Embed Size (px)
Citation preview

Molecular Cell
Article
Long Noncoding RNA TARIDDirects Demethylation and Activationof the Tumor Suppressor TCF21 via GADD45AKhelifa Arab,1,2 Yoon Jung Park,2,13 Anders M. Lindroth,2,14 Andrea Schafer,1 Christopher Oakes,2 Dieter Weichenhan,2
Annekatrin Lukanova,3,4 Eva Lundin,4 Angela Risch,2,5 Michael Meister,5,6 Hendrik Dienemann,5,7 Gerhard Dyckhoff,8
Christel Herold-Mende,8,9 Ingrid Grummt,10,* Christof Niehrs,1,11,* and Christoph Plass2,12,*1Institute of Molecular Biology (IMB), 55128 Mainz, Germany2Division of Epigenomics and Cancer Risk Factors, German Cancer Research Center (DKFZ), 69120 Heidelberg, Germany3Division of Cancer Epidemiology, German Cancer Research Center (DKFZ), 69120 Heidelberg, Germany4Medical Biosciences, Department of Pathology, Umea University, 90185 Umea, Sweden5TranslationalLungResearchCenterHeidelberg (TLRC-H),Memberof theGermanCenter for LungResearch (DZL), 69120Heidelberg,Germany6Translational Research Unit, Thoraxklinik-Heidelberg gGmbH University of Heidelberg, 69120 Heidelberg, Germany7Department of Thoracic Surgery, Thoraxklinik-Heidelberg gGmbH, University of Heidelberg, 69120 Heidelberg, Germany8Department of Otorhinolaryngology, Head and Neck Surgery, University of Heidelberg, 69120 Heidelberg, Germany9Division of Neurosurgical Research, Department of Neurosurgery, University of Heidelberg, 69120 Heidelberg, Germany10Division of Molecular Biology of the Cell II, DKFZ-ZMBH Alliance, 69120 Heidelberg, Germany11Division of Molecular Embryology, DKFZ-ZMBH Alliance, 69120 Heidelberg, Germany12The German Cancer Consortium, 69120 Heidelberg, Germany13Present address: Department of Nutritional Science and Food Management, Ewha Womans University, 120-750 Seoul, Republic of Korea14Present address: Graduate School of Cancer Science and Policy, National Cancer Center, 410-769 Goyang-si, Republic of Korea
*Correspondence: [email protected] (I.G.), [email protected] (C.N.), [email protected] (C.P.)
http://dx.doi.org/10.1016/j.molcel.2014.06.031
SUMMARY
DNAmethylation is a dynamic and reversible processthat governs gene expression during developmentand disease. Several examples of active DNAdemethylation have been documented, involvinggenome-wide and gene-specific DNAdemethylation.How demethylating enzymes are targeted to specificgenomic loci remains largely unknown.We show thatan antisense lncRNA, termed TARID (for TCF21antisense RNA inducing demethylation), activatesTCF21 expression by inducing promoter demethyla-tion. TARID interacts with both the TCF21 promoterand GADD45A (growth arrest and DNA-damage-inducible, alpha), a regulator of DNA demethylation.GADD45A in turn recruits thymine-DNA glycosylasefor base excision repair-mediated demethylationinvolvingoxidationof 5-methylcytosine to 5-hydroxy-methylcytosine in the TCF21 promoter by ten-eleventranslocation methylcytosine dioxygenase proteins.The results reveal a function of lncRNAs, serving asa genomic address label for GADD45A-mediateddemethylation of specific target genes.
INTRODUCTION
Long non-coding RNAs (lncRNAs) are key regulators of chromatin
structure, affecting epigenetic states and expression levels of
many target genes (Rinn and Chang, 2012; Ulitsky and Bartel,
604 Molecular Cell 55, 604–614, August 21, 2014 ª2014 Elsevier Inc.
2013). lncRNAs mediate chromatin-based gene regulation
through interactionswith histonemodifiers, chromatin remodeling
complexes, transcriptional regulators, or theDNAmethylationma-
chinery. Like other epigenetic modifications, DNA methylation is
dynamic and reversible. DNAdemethylation occurs through either
inhibition of the maintenance methyltransferase DNMT1 or by
active enzymatic reactions, leading to loss of methylation in
response to developmental or environmental signals (Bergman
andCedar,2013;WuandZhang,2011).ActiveDNAdemethylation
involvesoxidation ofmethyl groups via the TET family ofmethylcy-
tosine dioxygenases (Guo et al., 2011; He et al., 2011; Kriaucionis
andHeintz, 2009; Tahiliani et al., 2009). The DNA repairmachinery
playsan importantpart inDNAdemethylation, involvingactivation-
induced cytidine deaminase (AICDA) and thymine-DNA glycosy-
lase (TDG) (Bhutani et al., 2010; Cortazar et al., 2011; Kangaspe-
ska et al., 2008; Metivier et al., 2008; Rai et al., 2008). An adaptor
protein that tethers the nucleotide excision repair (NER) and
base excision repair (BER) machineries to sites of DNA demethy-
lation is the stress response protein GADD45A (growth arrest and
DNA-damage-inducible, alpha). GADD45A interacts with compo-
nents of DNA repair complexes, including XPG and TDG, thereby
recruiting DNA repair complexes to specific sites and resulting in
the replacement of methylated cytosines by unmethylated cyto-
sines (Barreto et al., 2007;Cortellino et al., 2011; Niehrs and Scha-
fer, 2012; Schmitz et al., 2009). While GADD45A binds to
promoters undergoing demethylation (Barreto et al., 2007; Cortel-
lino et al., 2011; Schafer et al., 2013; Schmitz et al., 2009), it re-
mains unresolved how specific promoters are targeted.
Epigenetic silencing of tumor suppressor genes is a well-
known phenomenon in tumorigenesis, but themolecular mecha-
nisms underlying transcriptional repression are poorly under-
stood. We have studied epigenetic regulation of the tumor

B
500 bp
10 20 30
pGL4
Luciferase activity (FL/RL)
D
A
CGI
TCF21 1kb
HN
SCC
OVC
T
N
T
N
T
N
a b c dAmplicon
NSC
LC
AS S
F1F2F3F4F5F6SV40
1 2 3
40
+427 -1672
TCF21 1kb
1 2 3
100%
0%D
NA
met
hyla
tion
+1
**
**
Luc
cons
truct
s
654
21
tota
l RN
A
poly
(A) R
NA
kb
C
T1T2T3T4T5
TARID
TCF21 1kb
12345
L i ii
RACERT-PCR
0.6
kb
Figure 1. TARID Is an Intergenic Long Noncod-
ing RNA
(A) DNA methylation analysis of TCF21 in tumor
samples. Heatmap of DNA methylation for CGI1,
CGI2, and CGI3 in amplicons (a–d) analyzed by
MassARRAY. Each square represents a single CpG or
a group of two or three CpG units analyzed, each row
representing a tissue sample.Methylation frequencies
extend from yellow (0%) to blue (100%). Samples
include tumor (T) or normal (N) tissue DNAs from pa-
tients with non-small cell lung cancer (NSCLC), head
and neck squamous cell carcinomas (HNSCC), and
ovarian cancers (OVC). A scheme representing the
TCF21 locus including the transcription start site
(arrow), coding exons (black boxes), 50 and 30 un-
translated regions (white boxes), and CpG islands 1–3
(green bars) is displayed above. See also Figure S1.
(B) Diagram showing the relative promoter activity of
TCF21 fragments cloned in sense (S) and antisense
(AS) orientation into the pGL4 reporter plasmid. pGL4-
SV40 (SV40) was used as positive control. The relative
luciferase activities (FL/RL) over vector control (pGL4)
are displayed. Data are represented as mean ± SD of
six independent experiments (n = 6). Constructs
yielding significantly increased promoter activity over
pGL4 (t test, p < 0.05) are marked (*).
