Embed Size (px)
Citation preview

Microsphere size effects on embryoid body incorporationand embryonic stem cell differentiation
Richard L. Carpenedo,1* Scott A. Seaman,1* Todd C. McDevitt1,21The Wallace H. Coulter Department of Biomedical Engineering, Georgia Institute of Technology/Emory University,Atlanta, Georgia2The Parker H. Petit Institute for Bioengineering and Bioscience, Georgia Institute of Technology, Atlanta, Georgia
Received 14 July 2009; revised 28 September 2009; accepted 22 October 2009Published online 8 March 2010 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jbm.a.32710
Abstract: Differentiation of pluripotent embryonic stemcells (ESCs) in vitro via multicellular spheroids calledembryoid bodies (EBs) is commonly performed to modelaspects of early mammalian development and initiate dif-ferentiation of cells for regenerative medicine technolo-gies. However, the three-dimensional nature of EBs posesunique challenges for directed ESC differentiation, includ-ing limited diffusion into EBs of morphogenic moleculescapable of specifying cell fate. Degradable polymer micro-spheres incorporated within EBs can present morphogenicmolecules to ESCs in a spatiotemporally controlled man-ner to more efficiently direct differentiation. In this study,the effect of microsphere size on incorporation into EBsand ESC differentiation in response to microsphere-mediated morphogen delivery were assessed. PLGAmicrospheres with mean diameters of 1, 3, or 11 lm were
fabricated and mixed with ESCs during EB formation.Smaller microspheres were incorporated more efficientlythroughout EBs than larger microspheres, and regardlessof size, retained for at least 10 days of differentiation.Retinoic acid release from incorporated microspheresinduced EB cavitation in a size-dependent manner, withsmaller microspheres triggering accelerated and morecomplete cavitation than larger particles. These resultsdemonstrate that engineering the size of microspheredelivery vehicles incorporated within stem cell environ-ments can be used to modulate the course of differentia-tion. � 2010 Wiley Periodicals, Inc. J Biomed Mater Res94A: 466–475, 2010
Key words: embryoid body; embryonic stem cell; micro-sphere; differentiation; retinoic acid
INTRODUCTION
Embryonic stem cells (ESCs) are pluripotent cellsderived from the inner cell mass of developing blas-tocysts with the inherent ability to differentiate intoall somatic cell types.1–4 The pluripotent nature ofESCs has attracted interest in these cells as a renew-able cell source for tissue engineering and other cellbased therapies, as well as for in vitro drug screening
and studies in mammalian development.5–7 Differen-tiation of ESCs can be induced using a variety ofmethods, including monolayer culture,8–10 co-culturewith stromal cells,11–13 and aggregation into multicel-lular spheroids called embryoid bodies (EBs).1,3,14
The use of EBs for ESC differentiation is particularlycommon because of their ability to differentiate tocells from all three germ lineages and similarities toembryonic development; however, the microenviron-ment within EBs is complex and dynamic, resultingin spontaneous and heterogeneous differentiationevents.15 Efforts to control and direct differentiationof ESCs comprising EBs have traditionally focusedon the addition of soluble morphogens to the culturemedia.16,17 While soluble treatment can improve theefficiency of directed differentiation to specific celltypes, a high degree of heterogeneity is still com-mon. Studies investigating the diffusion of moleculesinto tumor spheroids18–21 and EBs22–24 have shownthat diffusion into cell spheroids is hindered byspheroid size, extracellular matrix (ECM) content,and tight cell junctions. The inability of morphogens
Additional Supporting Information may be found in theonline version of this article.*These authors contributed equally to this work.Correspondence to: T. C. McDevitt; e-mail: todd.mcdevitt@
bme.gatech.eduContract grant sponsor: National Science Foundation;
contract grant number: CBET 0651739Contract grant sponsor: Undergraduate Research Schol-
ars program; contract grant number: GTEC, NSF EEC-9731463Contract grant sponsors: Presidential Undergraduate
Research Award, George Family Foundation fellowship
� 2010 Wiley Periodicals, Inc.

to effectively penetrate cell spheroids limits theefficacy of soluble morphogen delivery strategies forEB-based differentiation.
Recently, two approaches have been examined toovercome the diffusional limitations posed by EBs.One approach has been to disrupt ECM in EBsusing proteases to enhance morphogen penetrationinto EBs.23 However, ECM is an important compo-nent of the 3D microenvironment of EBs, and itsdisruption may negatively impact cell organization,viability, and differentiation. Additionally, thekinetics of ECM degradation and remodeling inEBs have not been characterized, indicating that thetimescale during which enhanced diffusion into EBsis permitted is unknown. Alternatively, the diffu-sional barriers posed by EBs have been circum-vented by incorporating biodegradable polymermicrospheres containing morphogenic moleculesdirectly within the interior of EBs.25,26 Such anapproach facilitates the presentation of morphogensto ESCs directly within the EB microenvironmentrather than relying on passive diffusion to transportmorphogens into the EB interior. ESCs comprisingEBs have been shown to tolerate the incorporationof PLGA microspheres without compromised viabil-ity.25,26 Delivery of retinoic acid (RA) via incorpo-rated microspheres has been shown to inducemorphological and phenotypic changes in EBs notobserved using traditional soluble delivery.25 Addi-tionally, microsphere parameters, including materialcomposition, molecular weight, degradation rate,and size, can be tailored to produce desirablerelease profiles and maintain bioactivity of variousencapsulated molecules. Although synthetic poly-mer particles of varying size (nano-micro) havebeen successfully mixed with ESCs and incorpo-rated within EBs, the effects of microsphere size onincorporation into EBs and differentiation due tomorphogen release have not been systematicallyexamined.
In this study, three distinct size populations ofPLGA microspheres (�1, 3, and 11 lm diameter)were fabricated and mixed with ESCs during EBformation under rotary orbital shaking conditions.The extent of microsphere incorporation and spatialdistribution within EBs were assessed as a functionof microsphere size. In addition, the release andcell uptake of a fluorescent dye (CellTracker Red)encapsulated within microspheres was assessedover the course of 10 days of differentiation. Mor-phological changes within EBs in response to deliv-ery of RA using the different size microsphereswere also examined. The results of this study indi-cate that microsphere size is an important parame-ter for microsphere-mediated delivery of morpho-genic factors within EBs for directed stem celldifferentiation.
