 50 ~tm.
Comparison. This specimen is very similar to Y. daleki from the Middle Cambrian of Australia in terms of its closely spaced ribs. However, the Siberian specimen is S-shaped in lateral view in contrast to the C-shaped Y. daleki which has the early and later parts of the shell curved in the same direction with the snorkel on the concave part of the shell. As illustrated by Runnegar & Jell (1976), specimens of Y. daleki show great variability both in shape and in size. ANU 29068
(Runnegar & Jell 1976, fig. 11A 15, 16) is most similar to the Siberian specimen as well as to the holotype of Y. angustiplicata Hinz-Schallreuter, 1997, in terms of the size of the initial and adult shells and the frequency of fibs, but it is clearly C-shaped. The holotype of Y. daleki has a much larger protoconch and adult shell, a wider snorkel, and as much as ten or more times greater distance between ribs. The shape of the holotype changes from the slightly coiled initial shell to the straight adult shell with a tendency to curve in the opposite direction according to the change in the surface inclination between ribs beneath the snorkel (Fig. 11A 13). This morphological variation suggests that specimens ANU 29068 (Australian National University, Canberra) of Runnegar & Jell (1976, fig. l l A 15, 16) and AM F97397 (Australian Museum, Sydney) of Brock (1998, fig. 4 4-10) may not be conspecific with the holotype of Y. daleki.
Eotebenna Runnegar & Jell, 1976.
Type species. Eotebenna pontifex Runnegar & Jell, 1976.
Diagnosis. Small, bilaterally symmetrical, laterally compressed, univalve mollusc with sub-central spoon-like protoconch and elongated posterior snorkel that connects to the aperture by a narrow slit.
Species included. Eotebenna pontifex Rurmegar & Jell, 1976; E.papilio Runnegar & Jell, 1976; E. arctica Peel, 1989; E.pelmani (Vassiljeva, 1990); E. viviannae Peel, 1991 a.
Eotebenna viviannae Peel, 1991 (Fig. 8A-I)
1991a Eotebenna viviannae sp. nov. Peel, pp. 173-175, figs 19A-N.
1991b Eotebenna viviannae Peel; Peel, fig. 31.
Holotype. Geological Museum, Copenhagen, Denmark, MGUH 19.564; Andmmm Limestone, late Middle Cambrian, Bomholm, Denmark.
Material. Ten specimens (SMNH Mo 160588-97), in various states ofpreservation, from sample 8/32.
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009
ALCHERINGA SIBERIAN MIDDLE CAMBRIANMOLLUSCS 11
i
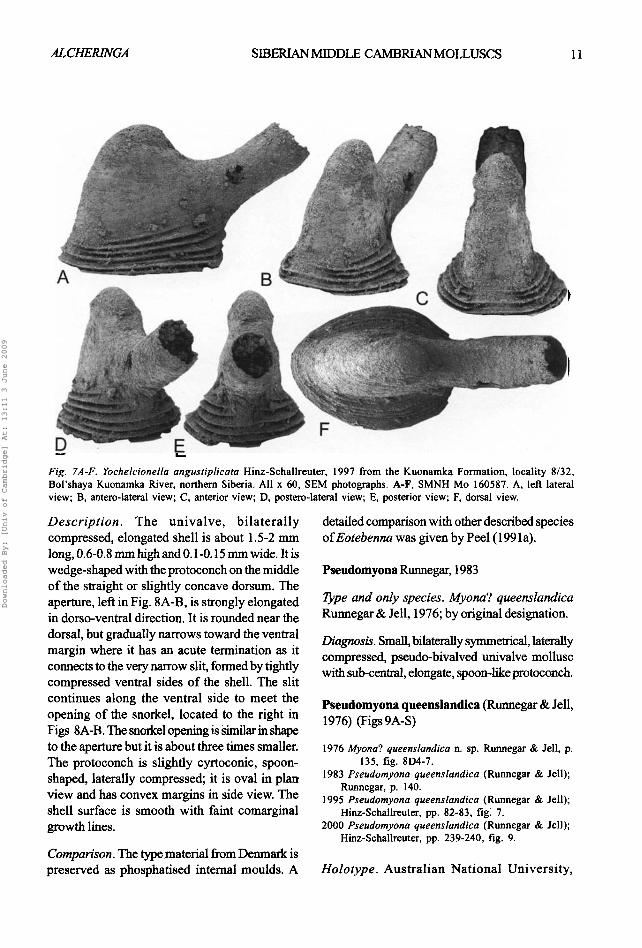
Fig. 7A-F. Yochelcionella angustiplicata Hinz-Sehallreuter, 1997 from the Kuonamka Formation, locality 8/32, Bol'shaya Kuonamka River, northern Siberia. All x 60, SEM photographs. A-F, SMNH Mo 160587. A, left lateral view; B, antero-lateral view; C, anterior view; D, postero-lateral view; E, posterior view; F. dorsal view.
Description. The univalve, bi la teral ly compressed, elongated shell is about 1.5-2 mm long, 0.6-0.8 nun high and 0.1-0.15 mm wide. It is wedge-shaped with the protoconch on the middle of the straight or slightly concave dorsum. The aperture, left in Fig. 8A-B, is strongly elongated in dorso-ventral direction. It is rounded near the dorsal, but gradually narrows toward the ventral margin where it has an acute termination as it connects to the very narrow slit, formed by tightly compressed ventral sides of the shell. The slit continues along the ventral side to meet the opening of the snorkel, located to the right in Figs 8A-B. The snorkel opening is Similar in shape to the aperture but it is about three times smaller. The protoconch is slightly cyrtoconic, spoon- shaped, laterally compressed; it is oval in plan view and has convex margins in side view. The shell surface is smooth with faint comarginal growth lines.