(C) RT-PCR products from cDNAs generated using
oligo(dT) (right, lane i) or a TARID-specific primer (lane
ii). The genomic structure ofTARID splice variants (T1–
T5) is shown at the right. Primers used for RACE are
indicated by green and red arrows.
(D) Northern blot of RNA from primary skin fibroblasts
using 10 mg of total RNA or 500 ng of poly(A)-enriched
RNA and a TARID-specific RNA probe.
Molecular Cell
LncRNA Demethylates the Tumor Suppressor TCF21
suppressor, transcription factor 21 (TCF21, also known as
CAPSULIN/POD1/EPICARDIN), a tissue-specific, basic helix-
loop-helix transcription factor regulating themetastasis suppres-
sor KISS1 (Arab et al., 2011). TCF21 is essential for embryonic
development (Cui et al., 2004; Lu et al., 2002; Quaggin et al.,
1999) and shows frequent aberrations in human cancers (Arab
et al., 2011; Smith et al., 2006; Tessema et al., 2008).TCF21 is en-
coded by three exons, each associated with a CpG island (CGI).
In cancer cells the promoter CGI is hypermethylated, leading to
epigenetic silencing of TCF21 (Arab et al., 2011; Richards et al.,
2011; Tessema et al., 2008). Here we provide evidence that the
third CGI of TCF21 harbors a promoter that directs transcription
of an unknown lncRNA in antisense orientation to TCF21. This
lncRNA, termed TARID, activates TCF21 transcription by
inducing TET protein-dependent DNA demethylation. We show
that TARID binds to the TCF21 promoter and recruits GADD45A
and TDG to direct base excision repair for demethylation. Our
work reveals a function of lncRNAs in epigenetic regulation to
serve as an address label for gene-specific DNA demethylation.
RESULTS
Concordant Promoter Methylation and Expressionof TCF21 and TARID
TCF21 is encoded by three exons, all being associated with a
CpG island, termed CGI1, CGI2, and CGI3 (Figure 1A). Aberrant
methylation of the CGI1 correlates with epigenetic silencing of
Mo
TCF21 in multiple human malignancies (Arab et al., 2011; Smith
et al., 2006). Analysis of DNAmethylation revealed that CGI1 and
CGI3, but not CGI2, were hypermethylated in samples from pa-
tients with non-small cell lung cancer, head and neck squamous
cell carcinomas, and ovarian cancers, suggesting a regulatory
mechanism that coordinates methylation and transcription of
CGI1 and CGI3 (Figures 1A and S1 available online). To examine
whether CGI3 might represent an alternative transcription start
site (TSS) of TCF21, we performed luciferase reporter assays
with constructs that cover CGI3 sequences in sense and anti-
sense orientation. A 554 bp fragment comprising the TCF21 frag-
ment in antisense orientation (F3) directed luciferase activity,
delineating the minimal active promoter and demonstrating
that CGI3 harbors a promoter that directs the synthesis of anti-
sense transcripts (Figure 1B). RT-PCR revealed that the TCF21
antisense transcripts are differentially spliced, yielding five
PCR products (T1–T5) comprising the TSS, the first exon 1 and
the last exon, but differing in the region that overlaps the
TCF21 promoter (Figure 1C). Exon-intron junctions in all splice
variants follow canonical splice-donor and splice-acceptor sites
(Table S1). Rapid amplification of cDNA ends (RACE) of these
transcripts, hereafter referred to as TARID (for TCF21 antisense
RNA inducing demethylation), identified the 50 end at
position +3,682 and the 30 end at position �4,399 relative to
the TCF21 transcription start site (Figure 1C). Accordingly, the
major transcript (T2) has a length of about 4.5 kb on northern
blots (Figure 1D). Consistent with a role as a nuclear lncRNA,
lecular Cell 55, 604–614, August 21, 2014 ª2014 Elsevier Inc. 605

TCF21TARID
RN
A le
vels
0.03
0.06
WT MUT
MUT 1kbTCF21
A
TARID1 2 10 20 100 LNA
(nM)100Ctrl
RN
A le
vels TCF21TARID
0.04
0.08
0.12
D
Amplicon a
500bp TCF21
ba‘
0.20.40.60.81.0
DN
A m
ethy
latio
n
Ctrl LNATARID LNA
B
C500bp
Fold
enr
ichm
ent (
Chi
p/Ig
G)
C1 C2 C3 C4 C5 C6
C1 C2 C3 C4 C5 C6 ACTB
TCF21
10
20
30
1020
30
40RNAPII
p300
*
*
Ctrl LNA TARID LNA
5 7 8 910 11 141 2 3-4CpG
100bp
CpG 1 2 3-4 5 7 8 9 10 11 14
0.20.40.60.81.0
DN
A m
ethy
latio
n
Figure 2. TARID Is Required for TCF21
Expression
(A) Relative levels of TARID and TCF21 mRNA in
HEK293 cells transfected with a BAC encoding
wild-type TCF21 (WT, HEK293TARIDWT) or amutant
BAC carrying a deletion of the TARID promoter
(MUT, HEK293TARIDMUT). Data are represented
as mean ± SD of three independent experiments
(n = 3).
(B) Relative levels of TARID and TCF21 mRNA
after locked nucleic acid (LNA)-mediated knock-
down of TARID in human primary skin fibroblasts
(48 hr). Scrambled control (Ctrl) LNA was used at
100 nM. RNA levels were normalized to HPRT1
and SDHA mRNAs. Data are represented as
mean ± SD of three independent experiments
(n = 3).
(C) Chromatin immunoprecipitation showing
RNAP II and p300 occupancy at the TCF21 locus
after knockdown of TARID in primary human skin
fibroblasts. Locations of amplicons (C1–C6) are
indicated in the scheme above. Values represent
the enrichment of bound protein fractions relative
to input. Data are represented as mean ± SD of
six independent experiments (n = 6). The asterisk
marks significant enrichment over control (t test,
p < 0.05).
(D) DNA methylation at the TCF21 promoter in
human primary skin fibroblasts after knockdown
of TARID with LNA (48 hr). The MassARRAY data
of amplicons comprising the TCF21 promoter is
shown below. Data are represented as mean ±
SD of three independent experiments (n = 3).
Molecular Cell
LncRNA Demethylates the Tumor Suppressor TCF21
TARID is localized preferentially in the nucleus and does not
encode an open reading frame longer than 85 amino acids
(Figures S2A and S2B). The coding potential was tested by Fick-
ett’s algorithm (Fickett, 1982), yielding test scores below values
commonly observed for coding transcripts (<0.95). Moreover,
computer-based prediction of the RNA structure suggested
that TARID exhibits a thermodynamically strong secondary
structure, a common feature of lncRNAs (Figures S2B and S2C).
In normal lung, oral, and ovarian epithelium, CGI1 and CGI3
are unmethylated and both TCF21 and TARID are expressed.
In corresponding tumor samples, CGI1 and CGI3 are methylated
and both TCF21 and TARID are silent. The correlation between
both TCF21 and TARID expression levels and methylation of
their corresponding promoter CGIs suggested that TARID acti-
vates TCF21 expression by decreasing promoter methylation
(Figures S3A and S3B).
TARID Activates TCF21 Transcription by InducingDemethylation of the TCF21 PromoterAs some lncRNAs alter epigenetic signatures through interac-
tions with chromatin remodeling enzymes (Gupta et al., 2010;
Li et al., 2013; Rinn and Chang, 2012; Zhao et al., 2008), we es-
tablished human embryonic kidney 293 cell lines (HEK293TARIDWT
and HEK293TARIDMUT) that harbor a bacterial artificial chromo-
some comprising the TCF21 locus or a mutated version lacking
the TARID promoter region (Figure S3C). In several subclones,
HEK293TARIDWT cells expressed both TCF21 and TARID,
whereas in HEK293TARIDMUT cells the level of both RNAs was
606 Molecular Cell 55, 604–614, August 21, 2014 ª2014 Elsevier Inc.
strongly reduced (Figure 2A). The positive correlation between
TARID and TCF21 mRNA suggested that expression of TCF21
is functionally linked to transcription of TARID (Figure S3D).
