Embed Size (px)
Citation preview

Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r.com/ locate /pa laeo
Marine biota response to clastic sediment influx: A quantitative approach
Stephen W. Lokier a,⁎, Moyra E.J. Wilson b,⁎, Lisa M. Burton c,⁎a Petroleum Geosciences Program, The Petroleum Institute, P.O. Box 2533, Abu Dhabi, United Arab Emiratesb Department of Applied Geology, Curtin University, P.O. Box U1987, Perth, WA 6845, Australiac StatoilHydro Petroleum ASA, Postboks 910, NO-9481 Harstad, Norway
⁎ Corresponding authors.E-mail addresses: [email protected] (S.W. Lokier), m.w
(M.E.J. Wilson), [email protected] (L.M. Burton).
0031-0182/$ – see front matter © 2009 Elsevier B.V. Adoi:10.1016/j.palaeo.2009.07.007
a b s t r a c t
a r t i c l e i n f oArticle history:Received 13 October 2008Received in revised form 7 July 2009Accepted 13 July 2009Available online 21 July 2009
Keywords:SiliciclasticVolcaniclasticsCarbonatesCoralAlgaeLarger benthic foraminiferaSedimentationNutrientsJavaBorneoSpainCenozoic
There have been few quantitative studies of the effects of siliciclastic and volcaniclastic sediment influx oncarbonate producing organisms from the geological record. This study presents the results of quantitative field,petrographic and acid digestion analyses of samples from three Cenozoic shallow-water mixed carbonate-volcani/siliciclastic depositional environments—the Miocene Wonosari Formation of south Java, the MioceneBatu Putih Limestone of east Borneo, and the Eocene Collsuspina Limestone of northeast Spain. Carbonateproduction in these systems is dominated by larger benthic foraminifera, coralline algae and corals. Analyses ofthe quantity, grainsize and type of volcani/siliciclastic sediment within these mixed successions have allowedassessment of the influence of these factors on shallow-water carbonate producers.Larger benthic foraminifera and coralline algae exhibit the greatest tolerance to siliciclastic sediment influx(abundant up to 55 and 38 wt.%, respectively) at a range of grain sizes (to coarse sand). Platy corals are alsopresent within sediments containing a large proportion of insoluble material (b80%); however, these arelimited to clay-dominated horizonswith amaximum siliciclastic size offine sand grade. Branching andmassivecorals exhibit the least tolerance to large quantities of insolublematerial (usually b40% up tomedium sand, andusually b25% up to very fine sand, respectively). These variations probably reflect organism: 1) mobility, 2)ability to self clean, 3) morphology and 4) feedingmechanisms relative to: a) sediment settling, b) turbidity, c)abrasion, d) energy, e) water depth, f) light levels, and g) nutrients. The influx of volcaniclastic and siliciclasticgrains of comparable size does not appear to have any differential effect on carbonate producers; althoughcorals may be more susceptible to damage by angular volcanic shards.Geologically averaged rates of clastic accumulation and their potential influences on carbonate producers inferredhereinmaybedifficult to compare directlywith thedaily and seasonalfluctuations experiencedbymodern clasticinfluenced systems. However, this studymay aid our understanding of the likely effects of increased clastic influx(today commonly anthropogenically related) to marine carbonate environments over longer time periods.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
There is a long-standing hypothesis that either constant orpunctuated clastic input to the marine environment prohibits thedevelopment of carbonate producing organisms. This supposition is stillwidely promoted despite recent research (Mount, 1984; Doyle andRoberts, 1988; Woolfe and Larcombe, 1999; Wilson and Lokier, 2002;Sanders and Baron-Szabo, 2005) demonstrating that carbonate produc-tion has been common in areas with significant silici/volcaniclasticinflux throughout geologic time and continues today (Fig. 1). Fewstudies have attempted, however, to quantify the effects of silici/volcaniclastic sediment influxoncarbonateproducers in the rock record.This lack of data hinders our ability to understand the dynamicinteractions in these mixed sedimentary systems or to assess the futureof sediment-impacted reef systems.
ll rights reserved.
This paper provides quantitative data on theeffects of siliciclastic andvolcaniclastic sediment influx on Cenozoic carbonate producing biotafrom three locations that formed in different shallow-water environ-ments in a variety of tectonic settings (Fig. 1). The Neogene WonosariFormation of south Java, Indonesia accumulated as a large-scalecarbonate platform affected by volcaniclastic input in a tropical forearcsetting. The Batu Putih Limestone, also of Neogene age, formed asequatorial patch-reefs interbedded with proximal deposits of theMahakan Delta in the Indonesian portion of Borneo. The PaleogeneCollsuspina Limestone at Calders was deposited as subtropical shelfalsedimentswithin the south Pyrenean foreland basin system of northernSpain. These three locations were selected as they allow us to addressthe following specific objectives of this study. 1) To compare and assesspossible organism responses to varying quantities of non-carbonatesediment influx during different epochs of the Cenozoic. 2) To establishthe response of organisms to various sediment grain sizes. 3) Toascertain any differences in biotic responses to siliciclastic versusvolcaniclastic sediment influx. There will always be problems drawingdirect comparisons between geologically averaged rates of clastic influx

Fig. 1. Global map showing the distribution of Recent tropical and temperate carbonates (adapted from Wilson and Rosen, 1998; Spalding et al., 2001) together with areas of activevolcanic activity and figures for suspended sediment discharge from major drainage basins (adapted from Milliman and Meade, 1983). The three research localities are arrowed.
26 S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
and the daily and seasonal fluctuations experienced by modern clasticinfluenced systems. Notwithstanding this, the geological data doesallow evaluation of the presence and character of carbonate producingbiota to differing clastic content, grain sizes and grain types (silici-,versus volcaniclastic) during punctuated or more consistent clasticinflux. Associated with clastic influx there is the potential to inputnutrients, increase turbidity and decrease light levels (Larcombe et al.,2001;Wilson and Lokier, 2002; Sanders and Baron-Szabo, 2005). Thesefactors are difficult to infer from the rock record. However, carboncontent, indications of depositional energy together with grain sizes,and apparent changes in biota depth zonation with no evidence foractualwater depth changes were used to infer if these additional factorsmay have been important (Wilson and Lokier, 2002). The results anddiscussion sections focus on the occurrence of three major organismgroups (corals, larger benthic foraminifera and coralline algae) todifferent types of insoluble influx. Survivorship of the differentorganisms to differing rates, amounts and types of insoluble influx arealso discussed under different local environmental conditions andthrough comparison with modern systems.
2. Methodology
2.1. Fieldwork and sampling strategy
The three mixed carbonate–clastic formations in Java, Borneo andSpain were each evaluated as part of larger-scale studies to understandplatform evolution and factors affecting changes to depositionalenvironments and biota (Lokier, 2000;Wilson and Lokier, 2002; Burton,2004; Wilson, 2005). The depositional geometries, together with
temporal and spatial variations in facies and biota, were analysedthrough detailed logging and mapping. The percentage occurrence andgrowth forms of the carbonate producing biota were documentedduringfieldwork. Representative sampleswere taken fromallmeasuredsections for thin sectioning and further analysis. A subset of 8 measuredsections, showing clear siliciclastic–carbonate interactions, was selectedfor more detailed sampling and laboratory analysis. A total of 197samples from these sections were subjected to detailed thin sectiondescription, point counting, grain size analysis and acid digestion (Datarepository item: Appendix A).
2.2. Point counting
For each thin section, 200 data points were ‘counted’, with thisfigure found to give a statistically reliable representation of thecomponent percentages. An initial examination of the thin sectionwasmade to determine a suitable step increment, depending on meangrain size, that would accurately record all of the major componentsunder investigation (Harwood, 1988). All point counting results aregiven as volume percentage (vol.%). Where samples were not thinsectioned (typically for coral-rich units with a friable matrix),component percentages were on the basis of field observations(field estimates quantified through image analysis of outcropphotographs) and acid digestion results.
2.3. Acid digestion
A portion of each of the selected samples was subjected to aciddigestion to determine the percentage of carbonate and non-carbonate

