Embed Size (px)
Citation preview

This article was downloaded by: [Niwa]On: 30 April 2014, At: 21:14Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
New Zealand Journal of Marine andFreshwater ResearchPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnzm20
Mangrove (Avicennia marina subsp.australasica) litter production anddecomposition in a temperate estuaryRV Gladstone-Gallaghera, CJ Lundquistbc & CA Pilditcha
a Department of Biological Sciences, University of Waikato,Hamilton, New Zealandb National Institute of Water and Atmospheric Research Ltd(NIWA), Hamilton, New Zealandc Institute of Marine Science, University of Auckland, Auckland,New ZealandPublished online: 16 Sep 2013.
To cite this article: RV Gladstone-Gallagher, CJ Lundquist & CA Pilditch (2014) Mangrove(Avicennia marina subsp. australasica) litter production and decomposition in a temperateestuary, New Zealand Journal of Marine and Freshwater Research, 48:1, 24-37, DOI:10.1080/00288330.2013.827124
To link to this article: http://dx.doi.org/10.1080/00288330.2013.827124
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &

Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

RESEARCH ARTICLE
Mangrove (Avicennia marina subsp. australasica) litter production and
decomposition in a temperate estuary
RV Gladstone-Gallaghera*, CJ Lundquistb,c and CA Pilditcha
aDepartment of Biological Sciences, University of Waikato, Hamilton, New Zealand; bNational Institute ofWater and Atmospheric Research Ltd (NIWA), Hamilton, New Zealand; cInstitute of Marine Science,University of Auckland, Auckland, New Zealand
(Received 15 May 2013; accepted 5 July 2013)
Mangrove forests can provide important cross-boundary subsidies of organic matter to adjacenthabitats through the production, export, decomposition and assimilation of litter. We quantifiedtwo of these components in a temperate mangrove forest in Whangamata Harbour, New Zealand:1) litter production; and 2) decomposition rates as a function of tidal elevation, sediment type andburial depth. Litter traps sampled monthly for a year measured an annual detrital input of 3.24�5.38 t DW ha�1, of which 77% occurred in summer. Decomposition rates depended on litter type,with leaves decomposing faster (63 d to decay by 50%) than pneumatophore and wood material(316 and 460 d, respectively). Buried leaf and wood litter decomposed 1.3�1.4 times slower thanlitter on the sediment surface; however, tidal elevation and sediment type (mud vs. sand) had noeffect. The slow decay of litter (an order of magnitude slower than tropical mangrove litter)suggests that its incorporation into temperate marine food webs may be relatively slow.
Keywords: detritus; New Zealand; organic matter; litterfall; degradation; temperate mangroves;productivity; decay
Introduction
Mangroves can represent an important cross-
boundary subsidy of organic matter to coastal
and continental shelf food webs, through the
production of litterfall, its export and subse-
quent decomposition (Lugo & Snedaker 1974;
Saenger & Snedaker 1993; Polis et al. 1997).
The magnitude of this subsidy is, in part, a
function of litterfall production rates, as well as
the rate of degradation into biologically avail-
able detritus (Lugo & Snedaker 1974; Rice &
Tenore 1981; Robertson 1988). Litterfall pro-
duction rates decrease with increasing latitude
(alongside reduced tree height and tree species
diversity; Twilley et al. 1992; Saenger & Sne-
daker 1993; Harty 2009), but are also influ-
enced by site-specific environmental conditions
(Clarke 1994). Similarly, litter decomposition
rates decrease with increasing latitude (Mackey
& Smail 1996), but can vary within a system
due to tidal submersion period, sediment prop-
erties and macrofaunal community (Robertson
1988; Dick & Osunkoya 2000; Holmer & Olsen
2002; Onate-Pacalioga 2005; Proffitt & Devlin
2005). Assessing the potential role of mangroves
in cross-boundary subsidies therefore requires
site-specific data.To date, most studies have focused on
tropical mangroves and there is a paucity of data
from temperate systems (latitudes�308N and S;
reviewed by Morrisey et al. 2010). New Zealand
has a single mangrove species, Avicennia marina
subsp. australasica, which can be found in estu-
aries ranging from Ohiwa Harbour (38803?S;
*Corresponding author. Email: [email protected]
New Zealand Journal of Marine and Freshwater Research, 2014Vol. 48, No. 1, 24�37, http://dx.doi.org/10.1080/00288330.2013.827124
# 2013 The Royal Society of New Zealand
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

southern limit) to Northland (34827?S; Harty2009). However, litterfall production and de-composition studies have been restricted largelyto the Auckland region (Albright 1976; Woo-droffe 1982, 1985; Onate-Pacalioga 2005; Burnset al. unpublished data, as cited in Morriseyet al. 2010), with only one study from outsidethis area (Northland; May 1999). In manyestuaries, forest area is increasing (Morriseyet al. 2010), and there is a need to assess ratesat which litter enters local ecosystems.
In response to forest area increases, con-sents for large-scale mangrove removals (e.g.c. 110 ha in Tauranga Harbour) have beengranted in New Zealand. Estimates of decom-position rates are needed to help predict recoveryrates following mangrove removal measures thatleave mulched and sometimes buried vegetationin situ to decompose (Harty 2009; Lundquistet al. 2012). The goal of this study was to measureseasonal variations in litterfall and, for the firsttime in a single study, the effects of multipleenvironmental variables on its degradation. Wealso examined decomposition rates of mangrovewood and pneumatophores, since removal meth-odologies may leave behind significant quantitiesof this material.
In the Whangamata estuary, we measuredlitterfall accumulation rates every month fora year in mature and newly established forestto estimate the potential contribution to cross-boundary subsidies. Decomposition rates ofleaves, pneumatophores and woody materialwere measured as a function of tidal elevation,because submergence is known to acceleratedecay (e.g. Robertson 1988; Dick & Osunkoya2000). We also buried material (which occurswhen litter is retained within the forest) to testwhether anoxic conditions slowed decomposi-tion rates, as has been demonstrated elsewhere(Albright 1976; Van der Valk & Attiwill 1984;Fourqurean & Schrlau 2003). While the impor-tance of litter burial in temperate forests has notbeen investigated, in tropical mangrove systems,the burial and incorporation of litter in thesediments is an important pathway of organicmatter recycling within the forest (e.g. Bouillon
et al. 2004; Kristensen et al. 2008). Furthermore,some mangrove removal methods result in theburial of mulched decomposing litter (Lund-quist et al. 2012). The carbon and nitrogencontent of leaf litter during decompositionwas measured to see how conditions influencednutritional value to consumers (Fell et al. 1984;Nordhaus et al. 2011). Finally, because decom-position rates may be altered by sediment proper-ties and associated changes in faunal composition(Bosire et al. 2005; Onate-Pacalioga 2005; Proffitt& Devlin 2005), we compared decay in perme-able sandy sediment and nearby cohesive muddy-sand. Overall, this study makes a contributionto the relatively limited information availableon the production of mangrove detritus and thepotential environmental factors affecting its de-composition in a temperate estuary.