MATERIALS AND METHODS
Microsphere fabrication
Poly(lactic-co-glycolic acid) (PLGA, 50:50, 0.55–0.75dL/g, ester terminated, Absorbable Polymers Interna-tional) microspheres encapsulating either CellTrackerRed (Molecular Probes, Eugene, OR) or retinoic acid(all trans, Acros Organics, Geel, Belgium) were fabricatedusing a single emulsion solvent evaporation technique asdescribed previously.25 Briefly, PLGA dissolved indichloromethane (DCM) (20 mg/mL) was added to a 0.3%poly(vinyl alcohol) solution and different sized micro-spheres were formulated by homogenization at 1000, 5000,or 12,000 RPM (Polytron PT 3100, Kinematica, Bohemia,NY). The residual organic solvent was evaporated underlight stirring for 4 hours before the microspheres werelyophilized for 24–36 hours (Freezone 4.5, Labconco,Kansas City, MO) and stored at 2208C until use.
Scanning electron microscopy (SEM)
Microspheres suspended in dH2O were attached to SEMstubs with carbon double-sided adhesive discs (SPI Sup-plies, West Chester, PA), frozen at 2808C and lyophilizedfor �6 hours until stubs were completely dehydrated.After lyophilization, stubs were gold sputter coated usinga Polaron range sputter coater (Quorum Technologies, EastSussex, UK) and imaged using Hitachi S-800 SEM micro-scope. Microsphere size was quantified using ImageJ soft-ware to measure the diameter of a minimum of 100 micro-spheres for each group. The resulting distribution ofmicrosphere sizes was plotted as a histogram with anequal number of bins between the largest and smallest val-ues for each experimental group.
CellTracker release profile
Microspheres from the three size distributions were sus-pended in PBS (pH 7.4) in microcentrifuge tubes andplaced on a rotisserie rotator at 378C. After various timepoints, microspheres were collected via centrifugation(3000 rpm, 5 min) and the supernatant was removed andanalyzed via fluorescent spectroscopy (SpectraMax M2e,Molecular Devices, Sunnyvale, CA). The concentration ofreleased CellTracker Red was determined using a standardcurve. To determine initial CellTracker loading, micro-spheres were suspended in an 0.1M NaOH/ 5% sodiumdodecyl sulfate (SDS) solution, rotated overnight at 378C,and the lysate was analyzed in a fluorescent plate readerand compared with a standard curve of CellTracker inNaOH/SDS solution. The stability of CellTracker fluores-cence following overnight incubation in NaOH/SDS at378C was confirmed spectrofluorometrically. Briefly, Cell-Tracker Red was dissolved in NaOH/SDS solution andstandard curves were constructed both immediately andafter overnight incubation at 378C under rotation, with nosignificant difference observed between the two conditions.Based on the initial loading and the concentration of
MICROSPHERE SIZE EFFECTS ON EMBRYOID BODIES 467
Journal of Biomedical Materials Research Part A

released CellTracker, a profile of cumulative CellTrackerreleased for each microsphere population was constructedover the course of 14 days.
Embryonic stem cell culture
ECSs (D314) were grown on 0.1% gelatin coated tissueculture dishes in Dulbecco’s modified Eagle’s medium(DMEM) (Mediatech., Manassas, VA). The media was sup-plemented with 15% fetal bovine serum (Atlanta Biologi-cals, Lawrenceville, GA), 0.1 mM 2-mercaptoethanol(MP Biomedicals, LLC, Solon, OH), 100 U/mL penicillin(MP Biomedicals, LLC, Solon, OH), 100 lg/mL strepto-myocin (MP Biomedicals, LLC, Solon, OH), 0.25 lg/mLamphotericin (MP Biomedicals, LLC, Solon, OH), 13 MEMnonessential amino acid solution (Lonza, Walkersville,MD), 2 mM L-glutamine (Mediatech, Manassas, VA), and103 U/mL leukemia inhibitory factor (Millipore, Temecula,CA.) Cells were passaged routinely every 2–3 days beforereaching 70% confluence.
Embryoid body formation
Equal masses of the different size microspheres weresuspended in a 0.1% gelatin solution and placed on a rotis-serie rotator at 378C for 3 hours to coat the microsphereswith an adhesive molecule. Gelatin-coated microsphereswere collected by centrifugation at 3000 RPM for 5 min,and resuspended in differentiation media (ESC mediawithout LIF). Embryonic stem cell colonies were dissoci-ated into single cells with a 0.05% trypsin-EDTA solutionand resuspended in 10 mL of differentiation media (2 3105 cells/mL final concentration) with 2 mg of gelatin-coated microspheres in 100 mm bacteriological-grade Petridishes. Embryoid body formation was initiated by placingdishes on an orbital rotary shaker set at 40 rotations perminute (RPM).27 Phase images of EBs were acquired after2 days using a Nikon TE 2000 inverted microscopeequipped with a SpotFLEX (Diagnostic Instruments) cam-era. Image J software was used to quantify the cross-sectional area of EBs formed with and without micro-spheres.
Microsphere incorporation quantification
After 2 days of rotary suspension culture, EBs were col-lected by gravity-induced sedimentation in conical tubes,spent media containing unincorporated microspheres wasaspirated, and EBs were resuspended in 10 mL of freshmedia. Phase and fluorescent images were obtained asaforementioned. A fraction of the EBs (1/10) were countedusing a Nikon TE 2000 inverted microscope after beingsuspended in a 60:40 PBS:glycerol solution. The remainingEBs were suspended in 5% SDS 0.1M NaOH solution androtated overnight at 378C in order to lyse the EBs and hy-drolyze the microspheres to release encapsulated fluores-cent dye (CellTracker Red). The number of microspheresincorporated within EBs was determined using a standardcurve relating fluorescence intensity to a known number of
microspheres hydrolyzed in NaOH/SDS overnight at378C. Fluorescence readings were acquired with a micro-plate reader (SpectraMax M2e, 586 nm excitation, 613 nmemission) and average fluorescence readings were plottedagainst initial microsphere concentration to determine theaverage number of incorporated microspheres per EB.
Confocal microscopy analysis
EBs harvested after 2 and 7 days of differentiation werefixed in 10% formalin for 30 min at room temperature on arotisserie rotator, rinsed three times in PBS and permeabil-ized in a 1% Triton/PBS solution for 10 min with rotation.The fixed EB samples were stained with a 1:50 Phalloidin(FITC) (Alexis Biochemicals, San Diego, CA) and Hoechst(10 lg/mL) solution in PBS for 30 min. Samples werestored in PBS at 48C until imaging was performed on aZeiss LSM 510 Confocal Microscope.