27S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
material. The samples from Java and Spain were highly indurated andneeded to be milled to a fine powder prior to acid digestion. Thistechnique precluded meaningful quantitative analysis of the insolubleresidue. Samples from Borneo were more friable and lightly crushedallowing the insoluble residue to be sieved for grain size analysis afteracid digestion. Samples were weighed and placed in a beaker with 10%hydrochloric acid. The contents of the beaker were regularly agitatedandadditionalhydrochloric acidwasaddeduntil no further reactionwasobserved. The residuewasfiltered and rinsedwith distilledwater beforebeingoven dried at 40 °C. Thedried samplewasweighed and theweightpercentage (wt.%) of insoluble material and calcium carbonatecalculated. For some coral-rich units in Borneo acid digestion of justthe interstitial matrix (b0.5 cm) between corals (N0.5 cm) wasundertaken. The weight percent of insoluble material for these samplesfrom the acid digestions results has been recalibratedbyallowing for thepercentage of corals versus matrix recorded in the field.
2.4. Grain size analysis
Thewell-lithified nature of the studied deposits generally precludedthe use of sieve or laser particle size analysis. The insoluble residue ofjust the friable samples (not subjected tomilling prior to acid digestion,see Section 2.3) was sieved to quantify grain-size fractions present.During thin section analysis grain-size fractions were recorded withthose contributing b5% of the insoluble material noted as minorfractions. Disseminated insoluble clay-sized material was difficult todistinguish frommicrite during petrography. Point count results of clay-sizedmatrixwere comparedwithweight percent of insoluble clay-sizedmaterial to determine percentages of the carbonate and non-carbonatefractions. True grain dimensions are not easily quantified in twodimensional thin sections. To gain a measure of the largest size fractionthe apparent long and short grain diameter of the largest silici/volcaniclastic grains in each thin section was accurately measured, andtheir median (d50) calculated (Lewis andMcConchie,1997; after Harrelland Eriksson, 1979). Measurements were taken on ten grains and formost samples the size fractions commonly fell into the same grain sizeclass (for N10 grains therewas commonlya shift to lower size classes). Inthe case of volcaniclastic samples, pumice grains were ignored as thesegrains are easily deformed during compaction andwould therefore givespurious measurements. For samples that were not thin sectioned themedian grain diameter of the largest size fraction was determinedthrough binocular microscopy of insoluble material (after aciddigestion).
3. Study areas
3.1. Forearc carbonate platform, Java
The lower to middle Miocene carbonate lithologies of the WonosariFormation are exposed as a series of cone-karst hills over an area of~1900 km2 on the southern coast of central Java, Indonesia (Fig. 2). TheWonosari platform developed as a shallow-water tropical carbonateplatform in an active forearc setting. Northern and western areas werelargely protected from volcaniclastic sediments by the deep-waterWonosari Trough, this resulted in typically low (b1 wt.%) volumes ofinsoluble clasticmaterialwithinmost of the carbonate facies. The east ofthe platform experienced elevated volcaniclastic sedimentation due toinflux from proximal, and possibly attached volcanic centres (Lokier,2000). Volcaniclastic sediments were emplaced into the marineenvironment via a variety of sedimentary processes including airfall,pyroclastic flows and density flows. Distal airfall and sea-surfacepyroclastic flows both had the potential to bypass the sediment trap ofthe Wonosari Trough and deposit volcaniclastic sediments of a widerange of grain sizes and compositions. This episodic sedimentary influxhad adiscernable effect on the shallow-marine assemblages (Fig. 3) and,consequently, on carbonate platform development.
Field sections primarily crop out in karstic hills, as well as withinsome quarries and river sections and range in stratigraphic thicknessbetween 6 to 359 m. Of the 44 field sections examined (Lokier, 2000)3 were selected for more detailed quantitative analysis, these are theKali Sambi, Djatirago and Kali Baksoko sections (Figs. 2, 3, Table 1).
Shallow-water platform carbonates were deposited in the photiczone under a range of energy regimes. The admixed volcaniclasticcontent within the sediments is typically b1 wt.% but locally exceeds96 wt.%. Where volcaniclastic sedimentation is low the platform faciesare dominated by coralline algae, primarily as rhodoliths, and largerbenthic foraminifera (including Miogypsina, Lepidocyclinids, Hetero-steginids and imperforate foraminifera). Corals are rare and are limitedto locally-developed patch reefs b100 m across. The paucity of coralsmay result from a combination of unsuitable water-depth, unstablesubstrates, high-energy regimes, elevated nutrient levels and highturbidity (Lokier, 2000). Where clastic contents are elevated a distinctrelationship is observed between volcaniclastic content, carbonateproducing organisms and organism growth morphologies.
Coralline algae are present in lithologies containing up to 65 wt.%volcanic material. The incorporation of volcaniclastic grains betweenalgal laminae confirms that growth was contemporaneous withdeposition of their host sediment. A change in coralline algaemorphology, from concentric spheroidal forms to branching andcolumnar forms, reflects increased volcaniclastic influx (Wilson andLokier, 2002). Larger benthic foraminifera are common in sedimentscontaining up to 77 wt.% volcaniclastic material and are commonly thefirst biota observed above thicker volcaniclastic units. Foraminifera inthese volcaniclastic-rich sediments exhibit a change in test morphologyto thinner, larger diameter forms traditionally associatedwith growth indeeper water (Hallock, 1985; Hallock and Glenn, 1986). For example,Heterostegina tests reach up 1.5 cm in diameter in carbonate-dominatedunits but exceed 5 cm in diameter but only 1 mm thickness involcaniclastic-rich horizons. Test morphologies return to more globoseforms as volcaniclastic contents decrease. As there is no evidence for anincrease inwater depth on theWonosari Platform, thesemorphologicalchanges are, instead, interpreted as resulting fromvertical contraction ofthe photic zone due to the presence of suspended sediment.
3.2. Delta front patch reefs, Borneo
Mixed carbonate–siliciclastic development was common on theshallow shelves bordering land areas in SE Asia throughout theCenozoic (Wilson and Lokier, 2002; Wilson, 2002; Wilson and Hall, inpress). The humid equatorial climate resulted in significant terrestrialrunoff into shallow-marine areas, where the conditions are otherwisesuitable for prolific carbonate production (Wilson, 2008). Despiteconstant high siliciclastic input, modern and Miocene carbonatesformed as patch reefs on delta-front mouth bars (Wilson, 2005) andas more distal shelfal build-ups (Roberts and Sydow,1996) seaward ofthe Mahakam Delta in eastern Borneo (Fig. 2).
Data for this study was obtained from outcrops exposed in smallquarry sections, although subsurface cores and modern deposits werealso analysed as part of a broader study (BouDagher-Fadel andWilson,2000; Wilson and Lokier, 2002; Hook and Wilson, 2003; Wilson,2005). Twenty-three sections were studied in detail through 7 patchreefs. Out of these, 4 sections through the lower to predominantlymiddle Miocene Airputih and Dibelakan Parliament (DPR) patch reefsare discussed here (Figs. 2 and 4, Table 2). The deltaic succession isfolded into NNE–SSW trending anticlines and both patch reefs areexposed on the eastern limb of an anticline where deposits dipbetween 10 and 90°.
The modern Mahakam Delta has developed under conditions oflow wave-energy, moderate tides of about 3 m and a high but non-flooding fluvial discharge of between 500 and 5000 m3/s (Allen andChambers, 1998). It is inferred from outcrop and subsurface data thatthe Neogene delta developed under similar conditions, although

Fig. 2. Map of SE Asia (A) showing the locations of the south central Java (B) and eastern Borneo (C) study areas.
28 S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
fluvial flooding may have been more influential in the past. About8 million m3 of sediment are deposited in the delta annually with siltsand clays making up 70% and sands the remainder (Allen et al., 1976).
The proximal patch reefs had low relief and developed in b10 mwater depth. Patch reef deposits are b40 m thick, up to 4 km long andare commonly amalgamated, with siliciclastic units interdigitatingwith or separating vertically-stacked carbonate units. Admixedsiliciclastic content within the carbonates is never less than 6 wt.%and is often significantly higher constituting up to 80 wt.%. Coralsdominate the deposits, although larger benthic foraminifera, corallinealgae and echinoderm debris are also common (Table 2). There is acharacteristic zonation of lithologies, carbonate-producing organismsand growth forms that show a correlation with siliciclastic and/ororganic matter content (Fig. 4).
Initial carbonate development occurred as platy coral growth(Pachyseris and Leptoseris) on soft sediment. Siliciclastic content inthese sheetstones (cf. Insalaco,1998) is considerable (up to 60–80wt.%)and calcareous encrusters are rare. During times of lowered siliciclasticcontent, or in central areas of the patch reef, branching (Porites,Stylophora) and then head corals (Porites, Goniopora) become progres-sively dominant as the siliciclastic content decreases to around 25–35 wt.% and b10–15 wt.% respectively. Calcareous encrusters arecommon in these pillarstone or dome/mixstone facies. When siliciclas-tics onlap the margins of the patch reef, the reefs shrink in size and thegrowth forms change progressively back to branching and platy corals.These morphological changes are suggestive of apparent deepening inthe environment. However, there is no evidence for actual changes inwater depth, and instead the biota is inferred to be responding toreductions inwater claritywith increased suspended siliciclastic content(Wilson, 2005). Coralline algae and echinoids are most common in thesiliciclastic-rich deposits and highest abundances correlate with raisedcarbon contents in the deposits (up to 4%). Larger benthic foraminifera
packstones, interpreted as accumulating in shallow seagrass beds,commonly cap patch reef deposits, before a return to shallow-waterrippled siliciclastics (Wilson, 2005).
3.3. Foreland basin shelf carbonates, Spain
The middle to upper Eocene Collsuspina Limestones of the Calderssection are exposed in a series of east-west trending valleys coveringan area of approximately 4 km2 to the southwest of Vic, NE Spain(Fig. 5). Shallow-water carbonates dominated by corals, corallinealgae and benthic foraminifera developed on an actively progradinglate Paleogene siliciclastic shelf that delineated the southernmargin ofthe SE Pyrenean Foreland Basin System (Santisteban and Taberner,1988). These sediments reflect the final stages of fully marinesedimentation in the foreland prior to widespread accumulation ofcontinental fluvial sediments (Hendry et al., 1999). Carbonatedevelopment was limited to periods of reduced siliciclastic influxrelated to temporary shelf abandonment. However, there was aconstant input of clay and silt due to wave/current re-suspension andlongshore transport, in addition to continental runoff thought to havebeen influenced by the increasingly seasonal climatic regime(Cavagnetto and Anadon, 1996). Sixteen cliff and road sections wereexamined in detail in the area; the Calders section being selected fordetailed analysis (Fig. 6; Table 3).
The Calders section is characterised by a number of very low-anglecarbonate clinoformsN1 km across and up to ~25 m thick thatdevelopedon the seaward-margin of temporarilyabandoned siliciclasticdune foresets. Echinoids and large gastropods are the most commonbiotawithin siliciclastic sediments. Initial colonisation of the siliciclasticsubstrate is marked by the development of sea grass shoals (Burton,2004) that harboured an epiphytic biota dominated by large encrustingforaminifera (e.g. Gypsina, Haddonia, Chapmanina, Korobkovella and

Fig. 3. Measured section (A) and graph (B) showing the relationship between carbonate components and weight percentage non-carbonate material from Java. Measured section(A) is from the upper part of the Kali Sambi section and the total weight percent (carbonate and non-carbonate material) is compared with the relative abundances of biota. B is aquantified plot showing the relationship between the principal carbonate components and the total weight percentage of non-carbonate material for 3 sections (Kali Sambi (SAM),Kali Baksoko (BAK) and Djatirago (TAN)). In plot B where the sum of the carbonate components does not reach 100% the difference is due to a combination of non-carbonatematerialand fine-grained, fragmented bioclastic material.
29S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42

Table 1Brief summary of the three selected field sections from the Wonosari Platform, for more details see Wilson and Lokier (2002).
Section name,code, andcoordinates
Total sectionthickness
Age Nature ofoutcrop
Brief description of stratigraphy and lithofacies
(m)
Djatirago 359.5 Middle Miocene(Tf2)
Sea cliffs andkarstic hills
Punctuated transition from carbonate to volcaniclastic-dominated sedimentation. The lower 122.38 mcomprises relatively clean (less than 34 wt.% volcaniclastics), coralline algal and larger benthic foraminiferapackstones, rudstones and grainstones with subordinate coral bafflestones and wackestones. Between 122.38to 139.95 m fining-upward tuff units (69–98 wt.% volcaniclastics) contain rare gastropods and bivalves. Areturn to carbonate-dominated sedimentation is observed between 139.95 to 290.36 m (up to 25 wt.%volcaniclastics). The uppermost 69.10 m of the section sees a transition to volcaniclastic-dominated depositionwith tuffs (73–98 wt.% volcaniclastics) passing up-section into volcanic breccias. The onset of volcaniclasticsedimentation sees a marked decrease in coralline algae while larger benthic foraminifera continue to beabundant.
(TAN)S 08° 13.185′E 111° 07.180′
Kali Baksoko 122.5 Middle Miocene(Tf1)
River section Transition from volcaniclastic to carbonate-dominated sedimentation. The lower 37.43 m of this section isdominated by volcanic breccias, tuffs andmudstones (up to 96 wt.% volcaniclastics) with a small component oflarger benthic foraminifera. Above 37.43 m carbonate lithofacies dominate. Up-section there is a transitionfrom larger benthic foraminifera wackestones and packstones interbedded with coral framestones to corallinealgae packstones, rudstones and grainstones and platy coral boundstones (up to 46 wt.% volcaniclastics).These carbonate units are punctuated by, and locally interdigitate with, centimetre- to metre-scalevolcaniclastic units (up to 89 wt.% volcaniclastics).
(BAK)S 08° 08.912′E 111° 02.370′
Kali Sambi 177.3 Early–MiddleMiocene (Tf1–2)
Series of karstichills
Recording a transition from carbonate to volcaniclastic-dominated sedimentation. The lower 79 m comprisesrelatively clean packstones, rudstones and grainstones (5–6 wt.% volcaniclastics) dominated by coralline algaeand larger benthic foraminifera. An up-section increase in volcaniclastic sediment between 79 and 123 m (upto 20 wt.% volcaniclastics) is accompanied by increasing gastropod and echinoid abundance. Uppermost unitscomprise larger benthic foraminifera packstones, tuffaceous packstones and tuffs (up to 95 wt.%volcaniclastics) exhibiting a marked decrease in coralline algae abundance and an increase in larger benthicforaminifera, gastropods and branching or solitary corals.
(SAM)S 08° 12.214′E 110° 57.272′
All co-ordinates are to map datum WGS84.
30 S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
Gyroidinella). High-siliciclastic content (N65 wt.%) inhibited the devel-opment of a sessile benthic biota. With increased substrate stabilisationand waning siliciclastic content (b65 wt.%), larger benthic foraminifera
Fig. 4.Measured section,AirputihK (AK) fromBorneo showing the relationshipbetween carbon
(up to 65 vol.%) and coralline algae became the dominant biota. Skeletalcomponents are predominantly whole and unabraded and are morelikely to be a colonisation fauna rather than bioclasts exported from
ate components,weight percentagenon-carbonatematerial andnon-carbonate grain sizes.

