Embed Size (px)
Citation preview

Review ArticleMethyl Jasmonate Putative Mechanisms of Action onCancer Cells Cycle Metabolism and Apoptosis
Italo Mario Cesari Erika Carvalho Mariana Figueiredo RodriguesBruna dos Santos Mendonccedila Nivea Dias Amocircedo and Franklin David Rumjanek
Laboratorio de Bioquımica e Biologia Molecular do Cancer Instituto de Bioquımica MedicaUniversidade Federal do Rio de Janeiro Avenida Carlos Chagas Filho 373 Predio CCS Bloco E Sala 22 Ilha do FundaoCidade Universitaria 21941-902 Rio de Janeiro RJ Brazil
Correspondence should be addressed to Italo Mario Cesari italobo2hotmailcom
Received 12 May 2013 Revised 6 November 2013 Accepted 7 November 2013 Published 6 February 2014
Academic Editor Afshin Samali
Copyright copy 2014 Italo Mario Cesari et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Methyl jasmonate (MJ) an oxylipid that induces defense-related mechanisms in plants has been shown to be active against cancercells both in vitro and in vivo without affecting normal cells Here we review most of the described MJ activities in an attempt toget an integrated view and better understanding of its multifaceted modes of action MJ (1) arrests cell cycle inhibiting cell growthand proliferation (2) causes cell death through the intrinsicextrinsic proapoptotic p53-independent apoptotic and nonapoptotic(necrosis) pathways (3) detaches hexokinase from the voltage-dependent anion channel dissociating glycolytic andmitochondrialfunctions decreasing the mitochondrial membrane potential favoring cytochrome 119888 release and ATP depletion activating pro-apoptotic and inactivating antiapoptotic proteins (4) induces reactive oxygen species mediated responses (5) stimulates MAPK-stress signaling and redifferentiation in leukemia cells (6) inhibits overexpressed proinflammatory enzymes in cancer cells such asaldo-keto reductase 1 and 5-lipoxygenase and (7) inhibits cell migration and shows antiangiogenic and antimetastatic activitiesFinally MJ may act as a chemosensitizer to some chemotherapics helping to overcome drug resistant The complete lack of toxicityto normal cells and the rapidity by which MJ causes damage to cancer cells turn MJ into a promising anticancer agent that can beused alone or in combination with other agents
1 Introduction
Cancer cells do not resemble normal cells in terms ofmorphology and metabolic behavior [1] under this premisemuch effort is invested worldwide in order to develop anti-cancer therapies that can kill cancer cells without harmingnormal cells These therapies attempt to target differentiallyexpressed functional molecules in cancer and normal non-transformed cells For this purpose a myriad of new smallmolecular weight synthetic andor natural inhibitor com-pounds are being tested aiming at achieving selective anti-cancer clinical treatments Small molecular weight chemicalsfromplants (phytochemicals) often accomplishmultitargetedanticancer activities including cell cycle arrest inhibitionof cell growth proliferation and metastasis and promoteapoptosis and cell death [2]
Methyl jasmonate (MJ) our focus in this review isa natural cyclopentanone lipid (Figure 1) belonging to thejasmonates (JAs) family of plant oxylipin stress hormones(oxygenated fatty acids) JAs originate from 120572-linolenic acidreleased from the lipid pool of plant cells by lipases andare subsequently oxygenated by lipoxygenases (LOXs) toformhydroperoxide derivatives (cyclopentanones) [3ndash5]TheJAs family members consist mainly of jasmonic acid (JA)cis-jasmonate (CJ) and MJ [6] They occur ubiquitously inplants and their production is induced by different typesof environmental abiotic (UV radiation osmotic stress andtemperature variation) and biotic (pathogens predators)stresses mediating signaling that triggers defense responsesagainst those stresses [7] When JAs are synthesized inresponse to stress they induced the production of reactiveoxygen species (ROS) and of plant secondary metabolites
Hindawi Publishing CorporationInternational Journal of Cell BiologyVolume 2014 Article ID 572097 25 pageshttpdxdoiorg1011552014572097
2 International Journal of Cell Biology
O
O
OCH3
CH3
Figure 1 Chemical structure of methyl jasmonate (cyclopen-taneaceticacid 3-oxo-2-(2-penten-1-yl)- methyl ester)
such as for instance phenolic compounds which are used aschemical defenses [6ndash12] For instance ultraviolet- (UV-) Bradiation triggers JA accumulation [10 13] whereasMJ is pro-duced under herbivore attack and can act as a volatile signalrecognized by neighboring plants to induce the synthesis ofprotease inhibitors [8] JAs alter plantmitochondrial functionand induce reactive oxygen species (ROS) leading to plant celldeath in a way similar to mammalian apoptosis [12 14]
MJ has been shown to have powerful anticancer activitythrough multiple mechanisms [15ndash24] The purpose of thisreview is to gather most relevant findings concerning MJ-mediated anticancer activities in an attempt to integrate theminto a functional view that could help us to develop newanticancer research strategies and therapies
2 Effects of Jasmonates onMammalian Cancer Cells
In 2002 Fingrut and Flescher [15] discovered that MJstrongly and adversely affected human lymphoblasticleukemia cells by suppressing their proliferation andinducing cell death while nontransformed lymphocytesand normal keratinocytes were unaffected [15] In theseinitial studies they showed that JAs selectively targetedcancer cells in an ex vivo mixed population of leukemic andnormal peripheral blood mononuclear cells (PBMCs) froma patient with chronic lymphocytic leukemia (CLL) JAs alsoincreased the life span of T-cell lymphoma-bearingmice [17]Thereafter JAs including MJ and related synthetic analogswere found to inhibit in vitro cancer cell proliferation andto induce cell death in other human and murine cancer celltypes [16 19ndash24] including human breast [15 25] cervix[26ndash29] colon [30 31] colorectal [32] gastric [33] hepatoma[34 35] lung [19 36 37] lymphoma [15 17 18 38]melanoma[15 30 39 40] myeloid leukemia [41 42] neuroblastoma[43ndash45] prostate [15 46ndash48] and sarcoma [49] cancercells (Table 1) Other results have shown that JAs and theirsynthetic derivates exerted selected cytotoxic effects invivo towards metastatic melanoma [21 39] and inhibitedangiogenesis at high doses (it was the reverse at low doses)in the chorioallantoic membrane (CAM) of chicken embryo[40] (Table 2) In general MJ has been found to be superior
to CJ and JA in terms of cytotoxicity and induction ofapoptosis in human cancer cells [33 38 44] Independentlyif jasmonates are dissolved in an organic solvent or not mostexperiments in vitro with JAs and MJ have been reportedto exert their biological effects at similar low millimolar(mM) concentrations (Tables 1 and 3) excepting few caseswhere MJ and particularly some of its chemical derivativeswere active at micromolar (120583M) concentrations in vivo(Table 2) Differential dispersion andor availability ofsmall hydrophobic MJ lipid droplets after phase separationin culture media or solubility in biological fluids in vivomight explain these differences Nonetheless JAs havebeen found to be nontoxic at doses higher than the usualpharmacological doses employed for other compounds (nM120583M) for instance an iv injection of 236mg MJkg bodyweight in mice (equivalent to asymp50mM circulating concin a 25 g adult mouse) was not toxic [15] The nontoxic ivdose was however found lower by Reischer et al in mice(75mgkg) [39] On the other hand oral acute toxicitystudies performed with MJ on 10 Sherman Wistar ratsreported a LD
50gt 5 gkg showing a normal necropsy
analysis whereas the dermal acute dose was LD50gt 2 gkg
from studies in 10 albino rabbits showing again a normalnecropsy analysis [50] No skin irritation was observed whenMJ was topically applied at 2 gkg to 10 albino rats and at 1MJ in 6 albino New Zealand rabbits similarly there was noskin irritation when a 10 MJ solution was topically appliedon 10 guinea pigs and on 50 volunteers (the latter assessedby the Human Repeat Insult Patch Test) [50] Repeatedapplication of the 10 MJ solution at the same skin sitein guinea pigs and in 50 volunteers did not show dermalsensitization neither photoirritation nor photoallergy wasobserved in the volunteers No mucosal irritation wasobserved after a volume of 01mL of neat MJ was instilledinto the right eye of 6 white New Zealand rabbits (left eyeserved as control) [50] Finally when topically applied oncancerous and precancerous skin lesions of eight patientswith different pathologies MJ showed promising results 38patients with oral lichen planus exhibited positive responsesand 18 patient with leukoplakia had complete recovery (for18 months following the first treatment) while MJ treatmentof one patient with lentigo maligna of the face resulted in drytumor surface with reduction of the metaplastic area duringtreatment but the cancer reappeared three months later MJdid not cause significant local or systemic side effects [51](Table 2)Thenotable absence of toxicity ofMJ to normal cellsand to healthy human and animals at mM doses indicatesthat MJ may be used safely in cancer treatments at thesedoses [15 19 21 22 38 39 50] Furthermore based in toxicitystudies an exemption from the requirement of a tolerance forMJ was recently published by the US Federal EnvironmentalProtection Agency (EPA) [52] In this document MJ wasconsidered as a ubiquitous and naturally occurring planthormone regarded as a safe and natural part of the humandiet through commonly consumed fruits No toxicologicalendpoints were found for MJ through any route of exposureand stated that MJ is virtually nontoxic to humans andother nontarget organisms through all routes of exposureincluding oral As further support for these arguments it was
International Journal of Cell Biology 3
Table 1 Effect in vitro of natural and synthetic jasmonates on normal and cancer cells
Jasmonate (JAs conc range)vehicle Cell lines Effects References
JA MJ 05ndash30mMEtOH
Normal mononuclear cells fromblood (healthy donors) No cytotoxicity
[15 38]
Acute human T-lymphoblasticleukemia Molt-4 andandrogen-responsive humanprostate adenocarcinoma LNCap
JA 3mM 90 cytotoxicityMJ 05mM 875 cytotoxicityuarrapoptosis (uarrcaspase-3) uarrnecrosisPTPC opening arrest at G0G1
Human melanoma SK-28 JA darrproliferationMJ uarrcytotoxicity uarrcell death
Human breast carcinoma MCF-7 JA darrproliferationMJ uarrcytotoxicity uarrcell death
Murine lymphoma EL-4 MJ uarrcytotoxicity
MJ 30mMno vehicle reported
Acute human T-lymphoblasticleukemia Molt-4
uarrp38 uarrJNK uarrAP-1 howevercell death MAPK-independent [18]
JA CJ andMJ (mM) Prostate PC-3 HTB-81 darrproliferation [46]
MJ 00ndash04mMno vehicle reported
Human myeloidleukemia HL-60 cells
darrcell growth uarrMAPKuarrdifferentiation to mature cells [41]
MDDHJ(synthetic)
0ndash250 120583Mno vehicle reported
Human myeloidleukemia HL-60 cellsMonocytoid leukemiaU937 THP-1 cellsPromyelocytic leukemiaNB4 cellsLung adenocarcinomaPC9 PC14 cells
MDDHJ more potent than MJdarrcell growth uarrMAPKuarrdifferentiation to mature cells
[41]
MJ 05ndash50mM Human lung adenocarcinoma A549 darrproliferation uarrROS uarrapoptosis(uarrBaxBcl-Xs uarrcaspase-3) [36]
JA CJ andMJ 05ndash30mMEtOH
Acute human T-lymphoblasticleukemia Molt-4 cells
Molt-4 uarrmitochondrial membranedepolarization uarrcyt c releaseuarrswelling uarrcell death
[38]
Lymphocytes from CLL patientsCLL cells uarrmitochondrialmembrane depolarizationuarrcytotoxicity
Liver carcinoma Hep 3B cellsHep3B uarrmitochondrial membranedepolarization (PTPC mediated)uarrcyt c release uarrswelling uarrcell death
Human fibroblast 3T3 cells(nontransformed cell line)Normal blood
Nontransformed 3T3 cells nocytotoxicityNormal lymphocytes nocytotoxicity
JA MJ 025ndash30mMEtOH
B-lymphoma clone29M62 (wild type p53)B-lymphoma clone
wt p53 cells uarrapoptosismutant p53 uarrnonapoptotic celldeath [17]
29M610 (mutant p53 resistant totreatment)
MJ uarrsim90 ATP depletion in bothcell types2DG high Glc but not pyruvatedarrATP
CJ MJ 05ndash25mM Nonsmall cell lung cancer linesA549 and H520
darrproliferation cell cycle arrest atG2M phase uarrp38 and ERK12phosphorylation uarrBaxuarrp21uarrcaspase-3
[37]
4 International Journal of Cell Biology
Table 1 Continued
Jasmonate (JAs conc range)vehicle Cell lines Effects References
MJ 05ndash20mM Hormone-refractory prostateadenocarcinomas PC-3 DU-145 darrproliferation uarrapoptosis darr5-LOX [47]
MJ 1ndash26mMEtOHmdash(IC50)
Murine melanoma cellsB16F10 and B16 COLR(overexpressing Pgp MDR)
darrcell motility darrcell growth darrMDR [39]
TBrJA(synthetic)
40 120583MEtOH(MJ doses lower than
in vitro)
Melanoma B16-F10Breast MCF-7Pancreas Mia PCA-2D122 PBL
uarrcitoxicity (TBrJA≫MJ) [39]
MJ 05ndash30mMno vehicle reported
CT-26 (murine colon carcinoma)B16 (murine melanoma)BCL1 (murine B-cell leukemia)Molt-4 (human T-lymphoblasticleukemia cell line)
MJ (but not JA) detached HK1 andHK2 from VDAC1 in isolatedmitochondria from thefour cell linesMJ did not inhibit HKs activitydarrATP uarrcyt c release uarrmitochondriaswelling uarrcell death
[20 30]
MJ 10-20mM Human neuroblastomaBE(2)-C
Arrest at S-phase darrcell growthdarrXIAP mRNA darrsurvivin mRNAuarrapoptosis
[43]
MJ 05ndash30mMDMSOHuman breast cancer MCF-7Human melanocytic MDA-MB-435cells [74]
Arrest at G0G1and S-phase darrmembrane fluidity uarrapoptosisextrinsic (TNFR1 uarrcaspase-8)intrinsic [darrΔ120595
119898uarrcaspase-3 (only
MDA-MB-435)]
[25]
CJ MJ 20mMDMSO Hormone-independent prostatePC-3 DU-145 cells
Cell cycle arrest darrcell growthuarrapoptosis(uarrTNFR1 uarrcaspase-3)
[48]
JA CJ andMJ 10-20mMDMSO
Human neuroblastomaSH-SY5Y
Arrest at G2M phase darrcell growthuarrapoptosis (darrXIAP darrsurvivin)activities MJ gt JA gt CJ
[44]
Human embryonic kidney HEK 293cells Not affected by MJ
MJ 10-20mMDMSO Human neuroblastomaSK-N-SH BE(2)-C
Arrest at G0G1 phase darrcell viabilitydarrmRNA of PCNA uarrapoptosis(darrXIAP darrsurvivin)
[45]
MJ 05ndash30mMEtOH Sarcomas MCA-105 uarrpAkt (correlates with lowersensitivity to cytotoxicity by MJ) [49]
SaOS-2 (resistent to MJ) MJ + 2DG uarrcytotoxicity
MJ 10ndash50mMEtOH
Cervical cancer SiHA CaSki andHeLa cells having wt p53Cervical cancer C33A cells (withmutated p53)
darrcell cycle uarrapoptosis throughdifferent pathwaysdarrATP uarrlactate (in more glycolyticCaSki) PARP cleavage multiple celldeath pathways depending on levelsof p53 p21 Bcl-2 and Bax
[26]
JA MJ 025ndash40mMEtOH Acute myelogenous leukemia cellsHL-60 and KG1
uarrROS uarrMJ-induced mitochondrialmembrane depolarizationuarrMJ-induced Mit SOD darrAKRC1
[132]
MJ MDDHJ 015mMDMSO Leukemia HL-60 cellsuarrCa2+-binding protein S100Puarrdifferentiation uarrregulator ofG-protein signaling-16 (RGS16)
[42]
J7 (synthetic) IC50 15 120583MHuman cervical carcinomaHeLa cells
Cell cycle arrest at G2M phasedarrBcl-2 (caspase 9 3) DNA damage [27]
International Journal of Cell Biology 5
Table 1 Continued
Jasmonate (JAs conc range)vehicle Cell lines Effects References
J7 (synthetic) 50120583MDMSO Human hepatoma Hep3B
uarrBaxBcl-2 ratio uarrDR5 uarrcaspase-8darrBid uarrapoptosis correlated withuarrcaspase-9 uarrcaspase-3 darrXIAP darrcIAPdarrPARP ExtrinsicintrinsicMAPK
[34]
MJ 05ndash25mMno vehicle reported
CD138+ tumorcells from MM patients
HK2 release from mitochondriarapid darrATP uarrapoptosis [105]
MJ 025ndash10mMno vehicle reported Human colorectal cancer cells CRC
uarrTRAIL uarrcyt c releaseuarrcaspase cleavage darrsurvivindarrTCF transcriptional activity
[32]
MJ 00ndash20mMEtOH Cervical cancer cellsSiHa CaSki HeLa and C33A
uarrmitochondrial O2
minus (HeLa CaSki)darrsurvivin darrE6 E7 (HPV)uarrdifferent cell death pathways(independently of HPV)
[28]
J7 (synthetic) 0ndash50 120583M DMSO Human hepatoma HepG2uarrROS uarrTRAIL-mediated apoptosis(darrBid darrXIAP darrcIAP darrBcl-xLuarrcaspases)
[35]
MJ 30mM (IC50) novehicle reported
Human adenocarcinoma colonHT-39
Arrest at S-G2M uarrcytotoxicityuarrapoptosis [31]
JA MJ 10ndash30mMDMSO Canine macrophagic malignantDM62 cells darrcell growth (MJ gt JA) uarrcytotoxicity [256]
MJ 05ndash02mMDMSO Human gastric SGC-7901 MKN-45cell lines
darrmigration darrinvasion darrangiogenesisdarrMMP-14 [33]
Bcl-2 B-cell lymphoma-2 Bcl-xL B-cell lymphoma-extra large Bid BH3 interacting domain death agonist cIAP cellular inhibitor of apoptosis CJ cisjasmonicacid 2DG 2-deoxy-D-glucose Glc glucose J7 methyl 5-chloro-45-didehydrojasmonate JA jasmonic acid NSCLC nonsmall-cell lung carcinoma PgpP-glycoprotein MDDHJ methyl 45-didehydro-jasmonate MDR multidrug resistance MMP-14 matrix metalloprotease 14 PARP poly (ADP-ribose)polymerase PCNA proliferating cell nuclear antigen PTPC permeability transition pore complex ROS reactive oxygen species S100P protein SP100 TBrJA57910-tetrabromo jasmonate TNFR1 tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand XIAP X-linked inhibitor of apoptosis protein
Table 2 Effect in vivo of natural and synthetic jasmonates derivatives
Jasmonates Conc rangevehicle Organismtissuecells Effects ReferencesMJ (oral) 236mgkglipofundin Normal C57BL6 mice Nontoxic [15]
MJ (oral) 236mgkglipofundin C57BL6 mice injected ip withmurine T-lymphoma EL-4
uarrsurvival of treated mice ascompared to the inoculated control [15]
TBrJA40120583MEtOH
(lower doses than MJin vitro)
Murine melanomaB16-F10 darrlung metastasis [39]
MJ 1ndash10120583MEtOH Chicken CAM darrangiogenesis [40]MJ 1ndash10120583MEtOH Human endothelial cells (HUVEC) darrCOX-2PGE2 pathway [40]
MJ (ip) 1 gKg in 01mLvegetal oil ip
Multiple myeloma- (MM1S-)inoculatedin nodscid mice
uarrsurvival of treated mice ascompared to the inoculated control [105]
MJ (topical) 1 gmL oil Application on cancerous andprecancerous human skin lesions
No local or systemic side effects38 patients withpositive responses
[51]
CAMchorioallantoic membrane of chicken embryoHUVEC human umbilical vein and endothelial cells TBrJA synthetic 5 7 9 10-tetrabromo jasmonate
mentioned that MJ was also assessed and approved by Foodand Agriculture OrganizationWorld Health Organization(FAOWHO) as a food additive Toxicity results reportedin the EPA document [52] were found to agree with thosepreviously published by other groups [50 51]
21 Jasmonates (MJ) Can Cause Cancer Cell Cycle Arrest atDifferent Phases in Different Cancer Cell Types Inhibiting
Growth and Proliferation The eucaryotic cell cycle is dividedinto four phases G
1 S G
2 and M that occur in response
to growth factors or mitogens The DNA synthetic (S) andmitotic (M) phases are preceded by gap phases (G
1 G2)
Chromosome duplication occurs during S phase replicatedchromosomes are segregated into individual nuclei (mitosis)during M phase and the cell then splits in two Cell cycleprogression can be regulated atG
1 G2points by various intra-
cellular and extracellular signals If extracellular conditions
6 International Journal of Cell Biology
Table 3 Effect in vitro and in vivo of methyl jasmonate (MJ) combined with other anticancer agents
Jasmonate Drug concrangevehicle Model Effects References
(1) MJ + BCNU(nitrosourea)in vitro
MJ Fixed conc Pancreatic MIA PaCa-1 Mitochondriotoxicsynergic cytotoxicity
[170](2) MJ + 2DGadriamycin taxolBCNU or cisplatinin vitro
MJ05ndash20mMEtOHTaxol cisplatin1ndash10120583gmL
CT26 DA-3 GTRAMP C1 MCF7MIA PaCa-2 D122 and BCL1
Strong cooperativeeffects of MJ + 2DGand MJ + other drugs
(3) MJ + adriamycinin vivo
MJ iv20ndash150mgkgdissolved inlipofundinAdr (DOX) ip4mgkg
Balbc mice injected ip with 1 times 104chronic BCL1 cells lymphocyteleukemia cells
MJ + Adr significant prolongedsurvival effect
(4) MJ + PI3KAktinhibitorsin vitro
MJ 05ndash30mMEtOHDMSO
Sarcoma MCA-105 SaOS-2 darrMJ-induced activation of Aktuarrsynergic cytotoxicity
[49](5) MJ + 2DGin vitro
MJ 05ndash30mMEtOHDMSO Sarcoma MCA-105 SaOS-2 2DG darrpAkt darrMJ-ind pAkt
MJ + 2DG uarrsynergic cytotoxicity
(6) MJ + irradiationin vitro
05ndash20mMDMSO
Irradiated prostate PC-3 (radiationinduces Bcl-2 expression)
darrradiation-induced Bcl-2uarrradiation sensitivity PC-3uarrcaspase-3
[104]
(7) MJ andor POHandorcis-platin (CP)in vitro
Both testedat IC20
Human MDA-MB-435[74]
MJ + POH uarrTNFR1darrΔ120595119898uarrcytotoxicity cell cycle arrest
at G0G1 uarrapoptosis+ CP all effects enhanced
[251]
Human breastMDA-MB-231 uarrapoptosis
(8) MJ + TRAILin vitro
MJ 05mM +TRAIL(100ndash200 ngsdotmLminus1)
CRC cancer cells
darrsurvivin (IAP)darrWntTCF pathwayuarrTRAIL-induced apoptosisuarrcaspase activity
[32]
(9) MJ + Smacin vitro
MJ 05ndash20mMDMSO
Human bladder cancer EJ T24 Synergy uarrIAPs-bound caspase 3uarrapoptosis [254]
Human embryonic kidney HEK293 No cytotoxicity
(10) MJ + Smac7N(IAP antagonist)in vitro
MJ 05ndash20mMDMSO
Hormone-independent prostateDU-145 PC-3Human proximal tubular epithelialHK-2 cells (overexpressing IAPs)
Smac7N uarrMJ-induced apoptosis bycaspase-9-dependent (intrinsic) andindependent (extrinsic) pathways
[255]
(11) MJ + cisplatin(01ndash05 120583M)MJ + X-rays(025ndash3Gy)MJ + 120572-rays
MJ 01ndash10mMEtOH
Cervical cancer cellsSiHa CaSki HeLa and C33A
darr viabilitydarrsurvivaldarrIC50 radiation dose
[29]
(12) MJ + 5-FUin vitro
05mMMJ1 hthen + 5-FU
Human adenocarcinomacolon HT-39 darrIC50 5-FU (5 rarr 25mM) [31]
BCNU 13-bis-(2-chloroethyl)-1-nitrosourea 2DG 2-deoxy-D-glucose 5-FU 5-fluorouracile IAP inhibitors of apoptosis MM multiple myeloma POHperillyl alcohol smac second mitochondria-derived activator of caspases Smac7N a peptide that contains the N-terminal seven residues of smac TNFR1tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand
are unfavorable cells delay progress through G1and may
enter a quiescent state known asG0 in which they can remain
for days weeks or years before resuming proliferation Ifextracellular conditions are favorable and signals to growand divide are present cells in early G
1or G0progress
through a restriction checkpoint late in G1after which cells
are committed to DNA replication even if the extracellularsignals that stimulate cell growth and division are removed[53 54] Cell cycle activity is modulated by intracellularROS levels and regulates cell survival cell proliferation andapoptosis [53ndash58]The progression through theG
1 S G2 and
M phases is promoted by cyclin-dependent kinases (CDKs)
International Journal of Cell Biology 7
which are positively and negatively regulated by cyclins andcyclin kinase inhibitors (CKis) respectively and by ROSUnlike the passage through the S G
2 and M phases G
1
progression normally relies on stimulation by mitogens andcan be blocked by antiproliferative factors passage throughthe restriction checkpoint and entry into S phase is controlledby CDKs that are sequentially regulated by cyclins D Eand A [53 54 56ndash58] In general CDK activity requirescyclin binding and depends on both positive and negativeregulatory phosphorylations [54] ROS levels influence thephosphorylating and ubiquitinating enzyme activites thuscontrolling cell cycle progression [54 57 58]
A master gene regulating cell passage through the cycleis the Myc family of transcription factors it switches onthe simultaneous synthesis of thousands of different proteinsrequired for cell growth and cell cycle function [58 59] TocounteractMyc protein activity the cell cycle is also regulatedby the p53 protein [60 61] a DNA-binding transcriptionfactor that senses DNA damage (occurring after exposureto ionizing radiation UV light oxidative stress or otherDNA damaging agents) and activates genes to halt cell cycleprogression in G
1for repair or otherwise initiates apoptosis
[56 62] When the damage caused by cell stress is mild p53activatesmechanisms for cell cycle arrest and repair damagedDNA if the damage is irreparable p53 initiates apoptosis [63]In case ofmild cellular stress low levels of p53 can activate theTP53-induced glycolysis and apoptosis regulator (TIGAR)gene that decreases the rate of glycolysis and hence ROS levelsin the cell [64] Glycolysis is essential for cancer progressionbut TIGAR provides protection from ROS and apoptosisHowever if damage caused by stress is irreparable p53prevents proliferation of cells with altered DNA and inducesincreased levels of ROS to trigger cell death [56] Increasedlevels of active p53 inhibit the cell entry into the S-phaseor induce apoptosis [56] Loss or inactivation of p53 resultsin loss of cell-cycle arrest or inhibition of apoptosis afterDNAdamage or physiological stress [65] leading to increasedgenetic instability increased accumulation of mutations andultimately oncogenesis [61] When the cell cycle is no longerregulated by p53 cells remain cycling (supported by 119888-Mycoverexpression) subverting cell cycle exit maturation andterminal differentiation [66] Over 50 of all human cancersharbor mutations and genetic alterations in cyclins and p53genes directly affecting the function of critical cell cycleproteins rendering the p53 gene ineffective in mediating cellgrowth arrest and promoting cancer cell death [62 67 68]ROS production is under p53 regulation and in turn p53being a redox-sensitive protein is influenced by ROS levelsthus ROS can act as both an upstream signal that triggersp53 activation and as a downstream factor that mediatesapoptosis [69]Thus mutations in the p53 gene downregulateits activity and consequently ROS production [70]ThroughROS p53 can also directly control metabolic traits of cells[71] regulate mitochondrial membrane potential (Δ120595
119898) and
induce cytochrome 119888-independent apoptosis (blocked by Bcl-2) ROS-mediated disruption of Δ120595
119898constitutes a pivotal
step in the apoptotic pathway of p53 and this pathwaydoes not involve cytochrome 119888 release [72] ROS can thusregulate cell fate through p53 in a way that physiological ROS
levels trigger cell protective pathways while under cytotoxicoxidative stress p53 behaves more like a cell killer [70 73]
JAs (mainly MJ) have been found to arrest cell cycle ofdifferent cancer cell types at different phases (Table 1) G
0G1
in human acute lymphoblastic leukemia Molt-4 [14 38] andhuman neuroblastoma SK-N-SH BE(2)-C [45] G
0G1-S in
human breast MCF-7 andmelanocytic MDA-MB-435 cancercells [25 74] S in human neuroblastoma BE(2)-C [43] S-G2M in human adenocarcinoma colon HT-39 [31] G
2M
in nonsmall cell lung cancer (NSCLC) lines A549 and H520[37] human neuroblastoma SH-SY5Y [44] human cervicalcarcinoma and HeLa cells [27] At 01mM MJ arrested invitro human umbilical vein endothelial cells (HUVEC) at G
1
[40]The molecular mechanisms underlying the above jasmo-
nate-mediated cell cycle arrest in cancer and embryonicHUVEC cells in vitro are not yet completely understoodChecking the above data against the UMD TP53 mutationdatabase (httpwwwp53fr) and revising the article byBerglind et al [75] we could not find correlation between agiven MJ-mediated phase arrest of cell cycle and the TP53wild type or mutated status of a cancer cell type (data notshown) a result that would agree with the original observa-tion of Fingrut and Flescher [15] stating thatMJ-mediated celldeath is independent of the p53 status [17] However otherstudies suggest thatMJ could be acting by interfering differentcell cycle regulatorymechanisms In plants MJ is able to bindthe TGACCG motif which is present in cell cycle cyclin pro-moters [76] wemight speculateMJ recognition of TGACCG-like motifs in mammalian cells cyclin promoters howeverthis would need experimental confirmation In some plantsMJ impairs the G
2-to-M transition by repressing M-phase
gene activation [77] MJ impaired the G2-to-M transition in
the human neuroblastoma SH-SY5Y cells downregulatingthe expressions of the proliferating cell nuclear antigen(PCNA a processivity factor encircling DNA at sites ofreplication and repair) andN-Myc [45] Asmentioned aboveMYC is the master gene allowing cancer cells to move alongthe cell cycle and regulating most genes needed to meet themetabolic demands for cell proliferation and survival [78]Cancer cell cycle arrest at different phases is in itself closelydependent on the cell energy and as such it is extremelydependent on metabolic ATP availability and its use byhexokinase Downregulation of Myc byMJ [45] is interestingresult that must be confirmed particularly because there is alack of specific inhibitors against Myc [57]
In some cases of cell cycle arrest MJs have been foundto upregulate p21 [37] whereas in others to downregulatesurvivinmRNA and protein levels [43ndash45] (Table 1) Survivinis an important member of the inhibitor of apoptosis protein(IAP) family that also regulates entrance of the cell cycle to theM phase and is overexpressed in cancer cells acting linkingboth regulation of the cell cycle and inhibition of apoptosis[79]
22 Jasmonates (MJ) CanCauseCancer Cell Death by InducingApoptosis andor Necrosis Apoptosis can be initiated by twosignaling cascades namely the extrinsic and the intrinsic
8 International Journal of Cell Biology
pathway The classical extrinsic pathway is triggered by thebinding of extracellular signals like tumor necrosis factor(TNF) tumor necrosis factor-related apoptosis-inducing lig-and (TRAIL) and apoptosis antigen-1 (Fas) ligand (FASL)to death receptors such as tumor necrosis factor receptor 1(TNFR1) and FAS [80]This interaction leads to procaspase-8activation which cleaves itself into caspase-8Then caspase-8can cleave and activate procaspase-3 into caspase-3 thatcleaves many substrates inside the cell inducing the typicalmorphological changes seen in apoptosis The intrinsic path-way can be triggered by intracellular stress signals such asoxidative stress and DNA damage The diverse signals aresensed by mitochondria through the balance between pro-and antiapoptotic members of the Bcl-2 family of proteinsWhen proapoptotic signals overcome antiapoptotic signalsmitochondria becomepermeabilized leading to dissipation ofthe mitochondrial electrochemical potential release of inter-membrane proteins like cytochrome 119888 SmacDIABLO AIFandHTRA2OMI Cytochrome 119888 interacts with procaspase-9and Apaf-1 to form the apoptosome complex that leads tocaspase-3 activation [81] The extrinsic pathway can connectwith the intrinsic pathway through the cleavage by caspase-8of Bid into tBid (truncated Bid) which is a proapoptoticmember of the Bcl-2 family and can lead to mitochondriapermeabilization [82 83] The members of the Bcl-2 fam-ily regulate mitochondria permeabilization and comprisesantiapoptotic (Bcl-2 Bcl-xL) proapoptotic (Bax Bak) andproapoptotic BH-3 only proteins (Bad Bid Puma andNoxa)[82ndash84] One of the mechanisms of mitochondria perme-abilization is controlled by Bax and Bak proteins that arenormally expressed in every cell but are maintained inhibitedby the antiapoptotic members When there is a signal toapoptosis BH3-only proteins interact with the antiapoptoticmembers releasing BaxBak or directly activating themleading to their oligomerization and to mitochondrial outermembrane permeabilization (MOMP) Another mechanismfor mitochondria permeabilization is the formation of themitochondrial permeability transition (MPT) pore which iscomposed by the voltage-dependent anion channel VDACthe adenine nucleotide translocase (ANT) and cyclophilin D[85]TheMPT pore comprising VDAC1 is directly regulatedby Bad [86] In contrast VDAC2 has been identified as anantiapoptotic negative regulator of Bak [87ndash89]
One of the hallmarks of cancer is resistance to cell death[90] This resistance is acquired by a host of different geneticdefects in cancer cells such as those present in the p53tumor suppressor gene [62 68 91] Normal p53 responds togenotoxic oncogenic and other stress signals by inducingantiproliferative transcriptional programs leading to growtharrest (for cell repair) or apoptosis [92] p53 mediates cyclearrest by transcriptionally activating the p21 (CDKN1A) 14-3-3120590 (SFN) and GADD45120572 genes whereas p53-dependentapoptosis is triggered by turning on the synthesis of proteinsthat produce ROS [58 93] and transactivating differentmitochondrial proapoptotic genes as well as death receptorsfor the synthesis of the corresponding proteins [58 94]Whenp53 (and its target genes) expression levels are below a thresh-old they induce cell cycle arrest but not apoptosis above thisthreshold p53 trigger apoptosis [95] Lowering this threshold
with inhibitors of the antiapoptotic Bcl-2 family proteinssensitize cells to p53-induced apoptosis [95] By damagingcells chemotherapeutical drugs turn on p53rsquos transcriptionalfunction that upregulates the expression of proapoptotic pro-teins Defects in the p53 function or in the apoptotic cascadeslead to cell phenotypes resistant to chemotherapy Apoptosiscan also be induced by anticancer agents independently ofp53 by either directly modulating the apoptotic machineryor directly interacting with mitochondria [96] ROS levelsmight be playing a central role in mediating various forms ofnonapoptotic caspase-independent programmed cell deatheither through direct irreversible oxidative damage to keyproteins nucleic acid molecules and cellular structure or bythe activation of prodeath signal transduction programs [97]In a ROS-dependent nonapoptotic cell death pathway respi-ration through the mitochondrial electron transport system(ETS) chain is not essential and ROS is generated throughNADPH oxidases [97] In contrast to the mitochondrial ETSwhich generates ROS as a byproduct of respiration NADPHoxidases produceROS as their primary function [98] Raise ofROS is characteristic after ionizing radiation chemotherapyand targeted therapy treatments [58 99] and may be killingcancer cells through a nonapoptotic ROS-dependent celldeath pathway
Tumor cells maintain high ATPADP as well asNADHNAD+ ratios so as to assure that ATP levels arenever limited ATP depletion would represent then anunsustainable metabolic stress for tumor cells and it isassociated with necrotic death in some cell types [100]Recently it was shown that a signaling cascade centered onthe proteins RIP1 and RIP3 can trigger cell death by necrosisin many situations and the RIP1-RIP3 complex appears topromote ATP depletion during necroptosis by impingingon components of the MPT pore [101 102] Necrosis was aterm used to describe a process of accidental cell death inwhich cells swell and burst releasing its intracellular contentwhereas necroptosis modernly refers rather to an orderedcell death program different from apoptosis apoptosis is acaspase-dependent cell death process whereas necrosis iscaspase-independent process [102 103] Studies concerningnecrosis have focusedmainly on cell membrane permeabilityto propidium iodide (PI) in the flow citometry doublestaining with annexin-VPI
Fingrut et al [17] by studying the effect of MJ ontwo clones of a highly malignant B-cell lymphoma (oneharboring a normal wild type (wt) p53 gene while the otherexpressed a mutated inactive p53 protein) (Table 1) reportedthat MJ was equally toxic to both clones [17] In contrastthe p53-mutant cells were resistant to radiomimetics (drugshaving effects similar to those produced in radiotherapy)and chemotherapeutic drugs In this cancer cell model MJinduced mostly apoptotic cell death in the wt p53-expressinglymphoma cells while no signs of early apoptosis weredetected in the mutant p53-expressing lymphoma cells thatwere killed by MJ through a p53-independent nonapoptoticcell death mechanism However other cancer cell types canundergo apoptosis with or without functional p53 and aswe will see MJ can induce apoptotic andor nonapoptoticcell death in wt p53 as well as in TP53 mutated cells
International Journal of Cell Biology 9
(Table 1) Nonapoptotic cell death could be the consequenceof the severe bioenergetic damages induced by the directinteraction of MJ with mitochondria leading to increasedROS levels inhibition of ATP synthesis and ATP depletion[20 30] Jasmonates including MJ and some of its analogshave been found to be strongly proapoptotic in vitro indifferent types of cancer cells by both the extrinsic and theintrinsic pathway (Table 1) with no or little cytotoxicity tonormal cells [15 25 36 37 44 45] Thus MJ induced theexpression of TNFR1 in human breast MCF-7 and MDA-MB-435 cells [25 74] and in hormone-independent humanprostate PC-3 and DU 145 cancer cells promoting apoptosisby the extrinsic pathway in these cells [48] TRAIL (TNF-related apoptosis inducing ligand) is able to induce apoptosisin prostate cancer cell yet overexpression of antiapoptoticproteins and inhibition of proapoptotic proteins results indiminishing the TRAIL-mediated apoptosis pathway TRAILhas emerged as a potent anticancer agent in laboratory-based studies and preclinical trials and JAs have been foundto sensitize cancer cells to TRAIL-mediated apoptosis [32](Table 3) J7 a synthetic analogue of MJ enhanced TRAIL-mediated apoptosis through upregulation of ROS levels inhuman hepatomaHepG2 cells apoptosis was induced via Bidcleavage downregulation of X-linked inhibitor of apoptosisprotein (XIAP) cellular inhibitor of apoptosis-1 (cIAP-1)B-cell lymphoma-extra large (Bcl-xL) and activation ofcaspases [35] Other examples of MJ-mediated apoptoticandor nonapoptotic cell death in different cancer cell types(Table 1) are as follows (1) MJ induced ROS production inA549 lung adenocarcinoma cells activating Bcl-2-associated-X-protein (Bax) as well as Bcl-2-Like-1 (Bcl2L1) proapoptoticproteins in these cells [36] (2) MJ activated caspase-3 inleukemia Molt-4 prostate SK28 and LNCaP cancer cells[15] (3) MJ inhibited the antiapoptotic Bcl-2 protein andenhanced proapoptotic proteins in radio-resistant humanprostate cancer cell line PC-3 [104] (4) MJ suppressed theproliferation of the neuroblastoma cell lines SH-SY5Y SK-N-SH and BE(2)-C through a strong apoptosis-inducingeffect as measured by annexin VFITCPI MJ treatment alsodownregulated the expression of the antiapoptotic proteinsXIAP and survivin [44 45] (5) MJ-mediated cell death wasobserved on cervical cancer and myeloma cells and it wasreported to be due to mixed characteristics of apoptosis andnecrosis [26 105] So far studies showing MJ inducing theRIP1 signaling cascade have not been reported howeverit will be important to deep into these analyses to cleardistinguish between MJ-induced apoptosis andor necrosis
23 Methyl Jasmonate Induces the Production of ReactiveOxygen Species (ROS) Responses in Some Cancer Cell TypesLeading Also to Cell Death Intracellular ROS levels reg-ulate cell cycle activity cell metabolism and survival cellproliferation and apoptosis [53ndash58] Stressful conditionsproduce a transient excess of intracellular ROS in normaltissues however under chronic pathological and oxidativestress conditions ROS is continuously produced in excessand persistently causing damage to DNA cell membraneslipids proteins and inducing apoptosis to control and avoid
oxidative damages cells counteract excess ROS by enzymatic(superoxide dismutase (SOD) glutathione peroxidase (GPx)and catalase) andor nonenzymatic (glutathione thoredoxin)antioxidant elements [99 106] Controlling ROS is central tointracellular redox signaling and homeostasis on the otherhand uncontrolledunbalanced ROS production modifiesintracellular redox leading to apoptosis [58 82 99 106] ROScan be generated by different biological systems (1) director indirect extracellular stimulation of plasma membrane-bound NAD(P)H oxidases (NOX) can generate intracellularH2O2[57 107ndash110] (2) biochemical pathways associated to
fatty acid oxidation and lipoxygenases activities generateROS as normal byproducts of these activities [111] (3)ATP generation by oxidative phosphorylation (OxPhos) inmitochondria is accompanied by the production of ROS as aconsequence of electron leakage from the electron transportchain mitochondria are considered to be major sources ofROS inmammalian cells particularly under a variety of stress-ful conditions [82 99] Superoxide (O
2
minus) is mainly produced
from complexes I and III and it is rapidly dismutated to H2O2
and oxygen by SOD [112] H2O2serves both as a toxic oxidant
and as an essential signaling molecule regulating cellularbiological processes [113] A strong positive correlation existsbetween mitochondrial membrane potential (Δ120595
119898) and ROS
production [106 114 115] Mitochondria produce more ROSat high membrane potential and an increase in the Δ120595
119898
produced either by a closure of MPT pore or by inhibitionof the ATP synthase is associated with increased ROS pro-duction However in cases of mitochondrial dysfunctionslow Δ120595
119898and decreased activity of the respiratory chain
are accompanied by a simultaneous increase in ROS [116117] It has been reported that cancer cells have higher ROScontent compared to normal cells [118 119] probably due toabnormal respiration because of dysfunctional mutations inmitochondria [120] but they also have increased contents ofantioxidants (SOD catalase glutathione thioredoxin etc)to counteract ROS [58 121] Tumor cells cannot tolerateexcessive high ROS levels [118 122] and as a matter of factradiation therapy chemotherapies and targeted therapiesinduce apoptosis and kill some cancer cells by generatingexcess ROS [99 106 123 124] Thus inducing higher levelsof ROS in cancers cells can be exploited as an effectiveselective strategy to kill tumor cells over normal cells [119]However prolonged treatment with the same drug reducesROS levels in cancer cells [124] indicating that when theybecome drug-resistant tumor cells have lower ROS contentthan drug-sensitive cancer cells Evidence suggests that drug-resistant cells have a higher expression of catalase at theplasma membrane that keeps reduced ROS levels [121]
In plants MJ-mediated damage to mitochondria is fol-lowed by a rapid production of H
2O2[125] H
2O2is used as
a second messenger for the induction of genes and productsrelated to defense against both herbivores and pathogens[126] in addition H
2O2induces apoptosis and senescence
[127 128] On the other hand it has been signaled that theMJ-induced production of H
2O2in plants was prevented by
inhibitors of NAD(P)H oxidase [128 129] indicating anothermechanism of MJ-mediated production of ROS and a majorrole for this enzyme in MJ-induced ROS responses MJ
10 International Journal of Cell Biology
induces also an endogenous H2O2response in some cancer
cell lines (Table 1) [19 36 38 130ndash132] In A549 cancerlung cells the H
2O2response induced by MJ increased the
expression of the proapoptotic Bax and Bcl-Xs (although notbeing that of the antiapoptotic Bcl-2 and Bcl-xL proteins) andthis led to apoptosis this event was inhibited by catalase (aspecific inhibitor of H
2O2) [36] MJ increased the intracel-
lular levels of H2O2and superoxide (O
2) ions in human C6
glioma cells in vitro and induced the expression of heat shockprotein 72 (HSP72) a negative regulator of H
2O2-induced
oxidative stress in these cells in vitro MJ-induced HSP72expression was blocked by ROS inhibitors [130] ElevatedROS generation was observed in response to both MJ andJA levels in the acute myeloid leukemia HL-60 and KG1a celllines but other factors like the MJ-mediated generation ofmitochondrial SOD rather than ROS were determinant forapoptosis or differentiation in these cells [132] Suppression ofMJ-induced apoptosis by antioxidants like N-acetylcysteine(NAC) and catalase but not by inhibitors of hydroxyl radicalsand superoxide ions indicates that H
2O2is nonetheless one
of the important factors involved inMJ anticancer effects [19]
24 By Dissociating Hexokinase (HK) from the VoltageDependent Anion Channel (VDAC) on the Outer Mitochon-drial Membrane MJ Dissociates Glycolysis from OxidativePhosphorylation Causing Severe Bioenergetic Deregulations inCancer Cells Metabolism and cell growth are two cellularprocesses that are tighly linked and regulated [133] Theclassic bioenergetics phenotype of cancer cells of enhancedglycolysis under aerobic conditions was described by War-burg [134] However the ldquoWarburg effectrdquo does not neces-sarily imply mitochondrial dysfunction [135] To increaseglycolysis cancer cells upregulate the transcription of genesinvolved in the glycolytic pathway (ie glucose transportersglycolytic enzymes etc) This metabolic reprogrammingallows increased rate of ATP production synthesis of lipidsand a new redox balance Cancer cells actually use a com-bination of both glycolysis and mitochondrial respirationto produce energy and they may vary in regards to thepreferential use of these pathways being in some caseseither more glycolyticless oxidative or less glycolyticmoreoxidative depending on the prevalent normoxic or hypoxicenvironmental conditions and their capacities to expressadequate levels of oncogenes and tumor suppressor geneproducts for cell growth [136ndash139] or when either glycolysisor OxPhos is inhibited in whose case there can be partialcompensation by the other metabolic pathway [17] Shiftingthe balance between those two processes with glycolyticandor mitochondrial enzyme inhibitors might represent aninteresting way to look for new anticancer agents
The first step in the metabolism of glucose is catalyzedby the enzyme hexokinase (HK) which by phosphorylatingglucose makes it negative sequestering it into the cytoplasmin the form of glucose-6-phosphate (G6P) In normal tissuesthis crucial step is catalyzed by four different HK isoforms(HK1 HK2 HK3 and glucokinase) indicating that regulationof glucose phosphorylation can vary in different tissues underdifferent condition [140] HK1 and HK2 have overlapping
tissue expression but different subcellular distributions withHK1 associated mainly with mitochondria and HK2 associ-atedwith bothmitochondrial and cytoplasmic compartments[141] HK1 is widely and constitutively expressed whereasHK2 predominates during embryogenesis and is not widelyexpressed in adult tissues but only in a limited numberof normal adult tissues [141] their different subcellulardistributions and kinetic properties reflect their differentmetabolic roles [141 142] Under physiological conditionsHK1 is predominantly bound to the outer mitochondrialmembrane where it primarily channels glucose toward gly-colysis [140] whereas HK2 is mainly soluble and controlsglycogen formation [140 141] In malignant tissues HK1and HK2 isoforms are glycolytic and overexpressed (HK1more than HK2) (httpwwwproteinatlascom) [143] andare tightly bound to mitochondria HK1 and HK2 bindthrough theirN-terminal hydrophobic regions to the voltage-dependent calcium channel isoform1 (VDAC1) the prevalentpore protein in the outer mitochondrial membrane [2482 143ndash146] VDAC1 is also overexpressed in cancer cells[147] and the binding of HK to VDAC1 on the outer mito-chondrial membrane is a fundamental aspect of the aerobicglycolytic metabolism of cancer cells [82 135 147 148]HKVDAC association appears to protect tumor cells frommitochondrial outer membrane permeabilization (MOMP)which marks a point of no return leading to cell death[149] VDAC transports ADP and inorganic phosphate (Pi)the substrates needed for the production of ATP into themitochondria it also transports ATP out of themitochondriainto the cytoplasm and facilitates the availability of ATPto HK2 [143 150 151] A continuous supply of glucose toHK maintains this activity at a constant level and controlsthe flux through the mitochondrial electron transport chaininfluencing mitochondrial ROS production [152] HK bindsthrough hydrophobic interaction to VDAC1 the prevalentpore protein in the outer mitochondrial membrane [24 82143ndash146] thereby assuring and gaining a preferential accessto ATP [82 150 153] Translocation of HK2 from the cytosolto the mitochondria is regulated through phosphorylationof HK by Akt [141] HK2 only binds VDAC after phos-phorylation by Akt [151 152] Glycogen synthase kinase 3beta (GSK3120573) a major target of Akt kinases phosphorylatesVDAC [154ndash157] and dissociates HK [156] HK2 boundto mitochondria enhance its affinity to ATP and is lesssensitive to enzymatic inhibition by the G6P product [140141 158] HK2 can produce G6P from cytoplasmic glucoseand mitochondrial ATP without regulatory restraints evenunder hypoxia supporting rapid cell growth [142 143 159ndash161]HK1 andHK2 aremore tightly bound toVDAC in cancercells than in nonmalignant cells [156 162 163] HKVDA1interaction is relatively stable for HK1 while it is dependentupon G6P levels for HK2 [140 141] The higher affinity ofmitochondrial HK2 for glucose ATP and VDAC1 representsa high advantage for cancer cells survival and growth con-ferring to them an aerobic glycolytic phenotype underlyingthe Warburg effect [82 135 138 142 147 148 150 160]Differential interactions of HK1 and HK2 with mitochondriamay underlie different glycolytic profiles in cancer cells [164165] As rate-controlling enzymes in the glycolyic pathway
International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 International Journal of Cell Biology
O
O
OCH3
CH3
Figure 1 Chemical structure of methyl jasmonate (cyclopen-taneaceticacid 3-oxo-2-(2-penten-1-yl)- methyl ester)
