Embed Size (px)
Citation preview

Sky Journal of Medicinal Plant Research Vol. 2(3), pp.18 - 28, September, 2013 Available online http://www.skyjournals.org/SJMPR ISSN 2315-8786 ©2013 Sky Journals
Full Length Research
Micropropagation of Gymnema sylvestre R.Br.
Syed Naseer Shah1, Amjad M. Husaini2*, S. A. Ansari1 1Genetics and Plant Propagation Division, Tropical Forest Research Institute, Mandla Road, Jabalpur 482 021, India.
2Centre for Plant Biotechnology, Division of Biotechnology, SKUAST-K, Shalimar, Srinagar-191121, India
Accepted 29 August, 2013
Gymnema sylvestre R.Br (Hindi: Gurmar) is a medicinal plant of immense pharmaceutical value, but is fast disappearing from its natural habitat. The species is threatened with extinction due to its indiscriminate collection as raw material for pharmaceutical industry, where it is used for manufacturing drugs for diabetes, asthma, eye complaints, etc. An attempt has been made for development of in vitro propagation procedure for the species, involving four steps, viz., culture establishment, shoot multiplication, rooting and hardening. Aseptic cultures were established on MS (Murashige and Skoog) medium supplemented with 10.0 µM BA (N
6-
Benzyladenine) and 0.5 µM NAA (α- Naphthalene acetic acid) using nodal segments (1-1.5 cm) as explants from five year old plants of three different locations. Of the five nutrient media [MS (Murashige and Skoog Medium), WPM (Woody Plant Medium), B5 (Gamborg Medium), SH (Schenk and Hildebrandt Medium) and NN (Nitsch and Nitsch Medium)], five cytokinin sources [Ads(Adenine hemisulphate), BA, Kn (Kinetin), 2-iP (N
6-(2-isopentyl)
adenine) and TDZ (Thidiazuron)], six doses (0, 1.25, 2.5, 5.0, 10.0 and 20.0 µM) and their all possible interactions in two successive experiments, NN medium and 5.0 µM BA significantly proved optimum for in vitro shoot multiplication and resulted in 1.82 shoot number per explant 2.7 node number shoot
-1 and 5.2 node number per
explants at one month after inoculation. The in vitro multiplied shoots were tested for in vitro root induction on
different culture media (MS, WPM, B5, SH, NN), explant types (apical bud with two nodal segments or with three nodal segments, two nodal segments, one nodal segment) and auxin (IAA, IBA, NAA)/ non-auxin (coumarin) treatment (0, 5.0, 10.0 and 15.0 µM) in two separate experiments laid out in factorial randomized designs. Explants comprising apical bud with two nodal segments or three nodal segments without apical bud, inoculated on B5 media supplemented with 5.0µM NAA, screened out to be significantly excellent for induction and growth of adventitious roots, resulting in 85% rooting and 4.27 root number explant
-1 at 35 days after
inoculation. The in vitro propagated plants exhibited excellent growth. Therefore, the present study recommends a four step micropropagation procedure for in vitro production of G. sylvestre plants on a commercial scale to meet the requirement of pharmaceutical industries and save the species from extinction. Key words: Gymnema sylvestre R.Br, cytokinin, auxin.
INTRODUCTION Gymnema silvestre R.Br., is a member of Asclepiadaceae, is a vulnerable woody and highly branched, slow growing, perennial vine run over the top of tall trees found in tropical and subtropical regions (Anonymous, 1997). It is a potent anti-diabetic plant used in folk, Ayurvedic, sidda and homeopathic systems of medicine. It is also used in the treatment of asthma, eye *Corresponding author. E-mail: [email protected].
complaint, inflammations, prevent dental caries caused by Streptococcus mutans and skin cosmetics (Komalavalli and Rao, 2000). Multiple uses are attributed by the presence of the active principle, gymnemic acid. This species is fast disappearing and is threatened with extinction due to its indiscriminate collection and over exploitation from natural resources for commercial purposes and to meet the requirement of the pharmaceutical industry (Choudhury, 1988). The conventional propagation is hampered due to its poor seed viability, low rate of germination and poor rooting

Shah et al. 19
Figure 1. Explant collection, culture establishment and shoot multiplication in Gymnema sylvestre R. Br.; (a) mother plant, (b) a twig, (c) nodal explants, (d) the in vitro culture establishment and (e-f) the in vitro shoot multiplication.