Comparison. The type material from Denmark is preserved as phosphatised internal moulds. A
detailed comparison with other described species of Eotebenna was given by Peel (1991a).
Pseudomyona Runnegar, 1983
Type and only species. Myona? queenslandica Runnegar & Jell, 1976; by original designation.
Diagnosis. Small, bilaterally symmetrical, laterally compressed, pseudo-bivalved univalve mollusc with sub-central, elongate, spoon-like protoconch.
Pseudomyona queenslandica (Runnegar & Jell, 1976) ffigs 9A-S)
1976 Myona? queenslandica n. sp. Runnegar & Jell, p. 135, fig. 8D4-7.
1983 Pseudomyona queenslandica (Runnegar & Jell); Runnegar, p. 140.
1995 Pseudomyona queenslandica (Runnegar & Jell); Hinz-Schallreuter, pp. 82-83, figl 7.
2000 Pseudomyona queenslandica (Runnegar & Jell); Hinz-Schallreuter, pp. 239-240, fig. 9.
Holotype. Australian National University,
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

12 GUBANOV, KOUCHINSKY, pE-~,&BENGTSON ALCHERINGA
v
Fig. 8A-L Eotebenna viviannae Peel, 1991 from the Kuonamka Formation, locality 8/32, Bol'shaya Kuonamka River, northern Siberia. All x 60, SEM photographs. A-F, SMNH Mo 160588. A, left lateral view; B, oblique antero- lateral view; C. antero-dorsal view; D, dorso-posterior view; E, dorso-lateral view; F, antero-lateral view. G-I, SMNH Mo 160589. G, left lateral view; H, dorso-lateral view; I, dorsal view.
Canberra, A N U 29071 from L 132; Currant Bush Limestone, Late Templetonian-Floran, Middle Cambrian, western Queensland, Australia.
Material. Twenty - seven spec imens ( f igured specimens SMNH Mo160598-603), in various states o f preservation, from sample 8/32.
Description. The shell is small, up to 1.2 mm long and 0.8 mm high, bilaterally symmetrical, pseudo- bivalved with strongly incurved lateral margins, and a sWaight (in lateral view) dorsum. The spoon- shaped larval shell is low and elongated. It is situated on the dorsum closer to the posterior end o f the shell, with a smooth transition to the
adult shell anteriorly and posteriorly; lateral edges o f the p ro toconch bend inward pr io r to the junction with the teleconch. The angular lateral margins continue posteriorly to delimit a prominent carina that continues from the wider posterior part o f the protoconch to the margin (Fig. 9F).
Remarks. Three phosphat ised internal moulds recovered from the Currant Bush Limestone o f Aust ra l ia were descr ibed as b iva lved wi th a univalve protoconch by Runnegar & Jell (1976). On the basis o f the univalve protoconch, P. queenslandica was a s s igned to the ex t inc t mol luscan Class Rost roconchia Runnegar & Pojeta, 1974. Subsequently, it was removed to
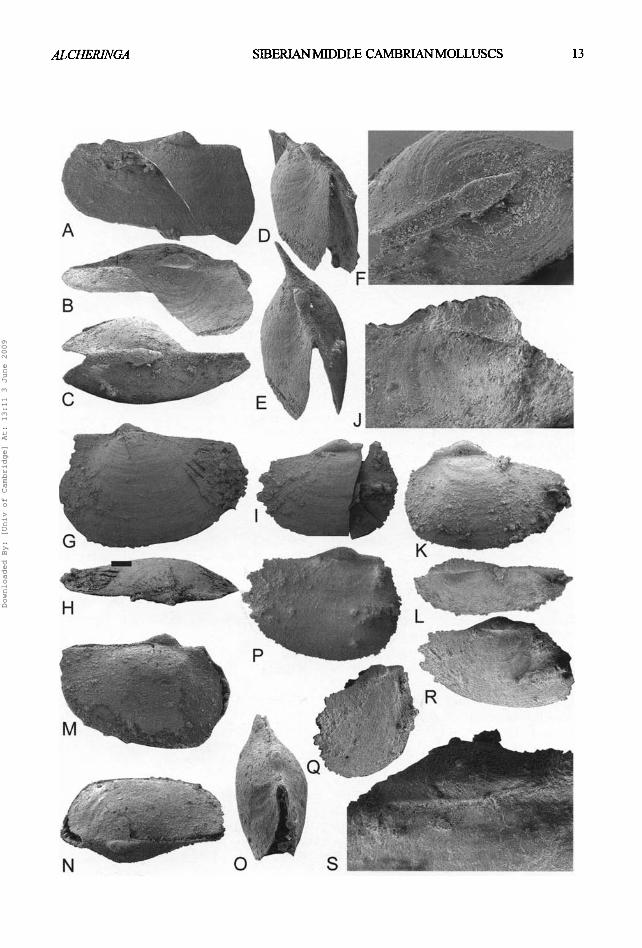
Fig. 9A-S. Pseudomyona queenslandica (Runnegar & Pojeta, 1976) from the Kuonamka Formation, locality 8/32, Bol'shaya Kuonamka River, northern Siberia. All x 40 except F, J and S. All SEM photographs. A-F, SMNH Mo. 160598, A, left lateral view, x 7.5; B, dorso-lateral view; C, dorsal view; D, antero-lateral view; E, antero-dorsal view; E close up of protoconch, x 80. G-H, SMNH Mo 160599. G, right lateral view; H, dorsal view. !, SMNH Mo 160600, left lateral view. J-L, SMNH Mo 160601. J, close up of protoconch, x 120. K, lateral view; L, latero-dorsal view. M-O, SMNH Mo 160602. M, left lateral view; N, dorso-lateral view; O, antero- dorsal view. P-S, SMNH Mo 160603. P, left lateral view; Q, antero-lateral view; R, dorso-lateral view; S, close up of protoconeh, x 175. [see page 13 opposite].