Indeed, knockdown of TARID in primary skin fibroblasts (ex-
pressing both TCF21 and TARID) led to decreased expression
of TCF21 (Figure 2B) and decreased occupancy of both
RNAP II and the histone acetyltransferase p300 at the TCF21
promoter (Figure 2C). In support of TARID being required for
maintaining the TCF21 promoter hypomethylated, knockdown
of TARID led to a 30% gain of DNA methylation around the
TSS of TCF21 (Figure 2D). To test whether elevated levels of
TARID would affect promoter methylation and TCF21 expres-
sion, we transfected all TARID splice variants (T1–T5) into three
cancer cell lines (H387, C8161, and A549) in which expression
of TCF21 is repressed by promoter hypermethylation. Expres-
sion of TCF21 was induced after transfection of transcripts T1
and T2, whereas transcripts T3, T4, and T5 did not activate
transcription. For comparison, TCF21 expression levels in
normal lung fibroblast, epithelial cells, and melanocytes are dis-
played in Figure 3A. Thus, sequences that promote transcrip-
tional activation of TCF21 are contained within exon 2 of TARID.
Transfection of TARID splice variant T2 resulted in a significant
increase of RNAP II, p300, and H3K4me3 at the TCF21 pro-
moter (Figure 3B). Moreover, ectopic TARID triggered demethy-
lation of the TCF21 promoter in all cancer cell lines assayed.
Demethylation was very specific, being restricted to a small re-
gion comprising the TSS, whereas no change was observed in
flanking sequences (Figure 3C). While TARID regulates TCF21,

A
T1 T2 T3 T4 T5TARID
0.02
0.01
Ctrl
TCF2
1 m
RN
A le
vel
B
*
252015105
15
5
10
40
30
20
10
*
* ** *
H3K4me3
p300
RNAPII
Fol
d en
richm
ent (
Chi
p/Ig
G)
H387C8161A549
C
5 7 8 910 11 141 2 3-4
A549
CpG
100bp
C8161
H387
DN
A m
ethy
latio
n 1.00.80.60.40.2
1.00.80.60.40.2
1.00.80.60.40.2
C1 C2 C3 C4 C5 C6 ACTB
EctopicRNA
T1T2T3T4T5
TARID
TCF21 1kb
500bp
C1 C2 C3 C4 C5 C6
TCF21
Ctrl RNATARID
CpG 1 2 3-4 5 7 8 9 10 11 14
Ctrl RNA TARID
Amplicon a
500bp TCF21
0.04
0.08
0.12
LF EC ML
TCF2
1 m
RN
A le
vel
Figure 3. TARID Activates TCF21 by Pro-
moter Demethylation
(A) Left: relative levels of TCF21 expression in lung
fibroblast (LF), epithelial cells (EC), and melano-
cytes (ML). Middle: TCF21 expression in H387,
C8161, and A549 cancer cells transfected with
in vitro-synthesized TARID splice variants (T1–T5).
Right: scheme of TARID splice variants high-
lighting the minimal region required for TCF21
activation (shaded). RNA levels were normalized to
HPRT1 and SDHA mRNAs. Data are represented
as mean ± SD of three independent experiments
(n = 3).
(B) ChIP assaymonitoring the association of RNAP
II, p300, and H3K4me3 with different regions of
TCF21 in H387 cells transfectedwith TARID (splice
variant T2) or control RNA (Ctrl RNA). Values
represent enrichment of bound protein fraction
relative to input. The locations of amplicons (C1–
C6) are indicated in the scheme above. Actin B
(ACTB) was used as positive control. Data are
represented as mean ± SD of four independent
experiments (n = 4, *p < 0.05).
(C) DNA methylation levels around the TSS of
TCF21 in H387, C8161, and A549 cells transfected
with TARID (splice variant T2) or with control RNA
(Ctrl RNA). The position of CpG residues in
amplicon a (indicated by lollipops) is shown above.
Data are represented as mean ± SD of three in-
dependent experiments (n = 3).
Molecular Cell
LncRNA Demethylates the Tumor Suppressor TCF21
the reverse was not true, since no significant change in TARID
expression and promoter methylation was observed upon
TCF21 siRNA knockdown (data not shown).
TARID-Mediated Demethylation Requires GADD45Aand TDGPrevious results have established that GADD45A, a stress
response protein involved in growth control, genomic stability,
DNA repair, and apoptosis, influences epigenetic gene regula-
tion by promoting active DNA demethylation (Barreto et al.,
2007). As GADD45A binds RNA and shows characteristics of
a ribonucleoprotein particle (Sytnikova et al., 2011), we
reasoned that TARID might guide GADD45A to the TCF21 pro-
moter. In support of GADD45A being involved in activation of
TCF21, knockdown of GADD45A inhibited TARID-mediated de-
methylation of the TCF21 promoter and prevented TCF21
expression in H387, C8161, and A549 tumor cells (Figures 4A
and 4B). GADD45A has been shown to recruit the nucleotide
excision repair (NER) or deamination-base excision repair
(BER) machinery, which removes methylated cytosine residues
(Barreto et al., 2007; Cortellino et al., 2011; Schmitz et al.,
2009). To examine whether demethylation of TCF21 is medi-
ated by NER or BER, we monitored TARID-mediated TCF21
expression after knockdown of XPA, XPC, XPF and XPG
(essential components of the NER pathway), and TDG
Molecular Cell 55, 604–614
(thymine-DNA glycosylase), an essential
component of the BER pathway. While
expression of TCF21 was clearly
reduced upon TDG knockdown, knock-
down of XPA, XPC, XPF, and XPG did not affect TCF21 expres-
sion (Figure 4C). To approve that active demethylation is
brought about by GADD45A-dependent recruitment of the
BER machinery, we treated cells with low doses of the BER in-
hibitors CRT0044876, betulinic acid, and ABT888 and moni-
tored TCF21 expression. All three BER inhibitors prevented
TARID-dependent expression of TCF21, whereas GADD45A
was not affected (Figure 4D). Moreover, coimmunoprecipitation
experiments confirmed the association of TDG with GADD45A
in H387 cells (Figure 4E). In addition, TARID-mediated DNA de-
methylation and expression of TCF21 were inhibited upon
knockdown of TDG (Figure 4F), indicating that TDG cooperates
with TARID to activate TCF21 expression.