ability of vegetative cuttings (Komalavalli and Rao 2000). Alternative in vitro propagation method is beneficial in accelerating large scale multiplication, improvement and conservation of this plant. MATERIALS AND METHODS
Plant material The explants were collected as twigs (Figure 1b) from the parental stock at Jabalpur area of MP (Figure 1a), brought alive to the laboratory and washed thoroughly for 15 min under running water for removing the debris from the surface. The washed twigs were defoliated and cut into nodal explants (approximately 1 - 1.5 cm long and 0.5 - 0.6 cm diameter) (Figure 1c). These explants were washed with 2% Cetrimide
® and kept for 10 min with
constant vigorous shaking (150 rpm) on an orbital shaker incubator to facilitate better surface contact with explants. The explants were rewashed 4 - 5 times with distilled water to remove traces of Cetrimide
®. The washed
explants were sterilized for 5 min with a composite sterilization treatment comprising HgCl2 (0.1%), Phytolon
®
(1.0%) and Streptomycin® (0.2%) in the laminar flow
cabinet. Finally, the surface sterilized nodal explants were rinsed 4 - 5 times with sterile distilled water for removal of sterilizing agent under laminar flow cabinet. The nodal segments were inoculated on MS medium (Murashige and Skoog, 1962) supplemented with 10.0
µM BA and 0.5 NAA µM for culture establishment (Figure 1d). Shoot multiplication Two sets of factorial randomized design experiments were conducted to investigate the effect of five basal media and six cytokinin sources and their all possible interactions on shoot number per explant, node number per shoot and node number per explant at 15 and 30 days of inoculation. Single nodal segments from established cultures were used for shoot multiplication experiments (Figure 1a-e). In the first experiment, five nutrient media [MS (Murashige and Skoog, 1962), WPM (Lloyd and McCown, 1980), B5 (Gamborg et al., 1968), SH (Schenk and Hildebrandt, 1972) and NN (Nitsch and Nitsch, 1969)] along with four doses of BA (0, 5.0, 10.0, and 20.0 µM) were tested either alone or in combination. In the second experiment, different cytokinins [Ads (adenine hemisulphate), BA (N
6-Benzyladenine), Kn
(Kinetin), 2-iP (N6-(2-isopentyl) adenine) and TDZ
(Thidiazuron)] in three doses (1.25, 2.5 and 5.00 µM) were screened out for their effect on shoot multiplication. In vitro rooting The in vitro multiplied shoots were tested for root induction in two separate experiments comprising: (1) different culture media (MS, B5, WPM, SH, and NN), explant types (Figure 2), [apical bud with two nodal segment (E1) or with three nodal segment (E2), two nodal segment (E3), one nodal segment (E4) and their all possible interactions at 10.0 µM IBA uniformly and (2)

20 Sky J. Med. Plants Res.
Figure 2. Different types of explant used for adventitious rooting in Gymnema sylvestre:
E1, Apical bud with two nodal segments, E2. Apical bud with three nodal segments, E3, Two nodal segments and E4, One nodal segment.
Figure 3. The in vitro adventitious root induction in Gymnema sylvestre R. Br.; root formation on liquid (a) and
semi-solid (b-c) medium and root system 30 days after inoculation.
four auxin (IAA, IBA ,NAA)/ non-auxin (coumarin) sources, four doses ( 0 , 5.0 , 10.0 and 15.0 µM) and their all possible interactions laid out in factorial randomized designs. Rooting (%) and root number per explant were recorded at 21, 28, 35 and 42 days in Experiment 1 and 21, 28 and 35 days in Experiment 2. Culture conditions and statistical analysis The inorganic salts used for preparation of culture medium were obtained from Qualigens Pvt. Ltd., India and phytohormones and B vitamins from Sigma Chemicals Pvt. Ltd., India. The medium contained 3% (w/v) sucrose, 0.8 % (w/v) agar (Hi-media chemical Ltd., India). The pH of the medium was adjusted to 6.0 before autoclaving for 15 min at 1.06 kg cm
-2 (121°C). Explants
were cultured in a 150 ml conical Borosil® flasks
containing 40 ml semi-solid medium. For in vitro shoot multiplication and rooting experiment, the cultures were
incubated at 25 ± 2°C under 16 h illuminations with fluorescent light (50 µmol Em
-2 s
-1). The experiments had
five replicates for in vitro shoot multiplication and three replicates for in vitro rooting (Figure 3). Each replicate had 10 propagules. The in vitro shoot multiplication data were recorded at 15 and 30 days and the in vitro rooting data at 21, 28, 35 and 42 days after inoculation. The data were subjected to two way (factor) analysis of variance for all the experiments with “F” test for ascertaining level of significance. If the data were found significant at p ≤ 0.05, LSD0.05 was computed for comparison of treatment means. Hardening and transplantation The in vitro raised plantlets were removed from rooting medium, washed with distilled water and transferred to fresh tubes containing MS inorganic for 5 - 7 days. The plantlets were subsequently transferred to root trainers

Shah et al. 21
Figure 4. Hardening and acclimatization of the in vitro raised plantlets of Gymnema
sylvestre R. Br.; plantlets transferred to root trainers (a) and covered with polythene (b) placed in the culture room, (c) hardened plantlets transferred to polythene bags and (d) growth of the plantlets in the open environment.