Table 2Brief summary of the four selected field sections from the Batu Putih Limestone, for more details see Wilson and Lokier (2002), Wilson (2005).
Section name,code andcoordinates
Total sectionthickness
Age Nature ofoutcrop
Brief description of stratigraphy and lithofacies
(m)
Airputih A 29 Early to predominantly MiddleMiocene (Tf1–2 and NN4–NN5).
Quarrysection
Section through the centre of the Airputih patch reef consisting of clay-rich carbonates (seeWilson, 2005, p. 126). Clay-rich platy coral sheetstones (up to 40–80% clays) interbedded withbioclastic floatstones (together totalling 9 m) overlie laminated shales. Up-section over the next12 m the siliciclastic content decreases to ~25 wt.% and then ~6 wt.% as the lithologies change tobranching coral pillarstones and head coral mix-domestone facies respectively. Over the next5 m coralline algae and coral floatstone sharply overlie the domestones and pass upwards intoclay-rich platy coral sheetstones (up to 40 wt.% siliciclastics). A larger benthic foraminiferapackstone forms the uppermost bed (1 m) of the patch reef and is overlain by siliciclastics.
(AA)S 00° 28.180′E 117° 07.014′
Airputih F 18.5 Early to predominantly MiddleMiocene (Tf1–2 and NN4–NN5).
Quarrysection
Southernmost section through the margin of the Airputih patch reef consisting of clay-richcarbonates (see Wilson, 2005, p. 126). Platy coral sheetstones (2.5 m) containing ~40 wt.% claysoverlie laminated silty sands and clays. A 1 m thick bed of sandy packstone (50 wt.%siliciclastics) passes up into 6 m of clay-rich branching coral and foraminifera wackestones(~30 wt.% clays). The upper 6.5 m of the patch reef consists predominantly of larger benthicforaminifera packstones interbedded with, and rich in clays (30–50 wt.%).
(AF)S 00° 28.860′E 117° 06.664′
Airputih K 12.4 Early to predominantly MiddleMiocene (Tf1–2 and NN4–NN5).
Quarrysection
Northernmost section through the margin of the Airputih patch reef where siliciclasticsinterdigitate with clay-rich carbonates (see Wilson, 2005, p. 124 and 126). Clay-rich platy coralsheetstones (up to 40–80 wt.% clays) overlie siliciclastics and pass up-section into branchingcoral pillarstones and then into head coral domestones (~10 wt.% clays) over 4.5 m. 1 m ofbranching coral pillarstone and rudstone is overlain by 1 m of interbedded clays and siltstones.There is an upward transition into sandy packstones and then, as the siliciclastic contentdecreases, platy coral sheetstones (~60%) pass upwards into branching and then head coralmixstones (~20%) over 3 m. A larger benthic foraminifera packstone, containing ~50 wt.% clays,forms the uppermost bed (30 cm) of the patch reef and is overlain by siliciclastics.
(AK)S 00° 28.068′E 117° 07.034′
Dibelakan 10 Early to predominantly MiddleMiocene (NN4–NN5)
Low reliefquarrysection
Section through amalgamated clay-rich patch reef (see Wilson, 2005, p. 124). The initial 3 m ofplaty coral sheetstones (80–30 wt.% clays) are overlain by 2 m of platy and head coral mixstones(~15 wt.% siliciclastics). 1 m of bioclastic wacke-floatstone is overlain by 2 m of predominantlyclays. The clays are overlain by 1.5 m of branching coral pillarstone and then platy coralsheetstone, which are the uppermost deposits exposed.
Parliament(DPR)S 00° 30.496′E 117° 06.177′
All co-ordinates are to map datum WGS84.
31S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
other already established adjacent carbonate producing areas. As withthe Java and Borneo study areas, flattened discoidal foraminifera testsare common (e.g. Operculina, Nummulites and Discocyclina) and mayindicate growth in a setting with a compressed photic zone resultingfrom high turbidity rather than from increased water depth.
As siliciclastic sediment content decreases (b38wt.%) larger benthicforaminifera were progressively replaced by coral and coralline algae-dominated facies (including Porites, Alveopora, Astreopora, Siderastreaand various Faviidae; Hendry et al., 1999). Coral rubble dominates andonly localised patches (b100m diameter) of in-situ coral framework areobserved. Encrusting organisms, particularly larger benthic foramini-fera, occupying cryptic habitats within the coral rubble were abundant.In shallow, high-energy settings with significant sand-grade siliciclastic
Fig. 5. Tectonic domains of the Pyrenean Foreland Basin system (after Puigdefàbregaset al., 1986) showing the location of the Calders section.
sediment (b27 wt.%), robust branching corals and solitary coralsdeveloped. Conversely, in protected settings with a high clay-gradesiliciclastic content (b39 wt.%) delicate vertically branching formsdominated. Renewed progradation of siliciclastic dune foresets resultedin burial of carbonate-producing biota and the abrupt cessation ofcarbonate production.
4. Results
In all of the study areas the biota are well-preserved and areinterpreted to be in-situ, i.e. inhabiting the environment at the time ofsiliciclastic or volcaniclastic sedimentation rather than transportedinto a siliciclastic-rich setting post mortem.
The presence of 2 vol.% or more biota (i.e. likely to be in-situ) wasrecorded in 80% of the analysed samples from the Wonosari Platformcompared with 84% from the Calders section and 95% from the Borneooutcrops. In all cases the dominant biota are larger benthicforaminifera, coral and coralline algae (Fig. 7), however, the relativedominance of these three groups varied between the three study areas(Fig. 8; Appendix A). Larger benthic foraminifera were initiallyquantified according to genera. However, no specific relationshipwas identified between genera and the amount of insoluble materialor siliciclastic grainsize, only the overall abundance of larger benthicforaminifera was found to vary. In the Wonosari Platform sectionslarger benthic foraminifera (8 genera) are the principal biota in 32% ofthe samples followed by coralline algae in 25% and coral in 18% of thesamples. Larger benthic foraminifera (7 genera) are also the dominantbiota in the Calders section where they are the primary component in48% of the studied samples with coral dominating 36% of samples andcoralline algae in 15%. In the Borneo sections the skeletal component isdominated by corals in 77% of the samples while larger benthicforaminifera (4 genera) dominated in 19% and coralline algae weredominant only in 4% of cases.

32 S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42

Table 3Brief summary of the Calders Section, co-ordinates are to map datum WGS84.
Section name,code andcoordinates
Total sectionthickness
Age Nature ofoutcrop
Brief description of stratigraphy and lithofacies
(m)
Calders 124 Middle to Late Eocene(Bartonian to Priabonian)
Exposedvalleysection
Multiple punctuated transitions from siliciclastic to carbonate-dominated sediment accumulation. Metre-scale, low-angle carbonate clinoforms developed on the seaward margin of temporarily abandoned duneforesets. The siliciclastic to carbonate transition is typically characterised by a well-sorted litharenite withabundant (up to 21 vol.%) epiphytic Gypsina that is in turn overlain by a mixed carbonate–siliciclasticlithology (14 –50 wt.% siliciclastics) dominated by large benthic foraminifera (predominantlyNummuliteswith Discocyclina and Operculina (up to 51 vol.%). The upper portion of the carbonate intervals comprisesa mixture of coral and coralline algae-dominated packstones, rudstones, boundstones andmixstones withvariable amounts of clay to sand-grade siliciclastic material (11–65 wt.%). The transition from carbonatesto siliciclastics is sharp but non-erosional.
(CAL)N 41° 46.997′E 001° 59.905′
33S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
4.1. Relationship between quantity of siliciclastic material andoccurrence of biota
There is a strong correlation between the amount of siliciclasticmaterial in the samples and the occurrence of the three dominantcarbonate producing organisms (Fig. 9). Larger benthic foraminiferawere identified in 157 of the samples evaluated during the study. Theseorganisms exhibit the greatest tolerance to the presence of siliciclasticsediments, beingobserved in samples containingup to96wt.% insolublematerials (Fig. 9A). Abundances of larger benthic foraminifera in excessof 10 vol.% are observed in samples containing up to 55 wt.% insolublematerial. The highest abundance of larger benthic foraminifera (up to60vol.%) is observed in samples fromSpain; all Java samples contain lessthan 28 vol.% larger benthic foraminifera.
Coralline algae are present in 138 of the studied samples, occurringin samples containing up to 90 wt.% insoluble material (Fig. 9B).Abundances of greater than 10 vol.% coralline algae are limited tosamples where the amount of insoluble material is 38 wt.% or less. Thehighest abundance of coralline algae (N40 vol.%) is observed insamples from Java; all samples from Borneo contain less than 25 vol.%coralline algae. Samples from Java contain coralline algae at thehighest concentrations of insoluble material (64 wt.% to 90 wt.%).
Corals are recorded from 113 of the samples analysed during thestudy (Fig. 9C). Only samples from Borneo contain coral where thequantity of insoluble material exceeds 37 wt.%. These corals areidentified as ‘first colonisers’ contributing up to 50 vol.% of samples inmuddy substrates containing up to 80 wt.% insoluble materials. Coralsaremost abundant in samples from Borneo and Spain (up to 58 vol.%);samples from Java exhibit a distinct paucity in corals with very fewoccurrences and no abundance exceeding 23 vol.%.
4.2. Relationship between siliciclastic grainsize and biota content
A wide range in siliciclastic grain sizes is observed in the threestudy areas (Fig. 10). The greatest distribution in grain size (clay tovery coarse sand grade) and the coarsest grains (1700 μm) areobserved in the volcaniclastic sediments contained within thecarbonate lithologies of the Wonosari Platform (Fig. 10A). Conversely,the deltaic clastic sediments of the Batu Putih Limestone contain thefinest grains and lowest range of grain sizes, from clay to fine sand(Fig. 10B). The siliciclastic material within the Collsuspina Limestonesranges in grain size between clay to coarse sand grade with finergrades (b300 μm) dominating (Fig. 10C). A distinct relationship isobserved between maximum siliciclastic grain size and the measuredabundance of coralline algae, larger benthic foraminifera and corals inthe three study areas (Fig. 11).
Fig. 6. Measured section (A) and graph (B) showing the relationship between carbonate com(A) is from the Calders 7 section and the total weight percent (carbonate and non-carbonshowing the relationship between the principal carbonate components and the total weightthe carbonate components does not reach 100% the difference is due to a combination of n
Coralline algae are present in 106 of the samples subjected to grainsize analysis (Fig. 11A). In these samples themajority of coralline algaeoccurrence (82%) is in samples that have coarsest median siliciclasticgrain sizes of fine sand grade or finer. The remaining 18% ofoccurrences are within samples where coarsest median grain sizesare between 300 μm to 890 μm (medium to coarse sand grade).Coralline algae contributing more than 10% of the sample volumewere observed in samples with maximum median grain sizes up to580 μm (coarse sand grade), the coarsest grained samples containedonly small (b2.3 vol.%) amounts of coralline algae.
Larger benthic foraminifera were identified from 126 of theanalysed samples (Fig. 11B). In total, 85% of samples containing largerbenthic foraminifera have maximummedian siliciclastic grain sizes offine sand grade or smaller. Larger benthic foraminifera contribute upto 36 vol.% in samples with maximum median siliciclastic grain sizesup to 140 μm (fine sand grade). The coarsest samples with largerbenthic foraminifera are of coarse sand grade (970 μm). An abundanceof in-situ larger benthic foraminifera of N10 vol.% is only observed insamples where maximum median grain sizes are less than 520 μm(coarse sand grade).
Corals have been observed in 86 of the samples that were analysedfor maximummedian siliciclastic grain size (Fig. 11C). Of these samples94% have maximum grain sizes of fine sand grade or less with coralscontributing more than 50 vol.% to samples with grain sizes of up to180 μm. Corals were not observed in samples where the maximummedian siliciclastic grain size exceeded 560 μm (coarse sand grade).
4.3. Overall relationship between dominant carbonate producers andsiliciclastic sediment
Larger benthic foraminifera, coralline algae and the three principalgrowth forms of corals (branching, platy, massive) all display arelationship to siliciclastic sediment grain size and the quantity ofinsoluble sediment present in the samples (Fig. 12). Larger benthicforaminifera are themost siliciclastic-tolerant of the groups of carbonateproducing organisms observed in this study (Fig. 12A). In total, 93samples contain a total volume of larger benthic foraminifera of 3 vol.%or greater. These samples have siliciclastic grain sizes up to 880 μm(coarse sand grade) and contain up to 76.4 wt.% insoluble sediment.Coralline algae are found at abundances of greater than 3 vol.% in a totalof 80 samples (Fig.12B). These samples have insolublematerial contentsup to 60wt.% andmedian largest siliciclastic grain sizes of up to 580 μm(coarse sand grade). Branching coral morphologies account for 55% ofthe samples containing corals. Branching corals are observed in samplescontaining up to 55 wt.% insoluble sediment and with siliciclastic grainsizes of up to 560 μm (Fig. 12C).
ponents and weight percentage non-carbonate material from Spain. Measured sectionate material) is compared with the relative abundances of biota. B is a quantified plotpercentage of non-carbonate material for all Calders sections. In plot B where the sum ofon-carbonate material and fine-grained, fragmented bioclastic material.