TARID Is Bound by GADD45A and Interacts with theTCF21 PromoterTo examine whether TARID associates with GADD45A, we
incubated bead-bound GADD45A with radiolabeled RNA
comprising different regions of TARID and monitored
GADD45A-associated RNA (Figures 5A and S4). These pull-
down assays revealed that GADD45A interacts with full-length
TARID (RNA1) and with a truncated version comprising exon 2,
the region that is complementary to the TSSofTCF21. Fragments
comprising the first exon of TARID, intronic sequences, and
fragments lacking the TSS overlapping region did not bind
, August 21, 2014 ª2014 Elsevier Inc. 607
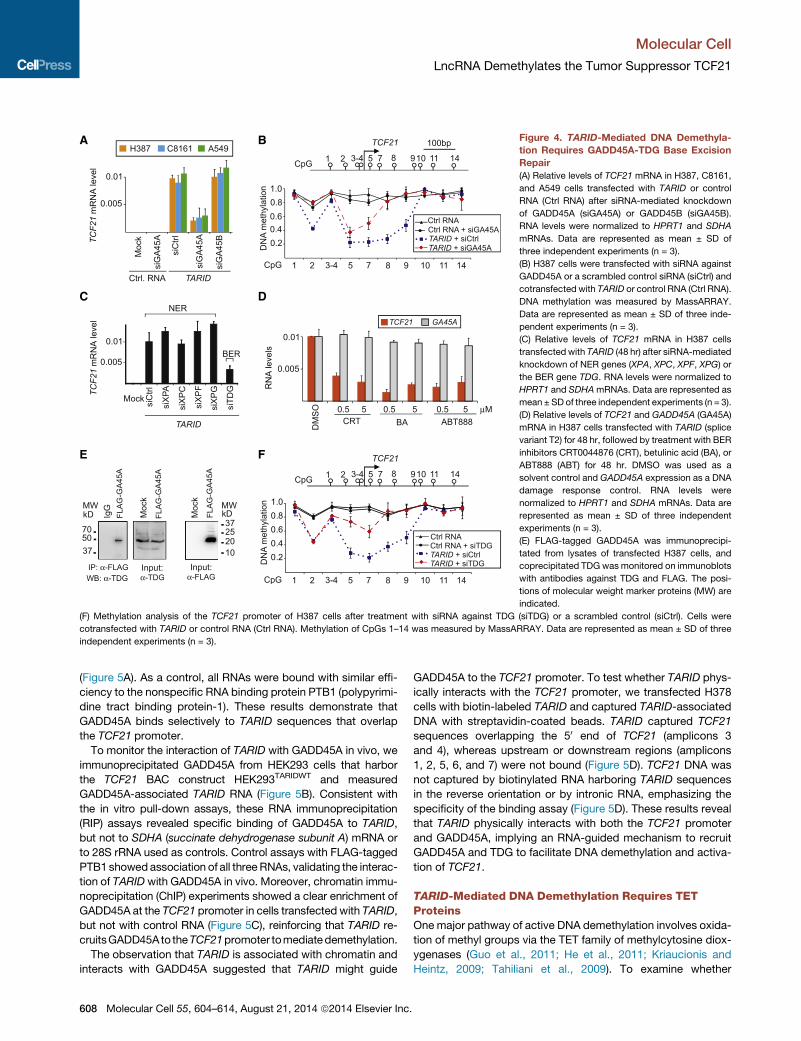
A B
C D
E F
Figure 4. TARID-Mediated DNA Demethyla-
tion Requires GADD45A-TDG Base Excision
Repair
(A) Relative levels of TCF21 mRNA in H387, C8161,
and A549 cells transfected with TARID or control
RNA (Ctrl RNA) after siRNA-mediated knockdown
of GADD45A (siGA45A) or GADD45B (siGA45B).
RNA levels were normalized to HPRT1 and SDHA
mRNAs. Data are represented as mean ± SD of
three independent experiments (n = 3).
(B) H387 cells were transfected with siRNA against
GADD45A or a scrambled control siRNA (siCtrl) and
cotransfected with TARID or control RNA (Ctrl RNA).
DNA methylation was measured by MassARRAY.
Data are represented as mean ± SD of three inde-
pendent experiments (n = 3).
(C) Relative levels of TCF21 mRNA in H387 cells
transfectedwith TARID (48 hr) after siRNA-mediated
knockdown of NER genes (XPA, XPC, XPF, XPG) or
the BER gene TDG. RNA levels were normalized to
HPRT1 and SDHAmRNAs. Data are represented as
mean ± SDof three independent experiments (n = 3).
(D) Relative levels of TCF21 and GADD45A (GA45A)
mRNA in H387 cells transfected with TARID (splice
variant T2) for 48 hr, followed by treatment with BER
inhibitors CRT0044876 (CRT), betulinic acid (BA), or
ABT888 (ABT) for 48 hr. DMSO was used as a
solvent control and GADD45A expression as a DNA
damage response control. RNA levels were
normalized to HPRT1 and SDHA mRNAs. Data are
represented as mean ± SD of three independent
experiments (n = 3).
(E) FLAG-tagged GADD45A was immunoprecipi-
tated from lysates of transfected H387 cells, and
coprecipitated TDG was monitored on immunoblots
with antibodies against TDG and FLAG. The posi-
tions of molecular weight marker proteins (MW) are
indicated.
(F) Methylation analysis of the TCF21 promoter of H387 cells after treatment with siRNA against TDG (siTDG) or a scrambled control (siCtrl). Cells were
cotransfected with TARID or control RNA (Ctrl RNA). Methylation of CpGs 1–14 was measured by MassARRAY. Data are represented as mean ± SD of three
independent experiments (n = 3).
Molecular Cell
LncRNA Demethylates the Tumor Suppressor TCF21
(Figure 5A). As a control, all RNAs were bound with similar effi-
ciency to the nonspecific RNA binding protein PTB1 (polypyrimi-
dine tract binding protein-1). These results demonstrate that
GADD45A binds selectively to TARID sequences that overlap
the TCF21 promoter.
To monitor the interaction of TARID with GADD45A in vivo, we
immunoprecipitated GADD45A from HEK293 cells that harbor
the TCF21 BAC construct HEK293TARIDWT and measured
GADD45A-associated TARID RNA (Figure 5B). Consistent with
the in vitro pull-down assays, these RNA immunoprecipitation
(RIP) assays revealed specific binding of GADD45A to TARID,
but not to SDHA (succinate dehydrogenase subunit A) mRNA or
to 28S rRNA used as controls. Control assays with FLAG-tagged
PTB1showedassociation of all threeRNAs, validating the interac-
tion of TARID with GADD45A in vivo. Moreover, chromatin immu-
noprecipitation (ChIP) experiments showed a clear enrichment of
GADD45A at the TCF21 promoter in cells transfected with TARID,
but not with control RNA (Figure 5C), reinforcing that TARID re-
cruitsGADD45Ato theTCF21promoter tomediatedemethylation.
The observation that TARID is associated with chromatin and
interacts with GADD45A suggested that TARID might guide
608 Molecular Cell 55, 604–614, August 21, 2014 ª2014 Elsevier Inc.
GADD45A to the TCF21 promoter. To test whether TARID phys-
ically interacts with the TCF21 promoter, we transfected H378
cells with biotin-labeled TARID and captured TARID-associated
DNA with streptavidin-coated beads. TARID captured TCF21
sequences overlapping the 50 end of TCF21 (amplicons 3
and 4), whereas upstream or downstream regions (amplicons
1, 2, 5, 6, and 7) were not bound (Figure 5D). TCF21 DNA was
not captured by biotinylated RNA harboring TARID sequences
in the reverse orientation or by intronic RNA, emphasizing the
specificity of the binding assay (Figure 5D). These results reveal
that TARID physically interacts with both the TCF21 promoter
and GADD45A, implying an RNA-guided mechanism to recruit
GADD45A and TDG to facilitate DNA demethylation and activa-
tion of TCF21.
TARID-Mediated DNA Demethylation Requires TETProteinsOnemajor pathway of active DNA demethylation involves oxida-
tion of methyl groups via the TET family of methylcytosine diox-
ygenases (Guo et al., 2011; He et al., 2011; Kriaucionis and
Heintz, 2009; Tahiliani et al., 2009). To examine whether

D
A C
Rel
ativ
e fo
ld e
nric
hmen
t(o
ver m
ock)
1 2 3 4 5 6 7 ACTB
B
12 3
5 6
4
1kbTCF21
GA
DD
45A
1
2
3
4
5
TARIDTARID rev
Ctrl RNA
2 34 5 6 7
1kbTCF21
1
500bp
TARID revCtrl RNA
TARID
GADD45A PTB1
C1 C2 C4 C6
GADD45A-ChIP
C1 C2 C4 C6 GAPDH MYOD1
Bou
nd R
NA
(% to
Inpu
t)
TCF21
4
8
12
16 *
*
**
Fold
enr
ichm
ent (
Chi
P/Ig
G)
Ctrl RNATARID
DNA capture assay
GADD45A IgG
RNA 1 2 3 4 5 6
1
2
3
4
5
RNAs
RNA
28S rRNATARID SDHA mRNA
Bou
nd R
NA
(% to
inpu
t)
Bou
nd R
NA
(% to
inpu
t)
PTB1 IgG
1
2
3
4
5
6
1 2 3 4 5 6
1
2
3
4
5
6
Inpu
t 5%
PTB
1
Figure 5. TARID Targets GADD45A to the
TCF21 Promoter
(A) Pull-down assay using immobilized FLAG-tag-
gedGADD45AorPTB1 incubatedwith radiolabeled
RNAs. Genomic regions encoding RNA fragments
1–6 relative to TCF21 is shown above. Bound RNA
was quantified by liquid scintillation counting (left)
followed by gel electrophoresis and autoradiog-
raphy (right). Seealso FigureS4. Theminimal region
in TARID that interacts with GADD45A is shaded.