containing autoclaved soilrite (Figure 4a) and covered with perforated polythene to maintain humidity, which were kept under culture room conditions for about 15 days. Subsequently, they were transferred to perforated polythene bags (Figure 4b) and kept initially in culture room for 5 days and finally transferred to natural condition (Figure 4c-d). RESULTS In vitro shoot multiplication Experiment 1 Shoot Number per explant: The shoot number was significantly affected by the basal medium at 15 days and by the medium supplemented with BA on all possible interactions at 30 days after inoculation (Figure 5a). Shoot number per explant was significantly maximum and on par, in MS, B5, SH and NN media at 15 days and B5 and NN media at 30 days after inoculation. On the other hand, the cultures maintained on WPM performed significantly poor for shoot number per explant
at both
stages of sampling. NN medium significantly enhanced shoot number per explant by 177% at 15 days and 236% at 30 days in comparison to WPM at respective stages of sampling. BA doses ranging from 5.0 to 20.0 µM were on par, but differed significantly with the control for shoot number per explant at 30 days after inoculation. Dose at 5.0 µM significantly enhanced shoot number per explant by 33% in comparison to the control. Interaction between basal medium and BA dose was significant for shoot number per explant only at 30 days. Interaction NN*BA at 5.0 µM produced significantly maximum shoot number per explant but was on par with interactions B5*BA at 10.0 – 20.0 µM, and NN*BA at 10.0 – 20.0 µM. The interaction WPM*BA at 5.0 µM produced significantly minimum shoot number per explant but was statistically equalled by interaction WPM*BA at 10.0 µM. Shoot number per explant obtained in interaction NN*BA at 5.0 µM was 100% higher than that obtained in interaction WPM*BA at 5.0 µM. Node number per shoot: Node number per shoot
was
significantly influenced only by basal medium at 15 and 30 days after inoculation (Figure 5b). NN medium produced significantly maximum node number per shoot,
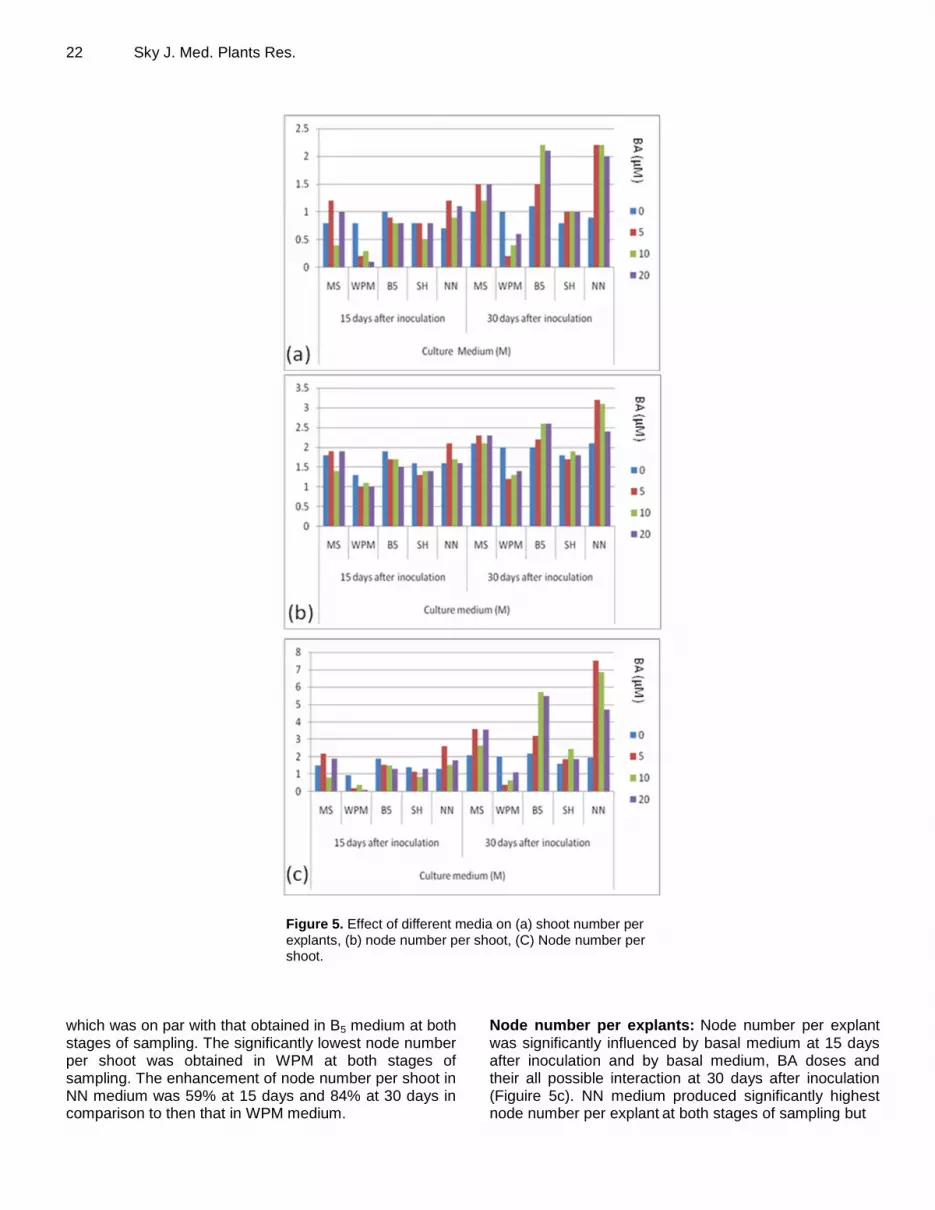
22 Sky J. Med. Plants Res.
Figure 5. Effect of different media on (a) shoot number per
explants, (b) node number per shoot, (C) Node number per shoot.
which was on par with that obtained in B5 medium at both stages of sampling. The significantly lowest node number per shoot was obtained in WPM at both stages of sampling. The enhancement of node number per shoot in NN medium was 59% at 15 days and 84% at 30 days in comparison to then that in WPM medium.