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009
ALCHERINGA SIBER/AN MIDDLE CAMBRIANMOLLUSCS 13
I t \
IvI
r~ L)
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009
14 GUBANOV, KOUCHINSKY, Ph-~ .&BENGTSON ALCI-1ERINGA
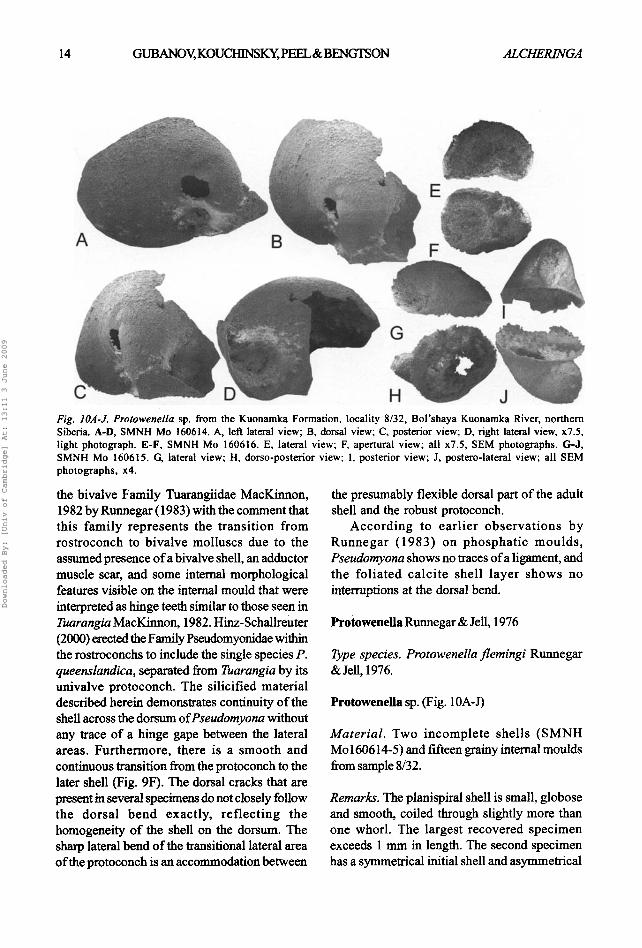
Fig. IOA-J. Protowenella sp. from the Kuonamka Formation, locality 8/32, Bol'shaya Kuonamka River, northern Siberia. A-D, SMNH Mo 160614. A, left lateral view; B. dorsal view; C, posterior view; D, right lateral view, x7.5, light photograph. E-F, SMNH Mo 160616. E, lateral view; F, apertural view; all x7.5, SEM photographs. G-J, SMNH Mo 160615. G, lateral view; H, dorso-posterior view; I, posterior view; J, postero-lateral view; all SEM photographs, x4.
the bivalve Family Tuarangiidae MacKinnon, 1982 by Runnegar (1983) with the comment that this family represents the transition from rostroconch to bivalve molluscs due to the assumed presence of a bivalve shell, an adductor muscle scar, and some internal morphological features visible on the internal mould that were interpreted as hinge teeth similar to those seen in Tuarangia MacKinnon, 1982. Hinz-Schallreuter (2000) erected the Family Pseudomyonidae within the rostroconchs to include the single species P. queenslandica, separated from Tuarangia by its univalve protoconch. The silicified material described herein demonstrates continuity of the shell across the dorsum of Pseudomyona without any trace of a hinge gape between the lateral areas. Furthermore, there is a smooth and continuous transition from the protoconch to the later shell (Fig. 9F). The dorsal cracks that are present in several specimens do not closely follow the dorsal bend exactly, ref lect ing the homogeneity of the shell on the dorsum. The sharp lateral bend of the transitional lateral area of the protoconch is an accommodation between
the presumably flexible dorsal part of the adult shell and the robust protoconch.
According to earl ier observat ions by Runnegar (1983) on phosphat ic moulds, Pseudomyona shows no traces of a ligament, and the fol iated calci te shell layer shows no interruptions at the dorsal bend.
Protowenella Runnegar & Jell, 1976
Type species. Protowenella flemingi Runnegar & Jell, 1976.
Protowenella sp. (Fig. 10A-J)
Material. Two incomplete shells (SMNH Mo 160614-5) and fifteen grainy internal moulds from sample 8/32.
Remarks. The planispiral shell is small, globose and smooth, coiled through slightly more than one whorl. The largest recovered specimen exceeds 1 mm in length. The second specimen has a symmetrical initial shell and asymmetrical
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

ALCHERINGA SIBERIAN MIDDLE CAMBRIAN MOLLUSCS 15
later whorl that could be a result of deformation. Internal moulds are poorly preserved and do not reveal any further details.