Materials and methods
Study site
The study was conducted at two sites in Whan-gamata Harbour, North Island, New Zealand.Mangroves cover an area of 101 ha (approxi-mately 22% of the harbour area), which hasincreased from 31 ha since the 1940s (Singleton2007). The mangrove forest where our study siteswere located covered an area of 27 ha andextended approximately 200 m out from theshoreline onto the intertidal flat. On the inter-tidal flat seaward of the forest, site 1 (sand;37810?43.4ƒS, 175851?37.4ƒE) was located ap-proximately 50 m from site 2 (muddy sand;37810?39.9ƒS, 175851?36.8ƒE). Macrofaunal com-munity structure was different between the twosites (measured at the mid-tidal position), drivenprimarily by differences in the relative abun-dances of some polychaete species (Gladstone-Gallagher 2012). Sediment at site 1 had a lowermud (particlesB63 mm) content (14.4% vs.29.9%) and a greater median grain size (197.6mm vs. 130.8 mm) than the adjacent site 2.Landward of the sites, 40 m into the forest, treeheights ranged from 1.5�3 m, compared to 1.2�1.9 m at the forest edge. Mean tree density(trees�1 m height) within the mangrove forest
Temperate mangrove litter production and decomposition 25
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

and on the edge of the forest was 14 (93; 9SEM, n�6) and 7 (91; n�6) trees per 100 m2,respectively.
Litterfall production
Litter traps were constructed using nylon shadecloth (2 mm mesh), and placed under themangrove canopy to quantify litterfall (as inWoodroffe 1985; May 1999). Traps were con-ical (0.5 m depth), with a 0.25 m2 opening,designed to minimise litter loss. In total therewere 24 traps: 12 replicate traps (3 m2 samplearea) were randomly positioned under the man-grove trees at the edge of the forest (hereinreferred to as ‘edge’); and another 12 wereplaced 40 m within the forest (herein referredto as ‘within’). At both the edge and withinlocations, traps were randomly distributed in aline parallel to the shore, to encompass bothsites 1 and 2. Trap openings were placed abovethe high tide water level to minimise litter lossduring tidal inundation. Traps were sampledmonthly from February 2011 to January 2012.Samples were rinsed with freshwater to removesediment and salt, and then dried to constantdry weight (DW) at 60 8C. Litter was separatedinto leaf, wood, fruit/seed and inflorescences,and the DW of each determined.
Decomposition rates
Decomposition rates of mangrove wood, pneu-matophores and leaf litter were measured usinglitter bags (16�16 cm) made from 2 mm meshnylon shade cloth (as in Woodroffe 1982). The2 mm mesh size allowed small invertebratesaccess to the decomposing litter, whilst exclud-ing larger macrofauna. The litter bag methodhas been criticised for excluding some macro-invertebrates, which may aid in the breakdownof litter into detritus (Fell et al. 1984), though atemperate study found similar decay rates be-tween bags that allowed and bags that excludedentry of invertebrates (Goulter & Allaway 1979).Although litter bags may underestimate the
conditions of natural decay, they are appropri-ate for determining relative decay rates.
The litter decomposition experiment beganin summer (January 2011), to coincide with theperiod when the majority of mangrove litter isproduced (see Results). Yellow senescent leaves(ready to abscise), wood (branch diameter 5�10mm) and pneumatophores were collected frommangrove trees at Whangamata Harbour. Fi-brous root material was removed from pneuma-tophores. Leaves, wood and pneumatophoreswere rinsed under freshwater to remove sedi-ment and salt, air dried for 48 h (at constanttemperature and humidity), weighed into sub-samples (5 g leaves, 4 g pneumatophores, 7 gwood) and placed separately into the decom-position bags. A similar volume of litter wasplaced in each bag but, because of differences inwater content and density, the initial weightvaried slightly among litter types. To determineinitial DW, 20 subsamples of each litter typewere dried at 60 8C to constant DW.
At each site the decomposition bags wereplaced at four tidal positions: low-tide (T-L);mid-tide (T-M); mangrove edge (pneumato-phore zone with no canopy cover; M-E); andunder the mangrove canopy (M-C). Addition-ally, at the M-E and M-C positions, bags wereburied (10�15 cm depth) in order to test theeffect of burial on decomposition rates (b�buried, s�sediment surface). During periodsof tidal inundation, bags positioned at the T-Lposition were submerged for 1�2.5 h longerthan bags at the M-C position. The effect oftidal position on pneumatophore decomposi-tion was not tested (bags only placed at M-Cand M-E). At each tidal position, decomposi-tion bags were tied to a central stake and thenpegged down (four replicate stakes at each tidalposition).
Four replicate bags of each litter type (n�4;one bag of each litter type per stake) werecollected from each position on d 11, 24, 38, 51,81, 169 and 357 after deployment. Only leafsamples were collected on d 11 and 38, andwood samples were also collected on d 262.Following each collection, samples were rinsed
26 RV Gladstone-Gallagher et al.
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

with freshwater over a 500 mm sieve, placed infoil dishes and oven dried to constant DW at60 8C. Decomposition was characterised by DWloss over time. Dried leaf samples for M-C-s,M-C-b, M-E-s, M-E-b and T-L-s were analysedfor total carbon (C) and total nitrogen (N) bygrinding to a fine powder using a ball mill(Retsch manual mixer mill type MM2000, Ret-sch GmbH & Co.KG, Germany) and then ana-lysed using an Elementar Vario EL cube C andN analyser (Elementar Analysensyteme GmbH,Germany). C and N content were analysed in leafsamples up to d 169, as after this time there wasinsufficient material for analysis.
