Embed Size (px)
Citation preview

This is an electronic reprint of the original article.This reprint may differ from the original in pagination and typographic detail.
Powered by TCPDF (www.tcpdf.org)
This material is protected by copyright and other intellectual property rights, and duplication or sale of all or part of any of the repository collections is not permitted, except that material may be duplicated by you for your research use or educational purposes in electronic or print form. You must obtain permission for any other use. Electronic or print copies may not be offered, whether for sale or otherwise to anyone who is not an authorised user.
Bhattacharjee, Rahul; Das, Sabya Sachi; Biswal, Smruti Sudha; Nath, Arijit; Das, Debangshi;Basu, Asmita; Malik, Sumira; Kumar, Lamha; Kar, Sulagna; Singh, Sandeep Kumar;Upadhye, Vijay Jagdish; Iqbal, Danish; Almojam, Suliman; Roychoudhury, Shubhadeep;Ojha, Shreesh; Ruokolainen, Janne; Jha, Niraj Kumar; Kesari, Kavindra KumarMechanistic role of HPV-associated early proteins in cervical cancer
DOI:10.1016/j.critrevonc.2022.103675
Published: 01/06/2022
Document VersionPublisher's PDF, also known as Version of record
Published under the following license:CC BY
Please cite the original version:Bhattacharjee, R., Das, S. S., Biswal, S. S., Nath, A., Das, D., Basu, A., Malik, S., Kumar, L., Kar, S., Singh, S.K., Upadhye, V. J., Iqbal, D., Almojam, S., Roychoudhury, S., Ojha, S., Ruokolainen, J., Jha, N. K., & Kesari, K.K. (2022). Mechanistic role of HPV-associated early proteins in cervical cancer: Molecular pathways andtargeted therapeutic strategies. Critical Reviews in Oncology/Hematology, 174, 1-17. [103675].https://doi.org/10.1016/j.critrevonc.2022.103675

Critical Reviews in Oncology / Hematology 174 (2022) 103675
Available online 4 April 20221040-8428/© 2022 The Author(s). Published by Elsevier B.V. This is an open access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/).
Mechanistic role of HPV-associated early proteins in cervical cancer: Molecular pathways and targeted therapeutic strategies
Rahul Bhattacharjee a,1, Sabya Sachi Das b,1, Smruti Sudha Biswal c, Arijit Nath a, Debangshi Das d, Asmita Basu a, Sumira Malik e, Lamha Kumar f, Sulagna Kar a, Sandeep Kumar Singh b, Vijay Jagdish Upadhye g, Danish Iqbal h, Suliman Almojam i, Shubhadeep Roychoudhury j, Shreesh Ojha k, Janne Ruokolainen l, Niraj Kumar Jha m,n,*,1, Kavindra Kumar Kesari l,o,**,2
a School of Biotechnology, Kalinga Institute of Industrial Technology (KIIT), Deemed to be University, Bhubaneswar, Odisha 751024, India b Department of Pharmaceutical Sciences and Technology, Birla Institute of Technology, Mesra, Ranchi, Jharkhand 835215, India c Department of Life Science, National Institute of Technology Rourkela, Rourkela 769008, India d Department of Biotechnology, Haldia Institute of Technology, Affiliated to Maulana Abul Kalam Azad University of Technology, Haldia, West Bengal 721657, India e Amity Institute of Biotechnology, Amity University Jharkhand, Ranchi, Jharkhand 834001, India f School of Biology, Indian Institute of Science Education and Research, Thiruvananthapuram, India g Center of Research for Development (CR4D), Parul Institute of Applied Sciences (PIAS), PO Limda, Tal Waghodia 391760 Vadodara Gujarat h Department of Medical Laboratory Sciences, College of Applied Medical Sciences, Majmaah University, Al-Majmaah 11952, Saudi Arabia i Vector-borne & Zoonotic Diseases General Directorate, Ministry of Health, Riyadh 11176, Saudi Arabia j Department of Life Science and Bioinformatics, Assam University, Silchar, India k Department of Pharmacology and Therapeutics, College of Medicine and Health Sciences, United Arab Emirates University, P.O. Box: 15551 Al Ain, United Arab Emirates l Department of Applied Physics, School of Science, Aalto University, Espoo 00076, Finland m Department of Biotechnology, School of Engineering & Technology (SET), Sharda University, Greater Noida 201306, India n Department of Biotechnology, School of Applied & Life Sciences (SALS), Uttaranchal University, Dehradun 248007, India o Department of Bio-products and Bio-systems, School of Chemical Engineering, Aalto University, Espoo 00076, Finland
A R T I C L E I N F O
Keywords: HPV E proteins Cervical cancer Signaling pathways Therapeutic targets Molecular mechanisms
A B S T R A C T
Purpose: Cervical cancer (CC), one of the major causes of death of women throughout the world is primarily caused due to Human Papilloma Virus (HPV) 16 and 18. The early region (E) oncoproteins of HPV are associated with the etiopathogenesis and contribute to the progression of cancer. The present article comprehensively discussed the structural organization and biological functions of all E proteins of HPV and their contribution to progression of CC with an intent to decipher the pathological hallmarks and their relationship. Additionally, the role of E proteins in reference to therapeutics will also be presented. Methods: A systematic search has been carried out for articles published in PubMed database by using combi-nations of different keywords with Boolean operators (AND, OR, NOT) including cervical cancer, HPV, E pro-teins, and signaling. Results: From the analysis of literature review, its apparent that E proteins are the major contributor to disease progression. E1, E2, and E4 forms are mainly associated with viral integration, replication, and transcription whereas E6 and E7 act as an oncoprotein and are associated with the progression of cancer. E5 regulates cell proliferation, apoptosis, and facilitates the activity of E6 and E7. Additionally, E proteins were observed asso-ciated with numerous cell signaling pathways including PI3K/AKT, Wnt, Notch and reasonably contribute to the initiation of malignancy, cell proliferation, metastasis, and drug resistance. Knowing the role and interplay of each protein in initiation to progression of CC, their therapeutic significance has been elucidated. The present study observations demonstrate that E6 and E7 are the major cause of HPV-mediated CC progression. E1, E2, and
* Corresponding author at: Department of Biotechnology, School of Engineering & Technology (SET), Sharda University, Greater Noida 201306, India. ** Corresponding author at: Department of Applied Physics, School of Science, Aalto University, Espoo 00076, Finland.
E-mail addresses: [email protected] (N.K. Jha), [email protected] (K.K. Kesari). 1 These authors contributed equally to this work. 2 ORCID: 0000-0003-3622-9555
Contents lists available at ScienceDirect
Critical Reviews in Oncology / Hematology
journal homepage: www.elsevier.com/locate/critrevonc
https://doi.org/10.1016/j.critrevonc.2022.103675 Received 3 January 2022; Received in revised form 26 March 2022; Accepted 30 March 2022

Critical Reviews in Oncology / Hematology 174 (2022) 103675
2
E5 also act as a backbone for E6 and E7 and most of the current approaches have targeted E6 and E7 mediated action only. Conclusion: The present review illustrates the structural organization as well as function and regulation of all early proteins of HPV and their association with several cellular signaling pathways. The observations provide clue on the regulatory aspect of these proteins in initiation to progression and reasonably represent that targeting these proteins could be a novel therapeutic strategy for CC. In particular, its seemingly appears that inhibition of the activity of E6 and E7 oncoproteins may be a better selective target to delay the progression of CC. The review reaffirms the role of E proteins and encourages future studies on developing diagnostics, and most importantly therapeutics strategies targeting E6 and E7 oncoproteins to tackle CC related morbidity and mortality.
1. Introduction
One of the most common cancers in women is cervical cancer (CC). There were approximately 570,000 new cases of CC and 311,000 deaths occurred because of CC worldwide in 2018 according to WHO (Adiga et al., 2021; Arbyn et al., 2020; Bray et al., 2018; Galvao and Coimbra, 2020; Razavi, Z. S. et al., 2021). The average age at diagnosis and death of CC has been reported 53 years and 59 years, respectively. According to GLOBOCAN reports, it has been observed that approximately 85% of all CC deaths occurred in low-income countries. The death asssociated with CC in these countries are reported 18 times higher than the developed countries (Bray et al., 2018). Women from Eastern and Southern Africa has been reportd to exhibit higher incidences (Arbyn et al., 2020). Whereas, more than a third of the CC cases globally were reported in Asian countries, China, and India (Bray et al., 2018; Ferlay et al., 2019). India reported approximately 74,000 deaths and 132,000 new cases per year. Majority of the causes of mortality due to CC in developing and underdeveloped countries has been found associated with the shortage of efficient treatment and diagnosis, lack of proper screening programs and awareness, along with increased exposure to the risk factors (Bobdey et al., 2016).
Human Papillomavirus (HPV) has been identified as a primary reason behind CC. Despite being a preventable disease by using thera-peutic strategies, CC has been one of the major gynecological malig-nancies around the world (Paavonen, 2007). Recently, Adiga and co-workers have reviewed and discussed the various aspects of disease including pathogenesis, sequelae, major molecular mechanisms and possible therapeutic targets and associated challenges with CC (Adiga et al., 2021). Although persistent HPV infection is one of the primary requisite for induction of CC, but only HPV cannot develop the CC; early coitus, promiscuity, usage of oral contraceptives, early age pregnancy, tobacco-smoking, unhygienic conditions also contribute to the pro-gression of CC. German virologist Harald Zur Hausen and his team have first established the relationship between HPV and CC in 1983 (Zur, 2002). Further, it has been reported that HPV infection is often seen in healthy women but rapid immune clearance commonly clears it within 1–2 years (Asiaf et al., 2014). However, sexually transmitted persistent HPV infection is the common causative agent of CC in most of the cases (84%) (Organization, 2017). Among 300 different HPV genotypes, the strains that are mostly associated with the progression of CC and adenocarcinoma are HPV16 and HPV18 (Chaturvedi, 2010; Tian, R. et al., 2022). High risk HPV (HR-HPV) types have been shown associated to the progression of CC and precancerous lesions besides HPV16 and 18 include 68, 66, 59, 58, 56, 52, 51, 45, 39, 35, 33, 33 and 31 (Fani et al., 2020).
In this review article, the structural organization, and biological functions of all the E proteins of HPV and their role in the initiation and progression of CC and their therapeutic significance have been comprehensively discussed. Additionally, using synoptic tables and schematic illustrations we have also discussed the signaling networking cascade of the HPV infection for CC via early proteins and the crosstalk between early proteins. Table 1 summarizes the mechanism of early protein for CC progression, the pathogenic hallmarks, influence over cellular signaling and their therapeutic potential.
2. HPV genome
HPV, a papillomaviridae family virus, belongs to a category of double-stranded DNA, non-enveloped, small (50–55 nm) viruses. HPV is divided into five sub-categories (alpha, beta, gamma, mu, nu) based on the nucleotide sequence of the L1 ORF region (de Villiers et al., 2004). Among these, alpha categories were found to be carcinogenic. HPV16 is the most frequently detected genotype in CC cases, followed by HPV18 according to reports from the International Agency for Research on Cancer (IARC) (Serrano et al., 2018).
The HPV16 genome stretches to a length of 7.9 kb. The HPV genome is contained in an icosahedral capsid that is further sub-divided into three regions: the early (E) region comprising different types of pre-cursor proteins including E1, E2, E4, E5, E6, and E7 that are required in the early stages of infection and associated with viral DNA replication, cell cycle control, and oncogenesis (Williams et al., 2011). The major and minor viral capsid structure is coded by the late (L) region proteins which comprise of two parts, L1 and L2. Viral replication and tran-scription are controlled by a non-coding region, the Long Control Region (LCR) (Graham, 2006). Among the six E proteins, E6 and E7 are the main regulators contributes to the pathogenicty of the virus. The cell repairing mechanisms, especially the functions of the tumor suppressor proteins – p53 and retinoblastoma are dysregulated by the expression of the E6 and E7 viral oncoproteins (Crook et al., 1991; Jones and Münger, 1996).
3. Role of E proteins in HPV integration
The initiation of the HPV infection is from the basal layer of the squamous epithelium. The viral replication process and transcription of other E proteins are regulated by E1 and E2 proteins (Pal and Kundu, 2020a). Initially, the copy number tends less in number; however, the copy number increase with the differentiation of basal cells to form the epithelial suprabasal layer and spread the infection to the adjacent cells. The integration of the HPV genome with the host genome is well re-ported (83% HPV-positive cases), but it has been also observed in an episomal form (Network, 2017). E2 disruption occurs due to the inte-gration of the viral genome that activates the E6 and E7 oncoproteins. The viral genome is multiplied by the active assistance of E6 and E7 with the help of cellular machinery. During the process of viral replication, the cells can be tricked to become oncogenic (Pal and Kundu, 2020a). The collateral damage to viral replication is the development of HPV-mediated tumors.
4. Role and mechanism of HPV early-region proteins
4.1. E1 and E2 proteins
E1 protein has been reported highly conserved among the types of HPV and commonly decoded during the early expression of HPV infec-tion (Bergvall et al., 2013). E2 proteins are multifunctional proteins mostly associated with viral replication and transcription. Depending on the promoter, it either acts as an activator of transcription or as a repressor of the viral genome (Grm et al., 2005). E2 proteins consist of two functional domains; the N terminus conserved domain, required for
R. Bhattacharjee et al.
Critical Reviews in Oncology / Hematology 174 (2022) 103675
3
Table 1 Synopsis of the role of HPV Early proteins in CC and their therapeutic targets.
Early Protein
Pre- clinical
Pathways Cancer Hallmarks Mechanisms Therapeutic Potential References
E1 In vitro and in vivo
NF-κB NA Helps in initiation and regulation of HPV replication. Acts as a helicase and recruits replication factors.
ATP acts as an allosteric modulator of E1-E2 interactions. Indandione is a small class of molecules that act as a potential inhibitor of the E1-E2 interaction.
(Berg and Stenlund, 1997; Bergvall et al., 2013; Castro-Munoz et al., 2019; Chojnacki and Melendy, 2018; D’Abramo and Archambault, 2011; Goodwin and DiMaio, 2000; Moody and Laimins, 2010; White et al., 2001; White et al., 2003)
E2 In vitro and in vivo
NA NA Aids in initiation and regulation of HPV replication. Enable the binding of E1 protein. Disruption of E2 leads to CC progression. Negative transcription regulator of E6.
ATP acts as an allosteric modulator of E1-E2 interactions. Indandione is a small class of molecules that act as a potential inhibitor of the E1-E2 interaction.
(Chojnacki and Melendy, 2018; Cruz-Gregorio et al., 2018; Goodwin and DiMaio, 2000; Gupta and Mania-Pramanik, 2019; Moody and Laimins, 2010; Zahra et al., 2021) (Berg and Stenlund, 1997; D’Abramo and Archambault, 2011; White et al., 2001; White et al., 2003)
E4 In vitro and in vivo
NA NA Involved in viral release, transmission and PTM. Associates with keratin and could manipulate cytokeratin network, essential in the release of the virus.
Controlling the phosphorylation of E4 modulates its activity and is unable to affect the host.
(Doorbar, 2013; DOORBAR et al., 1996; Khan et al., 2011; McIntosh et al., 2010; Roberts et al., 2003; Rogel-Gaillard et al., 1993; Wang et al., 2009) (Kim et al., 2009)
E5 In vitro MAPK-ERK pathways
Angiogenesis, evading cell death, cellular proliferation
Major oncoprotein in HPV mediated carcinogenesis. Supportive role in E6 and E7 modulation. Suppresses p21 expression and initiates CC progression.
E5 inhibition by inhibiting the NF- kB pathway with inhibitors like narasin or flurosalan or celecoxib. Specific cytotoxic T cells are designed specifically for HPV E5 proteins with CpG- oligodeoxynucleotide and targets and degrade them so that their function is lost permanently.
(Crook et al., 1991; Ganguly, 2012; Jones and Münger, 1996; Kabsch and Alonso, 2002; Kim et al., 2006; Regan and Laimins, 2008; Stoppler et al., 1996) ( Dinneen and Ceresa, 2004) ( Farley et al., 2006; Sima et al., 2008; Wang et al., 2010)
E6 In vitro and in vivo
PI3K, AKT, Wnt, Notch pathways and EMT
Cellular proliferation, invasion metastasis, genomic instability, cellular immortality, cell cycle arrest, tumor- promoting inflammation, resisting cell death
Neoplastic effect on HPV- infected cells by promoting ubiquitin-dependent proteasome degradation of p53 and evades cell death. Targets degradation of apoptotic signaling cascade molecules through the ubiquitin-proteasome pathway leading to tumor development by inhibiting apoptosis. Causes dysregulated cell proliferation by downregulating transcriptional co-activator p300/CBP, essential for cell cycle. E6 is associated with the production of ROS that induces DNA breakdown and downregulates tumor suppressor genes. Downregulates IRF-3 of IFN-β and makes the immune response vulnerable against antigens of HPV. E6 inhibits the X-ray repair cross-complementing 1 is a scaffold DNA repair protein leading to increased mutation.