such as for instance phenolic compounds which are used aschemical defenses [6ndash12] For instance ultraviolet- (UV-) Bradiation triggers JA accumulation [10 13] whereasMJ is pro-duced under herbivore attack and can act as a volatile signalrecognized by neighboring plants to induce the synthesis ofprotease inhibitors [8] JAs alter plantmitochondrial functionand induce reactive oxygen species (ROS) leading to plant celldeath in a way similar to mammalian apoptosis [12 14]
MJ has been shown to have powerful anticancer activitythrough multiple mechanisms [15ndash24] The purpose of thisreview is to gather most relevant findings concerning MJ-mediated anticancer activities in an attempt to integrate theminto a functional view that could help us to develop newanticancer research strategies and therapies
2 Effects of Jasmonates onMammalian Cancer Cells
In 2002 Fingrut and Flescher [15] discovered that MJstrongly and adversely affected human lymphoblasticleukemia cells by suppressing their proliferation andinducing cell death while nontransformed lymphocytesand normal keratinocytes were unaffected [15] In theseinitial studies they showed that JAs selectively targetedcancer cells in an ex vivo mixed population of leukemic andnormal peripheral blood mononuclear cells (PBMCs) froma patient with chronic lymphocytic leukemia (CLL) JAs alsoincreased the life span of T-cell lymphoma-bearingmice [17]Thereafter JAs including MJ and related synthetic analogswere found to inhibit in vitro cancer cell proliferation andto induce cell death in other human and murine cancer celltypes [16 19ndash24] including human breast [15 25] cervix[26ndash29] colon [30 31] colorectal [32] gastric [33] hepatoma[34 35] lung [19 36 37] lymphoma [15 17 18 38]melanoma[15 30 39 40] myeloid leukemia [41 42] neuroblastoma[43ndash45] prostate [15 46ndash48] and sarcoma [49] cancercells (Table 1) Other results have shown that JAs and theirsynthetic derivates exerted selected cytotoxic effects invivo towards metastatic melanoma [21 39] and inhibitedangiogenesis at high doses (it was the reverse at low doses)in the chorioallantoic membrane (CAM) of chicken embryo[40] (Table 2) In general MJ has been found to be superior
to CJ and JA in terms of cytotoxicity and induction ofapoptosis in human cancer cells [33 38 44] Independentlyif jasmonates are dissolved in an organic solvent or not mostexperiments in vitro with JAs and MJ have been reportedto exert their biological effects at similar low millimolar(mM) concentrations (Tables 1 and 3) excepting few caseswhere MJ and particularly some of its chemical derivativeswere active at micromolar (120583M) concentrations in vivo(Table 2) Differential dispersion andor availability ofsmall hydrophobic MJ lipid droplets after phase separationin culture media or solubility in biological fluids in vivomight explain these differences Nonetheless JAs havebeen found to be nontoxic at doses higher than the usualpharmacological doses employed for other compounds (nM120583M) for instance an iv injection of 236mg MJkg bodyweight in mice (equivalent to asymp50mM circulating concin a 25 g adult mouse) was not toxic [15] The nontoxic ivdose was however found lower by Reischer et al in mice(75mgkg) [39] On the other hand oral acute toxicitystudies performed with MJ on 10 Sherman Wistar ratsreported a LD
50gt 5 gkg showing a normal necropsy
analysis whereas the dermal acute dose was LD50gt 2 gkg
from studies in 10 albino rabbits showing again a normalnecropsy analysis [50] No skin irritation was observed whenMJ was topically applied at 2 gkg to 10 albino rats and at 1MJ in 6 albino New Zealand rabbits similarly there was noskin irritation when a 10 MJ solution was topically appliedon 10 guinea pigs and on 50 volunteers (the latter assessedby the Human Repeat Insult Patch Test) [50] Repeatedapplication of the 10 MJ solution at the same skin sitein guinea pigs and in 50 volunteers did not show dermalsensitization neither photoirritation nor photoallergy wasobserved in the volunteers No mucosal irritation wasobserved after a volume of 01mL of neat MJ was instilledinto the right eye of 6 white New Zealand rabbits (left eyeserved as control) [50] Finally when topically applied oncancerous and precancerous skin lesions of eight patientswith different pathologies MJ showed promising results 38patients with oral lichen planus exhibited positive responsesand 18 patient with leukoplakia had complete recovery (for18 months following the first treatment) while MJ treatmentof one patient with lentigo maligna of the face resulted in drytumor surface with reduction of the metaplastic area duringtreatment but the cancer reappeared three months later MJdid not cause significant local or systemic side effects [51](Table 2)Thenotable absence of toxicity ofMJ to normal cellsand to healthy human and animals at mM doses indicatesthat MJ may be used safely in cancer treatments at thesedoses [15 19 21 22 38 39 50] Furthermore based in toxicitystudies an exemption from the requirement of a tolerance forMJ was recently published by the US Federal EnvironmentalProtection Agency (EPA) [52] In this document MJ wasconsidered as a ubiquitous and naturally occurring planthormone regarded as a safe and natural part of the humandiet through commonly consumed fruits No toxicologicalendpoints were found for MJ through any route of exposureand stated that MJ is virtually nontoxic to humans andother nontarget organisms through all routes of exposureincluding oral As further support for these arguments it was
International Journal of Cell Biology 3
Table 1 Effect in vitro of natural and synthetic jasmonates on normal and cancer cells
Jasmonate (JAs conc range)vehicle Cell lines Effects References
JA MJ 05ndash30mMEtOH
Normal mononuclear cells fromblood (healthy donors) No cytotoxicity
[15 38]
Acute human T-lymphoblasticleukemia Molt-4 andandrogen-responsive humanprostate adenocarcinoma LNCap
JA 3mM 90 cytotoxicityMJ 05mM 875 cytotoxicityuarrapoptosis (uarrcaspase-3) uarrnecrosisPTPC opening arrest at G0G1
Human melanoma SK-28 JA darrproliferationMJ uarrcytotoxicity uarrcell death
Human breast carcinoma MCF-7 JA darrproliferationMJ uarrcytotoxicity uarrcell death
Murine lymphoma EL-4 MJ uarrcytotoxicity
MJ 30mMno vehicle reported
Acute human T-lymphoblasticleukemia Molt-4
uarrp38 uarrJNK uarrAP-1 howevercell death MAPK-independent [18]
JA CJ andMJ (mM) Prostate PC-3 HTB-81 darrproliferation [46]
MJ 00ndash04mMno vehicle reported
Human myeloidleukemia HL-60 cells
darrcell growth uarrMAPKuarrdifferentiation to mature cells [41]
MDDHJ(synthetic)
0ndash250 120583Mno vehicle reported
Human myeloidleukemia HL-60 cellsMonocytoid leukemiaU937 THP-1 cellsPromyelocytic leukemiaNB4 cellsLung adenocarcinomaPC9 PC14 cells
MDDHJ more potent than MJdarrcell growth uarrMAPKuarrdifferentiation to mature cells
[41]
MJ 05ndash50mM Human lung adenocarcinoma A549 darrproliferation uarrROS uarrapoptosis(uarrBaxBcl-Xs uarrcaspase-3) [36]
JA CJ andMJ 05ndash30mMEtOH
Acute human T-lymphoblasticleukemia Molt-4 cells
Molt-4 uarrmitochondrial membranedepolarization uarrcyt c releaseuarrswelling uarrcell death
[38]
Lymphocytes from CLL patientsCLL cells uarrmitochondrialmembrane depolarizationuarrcytotoxicity
Liver carcinoma Hep 3B cellsHep3B uarrmitochondrial membranedepolarization (PTPC mediated)uarrcyt c release uarrswelling uarrcell death
Human fibroblast 3T3 cells(nontransformed cell line)Normal blood
Nontransformed 3T3 cells nocytotoxicityNormal lymphocytes nocytotoxicity
JA MJ 025ndash30mMEtOH
B-lymphoma clone29M62 (wild type p53)B-lymphoma clone
wt p53 cells uarrapoptosismutant p53 uarrnonapoptotic celldeath [17]
29M610 (mutant p53 resistant totreatment)
MJ uarrsim90 ATP depletion in bothcell types2DG high Glc but not pyruvatedarrATP
CJ MJ 05ndash25mM Nonsmall cell lung cancer linesA549 and H520
darrproliferation cell cycle arrest atG2M phase uarrp38 and ERK12phosphorylation uarrBaxuarrp21uarrcaspase-3
[37]
4 International Journal of Cell Biology
Table 1 Continued
Jasmonate (JAs conc range)vehicle Cell lines Effects References
MJ 05ndash20mM Hormone-refractory prostateadenocarcinomas PC-3 DU-145 darrproliferation uarrapoptosis darr5-LOX [47]
MJ 1ndash26mMEtOHmdash(IC50)
Murine melanoma cellsB16F10 and B16 COLR(overexpressing Pgp MDR)
darrcell motility darrcell growth darrMDR [39]
TBrJA(synthetic)
40 120583MEtOH(MJ doses lower than
in vitro)
Melanoma B16-F10Breast MCF-7Pancreas Mia PCA-2D122 PBL
uarrcitoxicity (TBrJA≫MJ) [39]
MJ 05ndash30mMno vehicle reported
CT-26 (murine colon carcinoma)B16 (murine melanoma)BCL1 (murine B-cell leukemia)Molt-4 (human T-lymphoblasticleukemia cell line)
MJ (but not JA) detached HK1 andHK2 from VDAC1 in isolatedmitochondria from thefour cell linesMJ did not inhibit HKs activitydarrATP uarrcyt c release uarrmitochondriaswelling uarrcell death
[20 30]
MJ 10-20mM Human neuroblastomaBE(2)-C
Arrest at S-phase darrcell growthdarrXIAP mRNA darrsurvivin mRNAuarrapoptosis
[43]
MJ 05ndash30mMDMSOHuman breast cancer MCF-7Human melanocytic MDA-MB-435cells [74]
Arrest at G0G1and S-phase darrmembrane fluidity uarrapoptosisextrinsic (TNFR1 uarrcaspase-8)intrinsic [darrΔ120595
119898uarrcaspase-3 (only
MDA-MB-435)]
[25]
CJ MJ 20mMDMSO Hormone-independent prostatePC-3 DU-145 cells
Cell cycle arrest darrcell growthuarrapoptosis(uarrTNFR1 uarrcaspase-3)
[48]
JA CJ andMJ 10-20mMDMSO
Human neuroblastomaSH-SY5Y
Arrest at G2M phase darrcell growthuarrapoptosis (darrXIAP darrsurvivin)activities MJ gt JA gt CJ
[44]
Human embryonic kidney HEK 293cells Not affected by MJ
MJ 10-20mMDMSO Human neuroblastomaSK-N-SH BE(2)-C
Arrest at G0G1 phase darrcell viabilitydarrmRNA of PCNA uarrapoptosis(darrXIAP darrsurvivin)
[45]
MJ 05ndash30mMEtOH Sarcomas MCA-105 uarrpAkt (correlates with lowersensitivity to cytotoxicity by MJ) [49]
SaOS-2 (resistent to MJ) MJ + 2DG uarrcytotoxicity
MJ 10ndash50mMEtOH
Cervical cancer SiHA CaSki andHeLa cells having wt p53Cervical cancer C33A cells (withmutated p53)
darrcell cycle uarrapoptosis throughdifferent pathwaysdarrATP uarrlactate (in more glycolyticCaSki) PARP cleavage multiple celldeath pathways depending on levelsof p53 p21 Bcl-2 and Bax
[26]
JA MJ 025ndash40mMEtOH Acute myelogenous leukemia cellsHL-60 and KG1
uarrROS uarrMJ-induced mitochondrialmembrane depolarizationuarrMJ-induced Mit SOD darrAKRC1
[132]
MJ MDDHJ 015mMDMSO Leukemia HL-60 cellsuarrCa2+-binding protein S100Puarrdifferentiation uarrregulator ofG-protein signaling-16 (RGS16)
[42]
J7 (synthetic) IC50 15 120583MHuman cervical carcinomaHeLa cells
Cell cycle arrest at G2M phasedarrBcl-2 (caspase 9 3) DNA damage [27]
International Journal of Cell Biology 5
Table 1 Continued
Jasmonate (JAs conc range)vehicle Cell lines Effects References
J7 (synthetic) 50120583MDMSO Human hepatoma Hep3B
uarrBaxBcl-2 ratio uarrDR5 uarrcaspase-8darrBid uarrapoptosis correlated withuarrcaspase-9 uarrcaspase-3 darrXIAP darrcIAPdarrPARP ExtrinsicintrinsicMAPK
[34]
MJ 05ndash25mMno vehicle reported
CD138+ tumorcells from MM patients
HK2 release from mitochondriarapid darrATP uarrapoptosis [105]
MJ 025ndash10mMno vehicle reported Human colorectal cancer cells CRC
uarrTRAIL uarrcyt c releaseuarrcaspase cleavage darrsurvivindarrTCF transcriptional activity
[32]
MJ 00ndash20mMEtOH Cervical cancer cellsSiHa CaSki HeLa and C33A
uarrmitochondrial O2
minus (HeLa CaSki)darrsurvivin darrE6 E7 (HPV)uarrdifferent cell death pathways(independently of HPV)
[28]
J7 (synthetic) 0ndash50 120583M DMSO Human hepatoma HepG2uarrROS uarrTRAIL-mediated apoptosis(darrBid darrXIAP darrcIAP darrBcl-xLuarrcaspases)
[35]
MJ 30mM (IC50) novehicle reported
Human adenocarcinoma colonHT-39
Arrest at S-G2M uarrcytotoxicityuarrapoptosis [31]
JA MJ 10ndash30mMDMSO Canine macrophagic malignantDM62 cells darrcell growth (MJ gt JA) uarrcytotoxicity [256]
MJ 05ndash02mMDMSO Human gastric SGC-7901 MKN-45cell lines
darrmigration darrinvasion darrangiogenesisdarrMMP-14 [33]
Bcl-2 B-cell lymphoma-2 Bcl-xL B-cell lymphoma-extra large Bid BH3 interacting domain death agonist cIAP cellular inhibitor of apoptosis CJ cisjasmonicacid 2DG 2-deoxy-D-glucose Glc glucose J7 methyl 5-chloro-45-didehydrojasmonate JA jasmonic acid NSCLC nonsmall-cell lung carcinoma PgpP-glycoprotein MDDHJ methyl 45-didehydro-jasmonate MDR multidrug resistance MMP-14 matrix metalloprotease 14 PARP poly (ADP-ribose)polymerase PCNA proliferating cell nuclear antigen PTPC permeability transition pore complex ROS reactive oxygen species S100P protein SP100 TBrJA57910-tetrabromo jasmonate TNFR1 tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand XIAP X-linked inhibitor of apoptosis protein
Table 2 Effect in vivo of natural and synthetic jasmonates derivatives
Jasmonates Conc rangevehicle Organismtissuecells Effects ReferencesMJ (oral) 236mgkglipofundin Normal C57BL6 mice Nontoxic [15]
MJ (oral) 236mgkglipofundin C57BL6 mice injected ip withmurine T-lymphoma EL-4
uarrsurvival of treated mice ascompared to the inoculated control [15]
TBrJA40120583MEtOH
(lower doses than MJin vitro)
Murine melanomaB16-F10 darrlung metastasis [39]
MJ 1ndash10120583MEtOH Chicken CAM darrangiogenesis [40]MJ 1ndash10120583MEtOH Human endothelial cells (HUVEC) darrCOX-2PGE2 pathway [40]
MJ (ip) 1 gKg in 01mLvegetal oil ip
Multiple myeloma- (MM1S-)inoculatedin nodscid mice
uarrsurvival of treated mice ascompared to the inoculated control [105]
MJ (topical) 1 gmL oil Application on cancerous andprecancerous human skin lesions
No local or systemic side effects38 patients withpositive responses
[51]
CAMchorioallantoic membrane of chicken embryoHUVEC human umbilical vein and endothelial cells TBrJA synthetic 5 7 9 10-tetrabromo jasmonate
mentioned that MJ was also assessed and approved by Foodand Agriculture OrganizationWorld Health Organization(FAOWHO) as a food additive Toxicity results reportedin the EPA document [52] were found to agree with thosepreviously published by other groups [50 51]
21 Jasmonates (MJ) Can Cause Cancer Cell Cycle Arrest atDifferent Phases in Different Cancer Cell Types Inhibiting
Growth and Proliferation The eucaryotic cell cycle is dividedinto four phases G
1 S G
2 and M that occur in response
to growth factors or mitogens The DNA synthetic (S) andmitotic (M) phases are preceded by gap phases (G
1 G2)
Chromosome duplication occurs during S phase replicatedchromosomes are segregated into individual nuclei (mitosis)during M phase and the cell then splits in two Cell cycleprogression can be regulated atG
1 G2points by various intra-
cellular and extracellular signals If extracellular conditions
6 International Journal of Cell Biology
Table 3 Effect in vitro and in vivo of methyl jasmonate (MJ) combined with other anticancer agents
Jasmonate Drug concrangevehicle Model Effects References
(1) MJ + BCNU(nitrosourea)in vitro
MJ Fixed conc Pancreatic MIA PaCa-1 Mitochondriotoxicsynergic cytotoxicity
[170](2) MJ + 2DGadriamycin taxolBCNU or cisplatinin vitro
MJ05ndash20mMEtOHTaxol cisplatin1ndash10120583gmL
CT26 DA-3 GTRAMP C1 MCF7MIA PaCa-2 D122 and BCL1
Strong cooperativeeffects of MJ + 2DGand MJ + other drugs
(3) MJ + adriamycinin vivo
MJ iv20ndash150mgkgdissolved inlipofundinAdr (DOX) ip4mgkg
Balbc mice injected ip with 1 times 104chronic BCL1 cells lymphocyteleukemia cells
MJ + Adr significant prolongedsurvival effect
(4) MJ + PI3KAktinhibitorsin vitro
MJ 05ndash30mMEtOHDMSO
Sarcoma MCA-105 SaOS-2 darrMJ-induced activation of Aktuarrsynergic cytotoxicity
[49](5) MJ + 2DGin vitro
MJ 05ndash30mMEtOHDMSO Sarcoma MCA-105 SaOS-2 2DG darrpAkt darrMJ-ind pAkt
MJ + 2DG uarrsynergic cytotoxicity
(6) MJ + irradiationin vitro
05ndash20mMDMSO
Irradiated prostate PC-3 (radiationinduces Bcl-2 expression)
darrradiation-induced Bcl-2uarrradiation sensitivity PC-3uarrcaspase-3
[104]
(7) MJ andor POHandorcis-platin (CP)in vitro
Both testedat IC20
Human MDA-MB-435[74]
MJ + POH uarrTNFR1darrΔ120595119898uarrcytotoxicity cell cycle arrest
at G0G1 uarrapoptosis+ CP all effects enhanced
[251]
Human breastMDA-MB-231 uarrapoptosis
(8) MJ + TRAILin vitro
MJ 05mM +TRAIL(100ndash200 ngsdotmLminus1)
CRC cancer cells
darrsurvivin (IAP)darrWntTCF pathwayuarrTRAIL-induced apoptosisuarrcaspase activity
[32]
(9) MJ + Smacin vitro
MJ 05ndash20mMDMSO
Human bladder cancer EJ T24 Synergy uarrIAPs-bound caspase 3uarrapoptosis [254]
Human embryonic kidney HEK293 No cytotoxicity
(10) MJ + Smac7N(IAP antagonist)in vitro
MJ 05ndash20mMDMSO
Hormone-independent prostateDU-145 PC-3Human proximal tubular epithelialHK-2 cells (overexpressing IAPs)
Smac7N uarrMJ-induced apoptosis bycaspase-9-dependent (intrinsic) andindependent (extrinsic) pathways
[255]
(11) MJ + cisplatin(01ndash05 120583M)MJ + X-rays(025ndash3Gy)MJ + 120572-rays
MJ 01ndash10mMEtOH
Cervical cancer cellsSiHa CaSki HeLa and C33A
darr viabilitydarrsurvivaldarrIC50 radiation dose
[29]
(12) MJ + 5-FUin vitro
05mMMJ1 hthen + 5-FU
Human adenocarcinomacolon HT-39 darrIC50 5-FU (5 rarr 25mM) [31]
BCNU 13-bis-(2-chloroethyl)-1-nitrosourea 2DG 2-deoxy-D-glucose 5-FU 5-fluorouracile IAP inhibitors of apoptosis MM multiple myeloma POHperillyl alcohol smac second mitochondria-derived activator of caspases Smac7N a peptide that contains the N-terminal seven residues of smac TNFR1tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand
are unfavorable cells delay progress through G1and may
enter a quiescent state known asG0 in which they can remain
for days weeks or years before resuming proliferation Ifextracellular conditions are favorable and signals to growand divide are present cells in early G
1or G0progress
through a restriction checkpoint late in G1after which cells
are committed to DNA replication even if the extracellularsignals that stimulate cell growth and division are removed[53 54] Cell cycle activity is modulated by intracellularROS levels and regulates cell survival cell proliferation andapoptosis [53ndash58]The progression through theG
1 S G2 and
M phases is promoted by cyclin-dependent kinases (CDKs)
International Journal of Cell Biology 7
which are positively and negatively regulated by cyclins andcyclin kinase inhibitors (CKis) respectively and by ROSUnlike the passage through the S G
2 and M phases G
1
progression normally relies on stimulation by mitogens andcan be blocked by antiproliferative factors passage throughthe restriction checkpoint and entry into S phase is controlledby CDKs that are sequentially regulated by cyclins D Eand A [53 54 56ndash58] In general CDK activity requirescyclin binding and depends on both positive and negativeregulatory phosphorylations [54] ROS levels influence thephosphorylating and ubiquitinating enzyme activites thuscontrolling cell cycle progression [54 57 58]
A master gene regulating cell passage through the cycleis the Myc family of transcription factors it switches onthe simultaneous synthesis of thousands of different proteinsrequired for cell growth and cell cycle function [58 59] TocounteractMyc protein activity the cell cycle is also regulatedby the p53 protein [60 61] a DNA-binding transcriptionfactor that senses DNA damage (occurring after exposureto ionizing radiation UV light oxidative stress or otherDNA damaging agents) and activates genes to halt cell cycleprogression in G
1for repair or otherwise initiates apoptosis
[56 62] When the damage caused by cell stress is mild p53activatesmechanisms for cell cycle arrest and repair damagedDNA if the damage is irreparable p53 initiates apoptosis [63]In case ofmild cellular stress low levels of p53 can activate theTP53-induced glycolysis and apoptosis regulator (TIGAR)gene that decreases the rate of glycolysis and hence ROS levelsin the cell [64] Glycolysis is essential for cancer progressionbut TIGAR provides protection from ROS and apoptosisHowever if damage caused by stress is irreparable p53prevents proliferation of cells with altered DNA and inducesincreased levels of ROS to trigger cell death [56] Increasedlevels of active p53 inhibit the cell entry into the S-phaseor induce apoptosis [56] Loss or inactivation of p53 resultsin loss of cell-cycle arrest or inhibition of apoptosis afterDNAdamage or physiological stress [65] leading to increasedgenetic instability increased accumulation of mutations andultimately oncogenesis [61] When the cell cycle is no longerregulated by p53 cells remain cycling (supported by 119888-Mycoverexpression) subverting cell cycle exit maturation andterminal differentiation [66] Over 50 of all human cancersharbor mutations and genetic alterations in cyclins and p53genes directly affecting the function of critical cell cycleproteins rendering the p53 gene ineffective in mediating cellgrowth arrest and promoting cancer cell death [62 67 68]ROS production is under p53 regulation and in turn p53being a redox-sensitive protein is influenced by ROS levelsthus ROS can act as both an upstream signal that triggersp53 activation and as a downstream factor that mediatesapoptosis [69]Thus mutations in the p53 gene downregulateits activity and consequently ROS production [70]ThroughROS p53 can also directly control metabolic traits of cells[71] regulate mitochondrial membrane potential (Δ120595
119898) and
induce cytochrome 119888-independent apoptosis (blocked by Bcl-2) ROS-mediated disruption of Δ120595
119898constitutes a pivotal
step in the apoptotic pathway of p53 and this pathwaydoes not involve cytochrome 119888 release [72] ROS can thusregulate cell fate through p53 in a way that physiological ROS
levels trigger cell protective pathways while under cytotoxicoxidative stress p53 behaves more like a cell killer [70 73]
JAs (mainly MJ) have been found to arrest cell cycle ofdifferent cancer cell types at different phases (Table 1) G
0G1
in human acute lymphoblastic leukemia Molt-4 [14 38] andhuman neuroblastoma SK-N-SH BE(2)-C [45] G
0G1-S in
human breast MCF-7 andmelanocytic MDA-MB-435 cancercells [25 74] S in human neuroblastoma BE(2)-C [43] S-G2M in human adenocarcinoma colon HT-39 [31] G
2M
in nonsmall cell lung cancer (NSCLC) lines A549 and H520[37] human neuroblastoma SH-SY5Y [44] human cervicalcarcinoma and HeLa cells [27] At 01mM MJ arrested invitro human umbilical vein endothelial cells (HUVEC) at G
1
[40]The molecular mechanisms underlying the above jasmo-
nate-mediated cell cycle arrest in cancer and embryonicHUVEC cells in vitro are not yet completely understoodChecking the above data against the UMD TP53 mutationdatabase (httpwwwp53fr) and revising the article byBerglind et al [75] we could not find correlation between agiven MJ-mediated phase arrest of cell cycle and the TP53wild type or mutated status of a cancer cell type (data notshown) a result that would agree with the original observa-tion of Fingrut and Flescher [15] stating thatMJ-mediated celldeath is independent of the p53 status [17] However otherstudies suggest thatMJ could be acting by interfering differentcell cycle regulatorymechanisms In plants MJ is able to bindthe TGACCG motif which is present in cell cycle cyclin pro-moters [76] wemight speculateMJ recognition of TGACCG-like motifs in mammalian cells cyclin promoters howeverthis would need experimental confirmation In some plantsMJ impairs the G
2-to-M transition by repressing M-phase
gene activation [77] MJ impaired the G2-to-M transition in
the human neuroblastoma SH-SY5Y cells downregulatingthe expressions of the proliferating cell nuclear antigen(PCNA a processivity factor encircling DNA at sites ofreplication and repair) andN-Myc [45] Asmentioned aboveMYC is the master gene allowing cancer cells to move alongthe cell cycle and regulating most genes needed to meet themetabolic demands for cell proliferation and survival [78]Cancer cell cycle arrest at different phases is in itself closelydependent on the cell energy and as such it is extremelydependent on metabolic ATP availability and its use byhexokinase Downregulation of Myc byMJ [45] is interestingresult that must be confirmed particularly because there is alack of specific inhibitors against Myc [57]
In some cases of cell cycle arrest MJs have been foundto upregulate p21 [37] whereas in others to downregulatesurvivinmRNA and protein levels [43ndash45] (Table 1) Survivinis an important member of the inhibitor of apoptosis protein(IAP) family that also regulates entrance of the cell cycle to theM phase and is overexpressed in cancer cells acting linkingboth regulation of the cell cycle and inhibition of apoptosis[79]
22 Jasmonates (MJ) CanCauseCancer Cell Death by InducingApoptosis andor Necrosis Apoptosis can be initiated by twosignaling cascades namely the extrinsic and the intrinsic
8 International Journal of Cell Biology
pathway The classical extrinsic pathway is triggered by thebinding of extracellular signals like tumor necrosis factor(TNF) tumor necrosis factor-related apoptosis-inducing lig-and (TRAIL) and apoptosis antigen-1 (Fas) ligand (FASL)to death receptors such as tumor necrosis factor receptor 1(TNFR1) and FAS [80]This interaction leads to procaspase-8activation which cleaves itself into caspase-8Then caspase-8can cleave and activate procaspase-3 into caspase-3 thatcleaves many substrates inside the cell inducing the typicalmorphological changes seen in apoptosis The intrinsic path-way can be triggered by intracellular stress signals such asoxidative stress and DNA damage The diverse signals aresensed by mitochondria through the balance between pro-and antiapoptotic members of the Bcl-2 family of proteinsWhen proapoptotic signals overcome antiapoptotic signalsmitochondria becomepermeabilized leading to dissipation ofthe mitochondrial electrochemical potential release of inter-membrane proteins like cytochrome 119888 SmacDIABLO AIFandHTRA2OMI Cytochrome 119888 interacts with procaspase-9and Apaf-1 to form the apoptosome complex that leads tocaspase-3 activation [81] The extrinsic pathway can connectwith the intrinsic pathway through the cleavage by caspase-8of Bid into tBid (truncated Bid) which is a proapoptoticmember of the Bcl-2 family and can lead to mitochondriapermeabilization [82 83] The members of the Bcl-2 fam-ily regulate mitochondria permeabilization and comprisesantiapoptotic (Bcl-2 Bcl-xL) proapoptotic (Bax Bak) andproapoptotic BH-3 only proteins (Bad Bid Puma andNoxa)[82ndash84] One of the mechanisms of mitochondria perme-abilization is controlled by Bax and Bak proteins that arenormally expressed in every cell but are maintained inhibitedby the antiapoptotic members When there is a signal toapoptosis BH3-only proteins interact with the antiapoptoticmembers releasing BaxBak or directly activating themleading to their oligomerization and to mitochondrial outermembrane permeabilization (MOMP) Another mechanismfor mitochondria permeabilization is the formation of themitochondrial permeability transition (MPT) pore which iscomposed by the voltage-dependent anion channel VDACthe adenine nucleotide translocase (ANT) and cyclophilin D[85]TheMPT pore comprising VDAC1 is directly regulatedby Bad [86] In contrast VDAC2 has been identified as anantiapoptotic negative regulator of Bak [87ndash89]
One of the hallmarks of cancer is resistance to cell death[90] This resistance is acquired by a host of different geneticdefects in cancer cells such as those present in the p53tumor suppressor gene [62 68 91] Normal p53 responds togenotoxic oncogenic and other stress signals by inducingantiproliferative transcriptional programs leading to growtharrest (for cell repair) or apoptosis [92] p53 mediates cyclearrest by transcriptionally activating the p21 (CDKN1A) 14-3-3120590 (SFN) and GADD45120572 genes whereas p53-dependentapoptosis is triggered by turning on the synthesis of proteinsthat produce ROS [58 93] and transactivating differentmitochondrial proapoptotic genes as well as death receptorsfor the synthesis of the corresponding proteins [58 94]Whenp53 (and its target genes) expression levels are below a thresh-old they induce cell cycle arrest but not apoptosis above thisthreshold p53 trigger apoptosis [95] Lowering this threshold
with inhibitors of the antiapoptotic Bcl-2 family proteinssensitize cells to p53-induced apoptosis [95] By damagingcells chemotherapeutical drugs turn on p53rsquos transcriptionalfunction that upregulates the expression of proapoptotic pro-teins Defects in the p53 function or in the apoptotic cascadeslead to cell phenotypes resistant to chemotherapy Apoptosiscan also be induced by anticancer agents independently ofp53 by either directly modulating the apoptotic machineryor directly interacting with mitochondria [96] ROS levelsmight be playing a central role in mediating various forms ofnonapoptotic caspase-independent programmed cell deatheither through direct irreversible oxidative damage to keyproteins nucleic acid molecules and cellular structure or bythe activation of prodeath signal transduction programs [97]In a ROS-dependent nonapoptotic cell death pathway respi-ration through the mitochondrial electron transport system(ETS) chain is not essential and ROS is generated throughNADPH oxidases [97] In contrast to the mitochondrial ETSwhich generates ROS as a byproduct of respiration NADPHoxidases produceROS as their primary function [98] Raise ofROS is characteristic after ionizing radiation chemotherapyand targeted therapy treatments [58 99] and may be killingcancer cells through a nonapoptotic ROS-dependent celldeath pathway
Tumor cells maintain high ATPADP as well asNADHNAD+ ratios so as to assure that ATP levels arenever limited ATP depletion would represent then anunsustainable metabolic stress for tumor cells and it isassociated with necrotic death in some cell types [100]Recently it was shown that a signaling cascade centered onthe proteins RIP1 and RIP3 can trigger cell death by necrosisin many situations and the RIP1-RIP3 complex appears topromote ATP depletion during necroptosis by impingingon components of the MPT pore [101 102] Necrosis was aterm used to describe a process of accidental cell death inwhich cells swell and burst releasing its intracellular contentwhereas necroptosis modernly refers rather to an orderedcell death program different from apoptosis apoptosis is acaspase-dependent cell death process whereas necrosis iscaspase-independent process [102 103] Studies concerningnecrosis have focusedmainly on cell membrane permeabilityto propidium iodide (PI) in the flow citometry doublestaining with annexin-VPI
Fingrut et al [17] by studying the effect of MJ ontwo clones of a highly malignant B-cell lymphoma (oneharboring a normal wild type (wt) p53 gene while the otherexpressed a mutated inactive p53 protein) (Table 1) reportedthat MJ was equally toxic to both clones [17] In contrastthe p53-mutant cells were resistant to radiomimetics (drugshaving effects similar to those produced in radiotherapy)and chemotherapeutic drugs In this cancer cell model MJinduced mostly apoptotic cell death in the wt p53-expressinglymphoma cells while no signs of early apoptosis weredetected in the mutant p53-expressing lymphoma cells thatwere killed by MJ through a p53-independent nonapoptoticcell death mechanism However other cancer cell types canundergo apoptosis with or without functional p53 and aswe will see MJ can induce apoptotic andor nonapoptoticcell death in wt p53 as well as in TP53 mutated cells
International Journal of Cell Biology 9
(Table 1) Nonapoptotic cell death could be the consequenceof the severe bioenergetic damages induced by the directinteraction of MJ with mitochondria leading to increasedROS levels inhibition of ATP synthesis and ATP depletion[20 30] Jasmonates including MJ and some of its analogshave been found to be strongly proapoptotic in vitro indifferent types of cancer cells by both the extrinsic and theintrinsic pathway (Table 1) with no or little cytotoxicity tonormal cells [15 25 36 37 44 45] Thus MJ induced theexpression of TNFR1 in human breast MCF-7 and MDA-MB-435 cells [25 74] and in hormone-independent humanprostate PC-3 and DU 145 cancer cells promoting apoptosisby the extrinsic pathway in these cells [48] TRAIL (TNF-related apoptosis inducing ligand) is able to induce apoptosisin prostate cancer cell yet overexpression of antiapoptoticproteins and inhibition of proapoptotic proteins results indiminishing the TRAIL-mediated apoptosis pathway TRAILhas emerged as a potent anticancer agent in laboratory-based studies and preclinical trials and JAs have been foundto sensitize cancer cells to TRAIL-mediated apoptosis [32](Table 3) J7 a synthetic analogue of MJ enhanced TRAIL-mediated apoptosis through upregulation of ROS levels inhuman hepatomaHepG2 cells apoptosis was induced via Bidcleavage downregulation of X-linked inhibitor of apoptosisprotein (XIAP) cellular inhibitor of apoptosis-1 (cIAP-1)B-cell lymphoma-extra large (Bcl-xL) and activation ofcaspases [35] Other examples of MJ-mediated apoptoticandor nonapoptotic cell death in different cancer cell types(Table 1) are as follows (1) MJ induced ROS production inA549 lung adenocarcinoma cells activating Bcl-2-associated-X-protein (Bax) as well as Bcl-2-Like-1 (Bcl2L1) proapoptoticproteins in these cells [36] (2) MJ activated caspase-3 inleukemia Molt-4 prostate SK28 and LNCaP cancer cells[15] (3) MJ inhibited the antiapoptotic Bcl-2 protein andenhanced proapoptotic proteins in radio-resistant humanprostate cancer cell line PC-3 [104] (4) MJ suppressed theproliferation of the neuroblastoma cell lines SH-SY5Y SK-N-SH and BE(2)-C through a strong apoptosis-inducingeffect as measured by annexin VFITCPI MJ treatment alsodownregulated the expression of the antiapoptotic proteinsXIAP and survivin [44 45] (5) MJ-mediated cell death wasobserved on cervical cancer and myeloma cells and it wasreported to be due to mixed characteristics of apoptosis andnecrosis [26 105] So far studies showing MJ inducing theRIP1 signaling cascade have not been reported howeverit will be important to deep into these analyses to cleardistinguish between MJ-induced apoptosis andor necrosis
23 Methyl Jasmonate Induces the Production of ReactiveOxygen Species (ROS) Responses in Some Cancer Cell TypesLeading Also to Cell Death Intracellular ROS levels reg-ulate cell cycle activity cell metabolism and survival cellproliferation and apoptosis [53ndash58] Stressful conditionsproduce a transient excess of intracellular ROS in normaltissues however under chronic pathological and oxidativestress conditions ROS is continuously produced in excessand persistently causing damage to DNA cell membraneslipids proteins and inducing apoptosis to control and avoid
oxidative damages cells counteract excess ROS by enzymatic(superoxide dismutase (SOD) glutathione peroxidase (GPx)and catalase) andor nonenzymatic (glutathione thoredoxin)antioxidant elements [99 106] Controlling ROS is central tointracellular redox signaling and homeostasis on the otherhand uncontrolledunbalanced ROS production modifiesintracellular redox leading to apoptosis [58 82 99 106] ROScan be generated by different biological systems (1) director indirect extracellular stimulation of plasma membrane-bound NAD(P)H oxidases (NOX) can generate intracellularH2O2[57 107ndash110] (2) biochemical pathways associated to
fatty acid oxidation and lipoxygenases activities generateROS as normal byproducts of these activities [111] (3)ATP generation by oxidative phosphorylation (OxPhos) inmitochondria is accompanied by the production of ROS as aconsequence of electron leakage from the electron transportchain mitochondria are considered to be major sources ofROS inmammalian cells particularly under a variety of stress-ful conditions [82 99] Superoxide (O
2
minus) is mainly produced
from complexes I and III and it is rapidly dismutated to H2O2
and oxygen by SOD [112] H2O2serves both as a toxic oxidant
and as an essential signaling molecule regulating cellularbiological processes [113] A strong positive correlation existsbetween mitochondrial membrane potential (Δ120595
119898) and ROS
production [106 114 115] Mitochondria produce more ROSat high membrane potential and an increase in the Δ120595
119898
produced either by a closure of MPT pore or by inhibitionof the ATP synthase is associated with increased ROS pro-duction However in cases of mitochondrial dysfunctionslow Δ120595
119898and decreased activity of the respiratory chain
are accompanied by a simultaneous increase in ROS [116117] It has been reported that cancer cells have higher ROScontent compared to normal cells [118 119] probably due toabnormal respiration because of dysfunctional mutations inmitochondria [120] but they also have increased contents ofantioxidants (SOD catalase glutathione thioredoxin etc)to counteract ROS [58 121] Tumor cells cannot tolerateexcessive high ROS levels [118 122] and as a matter of factradiation therapy chemotherapies and targeted therapiesinduce apoptosis and kill some cancer cells by generatingexcess ROS [99 106 123 124] Thus inducing higher levelsof ROS in cancers cells can be exploited as an effectiveselective strategy to kill tumor cells over normal cells [119]However prolonged treatment with the same drug reducesROS levels in cancer cells [124] indicating that when theybecome drug-resistant tumor cells have lower ROS contentthan drug-sensitive cancer cells Evidence suggests that drug-resistant cells have a higher expression of catalase at theplasma membrane that keeps reduced ROS levels [121]
In plants MJ-mediated damage to mitochondria is fol-lowed by a rapid production of H
2O2[125] H
2O2is used as
a second messenger for the induction of genes and productsrelated to defense against both herbivores and pathogens[126] in addition H
2O2induces apoptosis and senescence
[127 128] On the other hand it has been signaled that theMJ-induced production of H
2O2in plants was prevented by
inhibitors of NAD(P)H oxidase [128 129] indicating anothermechanism of MJ-mediated production of ROS and a majorrole for this enzyme in MJ-induced ROS responses MJ
10 International Journal of Cell Biology
induces also an endogenous H2O2response in some cancer
cell lines (Table 1) [19 36 38 130ndash132] In A549 cancerlung cells the H
2O2response induced by MJ increased the
expression of the proapoptotic Bax and Bcl-Xs (although notbeing that of the antiapoptotic Bcl-2 and Bcl-xL proteins) andthis led to apoptosis this event was inhibited by catalase (aspecific inhibitor of H
2O2) [36] MJ increased the intracel-
lular levels of H2O2and superoxide (O
2) ions in human C6
glioma cells in vitro and induced the expression of heat shockprotein 72 (HSP72) a negative regulator of H
2O2-induced
oxidative stress in these cells in vitro MJ-induced HSP72expression was blocked by ROS inhibitors [130] ElevatedROS generation was observed in response to both MJ andJA levels in the acute myeloid leukemia HL-60 and KG1a celllines but other factors like the MJ-mediated generation ofmitochondrial SOD rather than ROS were determinant forapoptosis or differentiation in these cells [132] Suppression ofMJ-induced apoptosis by antioxidants like N-acetylcysteine(NAC) and catalase but not by inhibitors of hydroxyl radicalsand superoxide ions indicates that H
2O2is nonetheless one
of the important factors involved inMJ anticancer effects [19]
24 By Dissociating Hexokinase (HK) from the VoltageDependent Anion Channel (VDAC) on the Outer Mitochon-drial Membrane MJ Dissociates Glycolysis from OxidativePhosphorylation Causing Severe Bioenergetic Deregulations inCancer Cells Metabolism and cell growth are two cellularprocesses that are tighly linked and regulated [133] Theclassic bioenergetics phenotype of cancer cells of enhancedglycolysis under aerobic conditions was described by War-burg [134] However the ldquoWarburg effectrdquo does not neces-sarily imply mitochondrial dysfunction [135] To increaseglycolysis cancer cells upregulate the transcription of genesinvolved in the glycolytic pathway (ie glucose transportersglycolytic enzymes etc) This metabolic reprogrammingallows increased rate of ATP production synthesis of lipidsand a new redox balance Cancer cells actually use a com-bination of both glycolysis and mitochondrial respirationto produce energy and they may vary in regards to thepreferential use of these pathways being in some caseseither more glycolyticless oxidative or less glycolyticmoreoxidative depending on the prevalent normoxic or hypoxicenvironmental conditions and their capacities to expressadequate levels of oncogenes and tumor suppressor geneproducts for cell growth [136ndash139] or when either glycolysisor OxPhos is inhibited in whose case there can be partialcompensation by the other metabolic pathway [17] Shiftingthe balance between those two processes with glycolyticandor mitochondrial enzyme inhibitors might represent aninteresting way to look for new anticancer agents
The first step in the metabolism of glucose is catalyzedby the enzyme hexokinase (HK) which by phosphorylatingglucose makes it negative sequestering it into the cytoplasmin the form of glucose-6-phosphate (G6P) In normal tissuesthis crucial step is catalyzed by four different HK isoforms(HK1 HK2 HK3 and glucokinase) indicating that regulationof glucose phosphorylation can vary in different tissues underdifferent condition [140] HK1 and HK2 have overlapping
tissue expression but different subcellular distributions withHK1 associated mainly with mitochondria and HK2 associ-atedwith bothmitochondrial and cytoplasmic compartments[141] HK1 is widely and constitutively expressed whereasHK2 predominates during embryogenesis and is not widelyexpressed in adult tissues but only in a limited numberof normal adult tissues [141] their different subcellulardistributions and kinetic properties reflect their differentmetabolic roles [141 142] Under physiological conditionsHK1 is predominantly bound to the outer mitochondrialmembrane where it primarily channels glucose toward gly-colysis [140] whereas HK2 is mainly soluble and controlsglycogen formation [140 141] In malignant tissues HK1and HK2 isoforms are glycolytic and overexpressed (HK1more than HK2) (httpwwwproteinatlascom) [143] andare tightly bound to mitochondria HK1 and HK2 bindthrough theirN-terminal hydrophobic regions to the voltage-dependent calcium channel isoform1 (VDAC1) the prevalentpore protein in the outer mitochondrial membrane [2482 143ndash146] VDAC1 is also overexpressed in cancer cells[147] and the binding of HK to VDAC1 on the outer mito-chondrial membrane is a fundamental aspect of the aerobicglycolytic metabolism of cancer cells [82 135 147 148]HKVDAC association appears to protect tumor cells frommitochondrial outer membrane permeabilization (MOMP)which marks a point of no return leading to cell death[149] VDAC transports ADP and inorganic phosphate (Pi)the substrates needed for the production of ATP into themitochondria it also transports ATP out of themitochondriainto the cytoplasm and facilitates the availability of ATPto HK2 [143 150 151] A continuous supply of glucose toHK maintains this activity at a constant level and controlsthe flux through the mitochondrial electron transport chaininfluencing mitochondrial ROS production [152] HK bindsthrough hydrophobic interaction to VDAC1 the prevalentpore protein in the outer mitochondrial membrane [24 82143ndash146] thereby assuring and gaining a preferential accessto ATP [82 150 153] Translocation of HK2 from the cytosolto the mitochondria is regulated through phosphorylationof HK by Akt [141] HK2 only binds VDAC after phos-phorylation by Akt [151 152] Glycogen synthase kinase 3beta (GSK3120573) a major target of Akt kinases phosphorylatesVDAC [154ndash157] and dissociates HK [156] HK2 boundto mitochondria enhance its affinity to ATP and is lesssensitive to enzymatic inhibition by the G6P product [140141 158] HK2 can produce G6P from cytoplasmic glucoseand mitochondrial ATP without regulatory restraints evenunder hypoxia supporting rapid cell growth [142 143 159ndash161]HK1 andHK2 aremore tightly bound toVDAC in cancercells than in nonmalignant cells [156 162 163] HKVDA1interaction is relatively stable for HK1 while it is dependentupon G6P levels for HK2 [140 141] The higher affinity ofmitochondrial HK2 for glucose ATP and VDAC1 representsa high advantage for cancer cells survival and growth con-ferring to them an aerobic glycolytic phenotype underlyingthe Warburg effect [82 135 138 142 147 148 150 160]Differential interactions of HK1 and HK2 with mitochondriamay underlie different glycolytic profiles in cancer cells [164165] As rate-controlling enzymes in the glycolyic pathway
International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
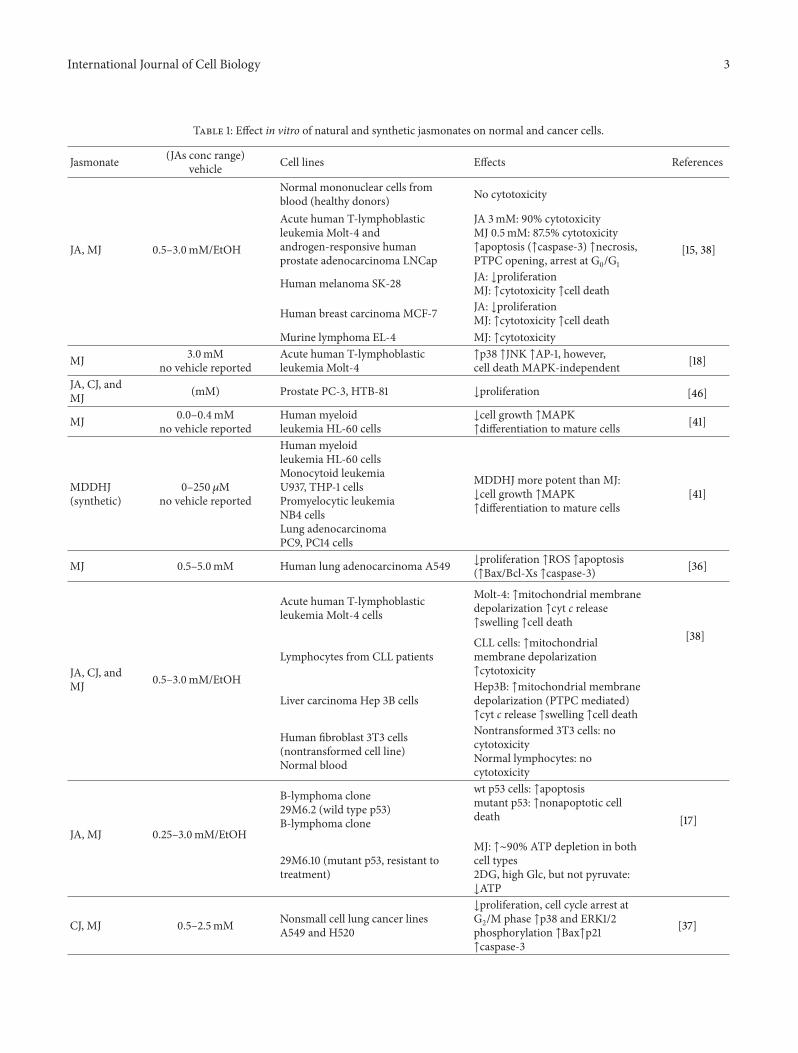
International Journal of Cell Biology 3
Table 1 Effect in vitro of natural and synthetic jasmonates on normal and cancer cells
Jasmonate (JAs conc range)vehicle Cell lines Effects References
JA MJ 05ndash30mMEtOH
Normal mononuclear cells fromblood (healthy donors) No cytotoxicity
[15 38]
Acute human T-lymphoblasticleukemia Molt-4 andandrogen-responsive humanprostate adenocarcinoma LNCap
JA 3mM 90 cytotoxicityMJ 05mM 875 cytotoxicityuarrapoptosis (uarrcaspase-3) uarrnecrosisPTPC opening arrest at G0G1
Human melanoma SK-28 JA darrproliferationMJ uarrcytotoxicity uarrcell death
Human breast carcinoma MCF-7 JA darrproliferationMJ uarrcytotoxicity uarrcell death
Murine lymphoma EL-4 MJ uarrcytotoxicity
MJ 30mMno vehicle reported
Acute human T-lymphoblasticleukemia Molt-4
uarrp38 uarrJNK uarrAP-1 howevercell death MAPK-independent [18]
JA CJ andMJ (mM) Prostate PC-3 HTB-81 darrproliferation [46]
MJ 00ndash04mMno vehicle reported
Human myeloidleukemia HL-60 cells
darrcell growth uarrMAPKuarrdifferentiation to mature cells [41]
MDDHJ(synthetic)
0ndash250 120583Mno vehicle reported
Human myeloidleukemia HL-60 cellsMonocytoid leukemiaU937 THP-1 cellsPromyelocytic leukemiaNB4 cellsLung adenocarcinomaPC9 PC14 cells
MDDHJ more potent than MJdarrcell growth uarrMAPKuarrdifferentiation to mature cells
[41]
MJ 05ndash50mM Human lung adenocarcinoma A549 darrproliferation uarrROS uarrapoptosis(uarrBaxBcl-Xs uarrcaspase-3) [36]
JA CJ andMJ 05ndash30mMEtOH
Acute human T-lymphoblasticleukemia Molt-4 cells
Molt-4 uarrmitochondrial membranedepolarization uarrcyt c releaseuarrswelling uarrcell death
[38]
Lymphocytes from CLL patientsCLL cells uarrmitochondrialmembrane depolarizationuarrcytotoxicity
Liver carcinoma Hep 3B cellsHep3B uarrmitochondrial membranedepolarization (PTPC mediated)uarrcyt c release uarrswelling uarrcell death
Human fibroblast 3T3 cells(nontransformed cell line)Normal blood
Nontransformed 3T3 cells nocytotoxicityNormal lymphocytes nocytotoxicity
JA MJ 025ndash30mMEtOH
B-lymphoma clone29M62 (wild type p53)B-lymphoma clone
wt p53 cells uarrapoptosismutant p53 uarrnonapoptotic celldeath [17]
29M610 (mutant p53 resistant totreatment)
MJ uarrsim90 ATP depletion in bothcell types2DG high Glc but not pyruvatedarrATP
CJ MJ 05ndash25mM Nonsmall cell lung cancer linesA549 and H520
darrproliferation cell cycle arrest atG2M phase uarrp38 and ERK12phosphorylation uarrBaxuarrp21uarrcaspase-3
[37]
4 International Journal of Cell Biology
Table 1 Continued
Jasmonate (JAs conc range)vehicle Cell lines Effects References
MJ 05ndash20mM Hormone-refractory prostateadenocarcinomas PC-3 DU-145 darrproliferation uarrapoptosis darr5-LOX [47]
MJ 1ndash26mMEtOHmdash(IC50)
Murine melanoma cellsB16F10 and B16 COLR(overexpressing Pgp MDR)
darrcell motility darrcell growth darrMDR [39]
TBrJA(synthetic)
40 120583MEtOH(MJ doses lower than
in vitro)
Melanoma B16-F10Breast MCF-7Pancreas Mia PCA-2D122 PBL
uarrcitoxicity (TBrJA≫MJ) [39]
MJ 05ndash30mMno vehicle reported
CT-26 (murine colon carcinoma)B16 (murine melanoma)BCL1 (murine B-cell leukemia)Molt-4 (human T-lymphoblasticleukemia cell line)
MJ (but not JA) detached HK1 andHK2 from VDAC1 in isolatedmitochondria from thefour cell linesMJ did not inhibit HKs activitydarrATP uarrcyt c release uarrmitochondriaswelling uarrcell death
[20 30]
MJ 10-20mM Human neuroblastomaBE(2)-C
Arrest at S-phase darrcell growthdarrXIAP mRNA darrsurvivin mRNAuarrapoptosis
[43]
MJ 05ndash30mMDMSOHuman breast cancer MCF-7Human melanocytic MDA-MB-435cells [74]
Arrest at G0G1and S-phase darrmembrane fluidity uarrapoptosisextrinsic (TNFR1 uarrcaspase-8)intrinsic [darrΔ120595
119898uarrcaspase-3 (only
MDA-MB-435)]
[25]
CJ MJ 20mMDMSO Hormone-independent prostatePC-3 DU-145 cells
Cell cycle arrest darrcell growthuarrapoptosis(uarrTNFR1 uarrcaspase-3)
[48]
JA CJ andMJ 10-20mMDMSO
Human neuroblastomaSH-SY5Y
Arrest at G2M phase darrcell growthuarrapoptosis (darrXIAP darrsurvivin)activities MJ gt JA gt CJ
[44]
Human embryonic kidney HEK 293cells Not affected by MJ
MJ 10-20mMDMSO Human neuroblastomaSK-N-SH BE(2)-C
Arrest at G0G1 phase darrcell viabilitydarrmRNA of PCNA uarrapoptosis(darrXIAP darrsurvivin)
[45]
MJ 05ndash30mMEtOH Sarcomas MCA-105 uarrpAkt (correlates with lowersensitivity to cytotoxicity by MJ) [49]
SaOS-2 (resistent to MJ) MJ + 2DG uarrcytotoxicity
MJ 10ndash50mMEtOH