Fig. 8. Percentage of samples dominated by identified biota types in the three studyareas (where biota contribute 2% or greater of the volume of the sample).
Fig. 7. Representative field photographs of the three dominant carbonate producingbiota in the study. (A) larger benthic foraminifera in a crystal tuff (Java) containing up to70% insoluble volcaniclastic material, (B) complex spheroidal columnar rhodolithincorporating volcaniclastic sediments and encrusting larger benthic foraminiferawithin growth layers (Java), (C) a platy coral sheetstone from the base of a patch reefcontaining about 60% insoluble fine grained siliciclastics (Borneo).
34 S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
Platy coral morphologies are the second most common morphol-ogies observed during the study, accounting for 29% of coraloccurrence (Fig. 12D). These corals exhibit a high upper tolerancelimit to insolublematerial volume (80 wt.%). However, platy corals areonly found in samples where siliciclastic grain sizes are of fine sandgrade (250 μm) or less.
Massive corals account for 12% of the growth forms observed in thisstudy. Massive corals only occur in samples where the total amount ofinsolublematerial is less than37.9wt.% and the coarsestmediangrainsizeof siliciclastic material is of very fine sand grade or less (Fig. 12E).
Solitary corals are onlyobserved in samples from theBorneo sections(Fig. 12F). These corals are the least abundant in the study, onlycontributing 2% of all recognised coral morphologies. Solitary corals areobserved in samples containing up to 70 wt.% siliciclastic sediment butonly where median siliciclastic grainsize is silt grade or less.
5. Discussion
This study demonstrates that there are clear relationships betweenthe quantity and grainsize of siliciclastic or volcaniclastic sedimentwithin samples and the volume and nature of the dominant groups ofcarbonate-producing organisms. However, variations in biota-clasticinteractions between the three study areas are also apparent. Theserelationships and disparities are discussed below for each of the threeprincipal groups of carbonate producing organisms.
5.1. Environmental effects of silici/volcaniclastic sediment influx
An influxof siliciclastic sediment to a normally carbonate dominatedmarine system results in a change in thedepositional environment andaconsequent change in ecological niches. Where the rate of thissiliciclastic sediment influx is high then there is a risk of physical burialofmarine organisms. Episodes of rapid siliciclastic influx are a particularrisk for sessile benthic organisms as they are unable to extricatethemselves from the sediment. Where siliciclastic sediments aredeposited in relatively high-energy environments then externalprocesses, such as strong currents or wave activity, may resuspendsediments to exhume buried organisms and, in so doing, increaseturbidity (c.f. Woolfe and Larcombe, 1998; Larcombe andWoolfe, 1999;Wolanski et al., 2005). Under such conditions, organisms that are able totolerate short periods of burial are most likely to survive.

Fig. 9. Relationship between the total weight percentage of insoluble material andpercentage of (A) larger benthic foraminifera, (B) coralline algae, and (C) coral withinthe samples. Shaded areas denote samples where the amount of measured biota wasless than 10 vol.% of the sample.
35S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
The non-carbonate content of deposits from the Javan and Spanishsites varied from minimal (b1%) to N50% and abrupt shifts in clasticsedimentation with an organism response is inferred. In the Borneoexample, all deposits contain ~10% fine-grained siliciclastic or greaterand the biota appears adapted to background clastic influx (e.g. veryfew Acropora coral were present: a common clear-water species inIndonesia, Tomascik et al., 1997). Despite this, changes in biota arenoted associated with changing siliciclastic or organic matter contentin the deposits (Wilson and Lokier, 2002; Wilson, 2005).
At lower siliciclastic sediment concentrations organisms are still atrisk of being smothered, either in terms of their ability to photo-synthesise or due to feeding mechanisms becoming physically blockedor damaged (Woolfe and Larcombe, 1999). Many sessile benthicorganisms are able to clear small amounts of sediment from theirsurfaces (Rodgers, 1983; Peters and Pilson, 1985; Tomascik et al., 1997;Gleason, 1998; Van Der Zwaan et al., 1999; Rosen et al., 2002; Sandersand Baron-Szabo, 2005; Lirman and Manzello, 2009). However, ifsediment accumulation exceeds the organisms cleaning capacity thenmortality will ensue (Rodgers, 1983; Philipp and Fabricius, 2003; Perry,2005).
Where siliciclastic sediments are dominated by silt to clay gradeparticles, their suspension in the water column will increase turbiditywith a resultant reduction in the depth of the photic zone. This processwill be particularly significant in high-energy shallow-water environ-ments where currents and wave action will resuspend fine-grainedsediments (c.f.Woolfe and Larcombe,1998; LarcombeandWoolfe,1999;Wolanski et al., 2005). Over time, the subaqueous weathering ofvolcaniclastic minerals will produce significant quantities of clays thatwill also be susceptible to suspension and increase turbidity.
An influxof siliciclastic sedimentwill typically result in an increase inavailable nutrients in the marine environment. Where such an influxresults in themortalityofmarineorganisms then their decaying remainswill contribute additional nutrients to the trophic resources. Whereherbivore activity is high, corals dominate oligotrophic settings whilecoralline algae dominate under eutrophic conditions (Littler et al.,1991).A small increase in nutrient availability has been documented asresulting in an increase in coral tissue and skeletal growth (Tomasciket al., 1997; Edinger et al., 2000; Lough and Barnes, 2000). However, alarger shift in nutrient availability will either result in a change in thecommunities in the marine environment (Hallock et al., 1988, Perrinet al., 1995; Hallock, 2000) or a change in themode of feeding (Anthonyand Fabricius, 2000; Larcombe et al., 2001). An increase innutrientsmayalso produce a bloom of planktonic organisms that may provide a foodsource forotherorganisms. Suchaneventmayalso result in a short-termreduction in the depth of the photic zone (Hallock and Schlager, 1986;Zingonea and Enevoldsen, 2000; Mutti and Hallock, 2003; Okey et al.,2004) and the onset of anoxic bottom conditions (Wu, 1999; Hallock,2005), both potentially resulting in mass-mortality at the sea floor(Thangaraja et al., 2007).
5.2. Larger benthic foraminifera
Overall, larger benthic foraminifera are the most siliciclastic-tolerant of the carbonate-producing biota observed in the study. Theseorganisms occur in samples containing more than 95 wt.% insolublematerial and at grain sizes up to 970 μm. In some cases larger benthicforaminifera were the only biota within volcani/siliciclastic-richsamples. Their abundance may be attributed to a lack of competitors(or predators) in a stressed environmental setting, an ability tosurvive burial for a short period of time (Van Der Zwaan et al., 1999),to excavate themselves following shallow burial and an ability tocolonise barren substrates (Alve, 1999).
In total, 9 genera of larger benthic foraminifera were identifiedfrom the lower to middle Miocene Indonesian sections while 7 generawere documented from the middle to upper Eocene Calders section.These relatively high generic diversities may reflect deposition during

Fig. 10. The distribution of median grain sizes of the coarsest siliciclastic grains.
36 S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
two of the four Cenozoic periods of peak larger benthic foraminiferadiversity (Renema, 2002).
Larger benthic foraminifera are the dominant carbonate producingorganisms in samples from the Wonosari Platform and Calderssections. Although also common in the Borneo sections, they are
secondary to corals as carbonate contributors and are concentrated inreef-capping larger benthic foraminifera packstones. These organismshost photosynthetic symbionts within their tests (Hallock and Glenn,1986), to maintain these symbionts the test surface must be kept clearof sediment. Larger benthic foraminifera are able to clean smallamounts of sediment from their surface (Van Der Zwaan et al., 1999);allowing them to survive small episodic influxes of siliciclasticsediments. However, an increase in the duration or quantity ofsiliciclastic influx will result in a corresponding increase in the stressto larger benthic foraminifera. In both Java and Spain, siliciclasticsediment influx is interpreted to have been episodic. Conversely, inBorneo siliciclastic supply appears to have been more persistent witha near-constant influx to the marine system. The presence of reef-capping larger benthic foraminifera packstones is inferred to beassociated with the development of seagrass (Wilson, 2005).
Where siliciclastic sediment influx is very high this would result inphysical burial of the larger benthic foraminifera. A degree of mobilityallows many species to extricate themselves if buried beneath smallamounts of sediment. However, a large sediment influx, associatedwith episodic processes such as volcanic eruptions, would result inrapid burial and mortality. Just such a mass-mortality of benthicforaminifera occurred following the deposition of only 2–6 cm ofvolcanic ash in the South China Sea during the 1991 eruption ofMt. Pinatubo (Alve, 1999). A similar scenario is hypothesised for anumber of the thicker volcaniclastic units in the Wonosari area thatcontain escape burrows, but lack larger benthic foraminifera (Lokier,2000).
The presence of larger benthic foraminifera as the principal biota insediments succeeding the thicker volcani/siliciclastic units is con-sistent with their ability to rapidly colonise new habitats. Suchcolonisation is primarily achieved via transport in suspension duringthe planktonic stage of reproduction. Passive entrainment, duringhigh-energy events, such as storms, can also result in the transporta-tion of more mature individuals over considerable distances (Alve,1999). Rapid colonisation of a new or stressed habitat allows anorganism to benefit from available resources without the risk ofcompetitors or predators. A relatively low abundance of larger benthicforaminifera in the clay-rich basal sheetstones of the Borneo sectionsmay reflect an inability to survive where sediments probably had a‘soupy’ character.
An observed dominance of thinner, large-diameter larger benthicforaminifera tests in volcaniclastic and siliciclastic sediments isconsistent with a reduction in incident light (Hallock and Glenn,1986). This is inferred to be associated with a decrease in the depth ofthe photic zone due to increased turbidity from suspended finegrained sediment.
Increased larger benthic foraminifera abundance in samples withmaximummedian siliciclastic grain sizes of fine sand grade or smallerand an absencewhere grains are of very-coarse sand grade or higher isinferred to result from the increased probability of abrasion-damageby coarser grains.
Samples from the Calders section contain the largest volumes oflarger benthic foraminifera and a diverse coral fauna. This may reflectoligotrophic conditions (Hallock and Glenn, 1986) and relativelystable substrates in this setting.
5.3. Coralline algae
Coralline algae are identified in samples containing in excess of90 wt.% insoluble material and at grain sizes up to 890 μm. Thepresence of siliciclastic grains between the growth layers of corallinealgae proves that their development was penecontemporaneous withsiliciclastic sedimentation.
Coralline algae must remain within the photic zone if they are tocontinue to develop. If buried then photosynthesis will cease andmortality will ensue. In moderate to high-energy settings, buried