PDZ domains proteins including HDL protein, MUPP1, MAGI-1,2,3 are useful as therapeutic targets. Replacement of Lxx Ф Lsh leucine motifs with alanine results in E6 binding being terminated. RNA interference (RNAi) technology simultaneously knockdown both E6 and E7 expression.
(Filippova et al., 2002; Henderson et al., 2014; Kelley et al., 2005; Lee et al., 1997; Li et al., 1999; Nakagawa and Huibregtse, 2000; Nguyen et al., 2002; Patel et al., 1999; Sanchez-Perez et al., 1997; Shah et al., 2015; Thomas et al., 2002; Vieira et al., 2014; Warren et al., 2017) (Al Moustafa et al., 2004; Jung et al., 2013; Manzo-Merino et al., 2014) (Baleja et al., 2006; Be et al., 2001; Brehm et al., 1999; Brennan et al., 2009; DeMasi et al., 2005)
E7 In vitro PI3K/AKT, EMT
Replicative immortality, activating invasion and metastasis, evading growth suppressors, cell invasion, genomic instability, chronic inflammation
Promotes cervical dysplasia causing dysregulated cell cycle. Inactivates pRb and downregulates E2F for CC progression through conserved LXCXE motif of amino terminus 9. Causes deregulation of cyclin A/CDK2 and cyclin E/CDK2 by inactivation of p21 and p27 leading to increases in the
Various HDAC inhibitors inactivate E7. p600 induces anchorage-dependent integrin modulated cellular interaction and is an important target for targeted therapy. RNAi technology simultaneously knockdowns both E6 and E7 expression.
(Akagi et al., 2014; Brehm et al., 1999; Brennan et al., 2009; Cardeal et al., 2012; DeMasi et al., 2005; Dinneen and Ceresa, 2004; Duensing et al., 2001; Hasan et al., 2013; Hinds et al., 1992; Huh et al., 2005; Ivashkiv and Donlin, 2014; Kabsch and Alonso, 2002; Nguyen et al., 2002; Songock et al., 2017)
(continued on next page)
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
4
transactivation (TA) and DNA replication and the conserved C-terminal domain as a DNA binding domain that aids in the dimerization (Horner and DiMaio, 2007).
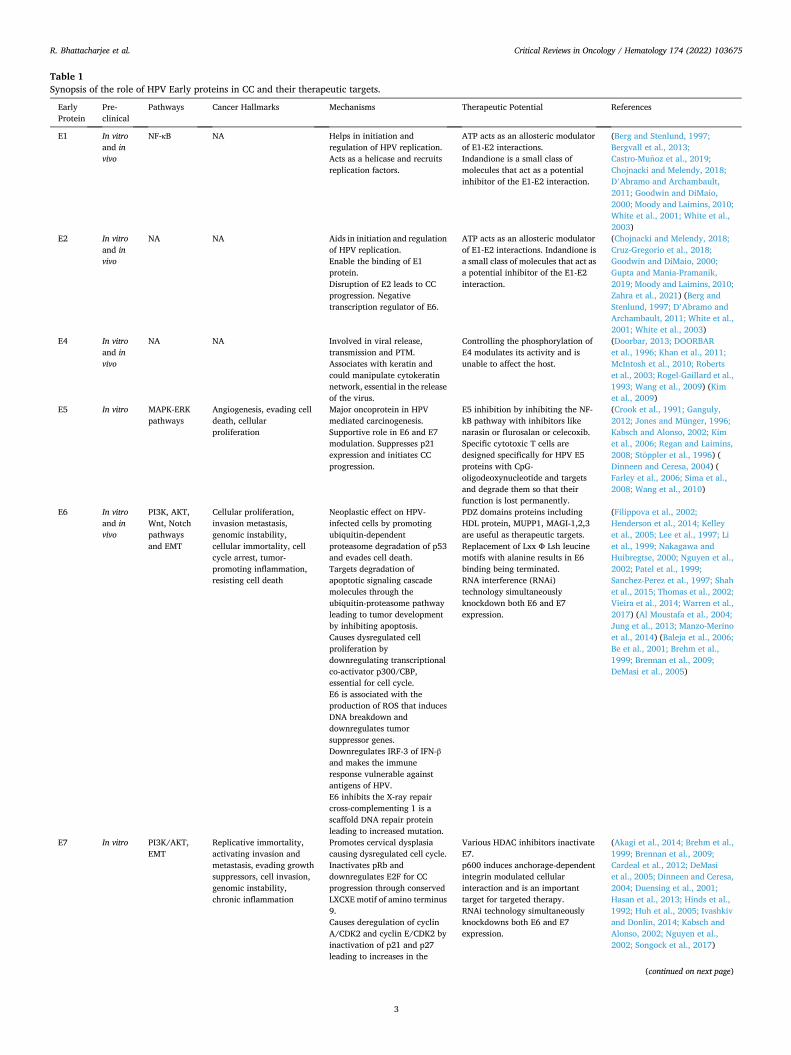
E1 and E2 proteins play crucial role in the initiation and regulation of HPV replication as illustrated in Fig. 1. Initially, E2 proteins bind to their binding sites (E2BS11 and E2BS12) at the origin of replication, which recruits a pair of E1 proteins to form a complex. Further, the E1 tetramer formed by displacing the E2 dimer at the E1 dimer, in an ATP-dependent manner. Thereafter, two E1 monomers bind to each half of the repli-cation origin and form an E1 double trimer complex. The subsequent binding of E1 monomers to the complex forms E1 double hexamer, which acts as a helicase to initiate the DNA unwinding for replication (Dinneen and Ceresa, 2004; Goodwin and DiMaio, 2000; Moody and Laimins, 2010). The E1 double hexamer complex starts unwinding the DNA in both directions and the replication process is initiated by recruiting the cellular replication factors like the single-stranded DNA binding protein RPA, topoisomerase I and DNA polymerase α-primase (Chojnacki and Melendy, 2018).
The functions of E2 proteins are disrupted by mutation or the inte-gration of DNA. The disruption of E2 blocks the viral replication that resulted in the aberrant viral gene expression, loss of control on E6 and E7 proteins, and ultimately leading to CC progression (Gupta and Mania-Pramanik, 2019). Further, E2 proteins of HPV18 have shown the association with the crest proteins of the mitochondria like ATP syn-thase, cytochrome (Cyt) c1, or Cyt C oxidase subunit II and also induce oxidative stress and the damage of the crest (Cruz-Gregorio et al., 2018). Oxidative stress appears as one of the major culprit in the pathogenesis of CC due to its role in the DNA damage that increase the chances of the incorporation of the HPV genome (Zahra et al., 2021).
E1 protein has been observed to regulate epigenetic modulation in the cells along with controlling replication machinery and interacting with other cellular components (Burley et al., 2020). Earlier studies have shown that HR-HPV infection increases the immune tolerance which makes a more susceptible microenvironment for further infection and disease progression (Castro-Munoz et al., 2019). A recent study, re-ported association of E1 proteins with immune response including TLR and interferon signaling (Castro-Munoz et al., 2019). It has also demonstrated the inhibitory role of E1 proteins on the antiviral immune responses that is reasoned to suppress the expression of IFNβ1 and IFNλ1 through the NF-κβ signaling pathway and intrfere in disease progression (Castro-Munoz et al., 2019).
4.2. E4 protein
Inside the E2 gene, in the region coding the flexible hinge domain of the E2 gene, is present the HPV E4 ORF. The HPV E4 protein is translated as an E1^E4 fusion protein by alternative splicing; as a result, the first few amino acids including initiation codon originate from the E1 (Wang et al., 2011). E4 associated with E1^E4 possesses a ‘leucine cluster’ motif in close proximity to the N-terminus of the protein, essential for the association with keratin. The C-terminus of the protein is involved in self-associations that produce amyloid fiber resembles structures and
helps in the manipulation of the cytokeratin network, essential for the viral release or transmission (McIntosh et al., 2010).
Commonly, E4 proteins have been expressed in the late stages of infection. In the virus replication cycle, the expression of the E4 protein is mostly confined to the middle and the upper layers of the epithelium. This protein appears as granules in the infected cells cytoplasm. These granules are considered responsible for the cytopathic effect wherein variations in morphology attributed to the type of HPV which infects the host (DOORBAR et al., 1996; Roberts et al., 2003; Rogel-Gaillard et al., 1993). The expression of E1^E4 occurs when the host cell initiates late viral promoter activation (Doorbar, 2013). Along with replication, E4 proteins also participate in post-translational modifications including phosphorylation. Phosphorylation of E4 occurs at the amino acid Thr 57 by S-phase associated p42 ERK and other MAP kinases which increase the stability of the protein and keratin binding capacity (McIntosh et al., 2010; Wang et al., 2009). After the viral genome amplification stage, calpain dissociates the E4 protein at the 17th and 18th amino acid, which leads to E4 mediated keratin disorganizations that help in a viral release (Khan et al., 2011).
4.3. E5 protein
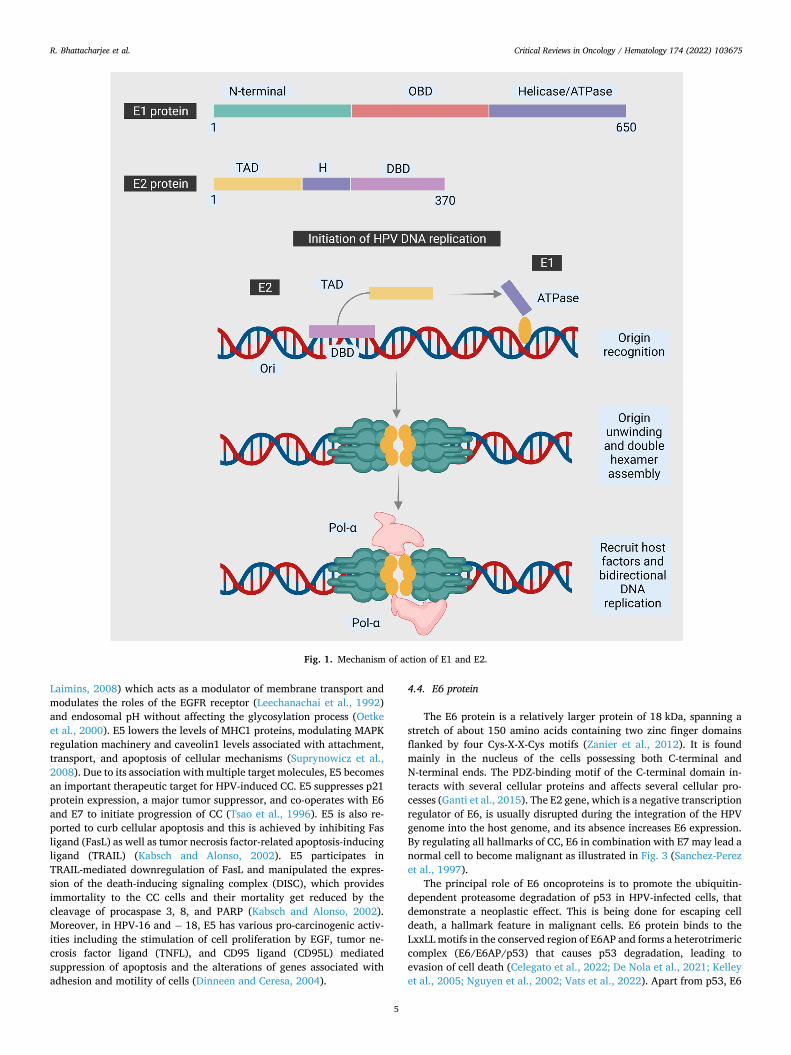
E5 is one of the major oncoproteins in HPV-mediated carcinogenesis, however, β-HPV does not encode for E5. It is a small size (10 kDa) 83 amino acids hydrophobic membrane-bound protein, associated with the endoplasmic reticulum and its gene located in the 3’ end of the early region of the viral genome expressed from a spliced mRNA (DiMaio and Petti, 2013). The protein contains three regions: the N-terminal domain, central domain, and C-terminal domain (DiMaio and Petti, 2013). The highly hydrophobic N-terminal is followed by the central and the C-terminal domain (Conrad et al., 1993; Moody and Laimins, 2010). E5 confers a supportive role in E6 and E7 modulation of both HR-HPV and LR-HPV and sometimes appears an oncogene as its lone presence may cause hyper-proliferation of the cells and cancer progression as illus-trated in Fig. 2 (Crook et al., 1991). E5 oncoprotein from HPV-16 and − 18 is present in the Golgi bodies or apparatus (GA), in the nuclear envelope and the endoplasmic reticulum (Disbrow et al., 2003), whereas it localizes entirely to the ER and with a lesser amount to the GA and early endosomes at physiological level of expression, as observed in primary human foreskin keratinocytes (Regan and Laimins, 2008).
E5 protein plays an important role in cell growth and regulation of signal transduction pathways in HPV infected cells (Regan and Laimins, 2008). It is commonly expressed in the early stages of HPV infection that is evident by the E5 protein-mediated increased immortalization of human keratocytes by E6 and E7, suggesting that E5 plays an important role in the initial stages of CC (Ganguly, 2012; Stoppler et al., 1996). Additionally, it is reported to induce angiogenesis during prolonged periods of HPV infection. Angiogenesis is a major player in cancer progression and E5 achieves this mediating ERK and PI3k-AKT pathways and EGFR signaling thus increasing VEGF, a major contributor to angiogenesis (Kim et al., 2006).
E5 binds to BAP31 (B cell receptor-associated protein 31) (Regan and
Table 1 (continued )
Early Protein
Pre- clinical
Pathways Cancer Hallmarks Mechanisms Therapeutic Potential References
spindle formation rate which brings uncontrollable cell cycle formation. Facilitates cell invasion by overexpression of MMP-9 leading to degradation of ECM. Binds to TLR9 leading to disrupted IFN-γ signaling and inhibiting the cyclic GMP-AMP- synthase causing inflammation.
R. Bhattacharjee et al.
Critical Reviews in Oncology / Hematology 174 (2022) 103675
5
Laimins, 2008) which acts as a modulator of membrane transport and modulates the roles of the EGFR receptor (Leechanachai et al., 1992) and endosomal pH without affecting the glycosylation process (Oetke et al., 2000). E5 lowers the levels of MHC1 proteins, modulating MAPK regulation machinery and caveolin1 levels associated with attachment, transport, and apoptosis of cellular mechanisms (Suprynowicz et al., 2008). Due to its association with multiple target molecules, E5 becomes an important therapeutic target for HPV-induced CC. E5 suppresses p21 protein expression, a major tumor suppressor, and co-operates with E6 and E7 to initiate progression of CC (Tsao et al., 1996). E5 is also re-ported to curb cellular apoptosis and this is achieved by inhibiting Fas ligand (FasL) as well as tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) (Kabsch and Alonso, 2002). E5 participates in TRAIL-mediated downregulation of FasL and manipulated the expres-sion of the death-inducing signaling complex (DISC), which provides immortality to the CC cells and their mortality get reduced by the cleavage of procaspase 3, 8, and PARP (Kabsch and Alonso, 2002). Moreover, in HPV-16 and − 18, E5 has various pro-carcinogenic activ-ities including the stimulation of cell proliferation by EGF, tumor ne-crosis factor ligand (TNFL), and CD95 ligand (CD95L) mediated suppression of apoptosis and the alterations of genes associated with adhesion and motility of cells (Dinneen and Ceresa, 2004).