Data analysis
Raw litterfall data were analysed using a singlet-test to determine significant differences in totalannual litterfall between mangroves on the edgeof the forest (i.e. newly established trees) andmangroves 40 m within the forest (i.e. estab-lished trees). Decomposition (% weight lossafter 357 d) was analysed using different fixedfactor multi-way analyses of variance (ANOVA)to test the effect of: firstly, site (1, 2), tidalposition (M-C-s, M-E-s, T-M-s, T-L-s) and littertype (wood, leaves); and, secondly, burial state(b, s), site (1, 2), forest position (M-C, M-E) andlitter type (leaf, wood, pneumatophore). Percen-tage weight loss data were arcsine transformedand the distribution of residuals analysed aftertransformation, to ensure data met the assump-tions of normality and homogeneity of variances.An additional fixed factor multi-way ANOVAtested the effect of tidal position and burialstate (M-C-s, M-C-b, M-E-s, M-E-b, T-L-s) onleaf litter C and N content (raw data conformedto statistical assumptions) after 169 d. Newman-Keuls post hoc tests were used to determinewhere significant differences occurred.
To characterise the decomposition process(as opposed to the final result tested above)and for comparison with previous studies (e.g.Robertson 1988; Mackey & Smail 1996; Onate-Pacalioga 2005), we fitted decay models to the
time series data to estimate t50 (time taken forlitter to decay to 50% of original weight). Asingle exponential decay model of X(t)�e�kt ,where X(t) is the proportion of mangrove mate-rial remaining after time t (days) and k (day�1) isthe decay constant, was used to describe the decayof surficial mangrove leaf litter. However, thedecomposition of buried leaves was more suitablydescribed by the asymptotic model (single r2B0.3,asymptotic r2�0.8), X(t)�C�(1�C)e�kt,which assumes there is a fast initial decay of easilybroken down labile material (k), followed by acompletely decay-resistant recalcitrant fraction(C) (Wieder &Lang 1982). This asymptotic modelassumes that litter will never decay completely,and therefore is unrealistic in nature; however, themodel can be useful to describe decompositionrates during the study period (Wieder & Lang1982). Wood and pneumatophore weight loss wassmall and data were highly variable. As a result,exponential decay models did not provide a goodfit (r2�0.3�0.6), so a linear decay rate (a) wasassumed between the initial (0 d) and final (357 d)sample date. Multi-way ANOVAs found nosignificant differences in weight loss (after 357 d)among tidal positions and sites (see Results),therefore decay constants and t50 values presentedin the Results section were estimated from meanspooled across sites and tidal positions.
Results
Litterfall production
Mean annual litterfall 40 m within the forest(538974 g DW m�2 yr�1;9SEM) was sig-nificantly higher (t-test, P�0.015, n�12) com-pared to the younger trees on the edge of theforest (324943 g DW m�2 yr�1). Annuallitterfall consisted of 60%�65% leaf, 9%�11%wood, 25%�26% fruit and 1%�3% inflores-cences, both on the edge and within the man-grove forest. At both locations the majority oflitterfall (77%) occurred during the warmermonths of November to February (Fig. 1). Leaffall occurred all year round, but was minimal inthe colder months (March�October). Wood fallremained relatively constant throughout the year,
Temperate mangrove litter production and decomposition 27
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

and so comprised a greater fraction (up to 60%)
of the monthly litterfall in winter. Fruit fall was
largest during summer months (12%�44% of
monthly litterfall), but low during the remainder
of the year (0%�29%of monthly litterfall). Inflo-
rescences were collected throughout the year,
although they represented a very small propor-
tion of the annual litterfall (1%�3%).
egdE
ni htiW
egdE
nihtiW
egdE
nihtiW
egdE
nihtiW
e gdE
nihtiW
egdE
n iht iW
e gdE
nihtiW
egdE
niht iW
egdE
nihtiW
egdE
niht iW
e gdE
nih tiW
egdE
nihtiW
JanFeb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
0
5
10
15
20
25
Mea
n m
onth
ly a
ir te
mpe
ratu
re (
°C)
0
20
40
60
80
100
120
140
160
180
Litte
rfal
l (g
DW
m–2
)
Mean monthly air temperature
Leaf Wood Fruit Inflorescences
Figure 1 Mean (�1 SEM, n�12) monthly (2011�2012) mangrove litterfall rates and composition, on theedge and 40 m within the forest. Secondary y-axis shows mean monthly air temperature (data obtained fromtemperature loggers deployed in Whangamata Harbour during the study).
Table 1 Summary of multi-way ANOVA comparing mean percentage weight loss after 357 d (arcsine
transformed) between sites (1, 2), litter types (wood, leaf), and tidal positions (M-C, M-E, T-M, T-L). Significantresults (PB0.05) are indicated in bold. Results of Newman-Keuls post hoc tests are shown as footnotes.
Source of variation d.f. Mean-square F-ratio P
Site 1 0.01 0.62 0.436Litter type 1 7.49 420.31 B0.0011
Tidal position 3 0.03 1.66 0.189
Site*Litter type 1 0.01 0.69 0.411Site*Tidal position 3 0.01 0.30 0.824Litter type*Tidal position 3 0.05 2.71 0.056
Site*Litter type*Tidal position 3 0.01 0.29 0.829Error 47 0.02
1Leaf�wood.
28 RV Gladstone-Gallagher et al.
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

Decomposition rates
A multi-way ANOVA revealed that, for wood
and leaf litter on the sediment surface, neither
site nor tidal position affected weight loss after
357 d of decomposition. There was, however, a
significant difference in weight loss with litter
type, where leaves lost on average 95% of their
weight after 357 d, while wood lost only 40%
(Table 1, Fig. 2). In a secondmulti-wayANOVA,
burial state (surface, buried), litter type (leaf,
wood, pneumatophore) and their interaction
were significant; however, other factors (site,
forest position) were not significant (Table 2,
Fig. 2). Post hoc testing revealed leaf litter
decomposition on the sediment surface (at both
M-C and M-E positions) lost the most weight
(95%), followed by pneumatophores (59%) and
wood (40%). Buried leaf and pneumatophore
material lost the same weight over 357 d (61%
and 58%, respectively), which was about 2 times
M-C-bM-C-s M-E-s
M-E-b T-L-sT-M-s
0 50 100 150 200350 400200 250 300 250 3001500 50 100 350 400
Time (d)Time (d)
0
20
40
60
80
100
0
20
40
60
80
100
80
100
0
20
40
60
ero
hp
otW
eig
ht re
main
ing (
%)
am
ue
nP
Site 1 Site 2BA
DC
E F
Weig
ht re
main
ing (
%)
Leaf
Weig
ht re
main
ing (
%)
Wood
Figure 2 Mangrove litter decomposition (expressed as the percentage of original weight remaining throughtime) at varying tidal positions and burial states. A, Leaves, site 1. B, Leaves, site 2. C, Wood, site 1. D,Wood, site 2. E, Pneumatophores, site 1. F, Pneumatophores, site 2. For clarity, each figure includes themaximum and minimum standard error (SEM).