Data are represented as mean ± SD of three inde-
pendent experiments (n = 3).
(B) RNA immunoprecipitation (RIP) of FLAG-tag-
ged GADD45A (top) and FLAG-tagged PTB1 (bot-
tom) transfected into HEK293TCF21WT cells. Levels
of TARID, SDHAmRNA, and 28S rRNA associated
with precipitated proteins were determined by
qRT-PCR and displayed as bound RNA relative to
input. Data are represented as mean ± SD of three
independent experiments (n = 3).
(C) Chromatin immunoprecipitation assays show-
ing the enrichment of GADD45A at TCF21 in H387
cells transfected with TARID or control RNA. Co-
precipitated DNA was analyzed by qPCR using
ampliconsC1–C5. Data are represented asmean±
SD of three independent experiments (n = 3, t test,
p < 0.05).
(D) DNA capture assay using H387 cells trans-
fected with biotin-labeled RNAs shown above.
Biotinylated RNA was captured with streptavidin-
coated beads, and associated TCF21 DNA
(amplicons 1–7) was analyzed by qPCR. Data are
represented as mean ± SD of three independent
experiments (n = 3, t test, p < 0.05).
Molecular Cell
LncRNA Demethylates the Tumor Suppressor TCF21
TARID-mediated demethylation of the TCF21 promoter is
brought about by TET proteins, we depleted H387 cells from
TET1, TET2, or TET3 by knocking down protein either alone or
in different combinations. Consistent with redundant functions
of individual TET proteins, depletion of all three TET proteins
was required to inhibit TARID-mediated demethylation and
TCF21 expression (Figures 6A and 6B). To monitor changes in
5-hydroxymethylcytosine (5hmC), we assayed 5hmC by quanti-
tative PCR (qPCR) following glucosyl-5-hydroxymethylcytosine-
sensitive MspI restriction digest (Kinney et al., 2011). We found
that hydroxymethylation of CpG#7 at the TCF21 promoter was
increased 5-fold in HEK293TARIDWT cells overexpressing TET1
(Figure S5). Significantly, ectopic TARID increased 5hmC
levels already 1–2 hr after TARID transfection (Figure 6C). In
support of TARID-induced demethylation involving hydroxyme-
thylation of CpG residues, 5hmC levels were 2-fold lower in
HEK293TARIDMUT compared to that in HEK293TARIDWT cells (Fig-
ure 6D, left). Moreover, 5hmC levels were reduced 3-fold after
knockdown of TARID in primary skin fibroblasts (Figure 6D,
right). These results indicate that TET protein(s)-mediated hy-
droxymethylation of CpG residues represents an intermediate
step of TARID-induced demethylation of the TCF21 promoter.
DISCUSSION
Two main mechanisms of DNA demethylation have been docu-
mented in mammals: global DNA demethylation and gene-spe-
Mo
cific DNA demethylation, the latter of which occurs at specific
promoter and enhancer sequences. One of the key questions
in gene-specific DNA demethylation is how locus specificity is
achieved. Because of their sequence specificity, noncoding
RNAs are excellent candidates for targeting protein complexes
to appropriate locations in the genome. In support of this view,
we show that lncRNAs can provide such specificity by binding
the adaptor protein GADD45A, which in turn recruits TDG and
TETs (seemodel Figure 7). TARID and GADD45Amediate deme-
thylation of TCF21 by utilizing BER and TDG, an essential
component of the BER pathway essential for DNA demethylation
(Cortazar et al., 2011; Pastor et al., 2013; Zhu et al., 2000). TDG
can efficiently excise the iterative oxidation products of 5mC and
5hmC, namely 5-formylcytosine (5fC), and 5-carboxylcytosine
(5caC), replacing 5-methyl cytosine with unmethylated cytosine
(He et al., 2011; Ito et al., 2010; Maiti and Drohat, 2011).
Demethylation of TCF21 is restricted to a few CpG dinucleo-
tides at the transcription start site, a characteristic feature of
gene-specific demethylation. Significantly, the part of TARID
that directs demethylation is associated with the TCF21 pro-
moter. The potential of RNA to bind to complementary DNA
sequences has led to the hypothesis that lncRNAs may play
important guiding roles in the establishment and transmission
of chromatin states. TARID may act as a scaffold to bring two
or more proteins into spatial proximity or may act as a guide to
recruit proteins, such as GADD45A to the TCF21 promoter.
This may occur through interactions of TARID with DNA or
lecular Cell 55, 604–614, August 21, 2014 ª2014 Elsevier Inc. 609
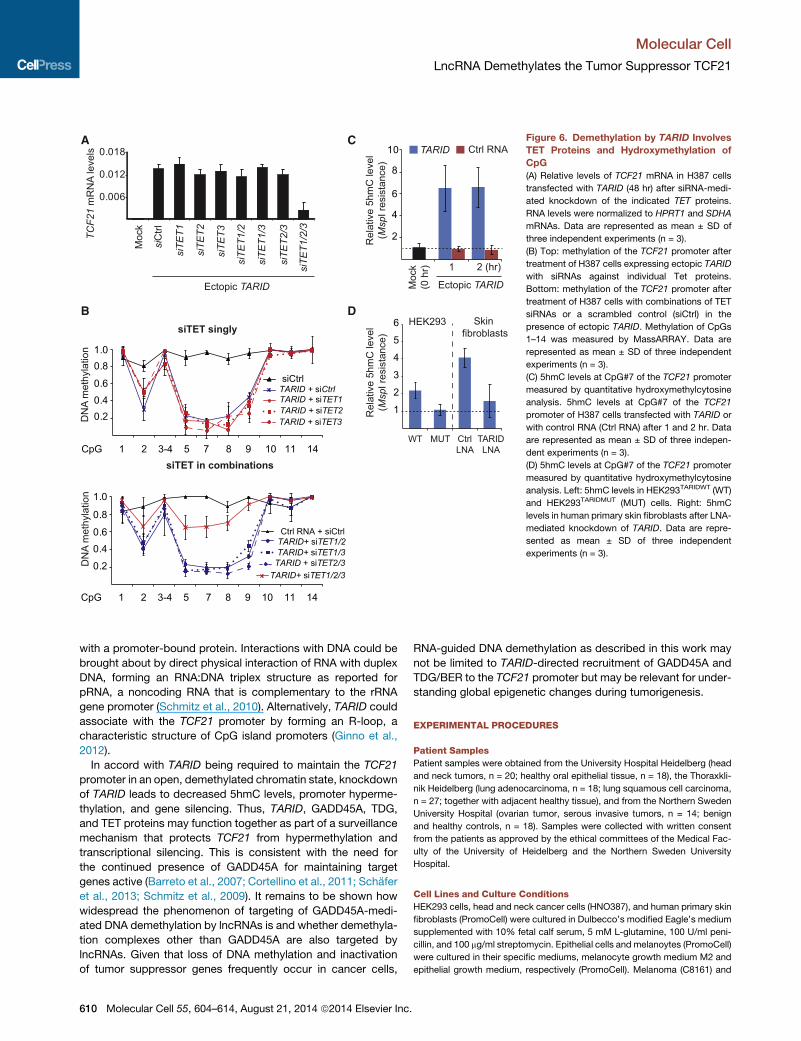
A
B
TCF2
1 m
RN
A le
vels
siC
trl
siTE
T1
Ectopic TARID
Moc
k
siTE
T2
siTE
T1/2
siTE
T1/3
siTE
T2/3
siTE
T1/2
/3
0.006
0.012
0.018
siTE
T3
0.20.4
0.6
0.81.0
TARID + siTET1TARID + siCtrlsiCtrl
TARID + siTET2TARID + siTET3D
NA
met
hyla
tion
siTET singly
siTET in combinations
TARID+ siTET1/3 TARID + siTET2/3TARID+ siTET1/2/3
Ctrl RNA + siCtrlTARID+ siTET1/2
CpG 1 2 3-4 5 7 8 9 10 11 14
CpG 1 2 3-4 5 7 8 9 10 11 14
0.2
0.4
0.6
0.8
1.0
DN
A m
ethy
latio
n
2
4
6
8
10
Rel
ativ
e 5h
mC
leve
l(M
spI r
esis
tanc
e)
Ectopic TARIDMoc
k
(0 h
r) 1 2 (hr)
Ctrl RNATARIDC
D
1
2
3
4
5
6
Rel
ativ
e 5h
mC
leve
l(M
spI r
esis
tanc
e)
HEK293
MUTWT
Skin fibroblasts
TARIDLNA
CtrlLNA
Figure 6. Demethylation by TARID Involves
TET Proteins and Hydroxymethylation of
CpG
(A) Relative levels of TCF21 mRNA in H387 cells
transfected with TARID (48 hr) after siRNA-medi-
ated knockdown of the indicated TET proteins.