4.4. E6 protein
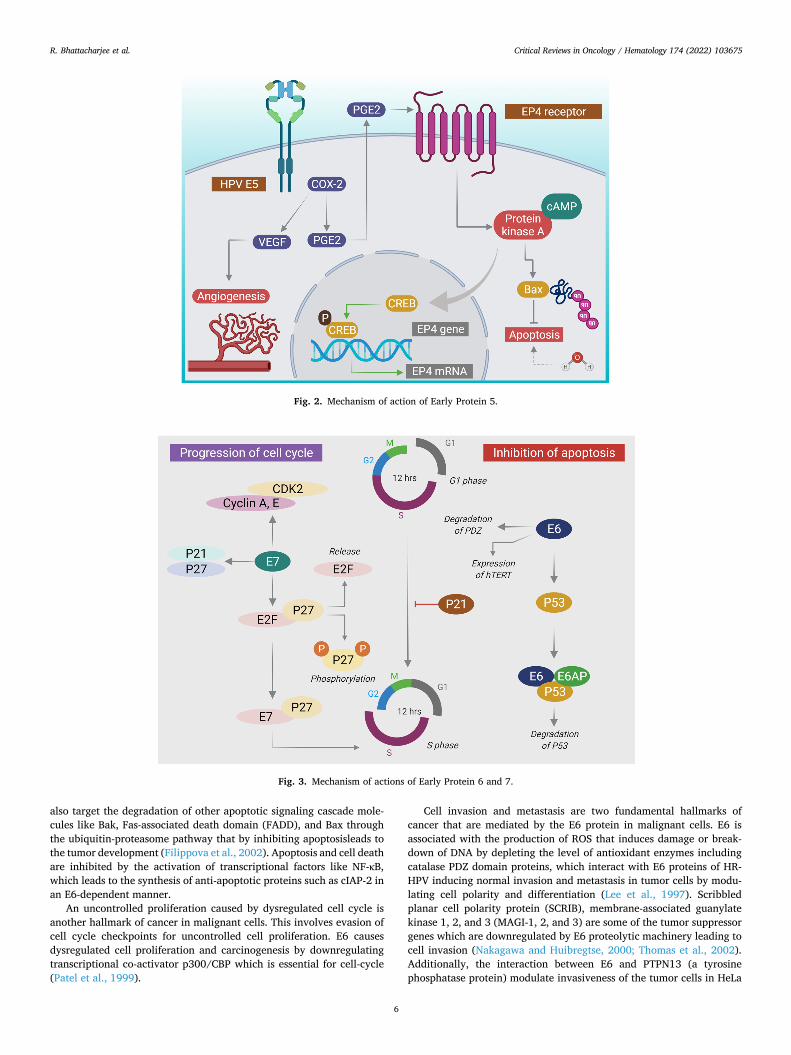
The E6 protein is a relatively larger protein of 18 kDa, spanning a stretch of about 150 amino acids containing two zinc finger domains flanked by four Cys-X-X-Cys motifs (Zanier et al., 2012). It is found mainly in the nucleus of the cells possessing both C-terminal and N-terminal ends. The PDZ-binding motif of the C-terminal domain in-teracts with several cellular proteins and affects several cellular pro-cesses (Ganti et al., 2015). The E2 gene, which is a negative transcription regulator of E6, is usually disrupted during the integration of the HPV genome into the host genome, and its absence increases E6 expression. By regulating all hallmarks of CC, E6 in combination with E7 may lead a normal cell to become malignant as illustrated in Fig. 3 (Sanchez-Perez et al., 1997).
The principal role of E6 oncoproteins is to promote the ubiquitin- dependent proteasome degradation of p53 in HPV-infected cells, that demonstrate a neoplastic effect. This is being done for escaping cell death, a hallmark feature in malignant cells. E6 protein binds to the LxxLL motifs in the conserved region of E6AP and forms a heterotrimeric complex (E6/E6AP/p53) that causes p53 degradation, leading to evasion of cell death (Celegato et al., 2022; De Nola et al., 2021; Kelley et al., 2005; Nguyen et al., 2002; Vats et al., 2022). Apart from p53, E6
Fig. 1. Mechanism of action of E1 and E2.
R. Bhattacharjee et al.
Critical Reviews in Oncology / Hematology 174 (2022) 103675
6
also target the degradation of other apoptotic signaling cascade mole-cules like Bak, Fas-associated death domain (FADD), and Bax through the ubiquitin-proteasome pathway that by inhibiting apoptosisleads to the tumor development (Filippova et al., 2002). Apoptosis and cell death are inhibited by the activation of transcriptional factors like NF-κB, which leads to the synthesis of anti-apoptotic proteins such as cIAP-2 in an E6-dependent manner.
An uncontrolled proliferation caused by dysregulated cell cycle is another hallmark of cancer in malignant cells. This involves evasion of cell cycle checkpoints for uncontrolled cell proliferation. E6 causes dysregulated cell proliferation and carcinogenesis by downregulating transcriptional co-activator p300/CBP which is essential for cell-cycle (Patel et al., 1999).
Cell invasion and metastasis are two fundamental hallmarks of cancer that are mediated by the E6 protein in malignant cells. E6 is associated with the production of ROS that induces damage or break-down of DNA by depleting the level of antioxidant enzymes including catalase PDZ domain proteins, which interact with E6 proteins of HR- HPV inducing normal invasion and metastasis in tumor cells by modu-lating cell polarity and differentiation (Lee et al., 1997). Scribbled planar cell polarity protein (SCRIB), membrane-associated guanylate kinase 1, 2, and 3 (MAGI-1, 2, and 3) are some of the tumor suppressor genes which are downregulated by E6 proteolytic machinery leading to cell invasion (Nakagawa and Huibregtse, 2000; Thomas et al., 2002). Additionally, the interaction between E6 and PTPN13 (a tyrosine phosphatase protein) modulate invasiveness of the tumor cells in HeLa
Fig. 2. Mechanism of action of Early Protein 5.
Fig. 3. Mechanism of actions of Early Protein 6 and 7.
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
7
cells (Kranjec et al., 2016; Lee et al., 2022). Moreover, downregulation of ECM accompanied with inhibition of cell adhesion protein Fibulin 1 and miR-23b in an E6 dependent manner result in tumor cell migration (Vats et al., 2021). E6 promotes metastasis and invasion in tumor cells by downregulating paxillin resulting in compromised actin fibers and consequently, the integrity of the epithelium (Wu et al., 2014).
Avoiding the host immune system is another hallmark in malignant cells, which demonstrated the inhibitory role of E1 proteins on the antiviral immune responses may take placele to that for prolonged replication of HPV to remain undetected inside the host epithelium for initiation of the immune evasion mechanism. It is facilitated by down-regulation of the Interferon Regulatory Factor-3 (IRF-3) of the IFN-β via E6 protein, thus making the immune responses vulnerable against an-tigens of HPV (Li et al., 1999). Furthermore, the E6 oncoprotein sup-presses the immunological response induced by Interferon α (IFN-α) by inhibiting phosphorylation of Tyrosine Kinase 2 (TYK2) in a STAT/TYK2 dependent manner, preventing IFNα and its receptor from interacting, resulting in prolonged HPV replication (Li et al., 1999). The therapeutic approach for the management of CC, mainly immunotherapy, is likely affected by the translational consequences of tumor stroma and a pro-fusion of tumor-sensitive lymphocytes in tumor microenvironment. However, the diagnosis of CC is contrarywise associated with micro-vessel density, allowing inhibition of angiogenesis with therapeutic agents like bevacizumab, which play an impotant role in targeting advanced as well as recurrent CC (De Nola et al., 2021).
Immortalization of the cell is one of the hallmarks that are facilitated by HPV in CC via E6 protein. The unbalanced expression of E6 causes expression of hTERT resulting in immortalization of the cell via degra-dation of NFX1–91 (Nuclear transcriptional factor X box binding 1–91) (Shah et al., 2015). Moreover, the overexpression of proto-oncogenic protein c-Myc and transcriptional factor Sp1 in an E6 dependent
manner leads to the recruitment of promoter that modulates epigenetic changes in hTERT along with phosphorylated DNA polymerase II ulti-mately leading to immortalization of cells altogether (Gewin et al., 2004; Zhang et al., 2017).
Genomic instability, a prominent hallmark of HPV is incorporated into the malignant cell by E6 protein by upregulating apolipoprotein B, an mRNA editing enzyme catalytic polypeptide 3 that caused replacing cytosine residue with uracil (Warren et al., 2017). This protein is prone to errors, and its overexpression leads to a greater mutation rate, resulting in genomic instability. E6 inhibits the scaffold DNA repair protein X-ray repair cross-complementing 1, causing an increase in mutation and thus increased genomic instability (Henderson et al., 2014; Vieira et al., 2014; Warren et al., 2017).
4.5. E7 protein
The E7 protein is smaller relative to E6 spanning across a length of around 100 amino acids, consisting of 3 conserved regions 1/2/3 (CR1/ 2/3). A minor portion of the CR1 region and more than a major portion of the CR2 region resembles the adenovirus in sequence and a large T antigen of SV40 (Pal and Kundu, 2020a; Phelps et al., 1988). The CR3 region is located at the carboxyl-terminal end of the protein. This area is conserved, and believed to encode a zinc finger domain bordered by two CXXC motifs separated by 29 amino acid residues (McINTYRE et al., 1993). Upon interaction with various host factors, the E7 protein is associated with various hallmarks in CC as illustrated in Fig. 5.
In HPV-induced CC, E7 oncoprotein plays a crucial role in promoting cervical dysplasia causing dysregulated cell cycle as illustrated in Fig. 4 (Moody and Laimins, 2010). The main targets of E7 are the inactivation of the pRb and the downregulation of E2F. It is well reported that E7 degrades the pRB protein, which is known to halt cell cycle by
Fig. 4. Events mediated and modulated by early protein 6 and 7 in CC.
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
8
suppressing the E2F factor induced unlimited cellular division and cancer progression. E7 induces proteasome degradation by ubiquitin-mediated pathways and low levels of E7 can also cause high damage leading to cancer progression through the conserved LXCXE motif of amino terminus 9 (Hinds et al., 1992). There are eight members in the Rb family and the majority of the proteins act like activators; (Chellappan et al., 1992; Liu et al., 2006), thus, E2F members can also be targeted for therapy with upregulation E2F repression. Whereas, p107 and p130 which are the pockets and the regulator of E2F are also degraded by E7 (Barrow-Laing et al., 2010). E7 causes deregulation of cyclin A/CDK2 and cyclin E/Cdk2 by inactivation of p21 and p27 leading to increases in the spindle formation rate which causes uncon-trollable cell cycle formation. Phosphorylation of these CDKs can cause inhibition of tissue transformation and morphogenesis (Huh et al., 2005). E2F6 is a transcriptional repressor of S phase-inducing genes which are inhibited by E7, leading to uncontrolled cell proliferation along the dysregulated cell cycle. Claspin belongs to a group of calcium-activated cysteine proteases and play role in a variety of cellular processes (Zhang et al., 2006). Overexpression of the cell cycle protein claspin in an E7 dependent manner leads to the facilitation of cell pro-liferation even during DNA damage.
Cell invasion is another hallmark modulated by E7 protein in an MMP-9 dependent manner. E7 promotes cell invasion by overexpression of MMP-9 leading to degradation of ECM and cell invasion (Cardeal et al., 2012). Genomic instability, another hallmark of CC is promoted by E7 through overexpression of centrosomes leading to aneuploidy. The dysregulated pRb and causes aberration at the centrosome through CDK2 resulting in degradation of the mitotic spindle and g-tubulin (Akagi et al., 2014; Duensing et al., 2001; Nguyen et al., 2002). Chronic inflammation is another hallmark modulated by E7 protein following binding to TLR9 that results in disrupted IFN-γ signaling and inhibition of the cyclic GMP-AMP-synthase (Hasan et al., 2013; Ivashkiv and
Donlin, 2014; Songock et al., 2017). The interplay of E6 and E7 triggers inflammation process in a pro-inflammatory cytokine-dependent manner by IL-6 and IL-18 leading to tumor progression by chemokines, and metalloprotease (Hammes et al., 2007; Ren et al., 2013; Richards et al., 2014).
A large cross-talk occurs between E6 and E7 oncoprotein during cervical carcinoma progression as illustrated in Fig. 4. This cross-talk between oncoprotein E6 and E7 is crucial for HPV-induced carcinoma and these oncoproteins largely control various cell processes. The angiogenic switch get modulated in malignant cells in an E6 and E7 interplay-dependent manner. This leads to the downregulation of mas-pin and thrombospondin-1 which results in simultaneous over-expression of IL-8 and VEGF (Toussaint-Smith et al., 2004). Tumor invasion and metastasis are triggered by the interplay between E6 and E7 protein as it induces downregulation of TIM-2 and RECK proteins which are responsible for MMP activity (Cardeal et al., 2012). The interplay of E6 and E7 is also involved in the dysregulation of cellular energetics via upregulation of GLUT1 and pyruvate kinase (Zwerschke et al., 1999).
5. HPV signaling pathway and targets
HPV E5, E6, and E7 oncoproteins can regulate and alter several cell- signaling pathways to cause cancer. These molecular pathways play major roles in malignant transformation, thus targeted therapy against these key signaling pathways has potential therapeutic implications. E5 oncoprotein stimulates the MAPK-ERK pathway (Branca et al., 2004). E6 oncoprotein alters phosphatidylinositol 3-kinase and protein kinase B (PI3K/AKT) pathway, Wnt pathway, Notch pathway, and E7 regulates the PI3K/AKT pathway. PI3K/AKT is particularly important due to its relation to initiation of malignancy, cell proliferation, metastasis, and also drug resistance (Manzo-Merino et al., 2014). Moreover, these
Fig. 5. Role of early roteins and its characteristics as cancer hallmarks.
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
9
oncoproteins also alter the NF-κB pathway, YY1 pathway (Guifen et al., 2011), and apoptotic pathways (intrinsic and extrinsic). Herein, we have discussed studies which suggest targeting the altered cell signaling pathways and epidermal growth factor receptor along with vascular endothelial growth factor as potential therapeutic approache against CC and illustrated in Fig. 6 and Fig. 7.
Activation of the PI3K/AKT signaling is the primary pathway implicated in cancer cell’s longevity involving the transduction of sig-nals by phosphorylation of AKT (Gupta et al., 2018). HPV-induced infection followed by the expression of either E6 or E7 activates the signaling pathway of PI3K/AKT by altering various cellular mecha-nisms, thus leads to carcinogenesis (Rodon et al., 2013). Earlier in-vestigations have well demonstrated that HPV 16/18 E6 and E7 proteins activate this pathway and thus in turn AKT activity results in the inac-tivation of pRB which play a role in causation of CC. p27 has been re-ported to be negatively regulated by the E7 protein of HPV 16 using AKT and increased retention of p27 in the cytoplasmic domains (Charette and McCance, 2007; Menges et al., 2006).
Cervical carcinomas and CIN lesions may be used to test ERK1 signaling in predicting CC incidence and it is suggested that the expression of ERK1 tends to be a early predictor of CC (Branca et al., 2004). The recently discovered proto-oncogene tyrosine-protein kinase SRC protein, together with phosphorylated ERK-1/2 and c-Fos, aided in the modulation of ERK signal transduction pathway expression to facilitate cell proliferation and prevent apoptosis in CC cells (Song et al., 2017). It has also been shown that ERK1/2 involves the expression of HIF-1α, VEGF, and IL-8 induced by HPV16 E6 oncoprotein (Gupta et al., 2018; Liu et al., 2016).
HPV16 oncoprotein E6 activates the Wnt / β-catenin pathway, vital for the transformation of HPV, epithelial cells immortalization, balancing tissue homeostasis and pathologic phenotype (Gupta et al., 2018; Moon et al., 2004). The altered Wnt/ β-catenin signaling pathway is believed to play role in the onset and maintenance of HPV-associated malignancies, and could be utilized as a therapeutic target for CC (Ramachandran et al., 2012).
The Yin Yang complex1 protein binds to the URR of HPV 18, thus controlling transcription of HPV E6 and E7 protein (Bauknecht and Shi, 1998). It also behaves as a negative regulator of p53 by interacting with mdm2 (Park and Atchison, 1991). It has recently been reported that YY1 has been found over expressed in CC and thus may play an important role in the progression of HPV infection associated CC.
There are reports which suggest that the activity of HER3 over-expression results in the poor prognosis of CC (Ojesina et al., 2014). Studies of full-exome sequencing suggest that activated HER2 is
associated with somatic mutation and incorporation of HPV into CC (Chavez-Blanco et al., 2004; Oh et al., 2015). HPV E6/E7 controls the expression of HER3 proteins and downstream PI3K signaling, that is suggestive of positive association between HER3 expression and HPV-induced CC (Brand et al., 2017). The knowledge of viral oncogene-driven methods reveal new molecular targets for tumors following infection with HPV.
Some reports showed that a possible binding site for STAT3 has been mapped to a 5 ’region of HPV16 LCR that regulates the expression of viral oncogenes, which may play a key role in epithelial cancer (Arany et al., 2002). Therefore, more research are needed to decipher the role of STAT3 in CC . Although some studies have shown that STAT3 is present in stem cells, a subclass of CC lesion, this additionally supports that STAT3 may play a key role in viral gene expression and HPV16-mediated carcinogenesis (Manzo-Merino et al., 2014; Martinez-Zapien et al., 2016; Menges et al., 2006).