Histological analysis
EBs with incorporated CellTracker Red and retinoic acidmicrospheres were harvested at days 7 and 10 of differen-tiation and fixed in 10% formalin for 30 min on a rotisserierotator at room temperature. Following fixation, the sam-ples were washed three times with PBS and re-suspendedin HistoGel (Richard-Allan Scientific, Kalamazoo, MI).Fixed samples were dehydrated in a series of graduatedalcohol solutions (70–100%) and xylene before being em-bedded in paraffin. Sections (5 lm) were stained with he-matoxylin and eosin (H&E) and imaged using a Nikon 80imicroscope with a SpotFLEX camera.
Statistical analysis
All experiments were performed in triplicate and datais reported as mean 6 standard deviation. Analysis ofvariance (ANOVA) and Tukey’s post hoc analysis wereperformed using Systat (v12, Systat Software).
RESULTS
Microsphere analysis
The morphology and size distribution of differentsized microspheres were assessed to determine therelative sizes of the microspheres fabricated at differ-ent homogenization speeds. As expected, micro-spheres were round with smooth surfaces and theaverage size of the microsphere populationsdecreased with increasing homogenization speed[Fig. 1(A–C)]. The lowest homogenization setting(1000 RPM) yielded microspheres with an averagediameter of 10.9 6 3.2 lm [Fig. 1(D)], whereas5000 RPM produced microspheres with an average
468 CARPENEDO, SEAMAN, AND MCDEVITT
Journal of Biomedical Materials Research Part A
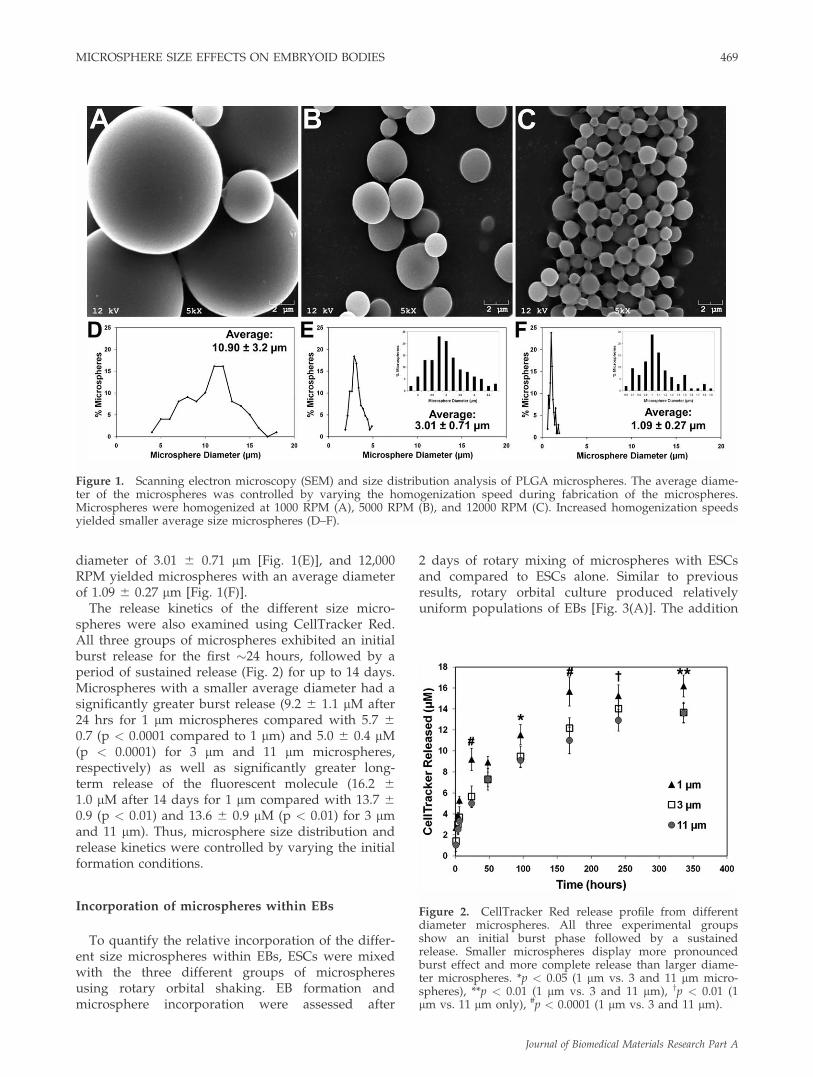
diameter of 3.01 6 0.71 lm [Fig. 1(E)], and 12,000RPM yielded microspheres with an average diameterof 1.09 6 0.27 lm [Fig. 1(F)].
The release kinetics of the different size micro-spheres were also examined using CellTracker Red.All three groups of microspheres exhibited an initialburst release for the first �24 hours, followed by aperiod of sustained release (Fig. 2) for up to 14 days.Microspheres with a smaller average diameter had asignificantly greater burst release (9.2 6 1.1 lM after24 hrs for 1 lm microspheres compared with 5.7 60.7 (p < 0.0001 compared to 1 lm) and 5.0 6 0.4 lM(p < 0.0001) for 3 lm and 11 lm microspheres,respectively) as well as significantly greater long-term release of the fluorescent molecule (16.2 61.0 lM after 14 days for 1 lm compared with 13.7 60.9 (p < 0.01) and 13.6 6 0.9 lM (p < 0.01) for 3 lmand 11 lm). Thus, microsphere size distribution andrelease kinetics were controlled by varying the initialformation conditions.
Incorporation of microspheres within EBs
To quantify the relative incorporation of the differ-ent size microspheres within EBs, ESCs were mixedwith the three different groups of microspheresusing rotary orbital shaking. EB formation andmicrosphere incorporation were assessed after
2 days of rotary mixing of microspheres with ESCsand compared to ESCs alone. Similar to previousresults, rotary orbital culture produced relativelyuniform populations of EBs [Fig. 3(A)]. The addition
Figure 1. Scanning electron microscopy (SEM) and size distribution analysis of PLGA microspheres. The average diame-ter of the microspheres was controlled by varying the homogenization speed during fabrication of the microspheres.Microspheres were homogenized at 1000 RPM (A), 5000 RPM (B), and 12000 RPM (C). Increased homogenization speedsyielded smaller average size microspheres (D–F).
Figure 2. CellTracker Red release profile from differentdiameter microspheres. All three experimental groupsshow an initial burst phase followed by a sustainedrelease. Smaller microspheres display more pronouncedburst effect and more complete release than larger diame-ter microspheres. *p < 0.05 (1 lm vs. 3 and 11 lm micro-spheres), **p < 0.01 (1 lm vs. 3 and 11 lm), yp < 0.01 (1lm vs. 11 lm only), #p < 0.0001 (1 lm vs. 3 and 11 lm).