Temperate mangrove litter production and decomposition 29
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

more weight than buried wood (30% loss).Pneumatophore weight loss was not affectedby burial state, with a buried and surface weightloss of 58% and 59%, respectively. However,buried wood and leaves lost significantly lessweight (30% and 61%, respectively) than theirsurficial counterparts (40% and 95%, respec-tively) (Table 2, Fig. 2).
The fitted decay models show differences indecomposition rates that are reflected in weightloss differences after 357 d between litter typesand burial treatments (Table 3). The t50 ofwood (460�613 d) was about 1.5 and 7 timeslonger than pneumatophores (317 d) and leaves(63�88 d), respectively. The mean t50 of buriedleaves and wood was approximately 1.3 and 1.4times greater (an additional 25 d and 153 d,respectively) than for surface equivalents, re-spectively (Table 3).
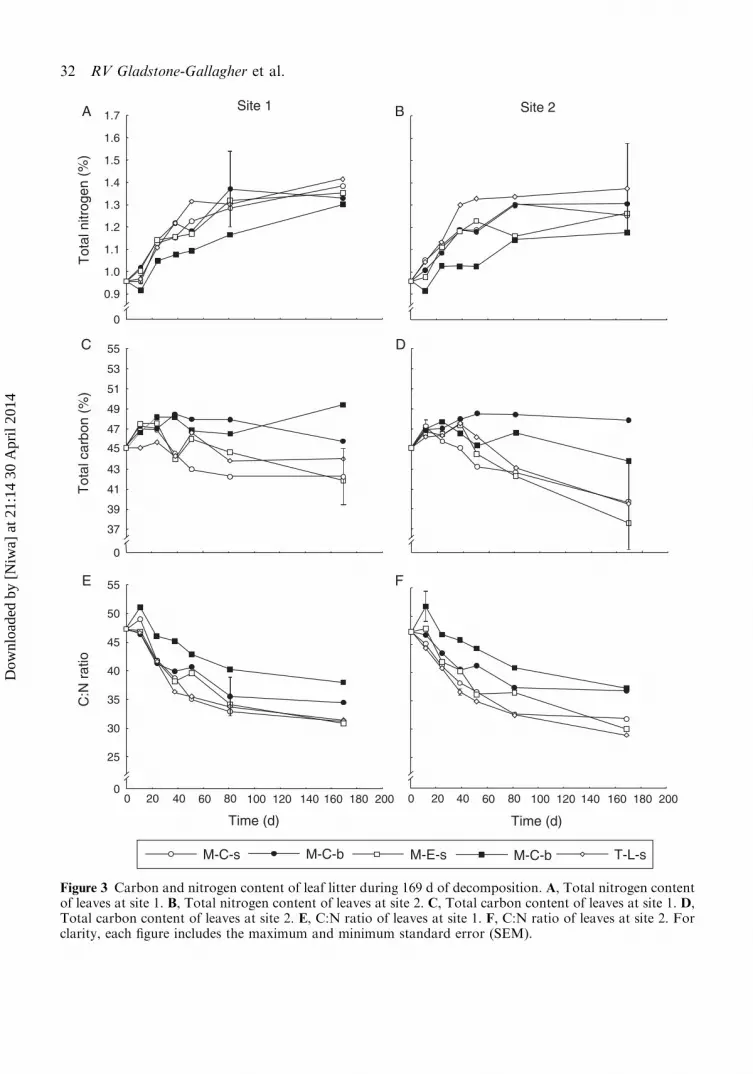
After 169 d of decomposition, average totalN increased in all leaf litter from 0.96% to1.31% (Fig. 3A, 3B), but there was no signifi-cant difference in final N content with site, tidal
position and burial state (Table 4). In contrast,
total C content was dependent on site and burial
state, but was unaffected by tidal position. C
content of leaf litter decreased in leaves decom-
posing on the sediment surface from 45.1%
(initial) to 42.7% and 39.0% (at sites 1 and 2,
respectively; Fig. 3C, 3D). However, C content
of buried leaves (47.6% site 1, 45.8% site 2, after
357 d) remained the same, or increased slightly,
during the decomposition process (Fig. 3C, 3D).
Consequently, the final C content of buried
leaves was significantly higher than for leaves
on the surface (Table 4). Leaves decomposing at
site 2 had a significantly lower final C content
(45.83% buried, 38.95% surface) than leaves at
site 1 (47.58% buried, 42.71% surface; Table 4).
As a result of N enrichment, the C:N ratio
decreased in all leaves from a mean of 47 on d
0 to 31 (surface) and 37 (buried) after 169 d
(Fig. 3E, 3F). Buried leaves had a significantly
higher C:N ratio (37) after 169 d compared to
leaves on the sediment surface (31). No significant
Table 2 Summary of multi-way ANOVA comparing mean percentage weight loss after 357 d (arcsine
transformed) between sites (1, 2), litter types (wood, leaf, pneumatophore), forest positions (M-C or M-E)and burial states (buried or surface). Significant results (PB0.05) are indicated in bold. Results of Newman-Keuls post hoc tests are shown as footnotes.
Source of variation d.f. Mean-square F-ratio P
Site 1 0.01 0.24 0.626Litter type 2 1.86 88.21 B0.001
Forest position 1 0.01 0.31 0.580Burial state 1 0.91 42.93 B0.001
Site*Litter type 2 0.04 1.68 0.195
Site*Forest position 1 0.04 1.98 0.165Litter type*Forest position 2 0.02 1.18 0.316Site*Burial state 1 0.07 3.32 0.073
Litter type*Burial state 2 0.48 22.94 B0.0011
Position*Burial state 1 0.06 2.78 0.101Site*Litter type*Forest position 2 0.03 1.48 0.235Site*Litter type*Burial state 2 0.02 1.04 0.359
Site* Forest position*Burial state 1 0.00 0.07 0.794Litter type* Forest position*Burial state 2 0.04 2.10 0.131Site*Litter type* Forest position*Burial state 2 0.01 0.26 0.774
Error 60 0.02
1Buried: leaf�pneum.�wood; Surface: leaf�pneum.�wood; Leaf: surface�buried; Wood: surface�buried; Pneum.:buried�surface.
30 RV Gladstone-Gallagher et al.
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

site effect was detected in leaf C:N ratios after169 d (Table 4).