RNA levels were normalized to HPRT1 and SDHA
mRNAs. Data are represented as mean ± SD of
three independent experiments (n = 3).
(B) Top: methylation of the TCF21 promoter after
treatment of H387 cells expressing ectopic TARID
with siRNAs against individual Tet proteins.
Bottom: methylation of the TCF21 promoter after
treatment of H387 cells with combinations of TET
siRNAs or a scrambled control (siCtrl) in the
presence of ectopic TARID. Methylation of CpGs
1–14 was measured by MassARRAY. Data are
represented as mean ± SD of three independent
experiments (n = 3).
(C) 5hmC levels at CpG#7 of the TCF21 promoter
measured by quantitative hydroxymethylcytosine
analysis. 5hmC levels at CpG#7 of the TCF21
promoter of H387 cells transfected with TARID or
with control RNA (Ctrl RNA) after 1 and 2 hr. Data
are represented as mean ± SD of three indepen-
dent experiments (n = 3).
(D) 5hmC levels at CpG#7 of the TCF21 promoter
measured by quantitative hydroxymethylcytosine
analysis. Left: 5hmC levels in HEK293TARIDWT (WT)
and HEK293TARIDMUT (MUT) cells. Right: 5hmC
levels in human primary skin fibroblasts after LNA-
mediated knockdown of TARID. Data are repre-
sented as mean ± SD of three independent
experiments (n = 3).
Molecular Cell
LncRNA Demethylates the Tumor Suppressor TCF21
with a promoter-bound protein. Interactions with DNA could be
brought about by direct physical interaction of RNA with duplex
DNA, forming an RNA:DNA triplex structure as reported for
pRNA, a noncoding RNA that is complementary to the rRNA
gene promoter (Schmitz et al., 2010). Alternatively, TARID could
associate with the TCF21 promoter by forming an R-loop, a
characteristic structure of CpG island promoters (Ginno et al.,
2012).
In accord with TARID being required to maintain the TCF21
promoter in an open, demethylated chromatin state, knockdown
of TARID leads to decreased 5hmC levels, promoter hyperme-
thylation, and gene silencing. Thus, TARID, GADD45A, TDG,
and TET proteins may function together as part of a surveillance
mechanism that protects TCF21 from hypermethylation and
transcriptional silencing. This is consistent with the need for
the continued presence of GADD45A for maintaining target
genes active (Barreto et al., 2007; Cortellino et al., 2011; Schafer
et al., 2013; Schmitz et al., 2009). It remains to be shown how
widespread the phenomenon of targeting of GADD45A-medi-
ated DNA demethylation by lncRNAs is and whether demethyla-
tion complexes other than GADD45A are also targeted by
lncRNAs. Given that loss of DNA methylation and inactivation
of tumor suppressor genes frequently occur in cancer cells,
610 Molecular Cell 55, 604–614, August 21, 2014 ª2014 Elsevier Inc.
RNA-guided DNA demethylation as described in this work may
not be limited to TARID-directed recruitment of GADD45A and
TDG/BER to the TCF21 promoter but may be relevant for under-
standing global epigenetic changes during tumorigenesis.
EXPERIMENTAL PROCEDURES
Patient Samples
Patient samples were obtained from the University Hospital Heidelberg (head
and neck tumors, n = 20; healthy oral epithelial tissue, n = 18), the Thoraxkli-
nik Heidelberg (lung adenocarcinoma, n = 18; lung squamous cell carcinoma,
n = 27; together with adjacent healthy tissue), and from the Northern Sweden
University Hospital (ovarian tumor, serous invasive tumors, n = 14; benign
and healthy controls, n = 18). Samples were collected with written consent
from the patients as approved by the ethical committees of the Medical Fac-
ulty of the University of Heidelberg and the Northern Sweden University
Hospital.
Cell Lines and Culture Conditions
HEK293 cells, head and neck cancer cells (HNO387), and human primary skin
fibroblasts (PromoCell) were cultured in Dulbecco’s modified Eagle’s medium
supplemented with 10% fetal calf serum, 5 mM L-glutamine, 100 U/ml peni-
cillin, and 100 mg/ml streptomycin. Epithelial cells and melanoytes (PromoCell)
were cultured in their specific mediums, melanocyte growth medium M2 and
epithelial growth medium, respectively (PromoCell). Melanoma (C8161) and

Figure 7. Model Depicting the Role of TARID in Regulation of TCF21
In cancer cells, the promoters of both TCF21 and TARID are frequently hypermethlyated, and both genes are silent. In normal cells, TARID lncRNA is expressed
and associates with the TCF21 promoter. TARID bindsGADD45A and recruits TDG together with TETs to induce promoter demethylation via base excision repair.
Demethylated TCF21 harbors RNAP II and active chromatinmarks H3K4me3, permitting TCF21 expression. Methylated and unmethylated CpGs are indicated by
black and white circles, respectively.
Molecular Cell
LncRNA Demethylates the Tumor Suppressor TCF21
lung cancer cells (A549) were cultured in RPMI 1640 medium. HEK293 cells
were transfected with either wild-type (RP11-706B15, carrying the TCF21
locus) or mutant (see below) BAC clone using the TransIT reagent (Mirus). Cells
were subjected to G418 selection (400 mg/ml; Sigma-Aldrich), and single cell-
derived clones were established by dilution seeding of G418-resistant col-
onies. DNAs and RNAs were isolated from cells or tissues using TRIzol (Life
Technologies).