The most important factor in controlling the EMT signaling pathway is the transformation of the growth factor-beta (TGF-β1). HPV16-E6/E7 prompts EMT-induced by FGF-2 and FGF-4 to increase invasive cancer capacity (Cheng et al., 2012). Simultaneous expression of E6/E7 and ErbB-2 reduces the E-cadherin function enhancing the expression of transcription factors including SLUG, SNAIL, and TWIST which activate the EMT pathway and consequently leads to the overexpression of cyclin D1 and c-Myc genes and expression of SFRP1/2 (Al Moustafa et al., 2004; Jung et al., 2013).
Protein 1 (AP-1) is a transcription factor activator that plays a critical role in the progression of HPV-associated CC. The availability of AP-1, which binds to the HPV URR, is required for the production of two key viral oncogenes, E6 and E7 (Rosl et al., 1997). If there is an occur-rence of mutation in the binding AP-1 sites inside URR, the expression E6 / E7 is completely abolished. Modified AP-1 complex with much more binding c-Fos and c-Jun and absence of Fra-1 expression in CC cell line HeLa is associated with the tumorigenic phenotype. The over-expression of c-fos acts as the catalyst for CC induction by ectopic expression of c-Fos in HeLa fibroblast hybrid tumorigenic. The upregu-lation of transcription factor c-fos resulted in negligible or very little expression of TCF transcription factor in CC cell line whereas its absence results in dysregulation of activity of Notch 1 in CC cells (Talora et al., 2002). Furthermore, AP-1 is a key regulator of the expression of HPV oncogenes E6 and E7, as well contribute in the development of resis-tance to chemotherapy and radiotherapy (Dhandapani et al., 2007). In the absence of HPV, functional heterodimerization of c-Fos and Fra-2 in the presence of c-Jun has been demonstrated to promote aggressive tumor phenotype with poor prognosis.
Fig. 6. Role of early protein (E6, E7) in regulating cellular signaling for CC progression.
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
10
The notch signaling pathway is responsible for the up-regulation of leukemia and other forms of cancer, but in HR-HPV it plays a different role and surprisingly suppresses the HPV-induced carcinoma. The downregulation of Notch signaling, which is a potent suppressor through oncoprotein E6 aggregates the tumorigenesis for CC progression (Zagouras et al., 1995). p53 regulation is an important factor for notch signaling and it maintains a reciprocal relationship. Upregulation of the notch is detrimental for HPV-induced carcinoma as it destroys p53 and possess potential to suppress notch signaling supports in limiting its progression (Talora et al., 2005). The notch pathway shows anti-tumorigenic activities with p53, it helps in cellular progression with the P13k-AKT pathway, thus aiding cervical tumorigenesis (Rangarajan et al., 2001; Veeraraghavalu et al., 2005). The notch is an important target for HR-HPV regulation since it is a key player in HPV-induced carcinoma and has a dual role in cervical tumorigenesis. Though, further comprehensive research are yet needed to establish the role of notch signaling in CC.
6. Therapeutic potentials of HPV early protein
E1 is the only oncoprotein with enzymatic activity like DNA helicase and ATPase activity, which supports the viral genome replication by opening up the replication fork before replication. The role of ATP in the above process focuses on assisting this opening up of the replication fork (Pappa et al., 2018). Also, it helps E1 to oligomerize into hexamers, which is essential for the interplay of E1 and E2 proteins. It play a sig-nificant role in the allosteric modulation of E1-E2 interactions. The E1 protein’s location where these activities occur is focused in the Cter-minal region which binds to the E2 protein (Berg and Stenlund, 1997; White et al., 2001). Thus, interfering with the helicase activity and ATP binding activity may help in developing novel antivial agents for HPV-induced CC. Following a thorough screening of around 500,000 molecules, hits of biphenylsulfonacetic acid analogs which had a biphenyl group substituted with a sulfonyl acetic acid entity were found as the lead molecules (White et al., 2005). The functionality of the stated molecules is dependent on the concentration of ATP as they affect ATP binding through allosteric mechanisms. The presence of tyrosine as a
replacement of Cys-486 in HPV 11 E1 enhanced the sensitivity to in-hibitors. The suggest that E1 ATPase domain may serve as a target for small molecules to impede HPV DNA replication and found promising in development of future therapeutic strategy (White et al., 2005).
While it is difficult to identify protein-protein interaction inhibitors in the form of small molecules, for HPV one such potential inhibitor of the E1-E2 interaction has been established. The congregation of the complex depends on the E1-E2 interaction with DNA and its therapeutic potential. In the E1-E2-ori ternary alongside a critical protein’s threshold level, the interplay is established between E2’s TAD and E1’s C terminal ATPase domain (White et al., 2003). Indandione is a small class of molecules that act as a potential inhibitor of the E1-E2 interaction. The basic mechanism how the inhibitors achieve their role is by bringing out significant changes in the movement of amino acids at the binding site (Talora et al., 2005; Zagouras et al., 1995). The amino acids whose residues mostly undergo a conformational change include Tyr-19, His-32, Leu-94, and Glu-100, ultimately resulting in the creation of a deep hydrophobic pocket to which the inhibitor’s indandione entity would firmly bind to. Also, the mutation of Glu-100 with alanine has shown increased sensitivity of the E2 protein to the indandione mole-cules (Talora et al., 2005; Zagouras et al., 1995). The potential of these indandione compounds in restricting the replication of the HPV genome in vivo will pave a path for utilizing the capacity of E2 protein for further consideration as a novel therapeutic target as well as a target for small molecules targeting the interaction of HPV with proteins. Further, in E2 TAD, the presence and characterization of inhibitor binding pockets showed potential in recognition of a novel target for therapy towards enhancing HPV infection treatment (D’Abramo and Archambault, 2011).
E4 protein also play role in the cell cycle arrest as it evident from the observation that the epithelial cells which showed an expression of E4 from HPV 16 and HPV 18 witnessed mitotic arrest at the G2 phase owing to the restriction of nuclear accumulation of cdk1 protein in the nucleus. Thus, it can be concluded that when phosphorylation of the E4 is controlled, its activity can be modulated and it is unable to affect the host. The vailable data is suggestive of that the epigenetic modulation of the E4 protein may serves as a novel therapeutic target for future drug
Fig. 7. Influence of early protein (E1) on cellular signaling for CC progression.
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
11
discovery in therapeutics (Knight et al., 2006). It has been speculated that HPV E5 oncoprotein induces cyclo-
oxygenases − 2(COX-2) with the help of the EGFR signaling pathway along with NF-kB and activator protein-1(AP-1) (Kim et al., 2009). E5 inhibition carried out by inhibiting the NF-kB pathway with inhibitors like narasin or flurosalan, mediating inhibition of AP-1 was much more effective in controlling the E5 oncoprotein function. Inhibitors that bring effect directly have shown toxic at higher levels and at lower levels they are non-functional (Kim et al., 2013). However, researches have suggested new inhibitor compounds like celecoxib (Farley et al., 2006). With this kind of inhibitor, E5 oncoprotein mediated HPV-induced carcinoma can be targeted following subsequent lowering of E5 levels. Thus, targeting E5 oncoprotein or its relevant pathways may inhibit HPV-induced carcinoma in cervical cells. Wang et al. reported that re-combinant oncolytic adenovirus M6 with an antisense E6-E7 gene selectively replicated in cancer cells, inhibiting tumor development and inducing apoptosis in HPV16-positive CC cells without causing cross-resistance or other problems (Wang et al., 2010). Other remedies include therapies like specific cytotoxic T cells which are designed specifically for HPV E5 proteins with CpG-oligodeoxynucleotide and may target and degrade them so that their function is lost permanently and leads loss of function of the viral E5 (Chen et al., 2004). RNA interference (RNAi) technology is another therapeutic strategy that target the HPV16-E7 region and help in the modulation of E6 and E7 proteins and knockdown both E6 and E7 expression simultaneously (Sima et al., 2008). This approach along with radioimmunotherapy have shown visible effects and showed potential of suppressing HPV- induced carcinoma along with a significant rise in p53 and Rb proteins (Sima et al., 2008; Wang et al., 2010).
HPV E6 oncoproteins are shown associated with p53 and degrada-tion of p53 which is dependent upon the activity of the E6/E6AP com-plex. The inhibition of this complex can be a promising therapeutic approach to reactivate the function of p53 tumor suppressor protein. PDZ domains proteins including HDL (human homolog of Drosophila discs large tumor suppressor protein, MUPP1 (tight junction) (Brennan et al., 2009), and MAGI-1,2,3 membrane-associated guanylate kinase (MAGUK) proteins (Thomas et al., 2002) are considered useful as novel therapeutic targets. It is associated with E6 and maintains cellular po-larity, adherence, and disrupting the cause of neoplasm formation. Among numerous approaches, one of them is to target the sequence alignment of the E6 binding motif Lxx Ф Lsh where L is leucine, s the small amino acid (glycine or alanine). xx denotes a dipeptide where one of the residues is Glu, Asp, Gln or Asn Ф is the hydrophobic residue (commonly leucine) and h usually glutamine, glutamate, asparagine or aspartate (Baleja et al., 2006). The studies of mutagenesis have shown that the replacement of each of these leucine motifs with alanine may result in E6 binding being terminated (Baleja et al., 2006; Be et al., 2001).
Several studies have shown that E7 oncoprotein can influence the E2F factor-induced transcription by interacting with various cellular proteins as well as class I histone deacetylases as illustrated in figure 14 (Brehm et al., 1999; DeMasi et al., 2005). Various HDAC inhibitors are being investigated in clinical trials to deactivate E7 as the upregulation of HDAC has a significant role in cancer progression (Baleja et al., 2006). In the reverse genetic experiments, it has been shown that mutations in E7-HDAC binding sequences in the HPV 31 genome can be diminished to facilitatr the episomal maintenance and also regulate the proliferation of the primary keratinocytes (Longworth and Laimins, 2004). Taken together, the results of the interaction between E7 and HDAC inhibitors can be considered as a promising antiviral target and can be used for future remedies. Another protein, p600, (600 kDa) induces anchorage-dependent integrin modulated cellular interaction and be-comes an important target both for therapy as well as for the E7 onco-protein mediated action (Akhatova et al., 2021; Huh et al., 2005).
7. Therapeutic strategies for targeting E proteins
E proteins are largely responsible for instigating the process of HPV- mediated CC, however, amongst all forms of E proteins, E6 and E7 are the most crucial oncoproteins. This is suggestive of that targeting E6 and E7 oncoproteins may be important in significant eradication of all CC cells by inhibiting these cancer hallmarks. Currently, the focus is on developing various therapeutic strategies for targeting E proteins as a treatment approach in HPV-mediated CC.
Nucleic acid-based therapeutic strategies depend on the usage of antisense RNA or DNA moieties to intrude the appearance and/or ac-tivities of E6 and E7 oncogenes, which can efficiently avoid the pro-gression of CC. Studies have shown that the hammerhead ribozymes significantly targeted the HPV16 E6/E7 transcripts and causes potent inhibition of the cell growth following apoptosis in vitro and also tumor growth in vivo in nude mice models (Zheng et al., 2002, 2004). In another study, it has been reported that a ribozyme targeting E6AP in the HeLa cells enhanced the process of apoptosis and mitomycin-C triggered DNA impairment (Kim et al., 2003). Reschner and co-workers developed antisense oligonucleotide conjugated with a ruthenium (photoreactive) complex that targeted E6 protein in HPV16-infected SiHa cells and significantly inhibited cell growth in monolayers as well as three-dimensional specimens (Reschner et al., 2013). Hu et al. first reported TALEN (transcription activator-like effector nucleases)-based genome editing of the E6/E7 oncogenes and demonstrated that it leads to the potent inhibition of the tumor growth, induce apoptosis, and decrease tumorigenic competence of HPV-infected cells, including HPV16 (SiHa) and HPV18 (HeLa) (Pal and Kundu, 2020b).
Conventionally, preventive vaccines exhibited a potential role in activating the immunity of the body to avoid the initiation and pro-gression of CC. However, recent clinical studies are directing the development of targeted therapeutic vaccines that can be useful and efficient in treating CC, even at progressive malignant phases. Certain preventive vaccines which exhibit significant effects against L1 or L2 capsid proteins, get deactivated or removed during the combination of the HPV genome within the cellular genome. On the other hand, the therapeutic vaccines efficiently target the E6 and E7 oncoproteins, and get expressed for a prolonged time in HPV’s life cycle, therefore. exhibit significant hallmarks of HPV-mediated CC (Chabeda et al., 2018).
Numerous therapeutic vaccines are being investigated and are presently under different stages of clinical trial (phase I/II/III) for CC. Few vaccines, such as ProCervix and PDS0101, have been developed specifically for persistent HPV-mediated infections and inferior grade squamous intraepithelial lesions (SIL). In addition, few vaccines such as pNGVL4a-CRT/E7 (detox), GX-188E, PepCan + Candin have been developed for targeting cervical intraepithelial lesions or higher grade SILs. Moreover, few highly modified vaccines such as TA-CIN + GPI- 0100, INO-3112, ADXS11-OO1, HPV16-SLP, and others have been developed for targeting HPV16/18-E6/E7 oncoproteins and treating CC of advanced stages (Yang et al., 2016). Akhatova et al. (2021) assessed and compared the prevailing DNA therapeutic vaccines under clinical trials (phase I/II), that expressed HPV E6 and E7 proteins for the treatment of CC relying on the viral clearance, clinical efficiency, immunogenicity, and adverse effects. It has been observed that 5 different DNA therapeutic vaccines (VGX-3100, GX-188E, pNGVL4a--Sig/E7(detox)/HSP70, pNGVL4a-CRT/E7(detox), and MEDI0457) pos-sesses potential to be efficient against CC and exhibit high endurance against CC (Akhatova et al., 2021).
The advancements in the genome editing tools have assisted the researchers in specifically targeting the HPV-E6/E7 regions of the HPV genome or associated mRNAs, that results in improved CC therapy. In recent studies, various approaches have been shown for competently silencing the E6/E7 expressions, including zinc finger nucleases, TAL-ENs, and the clustered regularly interspaced short palindromic repeat- associated nuclease (CRISPR/Cas9) RNA-guided endonucleases. These
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
12
approaches may aid in reducing the mortality rate associated with CC, particularly in the advanced stages of CC (Pal and Kundu, 2020b). One of the most important genome-editing tool, CRISPR-Cas9, have been initially detected as a part of adaptive immunity in the microbial systems to defend against extraneous nucleic acids (Makarova et al., 2015; Tenaillon et al., 2016; Wright et al., 2016). As per the reported studies, the CRISPR-Cas9 mediated E6 or E7 gene silencing is found highly applicable in inhibiting disease progression in CC. Moreover, the guide RNAs can be efficiently programmed following the specificity of the targeting exon segments of the E6 and E7 oncogenes. CRISPR-Cas9 facilitated silencing of E6/E7 oncogenes in HPV16-infected CasKi or SiHa carcinogenic cells and HPV18-infected HeLa cells exhibited reduced growth, arrest of cell cycle and cell death mediating regenera-tion of p53 and pRb (Hsu et al., 2014; Kennedy et al., 2014). A recent study showed that the gene knockout chain reaction (GKCR) Cas9/guide RNA (gRNA) cassettes can significantly integrate within the targeted loci through homology-independent targeted insertion (HITI). The study demonstrated that the GKCR method led to a higher proportion of in-sertions or deletions (indels) in the E6 and E7 oncogenes of HPV18. Furthermore, the targeting of HPV18 E6/E7 oncogenes using GKCR method resulted in upregulation of P53/RB proteins and inhibition of the propagation of HeLa cells. Thus, this technique may be promising in the treatment of HPV infections and associated CC (Tian, Rui et al., 2022).
An ideal tumor/carcinogenic progressive microenvironment neces-sitates deactivation of the immune system of the body, which comprises of the low amount of T cells and their repressed actions, alterations in the cytokine expressions and damaged antigen performance (Mandal et al., 2016; Sun et al., 2014; Varilla et al., 2013). E6/E7-specific hu-moral and cellular immunity are clinically effective leading to the incessant activities of these oncogenes during the disease development and progression. In one of the study, synthesized HPV-16/18 E6 and E7 DNA sequences were familiarized to trigger the immune system re-sponses employing a novel plasmid VGX-3100 (Bagarazzi et al., 2012; Trimble et al., 2015). Jin and co-workers showed that the T cells can be genetically modified and engineered for targeting the HPV16 E7, lead-ing to the deterioration of HPV16-infected CC in mouse xenograft models (Jin et al., 2018). In another study, it has been demonstrated that the metastatic CC undergo complete deterioration in response to a solo permeation of T cells targeting HPV16 E6/E7 (Stevanovic et al., 2015).