Cervical cancer SiHA CaSki andHeLa cells having wt p53Cervical cancer C33A cells (withmutated p53)
darrcell cycle uarrapoptosis throughdifferent pathwaysdarrATP uarrlactate (in more glycolyticCaSki) PARP cleavage multiple celldeath pathways depending on levelsof p53 p21 Bcl-2 and Bax
[26]
JA MJ 025ndash40mMEtOH Acute myelogenous leukemia cellsHL-60 and KG1
uarrROS uarrMJ-induced mitochondrialmembrane depolarizationuarrMJ-induced Mit SOD darrAKRC1
[132]
MJ MDDHJ 015mMDMSO Leukemia HL-60 cellsuarrCa2+-binding protein S100Puarrdifferentiation uarrregulator ofG-protein signaling-16 (RGS16)
[42]
J7 (synthetic) IC50 15 120583MHuman cervical carcinomaHeLa cells
Cell cycle arrest at G2M phasedarrBcl-2 (caspase 9 3) DNA damage [27]
International Journal of Cell Biology 5
Table 1 Continued
Jasmonate (JAs conc range)vehicle Cell lines Effects References
J7 (synthetic) 50120583MDMSO Human hepatoma Hep3B
uarrBaxBcl-2 ratio uarrDR5 uarrcaspase-8darrBid uarrapoptosis correlated withuarrcaspase-9 uarrcaspase-3 darrXIAP darrcIAPdarrPARP ExtrinsicintrinsicMAPK
[34]
MJ 05ndash25mMno vehicle reported
CD138+ tumorcells from MM patients
HK2 release from mitochondriarapid darrATP uarrapoptosis [105]
MJ 025ndash10mMno vehicle reported Human colorectal cancer cells CRC
uarrTRAIL uarrcyt c releaseuarrcaspase cleavage darrsurvivindarrTCF transcriptional activity
[32]
MJ 00ndash20mMEtOH Cervical cancer cellsSiHa CaSki HeLa and C33A
uarrmitochondrial O2
minus (HeLa CaSki)darrsurvivin darrE6 E7 (HPV)uarrdifferent cell death pathways(independently of HPV)
[28]
J7 (synthetic) 0ndash50 120583M DMSO Human hepatoma HepG2uarrROS uarrTRAIL-mediated apoptosis(darrBid darrXIAP darrcIAP darrBcl-xLuarrcaspases)
[35]
MJ 30mM (IC50) novehicle reported
Human adenocarcinoma colonHT-39
Arrest at S-G2M uarrcytotoxicityuarrapoptosis [31]
JA MJ 10ndash30mMDMSO Canine macrophagic malignantDM62 cells darrcell growth (MJ gt JA) uarrcytotoxicity [256]
MJ 05ndash02mMDMSO Human gastric SGC-7901 MKN-45cell lines
darrmigration darrinvasion darrangiogenesisdarrMMP-14 [33]
Bcl-2 B-cell lymphoma-2 Bcl-xL B-cell lymphoma-extra large Bid BH3 interacting domain death agonist cIAP cellular inhibitor of apoptosis CJ cisjasmonicacid 2DG 2-deoxy-D-glucose Glc glucose J7 methyl 5-chloro-45-didehydrojasmonate JA jasmonic acid NSCLC nonsmall-cell lung carcinoma PgpP-glycoprotein MDDHJ methyl 45-didehydro-jasmonate MDR multidrug resistance MMP-14 matrix metalloprotease 14 PARP poly (ADP-ribose)polymerase PCNA proliferating cell nuclear antigen PTPC permeability transition pore complex ROS reactive oxygen species S100P protein SP100 TBrJA57910-tetrabromo jasmonate TNFR1 tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand XIAP X-linked inhibitor of apoptosis protein
Table 2 Effect in vivo of natural and synthetic jasmonates derivatives
Jasmonates Conc rangevehicle Organismtissuecells Effects ReferencesMJ (oral) 236mgkglipofundin Normal C57BL6 mice Nontoxic [15]
MJ (oral) 236mgkglipofundin C57BL6 mice injected ip withmurine T-lymphoma EL-4
uarrsurvival of treated mice ascompared to the inoculated control [15]
TBrJA40120583MEtOH
(lower doses than MJin vitro)
Murine melanomaB16-F10 darrlung metastasis [39]
MJ 1ndash10120583MEtOH Chicken CAM darrangiogenesis [40]MJ 1ndash10120583MEtOH Human endothelial cells (HUVEC) darrCOX-2PGE2 pathway [40]
MJ (ip) 1 gKg in 01mLvegetal oil ip
Multiple myeloma- (MM1S-)inoculatedin nodscid mice
uarrsurvival of treated mice ascompared to the inoculated control [105]
MJ (topical) 1 gmL oil Application on cancerous andprecancerous human skin lesions
No local or systemic side effects38 patients withpositive responses
[51]
CAMchorioallantoic membrane of chicken embryoHUVEC human umbilical vein and endothelial cells TBrJA synthetic 5 7 9 10-tetrabromo jasmonate
mentioned that MJ was also assessed and approved by Foodand Agriculture OrganizationWorld Health Organization(FAOWHO) as a food additive Toxicity results reportedin the EPA document [52] were found to agree with thosepreviously published by other groups [50 51]
21 Jasmonates (MJ) Can Cause Cancer Cell Cycle Arrest atDifferent Phases in Different Cancer Cell Types Inhibiting
Growth and Proliferation The eucaryotic cell cycle is dividedinto four phases G
1 S G
2 and M that occur in response
to growth factors or mitogens The DNA synthetic (S) andmitotic (M) phases are preceded by gap phases (G
1 G2)
Chromosome duplication occurs during S phase replicatedchromosomes are segregated into individual nuclei (mitosis)during M phase and the cell then splits in two Cell cycleprogression can be regulated atG
1 G2points by various intra-
cellular and extracellular signals If extracellular conditions
6 International Journal of Cell Biology
Table 3 Effect in vitro and in vivo of methyl jasmonate (MJ) combined with other anticancer agents
Jasmonate Drug concrangevehicle Model Effects References
(1) MJ + BCNU(nitrosourea)in vitro
MJ Fixed conc Pancreatic MIA PaCa-1 Mitochondriotoxicsynergic cytotoxicity
[170](2) MJ + 2DGadriamycin taxolBCNU or cisplatinin vitro
MJ05ndash20mMEtOHTaxol cisplatin1ndash10120583gmL
CT26 DA-3 GTRAMP C1 MCF7MIA PaCa-2 D122 and BCL1
Strong cooperativeeffects of MJ + 2DGand MJ + other drugs
(3) MJ + adriamycinin vivo
MJ iv20ndash150mgkgdissolved inlipofundinAdr (DOX) ip4mgkg
Balbc mice injected ip with 1 times 104chronic BCL1 cells lymphocyteleukemia cells
MJ + Adr significant prolongedsurvival effect
(4) MJ + PI3KAktinhibitorsin vitro
MJ 05ndash30mMEtOHDMSO
Sarcoma MCA-105 SaOS-2 darrMJ-induced activation of Aktuarrsynergic cytotoxicity
[49](5) MJ + 2DGin vitro
MJ 05ndash30mMEtOHDMSO Sarcoma MCA-105 SaOS-2 2DG darrpAkt darrMJ-ind pAkt
MJ + 2DG uarrsynergic cytotoxicity
(6) MJ + irradiationin vitro
05ndash20mMDMSO
Irradiated prostate PC-3 (radiationinduces Bcl-2 expression)
darrradiation-induced Bcl-2uarrradiation sensitivity PC-3uarrcaspase-3
[104]
(7) MJ andor POHandorcis-platin (CP)in vitro
Both testedat IC20
Human MDA-MB-435[74]
MJ + POH uarrTNFR1darrΔ120595119898uarrcytotoxicity cell cycle arrest
at G0G1 uarrapoptosis+ CP all effects enhanced
[251]
Human breastMDA-MB-231 uarrapoptosis
(8) MJ + TRAILin vitro
MJ 05mM +TRAIL(100ndash200 ngsdotmLminus1)
CRC cancer cells
darrsurvivin (IAP)darrWntTCF pathwayuarrTRAIL-induced apoptosisuarrcaspase activity
[32]
(9) MJ + Smacin vitro
MJ 05ndash20mMDMSO
Human bladder cancer EJ T24 Synergy uarrIAPs-bound caspase 3uarrapoptosis [254]
Human embryonic kidney HEK293 No cytotoxicity
(10) MJ + Smac7N(IAP antagonist)in vitro
MJ 05ndash20mMDMSO
Hormone-independent prostateDU-145 PC-3Human proximal tubular epithelialHK-2 cells (overexpressing IAPs)
Smac7N uarrMJ-induced apoptosis bycaspase-9-dependent (intrinsic) andindependent (extrinsic) pathways
[255]
(11) MJ + cisplatin(01ndash05 120583M)MJ + X-rays(025ndash3Gy)MJ + 120572-rays
MJ 01ndash10mMEtOH
Cervical cancer cellsSiHa CaSki HeLa and C33A
darr viabilitydarrsurvivaldarrIC50 radiation dose
[29]
(12) MJ + 5-FUin vitro
05mMMJ1 hthen + 5-FU
Human adenocarcinomacolon HT-39 darrIC50 5-FU (5 rarr 25mM) [31]
BCNU 13-bis-(2-chloroethyl)-1-nitrosourea 2DG 2-deoxy-D-glucose 5-FU 5-fluorouracile IAP inhibitors of apoptosis MM multiple myeloma POHperillyl alcohol smac second mitochondria-derived activator of caspases Smac7N a peptide that contains the N-terminal seven residues of smac TNFR1tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand
are unfavorable cells delay progress through G1and may
enter a quiescent state known asG0 in which they can remain
for days weeks or years before resuming proliferation Ifextracellular conditions are favorable and signals to growand divide are present cells in early G
1or G0progress
through a restriction checkpoint late in G1after which cells
are committed to DNA replication even if the extracellularsignals that stimulate cell growth and division are removed[53 54] Cell cycle activity is modulated by intracellularROS levels and regulates cell survival cell proliferation andapoptosis [53ndash58]The progression through theG
1 S G2 and
M phases is promoted by cyclin-dependent kinases (CDKs)
International Journal of Cell Biology 7
which are positively and negatively regulated by cyclins andcyclin kinase inhibitors (CKis) respectively and by ROSUnlike the passage through the S G
2 and M phases G
1
progression normally relies on stimulation by mitogens andcan be blocked by antiproliferative factors passage throughthe restriction checkpoint and entry into S phase is controlledby CDKs that are sequentially regulated by cyclins D Eand A [53 54 56ndash58] In general CDK activity requirescyclin binding and depends on both positive and negativeregulatory phosphorylations [54] ROS levels influence thephosphorylating and ubiquitinating enzyme activites thuscontrolling cell cycle progression [54 57 58]
A master gene regulating cell passage through the cycleis the Myc family of transcription factors it switches onthe simultaneous synthesis of thousands of different proteinsrequired for cell growth and cell cycle function [58 59] TocounteractMyc protein activity the cell cycle is also regulatedby the p53 protein [60 61] a DNA-binding transcriptionfactor that senses DNA damage (occurring after exposureto ionizing radiation UV light oxidative stress or otherDNA damaging agents) and activates genes to halt cell cycleprogression in G
1for repair or otherwise initiates apoptosis
[56 62] When the damage caused by cell stress is mild p53activatesmechanisms for cell cycle arrest and repair damagedDNA if the damage is irreparable p53 initiates apoptosis [63]In case ofmild cellular stress low levels of p53 can activate theTP53-induced glycolysis and apoptosis regulator (TIGAR)gene that decreases the rate of glycolysis and hence ROS levelsin the cell [64] Glycolysis is essential for cancer progressionbut TIGAR provides protection from ROS and apoptosisHowever if damage caused by stress is irreparable p53prevents proliferation of cells with altered DNA and inducesincreased levels of ROS to trigger cell death [56] Increasedlevels of active p53 inhibit the cell entry into the S-phaseor induce apoptosis [56] Loss or inactivation of p53 resultsin loss of cell-cycle arrest or inhibition of apoptosis afterDNAdamage or physiological stress [65] leading to increasedgenetic instability increased accumulation of mutations andultimately oncogenesis [61] When the cell cycle is no longerregulated by p53 cells remain cycling (supported by 119888-Mycoverexpression) subverting cell cycle exit maturation andterminal differentiation [66] Over 50 of all human cancersharbor mutations and genetic alterations in cyclins and p53genes directly affecting the function of critical cell cycleproteins rendering the p53 gene ineffective in mediating cellgrowth arrest and promoting cancer cell death [62 67 68]ROS production is under p53 regulation and in turn p53being a redox-sensitive protein is influenced by ROS levelsthus ROS can act as both an upstream signal that triggersp53 activation and as a downstream factor that mediatesapoptosis [69]Thus mutations in the p53 gene downregulateits activity and consequently ROS production [70]ThroughROS p53 can also directly control metabolic traits of cells[71] regulate mitochondrial membrane potential (Δ120595
119898) and
induce cytochrome 119888-independent apoptosis (blocked by Bcl-2) ROS-mediated disruption of Δ120595
119898constitutes a pivotal
step in the apoptotic pathway of p53 and this pathwaydoes not involve cytochrome 119888 release [72] ROS can thusregulate cell fate through p53 in a way that physiological ROS
levels trigger cell protective pathways while under cytotoxicoxidative stress p53 behaves more like a cell killer [70 73]
JAs (mainly MJ) have been found to arrest cell cycle ofdifferent cancer cell types at different phases (Table 1) G
0G1
in human acute lymphoblastic leukemia Molt-4 [14 38] andhuman neuroblastoma SK-N-SH BE(2)-C [45] G
0G1-S in
human breast MCF-7 andmelanocytic MDA-MB-435 cancercells [25 74] S in human neuroblastoma BE(2)-C [43] S-G2M in human adenocarcinoma colon HT-39 [31] G
2M
in nonsmall cell lung cancer (NSCLC) lines A549 and H520[37] human neuroblastoma SH-SY5Y [44] human cervicalcarcinoma and HeLa cells [27] At 01mM MJ arrested invitro human umbilical vein endothelial cells (HUVEC) at G
1
[40]The molecular mechanisms underlying the above jasmo-
nate-mediated cell cycle arrest in cancer and embryonicHUVEC cells in vitro are not yet completely understoodChecking the above data against the UMD TP53 mutationdatabase (httpwwwp53fr) and revising the article byBerglind et al [75] we could not find correlation between agiven MJ-mediated phase arrest of cell cycle and the TP53wild type or mutated status of a cancer cell type (data notshown) a result that would agree with the original observa-tion of Fingrut and Flescher [15] stating thatMJ-mediated celldeath is independent of the p53 status [17] However otherstudies suggest thatMJ could be acting by interfering differentcell cycle regulatorymechanisms In plants MJ is able to bindthe TGACCG motif which is present in cell cycle cyclin pro-moters [76] wemight speculateMJ recognition of TGACCG-like motifs in mammalian cells cyclin promoters howeverthis would need experimental confirmation In some plantsMJ impairs the G
2-to-M transition by repressing M-phase
gene activation [77] MJ impaired the G2-to-M transition in
the human neuroblastoma SH-SY5Y cells downregulatingthe expressions of the proliferating cell nuclear antigen(PCNA a processivity factor encircling DNA at sites ofreplication and repair) andN-Myc [45] Asmentioned aboveMYC is the master gene allowing cancer cells to move alongthe cell cycle and regulating most genes needed to meet themetabolic demands for cell proliferation and survival [78]Cancer cell cycle arrest at different phases is in itself closelydependent on the cell energy and as such it is extremelydependent on metabolic ATP availability and its use byhexokinase Downregulation of Myc byMJ [45] is interestingresult that must be confirmed particularly because there is alack of specific inhibitors against Myc [57]
In some cases of cell cycle arrest MJs have been foundto upregulate p21 [37] whereas in others to downregulatesurvivinmRNA and protein levels [43ndash45] (Table 1) Survivinis an important member of the inhibitor of apoptosis protein(IAP) family that also regulates entrance of the cell cycle to theM phase and is overexpressed in cancer cells acting linkingboth regulation of the cell cycle and inhibition of apoptosis[79]
22 Jasmonates (MJ) CanCauseCancer Cell Death by InducingApoptosis andor Necrosis Apoptosis can be initiated by twosignaling cascades namely the extrinsic and the intrinsic
8 International Journal of Cell Biology
pathway The classical extrinsic pathway is triggered by thebinding of extracellular signals like tumor necrosis factor(TNF) tumor necrosis factor-related apoptosis-inducing lig-and (TRAIL) and apoptosis antigen-1 (Fas) ligand (FASL)to death receptors such as tumor necrosis factor receptor 1(TNFR1) and FAS [80]This interaction leads to procaspase-8activation which cleaves itself into caspase-8Then caspase-8can cleave and activate procaspase-3 into caspase-3 thatcleaves many substrates inside the cell inducing the typicalmorphological changes seen in apoptosis The intrinsic path-way can be triggered by intracellular stress signals such asoxidative stress and DNA damage The diverse signals aresensed by mitochondria through the balance between pro-and antiapoptotic members of the Bcl-2 family of proteinsWhen proapoptotic signals overcome antiapoptotic signalsmitochondria becomepermeabilized leading to dissipation ofthe mitochondrial electrochemical potential release of inter-membrane proteins like cytochrome 119888 SmacDIABLO AIFandHTRA2OMI Cytochrome 119888 interacts with procaspase-9and Apaf-1 to form the apoptosome complex that leads tocaspase-3 activation [81] The extrinsic pathway can connectwith the intrinsic pathway through the cleavage by caspase-8of Bid into tBid (truncated Bid) which is a proapoptoticmember of the Bcl-2 family and can lead to mitochondriapermeabilization [82 83] The members of the Bcl-2 fam-ily regulate mitochondria permeabilization and comprisesantiapoptotic (Bcl-2 Bcl-xL) proapoptotic (Bax Bak) andproapoptotic BH-3 only proteins (Bad Bid Puma andNoxa)[82ndash84] One of the mechanisms of mitochondria perme-abilization is controlled by Bax and Bak proteins that arenormally expressed in every cell but are maintained inhibitedby the antiapoptotic members When there is a signal toapoptosis BH3-only proteins interact with the antiapoptoticmembers releasing BaxBak or directly activating themleading to their oligomerization and to mitochondrial outermembrane permeabilization (MOMP) Another mechanismfor mitochondria permeabilization is the formation of themitochondrial permeability transition (MPT) pore which iscomposed by the voltage-dependent anion channel VDACthe adenine nucleotide translocase (ANT) and cyclophilin D[85]TheMPT pore comprising VDAC1 is directly regulatedby Bad [86] In contrast VDAC2 has been identified as anantiapoptotic negative regulator of Bak [87ndash89]
One of the hallmarks of cancer is resistance to cell death[90] This resistance is acquired by a host of different geneticdefects in cancer cells such as those present in the p53tumor suppressor gene [62 68 91] Normal p53 responds togenotoxic oncogenic and other stress signals by inducingantiproliferative transcriptional programs leading to growtharrest (for cell repair) or apoptosis [92] p53 mediates cyclearrest by transcriptionally activating the p21 (CDKN1A) 14-3-3120590 (SFN) and GADD45120572 genes whereas p53-dependentapoptosis is triggered by turning on the synthesis of proteinsthat produce ROS [58 93] and transactivating differentmitochondrial proapoptotic genes as well as death receptorsfor the synthesis of the corresponding proteins [58 94]Whenp53 (and its target genes) expression levels are below a thresh-old they induce cell cycle arrest but not apoptosis above thisthreshold p53 trigger apoptosis [95] Lowering this threshold
with inhibitors of the antiapoptotic Bcl-2 family proteinssensitize cells to p53-induced apoptosis [95] By damagingcells chemotherapeutical drugs turn on p53rsquos transcriptionalfunction that upregulates the expression of proapoptotic pro-teins Defects in the p53 function or in the apoptotic cascadeslead to cell phenotypes resistant to chemotherapy Apoptosiscan also be induced by anticancer agents independently ofp53 by either directly modulating the apoptotic machineryor directly interacting with mitochondria [96] ROS levelsmight be playing a central role in mediating various forms ofnonapoptotic caspase-independent programmed cell deatheither through direct irreversible oxidative damage to keyproteins nucleic acid molecules and cellular structure or bythe activation of prodeath signal transduction programs [97]In a ROS-dependent nonapoptotic cell death pathway respi-ration through the mitochondrial electron transport system(ETS) chain is not essential and ROS is generated throughNADPH oxidases [97] In contrast to the mitochondrial ETSwhich generates ROS as a byproduct of respiration NADPHoxidases produceROS as their primary function [98] Raise ofROS is characteristic after ionizing radiation chemotherapyand targeted therapy treatments [58 99] and may be killingcancer cells through a nonapoptotic ROS-dependent celldeath pathway
Tumor cells maintain high ATPADP as well asNADHNAD+ ratios so as to assure that ATP levels arenever limited ATP depletion would represent then anunsustainable metabolic stress for tumor cells and it isassociated with necrotic death in some cell types [100]Recently it was shown that a signaling cascade centered onthe proteins RIP1 and RIP3 can trigger cell death by necrosisin many situations and the RIP1-RIP3 complex appears topromote ATP depletion during necroptosis by impingingon components of the MPT pore [101 102] Necrosis was aterm used to describe a process of accidental cell death inwhich cells swell and burst releasing its intracellular contentwhereas necroptosis modernly refers rather to an orderedcell death program different from apoptosis apoptosis is acaspase-dependent cell death process whereas necrosis iscaspase-independent process [102 103] Studies concerningnecrosis have focusedmainly on cell membrane permeabilityto propidium iodide (PI) in the flow citometry doublestaining with annexin-VPI
Fingrut et al [17] by studying the effect of MJ ontwo clones of a highly malignant B-cell lymphoma (oneharboring a normal wild type (wt) p53 gene while the otherexpressed a mutated inactive p53 protein) (Table 1) reportedthat MJ was equally toxic to both clones [17] In contrastthe p53-mutant cells were resistant to radiomimetics (drugshaving effects similar to those produced in radiotherapy)and chemotherapeutic drugs In this cancer cell model MJinduced mostly apoptotic cell death in the wt p53-expressinglymphoma cells while no signs of early apoptosis weredetected in the mutant p53-expressing lymphoma cells thatwere killed by MJ through a p53-independent nonapoptoticcell death mechanism However other cancer cell types canundergo apoptosis with or without functional p53 and aswe will see MJ can induce apoptotic andor nonapoptoticcell death in wt p53 as well as in TP53 mutated cells
International Journal of Cell Biology 9
(Table 1) Nonapoptotic cell death could be the consequenceof the severe bioenergetic damages induced by the directinteraction of MJ with mitochondria leading to increasedROS levels inhibition of ATP synthesis and ATP depletion[20 30] Jasmonates including MJ and some of its analogshave been found to be strongly proapoptotic in vitro indifferent types of cancer cells by both the extrinsic and theintrinsic pathway (Table 1) with no or little cytotoxicity tonormal cells [15 25 36 37 44 45] Thus MJ induced theexpression of TNFR1 in human breast MCF-7 and MDA-MB-435 cells [25 74] and in hormone-independent humanprostate PC-3 and DU 145 cancer cells promoting apoptosisby the extrinsic pathway in these cells [48] TRAIL (TNF-related apoptosis inducing ligand) is able to induce apoptosisin prostate cancer cell yet overexpression of antiapoptoticproteins and inhibition of proapoptotic proteins results indiminishing the TRAIL-mediated apoptosis pathway TRAILhas emerged as a potent anticancer agent in laboratory-based studies and preclinical trials and JAs have been foundto sensitize cancer cells to TRAIL-mediated apoptosis [32](Table 3) J7 a synthetic analogue of MJ enhanced TRAIL-mediated apoptosis through upregulation of ROS levels inhuman hepatomaHepG2 cells apoptosis was induced via Bidcleavage downregulation of X-linked inhibitor of apoptosisprotein (XIAP) cellular inhibitor of apoptosis-1 (cIAP-1)B-cell lymphoma-extra large (Bcl-xL) and activation ofcaspases [35] Other examples of MJ-mediated apoptoticandor nonapoptotic cell death in different cancer cell types(Table 1) are as follows (1) MJ induced ROS production inA549 lung adenocarcinoma cells activating Bcl-2-associated-X-protein (Bax) as well as Bcl-2-Like-1 (Bcl2L1) proapoptoticproteins in these cells [36] (2) MJ activated caspase-3 inleukemia Molt-4 prostate SK28 and LNCaP cancer cells[15] (3) MJ inhibited the antiapoptotic Bcl-2 protein andenhanced proapoptotic proteins in radio-resistant humanprostate cancer cell line PC-3 [104] (4) MJ suppressed theproliferation of the neuroblastoma cell lines SH-SY5Y SK-N-SH and BE(2)-C through a strong apoptosis-inducingeffect as measured by annexin VFITCPI MJ treatment alsodownregulated the expression of the antiapoptotic proteinsXIAP and survivin [44 45] (5) MJ-mediated cell death wasobserved on cervical cancer and myeloma cells and it wasreported to be due to mixed characteristics of apoptosis andnecrosis [26 105] So far studies showing MJ inducing theRIP1 signaling cascade have not been reported howeverit will be important to deep into these analyses to cleardistinguish between MJ-induced apoptosis andor necrosis
23 Methyl Jasmonate Induces the Production of ReactiveOxygen Species (ROS) Responses in Some Cancer Cell TypesLeading Also to Cell Death Intracellular ROS levels reg-ulate cell cycle activity cell metabolism and survival cellproliferation and apoptosis [53ndash58] Stressful conditionsproduce a transient excess of intracellular ROS in normaltissues however under chronic pathological and oxidativestress conditions ROS is continuously produced in excessand persistently causing damage to DNA cell membraneslipids proteins and inducing apoptosis to control and avoid
oxidative damages cells counteract excess ROS by enzymatic(superoxide dismutase (SOD) glutathione peroxidase (GPx)and catalase) andor nonenzymatic (glutathione thoredoxin)antioxidant elements [99 106] Controlling ROS is central tointracellular redox signaling and homeostasis on the otherhand uncontrolledunbalanced ROS production modifiesintracellular redox leading to apoptosis [58 82 99 106] ROScan be generated by different biological systems (1) director indirect extracellular stimulation of plasma membrane-bound NAD(P)H oxidases (NOX) can generate intracellularH2O2[57 107ndash110] (2) biochemical pathways associated to
fatty acid oxidation and lipoxygenases activities generateROS as normal byproducts of these activities [111] (3)ATP generation by oxidative phosphorylation (OxPhos) inmitochondria is accompanied by the production of ROS as aconsequence of electron leakage from the electron transportchain mitochondria are considered to be major sources ofROS inmammalian cells particularly under a variety of stress-ful conditions [82 99] Superoxide (O
2
minus) is mainly produced
from complexes I and III and it is rapidly dismutated to H2O2
and oxygen by SOD [112] H2O2serves both as a toxic oxidant
and as an essential signaling molecule regulating cellularbiological processes [113] A strong positive correlation existsbetween mitochondrial membrane potential (Δ120595
119898) and ROS
production [106 114 115] Mitochondria produce more ROSat high membrane potential and an increase in the Δ120595
119898
produced either by a closure of MPT pore or by inhibitionof the ATP synthase is associated with increased ROS pro-duction However in cases of mitochondrial dysfunctionslow Δ120595
119898and decreased activity of the respiratory chain
are accompanied by a simultaneous increase in ROS [116117] It has been reported that cancer cells have higher ROScontent compared to normal cells [118 119] probably due toabnormal respiration because of dysfunctional mutations inmitochondria [120] but they also have increased contents ofantioxidants (SOD catalase glutathione thioredoxin etc)to counteract ROS [58 121] Tumor cells cannot tolerateexcessive high ROS levels [118 122] and as a matter of factradiation therapy chemotherapies and targeted therapiesinduce apoptosis and kill some cancer cells by generatingexcess ROS [99 106 123 124] Thus inducing higher levelsof ROS in cancers cells can be exploited as an effectiveselective strategy to kill tumor cells over normal cells [119]However prolonged treatment with the same drug reducesROS levels in cancer cells [124] indicating that when theybecome drug-resistant tumor cells have lower ROS contentthan drug-sensitive cancer cells Evidence suggests that drug-resistant cells have a higher expression of catalase at theplasma membrane that keeps reduced ROS levels [121]
In plants MJ-mediated damage to mitochondria is fol-lowed by a rapid production of H
2O2[125] H
2O2is used as
a second messenger for the induction of genes and productsrelated to defense against both herbivores and pathogens[126] in addition H
2O2induces apoptosis and senescence
[127 128] On the other hand it has been signaled that theMJ-induced production of H
2O2in plants was prevented by
inhibitors of NAD(P)H oxidase [128 129] indicating anothermechanism of MJ-mediated production of ROS and a majorrole for this enzyme in MJ-induced ROS responses MJ
10 International Journal of Cell Biology
induces also an endogenous H2O2response in some cancer
cell lines (Table 1) [19 36 38 130ndash132] In A549 cancerlung cells the H
2O2response induced by MJ increased the
expression of the proapoptotic Bax and Bcl-Xs (although notbeing that of the antiapoptotic Bcl-2 and Bcl-xL proteins) andthis led to apoptosis this event was inhibited by catalase (aspecific inhibitor of H
2O2) [36] MJ increased the intracel-
lular levels of H2O2and superoxide (O
2) ions in human C6
glioma cells in vitro and induced the expression of heat shockprotein 72 (HSP72) a negative regulator of H
2O2-induced
oxidative stress in these cells in vitro MJ-induced HSP72expression was blocked by ROS inhibitors [130] ElevatedROS generation was observed in response to both MJ andJA levels in the acute myeloid leukemia HL-60 and KG1a celllines but other factors like the MJ-mediated generation ofmitochondrial SOD rather than ROS were determinant forapoptosis or differentiation in these cells [132] Suppression ofMJ-induced apoptosis by antioxidants like N-acetylcysteine(NAC) and catalase but not by inhibitors of hydroxyl radicalsand superoxide ions indicates that H
2O2is nonetheless one
of the important factors involved inMJ anticancer effects [19]
24 By Dissociating Hexokinase (HK) from the VoltageDependent Anion Channel (VDAC) on the Outer Mitochon-drial Membrane MJ Dissociates Glycolysis from OxidativePhosphorylation Causing Severe Bioenergetic Deregulations inCancer Cells Metabolism and cell growth are two cellularprocesses that are tighly linked and regulated [133] Theclassic bioenergetics phenotype of cancer cells of enhancedglycolysis under aerobic conditions was described by War-burg [134] However the ldquoWarburg effectrdquo does not neces-sarily imply mitochondrial dysfunction [135] To increaseglycolysis cancer cells upregulate the transcription of genesinvolved in the glycolytic pathway (ie glucose transportersglycolytic enzymes etc) This metabolic reprogrammingallows increased rate of ATP production synthesis of lipidsand a new redox balance Cancer cells actually use a com-bination of both glycolysis and mitochondrial respirationto produce energy and they may vary in regards to thepreferential use of these pathways being in some caseseither more glycolyticless oxidative or less glycolyticmoreoxidative depending on the prevalent normoxic or hypoxicenvironmental conditions and their capacities to expressadequate levels of oncogenes and tumor suppressor geneproducts for cell growth [136ndash139] or when either glycolysisor OxPhos is inhibited in whose case there can be partialcompensation by the other metabolic pathway [17] Shiftingthe balance between those two processes with glycolyticandor mitochondrial enzyme inhibitors might represent aninteresting way to look for new anticancer agents
The first step in the metabolism of glucose is catalyzedby the enzyme hexokinase (HK) which by phosphorylatingglucose makes it negative sequestering it into the cytoplasmin the form of glucose-6-phosphate (G6P) In normal tissuesthis crucial step is catalyzed by four different HK isoforms(HK1 HK2 HK3 and glucokinase) indicating that regulationof glucose phosphorylation can vary in different tissues underdifferent condition [140] HK1 and HK2 have overlapping
tissue expression but different subcellular distributions withHK1 associated mainly with mitochondria and HK2 associ-atedwith bothmitochondrial and cytoplasmic compartments[141] HK1 is widely and constitutively expressed whereasHK2 predominates during embryogenesis and is not widelyexpressed in adult tissues but only in a limited numberof normal adult tissues [141] their different subcellulardistributions and kinetic properties reflect their differentmetabolic roles [141 142] Under physiological conditionsHK1 is predominantly bound to the outer mitochondrialmembrane where it primarily channels glucose toward gly-colysis [140] whereas HK2 is mainly soluble and controlsglycogen formation [140 141] In malignant tissues HK1and HK2 isoforms are glycolytic and overexpressed (HK1more than HK2) (httpwwwproteinatlascom) [143] andare tightly bound to mitochondria HK1 and HK2 bindthrough theirN-terminal hydrophobic regions to the voltage-dependent calcium channel isoform1 (VDAC1) the prevalentpore protein in the outer mitochondrial membrane [2482 143ndash146] VDAC1 is also overexpressed in cancer cells[147] and the binding of HK to VDAC1 on the outer mito-chondrial membrane is a fundamental aspect of the aerobicglycolytic metabolism of cancer cells [82 135 147 148]HKVDAC association appears to protect tumor cells frommitochondrial outer membrane permeabilization (MOMP)which marks a point of no return leading to cell death[149] VDAC transports ADP and inorganic phosphate (Pi)the substrates needed for the production of ATP into themitochondria it also transports ATP out of themitochondriainto the cytoplasm and facilitates the availability of ATPto HK2 [143 150 151] A continuous supply of glucose toHK maintains this activity at a constant level and controlsthe flux through the mitochondrial electron transport chaininfluencing mitochondrial ROS production [152] HK bindsthrough hydrophobic interaction to VDAC1 the prevalentpore protein in the outer mitochondrial membrane [24 82143ndash146] thereby assuring and gaining a preferential accessto ATP [82 150 153] Translocation of HK2 from the cytosolto the mitochondria is regulated through phosphorylationof HK by Akt [141] HK2 only binds VDAC after phos-phorylation by Akt [151 152] Glycogen synthase kinase 3beta (GSK3120573) a major target of Akt kinases phosphorylatesVDAC [154ndash157] and dissociates HK [156] HK2 boundto mitochondria enhance its affinity to ATP and is lesssensitive to enzymatic inhibition by the G6P product [140141 158] HK2 can produce G6P from cytoplasmic glucoseand mitochondrial ATP without regulatory restraints evenunder hypoxia supporting rapid cell growth [142 143 159ndash161]HK1 andHK2 aremore tightly bound toVDAC in cancercells than in nonmalignant cells [156 162 163] HKVDA1interaction is relatively stable for HK1 while it is dependentupon G6P levels for HK2 [140 141] The higher affinity ofmitochondrial HK2 for glucose ATP and VDAC1 representsa high advantage for cancer cells survival and growth con-ferring to them an aerobic glycolytic phenotype underlyingthe Warburg effect [82 135 138 142 147 148 150 160]Differential interactions of HK1 and HK2 with mitochondriamay underlie different glycolytic profiles in cancer cells [164165] As rate-controlling enzymes in the glycolyic pathway
International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
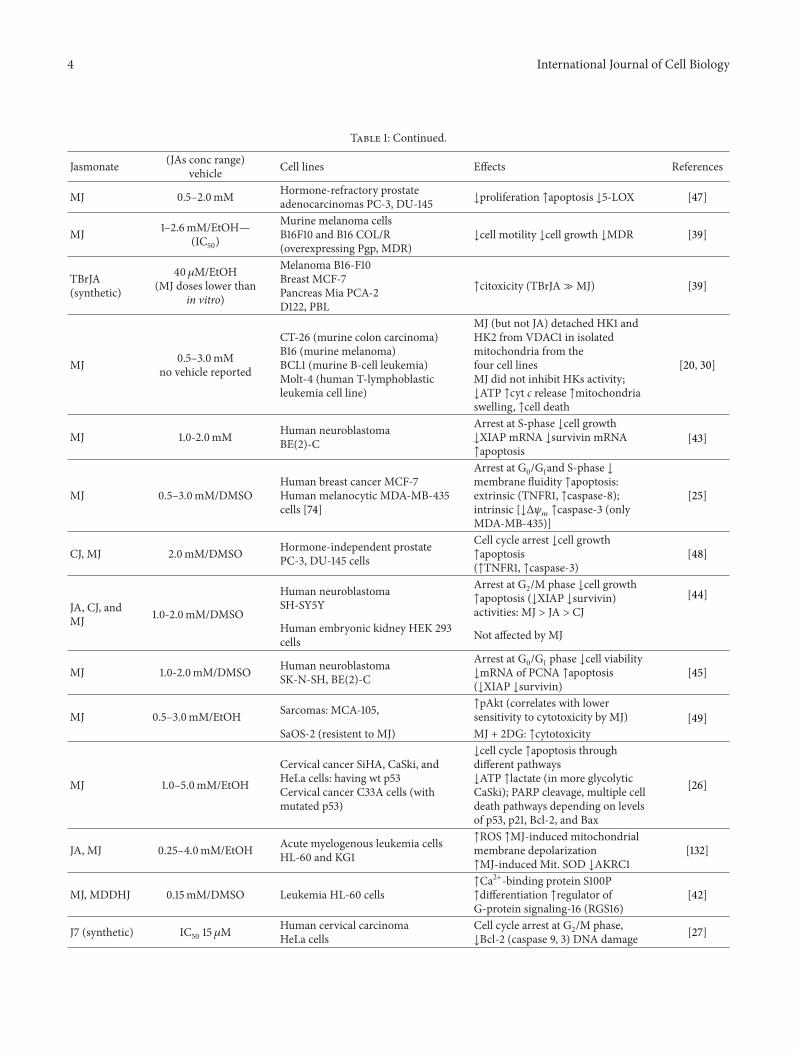
4 International Journal of Cell Biology
Table 1 Continued
Jasmonate (JAs conc range)vehicle Cell lines Effects References
MJ 05ndash20mM Hormone-refractory prostateadenocarcinomas PC-3 DU-145 darrproliferation uarrapoptosis darr5-LOX [47]
MJ 1ndash26mMEtOHmdash(IC50)
Murine melanoma cellsB16F10 and B16 COLR(overexpressing Pgp MDR)
darrcell motility darrcell growth darrMDR [39]
TBrJA(synthetic)
40 120583MEtOH(MJ doses lower than
in vitro)
Melanoma B16-F10Breast MCF-7Pancreas Mia PCA-2D122 PBL
uarrcitoxicity (TBrJA≫MJ) [39]
MJ 05ndash30mMno vehicle reported
CT-26 (murine colon carcinoma)B16 (murine melanoma)BCL1 (murine B-cell leukemia)Molt-4 (human T-lymphoblasticleukemia cell line)
MJ (but not JA) detached HK1 andHK2 from VDAC1 in isolatedmitochondria from thefour cell linesMJ did not inhibit HKs activitydarrATP uarrcyt c release uarrmitochondriaswelling uarrcell death
[20 30]
MJ 10-20mM Human neuroblastomaBE(2)-C
Arrest at S-phase darrcell growthdarrXIAP mRNA darrsurvivin mRNAuarrapoptosis
[43]
MJ 05ndash30mMDMSOHuman breast cancer MCF-7Human melanocytic MDA-MB-435cells [74]
Arrest at G0G1and S-phase darrmembrane fluidity uarrapoptosisextrinsic (TNFR1 uarrcaspase-8)intrinsic [darrΔ120595
119898uarrcaspase-3 (only
MDA-MB-435)]
[25]
CJ MJ 20mMDMSO Hormone-independent prostatePC-3 DU-145 cells
Cell cycle arrest darrcell growthuarrapoptosis(uarrTNFR1 uarrcaspase-3)
[48]
JA CJ andMJ 10-20mMDMSO
Human neuroblastomaSH-SY5Y
Arrest at G2M phase darrcell growthuarrapoptosis (darrXIAP darrsurvivin)activities MJ gt JA gt CJ
[44]
Human embryonic kidney HEK 293cells Not affected by MJ
MJ 10-20mMDMSO Human neuroblastomaSK-N-SH BE(2)-C
Arrest at G0G1 phase darrcell viabilitydarrmRNA of PCNA uarrapoptosis(darrXIAP darrsurvivin)
[45]
MJ 05ndash30mMEtOH Sarcomas MCA-105 uarrpAkt (correlates with lowersensitivity to cytotoxicity by MJ) [49]
SaOS-2 (resistent to MJ) MJ + 2DG uarrcytotoxicity
MJ 10ndash50mMEtOH
Cervical cancer SiHA CaSki andHeLa cells having wt p53Cervical cancer C33A cells (withmutated p53)
darrcell cycle uarrapoptosis throughdifferent pathwaysdarrATP uarrlactate (in more glycolyticCaSki) PARP cleavage multiple celldeath pathways depending on levelsof p53 p21 Bcl-2 and Bax
[26]
JA MJ 025ndash40mMEtOH Acute myelogenous leukemia cellsHL-60 and KG1
uarrROS uarrMJ-induced mitochondrialmembrane depolarizationuarrMJ-induced Mit SOD darrAKRC1
[132]
MJ MDDHJ 015mMDMSO Leukemia HL-60 cellsuarrCa2+-binding protein S100Puarrdifferentiation uarrregulator ofG-protein signaling-16 (RGS16)
[42]
J7 (synthetic) IC50 15 120583MHuman cervical carcinomaHeLa cells
Cell cycle arrest at G2M phasedarrBcl-2 (caspase 9 3) DNA damage [27]
International Journal of Cell Biology 5
Table 1 Continued
Jasmonate (JAs conc range)vehicle Cell lines Effects References
J7 (synthetic) 50120583MDMSO Human hepatoma Hep3B
uarrBaxBcl-2 ratio uarrDR5 uarrcaspase-8darrBid uarrapoptosis correlated withuarrcaspase-9 uarrcaspase-3 darrXIAP darrcIAPdarrPARP ExtrinsicintrinsicMAPK
[34]
MJ 05ndash25mMno vehicle reported
CD138+ tumorcells from MM patients
HK2 release from mitochondriarapid darrATP uarrapoptosis [105]
MJ 025ndash10mMno vehicle reported Human colorectal cancer cells CRC
uarrTRAIL uarrcyt c releaseuarrcaspase cleavage darrsurvivindarrTCF transcriptional activity
[32]
MJ 00ndash20mMEtOH Cervical cancer cellsSiHa CaSki HeLa and C33A
uarrmitochondrial O2
minus (HeLa CaSki)darrsurvivin darrE6 E7 (HPV)uarrdifferent cell death pathways(independently of HPV)
[28]
J7 (synthetic) 0ndash50 120583M DMSO Human hepatoma HepG2uarrROS uarrTRAIL-mediated apoptosis(darrBid darrXIAP darrcIAP darrBcl-xLuarrcaspases)
[35]
MJ 30mM (IC50) novehicle reported
Human adenocarcinoma colonHT-39
Arrest at S-G2M uarrcytotoxicityuarrapoptosis [31]
JA MJ 10ndash30mMDMSO Canine macrophagic malignantDM62 cells darrcell growth (MJ gt JA) uarrcytotoxicity [256]
MJ 05ndash02mMDMSO Human gastric SGC-7901 MKN-45cell lines
darrmigration darrinvasion darrangiogenesisdarrMMP-14 [33]
Bcl-2 B-cell lymphoma-2 Bcl-xL B-cell lymphoma-extra large Bid BH3 interacting domain death agonist cIAP cellular inhibitor of apoptosis CJ cisjasmonicacid 2DG 2-deoxy-D-glucose Glc glucose J7 methyl 5-chloro-45-didehydrojasmonate JA jasmonic acid NSCLC nonsmall-cell lung carcinoma PgpP-glycoprotein MDDHJ methyl 45-didehydro-jasmonate MDR multidrug resistance MMP-14 matrix metalloprotease 14 PARP poly (ADP-ribose)polymerase PCNA proliferating cell nuclear antigen PTPC permeability transition pore complex ROS reactive oxygen species S100P protein SP100 TBrJA57910-tetrabromo jasmonate TNFR1 tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand XIAP X-linked inhibitor of apoptosis protein
Table 2 Effect in vivo of natural and synthetic jasmonates derivatives
Jasmonates Conc rangevehicle Organismtissuecells Effects ReferencesMJ (oral) 236mgkglipofundin Normal C57BL6 mice Nontoxic [15]
MJ (oral) 236mgkglipofundin C57BL6 mice injected ip withmurine T-lymphoma EL-4
uarrsurvival of treated mice ascompared to the inoculated control [15]
TBrJA40120583MEtOH
(lower doses than MJin vitro)
Murine melanomaB16-F10 darrlung metastasis [39]
MJ 1ndash10120583MEtOH Chicken CAM darrangiogenesis [40]MJ 1ndash10120583MEtOH Human endothelial cells (HUVEC) darrCOX-2PGE2 pathway [40]
MJ (ip) 1 gKg in 01mLvegetal oil ip
Multiple myeloma- (MM1S-)inoculatedin nodscid mice
uarrsurvival of treated mice ascompared to the inoculated control [105]
MJ (topical) 1 gmL oil Application on cancerous andprecancerous human skin lesions
No local or systemic side effects38 patients withpositive responses
[51]
CAMchorioallantoic membrane of chicken embryoHUVEC human umbilical vein and endothelial cells TBrJA synthetic 5 7 9 10-tetrabromo jasmonate
mentioned that MJ was also assessed and approved by Foodand Agriculture OrganizationWorld Health Organization(FAOWHO) as a food additive Toxicity results reportedin the EPA document [52] were found to agree with thosepreviously published by other groups [50 51]
21 Jasmonates (MJ) Can Cause Cancer Cell Cycle Arrest atDifferent Phases in Different Cancer Cell Types Inhibiting
Growth and Proliferation The eucaryotic cell cycle is dividedinto four phases G
1 S G
2 and M that occur in response
to growth factors or mitogens The DNA synthetic (S) andmitotic (M) phases are preceded by gap phases (G
1 G2)
Chromosome duplication occurs during S phase replicatedchromosomes are segregated into individual nuclei (mitosis)during M phase and the cell then splits in two Cell cycleprogression can be regulated atG
1 G2points by various intra-
cellular and extracellular signals If extracellular conditions
6 International Journal of Cell Biology
Table 3 Effect in vitro and in vivo of methyl jasmonate (MJ) combined with other anticancer agents
Jasmonate Drug concrangevehicle Model Effects References
(1) MJ + BCNU(nitrosourea)in vitro
MJ Fixed conc Pancreatic MIA PaCa-1 Mitochondriotoxicsynergic cytotoxicity
[170](2) MJ + 2DGadriamycin taxolBCNU or cisplatinin vitro
MJ05ndash20mMEtOHTaxol cisplatin1ndash10120583gmL
CT26 DA-3 GTRAMP C1 MCF7MIA PaCa-2 D122 and BCL1
Strong cooperativeeffects of MJ + 2DGand MJ + other drugs
(3) MJ + adriamycinin vivo
MJ iv20ndash150mgkgdissolved inlipofundinAdr (DOX) ip4mgkg
Balbc mice injected ip with 1 times 104chronic BCL1 cells lymphocyteleukemia cells
MJ + Adr significant prolongedsurvival effect
(4) MJ + PI3KAktinhibitorsin vitro
MJ 05ndash30mMEtOHDMSO
Sarcoma MCA-105 SaOS-2 darrMJ-induced activation of Aktuarrsynergic cytotoxicity
[49](5) MJ + 2DGin vitro
MJ 05ndash30mMEtOHDMSO Sarcoma MCA-105 SaOS-2 2DG darrpAkt darrMJ-ind pAkt
MJ + 2DG uarrsynergic cytotoxicity
(6) MJ + irradiationin vitro
05ndash20mMDMSO
Irradiated prostate PC-3 (radiationinduces Bcl-2 expression)
darrradiation-induced Bcl-2uarrradiation sensitivity PC-3uarrcaspase-3
[104]
(7) MJ andor POHandorcis-platin (CP)in vitro
Both testedat IC20
Human MDA-MB-435[74]
MJ + POH uarrTNFR1darrΔ120595119898uarrcytotoxicity cell cycle arrest
at G0G1 uarrapoptosis+ CP all effects enhanced
[251]
Human breastMDA-MB-231 uarrapoptosis
(8) MJ + TRAILin vitro
MJ 05mM +TRAIL(100ndash200 ngsdotmLminus1)
CRC cancer cells
darrsurvivin (IAP)darrWntTCF pathwayuarrTRAIL-induced apoptosisuarrcaspase activity
[32]
(9) MJ + Smacin vitro
MJ 05ndash20mMDMSO
Human bladder cancer EJ T24 Synergy uarrIAPs-bound caspase 3uarrapoptosis [254]
Human embryonic kidney HEK293 No cytotoxicity
(10) MJ + Smac7N(IAP antagonist)in vitro
MJ 05ndash20mMDMSO
Hormone-independent prostateDU-145 PC-3Human proximal tubular epithelialHK-2 cells (overexpressing IAPs)
Smac7N uarrMJ-induced apoptosis bycaspase-9-dependent (intrinsic) andindependent (extrinsic) pathways
[255]
(11) MJ + cisplatin(01ndash05 120583M)MJ + X-rays(025ndash3Gy)MJ + 120572-rays
MJ 01ndash10mMEtOH
Cervical cancer cellsSiHa CaSki HeLa and C33A
darr viabilitydarrsurvivaldarrIC50 radiation dose
[29]
(12) MJ + 5-FUin vitro
05mMMJ1 hthen + 5-FU
Human adenocarcinomacolon HT-39 darrIC50 5-FU (5 rarr 25mM) [31]
BCNU 13-bis-(2-chloroethyl)-1-nitrosourea 2DG 2-deoxy-D-glucose 5-FU 5-fluorouracile IAP inhibitors of apoptosis MM multiple myeloma POHperillyl alcohol smac second mitochondria-derived activator of caspases Smac7N a peptide that contains the N-terminal seven residues of smac TNFR1tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand
are unfavorable cells delay progress through G1and may
enter a quiescent state known asG0 in which they can remain
for days weeks or years before resuming proliferation Ifextracellular conditions are favorable and signals to growand divide are present cells in early G
1or G0progress
through a restriction checkpoint late in G1after which cells
are committed to DNA replication even if the extracellularsignals that stimulate cell growth and division are removed[53 54] Cell cycle activity is modulated by intracellularROS levels and regulates cell survival cell proliferation andapoptosis [53ndash58]The progression through theG
1 S G2 and
M phases is promoted by cyclin-dependent kinases (CDKs)
International Journal of Cell Biology 7
which are positively and negatively regulated by cyclins andcyclin kinase inhibitors (CKis) respectively and by ROSUnlike the passage through the S G
2 and M phases G
1
progression normally relies on stimulation by mitogens andcan be blocked by antiproliferative factors passage throughthe restriction checkpoint and entry into S phase is controlledby CDKs that are sequentially regulated by cyclins D Eand A [53 54 56ndash58] In general CDK activity requirescyclin binding and depends on both positive and negativeregulatory phosphorylations [54] ROS levels influence thephosphorylating and ubiquitinating enzyme activites thuscontrolling cell cycle progression [54 57 58]
A master gene regulating cell passage through the cycleis the Myc family of transcription factors it switches onthe simultaneous synthesis of thousands of different proteinsrequired for cell growth and cell cycle function [58 59] TocounteractMyc protein activity the cell cycle is also regulatedby the p53 protein [60 61] a DNA-binding transcriptionfactor that senses DNA damage (occurring after exposureto ionizing radiation UV light oxidative stress or otherDNA damaging agents) and activates genes to halt cell cycleprogression in G
1for repair or otherwise initiates apoptosis
[56 62] When the damage caused by cell stress is mild p53activatesmechanisms for cell cycle arrest and repair damagedDNA if the damage is irreparable p53 initiates apoptosis [63]In case ofmild cellular stress low levels of p53 can activate theTP53-induced glycolysis and apoptosis regulator (TIGAR)gene that decreases the rate of glycolysis and hence ROS levelsin the cell [64] Glycolysis is essential for cancer progressionbut TIGAR provides protection from ROS and apoptosisHowever if damage caused by stress is irreparable p53prevents proliferation of cells with altered DNA and inducesincreased levels of ROS to trigger cell death [56] Increasedlevels of active p53 inhibit the cell entry into the S-phaseor induce apoptosis [56] Loss or inactivation of p53 resultsin loss of cell-cycle arrest or inhibition of apoptosis afterDNAdamage or physiological stress [65] leading to increasedgenetic instability increased accumulation of mutations andultimately oncogenesis [61] When the cell cycle is no longerregulated by p53 cells remain cycling (supported by 119888-Mycoverexpression) subverting cell cycle exit maturation andterminal differentiation [66] Over 50 of all human cancersharbor mutations and genetic alterations in cyclins and p53genes directly affecting the function of critical cell cycleproteins rendering the p53 gene ineffective in mediating cellgrowth arrest and promoting cancer cell death [62 67 68]ROS production is under p53 regulation and in turn p53being a redox-sensitive protein is influenced by ROS levelsthus ROS can act as both an upstream signal that triggersp53 activation and as a downstream factor that mediatesapoptosis [69]Thus mutations in the p53 gene downregulateits activity and consequently ROS production [70]ThroughROS p53 can also directly control metabolic traits of cells[71] regulate mitochondrial membrane potential (Δ120595
119898) and
induce cytochrome 119888-independent apoptosis (blocked by Bcl-2) ROS-mediated disruption of Δ120595
119898constitutes a pivotal
step in the apoptotic pathway of p53 and this pathwaydoes not involve cytochrome 119888 release [72] ROS can thusregulate cell fate through p53 in a way that physiological ROS
levels trigger cell protective pathways while under cytotoxicoxidative stress p53 behaves more like a cell killer [70 73]
JAs (mainly MJ) have been found to arrest cell cycle ofdifferent cancer cell types at different phases (Table 1) G
0G1
in human acute lymphoblastic leukemia Molt-4 [14 38] andhuman neuroblastoma SK-N-SH BE(2)-C [45] G
0G1-S in
human breast MCF-7 andmelanocytic MDA-MB-435 cancercells [25 74] S in human neuroblastoma BE(2)-C [43] S-G2M in human adenocarcinoma colon HT-39 [31] G
2M
in nonsmall cell lung cancer (NSCLC) lines A549 and H520[37] human neuroblastoma SH-SY5Y [44] human cervicalcarcinoma and HeLa cells [27] At 01mM MJ arrested invitro human umbilical vein endothelial cells (HUVEC) at G
1
[40]The molecular mechanisms underlying the above jasmo-