Discussion
Litterfall production
Annual litterfall estimates in WhangamataHarbour are equivalent to 3.24�5.38 t DW ha�1
yr�1 (324�538 g DW m�2 yr�1), and are withinthe range reported in the literature for other sitesin New Zealand (1.30�8.10 t DW ha�1 yr�1;Woodroffe 1985; May 1999; Onate-Pacalioga2005). They are also similar to mangrove forestsat other temperate latitudes (0.11�11.68 t DWha�1 yr�1), but are at the lower end of thoseobserved for tropical forests (3.74�18.7 t DWha�1 yr�1; reviewed by Morrisey et al. 2010).Litter composition was consistent with otherstudies, where leaves constitute the largestproportion of the litter (60%�65%; e.g. Woo-droffe 1985; May 1999; Imgraben & Dittmann2008). Litter production of younger trees on theedge of the Whangamata mangrove forest wasroughly half that of the established trees withinthe forest, consistent with previous studies thathave shown productivity to be spatially variablewithin a forest and can vary with tree height(reviewed by Morrisey et al. 2010).
The seasonal variation in litterfall, charac-terised by a pulse in summer months and mini-mal production during the rest of the year, hasbeen reported elsewhere and is attributed toseasonal changes in temperature, rainfall andevapotranspiration (Woodroffe 1985; Clarke1994; May 1999; Onate-Pacalioga 2005; Sanchez-Andres et al. 2010). Conversely, in some tropi-cal regions there are multiple peaks in litterproduction throughout the year (Duke 1990).While litterfall measurements do not measurethe increase in plant biomass, they are regardedas an important component of primary produc-tivity in determining organic matter contributionto the estuary (Woodroffe 1982; Clarke 1994). Intemperate systems, cross-boundary subsidies arehighly seasonal compared to tropical systems,and therefore are likely to function differentlyT
able3Fitteddecaymodelsusedto
describelitter
decompositionasafunctionoftypeandburialposition.t 50values
representthetime(days)
for
litter
todecayto
50%
oforiginalweight(calculatedfrom
decaymodelequationsandrate
constants).Alsoshownare
thedecayconstants(and95%
confidence
intervals)andanestimate
ofthemodel
fit(r2values).Meansare
calculatedfrom
meanspooledacross
sitesandtidalpositions(w
here
ANOVA
foundnosignificantdifference
intotalweightloss).Footnotesindicate
themodel
equationsthatwerefitted.
Model
k9SEM
95%
CI
C9SEM
95%
CI
a9SEM
95%
CI
r29SEM
t 509SEM
95%
CI
Leaves-surface
Single1
0.011190.0005
0.0102�0
.0120
��
��
0.9090.01
6393
58�6
8
Leaves-buried
Asymptotic2
0.032790.0042
0.0256�0
.0399
0.463290.0134
0.4405�0
.4860
��
0.8390.06
8896
78�9
7
Wood-surface
Linear3
��
��
0.001190.0001
0.001�0
.0012
0.8690.04
460928
409�5
11
Wood-buried
Linear3
��
��
0.000890.0001
0.0007�0
.0009
0.8290.09
613943
539�6
86
Pneum.
Linear3
��
��
0.001690.0001
0.0014�0
.0018
0.9190.05
317930
263�3
73
1X(t)
�e
�kt ;
2X(t)�C
�(1
�C)e
�kt ;
3Y
�aX
�1.
Temperate mangrove litter production and decomposition 31
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014
E
C D
F
A B
0
0.9
1.0
1.1
1.2
1.3
1.4
1.5
1.6
1.7
Tot
al n
itrog
en (
%)
43
45
47
49
37
39
41
51
53
55
0
Tot
al c
arbo
n (%
)
25
30
35
40
45
50
55
0
C:N
rat
io
120 140 160 180 2000 20 40 60 80 100 120 140 160 180 2000 20 40 60 80 100
Time (d) Time (d)
Site 1 Site 2
M-C-s M-C-b M-E-s M-C-b T-L-s
Figure 3 Carbon and nitrogen content of leaf litter during 169 d of decomposition. A, Total nitrogen contentof leaves at site 1. B, Total nitrogen content of leaves at site 2. C, Total carbon content of leaves at site 1. D,Total carbon content of leaves at site 2. E, C:N ratio of leaves at site 1. F, C:N ratio of leaves at site 2. Forclarity, each figure includes the maximum and minimum standard error (SEM).
32 RV Gladstone-Gallagher et al.
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014
with regard to consumer response and nutrientcycling.
Although temperate mangrove forests are lessproductive than those in the tropics, our resultssuggest that mangrove detrital production issimilar to that produced by seagrass (Zosteramuelleri). Above-ground seagrass production inthe Whangamata Harbour has been estimatedat 1.1�1.6 g DW m�2 d�1 (Turner 2007) which,assuming a 71 ha area (Singleton 2007), suggestsan annual production rate for the harbour of 370t DW. Scaling up mangrove litter productionrates measured in this study to the 101 ha offorest (Singleton 2007) gives a comparable esti-mate of 327�543 t DW yr�1. Seagrass beds inboth tropical and temperate coastal systems arean important detrital subsidy at the base ofcoastal food webs (e.g. Doi et al. 2009). Oursimple comparison suggests mangroves couldpotentially provide an important source ofdetritus to temperate coastal systems; however,this will be dependent on how much of theproductivity is exported from the forest, as wellas its rate of decay and assimilation.
Decomposition rates
Burial significantly reduced the rate and amountof leaf weight and carbon loss, and differencesin the fitted decay models indicate distinct differ-ences in the decay processes. A large proportionof leaf carbon is locked up in the recalcitrant
decay resistant fraction of the leaf (Davis et al.2003), which resisted decay to a greater extentin buried leaves (asymptotic vs. single decaymodels). The asymptotic decay model (used todescribe buried litter decay) has been associatedwith litter decomposition that excludes faunalactivity (Wieder & Lang 1982), as macrofaunamay aid in degradation of the recalcitrant por-tion of the leaf (Onate-Pacalioga 2005; Proffitt &Devlin 2005). Accordingly, anoxic litter decom-position is likely to be primarily through bacter-ial breakdown, as biotic and abiotic variablesthat accelerate litter fragmentation (macrofauna,climate and tidal inundation; Robertson 1988;Mackey & Smail 1996; Woitchik et al. 1997;Davis et al. 2003; Proffitt & Devlin 2005) will beminimal at depths of 10�15 cm in the sediment.Our findings are consistent with results fromsubtropical Florida, where carbon loss wasgreater in surficial compared to buried leaves,although no differences in mass loss wereobserved (Fourqurean & Schrlau 2003). Leaflitter that is retained within the mangrove forestand buried due to sedimentation could result innutrient recycling within the forest. However,the recycling of buried litter is likely to be slowin New Zealand forests due to the slow rateof weight loss and carbon decay under anoxicconditions.