Remarks on preservation and diagenesis
Silicification is common in the Kuonamka Formation but absent in the basal limestones of the overlying Olenyok Formation and the underlying limestones of the Emyaksin Formation. Bakhturov et al. (1988) suggested that spicules of siliceous sponges, which locally constitute 5- 10% percent of the rock, may have been the main source of silica, although terrigenous and volcanic input may also have been important (Akul'shina et al. 1972, Bakhturov et al. 1988), if only as the ultimate source of the silica used by the sponges.
Bedded black bituminous cherts and shales of the Kuonamka Formation reflect slow sedimentation rates and anoxic environments close to the sediment-water interface (Bakhturov et al. 1988). Pyrite in limestone samples of the Kuonamka Formation suggests recurrent anoxic conditions. Pyrite is commonly associated with diagenetic silicification, but the silicification of carbonate shells seems to be mainly governed by bacterial oxidation of organic matter rather than by sulphate reduction (Carson 1991). The richest silicified assemblage of fossils (sample 8/ 32) is associated with the marine hardground surface penetrated by borings (the 'Matokuonamka marker bed') at the top of member 7 in section 8 (Fig. 2). This bed shows the lowest concentration of bitumen anywhere within the Kuonamka Formation (Bakhturov et al. 1988); the early diagenetic silicification may have been facilitated by the higher permeability caused by bioturbation.
Silica-replaced fossils are well known through the Phanerozoic (Schubert et al. 1997). Skeletal fossils selectively replaced by silica are common for example in Early and Middle Cambrian limestones of New South Wales and Queensland, from which rich molluscan fauna have been
reported (Runnegar & Jell 1976, Brock 1998). Shell walls replaced by silica do not usually preserve original microstructure, but reflect shell shapes and ornamentation of the shell exterior (Rurmegar & Pojeta 1985). Preservation of shell shapes is mainly due to replication, with silica filling voids after dissolved aragonite or calcite. Rarely, silica precipitation and calcite dissolution proceed at equal rates, and silicification preserves original microstructure and shell layers by preferred orientation and external morphology of quartz crystals (Schmitt & Boyd 1981, Boyd & Newell 1984).
Dissolution and subsequent filling of the cavity with silica is more rapid for aragonitic shells or shell layers than for calcitic ones. Usually silicification fills voids left after aragonite dissolution, but calcite is more stable and therefore more likely to undergo replacement by silica. Calcite was indicated as a primary mineral in walls of the Middle Cambrian Eotebenna pontifex Rurmegar & Jell, 1976, Pseudomyona queenslandica (Runnegar & Jell 1976), Tuarangia paparua MacKinnon, 1982 and T. gravgaerd- ensis Berg-Madsen, 1987 (Runnegar 1985, Berg- Madsen 1987). The first two genera have been recovered from the Kuonamka Formation. Walls of Pseudomyona were possibly composed of calcite with a foliated microstructure in the whole shell except for the myostracal layer (Runnegar 1983). Eotebenna also had a foliated calcitic inner shell layer (Runnegar & Jell 1980). Fine radial striation seen in some specimens of Pseudomyona, Yochelcionella and Anabaro- conus (Figs 5N, 6S, 9G, H) may reflect similar architecture of the periostracum in these forms.
Stratigraphic and palaeogeographic implications
The first descriptions of Middle Cambrian molluscs of Australian aspect from Bornholm, Denmark (Berg-Madsen & Peel 1978, Berg- Madsen 1987), facilitated stmtigraphic correlation between the two areas but presented a problem for existing palaeogeographic reconstructions
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009
16 GUBANOV, KOUCHINSKY, pEEIJ& BENGTSON ALCHERINGA
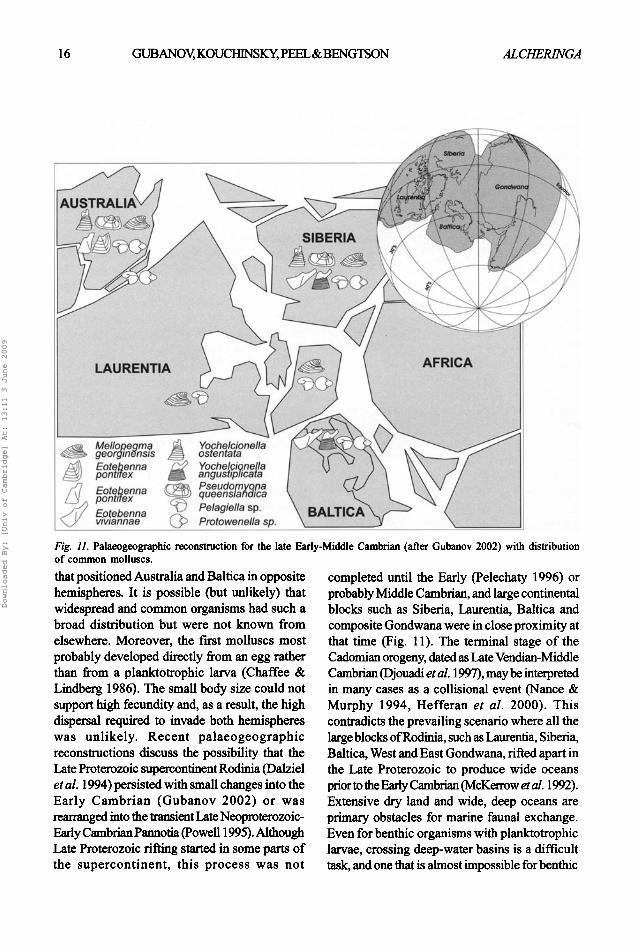
Fig. 11. Palaeogeographic reconstruction for the late Early-Middle Cambrian (after Gubanov 2002) with distribution of common molluscs. that positioned Australia and Baltica in opposite hemispheres. It is possible (but unlikely) that widespread and common organisms had such a broad distribution but were not known from elsewhere. Moreover, the lust molluscs most probably developed directly from an egg rather than from a planktotrophic larva (Chaffee & Lindberg 1986). The small body size could not support high fecundity and, as a result, the high dispersal required to invade both hemispheres was unlikely. Recent palaeogeographic reconstructions discuss the possibility that the Late Proterozoic supercontinent Rodinia (Dalziel et al. 1994) persisted with small changes into the Early Cambrian (Gubanov 2002) or was rearranged into the transient Late Neoproterozoic- Early Cambrian Pannotia (Powell 1995). Although Late Proterozoic firing started in some parts of the supercontinent, this process was not