nate-mediated cell cycle arrest in cancer and embryonicHUVEC cells in vitro are not yet completely understoodChecking the above data against the UMD TP53 mutationdatabase (httpwwwp53fr) and revising the article byBerglind et al [75] we could not find correlation between agiven MJ-mediated phase arrest of cell cycle and the TP53wild type or mutated status of a cancer cell type (data notshown) a result that would agree with the original observa-tion of Fingrut and Flescher [15] stating thatMJ-mediated celldeath is independent of the p53 status [17] However otherstudies suggest thatMJ could be acting by interfering differentcell cycle regulatorymechanisms In plants MJ is able to bindthe TGACCG motif which is present in cell cycle cyclin pro-moters [76] wemight speculateMJ recognition of TGACCG-like motifs in mammalian cells cyclin promoters howeverthis would need experimental confirmation In some plantsMJ impairs the G
2-to-M transition by repressing M-phase
gene activation [77] MJ impaired the G2-to-M transition in
the human neuroblastoma SH-SY5Y cells downregulatingthe expressions of the proliferating cell nuclear antigen(PCNA a processivity factor encircling DNA at sites ofreplication and repair) andN-Myc [45] Asmentioned aboveMYC is the master gene allowing cancer cells to move alongthe cell cycle and regulating most genes needed to meet themetabolic demands for cell proliferation and survival [78]Cancer cell cycle arrest at different phases is in itself closelydependent on the cell energy and as such it is extremelydependent on metabolic ATP availability and its use byhexokinase Downregulation of Myc byMJ [45] is interestingresult that must be confirmed particularly because there is alack of specific inhibitors against Myc [57]
In some cases of cell cycle arrest MJs have been foundto upregulate p21 [37] whereas in others to downregulatesurvivinmRNA and protein levels [43ndash45] (Table 1) Survivinis an important member of the inhibitor of apoptosis protein(IAP) family that also regulates entrance of the cell cycle to theM phase and is overexpressed in cancer cells acting linkingboth regulation of the cell cycle and inhibition of apoptosis[79]
22 Jasmonates (MJ) CanCauseCancer Cell Death by InducingApoptosis andor Necrosis Apoptosis can be initiated by twosignaling cascades namely the extrinsic and the intrinsic
8 International Journal of Cell Biology
pathway The classical extrinsic pathway is triggered by thebinding of extracellular signals like tumor necrosis factor(TNF) tumor necrosis factor-related apoptosis-inducing lig-and (TRAIL) and apoptosis antigen-1 (Fas) ligand (FASL)to death receptors such as tumor necrosis factor receptor 1(TNFR1) and FAS [80]This interaction leads to procaspase-8activation which cleaves itself into caspase-8Then caspase-8can cleave and activate procaspase-3 into caspase-3 thatcleaves many substrates inside the cell inducing the typicalmorphological changes seen in apoptosis The intrinsic path-way can be triggered by intracellular stress signals such asoxidative stress and DNA damage The diverse signals aresensed by mitochondria through the balance between pro-and antiapoptotic members of the Bcl-2 family of proteinsWhen proapoptotic signals overcome antiapoptotic signalsmitochondria becomepermeabilized leading to dissipation ofthe mitochondrial electrochemical potential release of inter-membrane proteins like cytochrome 119888 SmacDIABLO AIFandHTRA2OMI Cytochrome 119888 interacts with procaspase-9and Apaf-1 to form the apoptosome complex that leads tocaspase-3 activation [81] The extrinsic pathway can connectwith the intrinsic pathway through the cleavage by caspase-8of Bid into tBid (truncated Bid) which is a proapoptoticmember of the Bcl-2 family and can lead to mitochondriapermeabilization [82 83] The members of the Bcl-2 fam-ily regulate mitochondria permeabilization and comprisesantiapoptotic (Bcl-2 Bcl-xL) proapoptotic (Bax Bak) andproapoptotic BH-3 only proteins (Bad Bid Puma andNoxa)[82ndash84] One of the mechanisms of mitochondria perme-abilization is controlled by Bax and Bak proteins that arenormally expressed in every cell but are maintained inhibitedby the antiapoptotic members When there is a signal toapoptosis BH3-only proteins interact with the antiapoptoticmembers releasing BaxBak or directly activating themleading to their oligomerization and to mitochondrial outermembrane permeabilization (MOMP) Another mechanismfor mitochondria permeabilization is the formation of themitochondrial permeability transition (MPT) pore which iscomposed by the voltage-dependent anion channel VDACthe adenine nucleotide translocase (ANT) and cyclophilin D[85]TheMPT pore comprising VDAC1 is directly regulatedby Bad [86] In contrast VDAC2 has been identified as anantiapoptotic negative regulator of Bak [87ndash89]
One of the hallmarks of cancer is resistance to cell death[90] This resistance is acquired by a host of different geneticdefects in cancer cells such as those present in the p53tumor suppressor gene [62 68 91] Normal p53 responds togenotoxic oncogenic and other stress signals by inducingantiproliferative transcriptional programs leading to growtharrest (for cell repair) or apoptosis [92] p53 mediates cyclearrest by transcriptionally activating the p21 (CDKN1A) 14-3-3120590 (SFN) and GADD45120572 genes whereas p53-dependentapoptosis is triggered by turning on the synthesis of proteinsthat produce ROS [58 93] and transactivating differentmitochondrial proapoptotic genes as well as death receptorsfor the synthesis of the corresponding proteins [58 94]Whenp53 (and its target genes) expression levels are below a thresh-old they induce cell cycle arrest but not apoptosis above thisthreshold p53 trigger apoptosis [95] Lowering this threshold
with inhibitors of the antiapoptotic Bcl-2 family proteinssensitize cells to p53-induced apoptosis [95] By damagingcells chemotherapeutical drugs turn on p53rsquos transcriptionalfunction that upregulates the expression of proapoptotic pro-teins Defects in the p53 function or in the apoptotic cascadeslead to cell phenotypes resistant to chemotherapy Apoptosiscan also be induced by anticancer agents independently ofp53 by either directly modulating the apoptotic machineryor directly interacting with mitochondria [96] ROS levelsmight be playing a central role in mediating various forms ofnonapoptotic caspase-independent programmed cell deatheither through direct irreversible oxidative damage to keyproteins nucleic acid molecules and cellular structure or bythe activation of prodeath signal transduction programs [97]In a ROS-dependent nonapoptotic cell death pathway respi-ration through the mitochondrial electron transport system(ETS) chain is not essential and ROS is generated throughNADPH oxidases [97] In contrast to the mitochondrial ETSwhich generates ROS as a byproduct of respiration NADPHoxidases produceROS as their primary function [98] Raise ofROS is characteristic after ionizing radiation chemotherapyand targeted therapy treatments [58 99] and may be killingcancer cells through a nonapoptotic ROS-dependent celldeath pathway
Tumor cells maintain high ATPADP as well asNADHNAD+ ratios so as to assure that ATP levels arenever limited ATP depletion would represent then anunsustainable metabolic stress for tumor cells and it isassociated with necrotic death in some cell types [100]Recently it was shown that a signaling cascade centered onthe proteins RIP1 and RIP3 can trigger cell death by necrosisin many situations and the RIP1-RIP3 complex appears topromote ATP depletion during necroptosis by impingingon components of the MPT pore [101 102] Necrosis was aterm used to describe a process of accidental cell death inwhich cells swell and burst releasing its intracellular contentwhereas necroptosis modernly refers rather to an orderedcell death program different from apoptosis apoptosis is acaspase-dependent cell death process whereas necrosis iscaspase-independent process [102 103] Studies concerningnecrosis have focusedmainly on cell membrane permeabilityto propidium iodide (PI) in the flow citometry doublestaining with annexin-VPI
Fingrut et al [17] by studying the effect of MJ ontwo clones of a highly malignant B-cell lymphoma (oneharboring a normal wild type (wt) p53 gene while the otherexpressed a mutated inactive p53 protein) (Table 1) reportedthat MJ was equally toxic to both clones [17] In contrastthe p53-mutant cells were resistant to radiomimetics (drugshaving effects similar to those produced in radiotherapy)and chemotherapeutic drugs In this cancer cell model MJinduced mostly apoptotic cell death in the wt p53-expressinglymphoma cells while no signs of early apoptosis weredetected in the mutant p53-expressing lymphoma cells thatwere killed by MJ through a p53-independent nonapoptoticcell death mechanism However other cancer cell types canundergo apoptosis with or without functional p53 and aswe will see MJ can induce apoptotic andor nonapoptoticcell death in wt p53 as well as in TP53 mutated cells
International Journal of Cell Biology 9
(Table 1) Nonapoptotic cell death could be the consequenceof the severe bioenergetic damages induced by the directinteraction of MJ with mitochondria leading to increasedROS levels inhibition of ATP synthesis and ATP depletion[20 30] Jasmonates including MJ and some of its analogshave been found to be strongly proapoptotic in vitro indifferent types of cancer cells by both the extrinsic and theintrinsic pathway (Table 1) with no or little cytotoxicity tonormal cells [15 25 36 37 44 45] Thus MJ induced theexpression of TNFR1 in human breast MCF-7 and MDA-MB-435 cells [25 74] and in hormone-independent humanprostate PC-3 and DU 145 cancer cells promoting apoptosisby the extrinsic pathway in these cells [48] TRAIL (TNF-related apoptosis inducing ligand) is able to induce apoptosisin prostate cancer cell yet overexpression of antiapoptoticproteins and inhibition of proapoptotic proteins results indiminishing the TRAIL-mediated apoptosis pathway TRAILhas emerged as a potent anticancer agent in laboratory-based studies and preclinical trials and JAs have been foundto sensitize cancer cells to TRAIL-mediated apoptosis [32](Table 3) J7 a synthetic analogue of MJ enhanced TRAIL-mediated apoptosis through upregulation of ROS levels inhuman hepatomaHepG2 cells apoptosis was induced via Bidcleavage downregulation of X-linked inhibitor of apoptosisprotein (XIAP) cellular inhibitor of apoptosis-1 (cIAP-1)B-cell lymphoma-extra large (Bcl-xL) and activation ofcaspases [35] Other examples of MJ-mediated apoptoticandor nonapoptotic cell death in different cancer cell types(Table 1) are as follows (1) MJ induced ROS production inA549 lung adenocarcinoma cells activating Bcl-2-associated-X-protein (Bax) as well as Bcl-2-Like-1 (Bcl2L1) proapoptoticproteins in these cells [36] (2) MJ activated caspase-3 inleukemia Molt-4 prostate SK28 and LNCaP cancer cells[15] (3) MJ inhibited the antiapoptotic Bcl-2 protein andenhanced proapoptotic proteins in radio-resistant humanprostate cancer cell line PC-3 [104] (4) MJ suppressed theproliferation of the neuroblastoma cell lines SH-SY5Y SK-N-SH and BE(2)-C through a strong apoptosis-inducingeffect as measured by annexin VFITCPI MJ treatment alsodownregulated the expression of the antiapoptotic proteinsXIAP and survivin [44 45] (5) MJ-mediated cell death wasobserved on cervical cancer and myeloma cells and it wasreported to be due to mixed characteristics of apoptosis andnecrosis [26 105] So far studies showing MJ inducing theRIP1 signaling cascade have not been reported howeverit will be important to deep into these analyses to cleardistinguish between MJ-induced apoptosis andor necrosis
23 Methyl Jasmonate Induces the Production of ReactiveOxygen Species (ROS) Responses in Some Cancer Cell TypesLeading Also to Cell Death Intracellular ROS levels reg-ulate cell cycle activity cell metabolism and survival cellproliferation and apoptosis [53ndash58] Stressful conditionsproduce a transient excess of intracellular ROS in normaltissues however under chronic pathological and oxidativestress conditions ROS is continuously produced in excessand persistently causing damage to DNA cell membraneslipids proteins and inducing apoptosis to control and avoid
oxidative damages cells counteract excess ROS by enzymatic(superoxide dismutase (SOD) glutathione peroxidase (GPx)and catalase) andor nonenzymatic (glutathione thoredoxin)antioxidant elements [99 106] Controlling ROS is central tointracellular redox signaling and homeostasis on the otherhand uncontrolledunbalanced ROS production modifiesintracellular redox leading to apoptosis [58 82 99 106] ROScan be generated by different biological systems (1) director indirect extracellular stimulation of plasma membrane-bound NAD(P)H oxidases (NOX) can generate intracellularH2O2[57 107ndash110] (2) biochemical pathways associated to
fatty acid oxidation and lipoxygenases activities generateROS as normal byproducts of these activities [111] (3)ATP generation by oxidative phosphorylation (OxPhos) inmitochondria is accompanied by the production of ROS as aconsequence of electron leakage from the electron transportchain mitochondria are considered to be major sources ofROS inmammalian cells particularly under a variety of stress-ful conditions [82 99] Superoxide (O
2
minus) is mainly produced
from complexes I and III and it is rapidly dismutated to H2O2
and oxygen by SOD [112] H2O2serves both as a toxic oxidant
and as an essential signaling molecule regulating cellularbiological processes [113] A strong positive correlation existsbetween mitochondrial membrane potential (Δ120595
119898) and ROS
production [106 114 115] Mitochondria produce more ROSat high membrane potential and an increase in the Δ120595
119898
produced either by a closure of MPT pore or by inhibitionof the ATP synthase is associated with increased ROS pro-duction However in cases of mitochondrial dysfunctionslow Δ120595
119898and decreased activity of the respiratory chain
are accompanied by a simultaneous increase in ROS [116117] It has been reported that cancer cells have higher ROScontent compared to normal cells [118 119] probably due toabnormal respiration because of dysfunctional mutations inmitochondria [120] but they also have increased contents ofantioxidants (SOD catalase glutathione thioredoxin etc)to counteract ROS [58 121] Tumor cells cannot tolerateexcessive high ROS levels [118 122] and as a matter of factradiation therapy chemotherapies and targeted therapiesinduce apoptosis and kill some cancer cells by generatingexcess ROS [99 106 123 124] Thus inducing higher levelsof ROS in cancers cells can be exploited as an effectiveselective strategy to kill tumor cells over normal cells [119]However prolonged treatment with the same drug reducesROS levels in cancer cells [124] indicating that when theybecome drug-resistant tumor cells have lower ROS contentthan drug-sensitive cancer cells Evidence suggests that drug-resistant cells have a higher expression of catalase at theplasma membrane that keeps reduced ROS levels [121]
In plants MJ-mediated damage to mitochondria is fol-lowed by a rapid production of H
2O2[125] H
2O2is used as
a second messenger for the induction of genes and productsrelated to defense against both herbivores and pathogens[126] in addition H
2O2induces apoptosis and senescence
[127 128] On the other hand it has been signaled that theMJ-induced production of H
2O2in plants was prevented by
inhibitors of NAD(P)H oxidase [128 129] indicating anothermechanism of MJ-mediated production of ROS and a majorrole for this enzyme in MJ-induced ROS responses MJ
10 International Journal of Cell Biology
induces also an endogenous H2O2response in some cancer
cell lines (Table 1) [19 36 38 130ndash132] In A549 cancerlung cells the H
2O2response induced by MJ increased the
expression of the proapoptotic Bax and Bcl-Xs (although notbeing that of the antiapoptotic Bcl-2 and Bcl-xL proteins) andthis led to apoptosis this event was inhibited by catalase (aspecific inhibitor of H
2O2) [36] MJ increased the intracel-
lular levels of H2O2and superoxide (O
2) ions in human C6
glioma cells in vitro and induced the expression of heat shockprotein 72 (HSP72) a negative regulator of H
2O2-induced
oxidative stress in these cells in vitro MJ-induced HSP72expression was blocked by ROS inhibitors [130] ElevatedROS generation was observed in response to both MJ andJA levels in the acute myeloid leukemia HL-60 and KG1a celllines but other factors like the MJ-mediated generation ofmitochondrial SOD rather than ROS were determinant forapoptosis or differentiation in these cells [132] Suppression ofMJ-induced apoptosis by antioxidants like N-acetylcysteine(NAC) and catalase but not by inhibitors of hydroxyl radicalsand superoxide ions indicates that H
2O2is nonetheless one
of the important factors involved inMJ anticancer effects [19]
24 By Dissociating Hexokinase (HK) from the VoltageDependent Anion Channel (VDAC) on the Outer Mitochon-drial Membrane MJ Dissociates Glycolysis from OxidativePhosphorylation Causing Severe Bioenergetic Deregulations inCancer Cells Metabolism and cell growth are two cellularprocesses that are tighly linked and regulated [133] Theclassic bioenergetics phenotype of cancer cells of enhancedglycolysis under aerobic conditions was described by War-burg [134] However the ldquoWarburg effectrdquo does not neces-sarily imply mitochondrial dysfunction [135] To increaseglycolysis cancer cells upregulate the transcription of genesinvolved in the glycolytic pathway (ie glucose transportersglycolytic enzymes etc) This metabolic reprogrammingallows increased rate of ATP production synthesis of lipidsand a new redox balance Cancer cells actually use a com-bination of both glycolysis and mitochondrial respirationto produce energy and they may vary in regards to thepreferential use of these pathways being in some caseseither more glycolyticless oxidative or less glycolyticmoreoxidative depending on the prevalent normoxic or hypoxicenvironmental conditions and their capacities to expressadequate levels of oncogenes and tumor suppressor geneproducts for cell growth [136ndash139] or when either glycolysisor OxPhos is inhibited in whose case there can be partialcompensation by the other metabolic pathway [17] Shiftingthe balance between those two processes with glycolyticandor mitochondrial enzyme inhibitors might represent aninteresting way to look for new anticancer agents
The first step in the metabolism of glucose is catalyzedby the enzyme hexokinase (HK) which by phosphorylatingglucose makes it negative sequestering it into the cytoplasmin the form of glucose-6-phosphate (G6P) In normal tissuesthis crucial step is catalyzed by four different HK isoforms(HK1 HK2 HK3 and glucokinase) indicating that regulationof glucose phosphorylation can vary in different tissues underdifferent condition [140] HK1 and HK2 have overlapping
tissue expression but different subcellular distributions withHK1 associated mainly with mitochondria and HK2 associ-atedwith bothmitochondrial and cytoplasmic compartments[141] HK1 is widely and constitutively expressed whereasHK2 predominates during embryogenesis and is not widelyexpressed in adult tissues but only in a limited numberof normal adult tissues [141] their different subcellulardistributions and kinetic properties reflect their differentmetabolic roles [141 142] Under physiological conditionsHK1 is predominantly bound to the outer mitochondrialmembrane where it primarily channels glucose toward gly-colysis [140] whereas HK2 is mainly soluble and controlsglycogen formation [140 141] In malignant tissues HK1and HK2 isoforms are glycolytic and overexpressed (HK1more than HK2) (httpwwwproteinatlascom) [143] andare tightly bound to mitochondria HK1 and HK2 bindthrough theirN-terminal hydrophobic regions to the voltage-dependent calcium channel isoform1 (VDAC1) the prevalentpore protein in the outer mitochondrial membrane [2482 143ndash146] VDAC1 is also overexpressed in cancer cells[147] and the binding of HK to VDAC1 on the outer mito-chondrial membrane is a fundamental aspect of the aerobicglycolytic metabolism of cancer cells [82 135 147 148]HKVDAC association appears to protect tumor cells frommitochondrial outer membrane permeabilization (MOMP)which marks a point of no return leading to cell death[149] VDAC transports ADP and inorganic phosphate (Pi)the substrates needed for the production of ATP into themitochondria it also transports ATP out of themitochondriainto the cytoplasm and facilitates the availability of ATPto HK2 [143 150 151] A continuous supply of glucose toHK maintains this activity at a constant level and controlsthe flux through the mitochondrial electron transport chaininfluencing mitochondrial ROS production [152] HK bindsthrough hydrophobic interaction to VDAC1 the prevalentpore protein in the outer mitochondrial membrane [24 82143ndash146] thereby assuring and gaining a preferential accessto ATP [82 150 153] Translocation of HK2 from the cytosolto the mitochondria is regulated through phosphorylationof HK by Akt [141] HK2 only binds VDAC after phos-phorylation by Akt [151 152] Glycogen synthase kinase 3beta (GSK3120573) a major target of Akt kinases phosphorylatesVDAC [154ndash157] and dissociates HK [156] HK2 boundto mitochondria enhance its affinity to ATP and is lesssensitive to enzymatic inhibition by the G6P product [140141 158] HK2 can produce G6P from cytoplasmic glucoseand mitochondrial ATP without regulatory restraints evenunder hypoxia supporting rapid cell growth [142 143 159ndash161]HK1 andHK2 aremore tightly bound toVDAC in cancercells than in nonmalignant cells [156 162 163] HKVDA1interaction is relatively stable for HK1 while it is dependentupon G6P levels for HK2 [140 141] The higher affinity ofmitochondrial HK2 for glucose ATP and VDAC1 representsa high advantage for cancer cells survival and growth con-ferring to them an aerobic glycolytic phenotype underlyingthe Warburg effect [82 135 138 142 147 148 150 160]Differential interactions of HK1 and HK2 with mitochondriamay underlie different glycolytic profiles in cancer cells [164165] As rate-controlling enzymes in the glycolyic pathway
International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
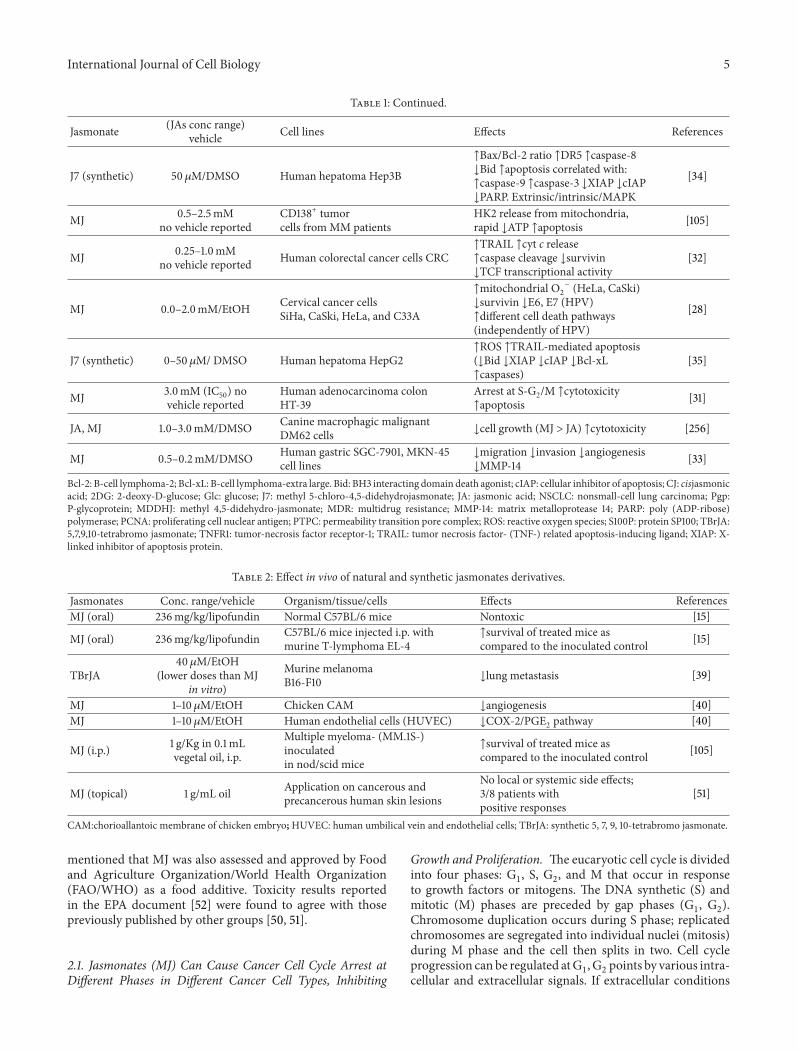
International Journal of Cell Biology 5
Table 1 Continued
Jasmonate (JAs conc range)vehicle Cell lines Effects References
J7 (synthetic) 50120583MDMSO Human hepatoma Hep3B
uarrBaxBcl-2 ratio uarrDR5 uarrcaspase-8darrBid uarrapoptosis correlated withuarrcaspase-9 uarrcaspase-3 darrXIAP darrcIAPdarrPARP ExtrinsicintrinsicMAPK
[34]
MJ 05ndash25mMno vehicle reported
CD138+ tumorcells from MM patients
HK2 release from mitochondriarapid darrATP uarrapoptosis [105]
MJ 025ndash10mMno vehicle reported Human colorectal cancer cells CRC
uarrTRAIL uarrcyt c releaseuarrcaspase cleavage darrsurvivindarrTCF transcriptional activity
[32]
MJ 00ndash20mMEtOH Cervical cancer cellsSiHa CaSki HeLa and C33A
uarrmitochondrial O2
minus (HeLa CaSki)darrsurvivin darrE6 E7 (HPV)uarrdifferent cell death pathways(independently of HPV)
[28]
J7 (synthetic) 0ndash50 120583M DMSO Human hepatoma HepG2uarrROS uarrTRAIL-mediated apoptosis(darrBid darrXIAP darrcIAP darrBcl-xLuarrcaspases)
[35]
MJ 30mM (IC50) novehicle reported
Human adenocarcinoma colonHT-39
Arrest at S-G2M uarrcytotoxicityuarrapoptosis [31]
JA MJ 10ndash30mMDMSO Canine macrophagic malignantDM62 cells darrcell growth (MJ gt JA) uarrcytotoxicity [256]
MJ 05ndash02mMDMSO Human gastric SGC-7901 MKN-45cell lines
darrmigration darrinvasion darrangiogenesisdarrMMP-14 [33]
Bcl-2 B-cell lymphoma-2 Bcl-xL B-cell lymphoma-extra large Bid BH3 interacting domain death agonist cIAP cellular inhibitor of apoptosis CJ cisjasmonicacid 2DG 2-deoxy-D-glucose Glc glucose J7 methyl 5-chloro-45-didehydrojasmonate JA jasmonic acid NSCLC nonsmall-cell lung carcinoma PgpP-glycoprotein MDDHJ methyl 45-didehydro-jasmonate MDR multidrug resistance MMP-14 matrix metalloprotease 14 PARP poly (ADP-ribose)polymerase PCNA proliferating cell nuclear antigen PTPC permeability transition pore complex ROS reactive oxygen species S100P protein SP100 TBrJA57910-tetrabromo jasmonate TNFR1 tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand XIAP X-linked inhibitor of apoptosis protein
Table 2 Effect in vivo of natural and synthetic jasmonates derivatives
Jasmonates Conc rangevehicle Organismtissuecells Effects ReferencesMJ (oral) 236mgkglipofundin Normal C57BL6 mice Nontoxic [15]
MJ (oral) 236mgkglipofundin C57BL6 mice injected ip withmurine T-lymphoma EL-4
uarrsurvival of treated mice ascompared to the inoculated control [15]
TBrJA40120583MEtOH
(lower doses than MJin vitro)
Murine melanomaB16-F10 darrlung metastasis [39]
MJ 1ndash10120583MEtOH Chicken CAM darrangiogenesis [40]MJ 1ndash10120583MEtOH Human endothelial cells (HUVEC) darrCOX-2PGE2 pathway [40]
MJ (ip) 1 gKg in 01mLvegetal oil ip
Multiple myeloma- (MM1S-)inoculatedin nodscid mice
uarrsurvival of treated mice ascompared to the inoculated control [105]
MJ (topical) 1 gmL oil Application on cancerous andprecancerous human skin lesions
No local or systemic side effects38 patients withpositive responses
[51]
CAMchorioallantoic membrane of chicken embryoHUVEC human umbilical vein and endothelial cells TBrJA synthetic 5 7 9 10-tetrabromo jasmonate
mentioned that MJ was also assessed and approved by Foodand Agriculture OrganizationWorld Health Organization(FAOWHO) as a food additive Toxicity results reportedin the EPA document [52] were found to agree with thosepreviously published by other groups [50 51]
21 Jasmonates (MJ) Can Cause Cancer Cell Cycle Arrest atDifferent Phases in Different Cancer Cell Types Inhibiting
Growth and Proliferation The eucaryotic cell cycle is dividedinto four phases G
1 S G
2 and M that occur in response
to growth factors or mitogens The DNA synthetic (S) andmitotic (M) phases are preceded by gap phases (G
1 G2)
Chromosome duplication occurs during S phase replicatedchromosomes are segregated into individual nuclei (mitosis)during M phase and the cell then splits in two Cell cycleprogression can be regulated atG
1 G2points by various intra-
cellular and extracellular signals If extracellular conditions
6 International Journal of Cell Biology
Table 3 Effect in vitro and in vivo of methyl jasmonate (MJ) combined with other anticancer agents
Jasmonate Drug concrangevehicle Model Effects References
(1) MJ + BCNU(nitrosourea)in vitro
MJ Fixed conc Pancreatic MIA PaCa-1 Mitochondriotoxicsynergic cytotoxicity
[170](2) MJ + 2DGadriamycin taxolBCNU or cisplatinin vitro
MJ05ndash20mMEtOHTaxol cisplatin1ndash10120583gmL
CT26 DA-3 GTRAMP C1 MCF7MIA PaCa-2 D122 and BCL1
Strong cooperativeeffects of MJ + 2DGand MJ + other drugs
(3) MJ + adriamycinin vivo
MJ iv20ndash150mgkgdissolved inlipofundinAdr (DOX) ip4mgkg
Balbc mice injected ip with 1 times 104chronic BCL1 cells lymphocyteleukemia cells
MJ + Adr significant prolongedsurvival effect
(4) MJ + PI3KAktinhibitorsin vitro
MJ 05ndash30mMEtOHDMSO
Sarcoma MCA-105 SaOS-2 darrMJ-induced activation of Aktuarrsynergic cytotoxicity
[49](5) MJ + 2DGin vitro
MJ 05ndash30mMEtOHDMSO Sarcoma MCA-105 SaOS-2 2DG darrpAkt darrMJ-ind pAkt
MJ + 2DG uarrsynergic cytotoxicity
(6) MJ + irradiationin vitro
05ndash20mMDMSO
Irradiated prostate PC-3 (radiationinduces Bcl-2 expression)
darrradiation-induced Bcl-2uarrradiation sensitivity PC-3uarrcaspase-3
[104]
(7) MJ andor POHandorcis-platin (CP)in vitro
Both testedat IC20
Human MDA-MB-435[74]
MJ + POH uarrTNFR1darrΔ120595119898uarrcytotoxicity cell cycle arrest
at G0G1 uarrapoptosis+ CP all effects enhanced
[251]
Human breastMDA-MB-231 uarrapoptosis
(8) MJ + TRAILin vitro
MJ 05mM +TRAIL(100ndash200 ngsdotmLminus1)
CRC cancer cells
darrsurvivin (IAP)darrWntTCF pathwayuarrTRAIL-induced apoptosisuarrcaspase activity
[32]
(9) MJ + Smacin vitro
MJ 05ndash20mMDMSO
Human bladder cancer EJ T24 Synergy uarrIAPs-bound caspase 3uarrapoptosis [254]
Human embryonic kidney HEK293 No cytotoxicity
(10) MJ + Smac7N(IAP antagonist)in vitro
MJ 05ndash20mMDMSO
Hormone-independent prostateDU-145 PC-3Human proximal tubular epithelialHK-2 cells (overexpressing IAPs)
Smac7N uarrMJ-induced apoptosis bycaspase-9-dependent (intrinsic) andindependent (extrinsic) pathways
[255]
(11) MJ + cisplatin(01ndash05 120583M)MJ + X-rays(025ndash3Gy)MJ + 120572-rays
MJ 01ndash10mMEtOH
Cervical cancer cellsSiHa CaSki HeLa and C33A
darr viabilitydarrsurvivaldarrIC50 radiation dose
[29]
(12) MJ + 5-FUin vitro
05mMMJ1 hthen + 5-FU
Human adenocarcinomacolon HT-39 darrIC50 5-FU (5 rarr 25mM) [31]
BCNU 13-bis-(2-chloroethyl)-1-nitrosourea 2DG 2-deoxy-D-glucose 5-FU 5-fluorouracile IAP inhibitors of apoptosis MM multiple myeloma POHperillyl alcohol smac second mitochondria-derived activator of caspases Smac7N a peptide that contains the N-terminal seven residues of smac TNFR1tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand
are unfavorable cells delay progress through G1and may
enter a quiescent state known asG0 in which they can remain
for days weeks or years before resuming proliferation Ifextracellular conditions are favorable and signals to growand divide are present cells in early G
1or G0progress
through a restriction checkpoint late in G1after which cells
are committed to DNA replication even if the extracellularsignals that stimulate cell growth and division are removed[53 54] Cell cycle activity is modulated by intracellularROS levels and regulates cell survival cell proliferation andapoptosis [53ndash58]The progression through theG
1 S G2 and
M phases is promoted by cyclin-dependent kinases (CDKs)
International Journal of Cell Biology 7
which are positively and negatively regulated by cyclins andcyclin kinase inhibitors (CKis) respectively and by ROSUnlike the passage through the S G
2 and M phases G
1
progression normally relies on stimulation by mitogens andcan be blocked by antiproliferative factors passage throughthe restriction checkpoint and entry into S phase is controlledby CDKs that are sequentially regulated by cyclins D Eand A [53 54 56ndash58] In general CDK activity requirescyclin binding and depends on both positive and negativeregulatory phosphorylations [54] ROS levels influence thephosphorylating and ubiquitinating enzyme activites thuscontrolling cell cycle progression [54 57 58]
A master gene regulating cell passage through the cycleis the Myc family of transcription factors it switches onthe simultaneous synthesis of thousands of different proteinsrequired for cell growth and cell cycle function [58 59] TocounteractMyc protein activity the cell cycle is also regulatedby the p53 protein [60 61] a DNA-binding transcriptionfactor that senses DNA damage (occurring after exposureto ionizing radiation UV light oxidative stress or otherDNA damaging agents) and activates genes to halt cell cycleprogression in G
1for repair or otherwise initiates apoptosis
[56 62] When the damage caused by cell stress is mild p53activatesmechanisms for cell cycle arrest and repair damagedDNA if the damage is irreparable p53 initiates apoptosis [63]In case ofmild cellular stress low levels of p53 can activate theTP53-induced glycolysis and apoptosis regulator (TIGAR)gene that decreases the rate of glycolysis and hence ROS levelsin the cell [64] Glycolysis is essential for cancer progressionbut TIGAR provides protection from ROS and apoptosisHowever if damage caused by stress is irreparable p53prevents proliferation of cells with altered DNA and inducesincreased levels of ROS to trigger cell death [56] Increasedlevels of active p53 inhibit the cell entry into the S-phaseor induce apoptosis [56] Loss or inactivation of p53 resultsin loss of cell-cycle arrest or inhibition of apoptosis afterDNAdamage or physiological stress [65] leading to increasedgenetic instability increased accumulation of mutations andultimately oncogenesis [61] When the cell cycle is no longerregulated by p53 cells remain cycling (supported by 119888-Mycoverexpression) subverting cell cycle exit maturation andterminal differentiation [66] Over 50 of all human cancersharbor mutations and genetic alterations in cyclins and p53genes directly affecting the function of critical cell cycleproteins rendering the p53 gene ineffective in mediating cellgrowth arrest and promoting cancer cell death [62 67 68]ROS production is under p53 regulation and in turn p53being a redox-sensitive protein is influenced by ROS levelsthus ROS can act as both an upstream signal that triggersp53 activation and as a downstream factor that mediatesapoptosis [69]Thus mutations in the p53 gene downregulateits activity and consequently ROS production [70]ThroughROS p53 can also directly control metabolic traits of cells[71] regulate mitochondrial membrane potential (Δ120595
119898) and
induce cytochrome 119888-independent apoptosis (blocked by Bcl-2) ROS-mediated disruption of Δ120595
119898constitutes a pivotal
step in the apoptotic pathway of p53 and this pathwaydoes not involve cytochrome 119888 release [72] ROS can thusregulate cell fate through p53 in a way that physiological ROS
levels trigger cell protective pathways while under cytotoxicoxidative stress p53 behaves more like a cell killer [70 73]
JAs (mainly MJ) have been found to arrest cell cycle ofdifferent cancer cell types at different phases (Table 1) G
0G1
in human acute lymphoblastic leukemia Molt-4 [14 38] andhuman neuroblastoma SK-N-SH BE(2)-C [45] G
0G1-S in
human breast MCF-7 andmelanocytic MDA-MB-435 cancercells [25 74] S in human neuroblastoma BE(2)-C [43] S-G2M in human adenocarcinoma colon HT-39 [31] G
2M
in nonsmall cell lung cancer (NSCLC) lines A549 and H520[37] human neuroblastoma SH-SY5Y [44] human cervicalcarcinoma and HeLa cells [27] At 01mM MJ arrested invitro human umbilical vein endothelial cells (HUVEC) at G
1
[40]The molecular mechanisms underlying the above jasmo-
nate-mediated cell cycle arrest in cancer and embryonicHUVEC cells in vitro are not yet completely understoodChecking the above data against the UMD TP53 mutationdatabase (httpwwwp53fr) and revising the article byBerglind et al [75] we could not find correlation between agiven MJ-mediated phase arrest of cell cycle and the TP53wild type or mutated status of a cancer cell type (data notshown) a result that would agree with the original observa-tion of Fingrut and Flescher [15] stating thatMJ-mediated celldeath is independent of the p53 status [17] However otherstudies suggest thatMJ could be acting by interfering differentcell cycle regulatorymechanisms In plants MJ is able to bindthe TGACCG motif which is present in cell cycle cyclin pro-moters [76] wemight speculateMJ recognition of TGACCG-like motifs in mammalian cells cyclin promoters howeverthis would need experimental confirmation In some plantsMJ impairs the G
2-to-M transition by repressing M-phase
gene activation [77] MJ impaired the G2-to-M transition in
the human neuroblastoma SH-SY5Y cells downregulatingthe expressions of the proliferating cell nuclear antigen(PCNA a processivity factor encircling DNA at sites ofreplication and repair) andN-Myc [45] Asmentioned aboveMYC is the master gene allowing cancer cells to move alongthe cell cycle and regulating most genes needed to meet themetabolic demands for cell proliferation and survival [78]Cancer cell cycle arrest at different phases is in itself closelydependent on the cell energy and as such it is extremelydependent on metabolic ATP availability and its use byhexokinase Downregulation of Myc byMJ [45] is interestingresult that must be confirmed particularly because there is alack of specific inhibitors against Myc [57]
In some cases of cell cycle arrest MJs have been foundto upregulate p21 [37] whereas in others to downregulatesurvivinmRNA and protein levels [43ndash45] (Table 1) Survivinis an important member of the inhibitor of apoptosis protein(IAP) family that also regulates entrance of the cell cycle to theM phase and is overexpressed in cancer cells acting linkingboth regulation of the cell cycle and inhibition of apoptosis[79]
22 Jasmonates (MJ) CanCauseCancer Cell Death by InducingApoptosis andor Necrosis Apoptosis can be initiated by twosignaling cascades namely the extrinsic and the intrinsic
8 International Journal of Cell Biology
pathway The classical extrinsic pathway is triggered by thebinding of extracellular signals like tumor necrosis factor(TNF) tumor necrosis factor-related apoptosis-inducing lig-and (TRAIL) and apoptosis antigen-1 (Fas) ligand (FASL)to death receptors such as tumor necrosis factor receptor 1(TNFR1) and FAS [80]This interaction leads to procaspase-8activation which cleaves itself into caspase-8Then caspase-8can cleave and activate procaspase-3 into caspase-3 thatcleaves many substrates inside the cell inducing the typicalmorphological changes seen in apoptosis The intrinsic path-way can be triggered by intracellular stress signals such asoxidative stress and DNA damage The diverse signals aresensed by mitochondria through the balance between pro-and antiapoptotic members of the Bcl-2 family of proteinsWhen proapoptotic signals overcome antiapoptotic signalsmitochondria becomepermeabilized leading to dissipation ofthe mitochondrial electrochemical potential release of inter-membrane proteins like cytochrome 119888 SmacDIABLO AIFandHTRA2OMI Cytochrome 119888 interacts with procaspase-9and Apaf-1 to form the apoptosome complex that leads tocaspase-3 activation [81] The extrinsic pathway can connectwith the intrinsic pathway through the cleavage by caspase-8of Bid into tBid (truncated Bid) which is a proapoptoticmember of the Bcl-2 family and can lead to mitochondriapermeabilization [82 83] The members of the Bcl-2 fam-ily regulate mitochondria permeabilization and comprisesantiapoptotic (Bcl-2 Bcl-xL) proapoptotic (Bax Bak) andproapoptotic BH-3 only proteins (Bad Bid Puma andNoxa)[82ndash84] One of the mechanisms of mitochondria perme-abilization is controlled by Bax and Bak proteins that arenormally expressed in every cell but are maintained inhibitedby the antiapoptotic members When there is a signal toapoptosis BH3-only proteins interact with the antiapoptoticmembers releasing BaxBak or directly activating themleading to their oligomerization and to mitochondrial outermembrane permeabilization (MOMP) Another mechanismfor mitochondria permeabilization is the formation of themitochondrial permeability transition (MPT) pore which iscomposed by the voltage-dependent anion channel VDACthe adenine nucleotide translocase (ANT) and cyclophilin D[85]TheMPT pore comprising VDAC1 is directly regulatedby Bad [86] In contrast VDAC2 has been identified as anantiapoptotic negative regulator of Bak [87ndash89]
One of the hallmarks of cancer is resistance to cell death[90] This resistance is acquired by a host of different geneticdefects in cancer cells such as those present in the p53tumor suppressor gene [62 68 91] Normal p53 responds togenotoxic oncogenic and other stress signals by inducingantiproliferative transcriptional programs leading to growtharrest (for cell repair) or apoptosis [92] p53 mediates cyclearrest by transcriptionally activating the p21 (CDKN1A) 14-3-3120590 (SFN) and GADD45120572 genes whereas p53-dependentapoptosis is triggered by turning on the synthesis of proteinsthat produce ROS [58 93] and transactivating differentmitochondrial proapoptotic genes as well as death receptorsfor the synthesis of the corresponding proteins [58 94]Whenp53 (and its target genes) expression levels are below a thresh-old they induce cell cycle arrest but not apoptosis above thisthreshold p53 trigger apoptosis [95] Lowering this threshold
with inhibitors of the antiapoptotic Bcl-2 family proteinssensitize cells to p53-induced apoptosis [95] By damagingcells chemotherapeutical drugs turn on p53rsquos transcriptionalfunction that upregulates the expression of proapoptotic pro-teins Defects in the p53 function or in the apoptotic cascadeslead to cell phenotypes resistant to chemotherapy Apoptosiscan also be induced by anticancer agents independently ofp53 by either directly modulating the apoptotic machineryor directly interacting with mitochondria [96] ROS levelsmight be playing a central role in mediating various forms ofnonapoptotic caspase-independent programmed cell deatheither through direct irreversible oxidative damage to keyproteins nucleic acid molecules and cellular structure or bythe activation of prodeath signal transduction programs [97]In a ROS-dependent nonapoptotic cell death pathway respi-ration through the mitochondrial electron transport system(ETS) chain is not essential and ROS is generated throughNADPH oxidases [97] In contrast to the mitochondrial ETSwhich generates ROS as a byproduct of respiration NADPHoxidases produceROS as their primary function [98] Raise ofROS is characteristic after ionizing radiation chemotherapyand targeted therapy treatments [58 99] and may be killingcancer cells through a nonapoptotic ROS-dependent celldeath pathway
Tumor cells maintain high ATPADP as well asNADHNAD+ ratios so as to assure that ATP levels arenever limited ATP depletion would represent then anunsustainable metabolic stress for tumor cells and it isassociated with necrotic death in some cell types [100]Recently it was shown that a signaling cascade centered onthe proteins RIP1 and RIP3 can trigger cell death by necrosisin many situations and the RIP1-RIP3 complex appears topromote ATP depletion during necroptosis by impingingon components of the MPT pore [101 102] Necrosis was aterm used to describe a process of accidental cell death inwhich cells swell and burst releasing its intracellular contentwhereas necroptosis modernly refers rather to an orderedcell death program different from apoptosis apoptosis is acaspase-dependent cell death process whereas necrosis iscaspase-independent process [102 103] Studies concerningnecrosis have focusedmainly on cell membrane permeabilityto propidium iodide (PI) in the flow citometry doublestaining with annexin-VPI
Fingrut et al [17] by studying the effect of MJ ontwo clones of a highly malignant B-cell lymphoma (oneharboring a normal wild type (wt) p53 gene while the otherexpressed a mutated inactive p53 protein) (Table 1) reportedthat MJ was equally toxic to both clones [17] In contrastthe p53-mutant cells were resistant to radiomimetics (drugshaving effects similar to those produced in radiotherapy)and chemotherapeutic drugs In this cancer cell model MJinduced mostly apoptotic cell death in the wt p53-expressinglymphoma cells while no signs of early apoptosis weredetected in the mutant p53-expressing lymphoma cells thatwere killed by MJ through a p53-independent nonapoptoticcell death mechanism However other cancer cell types canundergo apoptosis with or without functional p53 and aswe will see MJ can induce apoptotic andor nonapoptoticcell death in wt p53 as well as in TP53 mutated cells
International Journal of Cell Biology 9
(Table 1) Nonapoptotic cell death could be the consequenceof the severe bioenergetic damages induced by the directinteraction of MJ with mitochondria leading to increasedROS levels inhibition of ATP synthesis and ATP depletion[20 30] Jasmonates including MJ and some of its analogshave been found to be strongly proapoptotic in vitro indifferent types of cancer cells by both the extrinsic and theintrinsic pathway (Table 1) with no or little cytotoxicity tonormal cells [15 25 36 37 44 45] Thus MJ induced theexpression of TNFR1 in human breast MCF-7 and MDA-MB-435 cells [25 74] and in hormone-independent humanprostate PC-3 and DU 145 cancer cells promoting apoptosisby the extrinsic pathway in these cells [48] TRAIL (TNF-related apoptosis inducing ligand) is able to induce apoptosisin prostate cancer cell yet overexpression of antiapoptoticproteins and inhibition of proapoptotic proteins results indiminishing the TRAIL-mediated apoptosis pathway TRAILhas emerged as a potent anticancer agent in laboratory-based studies and preclinical trials and JAs have been foundto sensitize cancer cells to TRAIL-mediated apoptosis [32](Table 3) J7 a synthetic analogue of MJ enhanced TRAIL-mediated apoptosis through upregulation of ROS levels inhuman hepatomaHepG2 cells apoptosis was induced via Bidcleavage downregulation of X-linked inhibitor of apoptosisprotein (XIAP) cellular inhibitor of apoptosis-1 (cIAP-1)B-cell lymphoma-extra large (Bcl-xL) and activation ofcaspases [35] Other examples of MJ-mediated apoptoticandor nonapoptotic cell death in different cancer cell types(Table 1) are as follows (1) MJ induced ROS production inA549 lung adenocarcinoma cells activating Bcl-2-associated-X-protein (Bax) as well as Bcl-2-Like-1 (Bcl2L1) proapoptoticproteins in these cells [36] (2) MJ activated caspase-3 inleukemia Molt-4 prostate SK28 and LNCaP cancer cells[15] (3) MJ inhibited the antiapoptotic Bcl-2 protein andenhanced proapoptotic proteins in radio-resistant humanprostate cancer cell line PC-3 [104] (4) MJ suppressed theproliferation of the neuroblastoma cell lines SH-SY5Y SK-N-SH and BE(2)-C through a strong apoptosis-inducingeffect as measured by annexin VFITCPI MJ treatment alsodownregulated the expression of the antiapoptotic proteinsXIAP and survivin [44 45] (5) MJ-mediated cell death wasobserved on cervical cancer and myeloma cells and it wasreported to be due to mixed characteristics of apoptosis andnecrosis [26 105] So far studies showing MJ inducing theRIP1 signaling cascade have not been reported howeverit will be important to deep into these analyses to cleardistinguish between MJ-induced apoptosis andor necrosis
23 Methyl Jasmonate Induces the Production of ReactiveOxygen Species (ROS) Responses in Some Cancer Cell TypesLeading Also to Cell Death Intracellular ROS levels reg-ulate cell cycle activity cell metabolism and survival cellproliferation and apoptosis [53ndash58] Stressful conditionsproduce a transient excess of intracellular ROS in normaltissues however under chronic pathological and oxidativestress conditions ROS is continuously produced in excessand persistently causing damage to DNA cell membraneslipids proteins and inducing apoptosis to control and avoid
oxidative damages cells counteract excess ROS by enzymatic(superoxide dismutase (SOD) glutathione peroxidase (GPx)and catalase) andor nonenzymatic (glutathione thoredoxin)antioxidant elements [99 106] Controlling ROS is central tointracellular redox signaling and homeostasis on the otherhand uncontrolledunbalanced ROS production modifiesintracellular redox leading to apoptosis [58 82 99 106] ROScan be generated by different biological systems (1) director indirect extracellular stimulation of plasma membrane-bound NAD(P)H oxidases (NOX) can generate intracellularH2O2[57 107ndash110] (2) biochemical pathways associated to
fatty acid oxidation and lipoxygenases activities generateROS as normal byproducts of these activities [111] (3)ATP generation by oxidative phosphorylation (OxPhos) inmitochondria is accompanied by the production of ROS as aconsequence of electron leakage from the electron transportchain mitochondria are considered to be major sources ofROS inmammalian cells particularly under a variety of stress-ful conditions [82 99] Superoxide (O
2
minus) is mainly produced
from complexes I and III and it is rapidly dismutated to H2O2
and oxygen by SOD [112] H2O2serves both as a toxic oxidant
and as an essential signaling molecule regulating cellularbiological processes [113] A strong positive correlation existsbetween mitochondrial membrane potential (Δ120595
119898) and ROS
production [106 114 115] Mitochondria produce more ROSat high membrane potential and an increase in the Δ120595
119898
produced either by a closure of MPT pore or by inhibitionof the ATP synthase is associated with increased ROS pro-duction However in cases of mitochondrial dysfunctionslow Δ120595
119898and decreased activity of the respiratory chain
are accompanied by a simultaneous increase in ROS [116117] It has been reported that cancer cells have higher ROScontent compared to normal cells [118 119] probably due toabnormal respiration because of dysfunctional mutations inmitochondria [120] but they also have increased contents ofantioxidants (SOD catalase glutathione thioredoxin etc)to counteract ROS [58 121] Tumor cells cannot tolerateexcessive high ROS levels [118 122] and as a matter of factradiation therapy chemotherapies and targeted therapiesinduce apoptosis and kill some cancer cells by generatingexcess ROS [99 106 123 124] Thus inducing higher levelsof ROS in cancers cells can be exploited as an effectiveselective strategy to kill tumor cells over normal cells [119]However prolonged treatment with the same drug reducesROS levels in cancer cells [124] indicating that when theybecome drug-resistant tumor cells have lower ROS contentthan drug-sensitive cancer cells Evidence suggests that drug-resistant cells have a higher expression of catalase at theplasma membrane that keeps reduced ROS levels [121]