Fresh mangrove leaves are a relatively lowquality food resource to marine consumers (hightannins and C:N ratios), and decomposition turns
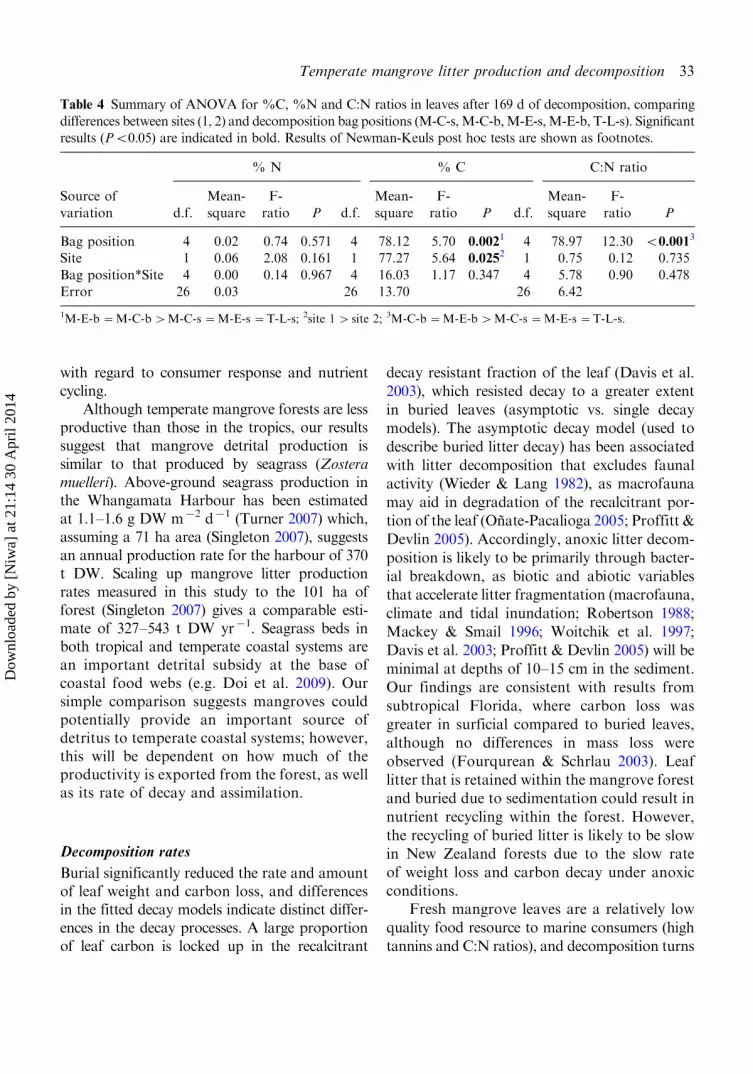
Table 4 Summary of ANOVA for %C, %N and C:N ratios in leaves after 169 d of decomposition, comparing
differences between sites (1, 2) and decomposition bag positions (M-C-s,M-C-b,M-E-s,M-E-b, T-L-s). Significantresults (PB0.05) are indicated in bold. Results of Newman-Keuls post hoc tests are shown as footnotes.
% N % C C:N ratio
Source of
variation d.f.
Mean-
square
F-
ratio P d.f.
Mean-
square
F-
ratio P d.f.
Mean-
square
F-
ratio P
Bag position 4 0.02 0.74 0.571 4 78.12 5.70 0.0021 4 78.97 12.30 B0.0013
Site 1 0.06 2.08 0.161 1 77.27 5.64 0.0252 1 0.75 0.12 0.735
Bag position*Site 4 0.00 0.14 0.967 4 16.03 1.17 0.347 4 5.78 0.90 0.478Error 26 0.03 26 13.70 26 6.42
1M-E-b �M-C-b�M-C-s �M-E-s �T-L-s; 2site 1�site 2; 3M-C-b �M-E-b�M-C-s �M-E-s �T-L-s.
Temperate mangrove litter production and decomposition 33
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

unpalatable leaf litter into biologically availabledetritus (via the microbial pathway; Robertson1988; Nordhaus et al. 2011). Leaf palatability toconsumers increases as a function of a reducedC:N ratio and enhanced bacterial colonisation(Fell et al. 1984; Nordhaus et al. 2011). Here, theC:N ratio in all leaves decreased from approxi-mately 45 to 35 after 3 months of decompositionas a result of nitrogen enrichment and carbonloss; however, this occurred to a greater extentin surficial leaves, as buried leaves resistedcarbon decay. Similar C:N decreases have beenmeasured in subtropical and tropical studies(Rice & Tenore 1981; Dick & Osunkoya 2000).However, in other tropical mangrove studies,C:N ratios decreased by half within 4 months(C:N decrease from 110 to c. 50, Fourqurean &Schrlau 2003; and from 75 toB37.5, Robertson1988).
Although wood comprises 9%�18% of an-nual mangrove litterfall (this study; May 1999),wood decomposition studies are lacking in tem-perate regions. As mangrove wood and rootmaterial is often left in situ following mangroveremovals in New Zealand estuaries (Lundquistet al. 2012), it is useful to describe the decay ofsuch material to determine site recovery. Modelpredictions (linear decay) suggest that man-grove wood and pneumatophores could takebetween 317 and 613 d to decompose to halftheir original weight. Even at lower latitudes(subtropical and tropical), mangrove wood de-cay is expected to take years (e.g. t50�179�421d; Steinke et al. 1983; Robertson & Daniel 1989;Mackey & Smail 1996). Wood decay was signi-ficantly slowed by burial, which can occur due tosedimentation or macroalgal blooms on clearedmangrove patches (Lundquist et al. 2012). Incontrast, pneumatophore decay rate was unaf-fected by burial, though this study consideredonly the decay of pneumatophores, whilst ex-cluding the fibrous root material. Just twotemperate studies have investigated mangroveroot decay; one found that pneumatophoresand fibrous roots decomposed at similar rates(Albright 1976), while the other showed thatpneumatophores decomposed significantly faster
(Van der Valk & Attiwill 1984). Thus, our resultsmay provide an underestimate of complete rootdegradation rates.