completed until the Early (Pelechaty 1996) or probably Middle Cambrian, and large continental blocks such as Siberia, Laurentia, Baltica and composite Gondwana were in close proximity at that time (Fig. 11). The terminal stage of the Cadomian orogeny, dated as Late Vendian-Middle Cambrian (Djouadi et al. 1997), may be interpreted in many cases as a coUisional event (Nance & Murphy 1994, Hefferan et al. 2000). This contradicts the prevailing scenario where all the large blocks of Rodinia, such as Laurentia, Siberia, Baltica, West and East Gondwana, fired apart in the Late Proterozoic to produce wide oceans prior to the Early Cambrian (McKerrow et al. 1992).
Extensive dry land and wide, deep oceans are primary obstacles for marine faunal exchange. Even for benthic organisms with planktotrophic larvae, crossing deep-water basins is a difficult task, and one that is almost impossible for benthic
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

ALCHERINGA SIBERIAN MIDDLE CAMBRIAN MOLLUSCS 17
organisms with direct development from an egg. The distribution of some late Vendian-Early Cambrian fossils supports a palaeogeographic model with closely juxtaposed continental blocks separated by shallow water basins, and not by wide, deep oceans (Gubanov 1998, Waggoner 1999). It seems also that there were no extensive areas of land but just relatively small islands and island chains along the coUisional zones. The fauna from northern Siberia described here may suggest that similar conditions persisted into the Middle Cambrian, permitting extensive exchange of benthic faunas between major continental blocks such as Siberia, Baltica, Laurentia and Australia. The high diversity of mollusc faunas from Siberia and Australia, in contrast to more restricted faunas from Greenland (JSP personal observation) and Baltica, possibly reflects climatic differentiation analogous to the Early Cambrian faunal zonation (Gubanov 2002), though there are insufficient data to support a firm conclusion.
The silicified molluscs described here from Siberia span three Middle Cambrian trilobite zones, namely Kuonamkites, Triplagnostus gibbus and Tomagnostus fissus - Paradoxides sacheri (Fig. 2); molluscs occur in the lower and upper of these zones. The two upper zones are also recognized in Baltica (Geyer & Shergold 2000), allowing correlation of the Exsulans Limestone and the Kalby Clay of Bornholm with the upper part of the Kuonamka Formation (unit 6 of Section 8 and unit 8 of section 1, Fig. 2). The Andrarum Limestone is late Middle Cambrian and somewhat younger than the Kuonamka Formation. The Triplagnostus gibbus trilobite zone (Jell 1975) is recognised world wide, including Australia (Shergold et al. 1990, Geyer & Shergold 2000) and Greenland 0neson & Peel 1997), where a similar molluscan fauna also occurs (JSP & Christian Skovsted, personal observation). The silicified molluscs described by Runnegar & Jell (1976) from the Coonigan Formation are coeval with Siberian molluscs from localities 8/18.5 - 23.3 and 1/22. The Currant Bush Limestone is slightly younger than the Member 7 of the Kuonamka Formation (locality 8/32) and
coeval with the overlying Olenyok Formation. Although the Middle Cambrian mollusc fauna
from Greenland is less diverse and mainly preserved as phosphatised steinkerns, it shares several taxa with Australia. 'Latouchella'-like steinkerns with characteristic furrows produced by ridges on the shell interior (Peel 1991b) resemble several Australian molluscs with silicified shells described as Latouchella merino, L. accordionata and Helcionella terraustralis from the Coonlgan Formation (Runnegar & Jell 1976) in which such ridges are present on the shell interior. Peel (1979) reported Protowenella from the same horizon in North Greenland, at that time commenting on the remarkable distribution from Australia, to Denmark and northernmost Greenland. Pelagiella from the same locality in Greenland shows more similarity to P. adunca He & Pei, 1981 (He et al. 1984) from the late Early Cambrian of Australia, North China (Bengtson et al. 1990, Parkhaev 2001) and Siberia (APG personal observation) than to P. corinthiana Runnegar & Jell, 1976 and P. deltoides Runnegar & Jell, 1976 from the Middle Cambrian Currant Bush Formation of Australia (Runnegar & Jell 1976). Furthermore, Mellopegma georginensis Runnegar & Jell, 1976, known from the Currant Bush Formation, the Gowers Formation and the Thorntonia Limestone (Glenn Brock, personal communication), also occurs in Greenland. Mellopegma georginensis has also been described from the upper part of the 'Anse Maranda Formation' of Quebec, Canada, but Landing et al. (2002) proposed an older, late Early Cambrian age for this interval. Similar poorly preserved phosphatized shells, preliminary determined as M. cf. georgiensis, have also found in the late Early Cambrian basal limestone of the Kuonamka Formation in the section 1 of the Malaya Kuonamka River.