In plants MJ-mediated damage to mitochondria is fol-lowed by a rapid production of H
2O2[125] H
2O2is used as
a second messenger for the induction of genes and productsrelated to defense against both herbivores and pathogens[126] in addition H
2O2induces apoptosis and senescence
[127 128] On the other hand it has been signaled that theMJ-induced production of H
2O2in plants was prevented by
inhibitors of NAD(P)H oxidase [128 129] indicating anothermechanism of MJ-mediated production of ROS and a majorrole for this enzyme in MJ-induced ROS responses MJ
10 International Journal of Cell Biology
induces also an endogenous H2O2response in some cancer
cell lines (Table 1) [19 36 38 130ndash132] In A549 cancerlung cells the H
2O2response induced by MJ increased the
expression of the proapoptotic Bax and Bcl-Xs (although notbeing that of the antiapoptotic Bcl-2 and Bcl-xL proteins) andthis led to apoptosis this event was inhibited by catalase (aspecific inhibitor of H
2O2) [36] MJ increased the intracel-
lular levels of H2O2and superoxide (O
2) ions in human C6
glioma cells in vitro and induced the expression of heat shockprotein 72 (HSP72) a negative regulator of H
2O2-induced
oxidative stress in these cells in vitro MJ-induced HSP72expression was blocked by ROS inhibitors [130] ElevatedROS generation was observed in response to both MJ andJA levels in the acute myeloid leukemia HL-60 and KG1a celllines but other factors like the MJ-mediated generation ofmitochondrial SOD rather than ROS were determinant forapoptosis or differentiation in these cells [132] Suppression ofMJ-induced apoptosis by antioxidants like N-acetylcysteine(NAC) and catalase but not by inhibitors of hydroxyl radicalsand superoxide ions indicates that H
2O2is nonetheless one
of the important factors involved inMJ anticancer effects [19]
24 By Dissociating Hexokinase (HK) from the VoltageDependent Anion Channel (VDAC) on the Outer Mitochon-drial Membrane MJ Dissociates Glycolysis from OxidativePhosphorylation Causing Severe Bioenergetic Deregulations inCancer Cells Metabolism and cell growth are two cellularprocesses that are tighly linked and regulated [133] Theclassic bioenergetics phenotype of cancer cells of enhancedglycolysis under aerobic conditions was described by War-burg [134] However the ldquoWarburg effectrdquo does not neces-sarily imply mitochondrial dysfunction [135] To increaseglycolysis cancer cells upregulate the transcription of genesinvolved in the glycolytic pathway (ie glucose transportersglycolytic enzymes etc) This metabolic reprogrammingallows increased rate of ATP production synthesis of lipidsand a new redox balance Cancer cells actually use a com-bination of both glycolysis and mitochondrial respirationto produce energy and they may vary in regards to thepreferential use of these pathways being in some caseseither more glycolyticless oxidative or less glycolyticmoreoxidative depending on the prevalent normoxic or hypoxicenvironmental conditions and their capacities to expressadequate levels of oncogenes and tumor suppressor geneproducts for cell growth [136ndash139] or when either glycolysisor OxPhos is inhibited in whose case there can be partialcompensation by the other metabolic pathway [17] Shiftingthe balance between those two processes with glycolyticandor mitochondrial enzyme inhibitors might represent aninteresting way to look for new anticancer agents
The first step in the metabolism of glucose is catalyzedby the enzyme hexokinase (HK) which by phosphorylatingglucose makes it negative sequestering it into the cytoplasmin the form of glucose-6-phosphate (G6P) In normal tissuesthis crucial step is catalyzed by four different HK isoforms(HK1 HK2 HK3 and glucokinase) indicating that regulationof glucose phosphorylation can vary in different tissues underdifferent condition [140] HK1 and HK2 have overlapping
tissue expression but different subcellular distributions withHK1 associated mainly with mitochondria and HK2 associ-atedwith bothmitochondrial and cytoplasmic compartments[141] HK1 is widely and constitutively expressed whereasHK2 predominates during embryogenesis and is not widelyexpressed in adult tissues but only in a limited numberof normal adult tissues [141] their different subcellulardistributions and kinetic properties reflect their differentmetabolic roles [141 142] Under physiological conditionsHK1 is predominantly bound to the outer mitochondrialmembrane where it primarily channels glucose toward gly-colysis [140] whereas HK2 is mainly soluble and controlsglycogen formation [140 141] In malignant tissues HK1and HK2 isoforms are glycolytic and overexpressed (HK1more than HK2) (httpwwwproteinatlascom) [143] andare tightly bound to mitochondria HK1 and HK2 bindthrough theirN-terminal hydrophobic regions to the voltage-dependent calcium channel isoform1 (VDAC1) the prevalentpore protein in the outer mitochondrial membrane [2482 143ndash146] VDAC1 is also overexpressed in cancer cells[147] and the binding of HK to VDAC1 on the outer mito-chondrial membrane is a fundamental aspect of the aerobicglycolytic metabolism of cancer cells [82 135 147 148]HKVDAC association appears to protect tumor cells frommitochondrial outer membrane permeabilization (MOMP)which marks a point of no return leading to cell death[149] VDAC transports ADP and inorganic phosphate (Pi)the substrates needed for the production of ATP into themitochondria it also transports ATP out of themitochondriainto the cytoplasm and facilitates the availability of ATPto HK2 [143 150 151] A continuous supply of glucose toHK maintains this activity at a constant level and controlsthe flux through the mitochondrial electron transport chaininfluencing mitochondrial ROS production [152] HK bindsthrough hydrophobic interaction to VDAC1 the prevalentpore protein in the outer mitochondrial membrane [24 82143ndash146] thereby assuring and gaining a preferential accessto ATP [82 150 153] Translocation of HK2 from the cytosolto the mitochondria is regulated through phosphorylationof HK by Akt [141] HK2 only binds VDAC after phos-phorylation by Akt [151 152] Glycogen synthase kinase 3beta (GSK3120573) a major target of Akt kinases phosphorylatesVDAC [154ndash157] and dissociates HK [156] HK2 boundto mitochondria enhance its affinity to ATP and is lesssensitive to enzymatic inhibition by the G6P product [140141 158] HK2 can produce G6P from cytoplasmic glucoseand mitochondrial ATP without regulatory restraints evenunder hypoxia supporting rapid cell growth [142 143 159ndash161]HK1 andHK2 aremore tightly bound toVDAC in cancercells than in nonmalignant cells [156 162 163] HKVDA1interaction is relatively stable for HK1 while it is dependentupon G6P levels for HK2 [140 141] The higher affinity ofmitochondrial HK2 for glucose ATP and VDAC1 representsa high advantage for cancer cells survival and growth con-ferring to them an aerobic glycolytic phenotype underlyingthe Warburg effect [82 135 138 142 147 148 150 160]Differential interactions of HK1 and HK2 with mitochondriamay underlie different glycolytic profiles in cancer cells [164165] As rate-controlling enzymes in the glycolyic pathway
International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
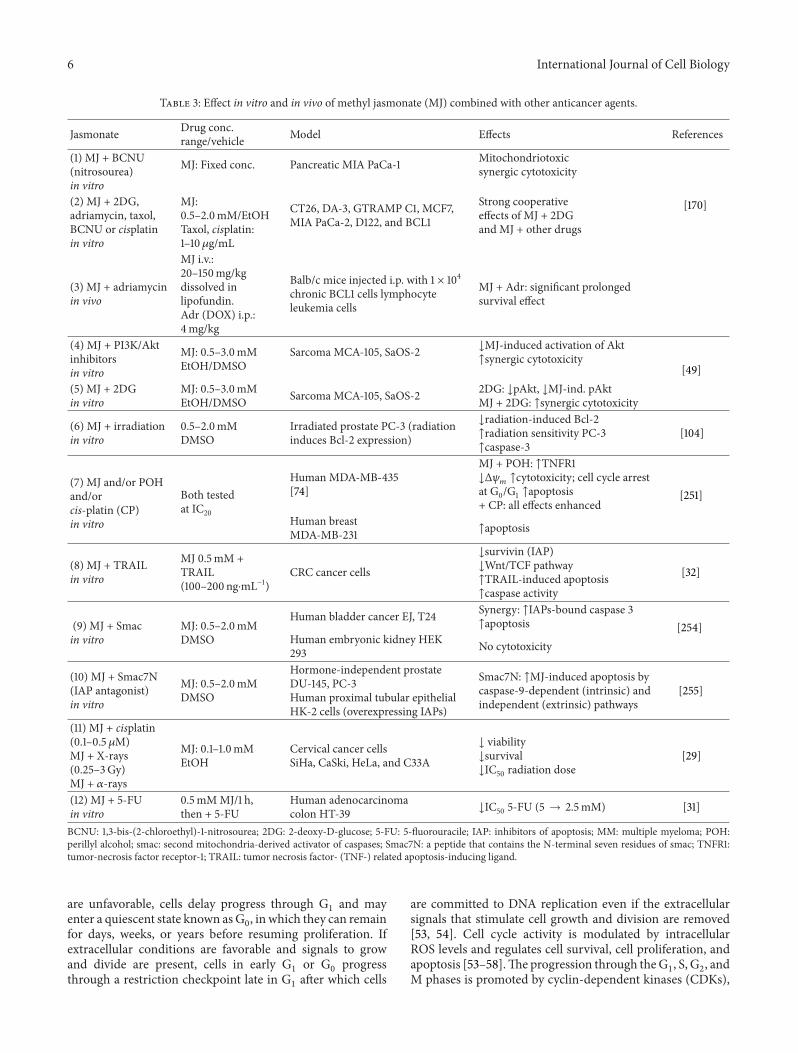
6 International Journal of Cell Biology
Table 3 Effect in vitro and in vivo of methyl jasmonate (MJ) combined with other anticancer agents
Jasmonate Drug concrangevehicle Model Effects References
(1) MJ + BCNU(nitrosourea)in vitro
MJ Fixed conc Pancreatic MIA PaCa-1 Mitochondriotoxicsynergic cytotoxicity
[170](2) MJ + 2DGadriamycin taxolBCNU or cisplatinin vitro
MJ05ndash20mMEtOHTaxol cisplatin1ndash10120583gmL
CT26 DA-3 GTRAMP C1 MCF7MIA PaCa-2 D122 and BCL1
Strong cooperativeeffects of MJ + 2DGand MJ + other drugs
(3) MJ + adriamycinin vivo
MJ iv20ndash150mgkgdissolved inlipofundinAdr (DOX) ip4mgkg
Balbc mice injected ip with 1 times 104chronic BCL1 cells lymphocyteleukemia cells
MJ + Adr significant prolongedsurvival effect
(4) MJ + PI3KAktinhibitorsin vitro
MJ 05ndash30mMEtOHDMSO
Sarcoma MCA-105 SaOS-2 darrMJ-induced activation of Aktuarrsynergic cytotoxicity
[49](5) MJ + 2DGin vitro
MJ 05ndash30mMEtOHDMSO Sarcoma MCA-105 SaOS-2 2DG darrpAkt darrMJ-ind pAkt
MJ + 2DG uarrsynergic cytotoxicity
(6) MJ + irradiationin vitro
05ndash20mMDMSO
Irradiated prostate PC-3 (radiationinduces Bcl-2 expression)
darrradiation-induced Bcl-2uarrradiation sensitivity PC-3uarrcaspase-3
[104]
(7) MJ andor POHandorcis-platin (CP)in vitro
Both testedat IC20
Human MDA-MB-435[74]
MJ + POH uarrTNFR1darrΔ120595119898uarrcytotoxicity cell cycle arrest
at G0G1 uarrapoptosis+ CP all effects enhanced
[251]
Human breastMDA-MB-231 uarrapoptosis
(8) MJ + TRAILin vitro
MJ 05mM +TRAIL(100ndash200 ngsdotmLminus1)
CRC cancer cells
darrsurvivin (IAP)darrWntTCF pathwayuarrTRAIL-induced apoptosisuarrcaspase activity
[32]
(9) MJ + Smacin vitro
MJ 05ndash20mMDMSO
Human bladder cancer EJ T24 Synergy uarrIAPs-bound caspase 3uarrapoptosis [254]
Human embryonic kidney HEK293 No cytotoxicity
(10) MJ + Smac7N(IAP antagonist)in vitro
MJ 05ndash20mMDMSO
Hormone-independent prostateDU-145 PC-3Human proximal tubular epithelialHK-2 cells (overexpressing IAPs)
Smac7N uarrMJ-induced apoptosis bycaspase-9-dependent (intrinsic) andindependent (extrinsic) pathways
[255]
(11) MJ + cisplatin(01ndash05 120583M)MJ + X-rays(025ndash3Gy)MJ + 120572-rays
MJ 01ndash10mMEtOH
Cervical cancer cellsSiHa CaSki HeLa and C33A
darr viabilitydarrsurvivaldarrIC50 radiation dose
[29]
(12) MJ + 5-FUin vitro
05mMMJ1 hthen + 5-FU
Human adenocarcinomacolon HT-39 darrIC50 5-FU (5 rarr 25mM) [31]
BCNU 13-bis-(2-chloroethyl)-1-nitrosourea 2DG 2-deoxy-D-glucose 5-FU 5-fluorouracile IAP inhibitors of apoptosis MM multiple myeloma POHperillyl alcohol smac second mitochondria-derived activator of caspases Smac7N a peptide that contains the N-terminal seven residues of smac TNFR1tumor-necrosis factor receptor-1 TRAIL tumor necrosis factor- (TNF-) related apoptosis-inducing ligand
are unfavorable cells delay progress through G1and may
enter a quiescent state known asG0 in which they can remain
for days weeks or years before resuming proliferation Ifextracellular conditions are favorable and signals to growand divide are present cells in early G
1or G0progress
through a restriction checkpoint late in G1after which cells
are committed to DNA replication even if the extracellularsignals that stimulate cell growth and division are removed[53 54] Cell cycle activity is modulated by intracellularROS levels and regulates cell survival cell proliferation andapoptosis [53ndash58]The progression through theG
1 S G2 and
M phases is promoted by cyclin-dependent kinases (CDKs)
International Journal of Cell Biology 7
which are positively and negatively regulated by cyclins andcyclin kinase inhibitors (CKis) respectively and by ROSUnlike the passage through the S G
2 and M phases G
1
progression normally relies on stimulation by mitogens andcan be blocked by antiproliferative factors passage throughthe restriction checkpoint and entry into S phase is controlledby CDKs that are sequentially regulated by cyclins D Eand A [53 54 56ndash58] In general CDK activity requirescyclin binding and depends on both positive and negativeregulatory phosphorylations [54] ROS levels influence thephosphorylating and ubiquitinating enzyme activites thuscontrolling cell cycle progression [54 57 58]
A master gene regulating cell passage through the cycleis the Myc family of transcription factors it switches onthe simultaneous synthesis of thousands of different proteinsrequired for cell growth and cell cycle function [58 59] TocounteractMyc protein activity the cell cycle is also regulatedby the p53 protein [60 61] a DNA-binding transcriptionfactor that senses DNA damage (occurring after exposureto ionizing radiation UV light oxidative stress or otherDNA damaging agents) and activates genes to halt cell cycleprogression in G
1for repair or otherwise initiates apoptosis
[56 62] When the damage caused by cell stress is mild p53activatesmechanisms for cell cycle arrest and repair damagedDNA if the damage is irreparable p53 initiates apoptosis [63]In case ofmild cellular stress low levels of p53 can activate theTP53-induced glycolysis and apoptosis regulator (TIGAR)gene that decreases the rate of glycolysis and hence ROS levelsin the cell [64] Glycolysis is essential for cancer progressionbut TIGAR provides protection from ROS and apoptosisHowever if damage caused by stress is irreparable p53prevents proliferation of cells with altered DNA and inducesincreased levels of ROS to trigger cell death [56] Increasedlevels of active p53 inhibit the cell entry into the S-phaseor induce apoptosis [56] Loss or inactivation of p53 resultsin loss of cell-cycle arrest or inhibition of apoptosis afterDNAdamage or physiological stress [65] leading to increasedgenetic instability increased accumulation of mutations andultimately oncogenesis [61] When the cell cycle is no longerregulated by p53 cells remain cycling (supported by 119888-Mycoverexpression) subverting cell cycle exit maturation andterminal differentiation [66] Over 50 of all human cancersharbor mutations and genetic alterations in cyclins and p53genes directly affecting the function of critical cell cycleproteins rendering the p53 gene ineffective in mediating cellgrowth arrest and promoting cancer cell death [62 67 68]ROS production is under p53 regulation and in turn p53being a redox-sensitive protein is influenced by ROS levelsthus ROS can act as both an upstream signal that triggersp53 activation and as a downstream factor that mediatesapoptosis [69]Thus mutations in the p53 gene downregulateits activity and consequently ROS production [70]ThroughROS p53 can also directly control metabolic traits of cells[71] regulate mitochondrial membrane potential (Δ120595
119898) and
induce cytochrome 119888-independent apoptosis (blocked by Bcl-2) ROS-mediated disruption of Δ120595
119898constitutes a pivotal
step in the apoptotic pathway of p53 and this pathwaydoes not involve cytochrome 119888 release [72] ROS can thusregulate cell fate through p53 in a way that physiological ROS
levels trigger cell protective pathways while under cytotoxicoxidative stress p53 behaves more like a cell killer [70 73]
JAs (mainly MJ) have been found to arrest cell cycle ofdifferent cancer cell types at different phases (Table 1) G
0G1
in human acute lymphoblastic leukemia Molt-4 [14 38] andhuman neuroblastoma SK-N-SH BE(2)-C [45] G
0G1-S in
human breast MCF-7 andmelanocytic MDA-MB-435 cancercells [25 74] S in human neuroblastoma BE(2)-C [43] S-G2M in human adenocarcinoma colon HT-39 [31] G
2M
in nonsmall cell lung cancer (NSCLC) lines A549 and H520[37] human neuroblastoma SH-SY5Y [44] human cervicalcarcinoma and HeLa cells [27] At 01mM MJ arrested invitro human umbilical vein endothelial cells (HUVEC) at G
1
[40]The molecular mechanisms underlying the above jasmo-
nate-mediated cell cycle arrest in cancer and embryonicHUVEC cells in vitro are not yet completely understoodChecking the above data against the UMD TP53 mutationdatabase (httpwwwp53fr) and revising the article byBerglind et al [75] we could not find correlation between agiven MJ-mediated phase arrest of cell cycle and the TP53wild type or mutated status of a cancer cell type (data notshown) a result that would agree with the original observa-tion of Fingrut and Flescher [15] stating thatMJ-mediated celldeath is independent of the p53 status [17] However otherstudies suggest thatMJ could be acting by interfering differentcell cycle regulatorymechanisms In plants MJ is able to bindthe TGACCG motif which is present in cell cycle cyclin pro-moters [76] wemight speculateMJ recognition of TGACCG-like motifs in mammalian cells cyclin promoters howeverthis would need experimental confirmation In some plantsMJ impairs the G
2-to-M transition by repressing M-phase
gene activation [77] MJ impaired the G2-to-M transition in
the human neuroblastoma SH-SY5Y cells downregulatingthe expressions of the proliferating cell nuclear antigen(PCNA a processivity factor encircling DNA at sites ofreplication and repair) andN-Myc [45] Asmentioned aboveMYC is the master gene allowing cancer cells to move alongthe cell cycle and regulating most genes needed to meet themetabolic demands for cell proliferation and survival [78]Cancer cell cycle arrest at different phases is in itself closelydependent on the cell energy and as such it is extremelydependent on metabolic ATP availability and its use byhexokinase Downregulation of Myc byMJ [45] is interestingresult that must be confirmed particularly because there is alack of specific inhibitors against Myc [57]
In some cases of cell cycle arrest MJs have been foundto upregulate p21 [37] whereas in others to downregulatesurvivinmRNA and protein levels [43ndash45] (Table 1) Survivinis an important member of the inhibitor of apoptosis protein(IAP) family that also regulates entrance of the cell cycle to theM phase and is overexpressed in cancer cells acting linkingboth regulation of the cell cycle and inhibition of apoptosis[79]
22 Jasmonates (MJ) CanCauseCancer Cell Death by InducingApoptosis andor Necrosis Apoptosis can be initiated by twosignaling cascades namely the extrinsic and the intrinsic
8 International Journal of Cell Biology
pathway The classical extrinsic pathway is triggered by thebinding of extracellular signals like tumor necrosis factor(TNF) tumor necrosis factor-related apoptosis-inducing lig-and (TRAIL) and apoptosis antigen-1 (Fas) ligand (FASL)to death receptors such as tumor necrosis factor receptor 1(TNFR1) and FAS [80]This interaction leads to procaspase-8activation which cleaves itself into caspase-8Then caspase-8can cleave and activate procaspase-3 into caspase-3 thatcleaves many substrates inside the cell inducing the typicalmorphological changes seen in apoptosis The intrinsic path-way can be triggered by intracellular stress signals such asoxidative stress and DNA damage The diverse signals aresensed by mitochondria through the balance between pro-and antiapoptotic members of the Bcl-2 family of proteinsWhen proapoptotic signals overcome antiapoptotic signalsmitochondria becomepermeabilized leading to dissipation ofthe mitochondrial electrochemical potential release of inter-membrane proteins like cytochrome 119888 SmacDIABLO AIFandHTRA2OMI Cytochrome 119888 interacts with procaspase-9and Apaf-1 to form the apoptosome complex that leads tocaspase-3 activation [81] The extrinsic pathway can connectwith the intrinsic pathway through the cleavage by caspase-8of Bid into tBid (truncated Bid) which is a proapoptoticmember of the Bcl-2 family and can lead to mitochondriapermeabilization [82 83] The members of the Bcl-2 fam-ily regulate mitochondria permeabilization and comprisesantiapoptotic (Bcl-2 Bcl-xL) proapoptotic (Bax Bak) andproapoptotic BH-3 only proteins (Bad Bid Puma andNoxa)[82ndash84] One of the mechanisms of mitochondria perme-abilization is controlled by Bax and Bak proteins that arenormally expressed in every cell but are maintained inhibitedby the antiapoptotic members When there is a signal toapoptosis BH3-only proteins interact with the antiapoptoticmembers releasing BaxBak or directly activating themleading to their oligomerization and to mitochondrial outermembrane permeabilization (MOMP) Another mechanismfor mitochondria permeabilization is the formation of themitochondrial permeability transition (MPT) pore which iscomposed by the voltage-dependent anion channel VDACthe adenine nucleotide translocase (ANT) and cyclophilin D[85]TheMPT pore comprising VDAC1 is directly regulatedby Bad [86] In contrast VDAC2 has been identified as anantiapoptotic negative regulator of Bak [87ndash89]
One of the hallmarks of cancer is resistance to cell death[90] This resistance is acquired by a host of different geneticdefects in cancer cells such as those present in the p53tumor suppressor gene [62 68 91] Normal p53 responds togenotoxic oncogenic and other stress signals by inducingantiproliferative transcriptional programs leading to growtharrest (for cell repair) or apoptosis [92] p53 mediates cyclearrest by transcriptionally activating the p21 (CDKN1A) 14-3-3120590 (SFN) and GADD45120572 genes whereas p53-dependentapoptosis is triggered by turning on the synthesis of proteinsthat produce ROS [58 93] and transactivating differentmitochondrial proapoptotic genes as well as death receptorsfor the synthesis of the corresponding proteins [58 94]Whenp53 (and its target genes) expression levels are below a thresh-old they induce cell cycle arrest but not apoptosis above thisthreshold p53 trigger apoptosis [95] Lowering this threshold
with inhibitors of the antiapoptotic Bcl-2 family proteinssensitize cells to p53-induced apoptosis [95] By damagingcells chemotherapeutical drugs turn on p53rsquos transcriptionalfunction that upregulates the expression of proapoptotic pro-teins Defects in the p53 function or in the apoptotic cascadeslead to cell phenotypes resistant to chemotherapy Apoptosiscan also be induced by anticancer agents independently ofp53 by either directly modulating the apoptotic machineryor directly interacting with mitochondria [96] ROS levelsmight be playing a central role in mediating various forms ofnonapoptotic caspase-independent programmed cell deatheither through direct irreversible oxidative damage to keyproteins nucleic acid molecules and cellular structure or bythe activation of prodeath signal transduction programs [97]In a ROS-dependent nonapoptotic cell death pathway respi-ration through the mitochondrial electron transport system(ETS) chain is not essential and ROS is generated throughNADPH oxidases [97] In contrast to the mitochondrial ETSwhich generates ROS as a byproduct of respiration NADPHoxidases produceROS as their primary function [98] Raise ofROS is characteristic after ionizing radiation chemotherapyand targeted therapy treatments [58 99] and may be killingcancer cells through a nonapoptotic ROS-dependent celldeath pathway
Tumor cells maintain high ATPADP as well asNADHNAD+ ratios so as to assure that ATP levels arenever limited ATP depletion would represent then anunsustainable metabolic stress for tumor cells and it isassociated with necrotic death in some cell types [100]Recently it was shown that a signaling cascade centered onthe proteins RIP1 and RIP3 can trigger cell death by necrosisin many situations and the RIP1-RIP3 complex appears topromote ATP depletion during necroptosis by impingingon components of the MPT pore [101 102] Necrosis was aterm used to describe a process of accidental cell death inwhich cells swell and burst releasing its intracellular contentwhereas necroptosis modernly refers rather to an orderedcell death program different from apoptosis apoptosis is acaspase-dependent cell death process whereas necrosis iscaspase-independent process [102 103] Studies concerningnecrosis have focusedmainly on cell membrane permeabilityto propidium iodide (PI) in the flow citometry doublestaining with annexin-VPI
Fingrut et al [17] by studying the effect of MJ ontwo clones of a highly malignant B-cell lymphoma (oneharboring a normal wild type (wt) p53 gene while the otherexpressed a mutated inactive p53 protein) (Table 1) reportedthat MJ was equally toxic to both clones [17] In contrastthe p53-mutant cells were resistant to radiomimetics (drugshaving effects similar to those produced in radiotherapy)and chemotherapeutic drugs In this cancer cell model MJinduced mostly apoptotic cell death in the wt p53-expressinglymphoma cells while no signs of early apoptosis weredetected in the mutant p53-expressing lymphoma cells thatwere killed by MJ through a p53-independent nonapoptoticcell death mechanism However other cancer cell types canundergo apoptosis with or without functional p53 and aswe will see MJ can induce apoptotic andor nonapoptoticcell death in wt p53 as well as in TP53 mutated cells
International Journal of Cell Biology 9
(Table 1) Nonapoptotic cell death could be the consequenceof the severe bioenergetic damages induced by the directinteraction of MJ with mitochondria leading to increasedROS levels inhibition of ATP synthesis and ATP depletion[20 30] Jasmonates including MJ and some of its analogshave been found to be strongly proapoptotic in vitro indifferent types of cancer cells by both the extrinsic and theintrinsic pathway (Table 1) with no or little cytotoxicity tonormal cells [15 25 36 37 44 45] Thus MJ induced theexpression of TNFR1 in human breast MCF-7 and MDA-MB-435 cells [25 74] and in hormone-independent humanprostate PC-3 and DU 145 cancer cells promoting apoptosisby the extrinsic pathway in these cells [48] TRAIL (TNF-related apoptosis inducing ligand) is able to induce apoptosisin prostate cancer cell yet overexpression of antiapoptoticproteins and inhibition of proapoptotic proteins results indiminishing the TRAIL-mediated apoptosis pathway TRAILhas emerged as a potent anticancer agent in laboratory-based studies and preclinical trials and JAs have been foundto sensitize cancer cells to TRAIL-mediated apoptosis [32](Table 3) J7 a synthetic analogue of MJ enhanced TRAIL-mediated apoptosis through upregulation of ROS levels inhuman hepatomaHepG2 cells apoptosis was induced via Bidcleavage downregulation of X-linked inhibitor of apoptosisprotein (XIAP) cellular inhibitor of apoptosis-1 (cIAP-1)B-cell lymphoma-extra large (Bcl-xL) and activation ofcaspases [35] Other examples of MJ-mediated apoptoticandor nonapoptotic cell death in different cancer cell types(Table 1) are as follows (1) MJ induced ROS production inA549 lung adenocarcinoma cells activating Bcl-2-associated-X-protein (Bax) as well as Bcl-2-Like-1 (Bcl2L1) proapoptoticproteins in these cells [36] (2) MJ activated caspase-3 inleukemia Molt-4 prostate SK28 and LNCaP cancer cells[15] (3) MJ inhibited the antiapoptotic Bcl-2 protein andenhanced proapoptotic proteins in radio-resistant humanprostate cancer cell line PC-3 [104] (4) MJ suppressed theproliferation of the neuroblastoma cell lines SH-SY5Y SK-N-SH and BE(2)-C through a strong apoptosis-inducingeffect as measured by annexin VFITCPI MJ treatment alsodownregulated the expression of the antiapoptotic proteinsXIAP and survivin [44 45] (5) MJ-mediated cell death wasobserved on cervical cancer and myeloma cells and it wasreported to be due to mixed characteristics of apoptosis andnecrosis [26 105] So far studies showing MJ inducing theRIP1 signaling cascade have not been reported howeverit will be important to deep into these analyses to cleardistinguish between MJ-induced apoptosis andor necrosis
23 Methyl Jasmonate Induces the Production of ReactiveOxygen Species (ROS) Responses in Some Cancer Cell TypesLeading Also to Cell Death Intracellular ROS levels reg-ulate cell cycle activity cell metabolism and survival cellproliferation and apoptosis [53ndash58] Stressful conditionsproduce a transient excess of intracellular ROS in normaltissues however under chronic pathological and oxidativestress conditions ROS is continuously produced in excessand persistently causing damage to DNA cell membraneslipids proteins and inducing apoptosis to control and avoid
oxidative damages cells counteract excess ROS by enzymatic(superoxide dismutase (SOD) glutathione peroxidase (GPx)and catalase) andor nonenzymatic (glutathione thoredoxin)antioxidant elements [99 106] Controlling ROS is central tointracellular redox signaling and homeostasis on the otherhand uncontrolledunbalanced ROS production modifiesintracellular redox leading to apoptosis [58 82 99 106] ROScan be generated by different biological systems (1) director indirect extracellular stimulation of plasma membrane-bound NAD(P)H oxidases (NOX) can generate intracellularH2O2[57 107ndash110] (2) biochemical pathways associated to
fatty acid oxidation and lipoxygenases activities generateROS as normal byproducts of these activities [111] (3)ATP generation by oxidative phosphorylation (OxPhos) inmitochondria is accompanied by the production of ROS as aconsequence of electron leakage from the electron transportchain mitochondria are considered to be major sources ofROS inmammalian cells particularly under a variety of stress-ful conditions [82 99] Superoxide (O
2
minus) is mainly produced
from complexes I and III and it is rapidly dismutated to H2O2
and oxygen by SOD [112] H2O2serves both as a toxic oxidant
and as an essential signaling molecule regulating cellularbiological processes [113] A strong positive correlation existsbetween mitochondrial membrane potential (Δ120595
119898) and ROS
production [106 114 115] Mitochondria produce more ROSat high membrane potential and an increase in the Δ120595
119898
produced either by a closure of MPT pore or by inhibitionof the ATP synthase is associated with increased ROS pro-duction However in cases of mitochondrial dysfunctionslow Δ120595
119898and decreased activity of the respiratory chain
are accompanied by a simultaneous increase in ROS [116117] It has been reported that cancer cells have higher ROScontent compared to normal cells [118 119] probably due toabnormal respiration because of dysfunctional mutations inmitochondria [120] but they also have increased contents ofantioxidants (SOD catalase glutathione thioredoxin etc)to counteract ROS [58 121] Tumor cells cannot tolerateexcessive high ROS levels [118 122] and as a matter of factradiation therapy chemotherapies and targeted therapiesinduce apoptosis and kill some cancer cells by generatingexcess ROS [99 106 123 124] Thus inducing higher levelsof ROS in cancers cells can be exploited as an effectiveselective strategy to kill tumor cells over normal cells [119]However prolonged treatment with the same drug reducesROS levels in cancer cells [124] indicating that when theybecome drug-resistant tumor cells have lower ROS contentthan drug-sensitive cancer cells Evidence suggests that drug-resistant cells have a higher expression of catalase at theplasma membrane that keeps reduced ROS levels [121]
In plants MJ-mediated damage to mitochondria is fol-lowed by a rapid production of H
2O2[125] H
2O2is used as
a second messenger for the induction of genes and productsrelated to defense against both herbivores and pathogens[126] in addition H
2O2induces apoptosis and senescence
[127 128] On the other hand it has been signaled that theMJ-induced production of H
2O2in plants was prevented by
inhibitors of NAD(P)H oxidase [128 129] indicating anothermechanism of MJ-mediated production of ROS and a majorrole for this enzyme in MJ-induced ROS responses MJ
10 International Journal of Cell Biology
induces also an endogenous H2O2response in some cancer
cell lines (Table 1) [19 36 38 130ndash132] In A549 cancerlung cells the H
2O2response induced by MJ increased the
expression of the proapoptotic Bax and Bcl-Xs (although notbeing that of the antiapoptotic Bcl-2 and Bcl-xL proteins) andthis led to apoptosis this event was inhibited by catalase (aspecific inhibitor of H
2O2) [36] MJ increased the intracel-
lular levels of H2O2and superoxide (O
2) ions in human C6
glioma cells in vitro and induced the expression of heat shockprotein 72 (HSP72) a negative regulator of H
2O2-induced
oxidative stress in these cells in vitro MJ-induced HSP72expression was blocked by ROS inhibitors [130] ElevatedROS generation was observed in response to both MJ andJA levels in the acute myeloid leukemia HL-60 and KG1a celllines but other factors like the MJ-mediated generation ofmitochondrial SOD rather than ROS were determinant forapoptosis or differentiation in these cells [132] Suppression ofMJ-induced apoptosis by antioxidants like N-acetylcysteine(NAC) and catalase but not by inhibitors of hydroxyl radicalsand superoxide ions indicates that H
2O2is nonetheless one
of the important factors involved inMJ anticancer effects [19]
24 By Dissociating Hexokinase (HK) from the VoltageDependent Anion Channel (VDAC) on the Outer Mitochon-drial Membrane MJ Dissociates Glycolysis from OxidativePhosphorylation Causing Severe Bioenergetic Deregulations inCancer Cells Metabolism and cell growth are two cellularprocesses that are tighly linked and regulated [133] Theclassic bioenergetics phenotype of cancer cells of enhancedglycolysis under aerobic conditions was described by War-burg [134] However the ldquoWarburg effectrdquo does not neces-sarily imply mitochondrial dysfunction [135] To increaseglycolysis cancer cells upregulate the transcription of genesinvolved in the glycolytic pathway (ie glucose transportersglycolytic enzymes etc) This metabolic reprogrammingallows increased rate of ATP production synthesis of lipidsand a new redox balance Cancer cells actually use a com-bination of both glycolysis and mitochondrial respirationto produce energy and they may vary in regards to thepreferential use of these pathways being in some caseseither more glycolyticless oxidative or less glycolyticmoreoxidative depending on the prevalent normoxic or hypoxicenvironmental conditions and their capacities to expressadequate levels of oncogenes and tumor suppressor geneproducts for cell growth [136ndash139] or when either glycolysisor OxPhos is inhibited in whose case there can be partialcompensation by the other metabolic pathway [17] Shiftingthe balance between those two processes with glycolyticandor mitochondrial enzyme inhibitors might represent aninteresting way to look for new anticancer agents
The first step in the metabolism of glucose is catalyzedby the enzyme hexokinase (HK) which by phosphorylatingglucose makes it negative sequestering it into the cytoplasmin the form of glucose-6-phosphate (G6P) In normal tissuesthis crucial step is catalyzed by four different HK isoforms(HK1 HK2 HK3 and glucokinase) indicating that regulationof glucose phosphorylation can vary in different tissues underdifferent condition [140] HK1 and HK2 have overlapping
tissue expression but different subcellular distributions withHK1 associated mainly with mitochondria and HK2 associ-atedwith bothmitochondrial and cytoplasmic compartments[141] HK1 is widely and constitutively expressed whereasHK2 predominates during embryogenesis and is not widelyexpressed in adult tissues but only in a limited numberof normal adult tissues [141] their different subcellulardistributions and kinetic properties reflect their differentmetabolic roles [141 142] Under physiological conditionsHK1 is predominantly bound to the outer mitochondrialmembrane where it primarily channels glucose toward gly-colysis [140] whereas HK2 is mainly soluble and controlsglycogen formation [140 141] In malignant tissues HK1and HK2 isoforms are glycolytic and overexpressed (HK1more than HK2) (httpwwwproteinatlascom) [143] andare tightly bound to mitochondria HK1 and HK2 bindthrough theirN-terminal hydrophobic regions to the voltage-dependent calcium channel isoform1 (VDAC1) the prevalentpore protein in the outer mitochondrial membrane [2482 143ndash146] VDAC1 is also overexpressed in cancer cells[147] and the binding of HK to VDAC1 on the outer mito-chondrial membrane is a fundamental aspect of the aerobicglycolytic metabolism of cancer cells [82 135 147 148]HKVDAC association appears to protect tumor cells frommitochondrial outer membrane permeabilization (MOMP)which marks a point of no return leading to cell death[149] VDAC transports ADP and inorganic phosphate (Pi)the substrates needed for the production of ATP into themitochondria it also transports ATP out of themitochondriainto the cytoplasm and facilitates the availability of ATPto HK2 [143 150 151] A continuous supply of glucose toHK maintains this activity at a constant level and controlsthe flux through the mitochondrial electron transport chaininfluencing mitochondrial ROS production [152] HK bindsthrough hydrophobic interaction to VDAC1 the prevalentpore protein in the outer mitochondrial membrane [24 82143ndash146] thereby assuring and gaining a preferential accessto ATP [82 150 153] Translocation of HK2 from the cytosolto the mitochondria is regulated through phosphorylationof HK by Akt [141] HK2 only binds VDAC after phos-phorylation by Akt [151 152] Glycogen synthase kinase 3beta (GSK3120573) a major target of Akt kinases phosphorylatesVDAC [154ndash157] and dissociates HK [156] HK2 boundto mitochondria enhance its affinity to ATP and is lesssensitive to enzymatic inhibition by the G6P product [140141 158] HK2 can produce G6P from cytoplasmic glucoseand mitochondrial ATP without regulatory restraints evenunder hypoxia supporting rapid cell growth [142 143 159ndash161]HK1 andHK2 aremore tightly bound toVDAC in cancercells than in nonmalignant cells [156 162 163] HKVDA1interaction is relatively stable for HK1 while it is dependentupon G6P levels for HK2 [140 141] The higher affinity ofmitochondrial HK2 for glucose ATP and VDAC1 representsa high advantage for cancer cells survival and growth con-ferring to them an aerobic glycolytic phenotype underlyingthe Warburg effect [82 135 138 142 147 148 150 160]Differential interactions of HK1 and HK2 with mitochondriamay underlie different glycolytic profiles in cancer cells [164165] As rate-controlling enzymes in the glycolyic pathway
International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Cell Biology 7
which are positively and negatively regulated by cyclins andcyclin kinase inhibitors (CKis) respectively and by ROSUnlike the passage through the S G
2 and M phases G
1
progression normally relies on stimulation by mitogens andcan be blocked by antiproliferative factors passage throughthe restriction checkpoint and entry into S phase is controlledby CDKs that are sequentially regulated by cyclins D Eand A [53 54 56ndash58] In general CDK activity requirescyclin binding and depends on both positive and negativeregulatory phosphorylations [54] ROS levels influence thephosphorylating and ubiquitinating enzyme activites thuscontrolling cell cycle progression [54 57 58]
A master gene regulating cell passage through the cycleis the Myc family of transcription factors it switches onthe simultaneous synthesis of thousands of different proteinsrequired for cell growth and cell cycle function [58 59] TocounteractMyc protein activity the cell cycle is also regulatedby the p53 protein [60 61] a DNA-binding transcriptionfactor that senses DNA damage (occurring after exposureto ionizing radiation UV light oxidative stress or otherDNA damaging agents) and activates genes to halt cell cycleprogression in G
1for repair or otherwise initiates apoptosis
[56 62] When the damage caused by cell stress is mild p53activatesmechanisms for cell cycle arrest and repair damagedDNA if the damage is irreparable p53 initiates apoptosis [63]In case ofmild cellular stress low levels of p53 can activate theTP53-induced glycolysis and apoptosis regulator (TIGAR)gene that decreases the rate of glycolysis and hence ROS levelsin the cell [64] Glycolysis is essential for cancer progressionbut TIGAR provides protection from ROS and apoptosisHowever if damage caused by stress is irreparable p53prevents proliferation of cells with altered DNA and inducesincreased levels of ROS to trigger cell death [56] Increasedlevels of active p53 inhibit the cell entry into the S-phaseor induce apoptosis [56] Loss or inactivation of p53 resultsin loss of cell-cycle arrest or inhibition of apoptosis afterDNAdamage or physiological stress [65] leading to increasedgenetic instability increased accumulation of mutations andultimately oncogenesis [61] When the cell cycle is no longerregulated by p53 cells remain cycling (supported by 119888-Mycoverexpression) subverting cell cycle exit maturation andterminal differentiation [66] Over 50 of all human cancersharbor mutations and genetic alterations in cyclins and p53genes directly affecting the function of critical cell cycleproteins rendering the p53 gene ineffective in mediating cellgrowth arrest and promoting cancer cell death [62 67 68]ROS production is under p53 regulation and in turn p53being a redox-sensitive protein is influenced by ROS levelsthus ROS can act as both an upstream signal that triggersp53 activation and as a downstream factor that mediatesapoptosis [69]Thus mutations in the p53 gene downregulateits activity and consequently ROS production [70]ThroughROS p53 can also directly control metabolic traits of cells[71] regulate mitochondrial membrane potential (Δ120595
119898) and
induce cytochrome 119888-independent apoptosis (blocked by Bcl-2) ROS-mediated disruption of Δ120595
119898constitutes a pivotal
step in the apoptotic pathway of p53 and this pathwaydoes not involve cytochrome 119888 release [72] ROS can thusregulate cell fate through p53 in a way that physiological ROS
levels trigger cell protective pathways while under cytotoxicoxidative stress p53 behaves more like a cell killer [70 73]
JAs (mainly MJ) have been found to arrest cell cycle ofdifferent cancer cell types at different phases (Table 1) G
0G1
in human acute lymphoblastic leukemia Molt-4 [14 38] andhuman neuroblastoma SK-N-SH BE(2)-C [45] G
0G1-S in
human breast MCF-7 andmelanocytic MDA-MB-435 cancercells [25 74] S in human neuroblastoma BE(2)-C [43] S-G2M in human adenocarcinoma colon HT-39 [31] G
2M
in nonsmall cell lung cancer (NSCLC) lines A549 and H520[37] human neuroblastoma SH-SY5Y [44] human cervicalcarcinoma and HeLa cells [27] At 01mM MJ arrested invitro human umbilical vein endothelial cells (HUVEC) at G
1
[40]The molecular mechanisms underlying the above jasmo-
nate-mediated cell cycle arrest in cancer and embryonicHUVEC cells in vitro are not yet completely understoodChecking the above data against the UMD TP53 mutationdatabase (httpwwwp53fr) and revising the article byBerglind et al [75] we could not find correlation between agiven MJ-mediated phase arrest of cell cycle and the TP53wild type or mutated status of a cancer cell type (data notshown) a result that would agree with the original observa-tion of Fingrut and Flescher [15] stating thatMJ-mediated celldeath is independent of the p53 status [17] However otherstudies suggest thatMJ could be acting by interfering differentcell cycle regulatorymechanisms In plants MJ is able to bindthe TGACCG motif which is present in cell cycle cyclin pro-moters [76] wemight speculateMJ recognition of TGACCG-like motifs in mammalian cells cyclin promoters howeverthis would need experimental confirmation In some plantsMJ impairs the G
2-to-M transition by repressing M-phase
gene activation [77] MJ impaired the G2-to-M transition in
the human neuroblastoma SH-SY5Y cells downregulatingthe expressions of the proliferating cell nuclear antigen(PCNA a processivity factor encircling DNA at sites ofreplication and repair) andN-Myc [45] Asmentioned aboveMYC is the master gene allowing cancer cells to move alongthe cell cycle and regulating most genes needed to meet themetabolic demands for cell proliferation and survival [78]Cancer cell cycle arrest at different phases is in itself closelydependent on the cell energy and as such it is extremelydependent on metabolic ATP availability and its use byhexokinase Downregulation of Myc byMJ [45] is interestingresult that must be confirmed particularly because there is alack of specific inhibitors against Myc [57]
In some cases of cell cycle arrest MJs have been foundto upregulate p21 [37] whereas in others to downregulatesurvivinmRNA and protein levels [43ndash45] (Table 1) Survivinis an important member of the inhibitor of apoptosis protein(IAP) family that also regulates entrance of the cell cycle to theM phase and is overexpressed in cancer cells acting linkingboth regulation of the cell cycle and inhibition of apoptosis[79]
22 Jasmonates (MJ) CanCauseCancer Cell Death by InducingApoptosis andor Necrosis Apoptosis can be initiated by twosignaling cascades namely the extrinsic and the intrinsic
8 International Journal of Cell Biology
pathway The classical extrinsic pathway is triggered by thebinding of extracellular signals like tumor necrosis factor(TNF) tumor necrosis factor-related apoptosis-inducing lig-and (TRAIL) and apoptosis antigen-1 (Fas) ligand (FASL)to death receptors such as tumor necrosis factor receptor 1(TNFR1) and FAS [80]This interaction leads to procaspase-8activation which cleaves itself into caspase-8Then caspase-8can cleave and activate procaspase-3 into caspase-3 thatcleaves many substrates inside the cell inducing the typicalmorphological changes seen in apoptosis The intrinsic path-way can be triggered by intracellular stress signals such asoxidative stress and DNA damage The diverse signals aresensed by mitochondria through the balance between pro-and antiapoptotic members of the Bcl-2 family of proteinsWhen proapoptotic signals overcome antiapoptotic signalsmitochondria becomepermeabilized leading to dissipation ofthe mitochondrial electrochemical potential release of inter-membrane proteins like cytochrome 119888 SmacDIABLO AIFandHTRA2OMI Cytochrome 119888 interacts with procaspase-9and Apaf-1 to form the apoptosome complex that leads tocaspase-3 activation [81] The extrinsic pathway can connectwith the intrinsic pathway through the cleavage by caspase-8of Bid into tBid (truncated Bid) which is a proapoptoticmember of the Bcl-2 family and can lead to mitochondriapermeabilization [82 83] The members of the Bcl-2 fam-ily regulate mitochondria permeabilization and comprisesantiapoptotic (Bcl-2 Bcl-xL) proapoptotic (Bax Bak) andproapoptotic BH-3 only proteins (Bad Bid Puma andNoxa)[82ndash84] One of the mechanisms of mitochondria perme-abilization is controlled by Bax and Bak proteins that arenormally expressed in every cell but are maintained inhibitedby the antiapoptotic members When there is a signal toapoptosis BH3-only proteins interact with the antiapoptoticmembers releasing BaxBak or directly activating themleading to their oligomerization and to mitochondrial outermembrane permeabilization (MOMP) Another mechanismfor mitochondria permeabilization is the formation of themitochondrial permeability transition (MPT) pore which iscomposed by the voltage-dependent anion channel VDACthe adenine nucleotide translocase (ANT) and cyclophilin D[85]TheMPT pore comprising VDAC1 is directly regulatedby Bad [86] In contrast VDAC2 has been identified as anantiapoptotic negative regulator of Bak [87ndash89]
One of the hallmarks of cancer is resistance to cell death[90] This resistance is acquired by a host of different geneticdefects in cancer cells such as those present in the p53tumor suppressor gene [62 68 91] Normal p53 responds togenotoxic oncogenic and other stress signals by inducingantiproliferative transcriptional programs leading to growtharrest (for cell repair) or apoptosis [92] p53 mediates cyclearrest by transcriptionally activating the p21 (CDKN1A) 14-3-3120590 (SFN) and GADD45120572 genes whereas p53-dependentapoptosis is triggered by turning on the synthesis of proteinsthat produce ROS [58 93] and transactivating differentmitochondrial proapoptotic genes as well as death receptorsfor the synthesis of the corresponding proteins [58 94]Whenp53 (and its target genes) expression levels are below a thresh-old they induce cell cycle arrest but not apoptosis above thisthreshold p53 trigger apoptosis [95] Lowering this threshold
with inhibitors of the antiapoptotic Bcl-2 family proteinssensitize cells to p53-induced apoptosis [95] By damagingcells chemotherapeutical drugs turn on p53rsquos transcriptionalfunction that upregulates the expression of proapoptotic pro-teins Defects in the p53 function or in the apoptotic cascadeslead to cell phenotypes resistant to chemotherapy Apoptosiscan also be induced by anticancer agents independently ofp53 by either directly modulating the apoptotic machineryor directly interacting with mitochondria [96] ROS levelsmight be playing a central role in mediating various forms ofnonapoptotic caspase-independent programmed cell deatheither through direct irreversible oxidative damage to keyproteins nucleic acid molecules and cellular structure or bythe activation of prodeath signal transduction programs [97]In a ROS-dependent nonapoptotic cell death pathway respi-ration through the mitochondrial electron transport system(ETS) chain is not essential and ROS is generated throughNADPH oxidases [97] In contrast to the mitochondrial ETSwhich generates ROS as a byproduct of respiration NADPHoxidases produceROS as their primary function [98] Raise ofROS is characteristic after ionizing radiation chemotherapyand targeted therapy treatments [58 99] and may be killingcancer cells through a nonapoptotic ROS-dependent celldeath pathway