MICROSPHERE SIZE EFFECTS ON EMBRYOID BODIES 469
Journal of Biomedical Materials Research Part A
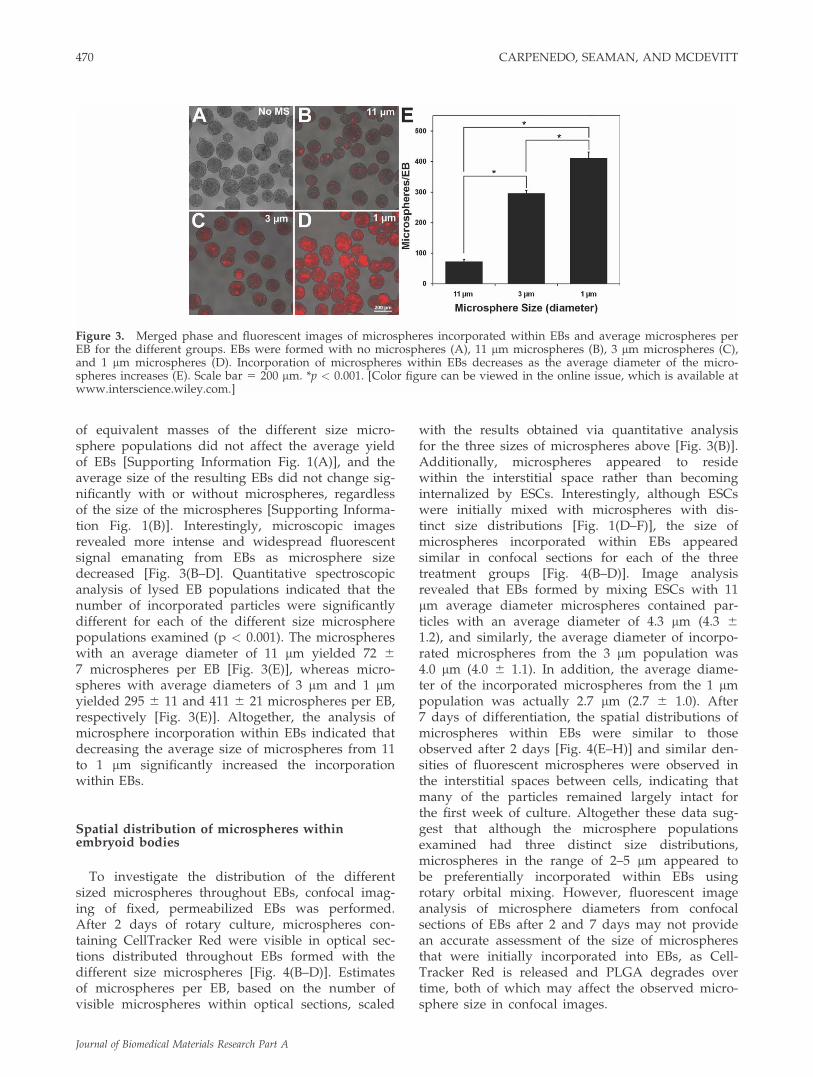
of equivalent masses of the different size micro-sphere populations did not affect the average yieldof EBs [Supporting Information Fig. 1(A)], and theaverage size of the resulting EBs did not change sig-nificantly with or without microspheres, regardlessof the size of the microspheres [Supporting Informa-tion Fig. 1(B)]. Interestingly, microscopic imagesrevealed more intense and widespread fluorescentsignal emanating from EBs as microsphere sizedecreased [Fig. 3(B–D]. Quantitative spectroscopicanalysis of lysed EB populations indicated that thenumber of incorporated particles were significantlydifferent for each of the different size microspherepopulations examined (p < 0.001). The microsphereswith an average diameter of 11 lm yielded 72 67 microspheres per EB [Fig. 3(E)], whereas micro-spheres with average diameters of 3 lm and 1 lmyielded 295 6 11 and 411 6 21 microspheres per EB,respectively [Fig. 3(E)]. Altogether, the analysis ofmicrosphere incorporation within EBs indicated thatdecreasing the average size of microspheres from 11to 1 lm significantly increased the incorporationwithin EBs.
Spatial distribution of microspheres withinembryoid bodies
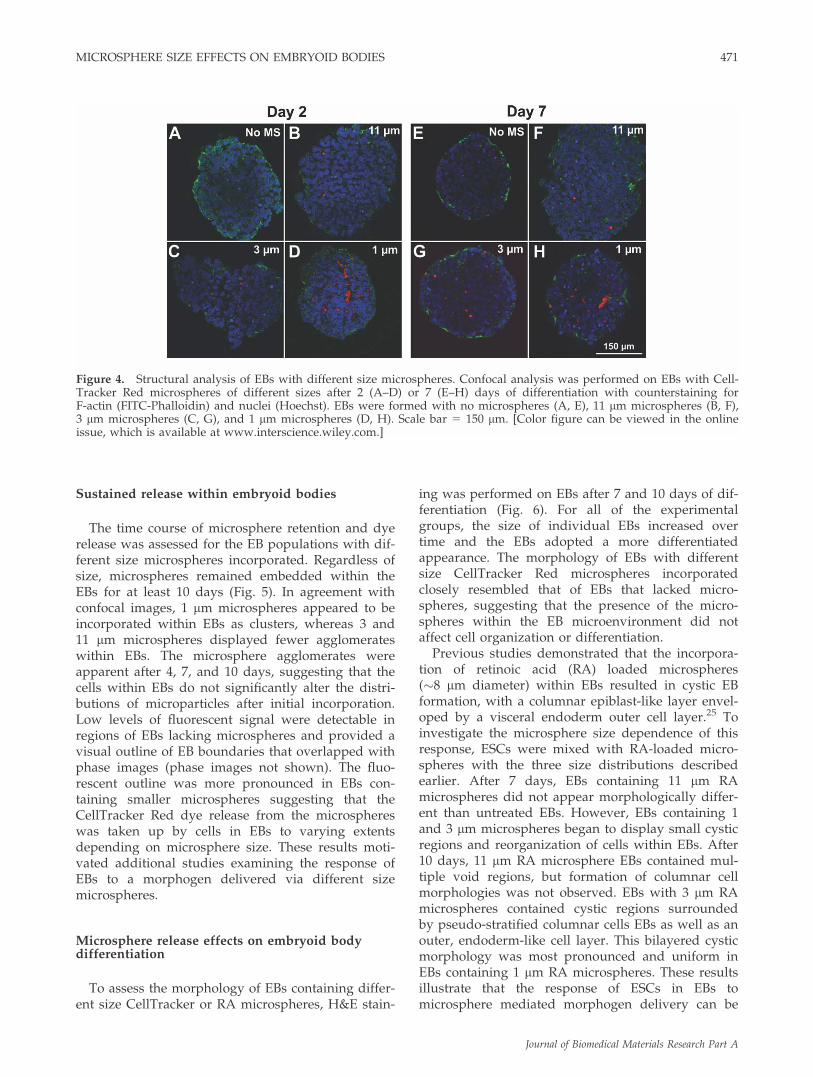
To investigate the distribution of the differentsized microspheres throughout EBs, confocal imag-ing of fixed, permeabilized EBs was performed.After 2 days of rotary culture, microspheres con-taining CellTracker Red were visible in optical sec-tions distributed throughout EBs formed with thedifferent size microspheres [Fig. 4(B–D)]. Estimatesof microspheres per EB, based on the number ofvisible microspheres within optical sections, scaled
with the results obtained via quantitative analysisfor the three sizes of microspheres above [Fig. 3(B)].Additionally, microspheres appeared to residewithin the interstitial space rather than becominginternalized by ESCs. Interestingly, although ESCswere initially mixed with microspheres with dis-tinct size distributions [Fig. 1(D–F)], the size ofmicrospheres incorporated within EBs appearedsimilar in confocal sections for each of the threetreatment groups [Fig. 4(B–D)]. Image analysisrevealed that EBs formed by mixing ESCs with 11lm average diameter microspheres contained par-ticles with an average diameter of 4.3 lm (4.3 61.2), and similarly, the average diameter of incorpo-rated microspheres from the 3 lm population was4.0 lm (4.0 6 1.1). In addition, the average diame-ter of the incorporated microspheres from the 1 lmpopulation was actually 2.7 lm (2.7 6 1.0). After7 days of differentiation, the spatial