Node number per explants: Node number per explant
was significantly influenced by basal medium at 15 days after inoculation and by basal medium, BA doses and their all possible interaction at 30 days after inoculation (Figuire 5c). NN medium produced significantly highest node number per explant
at both stages of sampling but

was statistically equaled by B5 medium at 15 days after inoculation. On the other hand, WPM medium resulted in significantly minimum node number per explant at both stages of sampling. NN medium had 341% and 304% more node number per explant than WPM medium. BA at 5.0 – 20.0 µM produced statistically similar value for node number per explant
but was significantly more than the
control. BA at 5.0 µM enhanced node number per explant by 69% in comparison to the control. Interaction between basal medium and BA dose was found significant only at 30 days after inoculation. Interaction of NN medium and BA at 5.0 µM resulted in maximum value for node number per explant which was, however, statistically equaled by interaction NN*BA at 10.0 µM and B5*BA at 5.0 – 10.0 µM. Interaction WPM*BA 5.0 µM recorded the lowest value for node number per explant and was statistically equaled by several other interactions, including WPM*BA at 0, 10.0 and 20.0 µM. Interaction NN*BA at 5.0 µM enhanced node number per explant by 1707% in comparison to WPM*BA at 5.0 µM. Experiment 2 Shoot number per explant: Cytokinin sources and their interaction with three doses significantly influenced shoot number per explant
after 30 days of inoculation (Figure
6a). Of the various cytokinin sources, BA proved significantly optimum for shoot number per explant. On the other hand, Ads recorded significantly lowest value for the parameter, which was, however, on par with that obtained in Kn. Shoot number per explant
obtained in BA
was 96% higher than that in Ads. However shoot number per explant
obtained in all three doses did not differ
significantly from each other. Interaction BA at 2.5 - 5.0 µM produced significantly highest shoot number per explant, whereas the remaining interactions, except BA and 2-iP at 1.25
µM produced the lowest values for the
parameter. An enhancement of 130% for shoot number per explant
was recorded in 2-iP at 5.0 µM in comparison
to Ads at 5.0 µM. Node number per shoot: The effect of different concentration of cytokinin were found to be non significant on node number per shoot at 30 days after inoculation. On the other hand, different cytokinin sources and their interactions with three doses were found to be significant on node number per shoot at 30 days after inoculation (Figure 6b). Of various cytokinin sources, BA produced significantly maximum node number per shoot which was on par with 2-iP.
Ads, however, resulted in
minimum node number per shoot but was statistically
equal to TDZ. Node number per shoot produced by BA was 61% higher than that by Ads. The interactions between different doses and cytokinin sources were found to be significant on node number per shoot at 30 days after inoculation. BA at 1.25 – 5.0 µM produced significantly maximum
Shah et al. 23 node number per shoot, which was statistically equaled by Kn at 2.5 – 5.0 µM and 2-iP at 1.25 and 5.0 µM. The remaining interactions recorded the significantly minimum values for the parameter. BA at 1.25 µM enhanced node number per shoot by 70 % in comparison to Ads at 1.25 µM. Node Number per explants: The effect of different doses and their interactions with cytokinin sources were found to be non significant on node number per explant
at
30 days after inoculation. However, different cytokinin sources significantly influenced node number per explant at 30 days after inoculation (Figure 6c). BA produced maximum node number per explant, which statistically equaled by that obtained in 2-iP and TDZ. Ads resulted in lowest node number per explant but
was on par with all
cytokinin sources, except BA. Node number per explant
was 283% more in BA than Ads. In Vitro Adventitious Rooting Experiment 1 Adventitious root (%): Explant type were significantly influenced adventitious root (%) only at 42 days of inoculation (Figure 7). Explant type E2, that is, apical bud with three nodal segment enhanced significantly highest adventitious root (%). Explant type E4 resulted in the lowest induction of adventitious root (%). This was, however, on par with that obtained in Explant type E1 and E3. Explant type E2 enhanced adventitious root (%) by 1031 % in comparison to E4. Basal medium significantly affected adventitious root (%) at all stages of sampling. Basal medium B5 invariably proved significantly optimum for adventitious root (%) at all stages of sampling (Figure 4). In contrast, basal medium SH did not induce adventitious root (%) at all stages of sampling (Figure 2). Also basal medium MS did not induce adventitious root (%) at 21 days of sampling. Interaction between explant type and basal medium was significant on adventitious rooting (%) only at 35 and 42 days of inoculation. Explant type E1 and E2 with basal medium B5
induced significantly highest adventitious root (%) at two stages of sampling. On the other hand, explant type E2, E3 and E4 with basal medium MS and SH resulted in significant lowest induction of adventitious root (%). Root number per explants: Root number per explant was significantly influenced by explant type at 42 days of inoculation and basal medium at all stages of sampling and interaction between explant type and basal medium at 42 days of inoculation (Figure 7). Explant type E2
produced maximum root number per explant and was statistically equaled by Explant type E1 at 42 days of inoculation. Explant type E4 had lowest root number per explant and was on par with Explant type E1 and E4 at 42

24 Sky J. Med. Plants Res.
Figure 6. Effect of fives cytokinins sources, three doses each in
(a) shoot number per explant (b) node number per shoot (c) node number per explant.