Acknowledgments We thank Anatolij V. Val'kov, Shane Pelechaty, and the late Vladimir V. Missarzhevsky for assistance in the field. Glenn A. Brock and Peter A. Jell are thanked for instructive reviews. Our
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

18 GUBANOV, KOUCHINSKY, PEI~L & BENGTSON ALCHERINGA
work has been financially supported by grants from the Royal Swedish Academy of Sciences (KVA) and the Swedish Natural Science Research Council (NFR, now VR).
R e f e r e n c e s
AKOL'SHn~A, E.P., EVTUSHE~KO, V.M. & PISAREVA, GM., 1972. O prirode plastichnykh glin v kembrijskikh otlozheniyakh bassejna r. Olenyok (On the nature of soft clays from the Cambrian deposits of the Olenyok River basin). Geologiya i geofizika 2, 147- 151 (Russian).
BAKHTUROV, S.E, EV'VOSHENKO, V.M. & PERELADOV, V.S., 1988. Kuonamskaya bituminoznaya karbonatno- slantsevaya formatsiya (The Kuonamka bituminous carbonate shale formation). Trudy Instituta Geolodii i Geofiziki, Sibirskoe otdelenL Akademiya nauk SSSR 671, 160 p. (Russian).
BENGTSON, S., CONWAY MORRIS, S., COOPER, B.J., JELL, P.A. & RUNNEGAR, B.N., 1990. Early Cambrian fossils from South Australia. Association of Australasian Palaeontologists, Memoir 9, 364 p.
BERG-MADSEN, V., 1987. Tuarangia from Bornholm (Denmark) and similarities in Baltoscandian and Australian late Middle Cambrian. Alcheringa 11, 245- 259.
BERG-MADSEN, V. & PEEL, J.S., 1987. Yochelcionella (Mollusca) from the late Middle Cambrian of Bornholm, Denmark. Bulletin of the Geological Society of Denmark 36, 259-261.
BLAKER, M.R. & PEEL, J.S., 1997. Lower Cambrian trilobites from North Greenland. Meddelelser om Gronland, Geoscience 35, 145 p.
BoYo, D.W. & NEWELL, ND., 1984. Vestigial shell structure in silicified pectinacean pelecypods. Contributions to geology, University of Wyoming 23, 1-8.
BROCK, G.A., 1998. Middle Cambrian molluscs from the southern New England Fold Belt, New South Wales, Australia. Geobios 31, 571-586.
CARSON, G.A., 1991. Silieification of fossils, 455-499. In Taphonomy: Releasing data locked in the fossil record, P.A. ALLtSON & D.E.G BRIGGS, eds, Plenum Press, New York, 560 p.
CHAFFEE, C. & LINDBERG, D.R., 1986. Larval biology of Early Cambrian molluscs: the implication of small body size. Bulletin of Marine Science 39, 536-549.
COBBOLD, E.S., 1921 The Cambrian horizons of Comley (Shropshire) and their Brachiopoda, Pteropoda, Gasteropoda and etc. The Quarterly Journal of the Geological Society of London 76, 325-386.
CUWER, (3., 1797. Tableau il~mentarie de l'historie naturelle des animaux. Baudouin, Paris, 710 p.
DALZrm, I., DALLA SALDA, L. & GAHAGAN, L., 1994. Paleozoic Laurentia-Gondwana interaction and the origin of the Appalachian-Andean mountain system. Geological Society of America Bulletin 106, 243-252.
DJOUADt, M.T., GLEtZES, G, FERRE, E., BOOCHEZ, J.L., CABY,
R. & LESQUER, A., 1997. Oblique magmatic structures of two epizonal granite plutons, Hoggar, Algeria: late-orogenic emplacement in a transcurrent orogen. Tectonophysics 279, 351-374.
Dz~ J., 1994. Evolution of 'small shelly fossils' assemblages. Acta Palaeontologica Polonica 3, 247-313.
ESAKOVA, N.V. & ZHEGALLO, E.A., 1996. Biostratigrafia I fauna nizhnego kembriya Mongolii (Biostratigraphy and fauna of Lower Cambrian of Mongolia). Nauka, Moscow, 216 p. (Russian).
GEYER, G., 1986. Mittelkambrische Mollusken aus Marokko und Spanien. Senckenbergiana lethaea 67, 55-118.
GEVER, G., 1994. Middle Cambrian molluscs from Idaho and early conchiferan evolution. New York State Museum Bulletin 481, 69-86.
GEYER, G. & SHERGOLD, J., 2000. The quest for internationally recognized divisions of Cambrian time. Episodes 23, 188-195.
GUBANOV, A.P., 1998. The Early Cambrian molluscan evolution and its palaeogeographic implications. Acta Universitatis Carolinae, Geologica 42, 419-422.
GUBANOV, A.P., 2002. Early Cambrian palaeogeography and the lberia-Siberia connection. Tectonophysics 347, 153-168.
GUaANOV, A.E & PEEL, J.S., 1999. Oelandiella, the earliest Cambrian helcionelloid mollusc from Siberia. Palaeontology 42, 211-222.
GRAaAU, A.W. & SmMER, H.W., 1909. North American Index Fossils. Invertebrates 1. New York, 853 p.