distributions ofmicrospheres within EBs were similar to thoseobserved after 2 days [Fig. 4(E–H)] and similar den-sities of fluorescent microspheres were observed inthe interstitial spaces between cells, indicating thatmany of the particles remained largely intact forthe first week of culture. Altogether these data sug-gest that although the microsphere populationsexamined had three distinct size distributions,microspheres in the range of 2–5 lm appeared tobe preferentially incorporated within EBs usingrotary orbital mixing. However, fluorescent imageanalysis of microsphere diameters from confocalsections of EBs after 2 and 7 days may not providean accurate assessment of the size of microspheresthat were initially incorporated into EBs, as Cell-Tracker Red is released and PLGA degrades overtime, both of which may affect the observed micro-sphere size in confocal images.
Figure 3. Merged phase and fluorescent images of microspheres incorporated within EBs and average microspheres perEB for the different groups. EBs were formed with no microspheres (A), 11 lm microspheres (B), 3 lm microspheres (C),and 1 lm microspheres (D). Incorporation of microspheres within EBs decreases as the average diameter of the micro-spheres increases (E). Scale bar 5 200 lm. *p < 0.001. [Color figure can be viewed in the online issue, which is available atwww.interscience.wiley.com.]
470 CARPENEDO, SEAMAN, AND MCDEVITT
Journal of Biomedical Materials Research Part A
Sustained release within embryoid bodies
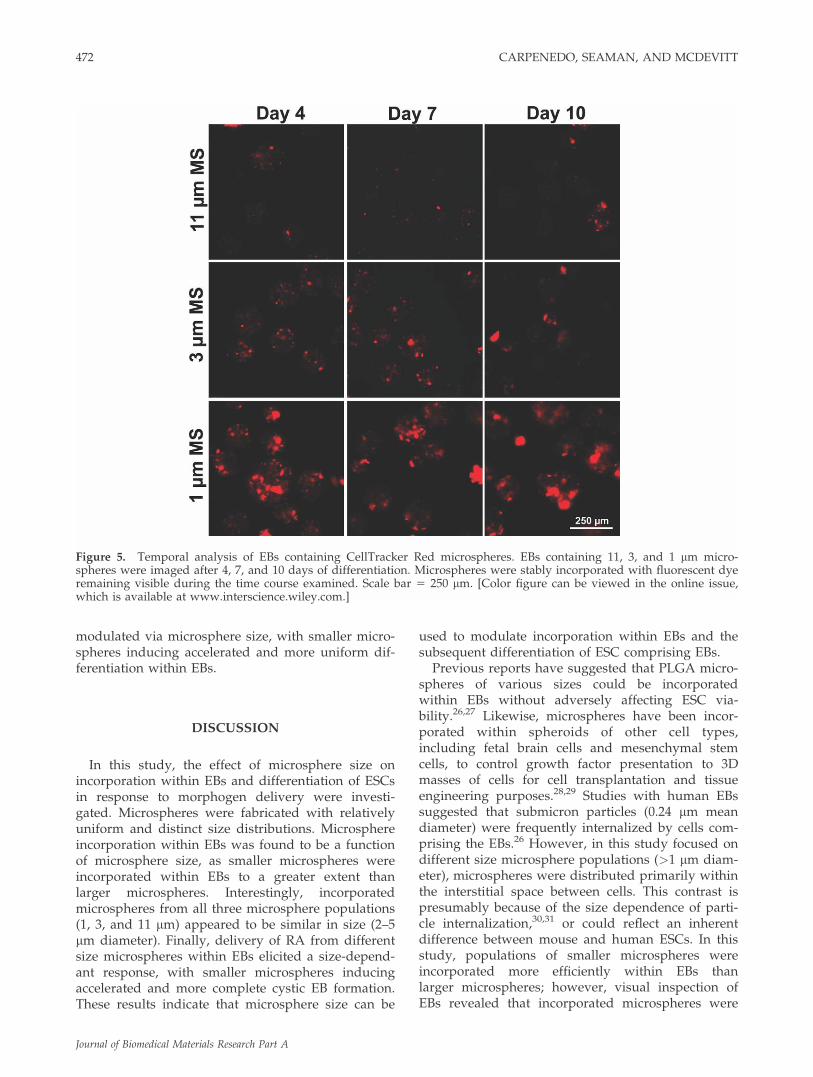
The time course of microsphere retention and dyerelease was assessed for the EB populations with dif-ferent size microspheres incorporated. Regardless ofsize, microspheres remained embedded within theEBs for at least 10 days (Fig. 5). In agreement withconfocal images, 1 lm microspheres appeared to beincorporated within EBs as clusters, whereas 3 and11 lm microspheres displayed fewer agglomerateswithin EBs. The microsphere agglomerates wereapparent after 4, 7, and 10 days, suggesting that thecells within EBs do not significantly alter the distri-butions of microparticles after initial incorporation.Low levels of fluorescent signal were detectable inregions of EBs lacking microspheres and provided avisual outline of EB boundaries that overlapped withphase images (phase images not shown). The fluo-rescent outline was more pronounced in EBs con-taining smaller microspheres suggesting that theCellTracker Red dye release from the microsphereswas taken up by cells in EBs to varying extentsdepending on microsphere size. These results moti-vated additional studies examining the response ofEBs to a morphogen delivered via different sizemicrospheres.