Plasmids and BAC Constructs
RP11-706B15 (TARIDWT), a BAC clone carrying a 183 kb fragment including
the TCF21 locus, was received from BACPAC Resources Center (http://
bacpac.chori.org/). RP11-706B15-MUT-NEO (TARIDMUT-NEO) was gener-
ated by recombination, replacing the TARID promoter (GRCh/hg19, position
chr6: 134,213,747–134,214,300, termed F3) by the Neomycin resistance
(Neo) cassette. The Neo cassette was deleted by arabinose-induced flpe re-
combinase, yielding RP11-706B15-MUT (TARIDMUT) lacking the TARID pro-
moter. For the preparation of stable cell lines, all three constructs were
engineered to contain a Neo cassette 22 kb downstream of F3. The TARID
expression vector was generated by RT-PCR amplification of TARID cDNA,
using RNA from primary human skin fibroblasts and cloned into the Gateway
pT-REX-DEST30 vector (Life Technologies). 50 RACE and 30 RACE for TARID
were conducted as described (Frohman, 1993) using 5 mg of total RNA. The
genomic localization of the TARID isoforms (T1–T5) at chromosome 6mapped
between positions 134,213,940 and 134,205,890. Plasmids for luciferase re-
porter assay were generated by PCR amplification of DNA from primary hu-
man skin fibroblasts using primers with built in KpnI and XhoI restriction sites
allowing directional cloning into pGL4.10[luc2] (Promega). The genomic coor-
dinates of the luciferase reporter inserts are listed in Table S2.
Mo
Chromatin Immunoprecipitation and Coimmunoprecipitation
Assays
Cells were crosslinked with 1% formaldehyde at room temperature for 10 min;
quenched with 0.5 M glycine; lysed in 1% SDS, 10 mM EDTA, 50 mM Tris-HCl
(pH 8.1); and sonicated to yield 150–250 bp DNA fragments. Chromatin was
diluted 10-fold with IP buffer (16.7 mM Tris-HCl [pH 8.1], 167 mM NaCl,
1.2mMEDTA, 0.01%SDS, 1%Triton X-100), precleared for 1 hr at 4�Con pro-
tein A/G agarose in the presence of 20 mg/ml sonicated salmon sperm DNA,
and incubated overnight with the respective antibodies. Protein-DNA com-
plexes were captured on protein A/G agarose followed by two washes in
low salt buffer (150mMNaCl, 50mMTris-HCl [pH 8.0], 5 mMMgCl2, 1% Triton
X-100), high salt buffer containing 500 mM NaCl, and with LiCl buffer (250 mM
LiCl, 10 mM Tris-HCl [pH 8.0], 5 mM EDTA, 0.5% Na-deoxycholate, 0.5%
Triton X-100) and TE buffer. After elution and reversal of the crosslink, DNA
was extracted and amplified by qPCR. The amount of precipitated DNA was
normalized to input DNA and to immunoglobulin G (IgG)-bound DNA. For
coimmunoprecipitation assays, FLAG-tagged GADD45A or FLAG-tagged
PTB overexpressed in HNO387 cells were affinity purified by binding to M2
beads (Sigma-Aldrich). After elution with the FLAG peptide, coprecipitated
proteins were detected by immunoblotting.
RNA Immunoprecipitation
HEK293 cells expressing TARID were transfected with GADD45 expression
vector (pCS2+-FLAG-GADD45A), harvested after 48 hr, and lysed in IP buffer
containing 200 mM NaCl, 5 mM MgCl2, 10 mM HEPES (pH 7.0), 0.5% Non-
idet P-40, 1 mM dithiothreitol (DTT), 100 U/ml RNasin. After digestion with
DNase I (5 U/ml, 20 min) and centrifugation, the supernatant was cleared
by addition of 50 ml protein A beads (Sigma-Aldrich) and rotation for 1 hr
lecular Cell 55, 604–614, August 21, 2014 ª2014 Elsevier Inc. 611

Molecular Cell
LncRNA Demethylates the Tumor Suppressor TCF21
at 4�C. The cleared lysate was incubated with M2 beads (Sigma-Aldrich) for
4 hr at 4�C. After stringent washing, coprecipitated RNA was extracted with
TRIzol and analyzed by qRT-PCR. IgGs were used as a negative control.
DNA Capture Assay
HNO387 cells were transfected with 2 pmoles of biotin- and 4-thiouridine-
labeled TARID using the TransIT transfection reagent (Mirus). After 24 hr, cells
were UV irradiated at 2,500 kJ (365 nm, 40 s) and lysed in hypotonic buffer
(10 mM HEPES [pH 7.9], 10 mM KCl, 340 mM sucrose, 10% glycerol, 1 mM
DTT, 1.5 mM MgCl2, 100 U/ml RNase inhibitor). Isolated nuclei were lysed in
10 mM Tris-HCl (pH 7.4), 150 mM NaCl, 5 mM MgCl2, 0.5% NP-40, 1 mM
DTT, 100 U/ml RNasin. Chromatin was sheered by sonication, centrifuged
(12,000 3 g, 10 min, 4�C), and the supernatant was incubated for 1.5 hr at
room temperature with streptavidin-coated magnetic beads (Pierce, Thermo
Scientific) that were blocked for 3 hr with salmon spermDNA (5 mg/10 ml beads)
and BSA (20 mg/10 ml beads). After treatment with proteinase K (50 ng/10 ml
beads) and stringent washing, RNA was isolated and RNA-associated DNA
was quantified by qPCR.
Locked Nucleic Acids, siRNAs, PCR Primers, Drugs, and Antibodies
Custom designed locked nucleic acids (LNAs) against TARID (GGTGATGG
AAAAGTGGAG) and a scrambled LNA (GGTGTATTGAATGACGTG) were syn-
thesized by Exiqon and transfected by electroporation using Amaxa (Lonza).
siRNAs against GADD45A, GADD45B, XPA, XPC, XPG, XPF, TDG, TET1,
TET2, TET3, and control siRNAs were purchased from Life Technologies
and transfected with Dharmafect (Thermo Scientific). All knockdown experi-
ments showed at least 70% reduction in target gene expression. Primers
were synthesized by Sigma-Aldrich. Sequences of siRNAs and primers used
in this study are listed in Tables S3 and S4. CRT 0044876 and betulinic acid
were purchased from Sigma-Aldrich, ABT-888 from tebu-bio, all the drugs
were diluted in DMSO and added at 0.5 and 10 mM final concentration to the
cell culture media and incubated for 48 hr. The following antibodies were
used: RNA polymerase II (Ab5408, Abcam), p300 (sc-585, Santa Cruz),
TDG (c10996, AssayBioTech), H3K4me3 (pAb-003-050, Diagenode), and
GADD45A (sc-797 Santa Cruz).
RNA Pull-Down Assays and Northern Blot Analysis
FLAG-GADD45A and FLAG-PTB were overexpressed in HEK293TARIDWT cells
and immobilized on FLAG epitope (M2) beads. Beads (10 ml) containing 800 ng
of immobilized proteins were washedwith high salt buffer, equilibrated in bind-
ing buffer (50 mM Tris-HCl [pH 7.9], 10% glycerol, 100 mM KCl, 5 mM MgCl2,
10 mM b-mercaptoethanol, 0.1% NP-40), and incubated with 4 pmoles of32P-labeled RNA (5,000 cpm/ng). Equimolar amounts of RNA and bound pro-
teins were incubated for 1.5 hr at room temperature. After stringent washing,
bound RNA was extracted and subjected to gel electrophoresis and autoradi-
ography. For northern blot analysis, 500 ng of poly(A)-enriched RNA from pri-
mary skin fibroblasts were subjected to gel electrophoresis. After blotting, UV
crosslinking, and prehybridization, the membrane was incubated overnight at
68�C in hybridization buffer (63 SSC, 53 Denhardt’s solution, 0.5% SDS,
400 mg yeast tRNA) with a 463 nt 32P-labeled RNA probe that is complemen-
tary to the 30 end TARID. The genomic coordinates of the RNA northern blot
probe are chr6: 134,207,789–134,208,251 (Table S4). After stringent washing,
hybridization signals were detected by phosphorimaging.