HE TINGGUI, PE! FANG & Fu GUANGHONG., 1984. Some small shelly fossils from the Lower Cambrian Xinji Formation in Fangcheng County, Henan Province. Acta Palaeontologica Sinica 23, 350-357 (Chinese).
HE TINGGUI & YANG XIANHE., 1982. Lower Cambrian Meishueun Stage of the western Yangtze stratigraphic region and its small shelly fossils. Bulletin of Chengdu Institute of Geology and Mineral Resources, Chinese Academy of Geological Sciences 3, 69-95. (Chinese).
HEFFERAN, K.P., ADMOU, H., KARSON, J.A. & SAQUAQUE, A., 2000. Anti-Atlas (Morocco) role in Neoprnterozoic Western Gondwana reconstruction. Precambrian Research 103, 89-96.
HICKS, H. 1872. On some undescribed fossils from the Menevian Group. Quarterly Journal of the Geological Society of London 28, 173-185.
Hmz, l., 1987. The Lower Cambrian mierofauna of Comley and Rushton, Shropshire/England. Palaeontographica A 198, 41-100.
HINz-SCHALLREUTER, I., 1997. Einsangstutzen oder Auspufl?. Das R~tsel urn Yochelcionella (Mollusca, Kambrium). Geschiebekunde aktuell 13, 105-122.
HINz-ScHALLREUTER, I., 2000. Middle Cambrian Bivalvia from Bornholm and a review of Cambrian bivalved Mollusca. Revista Espa~ola de Micropaleontologla 32, 225-242.
INESON, J.R. & PEEL, J.S., 1997. Cambrian shelf stratigraphy of North Greenland. Geology of Greenland Survey Bulletin 173, 120 p.
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

ALCHERINGA SIBERIAN M I D D L E C A M B R I A N M O L L U S C S 19
JELL, P.A., 1975. Australian Middle Cambrian endiscids with a review of the Superfamily. Palaeontographica A 150, 1-97.
LANOn~G, E. & BARrOWSKI, K.E., 1996. Oldest shelly fossils from the Taconie Allochthon and late Early Cambrian sea-levels in eastern Laurentia. Journal of Paleontology 70, 741-761.
LANDING, E., G~WR, G. & BARTOWSKI, K.E., 2002. Latest Early Cambrian small shelly fossils, trilobites, and Hatch Hill dysaerobic interval on the Qurbec continental slope. Journal of Paleontology 76, 287- 305.
MAcKINNON, D.I., 1982. Taurangia paparua n. gen. and n. sp., a late Middle Cambrian pelecypod from New Zealand. Journal of Paleontology 56, 589-598.
MACRON, D.I., 1985. New Zealand late Middle Cambrian molluscs and the origin of Rostroconehia and Bivalvia. Alcheringa 9, 65-81.
McKem~ow, W.S., SCOXESE, C.R. & B~SmR, M.D., 1992. Early Cambrian continental reconstructions. Journal of the Geological Society of London 149, 599-606.
MISSARZnEVSKV, V.V., 1989. Drevnejshie skeletnye okamenelosti i stratigrafiya pogranichnykh tolshch dokembriya i kembriya (Oldest skeletal fossils and stratigraphy of Precambrian and Cambrian boundary beds). Nauka, Moscow, 237 p. (Russian).
MISS^~EVSKV, V.V. & MAMBETOV, A.M., 1981. Stratigrafiya l fauna pogranichnykh sloev kembriya 1 dokembriya Malogo Karatau (Stratigraphy and fauna of Cambrian and Precambrian boundary beds of Maly Karatau). Nauka, Moscow, 92 p. (Russian).
NANCE, D.R. & MURPm¢, B.J., 1994. Contrasting basement isotopic signatures and the palinspastic restoration of peripheral orogens: Example from the Neoproterozoic Avalonia-Cadomian belt. Geology 22, 6 i 7-620.
PARKHAEV, P.Y., 2001. Molluscs and siphonoeonchs, 133- 210. In The Cambrian biostratigraphy of the Stansbury Basin, South Australia, E.M. ALEXANDER, J.B. JAGO, A.Y. ROZANOV & A.Y. ZHtmAVLEV, eds, IAPC Nauka/Interperiodica, Moscow.
PE~L, J.S., 1979. Protowenella (Mollusca) from the Cambrian of Peary Land, eastern North Greenland. Rapport Granlands Geologiske Undersogelse 91, 92 (only).
PE~, J.S., 1989. A Lower Cambrian Eotebenna (Mollusca) from Arctic North America. Canadian Journal of Earth Sciences 26, 1501-1503.
PEEL, J.S., 1991a. Functional morphology of the Class Helcionelloida nov., and the early evolution of the Mollusea. In The early evolution of Metazoa and the significance of problematic taxa, A. S~oNnrr^ & S.C. MORRIS, eds, Cambridge University Press and University of Camerino, 157-177.
P~EL, J.S., 1991b. The Classes Tergomya and Helcionelloida, and early molluscan evolution. Gronlands Geologiske Undersogelse Bulletin 161, 11-65.
PEt FANG, 1985. First discovery of Yochelcionella from
the Lower Cambrian of China and its significance. Acta Micropalaeontologica Sinica 2, 395-400.
PEL~CHATY S.M., 1996. Stratigraphic evidence for the Siberian-Laurentia connection and Early Cambrian rifting. Geology 24, 719-722.