Tumor cells maintain high ATPADP as well asNADHNAD+ ratios so as to assure that ATP levels arenever limited ATP depletion would represent then anunsustainable metabolic stress for tumor cells and it isassociated with necrotic death in some cell types [100]Recently it was shown that a signaling cascade centered onthe proteins RIP1 and RIP3 can trigger cell death by necrosisin many situations and the RIP1-RIP3 complex appears topromote ATP depletion during necroptosis by impingingon components of the MPT pore [101 102] Necrosis was aterm used to describe a process of accidental cell death inwhich cells swell and burst releasing its intracellular contentwhereas necroptosis modernly refers rather to an orderedcell death program different from apoptosis apoptosis is acaspase-dependent cell death process whereas necrosis iscaspase-independent process [102 103] Studies concerningnecrosis have focusedmainly on cell membrane permeabilityto propidium iodide (PI) in the flow citometry doublestaining with annexin-VPI
Fingrut et al [17] by studying the effect of MJ ontwo clones of a highly malignant B-cell lymphoma (oneharboring a normal wild type (wt) p53 gene while the otherexpressed a mutated inactive p53 protein) (Table 1) reportedthat MJ was equally toxic to both clones [17] In contrastthe p53-mutant cells were resistant to radiomimetics (drugshaving effects similar to those produced in radiotherapy)and chemotherapeutic drugs In this cancer cell model MJinduced mostly apoptotic cell death in the wt p53-expressinglymphoma cells while no signs of early apoptosis weredetected in the mutant p53-expressing lymphoma cells thatwere killed by MJ through a p53-independent nonapoptoticcell death mechanism However other cancer cell types canundergo apoptosis with or without functional p53 and aswe will see MJ can induce apoptotic andor nonapoptoticcell death in wt p53 as well as in TP53 mutated cells
International Journal of Cell Biology 9
(Table 1) Nonapoptotic cell death could be the consequenceof the severe bioenergetic damages induced by the directinteraction of MJ with mitochondria leading to increasedROS levels inhibition of ATP synthesis and ATP depletion[20 30] Jasmonates including MJ and some of its analogshave been found to be strongly proapoptotic in vitro indifferent types of cancer cells by both the extrinsic and theintrinsic pathway (Table 1) with no or little cytotoxicity tonormal cells [15 25 36 37 44 45] Thus MJ induced theexpression of TNFR1 in human breast MCF-7 and MDA-MB-435 cells [25 74] and in hormone-independent humanprostate PC-3 and DU 145 cancer cells promoting apoptosisby the extrinsic pathway in these cells [48] TRAIL (TNF-related apoptosis inducing ligand) is able to induce apoptosisin prostate cancer cell yet overexpression of antiapoptoticproteins and inhibition of proapoptotic proteins results indiminishing the TRAIL-mediated apoptosis pathway TRAILhas emerged as a potent anticancer agent in laboratory-based studies and preclinical trials and JAs have been foundto sensitize cancer cells to TRAIL-mediated apoptosis [32](Table 3) J7 a synthetic analogue of MJ enhanced TRAIL-mediated apoptosis through upregulation of ROS levels inhuman hepatomaHepG2 cells apoptosis was induced via Bidcleavage downregulation of X-linked inhibitor of apoptosisprotein (XIAP) cellular inhibitor of apoptosis-1 (cIAP-1)B-cell lymphoma-extra large (Bcl-xL) and activation ofcaspases [35] Other examples of MJ-mediated apoptoticandor nonapoptotic cell death in different cancer cell types(Table 1) are as follows (1) MJ induced ROS production inA549 lung adenocarcinoma cells activating Bcl-2-associated-X-protein (Bax) as well as Bcl-2-Like-1 (Bcl2L1) proapoptoticproteins in these cells [36] (2) MJ activated caspase-3 inleukemia Molt-4 prostate SK28 and LNCaP cancer cells[15] (3) MJ inhibited the antiapoptotic Bcl-2 protein andenhanced proapoptotic proteins in radio-resistant humanprostate cancer cell line PC-3 [104] (4) MJ suppressed theproliferation of the neuroblastoma cell lines SH-SY5Y SK-N-SH and BE(2)-C through a strong apoptosis-inducingeffect as measured by annexin VFITCPI MJ treatment alsodownregulated the expression of the antiapoptotic proteinsXIAP and survivin [44 45] (5) MJ-mediated cell death wasobserved on cervical cancer and myeloma cells and it wasreported to be due to mixed characteristics of apoptosis andnecrosis [26 105] So far studies showing MJ inducing theRIP1 signaling cascade have not been reported howeverit will be important to deep into these analyses to cleardistinguish between MJ-induced apoptosis andor necrosis
23 Methyl Jasmonate Induces the Production of ReactiveOxygen Species (ROS) Responses in Some Cancer Cell TypesLeading Also to Cell Death Intracellular ROS levels reg-ulate cell cycle activity cell metabolism and survival cellproliferation and apoptosis [53ndash58] Stressful conditionsproduce a transient excess of intracellular ROS in normaltissues however under chronic pathological and oxidativestress conditions ROS is continuously produced in excessand persistently causing damage to DNA cell membraneslipids proteins and inducing apoptosis to control and avoid
oxidative damages cells counteract excess ROS by enzymatic(superoxide dismutase (SOD) glutathione peroxidase (GPx)and catalase) andor nonenzymatic (glutathione thoredoxin)antioxidant elements [99 106] Controlling ROS is central tointracellular redox signaling and homeostasis on the otherhand uncontrolledunbalanced ROS production modifiesintracellular redox leading to apoptosis [58 82 99 106] ROScan be generated by different biological systems (1) director indirect extracellular stimulation of plasma membrane-bound NAD(P)H oxidases (NOX) can generate intracellularH2O2[57 107ndash110] (2) biochemical pathways associated to
fatty acid oxidation and lipoxygenases activities generateROS as normal byproducts of these activities [111] (3)ATP generation by oxidative phosphorylation (OxPhos) inmitochondria is accompanied by the production of ROS as aconsequence of electron leakage from the electron transportchain mitochondria are considered to be major sources ofROS inmammalian cells particularly under a variety of stress-ful conditions [82 99] Superoxide (O
2
minus) is mainly produced
from complexes I and III and it is rapidly dismutated to H2O2
and oxygen by SOD [112] H2O2serves both as a toxic oxidant
and as an essential signaling molecule regulating cellularbiological processes [113] A strong positive correlation existsbetween mitochondrial membrane potential (Δ120595
119898) and ROS
production [106 114 115] Mitochondria produce more ROSat high membrane potential and an increase in the Δ120595
119898
produced either by a closure of MPT pore or by inhibitionof the ATP synthase is associated with increased ROS pro-duction However in cases of mitochondrial dysfunctionslow Δ120595
119898and decreased activity of the respiratory chain
are accompanied by a simultaneous increase in ROS [116117] It has been reported that cancer cells have higher ROScontent compared to normal cells [118 119] probably due toabnormal respiration because of dysfunctional mutations inmitochondria [120] but they also have increased contents ofantioxidants (SOD catalase glutathione thioredoxin etc)to counteract ROS [58 121] Tumor cells cannot tolerateexcessive high ROS levels [118 122] and as a matter of factradiation therapy chemotherapies and targeted therapiesinduce apoptosis and kill some cancer cells by generatingexcess ROS [99 106 123 124] Thus inducing higher levelsof ROS in cancers cells can be exploited as an effectiveselective strategy to kill tumor cells over normal cells [119]However prolonged treatment with the same drug reducesROS levels in cancer cells [124] indicating that when theybecome drug-resistant tumor cells have lower ROS contentthan drug-sensitive cancer cells Evidence suggests that drug-resistant cells have a higher expression of catalase at theplasma membrane that keeps reduced ROS levels [121]
In plants MJ-mediated damage to mitochondria is fol-lowed by a rapid production of H
2O2[125] H
2O2is used as
a second messenger for the induction of genes and productsrelated to defense against both herbivores and pathogens[126] in addition H
2O2induces apoptosis and senescence
[127 128] On the other hand it has been signaled that theMJ-induced production of H
2O2in plants was prevented by
inhibitors of NAD(P)H oxidase [128 129] indicating anothermechanism of MJ-mediated production of ROS and a majorrole for this enzyme in MJ-induced ROS responses MJ
10 International Journal of Cell Biology
induces also an endogenous H2O2response in some cancer
cell lines (Table 1) [19 36 38 130ndash132] In A549 cancerlung cells the H
2O2response induced by MJ increased the
expression of the proapoptotic Bax and Bcl-Xs (although notbeing that of the antiapoptotic Bcl-2 and Bcl-xL proteins) andthis led to apoptosis this event was inhibited by catalase (aspecific inhibitor of H
2O2) [36] MJ increased the intracel-
lular levels of H2O2and superoxide (O
2) ions in human C6
glioma cells in vitro and induced the expression of heat shockprotein 72 (HSP72) a negative regulator of H
2O2-induced
oxidative stress in these cells in vitro MJ-induced HSP72expression was blocked by ROS inhibitors [130] ElevatedROS generation was observed in response to both MJ andJA levels in the acute myeloid leukemia HL-60 and KG1a celllines but other factors like the MJ-mediated generation ofmitochondrial SOD rather than ROS were determinant forapoptosis or differentiation in these cells [132] Suppression ofMJ-induced apoptosis by antioxidants like N-acetylcysteine(NAC) and catalase but not by inhibitors of hydroxyl radicalsand superoxide ions indicates that H
2O2is nonetheless one
of the important factors involved inMJ anticancer effects [19]
24 By Dissociating Hexokinase (HK) from the VoltageDependent Anion Channel (VDAC) on the Outer Mitochon-drial Membrane MJ Dissociates Glycolysis from OxidativePhosphorylation Causing Severe Bioenergetic Deregulations inCancer Cells Metabolism and cell growth are two cellularprocesses that are tighly linked and regulated [133] Theclassic bioenergetics phenotype of cancer cells of enhancedglycolysis under aerobic conditions was described by War-burg [134] However the ldquoWarburg effectrdquo does not neces-sarily imply mitochondrial dysfunction [135] To increaseglycolysis cancer cells upregulate the transcription of genesinvolved in the glycolytic pathway (ie glucose transportersglycolytic enzymes etc) This metabolic reprogrammingallows increased rate of ATP production synthesis of lipidsand a new redox balance Cancer cells actually use a com-bination of both glycolysis and mitochondrial respirationto produce energy and they may vary in regards to thepreferential use of these pathways being in some caseseither more glycolyticless oxidative or less glycolyticmoreoxidative depending on the prevalent normoxic or hypoxicenvironmental conditions and their capacities to expressadequate levels of oncogenes and tumor suppressor geneproducts for cell growth [136ndash139] or when either glycolysisor OxPhos is inhibited in whose case there can be partialcompensation by the other metabolic pathway [17] Shiftingthe balance between those two processes with glycolyticandor mitochondrial enzyme inhibitors might represent aninteresting way to look for new anticancer agents
The first step in the metabolism of glucose is catalyzedby the enzyme hexokinase (HK) which by phosphorylatingglucose makes it negative sequestering it into the cytoplasmin the form of glucose-6-phosphate (G6P) In normal tissuesthis crucial step is catalyzed by four different HK isoforms(HK1 HK2 HK3 and glucokinase) indicating that regulationof glucose phosphorylation can vary in different tissues underdifferent condition [140] HK1 and HK2 have overlapping
tissue expression but different subcellular distributions withHK1 associated mainly with mitochondria and HK2 associ-atedwith bothmitochondrial and cytoplasmic compartments[141] HK1 is widely and constitutively expressed whereasHK2 predominates during embryogenesis and is not widelyexpressed in adult tissues but only in a limited numberof normal adult tissues [141] their different subcellulardistributions and kinetic properties reflect their differentmetabolic roles [141 142] Under physiological conditionsHK1 is predominantly bound to the outer mitochondrialmembrane where it primarily channels glucose toward gly-colysis [140] whereas HK2 is mainly soluble and controlsglycogen formation [140 141] In malignant tissues HK1and HK2 isoforms are glycolytic and overexpressed (HK1more than HK2) (httpwwwproteinatlascom) [143] andare tightly bound to mitochondria HK1 and HK2 bindthrough theirN-terminal hydrophobic regions to the voltage-dependent calcium channel isoform1 (VDAC1) the prevalentpore protein in the outer mitochondrial membrane [2482 143ndash146] VDAC1 is also overexpressed in cancer cells[147] and the binding of HK to VDAC1 on the outer mito-chondrial membrane is a fundamental aspect of the aerobicglycolytic metabolism of cancer cells [82 135 147 148]HKVDAC association appears to protect tumor cells frommitochondrial outer membrane permeabilization (MOMP)which marks a point of no return leading to cell death[149] VDAC transports ADP and inorganic phosphate (Pi)the substrates needed for the production of ATP into themitochondria it also transports ATP out of themitochondriainto the cytoplasm and facilitates the availability of ATPto HK2 [143 150 151] A continuous supply of glucose toHK maintains this activity at a constant level and controlsthe flux through the mitochondrial electron transport chaininfluencing mitochondrial ROS production [152] HK bindsthrough hydrophobic interaction to VDAC1 the prevalentpore protein in the outer mitochondrial membrane [24 82143ndash146] thereby assuring and gaining a preferential accessto ATP [82 150 153] Translocation of HK2 from the cytosolto the mitochondria is regulated through phosphorylationof HK by Akt [141] HK2 only binds VDAC after phos-phorylation by Akt [151 152] Glycogen synthase kinase 3beta (GSK3120573) a major target of Akt kinases phosphorylatesVDAC [154ndash157] and dissociates HK [156] HK2 boundto mitochondria enhance its affinity to ATP and is lesssensitive to enzymatic inhibition by the G6P product [140141 158] HK2 can produce G6P from cytoplasmic glucoseand mitochondrial ATP without regulatory restraints evenunder hypoxia supporting rapid cell growth [142 143 159ndash161]HK1 andHK2 aremore tightly bound toVDAC in cancercells than in nonmalignant cells [156 162 163] HKVDA1interaction is relatively stable for HK1 while it is dependentupon G6P levels for HK2 [140 141] The higher affinity ofmitochondrial HK2 for glucose ATP and VDAC1 representsa high advantage for cancer cells survival and growth con-ferring to them an aerobic glycolytic phenotype underlyingthe Warburg effect [82 135 138 142 147 148 150 160]Differential interactions of HK1 and HK2 with mitochondriamay underlie different glycolytic profiles in cancer cells [164165] As rate-controlling enzymes in the glycolyic pathway
International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 International Journal of Cell Biology
pathway The classical extrinsic pathway is triggered by thebinding of extracellular signals like tumor necrosis factor(TNF) tumor necrosis factor-related apoptosis-inducing lig-and (TRAIL) and apoptosis antigen-1 (Fas) ligand (FASL)to death receptors such as tumor necrosis factor receptor 1(TNFR1) and FAS [80]This interaction leads to procaspase-8activation which cleaves itself into caspase-8Then caspase-8can cleave and activate procaspase-3 into caspase-3 thatcleaves many substrates inside the cell inducing the typicalmorphological changes seen in apoptosis The intrinsic path-way can be triggered by intracellular stress signals such asoxidative stress and DNA damage The diverse signals aresensed by mitochondria through the balance between pro-and antiapoptotic members of the Bcl-2 family of proteinsWhen proapoptotic signals overcome antiapoptotic signalsmitochondria becomepermeabilized leading to dissipation ofthe mitochondrial electrochemical potential release of inter-membrane proteins like cytochrome 119888 SmacDIABLO AIFandHTRA2OMI Cytochrome 119888 interacts with procaspase-9and Apaf-1 to form the apoptosome complex that leads tocaspase-3 activation [81] The extrinsic pathway can connectwith the intrinsic pathway through the cleavage by caspase-8of Bid into tBid (truncated Bid) which is a proapoptoticmember of the Bcl-2 family and can lead to mitochondriapermeabilization [82 83] The members of the Bcl-2 fam-ily regulate mitochondria permeabilization and comprisesantiapoptotic (Bcl-2 Bcl-xL) proapoptotic (Bax Bak) andproapoptotic BH-3 only proteins (Bad Bid Puma andNoxa)[82ndash84] One of the mechanisms of mitochondria perme-abilization is controlled by Bax and Bak proteins that arenormally expressed in every cell but are maintained inhibitedby the antiapoptotic members When there is a signal toapoptosis BH3-only proteins interact with the antiapoptoticmembers releasing BaxBak or directly activating themleading to their oligomerization and to mitochondrial outermembrane permeabilization (MOMP) Another mechanismfor mitochondria permeabilization is the formation of themitochondrial permeability transition (MPT) pore which iscomposed by the voltage-dependent anion channel VDACthe adenine nucleotide translocase (ANT) and cyclophilin D[85]TheMPT pore comprising VDAC1 is directly regulatedby Bad [86] In contrast VDAC2 has been identified as anantiapoptotic negative regulator of Bak [87ndash89]
One of the hallmarks of cancer is resistance to cell death[90] This resistance is acquired by a host of different geneticdefects in cancer cells such as those present in the p53tumor suppressor gene [62 68 91] Normal p53 responds togenotoxic oncogenic and other stress signals by inducingantiproliferative transcriptional programs leading to growtharrest (for cell repair) or apoptosis [92] p53 mediates cyclearrest by transcriptionally activating the p21 (CDKN1A) 14-3-3120590 (SFN) and GADD45120572 genes whereas p53-dependentapoptosis is triggered by turning on the synthesis of proteinsthat produce ROS [58 93] and transactivating differentmitochondrial proapoptotic genes as well as death receptorsfor the synthesis of the corresponding proteins [58 94]Whenp53 (and its target genes) expression levels are below a thresh-old they induce cell cycle arrest but not apoptosis above thisthreshold p53 trigger apoptosis [95] Lowering this threshold
with inhibitors of the antiapoptotic Bcl-2 family proteinssensitize cells to p53-induced apoptosis [95] By damagingcells chemotherapeutical drugs turn on p53rsquos transcriptionalfunction that upregulates the expression of proapoptotic pro-teins Defects in the p53 function or in the apoptotic cascadeslead to cell phenotypes resistant to chemotherapy Apoptosiscan also be induced by anticancer agents independently ofp53 by either directly modulating the apoptotic machineryor directly interacting with mitochondria [96] ROS levelsmight be playing a central role in mediating various forms ofnonapoptotic caspase-independent programmed cell deatheither through direct irreversible oxidative damage to keyproteins nucleic acid molecules and cellular structure or bythe activation of prodeath signal transduction programs [97]In a ROS-dependent nonapoptotic cell death pathway respi-ration through the mitochondrial electron transport system(ETS) chain is not essential and ROS is generated throughNADPH oxidases [97] In contrast to the mitochondrial ETSwhich generates ROS as a byproduct of respiration NADPHoxidases produceROS as their primary function [98] Raise ofROS is characteristic after ionizing radiation chemotherapyand targeted therapy treatments [58 99] and may be killingcancer cells through a nonapoptotic ROS-dependent celldeath pathway
Tumor cells maintain high ATPADP as well asNADHNAD+ ratios so as to assure that ATP levels arenever limited ATP depletion would represent then anunsustainable metabolic stress for tumor cells and it isassociated with necrotic death in some cell types [100]Recently it was shown that a signaling cascade centered onthe proteins RIP1 and RIP3 can trigger cell death by necrosisin many situations and the RIP1-RIP3 complex appears topromote ATP depletion during necroptosis by impingingon components of the MPT pore [101 102] Necrosis was aterm used to describe a process of accidental cell death inwhich cells swell and burst releasing its intracellular contentwhereas necroptosis modernly refers rather to an orderedcell death program different from apoptosis apoptosis is acaspase-dependent cell death process whereas necrosis iscaspase-independent process [102 103] Studies concerningnecrosis have focusedmainly on cell membrane permeabilityto propidium iodide (PI) in the flow citometry doublestaining with annexin-VPI
Fingrut et al [17] by studying the effect of MJ ontwo clones of a highly malignant B-cell lymphoma (oneharboring a normal wild type (wt) p53 gene while the otherexpressed a mutated inactive p53 protein) (Table 1) reportedthat MJ was equally toxic to both clones [17] In contrastthe p53-mutant cells were resistant to radiomimetics (drugshaving effects similar to those produced in radiotherapy)and chemotherapeutic drugs In this cancer cell model MJinduced mostly apoptotic cell death in the wt p53-expressinglymphoma cells while no signs of early apoptosis weredetected in the mutant p53-expressing lymphoma cells thatwere killed by MJ through a p53-independent nonapoptoticcell death mechanism However other cancer cell types canundergo apoptosis with or without functional p53 and aswe will see MJ can induce apoptotic andor nonapoptoticcell death in wt p53 as well as in TP53 mutated cells
International Journal of Cell Biology 9
(Table 1) Nonapoptotic cell death could be the consequenceof the severe bioenergetic damages induced by the directinteraction of MJ with mitochondria leading to increasedROS levels inhibition of ATP synthesis and ATP depletion[20 30] Jasmonates including MJ and some of its analogshave been found to be strongly proapoptotic in vitro indifferent types of cancer cells by both the extrinsic and theintrinsic pathway (Table 1) with no or little cytotoxicity tonormal cells [15 25 36 37 44 45] Thus MJ induced theexpression of TNFR1 in human breast MCF-7 and MDA-MB-435 cells [25 74] and in hormone-independent humanprostate PC-3 and DU 145 cancer cells promoting apoptosisby the extrinsic pathway in these cells [48] TRAIL (TNF-related apoptosis inducing ligand) is able to induce apoptosisin prostate cancer cell yet overexpression of antiapoptoticproteins and inhibition of proapoptotic proteins results indiminishing the TRAIL-mediated apoptosis pathway TRAILhas emerged as a potent anticancer agent in laboratory-based studies and preclinical trials and JAs have been foundto sensitize cancer cells to TRAIL-mediated apoptosis [32](Table 3) J7 a synthetic analogue of MJ enhanced TRAIL-mediated apoptosis through upregulation of ROS levels inhuman hepatomaHepG2 cells apoptosis was induced via Bidcleavage downregulation of X-linked inhibitor of apoptosisprotein (XIAP) cellular inhibitor of apoptosis-1 (cIAP-1)B-cell lymphoma-extra large (Bcl-xL) and activation ofcaspases [35] Other examples of MJ-mediated apoptoticandor nonapoptotic cell death in different cancer cell types(Table 1) are as follows (1) MJ induced ROS production inA549 lung adenocarcinoma cells activating Bcl-2-associated-X-protein (Bax) as well as Bcl-2-Like-1 (Bcl2L1) proapoptoticproteins in these cells [36] (2) MJ activated caspase-3 inleukemia Molt-4 prostate SK28 and LNCaP cancer cells[15] (3) MJ inhibited the antiapoptotic Bcl-2 protein andenhanced proapoptotic proteins in radio-resistant humanprostate cancer cell line PC-3 [104] (4) MJ suppressed theproliferation of the neuroblastoma cell lines SH-SY5Y SK-N-SH and BE(2)-C through a strong apoptosis-inducingeffect as measured by annexin VFITCPI MJ treatment alsodownregulated the expression of the antiapoptotic proteinsXIAP and survivin [44 45] (5) MJ-mediated cell death wasobserved on cervical cancer and myeloma cells and it wasreported to be due to mixed characteristics of apoptosis andnecrosis [26 105] So far studies showing MJ inducing theRIP1 signaling cascade have not been reported howeverit will be important to deep into these analyses to cleardistinguish between MJ-induced apoptosis andor necrosis
23 Methyl Jasmonate Induces the Production of ReactiveOxygen Species (ROS) Responses in Some Cancer Cell TypesLeading Also to Cell Death Intracellular ROS levels reg-ulate cell cycle activity cell metabolism and survival cellproliferation and apoptosis [53ndash58] Stressful conditionsproduce a transient excess of intracellular ROS in normaltissues however under chronic pathological and oxidativestress conditions ROS is continuously produced in excessand persistently causing damage to DNA cell membraneslipids proteins and inducing apoptosis to control and avoid
oxidative damages cells counteract excess ROS by enzymatic(superoxide dismutase (SOD) glutathione peroxidase (GPx)and catalase) andor nonenzymatic (glutathione thoredoxin)antioxidant elements [99 106] Controlling ROS is central tointracellular redox signaling and homeostasis on the otherhand uncontrolledunbalanced ROS production modifiesintracellular redox leading to apoptosis [58 82 99 106] ROScan be generated by different biological systems (1) director indirect extracellular stimulation of plasma membrane-bound NAD(P)H oxidases (NOX) can generate intracellularH2O2[57 107ndash110] (2) biochemical pathways associated to
fatty acid oxidation and lipoxygenases activities generateROS as normal byproducts of these activities [111] (3)ATP generation by oxidative phosphorylation (OxPhos) inmitochondria is accompanied by the production of ROS as aconsequence of electron leakage from the electron transportchain mitochondria are considered to be major sources ofROS inmammalian cells particularly under a variety of stress-ful conditions [82 99] Superoxide (O
2
minus) is mainly produced
from complexes I and III and it is rapidly dismutated to H2O2
and oxygen by SOD [112] H2O2serves both as a toxic oxidant
and as an essential signaling molecule regulating cellularbiological processes [113] A strong positive correlation existsbetween mitochondrial membrane potential (Δ120595
119898) and ROS
production [106 114 115] Mitochondria produce more ROSat high membrane potential and an increase in the Δ120595
119898
produced either by a closure of MPT pore or by inhibitionof the ATP synthase is associated with increased ROS pro-duction However in cases of mitochondrial dysfunctionslow Δ120595
119898and decreased activity of the respiratory chain
are accompanied by a simultaneous increase in ROS [116117] It has been reported that cancer cells have higher ROScontent compared to normal cells [118 119] probably due toabnormal respiration because of dysfunctional mutations inmitochondria [120] but they also have increased contents ofantioxidants (SOD catalase glutathione thioredoxin etc)to counteract ROS [58 121] Tumor cells cannot tolerateexcessive high ROS levels [118 122] and as a matter of factradiation therapy chemotherapies and targeted therapiesinduce apoptosis and kill some cancer cells by generatingexcess ROS [99 106 123 124] Thus inducing higher levelsof ROS in cancers cells can be exploited as an effectiveselective strategy to kill tumor cells over normal cells [119]However prolonged treatment with the same drug reducesROS levels in cancer cells [124] indicating that when theybecome drug-resistant tumor cells have lower ROS contentthan drug-sensitive cancer cells Evidence suggests that drug-resistant cells have a higher expression of catalase at theplasma membrane that keeps reduced ROS levels [121]
In plants MJ-mediated damage to mitochondria is fol-lowed by a rapid production of H
2O2[125] H
2O2is used as
a second messenger for the induction of genes and productsrelated to defense against both herbivores and pathogens[126] in addition H
2O2induces apoptosis and senescence
[127 128] On the other hand it has been signaled that theMJ-induced production of H
2O2in plants was prevented by
inhibitors of NAD(P)H oxidase [128 129] indicating anothermechanism of MJ-mediated production of ROS and a majorrole for this enzyme in MJ-induced ROS responses MJ
10 International Journal of Cell Biology
induces also an endogenous H2O2response in some cancer
cell lines (Table 1) [19 36 38 130ndash132] In A549 cancerlung cells the H
2O2response induced by MJ increased the
expression of the proapoptotic Bax and Bcl-Xs (although notbeing that of the antiapoptotic Bcl-2 and Bcl-xL proteins) andthis led to apoptosis this event was inhibited by catalase (aspecific inhibitor of H
2O2) [36] MJ increased the intracel-
lular levels of H2O2and superoxide (O
2) ions in human C6
glioma cells in vitro and induced the expression of heat shockprotein 72 (HSP72) a negative regulator of H
2O2-induced
oxidative stress in these cells in vitro MJ-induced HSP72expression was blocked by ROS inhibitors [130] ElevatedROS generation was observed in response to both MJ andJA levels in the acute myeloid leukemia HL-60 and KG1a celllines but other factors like the MJ-mediated generation ofmitochondrial SOD rather than ROS were determinant forapoptosis or differentiation in these cells [132] Suppression ofMJ-induced apoptosis by antioxidants like N-acetylcysteine(NAC) and catalase but not by inhibitors of hydroxyl radicalsand superoxide ions indicates that H
2O2is nonetheless one
of the important factors involved inMJ anticancer effects [19]
24 By Dissociating Hexokinase (HK) from the VoltageDependent Anion Channel (VDAC) on the Outer Mitochon-drial Membrane MJ Dissociates Glycolysis from OxidativePhosphorylation Causing Severe Bioenergetic Deregulations inCancer Cells Metabolism and cell growth are two cellularprocesses that are tighly linked and regulated [133] Theclassic bioenergetics phenotype of cancer cells of enhancedglycolysis under aerobic conditions was described by War-burg [134] However the ldquoWarburg effectrdquo does not neces-sarily imply mitochondrial dysfunction [135] To increaseglycolysis cancer cells upregulate the transcription of genesinvolved in the glycolytic pathway (ie glucose transportersglycolytic enzymes etc) This metabolic reprogrammingallows increased rate of ATP production synthesis of lipidsand a new redox balance Cancer cells actually use a com-bination of both glycolysis and mitochondrial respirationto produce energy and they may vary in regards to thepreferential use of these pathways being in some caseseither more glycolyticless oxidative or less glycolyticmoreoxidative depending on the prevalent normoxic or hypoxicenvironmental conditions and their capacities to expressadequate levels of oncogenes and tumor suppressor geneproducts for cell growth [136ndash139] or when either glycolysisor OxPhos is inhibited in whose case there can be partialcompensation by the other metabolic pathway [17] Shiftingthe balance between those two processes with glycolyticandor mitochondrial enzyme inhibitors might represent aninteresting way to look for new anticancer agents
The first step in the metabolism of glucose is catalyzedby the enzyme hexokinase (HK) which by phosphorylatingglucose makes it negative sequestering it into the cytoplasmin the form of glucose-6-phosphate (G6P) In normal tissuesthis crucial step is catalyzed by four different HK isoforms(HK1 HK2 HK3 and glucokinase) indicating that regulationof glucose phosphorylation can vary in different tissues underdifferent condition [140] HK1 and HK2 have overlapping
tissue expression but different subcellular distributions withHK1 associated mainly with mitochondria and HK2 associ-atedwith bothmitochondrial and cytoplasmic compartments[141] HK1 is widely and constitutively expressed whereasHK2 predominates during embryogenesis and is not widelyexpressed in adult tissues but only in a limited numberof normal adult tissues [141] their different subcellulardistributions and kinetic properties reflect their differentmetabolic roles [141 142] Under physiological conditionsHK1 is predominantly bound to the outer mitochondrialmembrane where it primarily channels glucose toward gly-colysis [140] whereas HK2 is mainly soluble and controlsglycogen formation [140 141] In malignant tissues HK1and HK2 isoforms are glycolytic and overexpressed (HK1more than HK2) (httpwwwproteinatlascom) [143] andare tightly bound to mitochondria HK1 and HK2 bindthrough theirN-terminal hydrophobic regions to the voltage-dependent calcium channel isoform1 (VDAC1) the prevalentpore protein in the outer mitochondrial membrane [2482 143ndash146] VDAC1 is also overexpressed in cancer cells[147] and the binding of HK to VDAC1 on the outer mito-chondrial membrane is a fundamental aspect of the aerobicglycolytic metabolism of cancer cells [82 135 147 148]HKVDAC association appears to protect tumor cells frommitochondrial outer membrane permeabilization (MOMP)which marks a point of no return leading to cell death[149] VDAC transports ADP and inorganic phosphate (Pi)the substrates needed for the production of ATP into themitochondria it also transports ATP out of themitochondriainto the cytoplasm and facilitates the availability of ATPto HK2 [143 150 151] A continuous supply of glucose toHK maintains this activity at a constant level and controlsthe flux through the mitochondrial electron transport chaininfluencing mitochondrial ROS production [152] HK bindsthrough hydrophobic interaction to VDAC1 the prevalentpore protein in the outer mitochondrial membrane [24 82143ndash146] thereby assuring and gaining a preferential accessto ATP [82 150 153] Translocation of HK2 from the cytosolto the mitochondria is regulated through phosphorylationof HK by Akt [141] HK2 only binds VDAC after phos-phorylation by Akt [151 152] Glycogen synthase kinase 3beta (GSK3120573) a major target of Akt kinases phosphorylatesVDAC [154ndash157] and dissociates HK [156] HK2 boundto mitochondria enhance its affinity to ATP and is lesssensitive to enzymatic inhibition by the G6P product [140141 158] HK2 can produce G6P from cytoplasmic glucoseand mitochondrial ATP without regulatory restraints evenunder hypoxia supporting rapid cell growth [142 143 159ndash161]HK1 andHK2 aremore tightly bound toVDAC in cancercells than in nonmalignant cells [156 162 163] HKVDA1interaction is relatively stable for HK1 while it is dependentupon G6P levels for HK2 [140 141] The higher affinity ofmitochondrial HK2 for glucose ATP and VDAC1 representsa high advantage for cancer cells survival and growth con-ferring to them an aerobic glycolytic phenotype underlyingthe Warburg effect [82 135 138 142 147 148 150 160]Differential interactions of HK1 and HK2 with mitochondriamay underlie different glycolytic profiles in cancer cells [164165] As rate-controlling enzymes in the glycolyic pathway
International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Cell Biology 9
(Table 1) Nonapoptotic cell death could be the consequenceof the severe bioenergetic damages induced by the directinteraction of MJ with mitochondria leading to increasedROS levels inhibition of ATP synthesis and ATP depletion[20 30] Jasmonates including MJ and some of its analogshave been found to be strongly proapoptotic in vitro indifferent types of cancer cells by both the extrinsic and theintrinsic pathway (Table 1) with no or little cytotoxicity tonormal cells [15 25 36 37 44 45] Thus MJ induced theexpression of TNFR1 in human breast MCF-7 and MDA-MB-435 cells [25 74] and in hormone-independent humanprostate PC-3 and DU 145 cancer cells promoting apoptosisby the extrinsic pathway in these cells [48] TRAIL (TNF-related apoptosis inducing ligand) is able to induce apoptosisin prostate cancer cell yet overexpression of antiapoptoticproteins and inhibition of proapoptotic proteins results indiminishing the TRAIL-mediated apoptosis pathway TRAILhas emerged as a potent anticancer agent in laboratory-based studies and preclinical trials and JAs have been foundto sensitize cancer cells to TRAIL-mediated apoptosis [32](Table 3) J7 a synthetic analogue of MJ enhanced TRAIL-mediated apoptosis through upregulation of ROS levels inhuman hepatomaHepG2 cells apoptosis was induced via Bidcleavage downregulation of X-linked inhibitor of apoptosisprotein (XIAP) cellular inhibitor of apoptosis-1 (cIAP-1)B-cell lymphoma-extra large (Bcl-xL) and activation ofcaspases [35] Other examples of MJ-mediated apoptoticandor nonapoptotic cell death in different cancer cell types(Table 1) are as follows (1) MJ induced ROS production inA549 lung adenocarcinoma cells activating Bcl-2-associated-X-protein (Bax) as well as Bcl-2-Like-1 (Bcl2L1) proapoptoticproteins in these cells [36] (2) MJ activated caspase-3 inleukemia Molt-4 prostate SK28 and LNCaP cancer cells[15] (3) MJ inhibited the antiapoptotic Bcl-2 protein andenhanced proapoptotic proteins in radio-resistant humanprostate cancer cell line PC-3 [104] (4) MJ suppressed theproliferation of the neuroblastoma cell lines SH-SY5Y SK-N-SH and BE(2)-C through a strong apoptosis-inducingeffect as measured by annexin VFITCPI MJ treatment alsodownregulated the expression of the antiapoptotic proteinsXIAP and survivin [44 45] (5) MJ-mediated cell death wasobserved on cervical cancer and myeloma cells and it wasreported to be due to mixed characteristics of apoptosis andnecrosis [26 105] So far studies showing MJ inducing theRIP1 signaling cascade have not been reported howeverit will be important to deep into these analyses to cleardistinguish between MJ-induced apoptosis andor necrosis
23 Methyl Jasmonate Induces the Production of ReactiveOxygen Species (ROS) Responses in Some Cancer Cell TypesLeading Also to Cell Death Intracellular ROS levels reg-ulate cell cycle activity cell metabolism and survival cellproliferation and apoptosis [53ndash58] Stressful conditionsproduce a transient excess of intracellular ROS in normaltissues however under chronic pathological and oxidativestress conditions ROS is continuously produced in excessand persistently causing damage to DNA cell membraneslipids proteins and inducing apoptosis to control and avoid
oxidative damages cells counteract excess ROS by enzymatic(superoxide dismutase (SOD) glutathione peroxidase (GPx)and catalase) andor nonenzymatic (glutathione thoredoxin)antioxidant elements [99 106] Controlling ROS is central tointracellular redox signaling and homeostasis on the otherhand uncontrolledunbalanced ROS production modifiesintracellular redox leading to apoptosis [58 82 99 106] ROScan be generated by different biological systems (1) director indirect extracellular stimulation of plasma membrane-bound NAD(P)H oxidases (NOX) can generate intracellularH2O2[57 107ndash110] (2) biochemical pathways associated to
fatty acid oxidation and lipoxygenases activities generateROS as normal byproducts of these activities [111] (3)ATP generation by oxidative phosphorylation (OxPhos) inmitochondria is accompanied by the production of ROS as aconsequence of electron leakage from the electron transportchain mitochondria are considered to be major sources ofROS inmammalian cells particularly under a variety of stress-ful conditions [82 99] Superoxide (O
2
minus) is mainly produced
from complexes I and III and it is rapidly dismutated to H2O2
and oxygen by SOD [112] H2O2serves both as a toxic oxidant
and as an essential signaling molecule regulating cellularbiological processes [113] A strong positive correlation existsbetween mitochondrial membrane potential (Δ120595
119898) and ROS
production [106 114 115] Mitochondria produce more ROSat high membrane potential and an increase in the Δ120595
119898
produced either by a closure of MPT pore or by inhibitionof the ATP synthase is associated with increased ROS pro-duction However in cases of mitochondrial dysfunctionslow Δ120595
119898and decreased activity of the respiratory chain
are accompanied by a simultaneous increase in ROS [116117] It has been reported that cancer cells have higher ROScontent compared to normal cells [118 119] probably due toabnormal respiration because of dysfunctional mutations inmitochondria [120] but they also have increased contents ofantioxidants (SOD catalase glutathione thioredoxin etc)to counteract ROS [58 121] Tumor cells cannot tolerateexcessive high ROS levels [118 122] and as a matter of factradiation therapy chemotherapies and targeted therapiesinduce apoptosis and kill some cancer cells by generatingexcess ROS [99 106 123 124] Thus inducing higher levelsof ROS in cancers cells can be exploited as an effectiveselective strategy to kill tumor cells over normal cells [119]However prolonged treatment with the same drug reducesROS levels in cancer cells [124] indicating that when theybecome drug-resistant tumor cells have lower ROS contentthan drug-sensitive cancer cells Evidence suggests that drug-resistant cells have a higher expression of catalase at theplasma membrane that keeps reduced ROS levels [121]
In plants MJ-mediated damage to mitochondria is fol-lowed by a rapid production of H
2O2[125] H
2O2is used as
a second messenger for the induction of genes and productsrelated to defense against both herbivores and pathogens[126] in addition H
2O2induces apoptosis and senescence
[127 128] On the other hand it has been signaled that theMJ-induced production of H
2O2in plants was prevented by
inhibitors of NAD(P)H oxidase [128 129] indicating anothermechanism of MJ-mediated production of ROS and a majorrole for this enzyme in MJ-induced ROS responses MJ
10 International Journal of Cell Biology
induces also an endogenous H2O2response in some cancer
cell lines (Table 1) [19 36 38 130ndash132] In A549 cancerlung cells the H
2O2response induced by MJ increased the
expression of the proapoptotic Bax and Bcl-Xs (although notbeing that of the antiapoptotic Bcl-2 and Bcl-xL proteins) andthis led to apoptosis this event was inhibited by catalase (aspecific inhibitor of H
2O2) [36] MJ increased the intracel-
lular levels of H2O2and superoxide (O
2) ions in human C6
glioma cells in vitro and induced the expression of heat shockprotein 72 (HSP72) a negative regulator of H
2O2-induced
oxidative stress in these cells in vitro MJ-induced HSP72expression was blocked by ROS inhibitors [130] ElevatedROS generation was observed in response to both MJ andJA levels in the acute myeloid leukemia HL-60 and KG1a celllines but other factors like the MJ-mediated generation ofmitochondrial SOD rather than ROS were determinant forapoptosis or differentiation in these cells [132] Suppression ofMJ-induced apoptosis by antioxidants like N-acetylcysteine(NAC) and catalase but not by inhibitors of hydroxyl radicalsand superoxide ions indicates that H
2O2is nonetheless one
of the important factors involved inMJ anticancer effects [19]
24 By Dissociating Hexokinase (HK) from the VoltageDependent Anion Channel (VDAC) on the Outer Mitochon-drial Membrane MJ Dissociates Glycolysis from OxidativePhosphorylation Causing Severe Bioenergetic Deregulations inCancer Cells Metabolism and cell growth are two cellularprocesses that are tighly linked and regulated [133] Theclassic bioenergetics phenotype of cancer cells of enhancedglycolysis under aerobic conditions was described by War-burg [134] However the ldquoWarburg effectrdquo does not neces-sarily imply mitochondrial dysfunction [135] To increaseglycolysis cancer cells upregulate the transcription of genesinvolved in the glycolytic pathway (ie glucose transportersglycolytic enzymes etc) This metabolic reprogrammingallows increased rate of ATP production synthesis of lipidsand a new redox balance Cancer cells actually use a com-bination of both glycolysis and mitochondrial respirationto produce energy and they may vary in regards to thepreferential use of these pathways being in some caseseither more glycolyticless oxidative or less glycolyticmoreoxidative depending on the prevalent normoxic or hypoxicenvironmental conditions and their capacities to expressadequate levels of oncogenes and tumor suppressor geneproducts for cell growth [136ndash139] or when either glycolysisor OxPhos is inhibited in whose case there can be partialcompensation by the other metabolic pathway [17] Shiftingthe balance between those two processes with glycolyticandor mitochondrial enzyme inhibitors might represent aninteresting way to look for new anticancer agents
The first step in the metabolism of glucose is catalyzedby the enzyme hexokinase (HK) which by phosphorylatingglucose makes it negative sequestering it into the cytoplasmin the form of glucose-6-phosphate (G6P) In normal tissuesthis crucial step is catalyzed by four different HK isoforms(HK1 HK2 HK3 and glucokinase) indicating that regulationof glucose phosphorylation can vary in different tissues underdifferent condition [140] HK1 and HK2 have overlapping
tissue expression but different subcellular distributions withHK1 associated mainly with mitochondria and HK2 associ-atedwith bothmitochondrial and cytoplasmic compartments[141] HK1 is widely and constitutively expressed whereasHK2 predominates during embryogenesis and is not widelyexpressed in adult tissues but only in a limited numberof normal adult tissues [141] their different subcellulardistributions and kinetic properties reflect their differentmetabolic roles [141 142] Under physiological conditionsHK1 is predominantly bound to the outer mitochondrialmembrane where it primarily channels glucose toward gly-colysis [140] whereas HK2 is mainly soluble and controlsglycogen formation [140 141] In malignant tissues HK1and HK2 isoforms are glycolytic and overexpressed (HK1more than HK2) (httpwwwproteinatlascom) [143] andare tightly bound to mitochondria HK1 and HK2 bindthrough theirN-terminal hydrophobic regions to the voltage-dependent calcium channel isoform1 (VDAC1) the prevalentpore protein in the outer mitochondrial membrane [2482 143ndash146] VDAC1 is also overexpressed in cancer cells[147] and the binding of HK to VDAC1 on the outer mito-chondrial membrane is a fundamental aspect of the aerobicglycolytic metabolism of cancer cells [82 135 147 148]HKVDAC association appears to protect tumor cells frommitochondrial outer membrane permeabilization (MOMP)which marks a point of no return leading to cell death[149] VDAC transports ADP and inorganic phosphate (Pi)the substrates needed for the production of ATP into themitochondria it also transports ATP out of themitochondriainto the cytoplasm and facilitates the availability of ATPto HK2 [143 150 151] A continuous supply of glucose toHK maintains this activity at a constant level and controlsthe flux through the mitochondrial electron transport chaininfluencing mitochondrial ROS production [152] HK bindsthrough hydrophobic interaction to VDAC1 the prevalentpore protein in the outer mitochondrial membrane [24 82143ndash146] thereby assuring and gaining a preferential accessto ATP [82 150 153] Translocation of HK2 from the cytosolto the mitochondria is regulated through phosphorylationof HK by Akt [141] HK2 only binds VDAC after phos-phorylation by Akt [151 152] Glycogen synthase kinase 3beta (GSK3120573) a major target of Akt kinases phosphorylatesVDAC [154ndash157] and dissociates HK [156] HK2 boundto mitochondria enhance its affinity to ATP and is lesssensitive to enzymatic inhibition by the G6P product [140141 158] HK2 can produce G6P from cytoplasmic glucoseand mitochondrial ATP without regulatory restraints evenunder hypoxia supporting rapid cell growth [142 143 159ndash161]HK1 andHK2 aremore tightly bound toVDAC in cancercells than in nonmalignant cells [156 162 163] HKVDA1interaction is relatively stable for HK1 while it is dependentupon G6P levels for HK2 [140 141] The higher affinity ofmitochondrial HK2 for glucose ATP and VDAC1 representsa high advantage for cancer cells survival and growth con-ferring to them an aerobic glycolytic phenotype underlyingthe Warburg effect [82 135 138 142 147 148 150 160]Differential interactions of HK1 and HK2 with mitochondriamay underlie different glycolytic profiles in cancer cells [164165] As rate-controlling enzymes in the glycolyic pathway
International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 International Journal of Cell Biology
induces also an endogenous H2O2response in some cancer
cell lines (Table 1) [19 36 38 130ndash132] In A549 cancerlung cells the H
2O2response induced by MJ increased the
expression of the proapoptotic Bax and Bcl-Xs (although notbeing that of the antiapoptotic Bcl-2 and Bcl-xL proteins) andthis led to apoptosis this event was inhibited by catalase (aspecific inhibitor of H
2O2) [36] MJ increased the intracel-
lular levels of H2O2and superoxide (O
2) ions in human C6
glioma cells in vitro and induced the expression of heat shockprotein 72 (HSP72) a negative regulator of H
2O2-induced