Previous investigations have linked litterdecay rates with differences in tidal submer-sion (Robertson 1988; Mackey & Smail 1996;Woitchik et al. 1997; Dick & Osunkoya 2000)and site characteristics, such as local macro-faunal assemblages (Bosire et al. 2005; Proffitt &Devlin 2005) and sediment properties (Holmer& Olsen 2002). However, expectations that man-grove litter decay rates would differ betweensites and tidal positions were not supported byour results. Differences in tidal submergenceperiods tested in previous studies were larger(e.g. submerged daily vs. submerged only insome tidal cycles; Robertson 1988; Mackey &Smail 1996; Woitchik et al. 1997; Dick &Osunkoya 2000) than those measured by thisexperiment (1�2.5 h difference in submergenceperiod between high and low tidal elevations).The relatively small difference in the submer-gence times compared in this study may havebeen too small to influence mangrove litterdecay. In addition, site-specific differences indecay rates have been observed in distinctlydifferent macrofaunal communities (Bosire et al.2005), but this study measured only differencesin the relative abundances of species ratherthan differences in species compositions (Glad-stone-Gallagher 2012).
We anticipated that litter decay rates woulddiffer with sediment type, because in tropicalforests rates were faster in sandy nutrient-poorsediments than in muddy nutrient-rich sedi-ments (k�0.013 vs. 0.0075 day�1; Holmer &Olsen 2002). Although, we detected no siteeffect in litter weight loss, the two sites ex-hibited differences in leaf carbon content. Leaflitter at the muddy sand site (site 2) lost signi-ficantly more carbon (after 169 d), which couldindicate increased degradation of the recalci-trant components of the leaf (Davis et al.2003), though this was not mirrored in massloss.
Mangrove leaf degradation is slow in tempe-rate New Zealand forests (t50�42�88 d; Albright
34 RV Gladstone-Gallagher et al.
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

1976; Woodroffe 1982; Onate-Pacalioga 2005;
this study), compared to tropical counterparts,
where t50 is often reached within one week (e.g.
Bosire et al. 2005; Sanchez-Andres et al. 2010).
Consequently, it is expected that organic matter
cycling and detrital production is faster in
tropical forests than in temperate New Zealand.
However, cross-boundary subsidies of organic
matter from mangrove forests are not only
controlled by the production and decomposi-
tion of litter, but also depend on whether
the litter is exported from the forest (Lugo &
Snedaker 1974; Polis et al. 1997). Some authors
have hypothesised that rapid decomposition
rates could be associated with a lower incidence
of litter export and a greater chance of nutri-
ent recycling within the mangrove forest, with
slow decomposition rates exhibiting the reverse
(Imgraben & Dittmann 2008; Adame & Lovelock
2011). Other studies, utilising stable isotopes, have
suggested that mangrove litter in temperate re-
gions is more likely to be consumed within the
forest rather than exported to adjacent food webs
(Guest & Connolly 2004; Alfaro et al. 2006).
Therefore, the fate of mangrove litter in temperate
forests requires further investigation through
the quantification of litter export to determine
the magnitude of the potential cross-boundary
subsidy.
Acknowledgements
Funding was provided by the New Zealand Ministry
for Science and Innovation Project No. CO1X1002,
the Waikato Regional Council and University of
Waikato Masters Research Scholarship. We thank
Sarah Hailes, Dudley Bell, Nikki Webb, Gillian
Gladstone, Maxine Gillard, Ray Foster, Martin
Gallagher, Simon Brown, Barry Greenfield, Lisa
McCartain, Pauline Robert, Daniel Pratt, Jamie
Armstrong, Arie Spyksma, Stacey Buchanan, Nicho-
las Wu, Rachel Harris, Clarisse Niemand, Dorothea
Kohlmeier, Kelly Carter, Phil Ross, Tim Rogers,
Anya Gladstone-Gallagher and Marenka Weis for
assistance with laboratory and field work. Also, we
thank three anonymous reviewers for their construc-
tive comments that improved the manuscript.
References
Adame MF, Lovelock CE 2011. Carbon and nu-trient exchange of mangrove forests with thecoastal ocean. Hydrobiologia 663: 23�50.
Albright LJ 1976. In situ degradation of mangrovetissues (note). New Zealand Journal of Marineand Freshwater Research 10: 385�389.
Alfaro AC, Thomas F, Sergent L, Duxbury M 2006.Identification of trophic interactions within anestuarine food web (northern New Zealand) usingfatty acid biomarkers and stable isotopes. Es-tuarine, Coastal and Shelf Science 70: 271�286.
Bosire JO, Dahdouh-Guebas F, Kairo JG, KazunguJ, Dehairs F, Koedam N 2005. Litter degrada-tion and CN dynamics in reforested mangroveplantations at Gazi Bay, Kenya. BiologicalConservation 126: 287�295.
Bouillon S,Moens T, KoedamN,Dahdouh-Guebas F,Baeyens W, Dehairs F 2004. Variability in theorigin of carbon substrates for bacterial commu-nities in mangrove sediments. FEMS Microbio-logy Ecology 49: 171�179.
Clarke PJ 1994. Baseline studies of temperate man-grove growth and reproduction: demographicand litterfall measures of leafing and flowering.Australian Journal of Botany 42: 37�48.
Davis SE, Corronado-Molina C, Childers DL, DayJr. JW 2003. Temporally dependent C, N andP dynamics associated with the decay ofRhizophora mangle L. leaf litter in oligotrophicmangrove wetlands of the southern Everglades.Aquatic Botany 75: 199�215.
Dick TM, Osunkoya OO 2000. Influence of tidalrestriction floodgates on decomposition of man-grove litter. Aquatic Botany 68: 273�280.
Doi H, Matsumasa M, Fujikawa M, Kanou K,Suzuki T, Kikuchi E 2009. Macroalgae andseagrass contribution to gastropods in sub-tropical and temperate tidal flats. Journal ofthe Marine Biological Association of the UnitedKingdom 89: 399�404.
Duke NC 1990. Phenological trends with latitude inthe mangrove tree Avicennia marina. Journal ofEcology 78: 113�133.
Fell JW, Master IM, Wiegert RG 1984. Litterdecomposition and nutrient enrichment. In:Snedaker SC, Snedaker JG eds. The mangroveecosystem: research methods. Paris, UNESCO.Pp. 239�251.
Fourqurean JW, Schrlau JE 2003. Changes in nu-trient content and stable isotope ratios of C andN during decomposition of seagrasses and man-grove leaves along a nutrient availability gradientin Florida Bay, USA. Chemistry and Ecology 19:373�390.