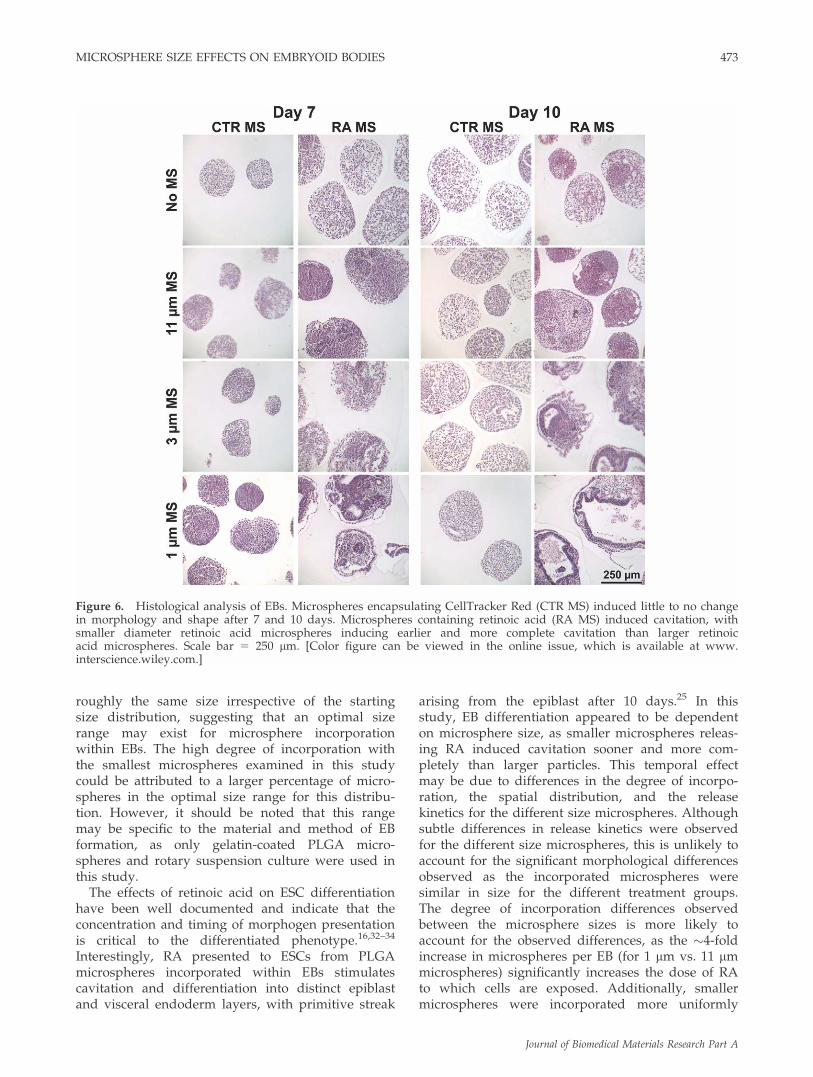
Microsphere release effects on embryoid bodydifferentiation
To assess the morphology of EBs containing differ-ent size CellTracker or RA microspheres, H&E stain-
ing was performed on EBs after 7 and 10 days of dif-ferentiation (Fig. 6). For all of the experimentalgroups, the size of individual EBs increased overtime and the EBs adopted a more differentiatedappearance. The morphology of EBs with differentsize CellTracker Red microspheres incorporatedclosely resembled that of EBs that lacked micro-spheres, suggesting that the presence of the micro-spheres within the EB microenvironment did notaffect cell organization or differentiation.
Previous studies demonstrated that the incorpora-tion of retinoic acid (RA) loaded microspheres(�8 lm diameter) within EBs resulted in cystic EBformation, with a columnar epiblast-like layer envel-oped by a visceral endoderm outer cell layer.25 Toinvestigate the microsphere size dependence of thisresponse, ESCs were mixed with RA-loaded micro-spheres with the three size distributions describedearlier. After 7 days, EBs containing 11 lm RAmicrospheres did not appear morphologically differ-ent than untreated EBs. However, EBs containing 1and 3 lm microspheres began to display small cysticregions and reorganization of cells within EBs. After10 days, 11 lm RA microsphere EBs contained mul-tiple void regions, but formation of columnar cellmorphologies was not observed. EBs with 3 lm RAmicrospheres contained cystic regions surroundedby pseudo-stratified columnar cells EBs as well as anouter, endoderm-like cell layer. This bilayered cysticmorphology was most pronounced and uniform inEBs containing 1 lm RA microspheres. These resultsillustrate that the response of ESCs in EBs tomicrosphere mediated morphogen delivery can be
Figure 4. Structural analysis of EBs with different size microspheres. Confocal analysis was performed on EBs with Cell-Tracker Red microspheres of different sizes after 2 (A–D) or 7 (E–H) days of differentiation with counterstaining forF-actin (FITC-Phalloidin) and nuclei (Hoechst). EBs were formed with no microspheres (A, E), 11 lm microspheres (B, F),3 lm microspheres (C, G), and 1 lm microspheres (D, H). Scale bar 5 150 lm. [Color figure can be viewed in the onlineissue, which is available at www.interscience.wiley.com.]
MICROSPHERE SIZE EFFECTS ON EMBRYOID BODIES 471
Journal of Biomedical Materials Research Part A
modulated via microsphere size, with smaller micro-spheres inducing accelerated and more uniform dif-ferentiation within EBs.
DISCUSSION
In this study, the effect of microsphere size onincorporation within EBs and differentiation of ESCsin response to morphogen delivery were investi-gated. Microspheres were fabricated with relativelyuniform and distinct size distributions. Microsphereincorporation within EBs was found to be a functionof microsphere size, as smaller microspheres wereincorporated within EBs to a greater extent thanlarger microspheres. Interestingly, incorporatedmicrospheres from all three microsphere populations(1, 3, and 11 lm) appeared to be similar in size (2–5lm diameter). Finally, delivery of RA from differentsize microspheres within EBs elicited a size-depend-ant response, with smaller microspheres inducingaccelerated and more complete cystic EB formation.These results indicate that microsphere size can be
used to modulate incorporation within EBs and thesubsequent differentiation of ESC comprising EBs.