Galvao and Coimbra discussed the role and significance of long non- coding RNAs (lncRNAs) in various types of cancer, including CC. They also summarized the facts associated with lncRNAs and recognized overall outline of the transcripts according to their role which they exhibit in CC (de Carvalho Galvao and Coimbra, 2020). Recently, it has been shown that non-coding RNAs (ncRNA), including LncRNAs, microRNAs, piRNAs (PIWI-interacting RNAs), and circular RNAs play significant role in monitoring various cellular function in the gyneco-logical cancers, including CC. Furthermore, numerous clinical trials are ongoing to identify the biomarkers and potential therapeutic effects of ncRNAs (Razavi, Zahra Sadat et al., 2021).
Celegato et al. developed a small moiety (Cpd12) that showed potet inhibition of the E6-mediated deprivation of p53 by disrupting the E6/ p53 binding in HPV16- and HPV18-positive CC cells. The authors re-ported that Cpd12 actively targeted CC cells altered through other high risk-HPV (HR-HPV) strains, including HPV68 and HPV45, and also against a HPV16-altered head/neck carcinogenic cell line. Thus, it was concluded that Cpd12 could be used in targeting a wide range of HPV- induced carcinomas (Celegato et al., 2021). Lee et al. reported direct interface between the PDZ binding motif (PBM) of HPV16 E6 and the PSD-95/Dlg/ZO-1 (PDZ) domain of human protein tyrosine phosphatase non-receptor type-4, established through structural and biochemical studies. Also, it was confirmed that the equivalent motifs from various other HR-HPV genotypes, such as HPV-18/31/33/45, bind to PTPN4-PDZ with similar affinities, signifying that PTPN4 acts as a common target of several pathogenic HPV genotypes (Lee et al., 2022).
Numerous phototherapeutic-based approaches have been reported for targeting HPV-mediated CC, both in vitro and in vivo. Studies showed that various herbal compounds directly abrogated HPV-E6/E7 activities and these compounds play a crucial role as a potential drug candidate which efficiently knockdown the oncoproteins and assist in CC regres-sion. Furthermore, numerous flavonoids have been recognized that specifically bind to the E6 proteins, leading to the inhibition of the p53 degradation which results a decrease in the viability of HPV-infected CC cells (Cherry et al., 2013). Numerous plant extracts and products such as latex from Ficus carica, flax-seed oil, Cudrania tricuspidate stem extracts reported to exhibit anti-oncogenic properties by inhibiting the expres-sions of E6 and E7 oncoproteins, following apoptosis (Deshpande et al., 2019; Ghanbari et al., 2019; Pertierra et al., 2017).
8. Conclusion and future perspectives
CC is a burgeoning issue plaguing the lives of many women across the world. While the causative organisms for the occurrence of CC remain to be a double-stranded DNA virus, the major aim as of now is to develop effective therapeutic approaches. Even though there has been the development of vaccines, the cost-effectiveness and accessibility continue to be a major issue. Here, we comprehensively reviewed the oncoproteins of HPV, their roles in various signaling pathways, cross- talk among the major oncoproteins, and targeting of cellular pathways for plausible development of novel therapeutic strategies.
The enzymatic role of E1 protein helps in HPV induced carcinoma with severe immunosuppressive nature concerning the immunological molecules which help in administration of HPV induced carcinoma with progression towards carcinogenesis. It can also be deduced that when E1 oncoprotein is targeted to control the prognosis of the HPV by regulating the enzymatic activity of E1 protein as the lead molecule for new drug molecules as if the enzymatic activity is dysregulated almost all the down-regulation of E1protein is controlled. The available data demon-strate that E2 has a dual role, therefore negative regulation of the E2 can easily be used to control the HPV and it can also serve as a potential target site. E4 may also play a role in arresting cell cycle as it has been evident that the epithelial cells which showed expression of E4 from HPV 16 and HPV 18 undergoes mitotic arrest at the G2 phase owing to the restriction of nuclear accumulation of CDK1 protein in the nucleus. This is suggestive of that if the phosphorylation of the E4 is controlled its activity is modulated and it is unable to affect the host. Thus, the epigenetic modulation of the E4 protein may serve as a novel therapetic target for future drug discovery. The inhibition of the E6/E6AP complex may represent a novel therapeutic approach to reactivate the function of p53 tumor suppressor protein. The interaction between E7 and HDAC inhibitors is also suggestive of promising therapeutic target for antiviral drugs in future.
HPV is regulated by several E proteins of which E5, E6, and E7 oncoprotein can regulate and alter several cellular signaling pathways to cause cancer. These molecular pathways play major roles in malignant transformation. E5 oncoprotein stimulates the MAPK-ERK pathway. E6 oncoprotein alters PI3K/AKT pathway, Wnt pathway, Notch pathway, and E7 regulates the PI3K/AKT pathway. Moreover, these oncoproteins also alter the NF-κB pathway, YY1 pathway, and apoptotic pathways (intrinsic and extrinsic). These altered pathways are the major cause of CC initiation. Targeting altered cell signaling pathways, as well as epidermal growth factor receptor and vascular endothelial growth fac-tor, has been recommended in several studies as a viable treatment method. Some inhibitors of these altered pathways have also been investigated in animal models and clinical studies. So, in essentiality, E5, E6, and E7 oncoproteins are the key players in cellular proliferation and cancer progression. Therefore, targeting these proteins and inactivating them in some aspects of their signaling pathways can prove to be promising anti-oncogenic therapeutic approaches.
Even though various advancement has been achieved to develop therapeutic strategies and techniques for targeting HPV-mediated CC,
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
13
however, tranlations of these aproaches in clinics are yet a major concern. The establishment of targeted delivery systems remains a crucial issue due to irregular particle size, surface charge, and lower membrane permeation (Bharti et al., 2018). Moreover, the application of genome editing tools in clinical usage have been a major concern due to safety, etical and economic issues. Nevertheless, the research findings are promising but more efficient research should be performed for the exploration of effective, targeted and affordable therapeutics to assist the populations affected with HPC-mediated CC.
Author contribution
KKK, NKJ and RB conceptualized, wrote, and edited the manuscript. RB, SSB, AN, DD, AB, SM, LK, SK, and SR performed the literature survey and drafted and edited the manuscript. NKJ ideated the schemes and performed artwork. SSD, SKS, VJU, DI, SA, SO and JR significantly contributed in editing and revision of the manuscript. KKK, NKJ and JR supervisied the work. All authors contributed to the article and approved the submitted version.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that influence the work reported in this paper.
Acknowledgements
Dr. Niraj Kumar Jha is thankful to Sharda University for the infra-structure and facility. The author would like to thank Deanship of Sci-entific Research at Majmaah University for supporting this work under project number No. R-2022-68. Kavindra Kumar Kesari and Janne Ruokolainen thanking Aalto University for providing open access sup-port. The authors would like to acknowledge the support from their respective institutes throughout the review writing process.
References
Adiga, D., Eswaran, S., Pandey, D., Sharan, K., Kabekkodu, S.P., 2021. Molecular landscape of recurrent cervical cancer. Crit. Rev. Oncol. Hematol. 157, 103178.
Akagi, K., Li, J., Broutian, T.R., Padilla-Nash, H., Xiao, W., Jiang, B., Rocco, J.W., Teknos, T.N., Kumar, B., Wangsa, D., 2014. Genome-wide analysis of HPV integration in human cancers reveals recurrent, focal genomic instability. Genome Res. 24 (2), 185–199.
Akhatova, A., Chan, C.K., Azizan, A., Aimagambetova, G., 2021. The efficacy of therapeutic DNA vaccines expressing the human papillomavirus E6 and E7 oncoproteins for treatment of cervical. Cancer System. Rev. Vaccines 10 (1).
Akhatova, A., Chan, C.K., Azizan, A., Aimagambetova, G., 2021. The efficacy of therapeutic DNA vaccines expressing the human papillomavirus E6 and E7 oncoproteins for treatment of cervical. Cancer System. Rev. Vaccines 10 (1), 53.
Al Moustafa, A.-E., Foulkes, W.D., Benlimame, N., Wong, A., Yen, L., Bergeron, J., Batist, G., Alpert, L., Alaoui-Jamali, M.A., 2004. E6/E7 proteins of HPV type 16 and ErbB-2 cooperate to induce neoplastic transformation of primary normal oral epithelial cells. Oncogene 23 (2), 350–358.
Arany, I., Grattendick, K.G., Tyring, S.K., 2002. Interleukin-10 induces transcription of the early promoter of human papillomavirus type 16 (HPV16) through the 5′- segment of the upstream regulatory region (URR). Antiviral Res. 55 (2), 331–339.
Arbyn, M., Weiderpass, E., Bruni, L., de Sanjose, S., Saraiya, M., Ferlay, J., Bray, F., 2020. Estimates of incidence and mortality of cervical cancer in 2018: a worldwide analysis. Lancet Glob. Health 8 (2), e191–e203.
Asiaf, A., Ahmad, S.T., Mohammad, S.O., Zargar, M.A., 2014. Review of the current knowledge on the epidemiology, pathogenesis, and prevention of human papillomavirus infection. Eur. J. Cancer Prevent. 23 (3), 206–224.
Bagarazzi, M.L., Yan, J., Morrow, M.P., Shen, X., Parker, R.L., Lee, J.C., Giffear, M., Pankhong, P., Khan, A.S., Broderick, K.E., Knott, C., Lin, F., Boyer, J.D., Draghia- Akli, R., White, C.J., Kim, J.J., Weiner, D.B., Sardesai, N.Y., 2012. Immunotherapy against HPV16/18 generates potent TH1 and cytotoxic cellular immune responses. Sci. Translat. Med. 4 (155), 155ra138.
Baleja, J.D., Cherry, J.J., Liu, Z., Gao, H., Nicklaus, M.C., Voigt, J.H., Chen, J.J., Androphy, E.J., 2006. Identification of inhibitors to papillomavirus type 16 E6 protein based on three-dimensional structures of interacting proteins. Antiviral Res. 72 (1), 49–59.
Barrow-Laing, L., Chen, W., Roman, A., 2010. Low- and high-risk human papillomavirus E7 proteins regulate p130 differently. Virology 400 (2), 233–239.
Bauknecht, T., Shi, Y., 1998. Overexpression of C/EBPβ represses human papillomavirus type 18 upstream regulatory region activity in HeLa cells by interfering with the binding of TATA-binding protein. J. Virol. 72 (3), 2113–2124.
Be, X., Hong, Y., Wei, J., Androphy, E.J., Chen, J.J., Baleja, J.D., 2001. Solution structure determination and mutational analysis of the papillomavirus E6 interacting peptide of E6AP. Biochemistry 40 (5), 1293–1299.
Berg, M., Stenlund, A., 1997. Functional interactions between papillomavirus E1 and E2 proteins. J. Virol. 71 (5), 3853–3863.
Bergvall, M., Melendy, T., Archambault, J., 2013. The E1 proteins. Virology 445 (1–2), 35–56.
Bharti, A.C., Singh, T., Bhat, A., Pande, D., Jadli, M., 2018. Therapeutic startegies for human papillomavirus infection and associated cancers. Front. Biosci.) 10 (1), 15–73.
Bobdey, S., Sathwara, J., Jain, A., Balasubramaniam, G., 2016. Burden of cervical cancer and role of screening in India. Indian journal of medical and paediatric oncology: official journal of Indian Society of Medical & Paediatric. Oncology 37 (4), 278.
Branca, M., Ciotti, M., Santini, D., Di Bonito, L., Benedetto, A., Giorgi, C., Paba, P., Favalli, C., Costa, S., Agarossi, A., 2004. Activation of the ERK/MAP kinase pathway in cervical intraepithelial neoplasia is related to grade of the lesion but not to high- risk human papillomavirus, virus clearance, or prognosis in cervical cancer. Am. J. Clin. Pathol. 122 (6), 902–911.
Brand, T.M., Iida, M., Corrigan, K.L., Braverman, C.M., Coan, J.P., Flanigan, B.G., Stein, A.P., Salgia, R., Rolff, J., Kimple, R.J., 2017. The receptor tyrosine kinase AXL mediates nuclear translocation of the epidermal growth factor receptor. Sci. Signal. 10 (460).
Bray, F., Ferlay, J., Soerjomataram, I., Siegel, R.L., Torre, L.A., Jemal, A., 2018. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 68 (6), 394–424.
Brehm, A., Nielsen, S.J., Miska, E.A., McCance, D.J., Reid, J.L., Bannister, A.J., Kouzarides, T., 1999. The E7 oncoprotein associates with Mi2 and histone deacetylase activity to promote cell growth. EMBO J. 18 (9), 2449–2458.
Brennan, K., Offiah, G., McSherry, E.A., Hopkins, A.M., 2009. Tight junctions: a barrier to the initiation and progression of breast cancer? J. Biomed. Biotechnol. 2010.
Burley, M., Roberts, S., Parish, J.L., 2020. Epigenetic regulation of human papillomavirus transcription in the productive virus life cycle. Seminars in Immunopathology. Springer, pp. 159–171.
Cardeal, L.Bd.S., Boccardo, E., Termini, L., Rabachini, T., Andreoli, M.A., di Loreto, C., Filho, A.L., Villa, L.L., Maria-Engler, S.S., 2012. HPV16 oncoproteins induce MMPs/ RECK-TIMP-2 imbalance in primary keratinocytes: possible implications in cervical carcinogenesis. PLoS One 7 (3), e33585.
Castro-Munoz, L.J., Manzo-Merino, J., Munoz-Bello, J.O., Olmedo-Nieva, L., Cedro- Tanda, A., Alfaro-Ruiz, L.A., Hidalgo-Miranda, A., Madrid-Marina, V., Lizano, M., 2019. The Human Papillomavirus (HPV) E1 protein regulates the expression of cellular genes involved in immune response. Sci. Rep. 9 (1), 1–13.
Celegato, M., Messa, L., Bertagnin, C., Mercorelli, B., Loregian, A., 2021. Targeted disruption of E6/p53 binding exerts broad activity and synergism with paclitaxel and topotecan against HPV-transformed cancer cells. Cancers 14 (1), 193.
Celegato, M., Messa, L., Bertagnin, C., Mercorelli, B., Loregian, A., 2022. Targeted disruption of E6/p53 binding exerts broad activity and synergism with paclitaxel and topotecan against HPV-transformed cancer cells. Cancers 14 (1), 193.
Chabeda, A., Yanez, R., Lamprecht, R., Meyers, A., Rybicki, E., Hitzeroth, I., 2018. Therapeutic vaccines for high-risk HPV-associated diseases. Papillomavirus Res. 5, 46–58. Epub 2017/12/27. https://doi. org/10.1016/j. pvr. 2017.12. 006. PubMed PMID.
Charette, S., McCance, D., 2007. The E7 protein from human papillomavirus type 16 enhances keratinocyte migration in an Akt-dependent manner. Oncogene 26 (52), 7386–7390.
Chaturvedi, A.K., 2010. Beyond cervical cancer: burden of other HPV-related cancers among men and women. J. Adol. Health 46 (4), S20–S26.
Chavez-Blanco, A., Perez-Sanchez, V., Gonzalez-Fierro, A., Vela-Chavez, T., Candelaria, M., Cetina, L., Vidal, S., Duenas-Gonzalez, A., 2004. HER2 expression in cervical cancer as a potential therapeutic target. BMC Cancer 4 (1), 1–6.
Chellappan, S., Kraus, V.B., Kroger, B., Munger, K., Howley, P.M., Phelps, W., Nevins, J., 1992. Adenovirus E1A, simian virus 40 tumor antigen, and human papillomavirus E7 protein share the capacity to disrupt the interaction between transcription factor E2F and the retinoblastoma gene product. Proceedings of the National Academy of Sciences 89(10), 4549–4553.
Chen, Y.-F., Lin, C.-W., Tsao, Y.-P., Chen, S.-L., 2004. Cytotoxic-T-lymphocyte human papillomavirus type 16 E5 peptide with CpG-oligodeoxynucleotide can eliminate tumor growth in C57BL/6 mice. J. Virol. 78 (3), 1333–1343.
Cheng, Y.-M., Chou, C.-Y., Hsu, Y.-C., Chen, M.-J., 2012. Influence of HPV16 E6/7 on the expression of FGF2 and FGFR type B in cervical carcinogenesis. Reprod. Sci. 19 (6), 580–586.