Temperate mangrove litter production and decomposition 35
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

Gladstone-Gallagher RV 2012. The production anddecay of mangrove (Avicennia marina subsp.australasica) detritus and its effects on coastalbenthic communities. Unpublished MSc thesis.University of Waikato, New Zealand. 86 p.
Goulter PFE, Allaway WG 1979. Litterfall anddecomposition in a mangrove stand, Avicenniamarina (Forsk.) Vierh., in Middle Harbour,Sydney. Australian Journal of Marine and Fresh-water Research 30: 541�546.
Guest MA, Connolly RM 2004. Fine-scale move-ment and assimilation of carbon in saltmarshand mangrove habitat by resident animals.Aquatic Ecology 38: 599�609.
Harty C 2009. Mangrove planning and managementin New Zealand and South East Australia � areflection on approaches. Ocean & Coastal Man-agement 52: 278�286.
Holmer M, Olsen AB 2002. Role of decompositionof mangrove and seagrass detritus in sedimentcarbon and nitrogen cycling in a tropicalmangrove forest. Marine Ecology Progress Series230: 87�101.
Imgraben S, Dittmann S 2008. Leaf litter dynamicsand litter consumption in two temperate SouthAustralian mangrove forests. Journal of SeaResearch 59: 83�93.
Kristensen E, Bouillon S, Dittmar T, Marchand C2008. Organic carbon dynamics in mangroveecosystems: a review. Aquatic Botany 89: 201�219.
Lugo AE, Snedaker SC 1974. The ecology of man-groves. Annual Review of Ecology and Systema-tics 5: 39�64.
Lundquist C, Hailes S, Cartner K, Carter K, GibbsM 2012. Physical and ecological impacts asso-ciated with mangrove removals using in situmechanical mulching in Tauranga Harbour.NIWA Technical Report 137. 106 p.
Mackey AP, Smail G 1996. The decomposition ofmangrove litter in a subtropical mangrove forest.Hydrobiologia 332: 93�98.
May JD 1999. Spatial variation in litter produc-tion by the mangrove Avicennia marina var.australasica in Rangaunu Harbour, Northland,New Zealand. New Zealand Journal of Marineand Freshwater Research 33: 163�172.
Morrisey DJ, Swales A, Dittmann S, Morrison MA,Lovelock CE, Beard CM 2010. The ecology andmanagement of temperate mangroves. Oceano-graphy and Marine Biology: An Annual Review48: 43�160.
Nordhaus I, Salewski T, Jennerjahn TC 2011. Foodpreferences of mangrove crabs related to leafnitrogen compounds in the Segara Anakan
Lagoon, Java, Indonesia. Journal of Sea Re-search 65: 414�426.
Onate-Pacalioga JA 2005. Leaf litter production,retention, and decomposition of Avicennia marinavar. australasica at Whangateau Estuary, North-land, New Zealand. Unpublished MSc thesis.University of Auckland, New Zealand. 108 p.
Polis GA, Anderson WB, Holt RD 1997. Toward anintegration of landscape and food web ecology:the dynamics of spatially subsidized food webs.Annual Review of Ecology and Systematics 28:289�316.
Proffitt CE, Devlin DJ 2005. Grazing by the inter-tidal gastropod Melampus coffeus greatly in-creases mangrove litter degradation rates. MarineEcology Progress Series 296: 209�218.
Rice DL, Tenore KR 1981. Dynamics of carbon andnitrogen during the decomposition of detritusderived from estuarine macrophytes. Estuarine,Coastal and Shelf Science 13: 681�690.
Robertson AI 1988. Decomposition of mangroveleaf litter in tropical Australia. Journal of Experi-mental Marine Biology and Ecology 116: 235�247.
Robertson AI, Daniel PA 1989. Decomposition andthe annual flux of detritus from fallen timber intropical mangrove forests. Limnology and Ocea-nography 34: 640�646.
Saenger P, Snedaker SC 1993. Pantropical trends inabove-ground biomass and annual litterfall.Oecologia 96: 293�299.
Sanchez-Andres R, Sanchez-Carrillo S, Alatorre LC,Cirujano S, Alvarez-Cobelas M 2010. Litterfalldynamics and nutrient decomposition of aridmangroves in the Gulf of California: their rolein sustaining ecosystem heterotrophy. Estuar-ine, Coastal and Shelf Science 89: 101�199.
Singleton P 2007. Draft Whangamata Harbour plan.Environment Waikato Internal Series 2007/14.84 p.
Steinke TD, Naidoo G, Charles LM 1983. Degrada-tion of mangrove leaf and stem tissues in situin Mgeni Estuary, South Africa. In: Teas HJ ed.Biology and ecology of mangroves. The Hague,The Netherlands, Dr W Junk Publishers.Pp. 141�149.
Turner SJ 2007. Growth and productivity of inter-tidal Zostera capricorni in New Zealand estu-aries. New Zealand Journal of Marine andFreshwater Research 41: 77�90.
Twilley RR, Chen RH, Hargis T 1992. Carbon sinksin mangroves and their implications to carbonbudget of tropical coastal ecosystems. Water,Air and Soil Pollution 64: 265�288.
Van der Valk AG, Attiwill PM 1984. Decompositionof leaf and root litter of Avicennia marina at
36 RV Gladstone-Gallagher et al.
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014

Westernport Bay, Victoria, Australia. AquaticBotany 18: 205�221.
Wieder RK, Lang GE 1982. A critique of theanalytical methods used in examining decom-position data obtained from litter bags. Ecology63: 1636�1642.
Woitchik AF, Ohowa B, Kazungu JM, Rao RG,Goeyens L, Dehairs F 1997. Nitrogen enrich-ment during decomposition of mangrove leaflitter in an eastern African coastal lagoon
(Kenya): relative importance of biological nitro-gen fixation. Biogeochemistry 39: 15�35.
Woodroffe CD 1982. Litter production and decom-position in theNewZealandmangrove,Avicenniamarina var. resinifera. New Zealand Journal ofMarine and Freshwater Research 16: 179�188.
Woodroffe CD 1985. Studies of a mangrove basin,Tuff Crater, New Zealand: I. Mangrove bio-mass and production of detritus. Estuarine,Coastal and Shelf Science 20: 265�280.
Temperate mangrove litter production and decomposition 37
Dow
nloa
ded
by [
Niw
a] a
t 21:
14 3
0 A
pril
2014





