Previous reports have suggested that PLGA micro-spheres of various sizes could be incorporatedwithin EBs without adversely affecting ESC via-bility.26,27 Likewise, microspheres have been incor-porated within spheroids of other cell types,including fetal brain cells and mesenchymal stemcells, to control growth factor presentation to 3Dmasses of cells for cell transplantation and tissueengineering purposes.28,29 Studies with human EBssuggested that submicron particles (0.24 lm meandiameter) were frequently internalized by cells com-prising the EBs.26 However, in this study focused ondifferent size microsphere populations (>1 lm diam-eter), microspheres were distributed primarily withinthe interstitial space between cells. This contrast ispresumably because of the size dependence of parti-cle internalization,30,31 or could reflect an inherentdifference between mouse and human ESCs. In thisstudy, populations of smaller microspheres wereincorporated more efficiently within EBs thanlarger microspheres; however, visual inspection ofEBs revealed that incorporated microspheres were
Figure 5. Temporal analysis of EBs containing CellTracker Red microspheres. EBs containing 11, 3, and 1 lm micro-spheres were imaged after 4, 7, and 10 days of differentiation. Microspheres were stably incorporated with fluorescent dyeremaining visible during the time course examined. Scale bar 5 250 lm. [Color figure can be viewed in the online issue,which is available at www.interscience.wiley.com.]
472 CARPENEDO, SEAMAN, AND MCDEVITT
Journal of Biomedical Materials Research Part A
roughly the same size irrespective of the startingsize distribution, suggesting that an optimal sizerange may exist for microsphere incorporationwithin EBs. The high degree of incorporation withthe smallest microspheres examined in this studycould be attributed to a larger percentage of micro-spheres in the optimal size range for this distribu-tion. However, it should be noted that this rangemay be specific to the material and method of EBformation, as only gelatin-coated PLGA micro-spheres and rotary suspension culture were used inthis study.
The effects of retinoic acid on ESC differentiationhave been well documented and indicate that theconcentration and timing of morphogen presentationis critical to the differentiated phenotype.16,32–34
Interestingly, RA presented to ESCs from PLGAmicrospheres incorporated within EBs stimulatescavitation and differentiation into distinct epiblastand visceral endoderm layers, with primitive streak
arising from the epiblast after 10 days.25 In thisstudy, EB differentiation appeared to be dependenton microsphere size, as smaller microspheres releas-ing RA induced cavitation sooner and more com-pletely than larger particles. This temporal effectmay be due to differences in the degree of incorpo-ration, the spatial distribution, and the releasekinetics for the different size microspheres. Althoughsubtle differences in release kinetics were observedfor the different size microspheres, this is unlikely toaccount for the significant morphological differencesobserved as the incorporated microspheres weresimilar in size for the different treatment groups.The degree of incorporation differences observedbetween the microsphere sizes is more likely toaccount for the observed differences, as the �4-foldincrease in microspheres per EB (for 1 lm vs. 11 lmmicrospheres) significantly increases the dose of RAto which cells are exposed. Additionally, smallermicrospheres were incorporated more uniformly
Figure 6. Histological analysis of EBs. Microspheres encapsulating CellTracker Red (CTR MS) induced little to no changein morphology and shape after 7 and 10 days. Microspheres containing retinoic acid (RA MS) induced cavitation, withsmaller diameter retinoic acid microspheres inducing earlier and more complete cavitation than larger retinoicacid microspheres. Scale bar 5 250 lm. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
MICROSPHERE SIZE EFFECTS ON EMBRYOID BODIES 473
Journal of Biomedical Materials Research Part A

throughout EBs than larger microspheres, whichmay contribute to the more pronounced and uniformcavitation in EBs with smaller microspheres com-pared with EBs with larger microspheres.
The effects of polymer microsphere size on releasekinetics have been studied for a variety of drugdelivery applications. In particular, the size of PLGAmicrospheres has been shown to strongly influencethe release kinetics of encapsulated molecules. Stud-ies performed with monodisperse PLGA micro-spheres demonstrate that smaller particles tend todisplay faster release rates with more pronouncedburst release as well as more complete release overthe time courses examined (weeks-months).35,36 Thisis generally attributed to the higher surface area-to-volume ratio of smaller particles, which allows mole-cules near the particle surface to be rapidly released.In this study, no significant differences in releaserate were observed for different size microspheres,presumably because of the relatively small differen-ces between the groups of microspheres examinedand the greater dispersity of the microsphere popu-lations than the monodisperse particle studies citedabove. However, the ability to control morphogenrelease by engineering microsphere size remains asimple approach to modulate the kinetics of morph-ogen presentation and stem cell differentiation.
This work demonstrates that microsphere size isan important parameter that influences incorporationwithin EBs, which in turn affects the differentiationof ESCs in response to morphogen delivery. Addi-tionally, these results provide evidence that simplemodulation of physical properties of deliveryvehicles can be utilized in the development ofdirected differentiation approaches. Moreover, theintegration of biomaterials into stem cell microenvir-onments to control cell fate decisions represents apowerful enabling technology for applications in tis-sue engineering, in vitro diagnostics, developmentalbiology, and stem cell bioprocessing.
References
1. Evans MJ, Kaufman MH. Establishment in culture of pluripo-tential cells from mouse embryos. Nature 1981;292:154–156.
2. Martin GR. Isolation of a pluripotent cell line from earlymouse embryos cultured in medium conditioned by terato-carcinoma stem cells. Proc Natl Acad Sci USA 1981;78:7634–7638.
3. Itskovitz-Eldor J, Schuldiner M, Karsenti D, Eden A, YanukaO, Amit M, Soreq H, Benvenisty N. Differentiation of humanembryonic stem cells into embryoid bodies compromising thethree embryonic germ layers. Mol Med 2000;6:88–95.
4. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA,Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell linesderived from human blastocysts. Science 1998;282:1145–1147.
5. Wobus AM, Boheler KR. Embryonic stem cells: Prospects fordevelopmental biology and cell therapy. Physiol Rev 2005;85:635–678.
6. Keller G. Embryonic stem cell differentiation: Emergence of anew era in biology and medicine. Genes Dev 2005;19:1129–1155.
7. Gorba T, Allsopp TE. Pharmacological potential of embryonicstem cells. Pharmacol Res 2003;47:269–278.
8. Tropepe V, Hitoshi S, Sirard C, Mak TW, Rossant J, van derKooy D. Direct neural fate specification from embryonic stemcells: A primitive mammalian neural stem cell stage acquiredthrough a default mechanism. Neuron 2001;30:65–78.
9. Ying QL, Stavridis M, Griffiths D, Li M, Smith A. Conversionof embryonic stem cells into neuroectodermal precursors inadherent monoculture. Nat Biotechnol 2003;21:183–186.