37S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
algae may be exhumed and growth reinitiated either by the samespecies or by a completely new species of coralline algae (Bosence,1983; Marshall et al., 1998).
The high volumes of coralline algae (and paucity of corals) insiliciclastic-rich samples from Java are consistent with deposition in ameso/eutrophic environment with unstable substrates and moderateenergy regimes. Under such conditions the unstable, regularly shiftingsubstrate would facilitate the development of concentric rhodolithsand assist in keeping the algal surface free of sediment. Rhodolithformation can be promotedwhere broken fragments of coralline algaebecome detached through physical or biological processes butcontinue to grow (Bosence, 1983). A modern analogue for theWonosari Formation is at Banua Wunu on the Sangihe Ridge northof Sulawesi. Here, the summit of a submarine volcano lies at a depth of6–10 m and is dominated by coralline algae, bryozoans and sponges(Tomascik et al., 1997) existing on an unstable substrate with currentvelocities up to 4 m/s. Coralline algae also dominate on the high-energy mixed carbonate–clastic shelf offshore NE Brazil (Testa andBosence, 1998).
Coralline algae are observed in samples withmaximum siliciclasticgrain sizes of clay to silt grade in samples from Calders and Borneo.Such an occurrence is perhaps surprising as fine-grained sedimentsare particularly susceptible to re-suspension, thereby enhancingturbidity and reducing the depth of the photic zone. However,coralline algae are tolerant of low light levels (Wilson and Vecsei,2005) and in both these sections, only developed in shallow waters ofless than 10 m depth. Further, clay to silt grade sediments are unlikelyto provide a firm substrate on which coralline algae growth may beinitiated.
In the Calders sections most of the coralline algae occur as thinencrusting forms associated with corals. Such growth forms arenormally associated with deeper water settings (Steneck, 1986) andare here interpreted to result from a turbidity-induced reduction inthe depth of the photic zone. In areas where fine-grained sedimentsdominate, corals may provide the only firm substrate on whichcoralline algae growth can be initiated. This raises the issue of how thecorals themselves could have become established in such areas. Twopossibilities arise: 1) Initiation of coral growth during episodes ofreduced fine-grained siliciclastic influx. 2) Transport of living coralsfrom another setting during high-energy events such as storms. Inboth cases, as long as coral growth equals or exceeds the rate ofrenewed sediment accumulation then the corals and their associatedencrusting organisms would survive. The encrustation of corals alsoresults in the coralline algae occupying an elevated bathymetricposition, thus reducing the effects of turbidity and sediment abrasion.
In the Borneo sections coralline algae are less common than in theother two study areas and, where they do occur, are always found inassociation with corals. Such a relationship may reflect the limitedavailability of a stable substrate in the fine-grained clay and silt-richsediments of this area. The low-energy conditions inferred for theBorneo sections would also have a negative impact on the developmentof coralline algae. Whereas corals and larger benthic foraminifera areable, to some extent, to clean themselves of small amounts of siliciclasticsediment, coralline algae must rely on currents to remove sediments.Thus, in low-energy environments, even quite small influxes of fine-grained sediments may be sufficient to terminate coralline algaedevelopment. Where sediment influx is relatively constant a lack ofcurrent activity would prevent the development of a stable substrateand inhibit recolonisation. An association between coralline algae and
Fig. 11. Relationship between the median size of the largest grains and volumepercentage (vol.%) of; (A) coralline algae, (B) larger benthic foraminifera, and (C) coralwithin the samples. Shaded area denotes grain sizes where measured biota is less than10% of the volume of the sample. Note: Any samples plotted at a grain size of 0 indicate alack of siliciclastic grains within the sample.
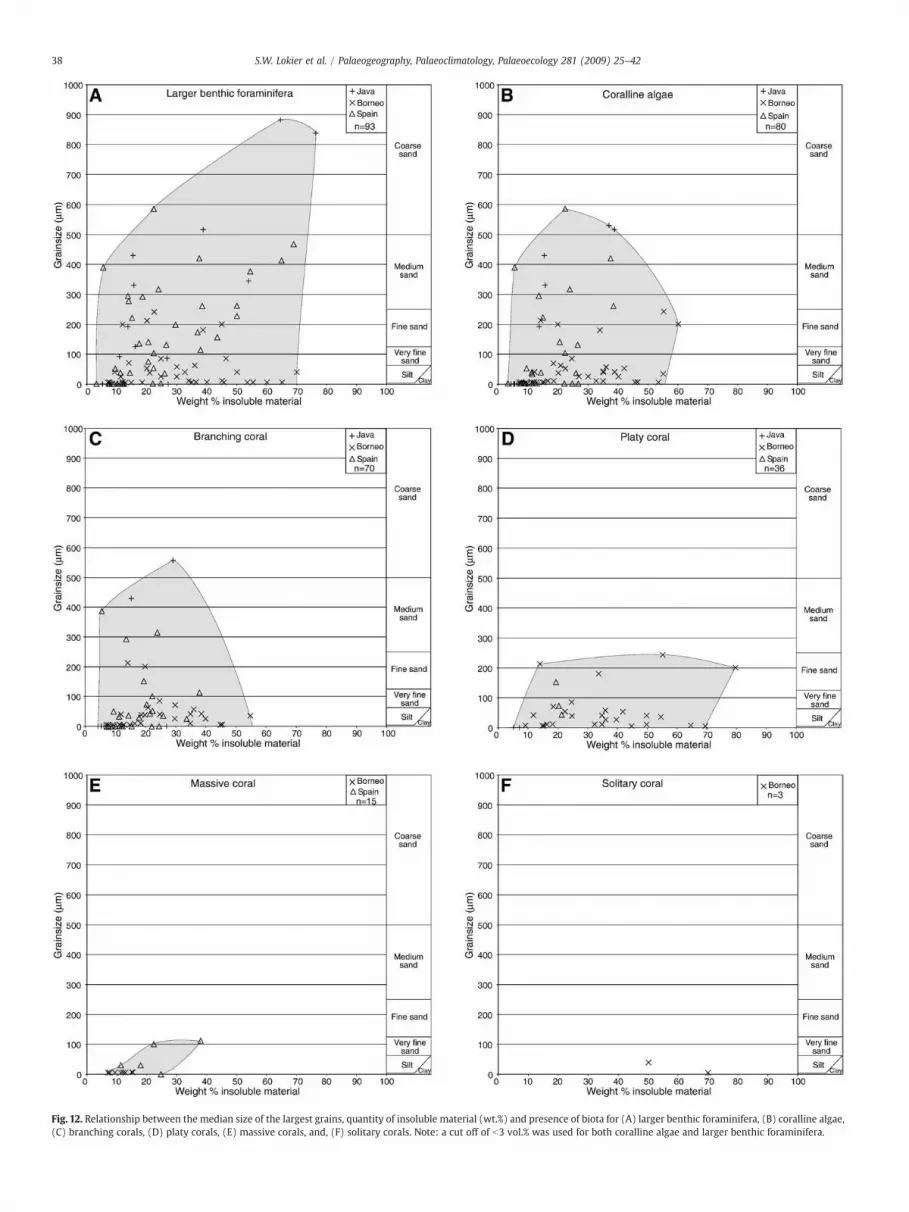
Fig. 12. Relationship between the median size of the largest grains, quantity of insoluble material (wt.%) and presence of biota for (A) larger benthic foraminifera, (B) coralline algae,(C) branching corals, (D) platy corals, (E) massive corals, and, (F) solitary corals. Note: a cut off of b3 vol.% was used for both coralline algae and larger benthic foraminifera.
38 S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42