oxidative stress in these cells in vitro MJ-induced HSP72expression was blocked by ROS inhibitors [130] ElevatedROS generation was observed in response to both MJ andJA levels in the acute myeloid leukemia HL-60 and KG1a celllines but other factors like the MJ-mediated generation ofmitochondrial SOD rather than ROS were determinant forapoptosis or differentiation in these cells [132] Suppression ofMJ-induced apoptosis by antioxidants like N-acetylcysteine(NAC) and catalase but not by inhibitors of hydroxyl radicalsand superoxide ions indicates that H
2O2is nonetheless one
of the important factors involved inMJ anticancer effects [19]
24 By Dissociating Hexokinase (HK) from the VoltageDependent Anion Channel (VDAC) on the Outer Mitochon-drial Membrane MJ Dissociates Glycolysis from OxidativePhosphorylation Causing Severe Bioenergetic Deregulations inCancer Cells Metabolism and cell growth are two cellularprocesses that are tighly linked and regulated [133] Theclassic bioenergetics phenotype of cancer cells of enhancedglycolysis under aerobic conditions was described by War-burg [134] However the ldquoWarburg effectrdquo does not neces-sarily imply mitochondrial dysfunction [135] To increaseglycolysis cancer cells upregulate the transcription of genesinvolved in the glycolytic pathway (ie glucose transportersglycolytic enzymes etc) This metabolic reprogrammingallows increased rate of ATP production synthesis of lipidsand a new redox balance Cancer cells actually use a com-bination of both glycolysis and mitochondrial respirationto produce energy and they may vary in regards to thepreferential use of these pathways being in some caseseither more glycolyticless oxidative or less glycolyticmoreoxidative depending on the prevalent normoxic or hypoxicenvironmental conditions and their capacities to expressadequate levels of oncogenes and tumor suppressor geneproducts for cell growth [136ndash139] or when either glycolysisor OxPhos is inhibited in whose case there can be partialcompensation by the other metabolic pathway [17] Shiftingthe balance between those two processes with glycolyticandor mitochondrial enzyme inhibitors might represent aninteresting way to look for new anticancer agents
The first step in the metabolism of glucose is catalyzedby the enzyme hexokinase (HK) which by phosphorylatingglucose makes it negative sequestering it into the cytoplasmin the form of glucose-6-phosphate (G6P) In normal tissuesthis crucial step is catalyzed by four different HK isoforms(HK1 HK2 HK3 and glucokinase) indicating that regulationof glucose phosphorylation can vary in different tissues underdifferent condition [140] HK1 and HK2 have overlapping
tissue expression but different subcellular distributions withHK1 associated mainly with mitochondria and HK2 associ-atedwith bothmitochondrial and cytoplasmic compartments[141] HK1 is widely and constitutively expressed whereasHK2 predominates during embryogenesis and is not widelyexpressed in adult tissues but only in a limited numberof normal adult tissues [141] their different subcellulardistributions and kinetic properties reflect their differentmetabolic roles [141 142] Under physiological conditionsHK1 is predominantly bound to the outer mitochondrialmembrane where it primarily channels glucose toward gly-colysis [140] whereas HK2 is mainly soluble and controlsglycogen formation [140 141] In malignant tissues HK1and HK2 isoforms are glycolytic and overexpressed (HK1more than HK2) (httpwwwproteinatlascom) [143] andare tightly bound to mitochondria HK1 and HK2 bindthrough theirN-terminal hydrophobic regions to the voltage-dependent calcium channel isoform1 (VDAC1) the prevalentpore protein in the outer mitochondrial membrane [2482 143ndash146] VDAC1 is also overexpressed in cancer cells[147] and the binding of HK to VDAC1 on the outer mito-chondrial membrane is a fundamental aspect of the aerobicglycolytic metabolism of cancer cells [82 135 147 148]HKVDAC association appears to protect tumor cells frommitochondrial outer membrane permeabilization (MOMP)which marks a point of no return leading to cell death[149] VDAC transports ADP and inorganic phosphate (Pi)the substrates needed for the production of ATP into themitochondria it also transports ATP out of themitochondriainto the cytoplasm and facilitates the availability of ATPto HK2 [143 150 151] A continuous supply of glucose toHK maintains this activity at a constant level and controlsthe flux through the mitochondrial electron transport chaininfluencing mitochondrial ROS production [152] HK bindsthrough hydrophobic interaction to VDAC1 the prevalentpore protein in the outer mitochondrial membrane [24 82143ndash146] thereby assuring and gaining a preferential accessto ATP [82 150 153] Translocation of HK2 from the cytosolto the mitochondria is regulated through phosphorylationof HK by Akt [141] HK2 only binds VDAC after phos-phorylation by Akt [151 152] Glycogen synthase kinase 3beta (GSK3120573) a major target of Akt kinases phosphorylatesVDAC [154ndash157] and dissociates HK [156] HK2 boundto mitochondria enhance its affinity to ATP and is lesssensitive to enzymatic inhibition by the G6P product [140141 158] HK2 can produce G6P from cytoplasmic glucoseand mitochondrial ATP without regulatory restraints evenunder hypoxia supporting rapid cell growth [142 143 159ndash161]HK1 andHK2 aremore tightly bound toVDAC in cancercells than in nonmalignant cells [156 162 163] HKVDA1interaction is relatively stable for HK1 while it is dependentupon G6P levels for HK2 [140 141] The higher affinity ofmitochondrial HK2 for glucose ATP and VDAC1 representsa high advantage for cancer cells survival and growth con-ferring to them an aerobic glycolytic phenotype underlyingthe Warburg effect [82 135 138 142 147 148 150 160]Differential interactions of HK1 and HK2 with mitochondriamay underlie different glycolytic profiles in cancer cells [164165] As rate-controlling enzymes in the glycolyic pathway
International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Cell Biology 11
HKs represent important targets for anticancer therapies anddrug development HK inhibition induces cancer cells deathand it is reportedly successful in the treatment of cancer[166ndash169] In addition the association between HKs andVDAC provides another therapeutic target Peptides as wellas small molecules that disrupt the interaction between HKandmitochondria can selectively kill tumor cells both in vitroand in vivo [149 169]
One of the major findings concerning MJ is that itwas able to detach HK1 and HK2 from VDAC in a time-and dose-dependent manner in the mitochondrial fractionof murine colon carcinoma CT-36 cells human leukemicMolt-4 and murine BCL1 cells and murine B16 melanomatumor cells by specifically binding to HK2 as judged byHK immunochemical surface plasmon resonance and planarlipid bilayer VDAC-activity analyses without inhibiting thekinase activity [30] These authors emphasized that the sus-ceptibility of cancer cells andmitochondria to jasmonateswasdependent onHK overexpression and its association tomito-chondria [30] Dissociation of HK2 from VDAC changedmitochondrial membrane permeability induced cytochrome119888 release inhibited ATP synthesis blocked OxPhos causeda drastic reduction of intracellular ATP levels and stoppedATP-drivenmembrane pumps causing an overall irreversiblebioenergetic damage leadingmitochondria to swell and burstand to cell death (Figure 2) [17 20 30] The HK2VDACdisruption event dissociated glycolysis from OxPhos and inglycolytic cells such as for instance CaSki cervical [26] andmultiple myeloma [105] cells and MJ treatment induced anincrease in lactate production whereas at the mitochondriallevel cell death by the intrinsic apoptotic and also by non-apoptotic pathways (including necrosis) was promoted [149]probably related to increased ROS levels [163] (Figure 2)On the other hand MJ treatment induced an increase inphospho(p)-Akt levels in sarcoma cell lines and activation ofthe PI3KAkt pathway attenuated the cytotoxic effect of MJ[49] This effect was blocked by PI3KAkt pathway inhibitorsand by 2-deoxy-D-glucose (2DG) (Table 3) inducing sensiti-zation toward MJ cytotoxicity in a synergistic manner [49]
MJ-induced ATP depletion was independent of pyru-vate (an OxPhos substrate) and oligomycin (an OxPhosinhibitor) [17] but it was inhibited by glucose (a glycoly-sis substrate) nonetheless 2-deoxy-D-glucose (a glycolysisinhibitor) synergistically enhanced the anticancer effect ofMJ[170] (Table 3)The above describedMJ-induced bioenergeticeffects were observed only in mitochondria isolated fromcancer cells but not in those isolated from normal cells[19 38 170] supporting the general idea that MJ interactswith molecular structures on the outer mitochondrial sur-face of cancer cells that may be exposed in a substantiallydifferent manner than in non-cancerous cells (for instancethe HK2VDAC complex) [24 30 149 163]
25 Jasmonates Stimulate the MAPK Stress Pathways andThisEffect Leads to Differentiation in Some Cancer Cell Types andto Apoptosis in Others MAPK pathways are critical for con-verting diverse extracellular stress signals (ROS mitogensmediators of inflammation and other stressors) to biological
responses such as cell growth survival proliferation inflam-matory responses apoptosis and differentiation [171] Themain MAPK subfamilies of signaling pathways include thestress-regulated c-Jun NH
2-terminal kinase (JNK) the p38-
MAPK and the extracellular signal-regulated kinase-12(ERK12) pathways ERK12 behaves mainly as mitogen-activated proliferationdifferentiation factors [172] whereasJNK and p38-MAPK pathways stimulates the AP-1 tran-scription factor and regulate responses for cell survival andinflammation andmediate cell death resulting from exposureto various stressors stimuli [173ndash175]
MJ was able to activate MAPK pathways in different can-cer cells types (Table 1) (1) In acute human T-lymphoblasticMolt-4 leukemic cells MJ activated JNK and p38-MAPKstress responses resulting in AP-1 activity however thissignaling did not mediate the observed apoptotic leukemiccell death [18] (2) In A549 lung carcinoma cells a similareffect was seen [19 36] The effect of MJ in these two caseswas independent of JNK and p38 activities and AP-1 did notmediate the apoptosis effect on these cells and is independentof RNA and protein synthesis [18] Thus MJ induced twoindependent processes in Molt-4 leukemic cells apoptoticdeath and a typical MAPKJNK and p38 stress response [18](3) MJ also activated the p38-MAPK and ERK12 pathways inmelanocytic MDA-MB-435 cells but not in the human ade-nocarcinoma breast cell lineMCF-7 [25 74] (4) MJ activatedthe MAPK pathway in the human myelocytic leukemia cellline HL-60 but the outcome of this event was cell differen-tiation rather than apoptosis [41] the differentiation effectwas inhibited by PD98059 a MEK1 inhibitor confirmingthe involvement of MAPKERK pathway in this MJ-mediateactivity [41] Acute myeloid leukemia (AML) is one of theworst forms of leukemia (AML blasts are immature myeloidcells) because it is largely resistant to chemotherapeuticdrugs and other forms of therapy [176] These cells can beinduced to differentiate into normal functional myeloid cellsby different chemicals unfortunately many of these chemi-cals are too toxic to be used clinically (httpwwwcancerorgacsgroupsciddocumentswebcontent003110-pdfpdf) MJ(04mM) stopped the growth of these blast cells and pro-moted their differentiation to normal cells as deduced fromthe expression of markers of differentiation such as NBTreduction (for myelomonocytic differentiation) morpholog-ical differentiation into granulocytes and expression of CD14(monocyte-specific) and CD15 (granulocyte-specific) surfaceantigens [41] MJ induced also the expression of the Ca2+-binding protein S100P in these cells reported to be linked toinduction of differentiation (Table 1) [42]
26 Methyl Jasmonate and Some of Its Synthetic DerivativesShowed Anti-Inflammatory Antiangiogenic and Antimeta-static Activities The relationship between inflammation andcancer has beenwidely accepted particularly chronic inflam-mation is a key component where ROS is generated creatinga tumormicroenvironment that promotes tumor progression[177] Relevant inflammation targets in cancer include COX-2 5-lipoxygenase (5-LOX) and inflammation factors (eginflammatory cytokines TNF IL-1 IL-6 and chemokines)
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
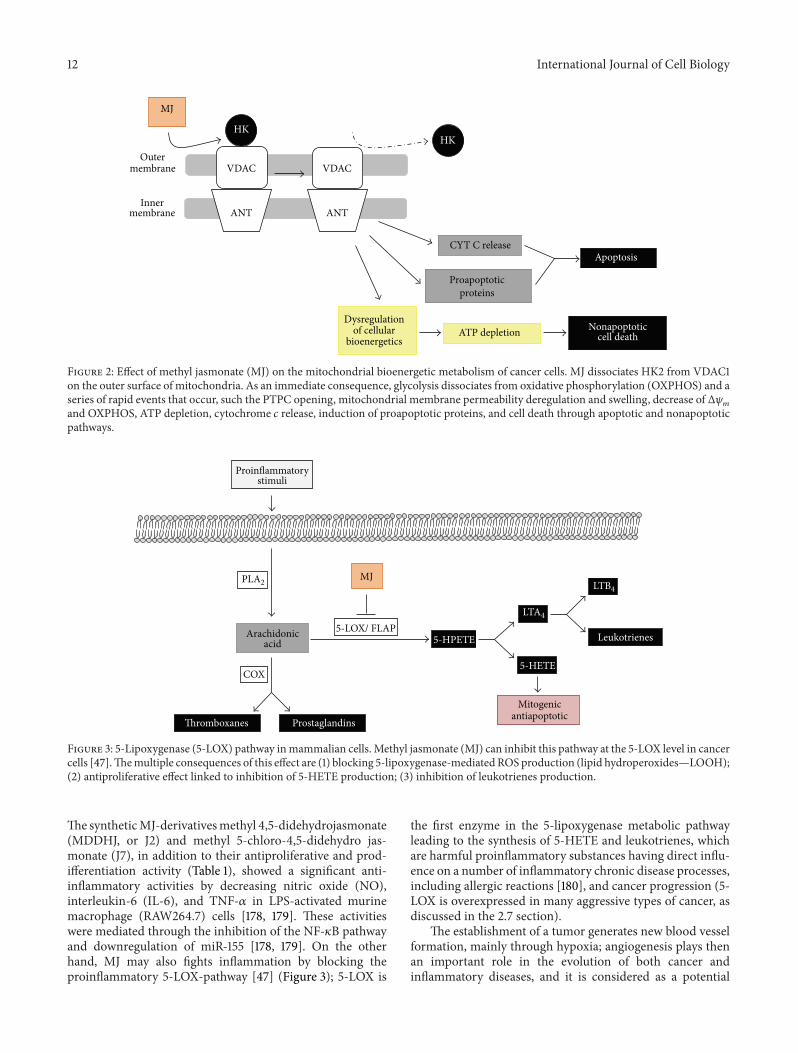
12 International Journal of Cell Biology
MJ
HKHK
VDAC
ANT
VDAC
ANT
Outermembrane
membraneInner
CYT C releaseApoptosis
Proapoptoticproteins
Dysregulationof cellular
bioenergeticsATP depletion Nonapoptotic
cell death
Figure 2 Effect of methyl jasmonate (MJ) on the mitochondrial bioenergetic metabolism of cancer cells MJ dissociates HK2 from VDAC1on the outer surface of mitochondria As an immediate consequence glycolysis dissociates from oxidative phosphorylation (OXPHOS) and aseries of rapid events that occur such the PTPC opening mitochondrial membrane permeability deregulation and swelling decrease of Δ120595
119898
and OXPHOS ATP depletion cytochrome 119888 release induction of proapoptotic proteins and cell death through apoptotic and nonapoptoticpathways
Proinflammatorystimuli
PLA2
Arachidonicacid
MJ
COX
5-LOX FLAP
ProstaglandinsThromboxanes
5-HPETE
5-HETE
Mitogenic
Leukotrienes
antiapoptotic
LTA4
LTB4
Figure 3 5-Lipoxygenase (5-LOX) pathway inmammalian cells Methyl jasmonate (MJ) can inhibit this pathway at the 5-LOX level in cancercells [47]Themultiple consequences of this effect are (1) blocking 5-lipoxygenase-mediated ROS production (lipid hydroperoxidesmdashLOOH)(2) antiproliferative effect linked to inhibition of 5-HETE production (3) inhibition of leukotrienes production
The syntheticMJ-derivativesmethyl 45-didehydrojasmonate(MDDHJ or J2) and methyl 5-chloro-45-didehydro jas-monate (J7) in addition to their antiproliferative and prod-ifferentiation activity (Table 1) showed a significant anti-inflammatory activities by decreasing nitric oxide (NO)interleukin-6 (IL-6) and TNF-120572 in LPS-activated murinemacrophage (RAW2647) cells [178 179] These activitieswere mediated through the inhibition of the NF-120581B pathwayand downregulation of miR-155 [178 179] On the otherhand MJ may also fights inflammation by blocking theproinflammatory 5-LOX-pathway [47] (Figure 3) 5-LOX is
the first enzyme in the 5-lipoxygenase metabolic pathwayleading to the synthesis of 5-HETE and leukotrienes whichare harmful proinflammatory substances having direct influ-ence on a number of inflammatory chronic disease processesincluding allergic reactions [180] and cancer progression (5-LOX is overexpressed in many aggressive types of cancer asdiscussed in the 27 section)
The establishment of a tumor generates new blood vesselformation mainly through hypoxia angiogenesis plays thenan important role in the evolution of both cancer andinflammatory diseases and it is considered as a potential
International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Cell Biology 13
target for cancer therapy [181] JAs inhibited angiogenesis athigh doses but the reverse was seen at lower doses in thechorioallantoic membrane (CAM) of chicken embryo [40]At present we have not found satisfactory explanations forthese results
Metastasis rather than the primary tumor is primarilyresponsible for anticancer treatment failure poor qualityof life and death in cancer patients Cancer metastasis ishighly complex and multistep process in nature involvingextracellular matrix degradation modified tumor cell adhe-sion active tumor cell migration altered tumor cell prolif-eration and altered cancer cell survival and angiogenesismodulation of these processes particularly the epithelial-to-mesenchymal transition (EMT) in which there is a profoundchange in transcriptional gene expression [57] enables thecells to escape the primary tumor microenvironment andspread locallydistally establishing a proliferative focus at asecondary site [182]MJ showed antimetastatic activity on themurine model of highly metastasic and drug-resistant B16-F10 melanoma cells [39] At low concentrations MJ was ableto reduced cell motility of melanoma cells and suppressedthe in vivo melanoma growth in the lung also the syntheticMJ derivative 57910-tetrabromo jasmonate prevented celladhesion and inhibited lung metastasis at a lower dose thanMJ (IC
50004mM vs IC
5026mM) (Tables 1 and 2) [24
39] In addition subcytotoxic concentrations of MJ (005ndash02mM) abolished migration invasion and angiogenesisof gastric cancer cells through downregulation of matrixmetalloproteinase 14 (MMP-14) [33] (Table 1)
27 Methyl Jasmonate Can Inhibit the Phase I Drug-Metabolizing Enzyme Aldo-Keto Reductase 1 (AKFR1) thatProvides Drug-Resistance in some Tumor Cells Aldo-ketoreductases (AKRs) are soluble cytoplasmic NAD(P)(H) oxi-doreductases that reduce aldehydes and ketones to yieldprimary and secondary alcohols respectively [183] Becausethis reaction permits subsequent conjugation reactions tooccur (eg sulfation and glucuronidation) AKRs can bereferred to as Phase I drug-metabolizing enzymes Thehuman AKRs can metabolize a vast range of substratesincluding drugs carcinogens and reactive aldehydes leadingto either their bioactivation or detoxification [183] AKRsare generally monomeric 34ndash37 kDa proteins present in allphyla the superfamily consists of 15 families which con-tains 151 members (httpwwwmedupenneduakr) Thir-teen human AKRs exist that use endogenous substrates(sugar and lipid aldehydes prostaglandins [184] retinalsand steroid hormones) and in many instances they regu-late nuclear receptor signaling [185] Exogenous substratesinclude metabolites implicated in chemical carcinogenesis[183] Human AKRs are highly polymorphic and there isinterindividual variation affecting susceptibility to nuclearreceptor signaling and chemical carcinogenesis [185] AKR1 member B1 (AKR1B1) and AKR 1 member B10 (AKR1B10)are overexpressed in human tumors such as liver breast andlung cancer and may play critical roles in the developmentand progression of cancer through carbonyl detoxificationretinoic acid homeostatic regulation and lipid metabolic
control as well as the activation of tobacco smoke car-cinogens [186] Members of the AKRF1 family are beingconsistently identified as potential biomarkers for varioustypes of cancer cells [187ndash189] Increased mRNA expressionofAKR1B1 AKR1C2 and carbonyl reductase (CR) and induc-tion of carbonyl-reducing enzymes AKR1B1 and AKR1C2can account for drug resistance to some anticancer agents[190ndash192] for instance the resistance of human stomachcarcinoma cells to daunorubicin [193] AKR1B1 was alsolinked to doxorubicin and cisplatin resistance in HeLa cervi-cal carcinoma cells because an AKR1B1 inhibitor enhancedthe cytotoxic effects of these anticancer agents [194] Allthis information clearly indicates that members of the AKRsuperfamily are emerging as important mediators of cancerpathology [132 192] and as new targets in cancer drugresistance therefore AKR1 inhibitors may represent a novelclass of antitumor agents
MJ can bind to members of the AKRF1 family [132] and acorrelation has been identified between cell sensitivity to MJand lower intracellular protein levels ofHK2 pVDAC23 andAKR1C1 [105]
28 Many Aggressive Cancer Cell Types Overexpress 5-Lipoxygenase (5-LOX) to Produce the Highly Proliferation-Stimulating Metabolite 5-HETE MJ Can Inhibit 5-LOXLipoxygenases (LOX linoleate oxygen oxidoreductase) are afamily of monomeric nonheme nonsulfur iron dioxygenasesthat catalyze the oxidation of polyunsaturated fatty acids(PUFAs) having a cis cis-14-pentadiene moiety into lipidhydroperoxides [195] LOX isozymes add a hydroperoxidegroup [minusOOH] at carbons 5 12 or 15 of AA and aredesignated 5- 12- or 15-lipoxygenases [196] The majorsubstrates for lipoxygenases in higher plants are linoleic acid(18 2) and linolenic acid (18 3) In animals LOXs catalysethe oxygenation of arachidonic (AA) (eicosatetraenoicC204 ETE) acid released from membrane phospholipidsby phospholipase A [197] and LOX-derived metabolitesparticipate as signals in several biochemical and physiologicalprocesses (httpenwikipediaorgwikiArachidonic acid)Arachidonic acid (AA) can be also converted by cyclo-oxygenase (COX) to prostaglandins (PGs) and othermetabolites LOX converts AA into 5-hydroperoxy-eicosatet-ranoate (5-HPETE) and then into 5-hydroxy-eicosatetranoicacid (5-HETE) and leukotrienes (B4) (Figure 3) TheAA-metabolites derived from COX or LOX reactionsmodulate many cellular processes and participate not onlyin various inflammatory reactions but also in proliferationand apoptosis [195] Our bodies respond to high levelsof dietary-generated AA (triggered by foods rich in 120596-6fatty acids and high-glycemic carbohydrates) by increasingthe expression of 5-LOX [198] 5-Lipoxygenase pathwaymetabolites 5-hydroxy-681114-eicosatetraenoic acid(5-HETE) and leukotriene LTB4 stimulates growth andincreases survival of cancer cells overexpressing 5-LOX[199ndash203] 5-HETE which enhances cell proliferation andactivates also antiapoptotic signaling (Figure 3) has beenimplicated in human cancer progression 5-HETE stronglystimulates the growth of hormone-responsive (LNCaP)
14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

14 International Journal of Cell Biology
and hormone-refractory (PC-3) human prostate cancer cells[204 205] 5-LOX is rate limiting for the synthesis of 5-HETEand subsequent LTs production thus inhibition of 5-LOXwould inhibit the generation of these proinflammatorymetabolites while promoting cell death [47 206ndash211]
Overexpression of 5-LOX has been shown in tissuesamples of primary tumor cells [212] in established cancercell lines and most human aggressive cancers such as breastMDA-MB-231 cells [208] PC-3 prostate cells [47 204 205207 213] bladder [15 214] esophagus [211 215] pancreas[216] gastric [217] and malignant pleural mesothelial cells[218] as well as lung [219] brain [15 220] colon [221] hepa-tocellular carcinoma (HCC) [222] neuroblastoma [223] col-orectal [224] glioma cells [225] andBarretrsquos adenocarcinoma[226] 5-LOX overexpression has been also found in oralcarcinoma [227] and canine osteosarcoma [228] all examplesstrongly suggesting the involvement of 5-LOX in cancerprogression Targeting 5-LOX with specific inhibitors orinhibiting its interaction with the 5-LOX-activating protein(FLAP) has resulted in decreased cell growth and increasedapoptosis in lung and breast cancer cell lines [201 229] Thepharmacological inhibition of 5-LOX by the specific syn-thetic inhibitorMK886 blocked 5-HETE production potentlysuppressed tumor cell growth induced cell cycle arrest andtriggered cell death via the intrinsic apoptotic pathway [204230 231] Inhibition of 5-LOX activity induced apoptosisin human prostate cancer cells [232] and human pancreaticcancer cells [230] and xenographs [200] Both the inhibitionof 5-LOX activity and the induction of apoptosis in prostatecancer cells were reversed by the addition of 5-HETE [204233 234]
LOX isozymes can be activated and upregulated by UVlight exposure bad fat diets oxidative stress ligand-receptoractivation (drugs Fas) ROS-inducing medications lipidhydroperoxides (LOOH) [198] and their secondary ROSderivatives (free radicals hydroxides ketones aldehydesetc) that can exert deleterious effects on membrane lipids(including mitochondrial membrane lipids) thus conform-ing the first critical step in oxidative stress leading to celldeath by the LOX-pathways Mitochondria that have beendamaged by lipid peroxidation decrease their membranepotential (Δ120595
119898) increasing cytochrome 119888 release and caspase
activation all these events ultimately leading to apoptosis[195 235] LOX-mediated apoptosis implies the following (1)ROS that (hydroperoxides) induced modifications of mem-brane properties (exposure of phosphatidylserine increasedlevels of cholesterol and consequently altered Ras expres-sion) (2) modifications of cytoskeleton and (3) modifi-cations in gene transcription (through nuclear factor NF-120581B poly(ADPribose) polymerase [195]) Apoptosis triggeredafter an oxidative stress caused by lipoxygenase activationis a common signal transduction pathway shared by animaland plant cells that has been conserved through evolution[235] The most interesting is the fact that MAPKERKp38MAPK and MAPKMEK stress pathways stimulate 5-LOX by phosphorylation [236ndash238] 5-LOX phosphorylationdoes not affect the enzyme catalytic activity but ratherregulates its interaction with other cellular components [239240] Thus for instance p38 MAPK can phosphorylate
5-LOX at Ser271 phosphorylation at this site is responsiblefor the stress-induced nuclear export of 5-LOX in CHO-K1cells (a cell line from Chinese hamster ovary) and HEK293(human embryonic kidney 293) cells [237] whereas in NIH3T3 (mouse embryonic fibroblastic) cells phosphorylationat Ser271 stimulates nuclear localization and subsequentlycellular 5-LOX activity [236]
5-LOX is an important target of JAs since these oxylipidsare synthesized in plants via 12-oxo-phytodienoic acid (12-oxo-PDA)mdashproduct of an oxidative cyclization of 120572-linolenicacid resulting in cyclopentanones [3 4] There are structuraland biogenic similarities of 12-oxo-PDA and jasmonates toanimal prostanglandins [3 15 38] It is not then a sur-prise to find that JAs can inhibit the lipoxygenase pathwayin animals MJ being itself a secondary cyclopentanoneproduct derived from of 120572-linolenic acid MJ significantlyinhibited proliferation and induced apoptosis and necrosisin extremely malignant human prostate cancer cells withinhours in dose- and kinetic-dependent manners showingspecific interaction with the 5-LOX pathway [47] (Table 1Figure 3) The mechanism of inhibition of 5-LOX activity byMJ is still unknown it is not known if JAs are able to directlyinteract with the catalytic site of 5-LOX or cause indirectinhibitory effects through interaction with an allosteric siteor its regulatory 5-lipoxygenase-activating (FLAP) proteinRelevant to this point is the fact that 5-LOX inhibition maycause cytotoxic and antiproliferative effects in cancer cellsindependently of suppression of the 5-LOX catalytic activity[239]
3 Therapeutic Advantages ofCombining Jasmonates with ConventionalAnticancer Drug Treatments
Conventional anticancer treatments (surgery radiationchemotherapy and targeted therapies) are all strong inducersof ROS and as discussed before ROS excess is a majormechanism for inducing cell death through apoptosis [58241] Excess ROS is highly proinflammatory to cancer neigh-boring normal cells tissues being mostly responsible forthe painful and aggressive side effects observed in can-cer patients [242] Many phytochemicals have been shownto have anti-inflammatory and also cancer-preventing andanticancer properties (linked to metabolism proliferationinvasion angiogenesis and metastasis activities) showing inaddition relatively low toxicity to normal cells [243] Thusa rational for a good clinical strategy is combining harshanticancer treatments with natural anti-inflammatory andcancer-preventing compounds to increase treatment efficacywhile lowering drug dose and toxicity [244ndash246] Howevercombination of components having opposite effects such asfor instance antioxidants (anti-ROS) may impair treatmentefficacy [57] Additive and synergic effects may actually bequite advantageous reducing the side effects often seen withsingle high drug doses treatments in addition to preventingresistant tumor cells to develop [245ndash247] Considerableamount of research has been done to identify the relevantmolecules that contribute to drug efflux and drug resistance
International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Cell Biology 15
DysregulationATP depletionof cellular
bioenergetics
Apoptosis
Apoptosis
Invasionmetastasis
Apoptosis Redifferentiation
MJ
MAPK
5-LOXFLAP
proteinsAntiapoptotic
proteinsProapoptotic
Nonapoptoticcell death
Nonapoptoticcell death
TNFR1
Cell cycle arrest at AKRF1
HKVDACdisruption
NF-120581B
uarr ROS
G0G1 G1S S and G2M
Figure 4 Methyl jasmonate mediated activities in cancer cells MJ and other jasmonates (JAs) (1) arrest cell cycle inhibiting cell growthand proliferation (2) cause cell death by triggering the intrinsic and extrinsic proapoptotic pathways and induce also p53-independentapoptosis and nonapoptotic cell death or necrosis (3) detach hexokinase (HK) from the voltage-dependent anion channel isoform 1 (VDAC1)dissociating glycolytic andmitochondrial functions decreasing themitochondrial membrane potential (Δ120595
119898) favoring cytochrome 119888 release
causing ATP depletion and activating proapoptotic and inactivating antiapoptotic proteins (4) induce reactive oxygen species (ROS)triggering andor mediating different cellular responses (5) stimulate MAPK-stress signaling pathways and induce redifferentiation inleukemia cells (6) inhibit overexpressed cancer cell enzymes such as aldo-keto reductase 1 (AKR1) and 5-lipoxygenase (5-LOX)
but the interdependence and cross-talk between metabolicpathways grant cancer cells a high metabolic plasticityfavoring their adaptation to new environmental and stressfulconditions [245 248] For these reasons phytochemicalsby being pleiotropic and multitarget have the great abilityto concomitantly modulate multiple metabolic and survivalpathways becoming selectively cytotoxic to cancer cellswithout inducing toxicity in normal cells [243] through theseproperties phytochemicals can regulate the death receptorpathways and overcome multidrug resistance (MDR) pro-teins as documented by multiple examples in laboratory andclinical practice [248]
By the way of its multiple and different mechanisms ofaction (Figure 4) MJ may display strong cooperative efficacywith other anticancer agents to induce death in severalcancer cells helping also to overcome multidrug resistance[19 23 170] Combination of MJ with any anticancer drugmay have a different rational synergism being the firstand the more important one to take into consideration
If synergism is present combined MJ-drug formulationsmay allow for less toxic drug concentrations while keepingan efficient treatment Chemical structure physicochemicalproperties and molecular mechanisms of action of selecteddrugs are also important parameters to be considered forcombination MJ is a small molecular weight fatty acid-derived cyclopentanone which is very hydrophobic and isnot charged it would act then better on membranes andamphipathic vehicles should be considered if combined withhydrophilic drugs JAs MJ and their derivatives have beencombined with ionizing radiation conventional chemother-apy drugs and also with other phytochemicals to enhancecancer cell cytotoxicity in anticancer treatments allowingfor lower effective doses while showing enhanced efficacy ininducing death of carcinoma cells (Table 3) [23 28 170] MJis very selective for cancer cells [17] it is not toxic for normalcells [15 50] and it induces cell death primarily by triggeringmitochondrial perturbation (detachment of HK from theouter mitochondria membrane [30] prooxidant activity
16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

16 International Journal of Cell Biology
etc) as deduced from previous discussion Most aggressivecancers show a glycolytic metabolic profile and combinationof glycolysis inhibitors with OxPhos inhibitors enhances theanticancer properties of this combinationThus combinationof MJ with 2-deoxy-D-glucose (glycolysis inhibitor) andwith four conventional chemotherapeutic drugs resulted insuper-additive cytotoxic effects on several types of cancercells (Table 3) MJ is clearly a mitochondriotoxic anticancercompound and compounds that directly targetmitochondriaoffer the advantage to inducemitochondrial outer membranepermeabilization independently of upstream signal transduc-tion elements that are frequently impaired in human cancersin this way mitochondrion-targeted agents may bypass someforms of drug resistance [96] and cause nonapoptotic celldeath
From Tables 1 and 2 we can infer that MJ acts on a widegamma of cancer types in vitro and in vivo and this propertyis probably due to its capacity to target multiple metabolicpathways in different cancer cell types leading to apoptoticand nonapoptotic cell death as already discussed aboveThe reported cooperative effects of MJ with other drugsare shown in Table 3 (1) BCNU (carmustine) an alkylatingnitrosourea agent with no cytotoxic effect on pancreaticcarcinoma cell line MIA PaCa-2 in vitro combined with MJshowed enhanced cell death [170] The underlying molecularmechanisms in this combination are probably related to theDNA damaging action of BCNU on mitochondria [249]rendering the above cells sensitive to MJ which also alter themitochondrial function [32] thus showing a highly additivecytotoxic effect of two combined mitochondriotoxic agentsThis combination acted also synergistically on murine B-cellleukemia (BCL1) cells [170] (2) MJ (a mitochondriotoxiccompound) combined with 2-deoxy-D-glucose (2DG) (aninhibitor of glycolysis) showed synergicadditive effects andcell death on various carcinoma cell types (Table 3) [170]the synergic effect was probably due to OxPhosATP biosyn-thesis inhibition by MJ and to glycolyticATP biosynthesisinhibition by 2DG [17] MJ enhanced also the toxic effects oftaxol [170] much by the same reasons discussed in (1) taxolinduces mitochondrial membrane depolarization resultingin translocation of apoptosis-inducing factor (AIF) but notcytochrome 119888 from the mitochondria to the cytosol (3)MJ enhanced significantly the in vivo antileukemic toxiceffect of the antibiotic adriamycin (doxorubicin) in a chroniclymphocyte leukemia (CLL) mouse model [170] adriamycinincreases the intracellular levels of ROS followed by mito-chondrial membrane depolarization cytochrome 119888 releaseand caspase 3 activation the IC
50value for adriamycin
dropped by half when combined with MJ [170 250] (4) MJtreatment increased the levels of pAkt in sarcoma cell linesMCA-105 and SaOS-2 however the treatment of both celllines with a combination of MJ and small molecular weightPI3KAkt inhibitors resulted in a synergistic cytotoxic effect[49] (5) Glucose attenuated the MJ-induced cytotoxicity inthese cells but the treatment with a combination of MJplus 2DG resulted in a synergistic cytotoxic effect [49] (6)MJ suppressed the 120574-radiation-induced expression of Bcl-2 in human prostate cancer cells enhancing the sensitivityof these cells to 120574-radiation [104] (7) MJ combined with
the monoterpene perillyl alcohol (POH) enhanced the cyto-toxicity of cisplatin in breastMDA-MB-231 cells and inMDA-MB-435 cancer cells [74 251] MJ and POH activate TNFR1(extrinsic apoptosis pathway) which is further increasedby the addition of cisplatin [251] MJ and POH decreasemitochondrialmembrane potential (intrinsic apoptosis path-ways) inducing cytochrome 119888 release and this effect isfurther enhanced by cisplatin whose cytotoxic activity seemsto be largely dependent on the glycolytic metabolism oftumor cells (cisplatin would redirect cancer cells to oxidativephosphorylation from the ldquoWarburg effectrdquo) [252] in thisexample the three drugs treatment was more effective thansingle drug or two drugs together [251] (8) Survivin (aninhibitor of apoptosis protein (IAP) member) negativelyregulates TRAIL-mediated apoptosis in colorectal cancer(CRC) cells butMJ by downregulating theWnttranscriptionfactor pathway prevented the overexpression of survivin thussensitizing cancer cells for TRAIL-mediated cytotoxicity [32]TRAIL is also able to induce apoptosis in prostate cancercells however overexpression of antiapoptotic proteins andinhibition of proapoptotic proteins results in lowering theTRAIL-mediated apoptosis [253] (9) MJ combined with theSmac peptide (inhibitor of IAPs) showed IAPs-inhibitingsynergy in human bladder EJ and T24 cells and humanembryonic kidney HEK 293 cells [254] (10) Hormone-refractory prostate cancer cells are relatively resistant to MJbecause the overexpression of IAP proteins decreases theproapoptotic activity ofMJ In these cases the IAP antagonistSmacN7 (a synthetic peptide comprising the first N-terminalseven residues of Smac) was successfully used to synergicallypotentiate MJ-mediated apoptosis [255] The SmacN7MJcombination induced sensitization to apoptosis through bothcaspase-9-dependent and caspase-9-independent pathways[255] (11) MJ worked synergically with cisplatin as well aswith X-rays irradiation on various cervical cancer cell linesenhancing the cytotoxic effects of these conventional thera-pies lowering the effective doses required to inhibit survivalof those cells [29] MJ worked also well with 120572-irradiationenhancing the selective reduction of cell viability and survivalof cervical cancer cells effected by 120572-rays [29] (12) MJcombined with 5-FU in human adenocarcinoma colon HT-39 cells drastically decreased the 5-FU IC
50values which
is important to reduce the unwanted side effects usuallyassociated with this drug [31] (13) MJ synergized also withbortezomib (a proteasome inhibitor) in vitro and prolongedsurvival of immunocompromised mice harboring diffuselesions of multiple myeloma (MM1S) cells as compared tovehicle-treated mice [105]
4 Concluding Remarks and Hypothesis
As inferred from the many in vitro and in vivo evidencesdiscussed here JAs are highly selective towards malignantcells (excepting hormone-independent prostate cancer cellsand some sarcoma cell lines which are relatively moreresistant to MJ than other cancer cells) not affecting normalcells Of particular interest is to know thatMJ is well toleratedand no meaningful local or systemic side effects have been
International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Cell Biology 17
detected in animal and humans trials after toxicology anddermatology studies for acute toxicity skin irritationmucousmembrane (eye) irritation skin sensitization phototoxicityand photoallergy not being toxic to healthy organisms ofdifferent species so far tested and emphasizing the safetyof MJ use in humans [50ndash52] JAs and particularly MJhave been shown to stimulate andor to inhibit multipleintracellular pathways and proteins (Figure 4) that may bedifferently expressed among different cancer cell types MJanticancer activities can be summarized as follows (1) MJis able to induce reactive oxygen species (ROS) that aresubsequent mediators of different intracellular responses (2)ROS may stimulate the MAPK-stress signaling pathwaysinducing ERK-dependent redifferentiation or apoptosis inleukemia cells (3) MJ induces cell cycle arrest at differentphases in different types of cancer cells interfering by thiswaycell growth proliferation and migration (4) MJ promotedthe detachment of hexokinase (HK) from to the voltage-dependent-anion-channel isoform 1 (VDAC1) in the outermitochondrial membrane dissociating glycolytic and mito-chondrial metabolic functions this event decreases the mito-chondrial membrane potential (Δ120595
119898) favoring cytochrome 119888
release causing ATP depletion activating proapoptotic andinactivating antiapoptotic proteins (5) MJ induces cell deathby triggering the intrinsicextrinsic proapoptotic and alsononapoptotic pathways (6) MJ inhibits metabolic enzymessuch as the aldo-keto reductase 1 (AKR1) and 5-lipoxygenase(5-LOX) that favor cancer cells survival and proliferationand that have been found overexpressed in aggressive cancercells (7) Jasmonate-mediated anti-inflammatory activitiesinvolving downregulation of NF-120581B transcription factoractivity [178 179] and inhibition of the 5-LOX pathway(8) MJ and some of synthetic derivatives inhibit cancercell invasion and metastasis through downregulation of theNF-120581B transcription factor and miR-155 [178 179] Thesepathways may all or partially be expressed in a given cancercell type at agiven time and MJ (JAs) can target all of themcausing a catastrophic cancer cellular failure and subsequentcell death (except when inducing redifferentiation) whilesparing normal cells As stated by Elia and Flescher [23] allMJ anticancer mechanisms are not mutually exclusive andmay occur concomitantly so affecting different cancer cellsor can be active in different time frames and concentrationranges [23] Thus MJ can probably act simultaneously on allthe above targets within the same cell population and cellsmay then die by apoptotic andor nonapoptotic mechanismsas it was observed by Kniazhanski et al [26] and Milrot et al[28] on cervical cancer cells after treatment withMJ (Table 1)
Integrating all the above knowledgewemight hypothesizethe jasmonates (mainly MJ) mechanisms of action leadingto death mammalian cancer cells By its highly hydrophobicciclopentanone lipid structure MJ may easily get across cel-lular membranes during this transit the prooxidant natureof the MJ oxylipin causes an oxidative event that perturbsthe lipid bilayer as reflected by a decrease in membranefluidity upon contact [25] this event stimulates NAD(P)Hoxidases (NOX) and generates reactive oxygen species (ROS)(mainly H
2O2) [24] both events linked to functions in the
MAPK stress pathways [25] ROS may subsequently act as
mediators of other cellular responses as well The interactionof MJ with the surface plasma membrane might also beresponsible for the increased expression of TNFR1 expressionand caspase-8 activation observed by Yeruva et al [25] insome cancer cell types activating this way the extrinsicpathway of apoptosis Intracellularly MJ may bind to lipidsand hydrophobic proteins and also interact with subcellu-lar organelles (mitochondria nucleus and peroxisomes) Apossible mechanism(s) of surface structures discriminationbetween normal and cancer cells mitochondria by MJ seemsto occur and it is probably related to the overexpressionand tight binding of hexokinase(s) (HK) to the VDAC1 porein the outer mitochondrial membranes (OMM) in cancercells [24 30] By binding to a hydrophobic HK region inthe HKVDAC1 complex on the OMM MJ induces HKdetachment from its VDAC anchor (Figure 2) When thishappen the BaxBcl-2 balance is altered antiapoptotic Bcl-2 proteins are released from the OMM and proapoptoticproteins occupy their sites inducing the intrinsic way of apop-tosis concomitantly the mitochondrial membrane potential(Δ120595119898) decreases cytochrome 119888 is released caspase-3 is
activated mitochondrial ATP depleted ROS generated andcell cycle arrested [23 25 30] causing a catastrophic energybreakdown and cell death Nonapoptotic cell death could bethe consequence of the severe bioenergetic damages inducedby the direct interaction of MJ with mitochondria leading toincreased ROS levels inhibition of ATP synthesis and ATPdepletion [20 30] However depending on cancer cell typesensitivity to MJ the extent of mitochondrial damage couldbe determinant for apoptotic andor nonapoptotic cell deathIn the nucleusMJmay interact with cell cycle regulatory pro-teins (cyclins CDKs and survivin) inducing cell cycle arrestat different phases in different cancer cell types (Table 1) thusinhibiting cell cancer growth proliferation and invasionMJ-induced downregulation of survivin severely affects cellcycle progression and induces apoptosis [32] MJ inducesTRAIL cytotoxicity but overexpression of survivin preventsthis activity [32] When targeting several cellular processesMJ induces ROS production ROS influences cell cycleprogression via phosphorylation and ubiquitination of CDKsand other cell cycle regulatory molecules [57]
The inhibition of multiple cancer cell activities by Jas-monates (MJ) becomes important when we think at cancer asa multifactorial disease that should be better treated throughmultitarget strategies Due to their multitarget and pleotropicproperties JAs and particularly MJ are capable of rapidlykilling many cancer cell types independently of factors suchas cellular mRNA transcription protein translation [18] andp53 expression [17] leaving untouched normal cells [15] Dueto these properties MJ may act also as a chemosensitizer tochemotherapics and help to overcome drug-resistance [1719] as seen inTable 3MJ significantly reduced the IC
50values
of chemotherapeutic agents with which it was combined Asit has been commented in this and other reviews JAs can beused alone or in synergic combination with other anticancerdrugs and might be seriously considered to be includedsafely in some of the current anticancer therapies Finally therapidity by which MJ causes damage to cancer cells turns itinto a promising anticancer agent of high therapeutic value
18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

18 International Journal of Cell Biology
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The research in our laboratory is financed by CNPq FAF andFAPERJ
References
[1] A Ertel A Verghese S W Byers M Ochs and A TozerenldquoPathway-specific differences between tumor cell lines andnormal and tumor tissue cellsrdquoMolecular Cancer vol 5 article55 2006
[2] X Mou S Kesari P Y Wen and X Huang ldquoCrude drugsas anticancer agentsrdquo International Journal of Clinical andExperimental Medicine vol 4 no 1 pp 17ndash25 2011
[3] R Liechti and E E Farmer ldquoThe Jasmonate pathwayrdquo Sciencevol 296 no 5573 pp 1649ndash1650 2002
[4] H Weber ldquoFatty acid-derived signals in plantsrdquo Trends in PlantScience vol 7 no 5 pp 217ndash224 2002
[5] W W Christie ldquoPlant oxylipins chemistry and biologyrdquo 2010httplipidlibraryaocsorglipidseicplantindexhtm
[6] G Sembdner and B Parthier ldquoThe biochemistry and thephysiological and molecular actions of jasmonatesrdquo AnnualReview of Plant Physiology and Plant Molecular Biology vol 44no 1 pp 569ndash589 1993
[7] N I Vasyukova S V Zinovieva Z V Udalova N G Gerasi-mova O L Ozeretskovskaya and M D Sonin ldquoJasmonic acidand tomato resistance to the root-knot nematode Meloidogyneincognitardquo Doklady Biological Sciences vol 428 pp 448ndash4502009
[8] E E Farmer and C A Ryan ldquoInterplant communicationairborne methyl jasmonate induces synthesis of proteinaseinhibitors in plant leavesrdquo Proceedings of the National Academyof Sciences of the United States of America vol 87 no 19 pp7713ndash7716 1990
[9] M De Vos V R Van Oosten R M P Van Poecke et alldquoSignal signature and transcriptome changes of Arabidopsisduring pathogen and insect attackrdquo Molecular Plant-MicrobeInteractions vol 18 no 9 pp 923ndash937 2005
[10] M M Izaguirre C A Mazza A Svatos I T Baldwin and CL Ballare ldquoSolar ultraviolet-B radiation and insect herbivorytrigger partially overlapping phenolic responses in Nicotianaattenuata and Nicotiana longiflorardquo Annals of Botany vol 99no 1 pp 103ndash109 2007
[11] R Karban ldquoPlant behaviour and communicationrdquo EcologyLetters vol 11 no 7 pp 727ndash739 2008
[12] A M D S Soares T F de Souza T Jacinto and O L TMachado ldquoEffect of methyl jasmonate on antioxidative enzymeactivities and on the contents of ROS and H
2O2in Ricinus
communis leavesrdquo Brazilian Journal of Plant Physiology vol 22no 3 pp 151ndash158 2010
[13] S A-H-Mackerness S L Surplus P Blake et al ldquoUltraviolet-B-induced stress and changes in gene expression in Arabidopsisthaliana role of signalling pathways controlled by jasmonicacid ethylene and reactive oxygen speciesrdquo Plant Cell ampEnvironment vol 22 no 11 pp 1413ndash1423 1999
[14] L Zhang and D Xing ldquoMethyl jasmonate induces productionof reactive oxygen species and alterations in mitochondrialdynamics that precede photosynthetic dysfunction and subse-quent cell deathrdquo Plant and Cell Physiology vol 49 no 7 pp1092ndash1111 2008
[15] O Fingrut and E Flescher ldquoPlant stress hormones suppressthe proliferation and induce apoptosis in human cancer cellsrdquoLeukemia vol 16 no 4 pp 608ndash616 2002
[16] E Flescher ldquoJasmonatesmdasha new family of anti-cancer agentsrdquoAnti-Cancer Drugs vol 16 no 9 pp 911ndash916 2005
[17] O Fingrut D Reischer R Rotem et al ldquoJasmonates inducenonapoptotic death in high-resistance mutant p53-expressingB-lymphoma cellsrdquoBritish Journal of Pharmacology vol 146 no6 pp 800ndash808 2005
[18] R Rotem O Fingrut J Moskovitz and E Flescher ldquoThe anti-cancer plant stress-proteinmethyl jasmonate induces activationof stress-regulated c-Jun N-terminal kinase and p38 proteinkinase in human lymphoid cellsrdquo Leukemia vol 17 no 11 pp2230ndash2234 2003
[19] E Flescher ldquoJasmonates in cancer therapyrdquo Cancer Letters vol245 no 1-2 pp 1ndash10 2007
[20] N Goldin A Heyfets D Reischer and E FlescherldquoMitochondria-mediated ATP depletion by anti-canceragents of the jasmonate familyrdquo Journal of Bioenergetics andBiomembranes vol 39 no 1 pp 51ndash57 2007
[21] S Cohen and E Flescher ldquoMethyl jasmonate a plant stresshormone as an anti-cancer drugrdquo Phytochemistry vol 70 no13-14 pp 1600ndash1609 2009
[22] D Reischer-Pelech and E Flescher ldquoJasmonates plant stresshormones as anticancer agentsrdquo in Emerging Trends in DietaryComponents for Preventing andCombatingDisease pp 303ndash3222012