days of inoculation. Explant type E2 enhanced root number explant
-1 by 657% in comparison to Explant type
E4 at 42 days of inoculation. Among different basal medium, B5 medium resulted in the highest root number
per explant at all stages of sampling and was equaled by WPM at initial stage of sampling, that is, 21 days of inoculation. On the other hand, MS medium induced the lowest root number per explant and was significantly

Shah et al. 25
Figure 7. Effect of basal media and explant type on (a) root (%) at 21 and 28 days after inoculation
(b) root (%) at 35 and 42 days after inoculation (c) root number per explant at 21 and 28 days after inocmulation (d) root number per explant at 35 and 42 days after inoculation.
equal to WPM, SH and NN medium. B5 medium significantly increased root number per explant by 800, 267 and 700% in comparison to MS medium at 28, 35 and 42 days of inoculation, respectively (Figure 7). Interaction between explant type E2 and basal medium B5 produced significantly highest root number per explant at 42 days of inoculation. The remaining interactions, except explant type E1 and basal medium B5 were on par with each other and produced significantly lowest shoot number per explant at 42 days of inoculation. Experiment 2 Rooting (%) IAA, IBA, NAA and coumarin significantly influenced rooting (%) at all stages of sampling (Figure 8). Administration of NAA significantly proved optimum at all stages of sampling. The lowest rooting (%) was recorded in IBA but was on par with that obtained in IAA and coumarin at all stages of sampling. Treatment with NAA enhanced rooting by 108%, 218% and 235% over IAA,
IBA and coumarin at 21 days, 890% and 1441% over IAA and coumarin at 28 days and 785, 2082 and 1009% over IAA, IBA and coumarin at 35 days of inoculation, respectively. Among various doses, administration of 5.0 µM proved maximum at first two stages (21 and 28 days) of sampling and that of 15.0 µM at the last stage (35 days) of sampling. However, the value obtained in 5.0 µM dose was on par with that of 10.0 µM at 28 and 35 days and 15.0 µM at 35 days of sampling. Administration of 5.0 µM dose enhanced rooting (%) by 618% and 460 % over untreated control at 28 and 35 days after sampling, respectively. As far interaction effect, 5.0 µM NAA proved significantly optimum for rooting (%), at all stages of sampling. On the other hand, 5.0 µM IAA, IBA and coumarin at all three stages of sampling did not induce rooting. However 5.0 µM dose of IAA, IBA and coumarin induced low level of rooting (%) at different sampling stages. Root number per explant: IAA, IBA, NAA and coumarin significantly influenced root number per explant at all stages of sampling (Figure 8). Of these, NAA invariably proved optimum at all stages of sampling. On the other

26 Sky J. Med. Plants Res.
Figure 8. Effect of different auxins and coumarin on (a) root (%) at 21 and 28 days after inoculation (b) root
(%) at 35 days after inoculation (c) root number per explants at 21 and 28 days after inoculation (d) root number per explants at 35 days after inoculation.
hand, IAA, IBA and coumarin resulted in significantly lowest root number per explant. NAA significantly enhanced root number per explant
by 518, 6700 and
871% at 21 days, 523 ,800 and 710% at 28 days and 8099, 1567 and 809% at 35 days in comparison to IAA, IBA and coumarin, respectively. Among various doses, 5.0 µM doses significantly enhanced root number per explant at 21 and 35 days of sampling, whereas 5.0 – 15 µM doses being on par produced maximum root number per explant
at 28 days of sampling. However, the control
produced significantly lowest root number per explant at all three stages of sampling. Administration of 5.0 µM dose had 625 and 510% more root number per explant than the control at 28 and 35 days of sampling, respectively. Interaction between root substances and their different doses was significant for root number per explant at all three stages of sampling. 5.0 µM NAA recorded significantly highest root number per explant at all three stages of sampling, which was 1250 and 1024% more than that of 5.0 µM coumarin at 21 and 35 days of sampling, respectively. However, 5.0 µM IAA and IBA did not produce root number per explant all three stages of
sampling and 5.0 µM coumarin at 28 days of sampling. Administration of 10.0 - 15.0 µM each of IAA, IBA, NAA and coumarin resulted in low root number per explant at all three stages of sampling, except 15.0 µM IBA and 10.0 µM coumarin at 21 and 28 days of sampling, which did not produce root number per explant. Discussion The micro-propagation of Gymnema sylvestre comprises four steps, viz., establishment of culture from nodal explants, shoot multiplication, root induction and hardening and acclimatization. Selection of medium is vital to success in tissue culture and the choice of the medium is dictated by the purpose of the tissue culture technology, which is to be employed for plant species or variety (Gamborg and Phillips, 1995; Smith, 2000). There are many reports comparing different media for their effect on in vitro shoot multiplication in different plants (Rugini, 1984; Mehta et al., 2000; Lu, 2005; Jain et al., 2009). In the present investigation on Gymnema sylvstre,

a number of culture media including MS, B5, WPM, SH and NN were employed. For shoot multiplication in Gymnema sylvestre, NN medium proved most effective. There are no earlier published reports on comparison of different culture media in Gymnema sylvestre and most of the workers have used MS medium for shoot multiplication (Reddy et al., 1998; Komalavalli and Rao, 2000; Devi and Srinvasan, 2008). The concentration of macro-nutrients in NN medium is low as compared to the other media tried. Among micro-nutrients, iodine and cobalt are absent in NN medium. Similarly, the amount of vitamins like nicotinic acid (vitamin B3) and thiamine (vitamin B1) is high as compared to other media. Also, NN medium incorporates biotin (vitamin B2) and folic acid (vitamin B4), which are totally absent in the other four media tried. NN medium has been found effective for shoot multiplication in Coluria geoids (Olszowska and Furmanowa, 1986), Corylus avellana (Nas and Read, 2001), Pelargonium graveolens (Temba and Deodhar, 2010). However, Arikat et al. (2004) could not observe any significant difference in shoot multiplication of Salvia fructicosa on MS, NN and B5 medium.