39S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
increased carbon content (up to 4%) may relate to increased growthrates of coralline algae associated with nutrient influx.
5.4. Corals
Of the three dominant carbonate-producing organisms, coralsexhibit the largest variability in occurrence between the three studyareas. They are the dominant carbonate producing organisms in theBorneo sections, but are relatively rare in samples from Java. Coralswere identified from sediments containing up to 80 wt.% (althoughmost were b55 wt.%) insoluble material and at maximum siliciclasticgrain sizes up to 560 μm.
Most high-diversity coral communities develop on firm substrates,under oligotrophic conditions in the upper part of the photic zone. Therelative scarcity of corals in the samples from the Wonosari Formation(b7%of samples contain coralswith abundances not exceeding23vol.%)is interpreted to result from the lack of a stable substrate, nutrient inputand from the amount and nature of volcaniclastic sediment input(Lokier, 2000). Most corals are sessile benthic organisms and, as such,are particularly at risk of burial. During burial, corals are susceptible toabrasion of soft tissues, particularly fromgrain sizes larger thanmediumsandgrade (Woolfe and Larcombe,1999). If the sediment is not removedfrom the coralwithin a fewhours thenphysical smotheringwill result indeath of the coral polyps (Heikoop et al., 1996; Scoffin,1997; Barnes andLough, 1999; Woolfe and Larcombe, 1999). Ingestion of angularvolcaniclastic grains, known to be damaging to polyps (James andKendall, 1992), may also hinder their development in the WonosariFormation.
At Manado, Sulawesi, Pleistocene coral biostromes were not re-established following deposition of a fine-grained tuff (Heikoop et al.,1996). In the Burdigalian of Iraq volcaniclastic sedimentation resultedin the cessation of coral development and a switch to coralline algaedominated sedimentation (Okhravi and Amini, 1998). Over 100 yearsafter the 1883 Krakatau eruption coral communities are still marginaldue to a lack of stable substrate (Tomascik et al., 1997). An abundanceof unconsolidated, moving rubble appears to inhibit colonisation(Sheppard et al., 2002), although if stabilised, rubble can be asubstrate for coral colonisation (Braga et al., 1990).
Solitary fungiid corals are common in the lowermost horizons ofvolcaniclastic sequences in the Wonosari Formation. The fungiids areefficient at cleaning their surfaces (Heikoop et al., 1996), canwithstandrelatively high sedimentation rates and are able to excavate themselvesif buried to a shallow depth (Goreau and Younge,1968; Hubbard,1972).Similaroccurrence of fungiids is reported fromPleistocenevolcaniclasticdeposits from Manado, Sulawesi (Heikoop et al., 1996).
There is a distinct relationship between coral growth morphologyand the volume and grain size of siliciclastic sediment, with most datafrom the Calders and Borneo sections (coral abundances up to 58%).Platy, branching and massive corals are present when wt.% insolubleswere b80% (fine sand or less), b55% (mainly b40%, medium sand orless) and b40% (mainly b25%, very fine sand or less), respectively.
In Calders, a diverse range of corals are very important in carbonateintervals but largely absent from siliciclastic intervals. The siliciclasticsare associated with moderate to high energies and shifting unstablesubstrates due to localised currents and siliciclastic sources along theshoreline (Burton, 3003). These conditions inhibit settling and nuclea-tion of larval corals (Hodgson,1990;Woolfe and Larcombe,1998,1999).The diversity in the Calders reefs (~10 genera), their development inshallow water, together with minimal carbon and low fine grainedsiliciclastic content for domestones (10% or less) is suggestive ofpredominantly oligotrophic conditions for the carbonate-rich intervals.Any sandy influx during periods of massive coral growth was limited tomaterial of very fine sand grade or less, that accumulated mainlybetween corals (surface clearing by polyps may have enhanced thisaccumulation). Corals present in lithologies containing 25–b40 wt.%fine-grained, non-carbonate sediment have delicate branching
morphologies. Steep upright surfaces on branching corals are lesssusceptible to sediment accumulation than planar or concave surfaces(Scoffin,1997; Sanders and Baron-Szabo, 2005). Corals adopt branchingmorphologies where sedimentation or competition for space is high(Grasso and Pedley, 1988). Platy coral development is evident in thesecond carbonate interval within lithologies containing up to 26 wt.%non-carbonate. With delicate coral growth forms and predominantlyclay-sized siliciclastic content it is inferred that the second intervaldeveloped under more turbid conditions, with re-suspension of finesediment and lower net sedimentation rates. In turbid waters, lightpenetration decreases and corals tend to adopt flattened plate-likemorphology that receive more incident light than vertical surfaces(Graus and Macintyre, 1976, Titlyanov and Latypov, 1991).
In the Borneo sections, generic diversity is moderate (at least 10genera; Wilson, 2005), and clay-rich or fine grained siliciclastics arepresent throughout (10–80% wt.%). These delta-front reefs areinterpreted to have developed under conditions of near-constantsiliciclastic input and oligo- to mesotrophy (Wilson, 2005). Coraldiversity is similar to other turbid-water reefs (similar to, or up toaround 2/3 that of, clear water reefs: Larcombe et al., 2001; Sandersand Baron-Szabo, 2005). As sediment input increases, in this low-energy setting growth forms respond to turbidity and available lightwith a lateral and vertical shrinkage of the patch reef and a change tolower light-level tolerant branching and then platy corals (Wilsonand Lokier, 2002; Wilson, 2005). Many of the corals present in theassemblage, such as Porites and Goniopora, are known for theirsediment-shedding ability and have been documented in low-energy, turbid environments today (Tomascik et al., 1997). Acro-pora, one of the most abundant corals in modern SE Asian reefs, israre, perhaps because of a preference for moderate to high-energy,generally clear-water settings (Tomascik et al., 1997).
Platy Leptoseris and Pachyseris are abundant in the initial, terminaland marginal, mud-rich (40–80 wt.% insolubles) patch reef deposits.These are common early colonisers in Indo-Pacific reefs followingenvironmental disturbance (Tomascik et al., 1997), efficient self-cleaners and can also change to more heterotrophic feeding (Insalaco,1996). A number of studies fromSEAsia andAustralia have shown reefaldevelopment over soft or sandy substrates (Umbgrove, 1947; Tudhopeand Scoffin, 1994; Tomascik et al., 1997; Hopley et al., 2007).Mechanisms for coral colonisation of soft substrates include settlingand growth of planulae on rock or bioclastic fragments (Tomascik et al.,1997), or through laminar growth forms spreading out over thesubstrate (Martin et al., 1989). In life at least some of the platy coralsgrew projecting from the sediment, as evidenced by encrustation ontheir underside. Angled, tiered, or whorled growth forms of platy coralsfacilitate the shedding of sediment (Rosen et al., 2002). Increasednutrient availability may be the explanation for the positive correlationbetween increased organic matter, siliciclastic content, increase inheterotrophs (e.g. Tubastrea), degree of bioerosion and encrustation byalgae and infaunal suspension feeders (Hallock and Schlager, 1986;Edinger et al., 2000) in the upper parts of the patch reefs.
5.5. Sedimentation rates and volumes
Survivorship of organisms in areas of clastic input is determined bythe volume and rate of sedimentation versus the growth rates andability to self-clean or extract. Although the geologically averagedrates of clastic influx are difficult to compare directly with the dailyand seasonal fluctuations experienced by modern systems, this studymay aid understanding of increased clastic influx over longer timescales. Foraminifera, having a degree of mobility, can self-extract froma thin layer of sediment covering. However, deposition of 2–6 cm ofash in the South China Sea during the 1991 Mt. Pinatubo eruptionresulted in mass mortality (Alve, 1999).
Average growth rates for modern larger benthic foraminifera varyfrom1.5 (Amphistegina), 3 (Marginopora), up to 4mm/yr (Heterostegina),

40 S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
and their longevity is typically 1–2 years (Hallock,1985). Growth rates forextinct foraminifera are more uncertain with estimates for equatorialgrowth rates of b1.5 mm/yr for Eocene Nummulites (Purton and Brasier,1999). However, Sarangi et al. (2001) have suggested growth rates for‘giant’ Eocene forms (up to 16 cm)were significantly higher, and disagreewith suggested longevity of up to 100 yrs (Purton and Brasier, 1999).Wilson (2000) suggested that thresholds for foraminiferal accumulationwere exceeded when sustained sediment influx equalled or exceededgrowth rates. Using the calculated growth rates (above) of Amphisteginaor Nummulites threshold rates for sedimentation resulting in demisewould be 1.5 m/ky. However, since the nummulitids accrete up to 10timesmore around their diameter than their thickness, this is likely to bean overestimate (perhaps by an order of magnitude). Accumulation onthe larger benthic foraminifera dominated Tonasa Platform in Indonesiaceased when sustained volcaniclastics influx exceeded 40–50 vol.%, i.e.when the calculated carbonate accumulation rates of between to 0.07–0.3 m/ky were exceeded (Wilson, 2000). Kumar and Saraswati (1997)also inferred that Lepidocyclina, Heterostegina and Operculina can survivewithin sediments with ≤40 vol.% siliciclastics, whereas Miogypsina cantolerate higher levels without detrimental effects. In the Calders sectionNummulitesandDiscocyclinawereable to survive in sediments containing65 wt.% siliciclastics.
Coralline algae cannot self-extract if coveredby sediment. However, ifremoved from light for a few months (due to overturning, or polarwinters) growth may continue slowly and/or resume if returned to thelight (Testa, pers. comm. 2008; Freiwald and Heinrich,1994). If currentsclear sediment from the surface of coralline algae growthmay resume ifburial was short, or the surface could be colonised by new growth ofcoralline algae. Due to this reinitiated growth, rhodolithsmay form overperiods of 100s to 10,000s of years (Bosence, 1983). On shorter time-scales, growth rates in rhodolith-formingbranching corallines have beenreported at0.1–1mm/yr for temperate settings off IrelandandNWSpain(Lithothamnion, Phymatolithon; Bosence and Wilson, 2003), 1.1 mm/yrin an area of polar upwelling off Norway (Freiwald and Heinrich, 1994)and 5 mm/yr for tropical Florida (Spongites; Bosence et al., 1985). Forrhodolith-rich maerl environments long-term accumulation rates ofbetween 0.08 to 1.4 m/ky were similar for temperate and tropicalsettings due to higher densities in temperate and nutrient-rich settings(Bosence andWilson, 2003). On the basis of these figures coralline algaeare likely to be able to withstand a similar order of magnitude rate ofsedimentation (up to 1.4 m/ky, but mostly b0.7 m/ky) as larger benthicforaminifera. The very similar co-occurrence patterns of wt.% corallinealgae and larger benthic foraminifera versus wt.% insoluble is suggestiveof similar resistance to comparable clastic influx. In theWonosari sectioncoralline algae appear more resilient to clastic influx, perhaps reflectinghigher growth rates in an area of nutrient upwelling. In comparison,larger benthic foraminifera show higher vol.% occurrence in the Calderssection where the conditions are inferred to have been moreoligotrophic.
With the exception of solitary forms (Goreau and Younge, 1968;Hubbard, 1972), corals are sessile and are reliant on either self-cleaning, sediment shedding or non-threshold levels of sedimentationfor survival. Wesseling et al. (1999) demonstrated that, while speciesof some coral genera (e.g. Porites, Galaxea and Heliopora) can surviveepisodic burial for up to 20 h, species of other genera (e.g. Acropora)will not recover. For the different growth forms massive, branchingand platy corals were generally not present when insoluble contentexceeded N25%, N40% and N80% respectively (Rodgers, 1983; Stafford-Smith and Ormond,1992;Wilson and Lokier, 2002). These values mayreflect an inability of the corals to self-clean, a reduction in light levels,an increase in nutrients, or an increase in abrasion or bioerosion ratherthan an inability to keep pace with sedimentation. Growth rates formassive corals are 5–10 mm/yr, up to 100–200 mm/yr for branchingcorals and up to 4 mm/yr for platy corals (James and Kendall, 1992;Insalaco, 1996; Edinger et al., 2000). No clear relationship is seenbetween possible growth rates and ability to survive sedimentation
for the different growth forms. However, not all of these rates areapplicable for sediment influenced reefs, particularly some of the highgrowth rates reported for branching corals, such as Acropora. Althoughvariable, some growth rates may decline in sediment-impacted reefs(Insalaco, 1996; Woolfe and Larcombe, 1999). In other cases a smallincrease in nutrients may enhance skeletal growth (Tomascik et al.,1997; Lough and Barnes, 2000) while a larger increase may promotebioerosion (Edinger et al., 2000).
6. Conclusions
Major shallow-water carbonate producing biota of larger benthicforaminifera, coralline algae and corals were found to be variablytolerant to clastic influx. Although mollusc and echinoderm debriswere also present in the mixed carbonate–clastic units they were notmajor sediment contributors. The three Cenozoic sections fromBorneo, Java and Spain allow the influence of siliciclastic orvolcaniclastic influx on shallow-water carbonate producers to bequantifiably assessed:
• Age — Within the Cenozoic there was no discernable variation intolerance of the main groups of organisms (corals, coralline algaeand larger benthic foraminifera) to insoluble clastic influx related todiffering ages of the deposits. Instead, survivorship of organisms toinsoluble material was dependent on local environmental condi-tions, likely including rate of influx, grain-size, nutrient influx,energy and water depth,
• Amounts of clastic influx — Larger benthic foraminifera and corallinealgae were the most resistant groups to clastic influx still presentwhen wt.% were up to 70% and 60%, respectively. Although platycoral development occurred in deposits where wt.% insolublematerial was up to 80%, this was only where grain sizes weredominated by clay, and no larger than fine sand grade. Branchingand massive corals were respectively the least tolerant of insolubleinflux, present when wt.% were less than 55% (usually b40%) andb40% (usually b25%) respectively. Modern studies indicate burialunder a few cm of sediment generally results in mortality, howeversome corals can recover if cleared after a few hours of burial andcoralline algae after a few months.
• Rates of influx— Both larger benthic foraminifera and coralline algaeare tolerant of sustained rates of clastic influx around 1 mm/yr overshort time-scales, and perhaps up to 1.5 m/ky (although generallyb0.7 and b0.3 m/ky, respectively) over longer time-scales. Thethresholds for foraminiferal and coralline algae accumulation weregenerally exceeded when sediment accumulation equalled orexceeded growth rates. There was no clear correlation betweencoral growth rates and survivorship under different rates of influxfor the different morphologies.
• Grain size— Larger benthic foraminifera and coralline algaewere themost tolerant to clastic influx with varying grain size (up to coarsegrain-size). This probably reflects the ability of the larger forami-nifera to self-extract and self clean if covered. The coralline algaelack soft parts prone to damage, and may grow to incorporatesediment within their structure (rhodoliths). The corals were lesstolerant of coarse grain sizes and platy and massive corals were onlypresent when grain sizes were less than fine sand, or very fine sand,respectively. Coarser grains may settle out between branchingcorals, whereas in clay-rich settings, high turbidity and correspond-ing reduced light levels may promote platy growth forms.
• Other environmental influences— Other factors such as energy, waterdepth, nutrients, turbidity, light penetration (many linked to clasticinflux) will all determine local environmental conditions andcarbonate production. Nutrient influx may increase the toleranceof coralline algae to clastic influx if it results in an increase in growthrates. Conversely the tolerance of larger benthic foraminifera orcorals may be reduced under increased nutrients associated with a