[23] U Elia and E Flescher ldquoCombined chemotherapy or biotherapywith jasmonates targeting energy metabolism for cancer treat-mentrdquo Current Pharmaceutical Biotechnology vol 14 no 3 pp331ndash341 2013
[24] Z Raviv S Cohen and D Reischer-Pelech ldquoThe anti-canceractivities of jasmonatesrdquo Cancer Chemotherapy and Pharmacol-ogy vol 71 no 2 pp 275ndash285 2013
[25] L Yeruva J A Elegbede and S W Carper ldquoMethyl jasmonatedecreases membrane fluidity and induces apoptosis throughtumor necrosis factor receptor 1 in breast cancer cellsrdquo Anti-Cancer Drugs vol 19 no 8 pp 766ndash776 2008
[26] T Kniazhanski A Jackman A Heyfets P Gonen E Flescherand L Sherman ldquoMethyl jasmonate induces cell death withmixed characteristics of apoptosis and necrosis in cervicalcancer cellsrdquo Cancer Letters vol 271 no 1 pp 34ndash46 2008
[27] J Zhao S Kang X Zhang et al ldquoApoptotic activity of a newjasmonate analogue is associated with its induction of DNAdamagerdquo Oncology Reports vol 24 no 3 pp 771ndash777 2010
[28] E Milrot A Jackman T Kniazhanski P Gonen E Flescherand L Sherman ldquoMethyl jasmonate reduces the survival ofcervical cancer cells and downregulates HPV E6 and E7 andsurvivinrdquo Cancer Letters vol 319 no 1 pp 31ndash38 2012
[29] E Milrot A Jackman E Flescher et al ldquoEnhanced killing ofcervical cancer cells by combinations of methyl jasmonate withcisplatin X or alpha radiationrdquo Investigational New Drugs vol31 no 2 pp 333ndash344 2013
[30] N Goldin L Arzoine A Heyfets et al ldquoMethyl jasmonatebinds to and detaches mitochondria-bound hexokinaserdquoOnco-gene vol 27 no 34 pp 4636ndash4643 2008
International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Cell Biology 19
[31] J Avila-Roman E Talero C de los Reyes Jimenez et alldquoMethyl jasmonate is a promising tool against colon cancerrdquo inProceedings of the 6th European Congress of Pharmacology 2012
[32] Z Raviv A Zilberberg S Cohen et al ldquoMethyl jasmonatedown-regulates survivin expression and sensitizes colon carci-noma cells towards TRAIL-induced cytotoxicityrdquo British Jour-nal of Pharmacology vol 164 no 5 pp 1433ndash1444 2011
[33] L Zheng D Li X Xiang et al ldquoMethyl jasmonate abolishesthe migration invasion and angiogenesis of gastric cancer cellsthrough down-regulation ofmatrixmetalloproteinase 14rdquo BMCCancer vol 13 article 74 2013
[34] C Park C-Y Jin G-Y Kim et al ldquoA methyl jasmonatederivative J-7 induces apoptosis in human hepatocarcinomaHep3B cells in vitrordquo Toxicology in Vitro vol 24 no 7 pp 1920ndash1926 2010
[35] C Park C-Y Jin H J Hwang et al ldquoJ7 a methyl jasmonatederivative enhances TRAIL-mediated apoptosis through up-regulation of reactive oxygen species generation in humanhepatoma HepG2 cellsrdquo Toxicology in Vitro vol 26 no 1 pp86ndash93 2012
[36] J H Kim S Y Lee S Y Oh et al ldquoMethyl jasmonate inducesapoptosis through induction of BaxBcl-XS and activation ofcaspase-3 via ROS production in A549 cellsrdquoOncology Reportsvol 12 no 6 pp 1233ndash1238 2004
[37] L Yeruva K J Pierre S W Carper J A Elegbede B J Toyand R C Wang ldquoJasmonates induce apoptosis and cell cyclearrest in non-small cell lung cancer linesrdquo Experimental LungResearch vol 32 no 10 pp 499ndash516 2006
[38] R Rotem A Heyfets O Fingrut D Blickstein M Shaklai andE Flescher ldquoJasmonates novel anticancer agents acting directlyand selectively on human cancer cell mitochondriardquo CancerResearch vol 65 no 5 pp 1984ndash1993 2005
[39] D Reischer AHeyfets S Shimony J Nordenberg Y Kashmanand E Flescher ldquoEffects of natural and novel synthetic jas-monates in experimental metastatic melanomardquo British Journalof Pharmacology vol 150 no 6 pp 738ndash749 2007
[40] J E F Pereira Lopes M R Barbosa C N Stella et al ldquoInvivo anti-angiogenic effects further support the promise of theantineoplasic activity of methyl jasmonaterdquo Brazilian Journal ofBiology vol 70 no 2 pp 443ndash449 2010
[41] Y Ishii H Kiyota S Sakai and Y Honma ldquoInduction ofdifferentiation of human myeloid leukemia cells by jasmonatesplant hormonesrdquo Leukemia vol 18 no 8 pp 1413ndash1419 2004
[42] H Tsumura M Akimoto H Kiyota Y Ishii H Ishikuraand Y Honma ldquoGene expression profiles in differentiatingleukemia cells induced by methyl jasmonate are similar tothose of cytokinins and methyl jasmonate analogs induce thedifferentiation of human leukemia cells in primary culturerdquoLeukemia vol 23 no 4 pp 753ndash760 2009
[43] G-S Jiang Q-S Tong F-Q Zeng et al ldquoMethyl jasmonateinduces apoptosis of human neuroblastoma cell line BE(2)-Cand its mechanismrdquo Yaoxue Xuebao vol 43 no 6 pp 584ndash5902008
[44] Q-S Tong G-S Jiang L-D Zheng et al ldquoNatural jas-monates of different structures suppress the growth of humanneuroblastoma cell line SH-SY5Y and its mechanismsrdquo ActaPharmacologica Sinica vol 29 no 7 pp 861ndash869 2008
[45] Q-S Tong G-S Jiang L-D Zheng et al ldquoMethyl jasmonatedownregulates expression of proliferating cell nuclear antigenand induces apoptosis in humanneuroblastoma cell linesrdquoAnti-Cancer Drugs vol 19 no 6 pp 573ndash581 2008
[46] D Samaila D E Ezekwudo K K Yimam and J A ElegbedeldquoBioactive plant compounds inhibited the proliferation andinduced apoptosis in human cancer cell lines in vitrordquo Trans-actions of the Integrated Bio-Medical Informatics amp EnablingTechnologies Symposiums vol 1 pp 34ndash42 2004
[47] D E Ezekwudo R C Wang and J A Elegbede ldquoMethyljasmonate induced apoptosis in human prostate carcinoma cellsvia 5-lipoxygenase dependent pathwayrdquo Journal of ExperimentalTherapeutics and Oncology vol 6 no 4 pp 267ndash277 2007
[48] L Yeruva K J Pierre M Bathina A Elegbede and S WCarper ldquoDelayed cytotoxic effects of methyl jasmonate and cis-jasmone induced apoptosis in prostate cancer cellsrdquo CancerInvestigation vol 26 no 9 pp 890ndash899 2008
[49] U Elia andE Flescher ldquoPI3KAkt pathway activation attenuatesthe cytotoxic effect of methyl jasmonate toward sarcoma cellsrdquoNeoplasia vol 10 no 11 pp 1303ndash1313 2008
[50] J Scognamiglio L Jones C S Letizia and A M ApildquoFragrance material review on methyl jasmonaterdquo Food andChemical Toxicology vol 50 supplement 3 pp S572ndashS576 2012
[51] B Palmieri T Iannitti S Capone and E Flescher ldquoA prelimi-nary study of the local treatment of preneoplastic andmalignantskin lesions using methyl jasmonaterdquo European Review forMedical and Pharmacological Sciences vol 15 no 3 pp 333ndash336 2011
[52] Environmental Protection Agency (EPA) ldquoMethyl Jasmonateexemption from the requirement of a tolerancerdquo Federal Regis-ter vol 78 no 74 pp 22789ndash22794 2013
[53] J A Pietenpol and Z A Stewart ldquoCell cycle checkpointsignaling cell cycle arrest versus apoptosisrdquo Toxicology vol 181-182 pp 475ndash481 2002
[54] S van den Heuvel ldquoCell-cycle regulationrdquo in WormBook TheC elegans Research Community WormBook 2005
[55] K Vermeulen Z N Berneman and D R Van Bockstaele ldquoCellcycle and apoptosisrdquoCell Proliferation vol 36 no 3 pp 165ndash1752003
[56] D Reisman P Takahashi A Polson and K Boggs ldquoTran-scriptional regulation of the p53 tumor suppressor gene inS-phase of the cell-cycle and the cellular response to DNAdamagerdquo Biochemistry Research International vol 2012 ArticleID 808934 5 pages 2012
[57] E H Verbon J A Post and J Boonstra ldquoThe influence ofreactive oxygen species on cell cycle progression inmammaliancellsrdquo Gene vol 511 no 1 pp 1ndash6 2012
[58] J Watson ldquoOxidants antioxidants and the current incurabilityof metastatic cancersrdquo Open Biology vol 3 no 1 Article ID120144 2013
[59] L Soucek J Whitfield C P Martins et al ldquoModelling Mycinhibition as a cancer therapyrdquo Nature vol 455 no 7213 pp679ndash683 2008
[60] A K Jain K Allton M Iacovino et al ldquoP53 regulates cell cycleand micrornas to promote differentiation of human embryonicstem cellsrdquoPLoS Biology vol 10 no 2 Article ID e1001268 2012
[61] M R Chorawala P M Oza and G B Shah ldquoMechanisms ofanticancer drugs resistance an overviewrdquo International Journalof Pharmaceutical Sciences and Drug Research vol 4 no 1 pp1ndash9 2012
[62] Y-A Suh S M Post A C Elizondo-Fraire et al ldquoMultiplestress signals activate mutant p53 in vivordquo Cancer Research vol71 no 23 pp 7168ndash7175 2011
[63] K Kuribayashi N Finnberg J R Jeffers G P Zambetti andW S El-Deiry ldquoThe relative contribution of pro-apoptotic
20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

20 International Journal of Cell Biology
p53-target genes in the triggering of apoptosis following DNAdamage in vitro and in vivordquo Cell Cycle vol 10 no 14 pp 2380ndash2389 2011
[64] A M Puzio-Kuter ldquoThe role of p53 in metabolic regulationrdquoGenes and Cancer vol 2 no 4 pp 385ndash391 2011
[65] K Suzuki and H Matsubara ldquoRecent advances in p53 researchand cancer treatmentrdquo Journal of Biomedicine and Biotechnol-ogy vol 2011 Article ID 978312 7 pages 2011
[66] M J Grossel and PWHinds ldquoFrom cell cycle to differentiationan expanding role for Cdk6rdquo Cell Cycle vol 5 no 3 pp 266ndash270 2006
[67] R G Jones and C B Thompson ldquoTumor suppressors and cellmetabolism a recipe for cancer growthrdquo Genes and Develop-ment vol 23 no 5 pp 537ndash548 2009
[68] M Olivier M Hollstein and P Hainaut ldquoTP53 mutations inhuman cancers origins consequences and clinical userdquo ColdSpring Harbor Perspectives in Biology vol 2 no 1 Article IDa001008 2010
[69] A Maillet and S Pervaiz ldquoRedox regulation of p53 redoxeffectors regulated by p53 a subtle balancerdquo Antioxidants andRedox Signaling vol 16 no 11 pp 1285ndash1294 2012
[70] A A Sablina A V Budanov G V Ilyinskaya L S Agapova JE Kravchenko and PM Chumakov ldquoThe antioxidant functionof the p53 tumor suppressorrdquoNatureMedicine vol 11 no 12 pp1306ndash1313 2005
[71] E Gottlieb and K H Vousden ldquoP53 regulation of metabolicpathwaysrdquoCold Spring Harbor Perspectives in Biology vol 2 no4 Article ID a001040 2010
[72] P-F Li R Dietz and R vonHarsdorf ldquoP53 regulatesmitochon-drial membrane potential through reactive oxygen species andinduces cytochrome c-independent apoptosis blocked by Bcl-2rdquo EMBO Journal vol 18 no 21 pp 6027ndash6036 1999
[73] B Liu P53 and reactive oxygen species a convoluted story[MS thesis] University of Kentucky 2007 httpuknowl-edgeukyedugradschool theses450
[74] J M Rae C J Creighton J M Meck B R Haddad and M DJohnson ldquoMDA-MB-435 cells are derived fromM14Melanomacellsmdasha loss for breast cancer but a boon for melanomaresearchrdquo Breast Cancer Research and Treatment vol 104 no1 pp 13ndash19 2007
[75] H Berglind Y Pawitan S Kato C Ishioka and T SoussildquoAnalysis of p53 mutation status in human cancer cell lines aparadigm for cell line cross-contaminationrdquo Cancer Biology ampTherapy vol 7 no 5 pp 699ndash708 2008
[76] E S Nejad H Askari and S Soltani ldquoRegulatory TGACG-motif may elicit the secondary metabolite production throughinhibition of active Cyclin-dependent kinaseCyclin complexrdquoPlant Omics Journal vol 5 no 6 pp 553ndash558 2012
[77] L Pauwels K Morreel E De Witte et al ldquoMappingmethyl jasmonate-mediated transcriptional reprogramming ofmetabolism and cell cycle progression in cultured Arabidopsiscellsrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 4 pp 1380ndash1385 2008
[78] C V Dang ldquoMYC on the path to cancerrdquoCell vol 149 no 1 pp22ndash35 2012
[79] G Nestal de Moraes K L Silva F D C Vasconcelos and RC Maia ldquoSurvivin overexpression correlates with an apoptosis-resistant phenotype in chronic myeloid leukemia cellsrdquo Oncol-ogy Reports vol 25 no 6 pp 1613ndash1619 2011
[80] A Rahman T Sultan and R Islam ldquoApoptosis and cancerinsights molecular mechanisms and treatmentsrdquo International
Journal of Biomolecules amp Biomedicine vol 2 no 1 pp 1ndash162012
[81] D-F Suen K L Norris and R J Youle ldquoMitochondrialdynamics and apoptosisrdquo Genes and Development vol 22 no12 pp 1577ndash1590 2008
[82] G Kroemer L Galluzzi and C Brenner ldquoMitochondrial mem-brane permeabilization in cell deathrdquo Physiological Reviews vol87 no 1 pp 99ndash163 2007
[83] T Bender and G C Martinou ldquoWhere killers meetmdashpermeabilization of the outer mitochondrial membrane duringapoptosisrdquoCold Spring Harbor Perspectives in Biology vol 5 no1 Article ID a011106 2013
[84] M H Kang and C P Reynolds ldquoBcI-2 inhibitors targetingmitochondrial apoptotic pathways in cancer therapyrdquo ClinicalCancer Research vol 15 no 4 pp 1126ndash1132 2009
[85] R Kumarswamy and S Chandna ldquoPutative partners in Baxmediated cytochrome-c release ANT CypD VDAC or none ofthemrdquoMitochondrion vol 9 no 1 pp 1ndash8 2009
[86] S S Roy M Madesh E Davies B Antonsson N Danial andG Hajnoczky ldquoBad targets the permeability transition poreindependent of Bax or Bak to switch between Ca2+-dependentcell survival and deathrdquo Molecular Cell vol 33 no 3 pp 377ndash388 2009
[87] S Shimizu M Narita and Y Tsujimoto ldquoBcl-2 family proteinsregulate the release of apoptogenic cytochrome c by the mito-chondrial channel VDACrdquo Nature vol 399 no 6735 pp 483ndash487 1999
[88] E H-Y Cheng T V Sheiko J K Fisher W J Craigenand S J Korsmeyer ldquoVDAC2 inhibits BAK activation andmitochondrial apoptosisrdquo Science vol 301 no 5632 pp 513ndash517 2003
[89] C P Baines R A Kaiser T Sheiko W J Craigen and J DMolkentin ldquoVoltage-dependent anion channels are dispensablefor mitochondrial-dependent cell deathrdquo Nature Cell Biologyvol 9 no 5 pp 550ndash555 2007
[90] P Kreuzaler and C J Watson ldquoKilling a cancer what are thealternativesrdquoNature Reviews Cancer vol 12 no 6 pp 411ndash4242012
[91] N Rivlin R Brosh M Oren and V Rotter ldquoMutations in thep53 tumor suppressor gene important milestones at the varioussteps of tumorigenesisrdquoGenes and Cancer vol 2 no 4 pp 466ndash474 2011
[92] K H Vousden and C Prives ldquoBlinded by the light the growingcomplexity of p53rdquo Cell vol 137 no 3 pp 413ndash431 2009
[93] H Wang K Luo L Z Tan et al ldquoP53-induced gene 3 mediatescell death induced by glutathione peroxidase 3rdquo The Journal ofBiological Chemistry vol 287 no 20 pp 16890ndash16902 2012
[94] T Ozaki and A Nakagawara ldquoP53 the attractive tumor sup-pressor in the cancer research fieldrdquo Journal of Biomedicine andBiotechnology vol 2011 Article ID 603925 13 pages 2011
[95] M Kracikova G Akiri A George R Sachidanandam and SA Aaronson ldquoA threshold mechanism mediates p53 cell fatedecision between growth arrest and apoptosisrdquo Cell Death ampDifferentiation vol 20 no 4 pp 576ndash588 2013
[96] S Fulda and G Kroemer ldquoMitochondria as therapeutic targetsfor the treatment of malignant diseaserdquoAntioxidants and RedoxSignaling vol 15 no 12 pp 2937ndash2949 2011
[97] J Honeychurch W Alduaij M Azizyan et al ldquoAntibody-induced nonapoptotic cell death in human lymphoma andleukemia cells is mediated through a novel reactive oxygenspecies-dependent pathwayrdquo Blood vol 119 no 15 pp 3523ndash3533 2012
International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Cell Biology 21
[98] J D Lambeth ldquoNOX enzymes and the biology of reactiveoxygenrdquo Nature Reviews Immunology vol 4 no 3 pp 181ndash1892004
[99] DGDeavall EAMartin JMHorner andRRoberts ldquoDrug-induced oxidative stress and toxicityrdquo Journal of Toxicology vol2012 Article ID 645460 13 pages 2012
[100] J Verrax N Dejeans B Sid C Glorieux and P B CalderonldquoIntracellular ATP levels determine cell death fate of cancer cellsexposed to both standard and redox chemotherapeutic agentsrdquoBiochemical Pharmacology vol 82 no 11 pp 1540ndash1548 2011
[101] N Festjens T Vanden Berghe and P Vandenabeele ldquoNecrosisa well-orchestrated form of cell demise signalling cascadesimportant mediators and concomitant immune responserdquoBiochimica et Biophysica Acta vol 1757 no 9-10 pp 1371ndash13872006
[102] K M Irrinki K Mallilankaraman R J Thapa et al ldquoRequire-ment of FADD NEMO and BAXBAK for aberrant mitochon-drial function in tumor necrosis factor alpha-induced necrosisrdquoMolecular and Cellular Biology vol 31 no 18 pp 3745ndash37582011
[103] P Vandenabeele L Galluzzi T Vanden Berghe and G Kroe-mer ldquoMolecularmechanisms of necroptosis an ordered cellularexplosionrdquoNature ReviewsMolecular Cell Biology vol 11 no 10pp 700ndash714 2010
[104] D Ezekwudo R Shashidharamurthy D Devineni E BozemanR Palaniappan and P Selvaraj ldquoInhibition of expression ofanti-apoptotic protein Bcl-2 and induction of cell death inradioresistant human prostate adenocarcinoma cell line (PC-3)by methyl jasmonaterdquo Cancer Letters vol 270 no 2 pp 277ndash285 2008
[105] S Klippel J Jakubikova J Delmore et al ldquoMethyl jasmonatedisplays in vitro and in vivo activity against multiple myelomacellsrdquoBritish Journal ofHaematology vol 159 no 3 pp 340ndash3512012
[106] G Manda M T Nechifor and T-M Neagu ldquoReactive oxygenspecies cancer and anti-cancer therapiesrdquo Current ChemicalBiology vol 3 no 1 pp 22ndash46 2009
[107] M S Wolin M Ahmad and S A Gupte ldquoOxidant andredox signaling in vascular oxygen sensing mechanisms basicconcepts current controversies and potential importance ofcytosolic NADPHrdquo American Journal of PhysiologymdashLung Cel-lular and Molecular Physiology vol 289 no 2 pp L159ndashL1732005
[108] M Ushio-Fukai ldquoLocalizing NADPH oxidase-derived ROSrdquoSciencersquos STKE vol 2006 no 349 article re8 2006
[109] K Chen S E Craige and J F Keaney Jr ldquoDownstream tar-gets and intracellular compartmentalization in Nox signalingrdquoAntioxidants amp Redox Signaling vol 11 no 10 pp 2467ndash24802009
[110] C E Paulsen and K S Carroll ldquoOrchestrating redox signalingnetworks through regulatory cysteine switchesrdquo ACS ChemicalBiology vol 5 no 1 pp 47ndash62 2010
[111] KWejksza C Lee-ChangM Bodogai et al ldquoCancer-producedmetabolites of 5-lipoxygenase induce tumor-evoked regulatoryB cells via peroxisome proliferator-activated receptor alphardquoThe Journal of Immunology vol 190 no 6 pp 2575ndash2584 2013
[112] A J Lambert and M D Brand ldquoReactive oxygen speciesproduction by mitochondriardquo Methods in Molecular Biologyvol 554 pp 165ndash181 2009
[113] M Malinouski Y Zhou V V Belousov D L Hatfield and VN Gladyshev ldquoHydrogen peroxide probes directed to different
cellular compartmentsrdquo PLoS ONE vol 6 no 1 Article IDe14564 2011
[114] S S Korshunov V P Skulachev and A A Starkov ldquoHighprotonic potential actuates a mechanism of production ofreactive oxygen species inmitochondriardquo FEBS Letters vol 416no 1 pp 15ndash18 1997
[115] J F Turrens ldquoMitochondrial formation of reactive oxygenspeciesrdquo Journal of Physiology vol 552 no 2 pp 335ndash344 2003
[116] M Le BrasM-V Clement S Pervaiz andC Brenner ldquoReactiveoxygen species and the mitochondrial signaling pathway of celldeathrdquoHistology and Histopathology vol 20 no 1 pp 205ndash2192005
[117] A Chatterjee S Dasgupta and D Sidransky ldquoMitochondrialsubversion in cancerrdquo Cancer Prevention Research vol 4 no 5pp 638ndash654 2011
[118] W Ladiges J Wanagat B Preston L Loeb and P RabinovitchldquoA mitochondrial view of aging reactive oxygen species andmetastatic cancerrdquo Aging Cell vol 9 no 4 pp 462ndash465 2010
[119] E Anso A R Mullen D W Felsher J M Mates R J DeBera-rdinis and N S Chandel ldquoMetabolic changes in cancer cellsupon suppression of MYCrdquo Cancer ampMetabolism vol 1 article7 2013
[120] S J Ralph S Rodrıguez-Enrıquez J Neuzil E Saave-dra and R Moreno-Sanchez ldquoThe causes of cancer revis-ited ldquomitochondrial malignancyrdquo and ROS-induced oncogenictransformationmdashwhy mitochondria are targets for cancer ther-apyrdquo Molecular Aspects of Medicine vol 31 no 2 pp 145ndash1702010
[121] W Bechtel and G Bauer ldquoCatalase protects tumor cells fromapoptosis induction by intercellular ROS signalingrdquo AnticancerResearch vol 29 no 11 pp 4541ndash4557 2009
[122] S J Ralph S Rodrıguez-Enrıquez J Neuzil and R Moreno-Sanchez ldquoBioenergetic pathways in tumor mitochondria astargets for cancer therapy and the importance of the ROS-induced apoptotic triggerrdquo Molecular Aspects of Medicine vol31 no 1 pp 29ndash59 2010
[123] T Ozben ldquoOxidative stress and apoptosis impact on cancertherapyrdquo Journal of Pharmaceutical Sciences vol 96 no 9 pp2181ndash2196 2007
[124] A K Maiti ldquoReactive oxygen species reduction is a key under-lying mechanism of drug resistance in cancer chemotherapyrdquoChemotherapy vol 1 pp 104ndash108 2012
[125] V Loyola-Vargas E Ruiz-May R Galaz-Avalos and C De-la-Pena ldquoThe role of jasmonic acid in root mitochondriadisruptionrdquo Plant Signaling amp Behavior vol 7 no 6 pp 611ndash6142012
[126] M L Orozco-Cardenas J Narvaez-Vasquez and C A RyanldquoHydrogen peroxide acts as a second messenger for the induc-tion of defense genes in tomato plants in response to woundingsystemin and methyl jasmonaterdquo Plant Cell vol 13 no 1 pp179ndash191 2001
[127] G P Bienert J K Schjoerring and T P Jahn ldquoMembranetransport of hydrogen peroxiderdquo Biochimica et Biophysica Actavol 1758 no 8 pp 994ndash1003 2006
[128] K T Hung Y T Hsu and C H Kao ldquoHydrogen peroxideis involved in methyl jasmonate-induced senescence of riceleavesrdquo Physiologia Plantarum vol 127 no 2 pp 293ndash303 2006
[129] F C Kupper E Gaquerel A Cosse et al ldquoFree fatty acidsand methyl jasmonate trigger defense reactions in laminariadigitatardquo Plant and Cell Physiology vol 50 no 4 pp 789ndash8002009
22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

22 International Journal of Cell Biology
[130] S Y Oh J H Kim M J Park et al ldquoInduction of heat shockprotein 72 in C6 glioma cells by methyl jasmonate throughROS-dependent heat shock factor 1 activationrdquo InternationalJournal of Molecular Medicine vol 16 no 5 pp 833ndash839 2005
[131] M Lopez-Lazaro ldquoIncrease in the 998779120595119898produced either by a
closure of the mitochondrial permeability transition pore or byinhibition of the ATP synthaserdquo Cancer Letts vol 252 pp 1ndash82007
[132] N J Davies R E Hayden P J Simpson et al ldquoAKR1C isoformsrepresent a novel cellular target for jasmonates alongside theirmitochondrial-mediated effectsrdquo Cancer Research vol 69 no11 pp 4769ndash4775 2009
[133] V Fritz and L Fajas ldquoMetabolism and proliferation sharecommon regulatory pathways in cancer cellsrdquoOncogene vol 29no 31 pp 4369ndash4377 2010
[134] O Warburg ldquoOn respiratory impairment in cancer cellsrdquoScience vol 124 no 3215 pp 269ndash270 1956
[135] S P Mathupala Y H Ko and P L Pedersen ldquoThe pivotalroles of mitochondria in cancer warburg and beyond andencouraging prospects for effective therapiesrdquo Biochimica etBiophysica Acta vol 1797 no 6-7 pp 1225ndash1230 2010
[136] M L Turcotte H Parliament A Franko and J Allalunis-Turner ldquoVariation in mitochondrial function in hypoxia-sensitive and hypoxia-tolerant human glioma cellsrdquo BritishJournal of Cancer vol 86 no 4 pp 619ndash624 2002
[137] C E Griguer C R Oliva and G Y Gillespie ldquoGlucosemetabolism heterogeneity in human and mouse malignantglioma cell linesrdquo Journal of Neuro-Oncology vol 74 no 2 pp123ndash133 2005
[138] A Wolf S Agnihotri and A Guha ldquoTargeting metabolicremodeling in glioblastoma multiformerdquo Oncotarget vol 1 no7 pp 552ndash562 2010
[139] RMajeed A Hamid Y Qurishi et al ldquoTherapeutic targeting ofcancer cell metabolism role of metabolic enzymes oncogenesand tumor suppressor genesrdquo Journal of Cancer Science ampTherapy vol 4 no 9 pp 281ndash291 2012
[140] J E Wilson ldquoIsozymes of mammalian hexokinase structuresubcellular localization and metabolic functionrdquo Journal ofExperimental Biology vol 206 no 12 pp 2049ndash2057 2003
[141] S John J N Weiss and B Ribalet ldquoSubcellular localization ofhexokinases I and II directs the metabolic fate of glucoserdquo PLoSONE vol 6 no 3 Article ID e17674 2011
[142] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase-2bound to mitochondria cancerrsquos stygian link to the ldquoWarburgeffectrdquo and a pivotal target for effective therapyrdquo Seminars inCancer Biology vol 19 no 1 pp 17ndash24 2009
[143] P L Pedersen S Mathupala A Rempel J F Geschwind and YH Ko ldquoMitochondrial bound type II hexokinase a key player inthe growth and survival of many cancers and an ideal prospectfor therapeutic interventionrdquoBiochimica et Biophysica Acta vol1555 no 1ndash3 pp 14ndash20 2002
[144] R A Nakashima P S Mangan M Colombini and P L Peder-sen ldquoHexokinase receptor complex in hepatomamitochondriaevidence fromNN1015840-dicyclohexylcarbodiimide-labeling studiesfor the involvement of the pore-forming protein VDACrdquoBiochemistry vol 25 no 5 pp 1015ndash1021 1986
[145] P L Pedersen ldquoVoltage dependent anion channels (VDACs) abrief introductionwith a focus on the outermitochondrial com-partmentrsquos roles together with hexokinase-2 in the ldquoWarburgeffectrdquo in cancerrdquo Journal of Bioenergetics and Biomembranesvol 40 no 3 pp 123ndash126 2008
[146] R P Goncalves N Buzhysnskyy and S Scheuring ldquoMinireview on the structure and supramolecular assembly ofVDACrdquo Journal of Bioenergetics and Biomembranes vol 40 no3 pp 133ndash138 2008
[147] V Shoshan-Barmatz and D Mizrachi ldquoVDAC1 from structureto cancer therapyrdquo Frontiers in Oncology vol 2 article 146 2012
[148] C Grills P V Jithesh J Blayney S-D Zhang andD A FennellldquoGene expressionmeta-analysis identifies VDAC1 as a predictorof poor outcome in early stage non-small cell lung cancerrdquo PLoSONE vol 6 no 1 Article ID e14635 2011
[149] LGalluzziOKeppN Tajeddine andGKroemer ldquoDisruptionof the hexokinase-VDAC complex for tumor therapyrdquo Onco-gene vol 27 no 34 pp 4633ndash4635 2008
[150] P L Pedersen ldquoWarburg me and Hexokinase 2 multiplediscoveries of key molecular events underlying one of cancersrsquomost common phenotypes the ldquoWarburg Effectrdquo ie elevatedglycolysis in the presence of oxygenrdquo Journal of Bioenergeticsand Biomembranes vol 39 no 3 pp 211ndash222 2007
[151] J J Lemasters and E Holmuhamedov ldquoVoltage-dependentanion channel (VDAC) asmitochondrial governatormdashthinkingoutside the boxrdquo Biochimica et Biophysica Acta vol 1762 no 2pp 181ndash190 2006
[152] R J Mailloux T Dumouchel C Aguer R Dekemp R Bean-lands and M-E Harper ldquoHexokinase II acts through UCP3 tosuppressmitochondrial reactive oxygen species production andmaintain aerobic respirationrdquo Biochemical Journal vol 437 no2 pp 301ndash311 2011
[153] K J Ahn J Kim M Yun J H Park and J D Lee ldquoEnzymaticproperties of theN-andC-terminal halves of humanhexokinaseIIrdquo BMB Reports vol 42 no 6 pp 350ndash355 2009
[154] NMajewski V Nogueira R B Robey andNHay ldquoAkt inhibitsapoptosis downstream of BID cleavage via a glucose-dependentmechanism involving mitochondrial hexokinasesrdquo Molecularand Cellular Biology vol 24 no 2 pp 730ndash740 2004
[155] R L Elstrom D E Bauer M Buzzai et al ldquoAkt stimulatesaerobic glycolysis in cancer cellsrdquo Cancer Research vol 64 no11 pp 3892ndash3899 2004
[156] JG Pastorino J BHoek andN Shulga ldquoActivation of glycogensynthase kinase 3120573 disrupts the binding of hexokinase IIto mitochondria by phosphorylating voltage-dependent anionchannel and potentiates chemotherapy-induced cytotoxicityrdquoCancer Research vol 65 no 22 pp 10545ndash10554 2005
[157] C Martel M Allouche D D Esposti et al ldquoGSK3-mediatedVDAC phosphorylation controls outer mitochondrial mem-brane permeability during lipid accumulationrdquoHepatology vol57 no 1 pp 93ndash102 2012
[158] C L Neary and J G Pastorino ldquoAkt inhibition promoteshexokinase 2 redistribution and glucose uptake in cancer cellsrdquoJournal of Cellular Physiology vol 228 no 9 pp 1943ndash19482013
[159] J G Pastorino and J B Hoek ldquoHexokinase II the integration ofenergymetabolism and control of apoptosisrdquoCurrentMedicinalChemistry vol 10 no 16 pp 1535ndash1551 2003
[160] S P Mathupala Y H Ko and P L Pedersen ldquoHexokinase IIcancerrsquos double-edged sword acting as both facilitator and gate-keeper ofmalignancy when bound tomitochondriardquoOncogenevol 25 no 34 pp 4777ndash4786 2006
[161] D B Zorov M Juhaszova Y Yaniv H B Nuss S Wang and SJ Sollott ldquoRegulation and pharmacology of the mitochondrialpermeability transition porerdquo Cardiovascular Research vol 83no 2 pp 213ndash225 2009
International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Cell Biology 23
[162] G Kroemer and J Pouyssegur ldquoTumor cell metabolism can-cerrsquos Achillesrsquo heelrdquoCancer Cell vol 13 no 6 pp 472ndash482 2008
[163] S Fulda L Galluzzi and G Kroemer ldquoTargeting mitochondriafor cancer therapyrdquo Nature Reviews Drug Discovery vol 9 no6 pp 447ndash464 2010
[164] A Wolf S Agnihotri J Micallef et al ldquoHexokinase 2 is akey mediator of aerobic glycolysis and promotes tumor growthin human glioblastoma multiformerdquo Journal of ExperimentalMedicine vol 208 no 2 pp 313ndash326 2011
[165] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences vol 109 no 50 pp 20491ndash20496 2012
[166] A L Simons I M Ahmad D M Mattson K J Dornfeldand D R Spitz ldquo2-Deoxy-D-glucose combined with cisplatinenhances cytotoxicity via metabolic oxidative stress in humanhead and neck cancer cellsrdquo Cancer Research vol 67 no 7 pp3364ndash3370 2007
[167] L S Ihrlund E Hernlund O Khan and M C Shoshan ldquo3-Bromopyruvate as inhibitor of tumour cell energy metabolismand chemopotentiator of platinum drugsrdquoMolecular Oncologyvol 2 no 1 pp 94ndash101 2008
[168] W Kim J-H Yoon J-M Jeong et al ldquoApoptosis-inducingantitumor efficacy of hexokinase II inhibitor in hepatocellularcarcinomardquo Molecular Cancer Therapeutics vol 6 no 9 pp2554ndash2562 2007
[169] F Chiara D Castellaro O Marin et al ldquoHexokinase IIdetachment from mitochondria triggers apoptosis through thepermeability transition pore independent of voltage-dependentanion channelsrdquo PLoS ONE vol 3 no 3 Article ID e1852 2008
[170] A Heyfets and E Flescher ldquoCooperative cytotoxicity of methyljasmonatewith anti-cancer drugs and 2-deoxy-d-glucoserdquoCan-cer Letters vol 250 no 2 pp 300ndash310 2007
[171] TThalhamerM AMcGrath andMMHarnett ldquoMAPKs andtheir relevance to arthritis and inflammationrdquo Rheumatologyvol 47 no 4 pp 409ndash414 2008
[172] S Tamura M Hanada M Ohnishi K Katsura M Sasakiand T Kobayashi ldquoRegulation of stress-activated protein kinasesignaling pathways by protein phosphatasesrdquo European Journalof Biochemistry vol 269 no 4 pp 1060ndash1066 2002
[173] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[174] Y T Ip and R J Davis ldquoSignal transduction by the c-Jun N-terminal kinase (JNK)mdashfrom inflammation to developmentrdquoCurrent Opinion in Cell Biology vol 10 no 2 pp 205ndash219 1998
[175] Y Shi and M Gaestel ldquoIn the cellular garden of forking pathshow p38 MAPKs signal for downstream assistancerdquo BiologicalChemistry vol 383 no 10 pp 1519ndash1536 2002
[176] J H Weisburg ldquoMultidrug resistance in acute myeloidleukemia potential new therapeuticsrdquo Journal of NuclearMedicine vol 49 no 9 pp 1405ndash1407 2008
[177] G Trinchieri ldquoInnate inflammation and cancer is it time forcancer preventionrdquo F1000Medicine Reports vol 3 no 1 article11 2011
[178] H T Dang H J Lee E S Yoo et al ldquoNew jasmonateanalogues as potential anti-inflammatory agentsrdquo Bioorganicand Medicinal Chemistry vol 16 no 24 pp 10228ndash10235 2008
[179] H-J Lee KMaengH-TDang et al ldquoAnti-inflammatory effectof methyl dehydrojasmonate (J2) is mediated by the NF-120581B
pathwayrdquo Journal ofMolecularMedicine vol 89 no 1 pp 83ndash902011
[180] H Ohnishi N Miyahara and E W Gelfand ldquoThe role ofleukotriene B4 in allergic diseasesrdquo Allergology Internationalvol 57 no 4 pp 291ndash298 2008
[181] V Mugoni and M M Santoro ldquoManipulating redox signalingto block tumor angiogenesisrdquo in Research Directions in TumorAngiogenesis chapter 3 pp 47ndash72 2013
[182] D X Nguyen P D Bos and J Massague ldquoMetastasis fromdissemination to organ-specific colonizationrdquo Nature ReviewsCancer vol 9 no 4 pp 274ndash284 2009
[183] Y Jin and T M Penning ldquoAldo-keto reductases and bioac-tivationdetoxicationrdquo Annual Review of Pharmacology andToxicology vol 47 pp 263ndash292 2007
[184] K Matsuura H Shiraishi A Hara et al ldquoIdentification of aprincipal mRNA species for human 3120572-hydroxysteroid dehy-drogenase isoform (AKR1C3) that exhibits high prostaglandinD2 11-ketoreductase activityrdquo Journal of Biochemistry vol 124no 5 pp 940ndash946 1998
[185] T M Penning and J E Drury ldquoHuman aldo-keto reductasesfunction gene regulation and single nucleotide polymor-phismsrdquo Archives of Biochemistry and Biophysics vol 464 no2 pp 241ndash250 2007
[186] T Matsunaga Y Wada S Endo M Soda O El-Kabbani andA Hara ldquoAldo-keto reductase 1B10 and its role in proliferationcapacity of drug-resistant cancersrdquo Frontiers in Pharmacologyvol 3 article 5 2012
[187] N-Y Hsu H-C Ho K-C Chow et al ldquoOverexpression ofdihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancerrdquo Cancer Research vol 61 no 6 pp 2727ndash2731 2001
[188] P Vihko A Herrala P Harkonen et al ldquoEnzymes as modula-tors in malignant transformationrdquo Journal of Steroid Biochem-istry and Molecular Biology vol 93 no 2-5 pp 277ndash283 2005
[189] H-L Tai T-S Lin H-H Huang et al ldquoOverexpression ofaldo-keto reductase 1C2 as a high-risk factor in bladder cancerrdquoOncology Reports vol 17 no 2 pp 305ndash311 2007
[190] H B Deng H K Parekh K-C Chow and H SimpkinsldquoIncreased expression of dihydrodiol dehydrogenase inducesresistance to cisplatin in human ovarian carcinoma cellsrdquo TheJournal of Biological Chemistry vol 277 no 17 pp 15035ndash150432002
[191] Y-J Chen C-C Yuan K-C Chow et al ldquoOverexpressionof dihydrodiol dehydrogenase is associated with cisplatin-based chemotherapy resistance in ovarian cancer patientsrdquoGynecologic Oncology vol 97 no 1 pp 110ndash117 2005
[192] W D Chen and Y Zhang ldquoRegulation of aldo-keto reductasesin human diseasesrdquo Frontiers in Pharmacology vol 3 article 352012
[193] W Ax M Soldan L Koch and E Maser ldquoDevelopmentof daunorubicin resistance in tumour cells by induction ofcarbonyl reductionrdquo Biochemical Pharmacology vol 59 no 3pp 293ndash300 2000
[194] E K Lee W T Regenold and P Shapiro ldquoInhibition of aldosereductase enhances Hela cell sensitivity to chemotherapeuticdrugs and involves activation of extracellular signal-regulatedkinasesrdquo Anti-Cancer Drugs vol 13 no 8 pp 859ndash868 2002
[195] M Maccarrone ldquoLipoxygenases apoptosis and the role ofantioxidantsrdquo in Photoprotection Photoinhibition Gene Regula-tion and Environment vol 21 ofAdvances in Photosynthesis andRespiration pp 321ndash332 2008
24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

24 International Journal of Cell Biology
[196] A R Brash ldquoLipoxygenases occurrence functions catalysisand acquisition of substraterdquo The Journal of Biological Chem-istry vol 274 no 34 pp 23679ndash23682 1999
[197] M C Meyer P Rastogi C S Beckett and J McHowatldquoPhospholipase A2 inhibitors as potential anti-inflammatoryagentsrdquo Current Pharmaceutical Design vol 11 no 10 pp 1301ndash1312 2005
[198] M M Bosma-den Boer M-L vanWetten and L PruimboomldquoChronic inflammatory diseases are stimulated by currentlifestyle howdiet stress levels andmedication prevent our bodyfrom recoveringrdquo Nutrition amp Metabolism vol 9 article 322012
[199] O Werz J Klemm B Samuelsson and O Radmark ldquo5-Lipoxygenase is phosphorylated by p38 kinase-dependentMAPKAP kinasesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 97 no 10 pp 5261ndash5266 2000
[200] W-G Tong X-Z Ding R C Witt and T E Adrian ldquoLipoxy-genase inhibitors attenuate growth of human pancreatic cancerxenografts and induce apoptosis through the mitochondrialpathwayrdquoMolecular Cancer Therapeutics vol 1 no 11 pp 929ndash935 2002
[201] W-G Tong X-Z Ding and T E Adrian ldquoThe mechanismsof lipoxygenase inhibitor-induced apoptosis in human breastcancer cellsrdquo Biochemical and Biophysical Research Communi-cations vol 296 no 4 pp 942ndash948 2002
[202] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[203] W-G Tong X-Z Ding M S Talamonti R H Bell and T EAdrian ldquoLTB4 stimulates growth of human pancreatic cancercells via MAPK and PI-3 kinase pathwaysrdquo Biochemical andBiophysical Research Communications vol 335 no 3 pp 949ndash956 2005
[204] J Ghosh and C E Myers ldquoArachidonic acid stimulates prostatecancer cell growth critical role of 5-lipoxygenaserdquo Biochemicaland Biophysical Research Communications vol 235 no 2 pp418ndash423 1997
[205] J T OrsquoFlaherty L C Rogers B A Chadwell et al ldquo5(S)-hydroxy-681114-EZZZ-eicosatetraenoate stimulates PC3 cellsignaling and growth by a receptor-dependent mechanismrdquoCancer Research vol 62 no 23 pp 6817ndash6819 2002
[206] Y-N Ye E S-L Liu V Y Shin W K-K Wu and C-HCho ldquoContributory role of 5-lipoxygenase and its associationwith angiogenesis in the promotion of inflammation-associatedcolonic tumorigenesis by cigarette smokingrdquo Toxicology vol203 no 1ndash3 pp 179ndash188 2004
[207] K M Anderson T Seed M Vos et al ldquo5-Lipoxygenaseinhibitors reduce PC-3 cell proliferation and initiate non-necrotic cell deathrdquo Prostate vol 37 no 3 pp 161ndash173 1998
[208] I Avis S H Hong A Martinez et al ldquoFive-lipoxygenaseinhibitors can mediate apoptosis in human breast cancer celllines through complex eicosanoid interactionsrdquo The FASEBJournal vol 15 no 11 pp 2007ndash2009 2001
[209] X M Fan S P Tu S K Lam et al ldquoFive-lipoxygenase-activating protein inhibitor MK-886 induces apoptosis in gas-tric cancer through upregulation of p27kip1 and baxrdquo Journal ofGastroenterology and Hepatology vol 19 no 1 pp 31ndash37 2004
[210] M Kovarikova J Hofmanova K Soucek and A KozubikldquoThe effects of TNF-alpha and inhibitors of arachidonic acid
metabolism on human colon HT-29 cells depend on differen-tiation statusrdquo Differentiation vol 72 no 1 pp 23ndash31 2004
[211] A Hoque S M Lippman T-T Wu et al ldquoIncreased 5-lipoxygenase expression and induction of apoptosis by itsinhibitors in esophageal cancer a potential target for preven-tionrdquo Carcinogenesis vol 26 no 4 pp 785ndash791 2005
[212] X Chen S Sood C S Yang N Li and Z Sun ldquoFive-lipoxygenase pathway of arachidonic acid metabolism in car-cinogenesis and cancer chemopreventionrdquo Current CancerDrug Targets vol 6 no 7 pp 613ndash622 2006
[213] S Gupta M Strivastva N Ahmad K Sakamoto D GBostwick and H Muktar ldquoLipoxygenase-5 is overexpressed inprostate adenocarcinomardquo Cancer vol 91 pp 737ndash743 2001
[214] R Yoshimura M Matsuyama K Tsuchida Y Kawahito HSano and T Nakatani ldquoExpression of lipoxygenase in humanbladder carcinoma and growth inhibition by its inhibitorsrdquoJournal of Urology vol 170 no 5 pp 1994ndash1999 2003
[215] H Zhi J Zhang G Hu et al ldquoThe deregulation of arachidonicacid metabolism-related genes in human esophageal squamouscell carcinomardquo International Journal of Cancer vol 106 no 3pp 327ndash333 2003
[216] R Hennig X-Z Ding W-G Tong et al ldquo5-Lipoxygenase andleukotriene B4 receptor are expressed in human pancreaticcancers but not in pancreatic ducts in normal tissuerdquo AmericanJournal of Pathology vol 161 no 2 pp 421ndash428 2002
[217] L-Y Zou J-Y Li F-L Chen Z-X Chen and X-Z WangldquoTumor 5-lipoxygenase expression correlates with gastric can-cer metastasis and its selective inhibitor induces cancer cellapoptosisrdquo Journal of Cancer Molecules vol 2 no 6 pp 227ndash233 2006
[218] M Romano A Catalano M Nutini et al ldquo5-Lipoxygenaseregulates malignant mesothelial cell survival involvement ofvascular endothelial growth factorrdquoThe FASEB Journal vol 15no 13 pp 2326ndash2336 2001
[219] I M Avis M Jett T Boyle et al ldquoGrowth control of lungcancer by interruption of 5-lipoxygenase-mediated growthfactor signalingrdquoThe Journal of Clinical Investigation vol 97 no3 pp 806ndash813 1996
[220] R J Boado W M Pardridge H V Vinters and K L BlackldquoDifferential expression of arachidonate 5-lipoxygenase tran-scripts in human brain tumors evidence for the expression ofa multitranscript familyrdquo Proceedings of the National Academyof Sciences of the United States of America vol 89 no 19 pp9044ndash9048 1992
[221] C V Rao A Rivenson B Simi et al ldquoChemoprevention ofcolon carcinogenesis by sulindac a nonsteroidal anti- inflam-matory agentrdquo Cancer Research vol 55 no 7 pp 1464ndash14721995
[222] X-M Xu J-J Deng G-J Yuan et al ldquo5-Lipoxygenase con-tributes to the progression of hepatocellular carcinomardquoMolec-ular Medicine Reports vol 4 no 6 pp 1195ndash1200 2011
[223] B Sveinbjornsson A Rasmuson N Baryawno et al ldquoExpres-sion of enzymes and receptors of the leukotriene pathway inhuman neuroblastoma promotes tumor survival and provides atarget for therapyrdquoThe FASEB Journal vol 22 no 10 pp 3525ndash3536 2008
[224] L T Soumaoro S Iida H Uetake et al ldquoExpression of 5-lipoxygenase in human colorectal cancerrdquo World Journal ofGastroenterology vol 12 no 39 pp 6355ndash6360 2006
[225] K Ishii M Zaitsu N Yonemitsu Y Kan Y Hamasaki and MMatsuo ldquo5-lipoxygenase pathway promotes cell proliferation in
International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Cell Biology 25
human glioma cell linesrdquoClinical Neuropathology vol 28 no 6pp 445ndash452 2009
[226] P C Boger J D Shutt and J R Neale ldquoIncreased expressionof the 5-lipoxygenase pathway and its cellular localization inBarrettrsquos adenocarcinomardquo Histopathology vol 61 pp 509ndash5172012
[227] Z Sun S Sood N Li et al ldquoInvolvement of the 5-lipoxy-genaseleukotriene A4 hydrolase pathway in 712-dimeth-ylbenz[a]anthracene (DMBA)-induced oral carcinogenesis inhamster cheek pouch and inhibition of carcinogenesis by itsinhibitorsrdquo Carcinogenesis vol 27 no 9 pp 1902ndash1908 2006
[228] R C Goupil J J Bushey J Peters-Kennedy and J J WakshlagldquoPrevalence of 5-lipoxygenase expression in canine osteosar-coma and the effects of a dual 5-lipoxygenasecyclooxygenaseinhibitor on osteosarcoma cells in vitro and in vivordquo VeterinaryPathology vol 49 no 5 pp 802ndash810 2012
[229] H J Hussey andM J Tisdale ldquoInhibition of tumour growth bylipoxygenase inhibitorsrdquo British Journal of Cancer vol 74 no 5pp 683ndash687 1996
[230] X-Z Ding P Iversen M W Cluck J A Knezetic and TE Adrian ldquoLipoxygenase inhibitors abolish proliferation ofhuman pancreatic cancer cellsrdquo Biochemical and BiophysicalResearch Communications vol 261 no 1 pp 218ndash223 1999
[231] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[232] J Ghosh and C E Myers ldquoInhibition of arachidonate 5-lipoxygenase triggers massive apoptosis in human prostatecancer cellsrdquo Proceedings of the National Academy of Sciencesof the United States of America vol 95 no 22 pp 13182ndash131871998
[233] T A Miller J Ghosh C E Myers and T L Macdonald ldquo5-HETE congeners asmodulators of cell proliferationrdquoBioorganicand Medicinal Chemistry Letters vol 10 no 17 pp 1913ndash19162000
[234] J Ghosh ldquoInhibition of arachidonate 5-lipoxygenase triggersprostate cancer cell death through rapid activation of c-Jun N-terminal kinaserdquoBiochemical and Biophysical Research Commu-nications vol 307 no 2 pp 342ndash349 2003
[235] M Maccarrone G Melino and A Finazzi-Agro ldquoLipoxyge-nases and their involvement in programmed cell deathrdquo CellDeath and Differentiation vol 8 no 8 pp 776ndash784 2001
[236] M Luo S Lee and T G Brock ldquoLeukotriene synthesis byepithelial cellsrdquo Histology and Histopathology vol 18 no 2 pp587ndash595 2003
[237] H Hanaka T Shimizu and T Izumi ldquoStress-induced nuclearexport of 5-lipoxygenaserdquo Biochemical and Biophysical ResearchCommunications vol 338 no 1 pp 111ndash116 2005
[238] O Werz and D Steinhilber ldquoTherapeutic options for 5-lipoxygenase inhibitorsrdquo Pharmacology and Therapeutics vol112 no 3 pp 701ndash718 2006
[239] A S Fischer J Metzner S D Steinbrink et al ldquo5-Lipoxygenaseinhibitors induce potent anti-proliferative and cytotoxic effectsin human tumour cells independently of suppression of 5-lipoxygenase activityrdquo British Journal of Pharmacology vol 161no 4 pp 936ndash949 2010
[240] A M Bair M V Turman C A Vaine R A Panettieri and RJ Soberman Jr ldquoThe nuclear membrane leukotriene syntheticcomplex is a signal integrator and transducerrdquo Molecular Biol-ogy of the Cell vol 23 no 22 pp 4456ndash4464 2012
[241] A Russo C L Espinoza S Caggia et al ldquoA new jasmonic acidstereoisomeric derivative induces apoptosis via reactive oxygenspecies in human prostate cancer cellsrdquo Cancer Letters vol 326no 2 pp 199ndash205 2012
[242] L J Marnett ldquoInflammation and cancer chemical approachestomechanisms imaging and treatmentrdquoThe Journal of OrganicChemistry vol 77 no 12 pp 5224ndash5238 2012
[243] P Pratheeshkumar C Sreekala Z Zhang et al ldquoCancerprevention with promising natural products mechanisms ofaction and molecular targetsrdquo Anti-Cancer Agents in MedicinalChemistry vol 12 no 10 pp 1159ndash1184 2012
[244] E L Kwak J W Clark and B Chabner ldquoTargeted agents therules of combinationrdquo Clinical Cancer Research vol 13 no 18pp 5232ndash5237 2007
[245] G Ghavami M R Kazemali and S Sardari ldquoInformaticsof drug synergism in naturally occurring anticancer agentsrdquoRecent Patents on Anti-Cancer Drug Discovery vol 6 no 1 pp26ndash44 2011
[246] A Mohan S Narayanan S Sethuraman and U M KrishnanldquoCombinations of plant polyphenols amp anti-cancer molecules anovel treatment strategy for cancer chemotherapyrdquoAnti-CancerAgents in Medicinal Chemistry vol 13 no 2 pp 281ndash295 2013
[247] R Takimoto J Kato T Terui et al ldquoAugmentation of antitu-mor effects of p53 gene therapy by combination with HDACinhibitorrdquo Cancer Biology and Therapy vol 4 no 4 pp 421ndash428 2005
[248] B S Vinod T T Maliekal and R J Anto ldquoPhytochemicalsas chemosensitizers from molecular mechanism to clinicalsignificancerdquo Antioxidants amp Redox Signaling vol 18 no 11 pp1307ndash1348 2013
[249] S Cai Y Xu R J Cooper et al ldquoMitochondrial targetingof human O6-methylguanine DNA methyltransferase protectsagainst cell killing by chemotherapeutic alkylating agentsrdquoCancer Research vol 65 no 8 pp 3319ndash3327 2005
[250] W P Tsang S P Y Chau S K Kong K P Fung and T TKwok ldquoReactive oxygen species mediate doxorubicin inducedp53-independent apoptosisrdquo Life Sciences vol 73 no 16 pp2047ndash2058 2003
[251] L Yeruva C Hall J A Elegbede and S W Carper ldquoPerillylalcohol andmethyl jasmonate sensitize cancer cells to cisplatinrdquoAnti-Cancer Drugs vol 21 no 1 pp 1ndash9 2010
[252] A Maccio and C Madeddu ldquoCisplatin an old drug with anewfound efficacymdashfrom mechanisms of action to cytotoxic-ityrdquo Expert Opinion on Pharmacotherapy vol 14 no 13 pp1839ndash1857 2013
[253] A A Farooqi G Butt and Z Razzaq ldquoAlgae extracts andmethyl jasmonate anti-cancer activities in prostate cancerchoreographers of lsquothe dance macabrersquordquo Cancer Cell Interna-tional vol 12 article 50 2012
[254] X-Y Xiao G-S Jiang L Wang L Lv and F-Q ZengldquoPredominant enhancement of apoptosis induced by methyljasmonate in bladder cancer cells therapeutic effect of theAntp-conjugated Smac peptiderdquo Anti-Cancer Drugs vol 22 no 9 pp853ndash863 2011
[255] G Jiang J Zhao X Xiao et al ldquoAN N-terminal Smac peptidesensitizes human prostate carcinoma cells tomethyl jasmonate-induced apoptosisrdquo Cancer Letters vol 302 no 1 pp 37ndash462011
[256] C C G Hernandes S C Franca A L Fachin M Marinsand M V Lourenco ldquoCytotoxic effect of jasmonate and methyljasmonate on a canine macrophage tumor cell linerdquo A RevistaBrasileira de Plantas Medicinais vol 14 no 1 pp 122ndash124 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology




![6-Methyl-1-({[(2 E )-2-methyl-3-phenylprop-2-en-1-yl]oxy}methyl)-1,2,3,4-tetrahydroquinazoline-2,4-dione](https://img.dokumen.tips/doc/110x75/6324d4e17fd2bfd0cb0325dc/6-methyl-1-2-e-2-methyl-3-phenylprop-2-en-1-yloxymethyl-1234-tetrahydroquinazoline-24-dione.jpg)

















![4-[(4-Methylbenzenesulfonamido)methyl]cyclohexanecarboxylic acid](https://img.dokumen.tips/doc/110x75/632f56bff9bd4ec430033593/4-4-methylbenzenesulfonamidomethylcyclohexanecarboxylic-acid.jpg)
![Methyl 2-{[(4-hydroxyphenyl)(methoxycarbonyl)methyl]aminocarbonyl}ethanoate hemihydrate](https://img.dokumen.tips/doc/110x75/635b87b19d85dc43cb077e8b/methyl-2-4-hydroxyphenylmethoxycarbonylmethylaminocarbonylethanoate-hemihydrate.jpg)
![Methyl 3-[(1-adamantylcarbonyloxy)aminocarbonyl]propanoate](https://img.dokumen.tips/doc/110x75/63239b3b03238a9ff60a820f/methyl-3-1-adamantylcarbonyloxyaminocarbonylpropanoate.jpg)


![A nonmerohedral twin of methyl 2-[methyl(methylsulfonyl)amino]benzoate](https://img.dokumen.tips/doc/110x75/634fd30e01c04712f406e21c/a-nonmerohedral-twin-of-methyl-2-methylmethylsulfonylaminobenzoate.jpg)