The nodal explant of Gymnema sylvestre differentiated into multiple shoots on cytokinin supplemented medium, and relatively low dose of BA (5µM) was found to be best for shoot multiplication. BA was found to be more effective than other cytokinins (Ads Kn, 2-iP, and TDZ) for in vitro shoot multiplication. BA has been used for shoot multiplication in many other species of Ascelpidieaceae family like Ceropegia jainii, C. bulbosa var. Bulbosa and C. bulbosa var. Lushii, (Vishwanath, 1998), Cryptolepis buchananii (Prasad et al., 2004), Tylophora asthmatica (Mukundan et al., 2002). There are earlier reports on micropropagation of other medicinal plants where BA has consistently been used for shoot multiplication. BA was found to be optimum for shoot multiplication in Curcuma aromatic (Nayak, 2000), Azadirachta indica (Quraishi et al., 2004) and Carissa carandas (Rai and Misra 2005). BA has also been effective in many woody species too, like Maytenus emarginata (Rathore et al., 1992), Prosopis cineraria (Shekhawat et al., 1993), Wrightia tomentosa (Purohit et al., 1994) Capparis deciduas (Deora and Shekhawat, 1995), Sapium sebiferum (Sirail and Dhar, 1996) and Acacia catechu (Kaur et al., 1998). The superiority of BA over other sources of cytokinins has been reported earlier and attributed to the group localized at N6 position of the cytokinins (Murashige, 1974; Street, 1997). The precise reason for greater efficacy of BA is not known. However, BA, because of its artificial nature, does not get readily metabolized (degraded) in the plant tissue (Schwarz, 1987).
For in vitro rooting, nutrient salts in the medium have a dramatic effect both on rooting (%) and root number per explants. In the present investigation, out of the five culture media tested, B5 medium proved to be the best for root induction. B5 medium is a low salt strength medium
Shah et al. 27 with only one source of nitrate as KNO3 and contains high concentration of thiamine (vitamin B1). Rooting improves in many woody and herbaceous species when the concentration of macro-salts was lowered to half or less (Webb and Street, 1977). Our results are in agreement with earlier workers, who have used B5 medium for in vitro rooting. For instance, Burnouf-Radosevich and Paupardin (1985) reported in vitro rooting in Chenopodium quinoai on B5 medium. Sharifi and Ebrahimzadeh (2010) compared MS and B5 medium and found that root formation was maximum on B5 medium in Crocus sativus. In Rhinacanthus nasutus also, root formation was accomplished on B5 medium (Panichayupakarananat and Meerungrueang, 2010).
Four types of explants were used for in vitro root induction in the present study. Apical bud with three nodal segments (E2 explants) resulted in maximum rooting (%) and root number per explants. The auxins stimulate root development by inducing root initials that differentiate cells of the young secondary phloem, cambium and pith tissue (Gianfagna, 1995). High concentration of thiamine (vitamin B1) included in B5 medium seems to have synergistically interacted with auxins for facilitation of rhizogenesis as also reported in teak by Ansari et al. (2002). In the present study NAA was found as the best auxin source for in vitro rooting. Superiority of NAA for in vitro rooting may be attributed to its synthetic nature and stability as it eludes the auxin oxidizing/ degrading enzyme systems of the plants (Jacobs, 1972). Lal et al. (1988) examined the effectiveness of various auxins on rooting of Picrorhiza kurroa and found that NAA was superior to IBA or IAA. Rooting of cultured shoots of Saussurea lappa was achieved on NAA (Arora and Bhojwani, 1989). Faria and Illg (1995) obtained rooting in shoots of Zingiber spectabile on NAA supplemented medium. Root induction was reported on NAA in many other medicinal plant species too, viz., Zingiber officinale (Babu et al., 1992), Tylophora indica (Bera and Roy, 1993), Rauvolfia micrantha (Sudha and Seeni, 1996), Decalepis arayalpathra (Sudha et al., 2005) and Dioscorea hispida (Behera et al., 2009).