Cherry, J.J., Rietz, A., Malinkevich, A., Liu, Y., Xie, M., Bartolowits, M., Davisson, V.J., Baleja, J.D., Androphy, E.J., 2013. Structure based identification and characterization of flavonoids that disrupt human papillomavirus-16 E6 function. PLoS One 8 (12), e84506.
Chojnacki, M., Melendy, T., 2018. The human papillomavirus DNA helicase E1 binds, stimulates, and confers processivity to cellular DNA polymerase epsilon. Nucl. Acids Res. 46 (1), 229–241.
Conrad, M., Bubb, V., Schlegel, R., 1993. The human papillomavirus type 6 and 16 E5 proteins are membrane-associated proteins which associate with the 16-kilodalton pore-forming protein. J. Virol. 67 (10), 6170–6178.
Crook, T., Vousden, K.H., Tidy, J.A., 1991. Degradation of p53 can be targeted by HPV E6 sequences distinct from those required for p53 binding and trans-activation. Cell 67 (3), 547–556.
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
14
Cruz-Gregorio, A., Manzo-Merino, J., Gonzalez-García, M.C., Pedraza-Chaverri, J., Medina-Campos, O.N., Valverde, M., Rojas, E., Rodríguez-Sastre, M.A., García- Cuellar, C.M., Lizano, M., 2018. Human papillomavirus types 16 and 18 early- expressed proteins differentially modulate the cellular redox state and DNA damage. Int. J. Biol. Sci. 14 (1), 21.
D’Abramo, C., Archambault, J., 2011. Small molecule inhibitors of human papillomavirus protein-protein interactions. Open Virol. J. 5, 80.
de Carvalho Galvao, M.L.T., Coimbra, E.C., 2020. Long noncoding RNAs (lncRNAs) in cervical carcinogenesis: new molecular targets, current prospects. Crit. Rev. Oncol. Hematol. 156, 103111.
De Nola, R., Loizzi, V., Cicinelli, E., Cormio, G., 2021. Dynamic crosstalk within the tumor microenvironment of uterine cervical carcinoma: baseline network, iatrogenic alterations, and translational implications. Crit. Rev. Oncol. Hematol. 162, 103343.
de Villiers, E.-M., Fauquet, C., Broker, T.R., Bernard, H.-U., HH, zur, 2004. Classification of papillomaviruses. Virology 324, 17–27.
DeMasi, J., Huh, K.-W., Nakatani, Y., Münger, K., Howley, P.M., 2005. Bovine papillomavirus E7 transformation function correlates with cellular p600 protein binding. Proceedings of the National Academy of Sciences 102(32), 11486–11491.
Deshpande, R., Raina, P., Shinde, K., Mansara, P., Karandikar, M., Kaul-Ghanekar, R., 2019. Flax seed oil reduced tumor growth, modulated immune responses and decreased HPV E6 and E7 oncoprotein expression in a murine model of ectopic cervical cancer. Prostaglandins Other Lipid Med. 143, 106332.
Dhandapani, K.M., Mahesh, V.B., Brann, D.W., 2007. Curcumin suppresses growth and chemoresistance of human glioblastoma cells via AP-1 and NFκB transcription factors. J. Neurochem. 102 (2), 522–538.
DiMaio, D., Petti, L.M., 2013. The E5 proteins. Virology 445 (1–2), 99–114. Dinneen, J.L., Ceresa, B.P., 2004. Continual expression of rab5 (Q79L) causes a ligand-
independent EGFR internalization and diminishes EGFR activity. Traffic 5 (8), 606–615.
Disbrow, G.L., Sunitha, I., Baker, C.C., Hanover, J., Schlegel, R., 2003. Codon optimization of the HPV-16 E5 gene enhances protein expression. Virology 311 (1), 105–114.
Doorbar, J., 2013. The E4 protein; structure, function and patterns of expression. Virology 445 (1–2), 80–98.
DOORBAR, J., MEDCALF, E., NAPTHINE, S., 1996. Analysis of HPV1 E4 complexes and their association with keratinsin vivo. Virology 218 (1), 114–126.
Duensing, S., Duensing, A., Crum, C.P., Münger, K., 2001. Human papillomavirus type 16 E7 oncoprotein-induced abnormal centrosome synthesis is an early event in the evolving malignant phenotype. Cancer Re. 61 (6), 2356–2360.
Fani, M., Mahmoodi, P., Emadzadeh, M., Avan, A., Karimi, E., Ferns, G.A., Rezayi, M., Amiri, I.S., 2020. Correlation of human papillomavirus 16 and 18 with cervical cancer and their diagnosis methods in Iranian women: a systematic review and meta- analysis. Curr. Problems Cancer 44 (1), 100493.
Farley, J.H., Truong, V., Goo, E., Uyehara, C., Belnap, C., Larsen, W.I., 2006. A randomized double-blind placebo-controlled phase II trial of the cyclooxygenase-2 inhibitor Celecoxib in the treatment of cervical dysplasia. Gynecol. Oncol. 103 (2), 425–430.
Ferlay, J., Colombet, M., Soerjomataram, I., Mathers, C., Parkin, D., Pineros, M., Znaor, A., Bray, F., 2019. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 144 (8), 1941–1953.
Filippova, M., Song, H., Connolly, J.L., Dermody, T.S., Duerksen-Hughes, P.J., 2002. The human papillomavirus 16 E6 protein binds to tumor necrosis factor (TNF) R1 and protects cells from TNF-induced apoptosis. J. Biol.BChem. 277 (24), 21730–21739.
Galvao, M., Coimbra, E.C., 2020. Long noncoding RNAs (lncRNAs) in cervical carcinogenesis: new molecular targets, current prospects. Crit. Rev. Oncol. Hematol. 156, 103111.
Ganguly, N., 2012. Human papillomavirus-16 E5 protein: oncogenic role and therapeutic value. Cell. Oncol. 35 (2), 67–76.
Ganti, K., Broniarczyk, J., Manoubi, W., Massimi, P., Mittal, S., Pim, D., Szalmas, A., Thatte, J., Thomas, M., Tomaic, V., Banks, L., 2015. The human papillomavirus E6 PDZ binding motif: from life cycle to malignancy. Viruses 7 (7), 3530–3551.
Gewin, L., Myers, H., Kiyono, T., Galloway, D.A., 2004. Identification of a novel telomerase repressor that interacts with the human papillomavirus type-16 E6/E6- AP complex. Genes Dev. 18 (18), 2269–2282.
Ghanbari, A., Gresley, Le, Naughton, A., Kuhnert, D., Sirbu, N., Ashrafi, G.H, D., 2019. Biological activities of Ficus carica latex for potential therapeutics in Human Papillomavirus (HPV) related cervical cancers. Sci. Rep. 9 (1), 1–11.
Goodwin, E.C., DiMaio, D., 2000. Repression of human papillomavirus oncogenes in HeLa cervical carcinoma cells causes the orderly reactivation of dormant tumor suppressor pathways. Proceedings of the National Academy of Sciences 97(23), 12513–12518.
Graham, S.V., 2006. Late events in the life cycle of human papillomaviruses. Papillomavirus Res. Nat. History Vaccines Beyond 193–211.
Grm, H.S., Massimi, P., Gammoh, N., Banks, L., 2005. Crosstalk between the human papillomavirus E2 transcriptional activator and the E6 oncoprotein. Oncogene 24 (33), 5149–5164.
Guifen, L., Fachun, W., Enliang, S., Xiaomu, L., Xiuwen, T., Zhenshan, L., 2011. Effects of chromium picolinate supplementation on the growth, carcass quality and gene expression of beef during the finishing period. Mol. Biol. Rep. 38 (7), 4469–4474.
Gupta, S., Kumar, P., Das, B.C., 2018. HPV: molecular pathways and targets. Curr. Problems Cancer 42 (2), 161–174.
Gupta, S.M., Mania-Pramanik, J., 2019. RETRACTED ARTICLE: molecular mechanisms in progression of HPV-associated cervical carcinogenesis. J. Biomed. Sci. 26 (1), 1–19.
Hammes, L.S., Tekmal, R.R., Naud, P., Edelweiss, M.I., Kirma, N., Valente, P.T., Syrjanen, K.J., Cunha-Filho, J.S., 2007. Macrophages, inflammation and risk of
cervical intraepithelial neoplasia (CIN) progression—clinicopathological correlation. Gynecol. Oncol. 105 (1), 157–165.
Hasan, U.A., Zannetti, C., Parroche, P., Goutagny, N., Malfroy, M., Roblot, G., Carreira, C., Hussain, I., Müller, M., Taylor-Papadimitriou, J., 2013. The human papillomavirus type 16 E7 oncoprotein induces a transcriptional repressor complex on the Toll-like receptor 9 promoter. J. Exp. Med. 210 (7), 1369–1387.
Henderson, S., Chakravarthy, A., Su, X., Boshoff, C., Fenton, T.R., 2014. APOBEC- mediated cytosine deamination links PIK3CA helical domain mutations to human papillomavirus-driven tumor development. Cell Rep. 7 (6), 1833–1841.
Hinds, P.W., Mittnacht, S., Dulic, V., Arnold, A., Reed, S.I., Weinberg, R.A., 1992. Regulation of retinoblastoma protein functions by ectopic expression of human cyclins. Cell 70 (6), 993–1006.
Horner, S.M., DiMaio, D., 2007. The DNA binding domain of a papillomavirus E2 protein programs a chimeric nuclease to cleave integrated human papillomavirus DNA in HeLa cervical carcinoma cells. J. Virol. 81 (12), 6254–6264.
Hsu, P.D., Lander, E.S., Zhang, F., 2014. Development and applications of CRISPR-Cas9 for genome engineering. Cell 157 (6), 1262–1278.
Huh, K.-W., DeMasi, J., Ogawa, H., Nakatani, Y., Howley, P.M., Münger, K., 2005. Association of the human papillomavirus type 16 E7 oncoprotein with the 600-kDa retinoblastoma protein-associated factor, p600. Proceedings of the National Academy of Sciences 102(32), 11492–11497.
Ivashkiv, L.B., Donlin, L.T., 2014. Regulation of type I interferon responses. Nat. Rev. Immunol. 14 (1), 36–49.
Jin, B.Y., Campbell, T.E., Draper, L.M., Stevanovic, S., Weissbrich, B., Yu, Z., Restifo, N. P., Rosenberg, S.A., Trimble, C.L., Hinrichs, C.S., 2018. Engineered T cells targeting E7 mediate regression of human papillomavirus cancers in a murine model. JCI insight 3 (8).
Jones, D.L., Münger, K., 1996. Interactions of the human papillomavirus E7 protein with cell cycle regulators. Seminars in Cancer Biology. Elsevier, pp. 327–337.
Jung, Y.-S., Kato, I., Kim, H.-R.C., 2013. A novel function of HPV16-E6/E7 in epithelial–mesenchymal transition. Biochem. Biophys. Res. Commun. 435 (3), 339–344.
Kabsch, K., Alonso, A., 2002. The human papillomavirus type 16 E5 protein impairs TRAIL-and FasL-mediated apoptosis in HaCaT cells by different mechanisms. J. Virol. 76 (23), 12162–12172.
Kelley, M.L., Keiger, K.E., Lee, C.J., Huibregtse, J.M., 2005. The global transcriptional effects of the human papillomavirus E6 protein in cervical carcinoma cell lines are mediated by the E6AP ubiquitin ligase. J. Virol. 79 (6), 3737–3747.
Kennedy, E.M., Kornepati, A.V., Goldstein, M., Bogerd, H.P., Poling, B.C., Whisnant, A. W., Kastan, M.B., Cullen, B.R., 2014. Inactivation of the human papillomavirus E6 or E7 gene in cervical carcinoma cells by using a bacterial CRISPR/Cas RNA-guided endonuclease. J. Virol. 88 (20), 11965–11972.
Khan, J., Davy, C.E., McIntosh, P.B., Jackson, D.J., Hinz, S., Wang, Q., Doorbar, J., 2011. Role of calpain in the formation of human papillomavirus type 16 E1^ E4 amyloid fibers and reorganization of the keratin network. J. Virol 85 (19), 9984–9997.
Kim, H.S., Kim, T., Kim, M.-K., Suh, D.H., Chung, H.H., Song, Y.S., 2013. Cyclooxygenase-1 and-2: molecular targets for cervical neoplasia. J. Cancer Prevent. 18 (2), 123.
Kim, S.-H., Juhnn, Y.-S., Kang, S., Park, S.-W., Sung, M.-W., Bang, Y.-J., Song, Y.-S., 2006. Human papillomavirus 16 E5 up-regulates the expression of vascular endothelial growth factor through the activation of epidermal growth factor receptor, MEK/ERK1, 2 and PI3K/Akt. Cell. Mol. Life Sci. CMLS 63 (7), 930–938.
Kim, S.-H., Oh, J.-M., No, J.-H., Bang, Y.-J., Juhnn, Y.-S., Song, Y.-S., 2009. Involvement of NF-κB and AP-1 in COX-2 upregulation by human papillomavirus 16 E5 oncoprotein. Carcinogenesis 30 (5), 753–757.
Kim, Y., Cairns, M.J., Marouga, R., Sun, L.-Q., 2003. E6AP gene suppression and characterization with in vitro selected hammerhead ribozymes. Cancer Gene Ther. 10 (9), 707–716.
Knight, G.L., Turnell, A.S., Roberts, S., 2006. Role for Wee1 in inhibition of G2-to-M transition through the cooperation of distinct human papillomavirus type 1 E4 proteins. J. Virol. 80 (15), 7416–7426.
Kranjec, C., Tomaic, V., Massimi, P., Nicolaides, L., Doorbar, J., Banks, L., 2016. The high-risk HPV E6 target scribble (hScrib) is required for HPV E6 expression in cervical tumour-derived cell lines. Papillomavirus Res. 2, 70–77.
Lee, H.S., Yun, H.Y., Lee, E.W., Shin, H.C., Kim, S.J., Ku, B., 2022. Structural and biochemical analysis of the PTPN4 PDZ domain bound to the C-terminal tail of the human papillomavirus E6 oncoprotein. J. Microbiol.
Lee, S.S., Weiss, R.S., Javier, R.T., 1997. Binding of human virus oncoproteins to hDlg/ SAP97, a mammalian homolog of the Drosophila discs large tumor suppressor protein. Proceedings of the National Academy of Sciences 94(13), 6670–6675.
Leechanachai, P., Banks, L., Moreau, F., Matlashewski, G., 1992. The E5 gene from human papillomavirus type 16 is an oncogene which enhances growth factor- mediated signal transduction to the nucleus. Oncogene 7 (1), 19.
Li, S., Labrecque, S., Gauzzi, M.C., Cuddihy, A.R., Wong, A.H., Pellegrini, S., Matlashewski, G.J., Koromilas, A.E., 1999. The human papilloma virus (HPV)-18 E6 oncoprotein physically associates with Tyk2 and impairs Jak-STAT activation by interferon-α. Oncogene 18 (42), 5727–5737.
Liu, F., Lin, B., Liu, X., Zhang, W., Zhang, E., Hu, L., Ma, Y., Li, X., Tang, X., 2016. ERK signaling pathway is involved in HPV-16 E6 but not E7 oncoprotein-induced HIF-1α protein accumulation in NSCLC cells. Oncol. Res. 23 (3), 109.
Liu, X., Clements, A., Zhao, K., Marmorstein, R., 2006. Structure of the human Papillomavirus E7 oncoprotein and its mechanism for inactivation of the retinoblastoma tumor suppressor. J. Biol. Chem. 281 (1), 578–586.
Longworth, M.S., Laimins, L.A., 2004. The binding of histone deacetylases and the integrity of zinc finger-like motifs of the E7 protein are essential for the life cycle of human papillomavirus type 31. J. Virol. 78 (7), 3533–3541.
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
15
Makarova, K.S., Wolf, Y.I., Alkhnbashi, O.S., Costa, F., Shah, S.A., Saunders, S.J., Barrangou, R., Brouns, S.J., Charpentier, E., Haft, D.H., Horvath, P., Moineau, S., Mojica, F.J., Terns, R.M., Terns, M.P., White, M.F., Yakunin, A.F., Garrett, R.A., van der Oost, J., Backofen, R., Koonin, E.V., 2015. An updated evolutionary classification of CRISPR-Cas systems. Nat. Rev. Microbiol. 13 (11), 722–736.