10. Hwang YS, Polak JM, Mantalaris A. In vitro direct chondro-genesis of murine embryonic stem cells by bypassing embry-oid body formation. Stem Cells Dev 2008;17:971–978.
11. Kaufman DS, Hanson ET, Lewis RL, Auerbach R, ThomsonJA. Hematopoietic colony-forming cells derived from humanembryonic stem cells. Proc Natl Acad Sci USA 2001;98:10716–10721.
12. Buttery LD, Bourne S, Xynos JD, Wood H, Hughes FJ,Hughes SP, Episkopou V, Polak JM. Differentiation of osteo-blasts and in vitro bone formation from murine embryonicstem cells. Tissue Eng 2001;7:89–99.
13. Zeng X, Cai J, Chen J, Luo Y, You ZB, Fotter E, Wang Y, Har-vey B, Miura T, Backman C, Chen GJ, Rao MS, Freed WJ. Do-paminergic differentiation of human embryonic stem cells.Stem Cells 2004;22:925–940.
14. Doetschman TC, Eistetter H, Katz M, Schmidt W, Kemler R.The in vitro development of blastocyst-derived embryonicstem cell lines: Formation of visceral yolk sac, blood islandsand myocardium. J Embryol Exp Morphol 1985;87:27–45.
15. Bratt-Leal AM, Carpenedo RL, McDevitt TC. Engineering theembryoid body microenvironment to direct embryonic stemcell differentiation. Biotechnol Prog 2009;25:43–51.
16. Bain G, Ray WJ, Yao M, Gottlieb DI. Retinoic acid promotesneural and represses mesodermal gene expression in mouseembryonic stem cells in culture. Biochem Biophys Res Com-mun 1996;223:691–694.
17. Schuldiner M, Yanuka O, Itskovitz-Eldor J, Melton DA, Ben-venisty N. Effects of eight growth factors on the differentia-tion of cells derived from human embryonic stem cells. ProcNatl Acad Sci USA 2000;97:11307–11312.
18. Hu G, Li D. Three-dimensional modeling of transport ofnutrients for multicellular tumor spheroid culture in a micro-channel. Biomed Microdevices 2007;9:315–323.
19. Thurber GM, Wittrup KD. Quantitative spatiotemporal analy-sis of antibody fragment diffusion and endocytic consump-tion in tumor spheroids. Cancer Res 2008;68:3334–3341.
20. Ivascu A, Kubbies M. Diversity of cell-mediated adhesions inbreast cancer spheroids. Int J Oncol 2007;31:1403–1413.
21. Davies Cde L, Berk DA, Pluen A, Jain RK. Comparison ofIgG diffusion and extracellular matrix composition in rhabdo-myosarcomas grown in mice versus in vitro as spheroidsreveals the role of host stromal cells. Br J Cancer 2002;86:1639–1644.
22. Wartenberg M, Gunther J, Hescheler J, Sauer H. The embry-oid body as a novel in vitro assay system for antiangiogenicagents. Lab Invest 1998;78:1301–1314.
23. Sachlos E, Auguste DT. Embryoid body morphology influen-ces diffusive transport of inductive biochemicals: A strategyfor stem cell differentiation. Biomaterials 2008;29:4471–4480.
24. Wartenberg M, Donmez F, Ling FC, Acker H, Hescheler J,Sauer H. Tumor-induced angiogenesis studied in confronta-tion cultures of multicellular tumor spheroids and embryoidbodies grown from pluripotent embryonic stem cells. FASEBJ 2001;15:995–1005.
25. Carpenedo RL, Bratt-Leal AM, Marklein RA, Seaman SA,Bowen NJ, McDonald JF, McDevitt TC. Homogeneous and
474 CARPENEDO, SEAMAN, AND MCDEVITT
Journal of Biomedical Materials Research Part A

organized differentiation within embryoid bodies induced bymicrosphere-mediated delivery of small molecules. Biomate-rials 2009;30:2507–2515.
26. Ferreira L, Squier T, Park H, Choe H, Kohane DS, Langer R.Human embryoid bodies containing nano- and microparticu-late delivery vehicles. Adv Mater 2008;20:2285–2291.
27. Carpenedo RL, Sargent CY, McDevitt TC. Rotary suspen-sion culture enhances the efficiency, yield, and homogene-ity of embryoid body differentiation. Stem Cells 2007;25:2224–2234.
28. Mahoney MJ, Saltzman WM. Transplantation of brain cellsassembled around a programmable synthetic microenviron-ment. Nat Biotechnol 2001;19:934–939.
29. Solorio LD, Fu AS, Hernandez-Irizarry R, Alsberg E. Chon-drogenic differentiation of human mesenchymal stem cellaggregates via controlled release of TGF-beta1 from incorpo-rated polymer microspheres. J Biomed Mater Res A 2010;92:1139–1144.
30. Desai MP, Labhasetwar V, Amidon GL, Levy RJ. Gastrointes-tinal uptake of biodegradable microparticles: Effect of particlesize. Pharm Res 1996;13:1838–1845.
31. Desai MP, Labhasetwar V, Walter E, Levy RJ, Amidon GL.The mechanism of uptake of biodegradable microparticles inCaco-2 cells is size dependent. Pharm Res 1997;14:1568–1573.
32. Goncalves MB, Boyle J, Webber DJ, Hall S, Minger SL,Corcoran JP. Timing of the retinoid-signalling pathway deter-mines the expression of neuronal markers in neural progeni-tor cells. Dev Biol 2005;278:60–70.
33. Okada Y, Shimazaki T, Sobue G, Okano H. Retinoic-acid-concentration-dependent acquisition of neural cell identityduring in vitro differentiation of mouse embryonic stem cells.Dev Biol 2004;275:124–142.
34. Pachernik J, Bryja V, Esner M, Kubala L, Dvorak P, HamplA. Neural differentiation of pluripotent mouse embryonalcarcinoma cells by retinoic acid: Inhibitory effect of serum.Physiol Res 2005;54:115–122.
35. Berkland C, King M, Cox A, Kim K, Pack DW. Precise con-trol of PLG microsphere size provides enhanced control ofdrug release rate. J Control Release 2002;82:137–147.
36. Varde NK, Pack DW. Influence of particle size and antacidon release and stability of plasmid DNA from uniform PLGAmicrospheres. J Control Release 2007;124:172–180.
MICROSPHERE SIZE EFFECTS ON EMBRYOID BODIES 475
Journal of Biomedical Materials Research Part A





