The procedure offers a potential system for improvement, conservation and mass propagation of Gymnema sylvestre from mature plant derived explants. Nitsch and Nitsch (1969) medium supplemented with 5.0 µM BA has been selected for efficient and rapid multiplication of in vitro shoot multiplication and B5
medium supplemented with 5.0 µM NAA for optimum induction of in vitro adventitious roots at 35 days in explants comprising apical bud with three nodes. Further, the hardening procedure reported here ensures 60 - 70% field survival of micropropagated plants of Gymnema sylvestre.
REFERENCES Anonymous (1997). Medicinal Plant Categorized new IUCN Red List

28 Sky J. Med. Plants Res. Criteria Under the Biodiversity Conservation” Priorisation Project. pp.
10 In: CBSG, India News. Ansari SA, Sharma S, Pant NC, Mandal A K (2002). Synergism
between IBA and Thiamines for induction and Growth of Adventitious Roots in Tectona grandis. J. of Sust. Forestr. 15:99-112.
Arikat NA, Jawad FM, Karam NS, Shibil RA (2004). Micropropagation and accumulation of essential oils in wild sage (Salvia Fructicosa Mill.). Sci. Hort., 100: 193-202.
Arora R, Bhojwani SS (1989). In vitro propagation and low temperature storage of Saussurea lappa C.B. Clarke an endangered medicinal plant. Plant Cell Rep,. 8:44-47.
Babu KN, Samsudeen K, Ratnabal MJ (1992). In vitro plant regeneration from leaf derived callus in ginger (Zingiber oficinale Ross). Plant Cell Tiss.Org. Cult., 29: 71-74.
Behera KK, Sahoo S, Prusti A (2009). Effect of plant growth on in vitro micropropagation of „Bitter Yam‟ (Dioscorea hispida Dennst.). Int. J. integrative Biol., 4: 50-54.
Bera TK, Roy SC (1993). Micropropagation of Tylophora indica (Burm.F.) Merr. By multiple bud formation from mature leaf explant without callus intervention. Bot. Bull. Academia Sinica 34: 83-87.
Choudhury BP (1988). Assessment and conservation of medicinal plants of Bhubaneswar and its neighborhood”. In: Indigenous Medicinal Plants, Today & Tomorrow Printers & Publishers, New Delhi, India. pp. 211-219
Deora NS, Shekhawat NS (1995). Micropropagation of Capparis deciduas (Forsk.) Edgew. : A tree of arid horticulture. Plant Cell Reports, 15: 278-281.
Devi CS, Srinivasan VM (2008). In vitro propagation of Gymnema sylvestre. Asian J. Plant Sciences, 7: 660-665.
Faria RT, IIIg RD (1995). Micropropagation of Zingiber spectabile Griff. Sci. Hort., 62: 135-137.
Gamborg OL, Miller RA, Ojima K (1968). Nutrient requirements of suspension cultures of soya bean root cells. Exp. Cell Res., 50: 151-158.
Gamborg OL, Phillips GC (1995). Media preparation and handling, in Gamborg, O.L. and Phillips, G.C. (eds.) Plant Cell, Tissue and Organ Culture-Fundamental methods.: Springer – Verlag, Berlin pp. 21-34.
Gianfagna T (1995). Natural and synthetic growth regulators and their use in horticultural and agronomic crops. In: Plant Hormones: Physiology, Biochemistry and Molecular Biology (ed Davies P.J.) Kluwer Academic Publishers, Dordrecht. pp.751-773.
Jacobs WP (1972). The movement of plant hormones: auxins, gibberellins and cytokinins”. In D.J. Carr (ed.), Plant Growth Substances.. Springer, New York. pp. 701-709
Jain N, Bairu MW, Stirk WA, Van Staden J (2009). The effect of medium, carbon source and explant on regeneration and control of shoot-tips necrosis in Harpagophytum procumbens. South Afr. J. Bot., 75: 117-121.
Kaur K, Verma B, Kant U (1998). Plants obtained from the Khair tree (Acacia catechu Willd.) using mature nodal segments. Plant cell Reports, 17: 427-429.
Komalavalli N, Rao MV (2000). In vitro propagation of Gymnema sylvestre-A multipurpose medicinal plant. Plant Cell Tissue. Organ Culture, 61: 97-105.
Lal NP, Ahuja S, Kukreja AK, Pandey B (1988). Clonal propagation of Picrorhiza Kurroa Royale ex. Benth. By shoot tip culture. Plant Cell Reports, 7: 202-205.
Lloyd G, McCown B (1980). Commercially feasible micropropagation of mountain laurel Kalmia labtifolia by use of shoot tip culture. Comb. Proc. Int. Plant prop.Soc., 30:421-427.
Lu MC (2005). Micropropagation of Vitis thunbergii Sieb. Et Zucc., a medicinal herb, through high-frequency shoot tip culture. Sci., Hort., 84:64-69.
Mehta UJ, Krishnamurthy KV, Hazra S (2000). Regeneration of plants via adventitious dud formation from mature zygotic embryo axis of tamarind (Tamarindus indica L.). Curr. Sci., 78: 1231-1234.
Mukundan U, Sivaram L, Kumar A (2002). Micropropagation of Tylophora asthamatica and Uraria picta. Plant cell Biotechnol. and Mol. Biol., 3:73-76.