POWELL, C.MCA., 1995. Are Neoproterozoic glacial deposits preserved on the margins of Laurentia related to the fragmentation of two supercontinents? Comment. Geology 23, 1053-1054.
ROnlSON, R.A., 1994. Agnostoid trilobites from the Henson Gletscher and Kap Stanton formations (Middle Cambrian), North Greenland. Bulletin Gronlands Geologiske Undersogelse 169, 25-77.
ROZANOV, A.Y., MISSARZHEVSKY, V.V., VOLKOVA, N.A., VORONOVA, L.G., K~VLOV, I.N., I~Lt£R, B.M., KOROLVtaC, I.K., LENDZION, K., MICHN1AK, R., PYCHOVA, N.G. & SIDOROV, A.D., 1969. Tommotskij yarus i problema nizhnej granitsy kembriya (The Tommotian Stage and the Cambrian lower boundary problem). Nauka, Moskva. 380 p. (Russian).
RUNNEGAR, B., 1983. Molluscan phylogeny revisited. Association of Australasian Palaeontologists, Memoir 1, 121-144.
RUNNEGAR, B., 1985. Shell microstructures of Cambrian molluscs replicated by phosphate. Alcheringa 9, 245- 257.
RUNNEGAR, B. & JELL, P.A., 1976. Australian Middle Cambrian molluscs and their bearing on early molluscan evolution. Aleheringa 1, 109-138.
RUNNEGAR, B. & JELL, P.A., 1980. Australian Middle Cambrian molluscs: corrections and additions. Alcheringa 4, 1 I-113.
RUNNEGAR, B. & POJETA, J., Jr, 1980. The monoplacophoran mollusk Yochelcionella identified from the Lower Cambrian of Pennsylvania. Journal of Paleontology 54, 635-636.
RUNNEOAR, B. & POJETA, J.Jr., 1985. Origin and diversification of the Mollusca, 1-57. In The Mollusca 10, Evolution, E.R. TRUI~M^N & M.R. CLARr,£, eds, Academic Press, Orlando.
ScnMrrr, J.G & Bovo, D.W., 1981. Patterns of silicification in Permian peleeypods and brachiopods from Wyoming. Journal of Sedimentary Petrology 51, 1297-1308.
SCHtm~RT, J.K., IOODER, D.L., & ERWrN, D.H., 1997. Silica- replaced fossils through the Phanerozoic. Geology 25, 1031-1034.
SHERGOLD, J.H., LAURIE, J.R. & SUN XIAOWEN, 1990. Classification and review of the trilobite order Agnostida Salter, 1864: an Australian perspective. Bureau of Mineral Resources (Australia) Report 296, 93 p.
VAL'KOV, A.K., 1975. Biostratigrafiya i khiolity kembriya severo-vostoka Sibirskoi platformy (Cambrian biostratigraphy and hyoliths of the northeastern part of the Siberian Platphorm). Moscow, Nauka, 139 p. (Russian).
VASSILJEVA, N.I., 1990. Novye rannekembriiskie bryukhonogie molluski Sibirskoi platformy (New
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009

20 GUBANOV, KOUCHINSKY, PEEI,&BENGTSON ALCHER1NGA
Early Cambrian gastropods of the Siberian Platform). In Voprosy sistematiki I biostratigrafii (Problems of the Systematics and Biostratigraphy), A.I. NmOLAEV, ed., Leningrad, VNIGRI, 4-21 (Russian).
VASSXLJEVA, N.I., 1996. Melkaya rakovinnaya fauna 1 biostratigrafiya nizhnego kembriya Sibirskoi platformy (Lower Cambrian Small Shelly Fossils and biostratigraphy of the Siberian Platform). St. Petersburg, VNIGRI, 139 p. (Russian).
VORONOVA, L.G, DROZDOVA, N.A., E~AKOVA, N.V., ZHEGALLO, E.A., ZHURAVLEV, A.Y., ROZANOV, A.Y., SAYUTINA, T.A.
USHATINSKAYA, G.T., 1987. Iskopaemye nizhnego kembriya got Makkenzi (Kanada) (Lower Cambrian fossils of the Mackenzie Mountains (Canada). Moscow, Nauka, 88 p. (Russian).
VOSTOKOVA, V.A., 1962. Kembrijskie gastropody Sibirskoj platformy i Tajmyra (Cambrian gastropods from Siberian Platform and Taimyr). Trudy Nauchno
Issledovatel'skogo Instituta Geologii Arktiki 28, 51- 74 (Russian).
WAGGONER, B., 1999. Biogeographic analyses of the Ediaeara biota: a conflict with paleotectonic reconstructions. Paleobiology 25, 440-458.
WAtCOTT, C.D., 1891. The fauna of the Lower Cambrian Olenellus Zone. U. S. Geological Survey Annual Report 10, 509-761.
WENZ, W., 1938. Gastropoda. Allgemeiner Tell und Prosobranchia. ! -720. In Handbuch der Paliiozoologie. Band 6. Borntrager, O.H. SCHtNDEWOLF, ed., Berlin, 1639p.
ZHOO BENHE & X1AO Lt6ONG, 1984. Early Cambrian monoplacophorans and gastropods from Huainan and Huoqiu counties, Anhui Province. Professional Papers of Stratigraphy and Palaeontology, Chinese Academy of Geological Sciences 13, 125-140 (Chinese).
Downloaded By: [Univ of Cambridge] At: 13:11 3 June 2009





