Mandal, R., Senbabaoglu, Y., Desrichard, A., Havel, J.J., Dalin, M.G., Riaz, N., Lee, K.W., Ganly, I., Hakimi, A.A., Chan, T.A., Morris, L.G., 2016. The head and neck cancer immune landscape and its immunotherapeutic implications. JCI Insight 1 (17), e89829.
Manzo-Merino, J., Contreras-Paredes, A., Vazquez-Ulloa, E., Rocha-Zavaleta, L., Fuentes- Gonzalez, A.M., Lizano, M., 2014. The role of signaling pathways in cervical cancer and molecular therapeutic targets. Arch. Med. Res. 45 (7), 525–539.
Martinez-Zapien, D., Ruiz, F.X., Poirson, J., Mitschler, A., Ramirez, J., Forster, A., Cousido-Siah, A., Masson, M., Pol, S.V., Podjarny, A., 2016. Structure of the E6/ E6AP/p53 complex required for HPV-mediated degradation of p53. Nature 529 (7587), 541–545.
McIntosh, P.B., Laskey, P., Sullivan, K., Davy, C., Wang, Q., Jackson, D.J., Griffin, H.M., Doorbar, J., 2010. E1^ E4-mediated keratin phosphorylation and ubiquitylation: A mechanism for keratin depletion in HPV16-infected epithelium. J. Cell Sci. 123 (16), 2810–2822.
McINTYRE, M.C., Frattini, M.G., Grossman, S.R., Laimins, L.A., 1993. Human papillomavirus type 18 E7 protein requires intact Cys-XX-Cys motifs for zinc binding, dimerization, and transformation but not for Rb binding. J. Virol. 67 (6), 3142–3150.
Menges, C.W., Baglia, L.A., Lapoint, R., McCance, D.J., 2006. Human papillomavirus type 16 E7 up-regulates AKT activity through the retinoblastoma protein. Cancer Res. 66 (11), 5555–5559.
Moody, C.A., Laimins, L.A., 2010. Human papillomavirus oncoproteins: pathways to transformation. Nat. Rev. Cancer 10 (8), 550–560.
Moon, R.T., Kohn, A.D., De Ferrari, G.V., Kaykas, A., 2004. WNT and β-catenin signalling: diseases and therapies. Nat. Rev. Gen. 5 (9), 691–701.
Nakagawa, S., Huibregtse, J.M., 2000. Human scribble (Vartul) is targeted for ubiquitin- mediated degradation by the high-risk papillomavirus E6 proteins and the E6AP ubiquitin-protein ligase. Mol. Cell. Biol. 20 (21), 8244–8253.
Network, C.G.A.R., 2017. Integrated genomic and molecular characterization of cervical cancer. Nature 543 (7645), 378.
Nguyen, M., Song, S., Liem, A., Androphy, E., Liu, Y., Lambert, P.F., 2002. A mutant of human papillomavirus type 16 E6 deficient in binding α-helix partners displays reduced oncogenic potential in vivo. J. Virol. 76 (24), 13039–13048.
Oetke, C., Auvinen, E., Pawlita, M., Alonso, A., 2000. Human papillomavirus type 16 E5 protein localizes to the Golgi apparatus but does not grossly affect cellular glycosylation. Arch. Virol. 145 (10), 2183–2191.
Oh, D.-Y., Kim, S., Choi, Y.-L., Cho, Y.J., Oh, E., Choi, J.-J., Jung, K., Song, J.-Y., Ahn, S. E., Kim, B.-G., 2015. HER2 as a novel therapeutic target for cervical cancer. Oncotarget 6 (34), 36219.
Ojesina, A.I., Lichtenstein, L., Freeman, S.S., Pedamallu, C.S., Imaz-Rosshandler, I., Pugh, T.J., Cherniack, A.D., Ambrogio, L., Cibulskis, K., Bertelsen, B., 2014. Landscape of genomic alterations in cervical carcinomas. Nature 506 (7488), 371–375.
Organization, W.H., 2017. Human papillomavirus vaccines: WHO position paper, May 2017–Recommendations. Vaccine 35 (43), 5753–5755.
Paavonen, J., 2007. Human papillomavirus infection and the development of cervical cancer and related genital neoplasias. Int. J. Infect. Dis. 11, S3–S9.
Pal, A., Kundu, R., 2020a. Human papillomavirus E6 and E7: the cervical cancer hallmarks and targets for therapy. Front. Microbiol. 10, 3116.
Pal, A., Kundu, R., 2020b. Human papillomavirus E6 and E7: the cervical cancer hallmarks and targets for therapy. Front. Microbiol. 3116.
Pappa, K.I., Kontostathi, G., Lygirou, V., Zoidakis, J., Anagnou, N.P., 2018. Novel structural approaches concerning HPV proteins: Insight into targeted therapies for cervical cancer. Oncol. Rep. 39 (4), 1547–1554.
Park, K., Atchison, M.L., 1991. Isolation of a candidate repressor/activator, NF-E1 (YY-1, delta), that binds to the immunoglobulin kappa 3’enhancer and the immunoglobulin heavy-chain mu E1 site. Proceedings of the National Academy of Sciences 88(21), 9804–9808.
Patel, D., Huang, S.M., Baglia, L.A., McCance, D.J., 1999. The E6 protein of human papillomavirus type 16 binds to and inhibits co-activation by CBP and p300. EMBO J. 18 (18), 5061–5072.
Pertierra, L.R., Hughes, K.A., Vega, G.C., Olalla-Tarraga, M.A., 2017. High resolution spatial mapping of human footprint across Antarctica and its implications for the strategic conservation of avifauna. PLoS one 12 (1), e0168280.
Phelps, W.C., Yee, C.L., Münger, K., Howley, P.M., 1988. The human papillomavirus type 16 E7 gene encodes transactivation and transformation functions similar to those of adenovirus E1A. Cell 53 (4), 539–547.
Ramachandran, I., Thavathiru, E., Ramalingam, S., Natarajan, G., Mills, W., Benbrook, D., Zuna, R., Lightfoot, S., Reis, A., Anant, S., 2012. Wnt inhibitory factor 1 induces apoptosis and inhibits cervical cancer growth, invasion and angiogenesis in vivo. Oncogene 31 (22), 2725–2737.
Rangarajan, A., Syal, R., Selvarajah, S., Chakrabarti, O., Sarin, A., Krishna, S., 2001. Activated Notch1 signaling cooperates with papillomavirus oncogenes in transformation and generates resistance to apoptosis on matrix withdrawal through PKB/Akt. Virology 286 (1), 23–30.
Razavi, Z.S., Tajiknia, V., Majidi, S., Ghandali, M., Mirzaei, H.R., Rahimian, N., Hamblin, M.R., Mirzaei, H., 2021. Gynecologic cancers and non-coding RNAs: epigenetic regulators with emerging roles. Crit. Rev. Oncol. Hematol. 157, 103192.
Razavi, Z.S., Tajiknia, V., Majidi, S., Ghandali, M., Mirzaei, H.R., Rahimian, N., Hamblin, M.R., Mirzaei, H., 2021. Gynecologic cancers and non-coding RNAs: Epigenetic regulators with emerging roles. Crit. Rev. Oncol. Hematol. 157, 103192.
Regan, J.A., Laimins, L.A., 2008. Bap31 is a novel target of the human papillomavirus E5 protein. J. Virol. 82 (20), 10042–10051.
Ren, C., Cheng, X., Lu, B., Yang, G., 2013. Activation of interleukin-6/signal transducer and activator of transcription 3 by human papillomavirus early proteins 6 induces fibroblast senescence to promote cervical tumourigenesis through autocrine and paracrine pathways in tumour microenvironment. Eur. J. Cancer 49 (18), 3889–3899.
Reschner, A., Bontems, S., Le Gac, S., Lambermont, J., Marcelis, L., Defrancq, E., Hubert, P., Moucheron, C., Mesmaeker, K.-D., Raes, M., 2013. Ruthenium oligonucleotides, targeting HPV16 E6 oncogene, inhibit the growth of cervical cancer cells under illumination by a mechanism involving p53. Gene Ther. 20 (4), 435–443.
Richards, K.H., Doble, R., Wasson, C.W., Haider, M., Blair, G.E., Wittmann, M., Macdonald, A., 2014. Human papillomavirus E7 oncoprotein increases production of the anti-inflammatory interleukin-18 binding protein in keratinocytes. J. Virol. 88 (8), 4173–4179.
Roberts, S., Hillman, M.L., Knight, G.L., Gallimore, P.H., 2003. The ND10 component promyelocytic leukemia protein relocates to human papillomavirus type 1 E4 intranuclear inclusion bodies in cultured keratinocytes and in warts. J. Virol. 77 (1), 673–684.
Rodon, J., Dienstmann, R., Serra, V., Tabernero, J., 2013. Development of PI3K inhibitors: lessons learned from early clinical trials. Nature reviews. Clin. Oncol. 10 (3), 143–153.
Rogel-Gaillard, C., Pehau-Arnaudet, G., Breitburd, F., Orth, G., 1993. Cytopathic effect in human papillomavirus type 1–Induced inclusion warts: In vitro analysis of the contribution of two forms of the viral E4 protein. J. invest. Dermatol. 101 (6), 843–851.
Rosl, F., Das, B.C., Lengert, M., Geletneky, K., Zur Hausen, H., 1997. Antioxidant-induced changes of the AP-1 transcription complex are paralleled by a selective suppression of human papillomavirus transcription. J. Virol. 71 (1), 362–370.
Sanchez-Perez, A.M., Soriano, S., Clarke, A.R., Gaston, K., 1997. Disruption of the human papillomavirus type 16 E2 gene protects cervical carcinoma cells from E2F-induced apoptosis. J. Gen. Virol. 78 (Pt 11), 3009–3018.
Serrano, B., Brotons, M., Bosch, F.X., Bruni, L., 2018. Epidemiology and burden of HPV- related disease. Best Pract. Res. Clin. Obs. Gynaecol. 47, 14–26.
Shah, M., Anwar, M.A., Park, S., Jafri, S.S., Choi, S., 2015. In silico mechanistic analysis of IRF3 inactivation and high-risk HPV E6 species-dependent drug response. Sci. Rep. 5 (1), 1–14.
Sima, N., Wang, W., Kong, D., Deng, D., Xu, Q., Zhou, J., Xu, G., Meng, L., Lu, Y., Wang, S., 2008. RNA interference against HPV16 E7 oncogene leads to viral E6 and E7 suppression in cervical cancer cells and apoptosis via upregulation of Rb and p53. Apoptosis 13 (2), 273–281.
Song, Z., Ding, L., Ren, Z., Sun, X., Yang, Q., Wang, L., Feng, M., Liu, C., Wang, J., 2017. Effects of Src on cervical cancer cells proliferation and apoptosis through ERK signal transduction pathway. Zhonghua Liu Xing Bing Xue Za Zhi Zhonghua Liuxingbingxue Zazhi 38 (9), 1246–1251.
Songock, W.K., Kim, S.-m, Bodily, J.M., 2017. The human papillomavirus E7 oncoprotein as a regulator of transcription. Virus Res. 231, 56–75.
Stevanovic, S., Draper, L.M., Langhan, M.M., Campbell, T.E., Kwong, M.L., Wunderlich, J.R., Dudley, M.E., Yang, J.C., Sherry, R.M., Kammula, U.S., 2015. Complete regression of metastatic cervical cancer after treatment with human papillomavirus–targeted tumor-infiltrating T cells. J. Clin. Oncol. 33 (14), 1543.
Stoppler, M.C., Straight, S.W., Tsao, G., Schlegel, R., Mccance, D.J., 1996. The E5 gene of HPV-16 enhances keratinocyte immortalization by full-length DNA. Virology 223 (1), 251–254.
Sun, K., Salmon, S., Yajjala, V.K., Bauer, C., Metzger, D.W., 2014. Expression of suppressor of cytokine signaling 1 (SOCS1) impairs viral clearance and exacerbates lung injury during influenza infection. PLoS Pathogens 10 (12), e1004560.
Suprynowicz, F., Disbrow, G., Krawczyk, E., Simic, V., Lantzky, K., Schlegel, R., 2008. HPV-16 E5 oncoprotein upregulates lipid raft components caveolin-1 and ganglioside GM1 at the plasma membrane of cervical cells. Oncogene 27 (8), 1071–1078.
Talora, C., Cialfi, S., Segatto, O., Morrone, S., Choi, J.K., Frati, L., Dotto, G.P., Gulino, A., Screpanti, I., 2005. Constitutively active Notch1 induces growth arrest of HPV- positive cervical cancer cells via separate signaling pathways. Exp. Cell Res. 305 (2), 343–354.
Talora, C., Sgroi, D.C., Crum, C.P., Dotto, G.P., 2002. Specific down-modulation of Notch1 signaling in cervical cancer cells is required for sustained HPV-E6/E7 expression and late steps of malignant transformation. Genes Dev. 16 (17), 2252–2263.
Tenaillon, O., Barrick, J.E., Ribeck, N., Deatherage, D.E., Blanchard, J.L., Dasgupta, A., Wu, G.C., Wielgoss, S., Cruveiller, S., Medigue, C., 2016. Tempo and mode of genome evolution in a 50,000-generation experiment. Nature 536 (7615), 165–170.
Thomas, M., Laura, R., Hepner, K., Guccione, E., Sawyers, C., Lasky, L., Banks, L., 2002. Oncogenic human papillomavirus E6 proteins target the MAGI-2 and MAGI-3 proteins for degradation. Oncogene 21 (33), 5088–5096.
Tian, R., Liu, J., Fan, W., Li, R., Cui, Z., Jin, Z., Huang, Z., Xie, H., Li, L., Huang, Z., 2022. Gene knock-out chain reaction enables high disruption efficiency of HPV18 E6/E7 genes in cervical cancer cells. Mol. Ther. Oncol. 24, 171–179.
Tian, R., Liu, J., Fan, W., Li, R., Cui, Z., Jin, Z., Huang, Z., Xie, H., Li, L., Huang, Z., Hu, Z., Zhou, P., Tian, X., 2022. Gene knock-out chain reaction enables high disruption efficiency of HPV18 E6/E7 genes in cervical cancer cells. Mol. Ther. Oncol. 24, 171–179.
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
16
Toussaint-Smith, E., Donner, D.B., Roman, A., 2004. Expression of human papillomavirus type 16 E6 and E7 oncoproteins in primary foreskin keratinocytes is sufficient to alter the expression of angiogenic factors. Oncogene 23 (17), 2988–2995.
Trimble, C.L., Morrow, M.P., Kraynyak, K.A., Shen, X., Dallas, M., Yan, J., Edwards, L., Parker, R.L., Denny, L., Giffear, M., Brown, A.S., Marcozzi-Pierce, K., Shah, D., Slager, A.M., Sylvester, A.J., Khan, A., Broderick, K.E., Juba, R.J., Herring, T.A., Boyer, J., Lee, J., Sardesai, N.Y., Weiner, D.B., Bagarazzi, M.L., 2015. Safety, efficacy, and immunogenicity of VGX-3100, a therapeutic synthetic DNA vaccine targeting human papillomavirus 16 and 18 E6 and E7 proteins for cervical intraepithelial neoplasia 2/3: a randomised, double-blind, placebo-controlled phase 2b trial. Lancet 386 (10008), 2078–2088.
Tsao, Y.-P., Li, L.-Y., Tsai, T.-C., Chen, S.-L., 1996. Human papillomavirus type 11 and 16 E5 represses p21 (WafI/SdiI/CipI) gene expression in fibroblasts and keratinocytes. J. Virol. 70 (11), 7535–7539.
Varilla, V., Atienza, J., Dasanu, C.A., 2013. Immune alterations and immunotherapy prospects in head and neck cancer. Expert Opin. Biol. Ther. 13 (9), 1241–1256.
Vats, A., Skrabar, N., Del Sal, G., Banks, L., 2022. Loss of the E6AP ubiquitin ligase induces p53-dependent phosphorylation of HPV-18 E6 in cells derived from cervical cancer. J. Virol. Jvi0150321.
Vats, A., Cerro, Trejo, Thomas, O., Banks, L, M., 2021. Human Papillomavirus E6 and E7: what remains? Tumour Virus Res. 11, 200213.
Veeraraghavalu, K., Subbaiah, V.K., Srivastava, S., Chakrabarti, O., Syal, R., Krishna, S., 2005. Complementation of human papillomavirus type 16 E6 and E7 by Jagged1- specific Notch1-phosphatidylinositol 3-kinase signaling involves pleiotropic oncogenic functions independent of CBF1; Su (H); Lag-1 activation. J. Virol. 79 (12), 7889–7898.