Murashige T (1997). Plant propagation through tissue cultures. Annual Rev. Plant Physiol., 25: 135-166.
Murashige T, Skoog F (1962). A revised medium for rapid growth and
bioassays with tobacco tissue cultures. Physiol. Plant. 15: 473-497. Nas MN, Read PE (2001). Micropropagation of hybrid hazelnut: medium
composition, physical state and iron source affect shoot morphogenesis, multiplication and explant vitality.V. International congress on Hazelnut. S.A. Mehlenbacher ISBN 978-90-66058-74-3 Volumes Corvallis, Oregon, United States of America.
Nayak S (2000). In vitro multiplication and microrhizome induction in Curcuma aromatic Salisb. Plant Growth Regulation, 32:41-47.
Nitsch JP, Nitsch C (1969). Haploid plants from pollen grains. Science 163: 85-87.
Olszowska O, Furmanowa M (1986). Micropropagation of Coluria geoides by axillary shoots. Planta Medica, 52: 521.
Panichayupakarananat P, Meerungrueang W (2010). Effect of medium composition and light on root and Rhinacanthus formation in Rhinacanthus nasutus cultures. J. Pharm., 48:1192-1197.
Prasad PJN, Chakradhar T, Pullaiah T (2004). Micropropagation of Cryptolepis buchanani Roem. & Schult. Int. J of Life Sci., 49: 57-65.
Purohit SD, Kudka G, Sharma P, Tak K (1994). In vitro propagation of an adult tree Wrightia tomentosa through enhanced axillary branching. Plant Sci., 103:67-72.
Quraishi A, Koche P, Sharma P, Mishra SK (2004). In vitro Clonal propagation of Neem (Azadirachta indica), Plant Cell, Tissue and Organ Culture,78: 281-284.
Radosevich MB, Paupardin C (1985). Vegetative propagation of Chenopodium quino by shoot tip culture. Am. J. Bot., 72: 278-283.
Rai R, Mishra KK (2005) Micropropagation of Karonda (Carissa carandas) through shoot multiplication. Sci. Hort., 103: 227-232.
Rathore TS, Deora NS, Shekhawat NS (1992). Cloning of Maytenus marginata (Willd.) Ding Hou-a tree of the Indian Desert, through tissue culture. Plant Cell Reports, 11: 449-451.
Reddy PS, Gopal GR, Sita GL (1998). In vitro multiplication of Gymnema sylvestre R.Br.-An important medicinal plant. Curr. Sci., 75:843-845.
Rugini E (1984). In vitro propagation of some olive (Olea europaea sativa L.) cultivars with different root-ability, and medium development using analytical data from developing shoots and embryos. Sci. Hotricul., 24:123-134.
Schenk RU, Hildebrandt AC (1972). Medium and technique for induction and growth of monocotyledonous and dicotyledonous plant cell culture”. Can. J. Bot., 50: 199-204.
Schwarz OJ (1987). Plant growth regulator effect in the in vitro propagation of three hardwood tree genera: Castanea, Juglans and Quercus. Plant growth Regulation, 16: 113-135.
Sharifi G, Ebrahimzadeh H (2010). Interaction of IBA and NNA with enzymes in root induction of Crocus sativus (L). Afr. J. Biotechnol., 9:217-225.
Shekhawat NS, Rathore TS, Singh RP, Deora NS, Rao SR (1993). Factors affecting in vitro cloning of Prosopis Cineraria. Plant Growth Regulation, 12: 273-280.
Sirail EA, Dhar U (1996). A highly efficient in vitro regeneration methodology for mature Chinese tallow tree (Sapium sebiferum Roxb.). Plant Cell Reports, 16: 83-87.
Smith RH (2000). Plant tissue culture techniques and experiments. Acad. Press, USA, pp. 43-54.
Street HE (1997). Plant tissue and cell culture, University of Calfornia Press, Berkely, pp. 1-614.
Sudha CG, Krishnan PN, Pushpangadan P, Seeni S (2005). In vitro multiplication of Decalepis arayalpathra, a critically endangered ethno
medical plant. In vitro Cellular and developmental Biology. Plant 41: 648-654.
Sudha CG, Seeni S (1996). In vitro propagation of Rauwolfia micrantha, a rare medicinal plant. Plant Cell Tissue Organ Culture, 44: 243-248
Temba PR, Deodhar AM (2010). Clonal propagation of different cultivars of Pelargonium graveolens (L‟ Herit.). Boitechnol., 9: 492-498.
Vishwanath MP (1998). Micropropagation studies in Ceropegia ssp. In vitro Cellular and Developmental Biology- Plant, 34: 240-243.
Webb KJ, Street HE (1977). Morphogenesis in vitro of Pinus and Picea. Acta Hort., 78: 259-269.






![Micropropagation of Dev-ringal [Thamnocalamus spathiflorus (Trin.) Munro] — a temperate bamboo, and comparison between in vitro propagated plants and seedlings](https://img.dokumen.tips/doc/110x75/6337acbed102fae1b6077c4d/micropropagation-of-dev-ringal-thamnocalamus-spathiflorus-trin-munro-a.jpg)






