41S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
change to more eutrophic conditions and higher bioerosion. Withincreasing turbidity, associated with fine grain influx, light penetra-tion decreases and a change to flatter morphologies reflects a needtomaximise light collection. This morphological changemay only bepossible if energies at the specific water depth are insufficient tocause breakage of more delicate growth forms. Moderate to highenergies may increase the survivorship of organism, such ascoralline algae, which are reliant on external factors to clearsediment from growth surfaces.
• Siliciclastic versus volcaniclastic influx — Influx of volcaniclastic andsiliciclastic material of comparable grain sizes appear to have similareffects on many carbonate producers, although corals may be moresusceptible to damage by angular volcanic shards.Wilson and Lokier(2002) have outlined differences in the platform context withfringing reefs around volcanoes or offshore platforms isolated fromsiliciclastic runoff but affected by punctuated volcaniclastic influx.
Acknowledgements
This research was undertaken at Royal Holloway College, London, aspart of PhD (S.W.L., Java) and postdoctoral (M.E.J.W., Borneo) studieswith the SE Asia Research Group. The research on Spainwas funded as aNERC (Award GT 04/99/ES/59) PhD studentship (L.M.B), when L.M.Band M.E.J.W were based at Durham University, UK. We gratefullyacknowledge the fundsand support of these institutes andorganisationsaswell as LASMORuntu, Unocal, AAPG and the Geological Research andDevelopment Centre, Indonesia for making fieldwork possible. BrianRosen, Marcelle Fadel, Esmerelda Caus and Ted Finch are thanked fortheir considerable help with coral, foraminifera and nannofossil workrespectively. Colleagues thanked for significanthelpduring this researchincludeDanBosence, Robert Hall, Diane Cameron, FauzieHasibuan, JohnChambers, Harry Alam, Viviane Testa,Maurice Tucker, Conxita Taberner,Jim Hendry and Dave Scott. Thierry Corrège and two anonymous re-viewers are thanked for their constructive comments on themanuscript.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.palaeo.2009.07.007.
References
Allen, G.P., Chambers, J.L.C., 1998. Sedimentation in the Modern and Miocene MahakamDelta. Indonesian Petroleum Association, Jakarta, Indonesia. 236 pp.
Allen, G.P., Laurier, D., Thouvenin, J., 1976. Sediment distribution patterns in the modernMahakam Delta: 5th Annual Convention of the Indonesian Petroleum Association,Proceedings, pp. 159–177.
Alve, E., 1999. Colonization of new habitats by benthic foraminifera: a review. Earth-Science Reviews 46, 167–185.
Anthony, K.R.N., Fabricius, K.E., 2000. Shifting roles of heterotrophy and autotrophy incoral energy budgets under varying turbidity. Journal of Experimental MarineBiology and Ecology 252, 221–253.
Barnes, D.J., Lough, J.M., 1999. Porites growth characteristics in a changed environment:Misima Island, Papua New Guinea. Coral Reefs 18, 213–218.
Bosence, D.W.J., 1983. Description and classification of rhodoliths (Rhoids, Rhodolites).In: Peryt, T.M. (Ed.), Coated Grains. Springer-Verlag, Berlin, pp. 217–224.
Bosence, D.W.J., Wilson, J., 2003. Maerl growth, carbonate production rates andaccumulation rates in the northeast Atlantic. Aquatic conservation: Marine andFreshwater Ecosystems 13, 21–31.
Bosence, D.W.J., Rowlands, R., Quine, M.J., 1985. Sedimentology and budget of a Recentcarbonate mound, Florida Keys. Sedimentology 32, 317–343.
BouDagher-Fadel, M.K., Wilson, M.E.J., 2000. A revision of some larger Foraminiferafrom the Miocene of East Kalimantan. Micropaleontology 46, 153–165.
Braga, J.C., Martin, J.M., Alcala, B., 1990. Coral reefs in coarse-terrigenous sedimentaryenvironments Upper Tortonian, Granada Basin, southern Spain. SedimentaryGeology 66, 135–150.
Burton, L.M., 2004. Carbonate–siliciclastic Interactions; Tertiary Examples from Spain.PhD Thesis, University of Durham, UK, 396 p.
Cavagnetto, C., Anadon, P., 1996. Preliminary palynological data on floristic and climaticchanges during the Middle Eocene–Early Oligocene of the eastern Ebro Basin,northeast Spain. Review of Palaeobotany and Palynology 92, 281–305.
Doyle, L.J., Roberts, H.H., 1988. Carbonate–clastic Transitions. In: Developments inSedimentology, vol. 42. Elsevier, Amsterdam. 304 pp.
Edinger, E.N., Limmon, G.V., Jamaluddin, J., Widjatmoko, W., Heikoop, J.M., Risk, M.J.,2000. Normal coral growth rates on dying reefs: are coral growth rates goodindicators of reef health? Marine Pollution Bulletin 40, 404–425.
Freiwald, A., Heinrich, R., 1994. Reefal coralline algal buildups within the Arctic circle:morphology and sedimentary dynamics under extreme environmental seasonality.Sedimentology 41, 963–984.
Gleason, D.F., 1998. Sedimentation and distributions of green and brown morphs of theCaribbean coral Porites astreoides. Lamarck Journal of Experimental Marine Biologyand Ecology 230, 73–89.
Goreau, T.F., Younge, C.M., 1968. Coral community onmuddy sand. Nature 217, 421–423.Grasso, M., Pedley, H.M., 1988. The sedimentology and development of the Terravecchia
Formation carbonates (upper Miocene) of North Central Sicily: possible eustaticinfluence on facies development. Sedimentary Geology 57, 131–149.
Graus, R.R., Macintyre, I.G., 1976. Light control of growth form in colonial reef corals:computer simulation. Science 193, 895–897.
Hallock, P., 1985. Why are larger foraminifera large? Paleobiology 11, 195–208.Hallock, P., 2000. Symbiont-bearing foraminifera: harbingers of global change?
Micropaleontology 46, 95–104.Hallock, P., 2005. Global change and modern coral reefs: New opportunities to
understand shallow-water carbonate depositional processes. Sedimentary Geology175, 19–33.
Hallock, P., Glenn, E.C., 1986. Larger foraminifera: a tool for paleoenvironmental analysisof Cenozoic carbonate depositional facies. Palaios 1, 55–64.
Hallock, P., Schlager, W., 1986. Nutrient excess and the demise of coral reefs and thedemise of coral reefs and carbonate platforms. PALAIOS 1, 389–398.
Hallock, P., Hine, A.C., Vargo, G.A., Elrod, J.A., Jaap, W.C., 1988. Platforms of theNicaraguan Rise: examples of the sensitivity of carbonate sedimentation to excesstrophic resources. Geology 16, 1104–1107.
Harrell, J.A., Eriksson, K.A.,1979. Empirical conversion equations for thin-section and sieve-derived size distribution parameters. Journal of Sedimentary Petrology 49, 273–280.
Harwood, G., 1988. Microscopic techniques: II. Principles of sedimentary petrography.In: Tucker, M.E. (Ed.), Techniques in Sedimentology. Blackwell Scientific Publica-tions, UK, pp. 108–173.
Heikoop, J.M., Tsujita, C.J., Heikoop, C.E., Risk, M., Dickin, A., 1996. Effects of volcanicashfall recorded in ancient marine benthic communities: comparison of anearshore and an offshore environment. Lethaia 29, 125–139.
Hendry, J.P., Taberner, C., Marshall, J.D., Pierre, C., Carey, P.F., 1999. Coral reef diagenesisrecords pore-fluid evolution and paleohydrology of a siliciclastic basin marginsuccession (Eocene South Pyrenean foreland basin, northeastern Spain). GeologicalSociety of America Bulletin 111, 395–411.
Hodgson, G., 1990. Sediment and the settlement of larvae of the reef coral Pocilloporadamicornis. Coral Reefs 9, 41–43.
Hook, J., Wilson, M.E.J., 2003. Stratigraphic relationships of aMiocenemixed carbonate–siliciclastic interval in the Badak field, East Kalimantan, Indonesia. 29th IndonesianPetroleum Association, Annual Convention, Proceedings, pp. 147–161.
Hopley, D., Smithers, S.G., Parnell, K.E., 2007. The Geomorphology of the Great BarrierReef. Cambridge University Press. 518 pp.
Hubbard, J.A.E.B., 1972. Diaseris distorta, an ‘Acrobatic’ Coral. Nature 236, 457–459.Insalaco, E., 1996. Upper Jurassic microsolenid biostromes of northern and central
Europe: facies and depositional environment. Palaeogeography, Palaeoclimatology,Palaeoecology 121, 169–194.
Insalaco, E., 1998. The descriptive nomenclature and classification of growth fabrics infossil scleractinian reefs. Sedimentary Geology 118, 159–186.
James, N.P., Kendall, P., 1992. Introduction to carbonate and evaporite facies models. In:Walker, R.G., James, N.P. (Eds.), Facies Models: Response to Sea Level Change.Geological Association of Canada, Ontario, pp. 265–275.
Kumar, A., Saraswati, P.K., 1997. Response of larger foraminifera to mixed carbonate–siliciclastic environments: an example from the Oligocene–Miocene sequence ofKutch, India. Palaeogeography, Palaeoclimatology, Palaeoecology 136, 53–65.
Larcombe, P., Woolfe, K.J., 1999. Increased sediment supply to the Great Barrier Reef willnot increase sediment accumulation at most coral reefs. Coral Reefs 18, 163–169.
Larcombe, P., Costen, A., Woolfe, K.J., 2001. The hydrodynamic and sedimentary settingof nearshore coral reefs, central Great Barrier Reef shelf, Australia: Paluma Shoals, acase study. Sedimentology 48, 811–835.
Lewis, D.W., McConchie, D., 1997. Analytical Sedimentology. Chapman & Hall, London.197 pp.
Lirman, D., Manzello, D., 2009. Patterns of resistance and resilience of the stress-tolerant coral Siderastrea radians (Pallas) to sub-optimal salinity and sedimentburial. Journal of Experimental Marine Biology and Ecology 369, 72–77.
Littler, M.M., Littler, D.S., Titlyanov, E.A., 1991. Producers of organic-matter on tropicalreefs and their relative dominance. Biologiya Morya 6, 3–14.
Lokier, S.W., 2000. The Miocene Wonosari Formation, Java, Indonesia: VolcaniclasticInfluences on Carbonate Platform Development. PhD Thesis, University of London,UK, 648 p.
Lough, J.M., Barnes, D.J., 2000. Environmental controls on growth of the massive coralPorites. Journal of Experimental Marine Biology and Ecology 245, 225–243.
Marshall, J.F., Tsuji, Y., Matsuda, H., Davies, P.J., Iryu, Y., Honda, N., Satoh, Y., 1998.Quaternary and Tertiary subtropical carbonate platform development on thecontinental margin of southern Queensland, Australia. In: Camoin, G.F., Davies, P.J.(Eds.), Reefs and Carbonate Platforms in the Pacific and Indian Oceans. In:International Association of Sedimentologists. Special Publication No. 25. BlackwellScientific Publications, Oxford, pp. 163–195.
Martin, J.M., Braga, J.C., Rivas, P., 1989. Coral successions in Upper Tortonian reefs in SESpain. Lethaia 22, 271–286.
Milliman, J.D., Meade, R.H., 1983. Worldwide delivery of river sediment to the oceans.Journal of Geology 91, 1–21.