Vieira, V.C., Leonard, B., White, E.A., Starrett, G.J., Temiz, N.A., Lorenz, L.D., Lee, D., Soares, M.A., Lambert, P.F., Howley, P.M., 2014. Human papillomavirus E6 triggers upregulation of the antiviral and cancer genomic DNA deaminase APOBEC3B. MBio 5 (6), e02234–02214.
Wang, Q., Kennedy, A., Das, P., McIntosh, P.B., Howell, S.A., Isaacson, E.R., Hinz, S.A., Davy, C., Doorbar, J., 2009. Phosphorylation of the human papillomavirus type 16 E1^ E4 protein at T57 by ERK triggers a structural change that enhances keratin binding and protein stability. J. Virol. 83 (8), 3668–3683.
Wang, W., Sima, N., Kong, D., Luo, A., Gao, Q., Liao, S., Li, W., Han, L., Wang, J., Wang, S., 2010. Selective targeting of HPV-16 E6/E7 in cervical cancer cells with a potent oncolytic adenovirus and its enhanced effect with radiotherapy in vitro and vivo. Cancer Lett. 291 (1), 67–75.
Wang, X., Meyers, C., Wang, H.-K., Chow, L.T., Zheng, Z.-M., 2011. Construction of a full transcription map of human papillomavirus type 18 during productive viral infection. J. Virol. 85 (16), 8080–8092.
Warren, C.J., Westrich, J.A., Doorslaer, K.V., Pyeon, D., 2017. Roles of APOBEC3A and APOBEC3B in human papillomavirus infection and disease progression. Viruses 9 (8), 233.
White, P.W., Faucher, A.-M., Massariol, M.-J., Welchner, E., Rancourt, J., Cartier, M., Archambault, J., 2005. Biphenylsulfonacetic acid inhibitors of the human papillomavirus type 6 E1 helicase inhibit ATP hydrolysis by an allosteric mechanism involving tyrosine 486. Antimicrob. Agents Chemother. 49 (12), 4834–4842.
White, P.W., Pelletier, A., Brault, K., Titolo, S., Welchner, E., Thauvette, L., Fazekas, M., Cordingley, M.G., Archambault, J., 2001. Characterization of recombinant HPV6 and 11 E1 helicases: effect of ATP on the interaction of E1 with E2 and mapping of a minimal helicase domain. J. Biol. Chem. 276 (25), 22426–22438.
White, P.W., Titolo, S., Brault, K., Thauvette, L., Pelletier, A., Welchner, E., Bourgon, L., Doyon, L., Ogilvie, W.W., Yoakim, C., 2003. Inhibition of human papillomavirus DNA replication by small molecule antagonists of the E1-E2 protein interaction. J. Biol. Chem. 278 (29), 26765–26772.
Williams, V.M., Filippova, M., Soto, U., Duerksen-Hughes, P.J., 2011. HPV-DNA integration and carcinogenesis: putative roles for inflammation and oxidative stress. Future Virol. 6 (1), 45–57.
Wright, A.V., Nunez, J.K., Doudna, J.A., 2016. Biology and applications of CRISPR systems: harnessing nature’s toolbox for genome engineering. Cell 164 (1–2), 29–44.
Wu, D.-W., Chuang, C.-Y., Lin, W.-L., Sung, W.-W., Cheng, Y.-W., Lee, H., 2014. Paxillin promotes tumor progression and predicts survival and relapse in oral cavity squamous cell carcinoma by microRNA-218 targeting. Carcinogenesis 35 (8), 1823–1829.
Yang, A., Jeang, J., Cheng, K., Cheng, T., Yang, B., Wu, T.-C., Hung, C.-F., 2016. Current state in the development of candidate therapeutic HPV vaccines. Exp. Rev. Vaccines 15 (8), 989–1007.
Zagouras, P., Stifani, S., Blaumueller, C.M., Carcangiu, M.L., Artavanis-Tsakonas, S., 1995. Alterations in Notch signaling in neoplastic lesions of the human cervix. Proceedings of the National Academy of Sciences 92(14), 6414–6418.
Zahra, K., Patel, S., Dey, T., Pandey, U., Mishra, S.P., 2021. A study of oxidative stress in cervical cancer-an institutional study. Biochem. Biophys. Rep. 25, 100881.
Zanier, K., ould M’hamed, ould Sidi, Boulade-Ladame, A., Rybin, C., Chappelle, V., Atkinson, A., Kieffer, A., Trave, G, B., 2012. Solution structure analysis of the HPV16 E6 oncoprotein reveals a self-association mechanism required for E6-mediated degradation of p53. Structure 20 (4), 604–617.
Zhang, B., Chen, W., Roman, A., 2006. The E7 proteins of low-and high-risk human papillomaviruses share the ability to target the pRB family member p130 for degradation. Proceedings of the National Academy of Sciences 103(2), 437–442.
Zhang, Y., Dakic, A., Chen, R., Dai, Y., Schlegel, R., Liu, X., 2017. Direct HPV E6/Myc interactions induce histone modifications, Pol II phosphorylation, and hTERT promoter activation. Oncotarget 8 (56), 96323.
Zheng, Y.-F., Rao, Z.-G., Zhang, J.-R., 2002. Effects of anti-HPV16 E6-ribozyme on the proliferation and apoptosis of human cervical cancer cell line CaSKi. Di 1 Jun Yi Da Xue Xue Bao Acad. J. First Med. Coll. of PLA 22 (6), 496–498.
Zheng, Y., Zhang, J., Rao, Z., 2004. Ribozyme targeting HPV16 E6E7 transcripts in cervical cancer cells suppresses cell growth and sensitizes cells to chemotherapy and radiotherapy. Cancer biology & therapy 3 (11), 1129–1134.
Zur, H.H., 2002. Papillomaviruses and cancer: from basic studies to clinical application. Nature reviews cancer 2 (5), 342–350.
Zwerschke, W., Mazurek, S., Massimi, P., Banks, L., Eigenbrodt, E., Jansen-Dürr, P., 1999. Modulation of type M2 pyruvate kinase activity by the human papillomavirus type 16 E7 oncoprotein. Proceedings of the National Academy of Sciences 96(4), 1291–1296.
Rahul Bhattacharjee is currently pursuing his doctoral studies at Sackler School of Medicine in Tel Aviv University on cancer immunology. He has done his Bachelors and Masters from from Kalinga Institute of Industrial Technology (KIIT deemed to be Uni-versity), Bhubaneswar, and did his master’s thesis work from Indian Institute of Chemical Biology (IICB), India. He did a specialization on immunology from Taiwan Medical Uni-versity. He has published reviews and research paper in reputed journals. His area of in-terest includes Cancer signaling and mechanism.
Sabya Sachi Das is pursuing his doctorate from the Department of Pharmaceutical Sci-ences and Technology, Birla Institute of Technology, Mesra, Ranchi, Jharkhand. His area of research majorly includes solid dispersions, polymer/lipid-based nano-drug delivery systems, metallic nanoparticles, herbal nanotherapeutics, targeted nanomedicines for diseases, including, cancer, neurological disorders, microbial diseases, and others. To date, he has published peer-reviewed scientific articles (>15) and also book chapters (>15).
Smruti Sudha Biswal is currently an Integrated Masters student at National Institute of Technology, Rourkela. She has been research intern at several renowned institutions like CSIR-IICB, Kolkata, NIBMG, Tamkang University, Taiwan in the field of cancer biology, neuroscience and developmental biology. Her current area of research is focused on un-derstanding hypoxia and its pathological context using Drosophila model as well as cancer therapuetics.
Arijit Nath is currently a part of the Pathology and Cancer Screening team as project fellow at Chittaranjan National Cancer Institute (CNCI), India. He received his master’s degree in Biotechnology in 2021 from Kalinga Institute of Industrial Technology (KIIT deemed to be University), Bhubaneswar. His research interests are mainly focused on investigating the molecular mechanisms of breast cancer particularly in TNBC.
Debangshi Das is currently pursuing her M.Tech degree in Environmental Biotechnology at Jadavpur University. She has received a B.Tech degree in Biotechnology from Haldia Institute of Technology affiliated to Maulana Abul Kalam Azad University of Technology. Her research interests include cancer biology and environmental microbiology.
Asmita Basu is presently working in research and development department at Biocon limited. She has received her master’s degree in Biotechnology from KIIT university in 2021. She is interested in understanding downstream processing of peptides and proteins.
Sumira Mallick is an Assistant Professor in Amity institute of Biotechnology, Jharkhand, Ranchi. She is a former, Assistant professor and Academic coordinator as well as Research Coordinator (Agriculture department, R and D dept) at Shivalik Institute of Professional Studies in Dehradun. She has received her Postdoctoral training from School of Biological Science and Biotech, Chonnam National University (CNU), Gwangju, South Korea. She received her Doctoral and Master’s degree from Hormone Research Centre, Gwangju, South Korea.
Lamha Kumar is currently pursuing her masters in science in biological sciences from the Indian Institute of Science Education and Research, Thivandrum. She did Bachelor of Science with majors in botany from the University of Delhi, India. Her area of interest is to understand the pathogenesis and therepautics of cancer.
Sulagna Kar is currently pursuing her integrated dual degree (B.Tech- M.Tech) in Biotechnology at Kalinga Institute of Industrial Technology (KIIT deemed to be Univer-sity), Bhubaneswar. She is doing her Masters dissertation work from Indian Council of Medical Research, Bhubaneswar, India. Her area of interest is understanding cellular signaling mechanisms in cervical cancer.
Sandeep Kumar Singh is an Associate Professor in the Department of Pharmaceutical Sciences and Technology at Birla Institute of Technology, Mesra, Ranchi, Jharkhand since 2006-till date. He has expertise in the field of Lipid-Based Drug Delivery System, Silk Nanoparticles, Self-Nanoemulsifying Drug Delivery System, In silico Pharmacokinetics Simulations, Quality by design (QbD). He contributed over 75 scholarly publications, including nearly original research papers, reviews and book chapters in peer reviewed and referred journals of repute. He serves as a reviewer for various high-impactreputed journals.
Vijay Jagdish Upadhye currently serving as Deputy Director Center of Research for Development (CR4D) and Associate Professor Parul Institute of Applied Sciences, Parul University, Vadodara, Gujarat. He has research experience of > 12 years on Immunodi-agnostics for Tuberculosis, Hepatitis and Infectious Diseases, Studies on Biomarkers Identification and exploring them on various diagnostic platforms, Macrophage and cell culture studies, Enzyme kinetics and protein purifications studies.
R. Bhattacharjee et al.

Critical Reviews in Oncology / Hematology 174 (2022) 103675
17
Dr. Danish Iqbal obtained his Ph.D. degree in 2015 from Integral University, India. Currently, he holds the position of Assistant Professor at the Majmaah University, Saudi Arabia. His research interest is in small organic molecules-based therapeutics to manage metabolic and infectious disorders. His major research theme is enzyme inhibition ki-netics. He is involved in ongoing research projects from different funding agencies in Saudi Arabia.
Suliman Almojam currently working as a Laboratory Manager at Vector-borne & Zoo-notic Diseases General Directorate, Ministry of Health, Riyadh, Saudi Arabia. He obtained his bachelor’s from King Saud University, Riyadh, Saudi Arabia in Applied medical sci-ences followed by a master’s in Molecular Biology from the University of Queensland, Australia. His research interest is cancer biology.
Shubhadeep Roychoudhury is an Assistant Professor in the Department of Life Science and Bioinformatics, Assam University, Silchar, India. He obtained Master and Doctoral degrees in Biotechnology from Slovakia, Europe. He completed several Postdoctoral re-searches including one as a Research Fellow in Andrology/Urology in the American Center for Reproductive Medicine, Department of Urology, Glickman Urological and Kidney Institute, Cleveland Clinic, Ohio, USA. He has been serving as visiting scientist and visiting Professor in institutes of repute including the Pedagogical University in Krakow in Poland, Mendel University in Brno in Czech Republic and Slovak University of Agriculture in Nitra, Slovakia. His teaching and research activities span the fields of reproductive health, reproductive and environmental toxicology, and oxidative stress within the context of male pathophysiology and reproductive dysfunction as well as herbal applications in management. He has published over 85 research papers in scientific journals, 30 book chapters, 4 books and presented over 50 papers at national and international scientific meetings. His Hirsch index (h-index) is 22 on Google Scholar, 19 on Scopus. He has con-ducted several research projects and received young scientist awards.
Shreesh Ojha is currently Associate Professor in the Department of Pharmacology and Therapeutics at the College of Medicine and Health Sciences, United Arab Emirates Uni-versity, Al Ain, United Arab Emirates. He holds a doctoral degree in Pharmacology from All India Institute of Medical Sciences, India following postdoctoral training from College of Medicine and BIO 5 Institutes of University of Arizona, Tucson, Arizona, USA. He also worked as Research Scientist in Clinical Pharmacology and Drug Development division with a pharmaceutical company, Ranbaxy Research Laboratories. His research expertise span from preclinical to clinical pharmacology across industries to academia. He is on the editorial board of many journals and reviewer for several reputed journals. Dr. Ojha has published more than 200 manuscripts including original research, reviews and book chapters. His scholarly work has recieved more than 6000 citations with H-index of 43 (google scholar). His research interest is natural products pharmacology and caanabinoid signaling in cancer, neurodegenerative and cardiovascular diseases. He is reciepient of many research grants including Sheikh Hamdan Award for Medical Sciences and Abu Dhabi Department of Education and Knowledge (ADEK), United Arab Emirates.
Janne Ruokolainen is a Professor in the Department of Applied Physics, Aalto University, Espoo, Finland. He gained his PhD (Tech) in Materials Physics from Helsinki University of Technology and Docentship in polymer physics and soft materials microscopy. His current research activities focus the characterization of molecular bioactive compounds extracted from plant species in neurodegenerative disease treatment. He has published over 250 scientific papers in peer reviewed and refereed journals. Since 1995, he has gained high bibliometric factor, where H-index 65 (google scholar). His awards and research honours include invitation to the Alfred Nobel Symposium (2005), Young Scientist Award by Foundation of Technology (2002), Prize for the best PhD Thesis in the field of technical sciences by Tekniikan Akateemisten Liitto and Tekniska Foreningen in Finland (TEK/TFiF) (2001), IUPAC Prize for Young Chemists – Honourable Mention Award (2001).
Niraj Kumar Jha is currently Assistant Professor in the Department of Biotechnology, Sharda University, Greater Noida, India. He is a former Assistant Professor of the Faculty of Biotechnology, Noida Institute of Engineering and Technology (NIET), affiliated with Abdul Kalam Technical University (AKTU), India. He holds postgraduate degree in Biotechnology from Vellore Institute of Technology, Tamilnadu, India and Doctorate in Biotechnology from Delhi Technological University with a research fellowship from Department of Biotechnology, Government of India. He has published over 100 quality scholarly work including original research, reviews and book chapters. He has presented his research work in prestigious international conferences and annual meetings of the professional scientific associations. He is editorial board member as well as guest editor of numerous peer reviewed and referred international journals of high impact and reputa-tion. His research interests include understanding the pathogenesis of cancer, neurode-generative, and cardiovascular diseases and deciphering the therapeutic and preventive role of smalll molecules of natural or synthetic origin.
Dr. Kavindra Kumar Kesari is currently working as Senior Scientist in the Department of Bioproducts and Biosystems, School of Chemical Engineering at the Aalto University, Espoo, Finland. He obtained Doctoral degree in Biotechnology from India. He received Junior and Senior Research fellowships during his Doctoral studies at Jawaharlal Nehru University, New Delhi, India. He served as an Assistant Professor position at Jaipur Na-tional University, Jaipur, India. He obtained his first Postdoctoral research from University of Eastern Finland, Kuopio, Finland. Currently, he is working on cell ultrastructure study with micro & macro spectroscopic techniques. His study explores the medicinal applica-tions of plant derived bioactive compounds and nanoparticles for disease treatment. Kesari, actively involved working in cell and cancer biology research with focus on qualitative and quantitative measurements by applying in vitro, in vivo & bio-computational methods. He has published over 100 papers in reputed scientific journals, 25 book chapters, 5 books and presented over 30 papers at national and international scientific meetings. His Hirsch index (h-index) as of March 2022 is 30 on Google Scholar. Kesari served as subject expert for many International committee and also Editorial board member and reviewer of repute. He has received four times young scientist award and international travel award. Received several grants and supervised master and doctoral students.
R. Bhattacharjee et al.



















![[PATTENT] Cervical plate system](https://img.dokumen.tips/doc/110x75/635f6512cdabb5e9220a1781/pattent-cervical-plate-system.jpg)