42 S.W. Lokier et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 281 (2009) 25–42
Mount, J.F., 1984. Mixing of siliciclastic and carbonate sediments in shallow shelfenvironments. Geology 12, 432–435.
Mutti, M., Hallock, P., 2003. Carbonate systems along nutrient and temperaturegradients: some sedimentological and geochemical constraints. InternationalJournal of Earth Sciences 92, 465–475.
Okey, T.A., Vargo, G.A., Mackinson, S., Vasconcellos, M., Mahmoudie, B., Meyer, C.A.,2004. Simulating community effects of sea floor shading plankton blooms over theWest Florida Shelf. Ecological Modelling 172, 339–359.
Okhravi, R., Amini, A., 1998. An example of mixed carbonate-pyroclastic sedimentation(Miocene, Central Basin, Iran). Sedimentary Geology 118, 37–54.
Perrin, C., Bosence, D., Rosen, B., 1995. Quantitative approaches to palaeozonation andpalaeobathymetry of corals and coralline algae in Cenozoic reefs. In: Bosence, D.W.J., Allison, P.A. (Eds.), Marine Palaeoenvironmental Analysis from Fossils: GeologicalSociety of London Special Publication No. 83, pp. 181–229.
Perry, C., 2005. Structure and development of detrital reef deposits in turbid nearshoreenvironments, Inhaca Island, Mozambique. Marine Geology 214, 143–161.
Peters, E.C., Pilson, M.E.Q., 1985. A comparative study of the effects of sedimentation onsymbiotic and asymbiotic colonies of the coral Astrangia danae Milne Edwards andHaime 1849. Journal of Experimental Marine Biology and Ecology 92, 215–230.
Philipp, E., Fabricius, K., 2003. Photophysiological stress in scleractinian corals inresponse to short-term sedimentation. Journal of Experimental Marine Biology andEcology 287, 57–78.
Puigdefàbregas, C., Muñoz, J.A., Marzo, M., 1986. Thrust belt development in the EasternPyrenees and related depositional sequences in the Southern Foreland Basin. In:Allen, P.A., Homewood, P. (Eds.), Foreland Basins International Association ofSedimentologists Special Publication 8, pp. 229–246.
Purton, L.M.A., Brasier, M.D., 1999. Giant protest Nummulites and its Eoceneenvironment: Life span and habitat insights from δ18O and δ13C data from Num-mulites and Venericardia, Hampshire Basin, UK. Geology 27, 711–714.
Renema, W., 2002. Larger foraminifera as marine environmental indicators. ScriptaGeologica 124 263 pp.
Roberts, H.H., Sydow, J., 1996. The Offshore Mahakam Delta: Stratigraphic Response oflate Pleistocene-to-modern sea level cycle: 25th Indonesian Petroleum Association,Annual Convention, Proceedings, pp. 147–161.
Rodgers, C.S., 1983. Sublethal and lethal effects of sediments applied to commonCaribbean reef corals in the field. Marine Pollution Bulletin 14, 278–382.
Rosen, B.R., Aillud, G.S., Bosellini, F.R., Clack, N., Insalaco, E., Valldeperas, F.X.,Wilson,M.E.J.,2002. Platy coral assemblages: 210 million years of response to the limiting effects ofdepth, light and turbidity. 9th International Coral Reefs Symposium, 2000, Proceed-ings, pp. 255–264.
Sanders, D., Baron-Szabo, R.C., 2005. Scleractinian assemblages under sediment input:their characteristics and relation to the nutrient input concept. Palaeogeography,Palaeoclimatology, Palaeoecology 216, 139–181.
Santisteban, C., Taberner, C., 1988. Sedimentary models of siliciclastic deposits and coralreef interactions. In: Doyle, L.J., Roberts, H.H. (Eds.), Carbonate-Clastic Transitions:Developments in. Sedimentology, vol. 42, pp. 35–76.
Sarangi, S., Sarkar, A., Sarin, M.M., Bhattacharya, S.K., Ebihara, M., Ray, A.K., 2001. Growthrate and life span of Eocene/Oligocene Nummulites tests: inferences from Sr/Caratio. Terra Nova 13, 264–269.
Scoffin, T.P.,1997. Preservation andCaCO3 budgetof reef-flat corals, Phuket, South Thailand.Proceedings 8th International Coral Reef Symposium, vol. 2, pp. 1795–1800.
Sheppard, C.R.C., Spalding, M., Bradshaw, C., Wilson, S., 2002. Erosion vs. recovery ofcoral reefs after 1998 El Niňo: Chagos reefs, Indian Ocean. Ambio 31, 40–48.
Spalding, M.D., Ravilious, C., Green, E.P., 2001. World Atlas of Coral Reefs. University ofCalifornia Press, Berkeley. 424 pp.
Stafford-Smith,M.G.,Ormond,R.F.G.,1992. Sediment rejectionmechanismsof42speciesofAustralian scleractinian corals. Australian Journal of Marine and Freshwater Research43, 683–705.
Steneck, R.S., 1986. The ecology of coralline algal crusts: convergent patterns andadaptive strategies. Annual Review of Ecology and Systematics 7, 273–303.
Testa, V., Bosence, D.W.J., 1998. Carbonate–siliciclastic sedimentation on a high-energy,ocean facing, tropical ramp, NE Brazil. In:Wright, V.P., Burchette, T.P. (Eds.), Carbonateramps: Geological Society, London, Special Publications, vol. 149, pp. 55–61.
Thangaraja, M., Al-Aisry, A., Al-Kharusi, L., 2007. Harmful algal blooms and their impactsin the middle and outer ROPME sea area. International Journal of Oceans andOceanography 2, 85–98.
Titlyanov, E.A., Latypov, Y.Y., 1991. Light-dependence in scleractinian distribution in thesublittoral zone of South China Sea Islands. Coral Reefs 10, 133–138.
Tomascik, T.,Mah, A.J., Nontji, A.,Moosa,M.K.,1997. The Ecology of the Indonesian Seas. In:The Ecology of Indonesia Series, vol. VII. Oxford University Press, p. Oxford. 1388 pp.
Tudhope, A.W., Scoffin, T.P., 1994. Growth and structure of fringing reefs in a muddyenvironment, South Thailand. Journal of Sedimentary Research A64, 752–764.
Umbgrove, J.H.F., 1947. Coral reefs of the East Indies. American Association of PetroleumGeologists Bulletin 58, 729–778.
Van Der Zwaan, G.J., Duijnstee, I.A.P., Dulk, M., Ernst, S.R., Jannink, N.T., Kouwenhoven, T.J.,1999. Benthic foraminifers: proxies or problems? A review of palaeoecologicalconcepts. Earth-Science Reviews 46, 213–236.
Wesseling, I., Uychiaoco, A.J., Alino, P.M., Aurin, T., Vermaat, J.E., 1999. Damage andrecovery of four Philippine corals from short-term sediment burial. Marine Ecology.Progress Series 176, 11–15.
Wilson, M.E.J., 2000. Tectonic and volcanic influences on the development anddiachronous termination of a tropical carbonate platform. Journal of SedimentaryResearch 70, 310–324.
Wilson, M.E.J., 2002. Cenozoic carbonates in SE Asia: implications for equatorialcarbonate development. Sedimentary Geology 147, 295–428.
Wilson, M.E.J., 2005. Equatorial delta-front patch reef development during theNeogene, Borneo. Journal of Sedimentary Research 75, 116–134.
Wilson, M.E.J., 2008. Global and regional influences on equatorial shallow marinecarbonates during the Cenozoic. Palaeogeography, Palaeoclimatology, Palaeoecology265, 262–274.
Wilson, M.E.J., Hall, R. in press. Tectonic influence on SE Asian carbonate systems andtheir reservoir quality. SEPM Special Publication.
Wilson, M.E.J., Lokier, S.W., 2002. Siliciclastic and volcaniclastic influences on equatorialcarbonates: insights from the Neogene of Indonesia. Sedimentology 49, 583–601.
Wilson, M.E.J., Rosen, B.R., 1998. Implications of paucity of corals in the Paleogene of SEAsia: plate tectonics or centre of origin? In: Hall, R., Holloway, J.D. (Eds.), Biogeographyand Geological Evolution of SE Asia. Backhuys Publishers, Netherlands, pp. 165–195.
Wilson, M.E.J., Vecsei, A., 2005. The apparent paradox of abundant foramol facies in lowlatitudes: their environmental significance and effect on platform development.Earth Science Reviews 69, 133–168.
Wolanski, E., Fabricius, K., Spagnol, S., Brinkman, R., 2005. Fine sediment budget on aninner-shelf coral-fringed island, Great Barrier Reef of Australia. Estuarine, Coastaland Shelf Science 65, 153–158.
Woolfe, K.J., Larcombe, P., 1998. Terrigenous sediment accumulation as a regionalcontrol on the distribution of reef carbonates. In: Camoin, G.F., Davies, P.J. (Eds.),Reefs and Carbonate Platforms in the Pacific and Indian Oceans: InternationalAssociation of Sedimentologists Special Publication, vol. 25, pp. 295–310.
Woolfe, K.J., Larcombe, P., 1999. Terrigenous sedimentation and coral reef growth: aconceptual framework. Marine Geology 155, 331–345.
Wu, R.S.S., 1999. Eutrophication, Water Borne Pathogens and Xenobiotic Compounds:Environmental Risks and Challenges. Marine Pollution Bulletin 39, 11–22.
Zingonea, A., Enevoldsen, H.O., 2000. The diversity of harmful algal blooms: a challengefor science and management. Ocean & Coastal Management 43, 725–748.





