Cell Fractionation
HEK293TARIDWT cells were harvested in cold PBS, washed twice, and incu-
bated in buffer A (HEPES-KOH 50 mM [pH 7.5], 10 mM KCl, 350 mM sucrose,
1 mM EDTA, 1 mM DTT, 0.1% Triton X-100) for 10 min on ice with occasional
shaking. The nuclei were collected by centrifugation (2,0003 g, 5 min), and the
supernatant was saved as the cytoplasmic fraction. The nuclei were washed
twice with buffer A without Triton X-100. RNA was purified using TRIzol and
checked by electrophoresis. Quantitative RT-PCR was used to measure the
enrichment of nuclear and cytoplasmic RNA.
Luciferase Reporter Assays
The Dual-Luciferase Reporter Assay System (Promega) was used to monitor
the promoter activity of the genomic inserts. Transfections were performed us-
612 Molecular Cell 55, 604–614, August 21, 2014 ª2014 Elsevier Inc.
ing TransIT reagent (MoBiTec). HEK293 cells were cotransfected with 50 ng of
the respective reporter constructs and 1 ng pRL-CMV in 384-well plates.
Firefly and Renilla luciferase activity were measured after 48 hr using Spectra
Max M5 plate reader (Molecular Device). The firefly luciferase signal was
normalized against the Renilla signal and against the empty vector pGL4.10
[luc2] (pGL4-basic) as control.
Quantitative DNA Methylation Analysis
DNA methylation was analyzed on bisulfite-converted genomic DNA (EZ DNA
Methylation Kit; Zymo Research) using the MassARRAY system (Sequenom)
as reported (Ehrich et al., 2005). DNA methylation levels were quantified from
mass spectra using Epityper software v.1.2 (Sequenom). The MassARRAY
primers for TCF21 locus are listed in Table S4.
Quantitative Hydroxymethylcytosine Analysis
The assay was performed as previously reported with minor modifications
(Kinney et al., 2011). Genomic DNA was purified with a commercial kit
(QIAGEN). A total of 500 ng of each DNA sample were resuspended in 50 ml
buffer (50 mM potassium acetate, 20 mM Tris-acetate, 10 mMmagnesium ac-
etate, 1 mM DTT [pH 7.9], and 100 mM uridine diphosphoglucose (NEB). DNA
was glucosylated with 4 units T4 phage beta glucosyltransferase (+ T4-BGT)
(NEB) or no enzyme (� T4-BGT). Samples were incubated at 37�C for 12 hr
and heat inactivated at 75�C for 10 min. One 1 unitMspI was added and incu-
bated for an additional 12 hr at 37�C, followed by heat inactivation at 75�C for
10min. DNAs were purified (QIAGEN kit), and the collected DNA was analyzed
by quantitative PCR. To normalize for differences ofMspI digestion efficiency,
the Cp values of the TCF21 promoter MspI site (CpG#7) was normalized to
qPCRs monitoring another MspI site located in CpG island 2. The ratio be-
tween (+T4-BGT) and (�T4-BGT) values was calculated, and the MspI resis-
tance is shown as 5-hydromethylcytosine relative level. A level of one is
considered background. In vitro methylation of TCF21 promoter plasmid
was carried as previously reported (Smith et al., 2006).
Quantitative RT-PCR
Relative expression was determined by qRT-PCR using the Universal Probe
Library System (Roche). cDNA was generated from 1 mg total RNA using oli-
go(dT) priming and Superscript II RT Kit (Invitrogen). Values were normalized
to the housekeeping genes HPRT1 and SDHA. Alternatively, gene-specific
primers were used (Table S3).
In Vitro Synthesis of TARID
DNA templates were generated by PCR using forward primers harboring the
T7 promoter. In vitro transcription assays contained 1 mg template DNA,
40 mM Tris-HCl (pH 7.9), 10 mM NaCl, 6 mM MgCl2, 10 mM DTT, 2 mM sper-
midine, 0.05% Tween-20, 0.5 mM each of ATP, GTP, CTP, and UTP, 0.5 U
RNase inhibitor, and 40 U T7 RNA polymerase (Promega). After incubation
for 16 hr at 37�C, RNA was purified and checked by gel electrophoresis. Biotin
and 4-thiouridine-labeled RNAs were synthesized using 1 mg template DNA in
40 mM Tris-HCl (pH 7.9), 10 mM NaCl, 6 mM MgCl2, 10 mM DTT, 2 mM sper-
midine, 0.05% Tween-20, 0.5 mM each of ATP, GTP, and CTP, 0.3 mM UTP,
0.05 mM biotin 16-UTP (Roche), and 0.05 mM 4-ThioUTP (Jena Bioscience),
0.5 U RNAsin, and 40 U T7 RNA polymerase (Promega). The genomic position
for RNA control intronic region (RNA6) used in RNApull-down experiments and
the capture assays is chr6: 134,211,494–134,212,061.
Bioinformatic and Statistical Analysis
Fickett’s test code algorithm was used to calculate Fickett’s TestCode scores
of TCF21 and TARID (Fickett, 1982). For prediction of secondary structure,
RNAz (http://rna.tbi.univie.ac.at/cgi-bin/RNAz.cgi) was used to calculate z
scores. To predict the transcription starting site in the TARID promoter, an on-
line tool was used (http://www.fruitfly.org/seq_tools/promoter.html). Fisher’s
exact test was used to test for differences in RNA expression or methylation
levels between two groups. The threshold for low and high methylation was
defined by the mean value plus two SD of the healthy tissues. The threshold
for low and high RNA expression was defined by values below and above
the mean of the cancerous tissues plus two SD. To test for the significance
of Spearman correlation coefficients, t distribution was calculated using Prism

Molecular Cell
LncRNA Demethylates the Tumor Suppressor TCF21
5 software (GraphPad). The statistical significance is considered to be signif-
icant if p < 0.05.
ACCESSION NUMBERS
The GenBank accession numbers for TARID T1–T5 reported in this paper are
KF484511, KF484512, KF484513, KF484514, and KF484515.
SUPPLEMENTAL INFORMATION
Supplemental Information includes five figures and four tables and can be
foundwith this article online at http://dx.doi.org/10.1016/j.molcel.2014.06.031.
ACKNOWLEDGMENTS
We thank Oliver Mucke and Jana Petersen for technical assistance. This work
was supported by funding from the Helmholtz Foundation, the German Con-
sortium for Cancer Research, and the National Institutes of Health, DE13123
to C.P. and by an ERC senior investigator grant N�249826-‘‘DNAdemethylase’’
to C.N. I.G.’s work has been supported by the DFG (GR475/22-1, SFB1036),
the excellence cluster CellNetworks, and the ERC (N�232645). Melanoma
cell line C8161 and pEF-FH-TET1 were kindly provided by Dr. Mary Hendrix
(Children’s Hospital of Chicago Research Center) and Dr. Anjana Rao (La Jolla
Institute for Allergy and Immunology), respectively.
Received: January 28, 2014
Revised: May 14, 2014
Accepted: June 24, 2014
Published: July 31, 2014
REFERENCES
Arab, K., Smith, L.T., Gast, A., Weichenhan, D., Huang, J.P., Claus, R.,
Hielscher, T., Espinosa, A.V., Ringel, M.D., Morrison, C.D., et al. (2011).
Epigenetic deregulation of TCF21 inhibits metastasis suppressor KISS1 in
metastatic melanoma. Carcinogenesis 32, 1467–1473.
Barreto, G., Schafer, A., Marhold, J., Stach, D., Swaminathan, S.K., Handa, V.,
Doderlein, G., Maltry, N., Wu, W., Lyko, F., and Niehrs, C. (2007). Gadd45a
promotes epigenetic gene activation by repair-mediated DNA demethylation.
Nature 445, 671–675.
Bergman, Y., and Cedar, H. (2013). DNA methylation dynamics in health and
disease. Nat. Struct. Mol. Biol. 20, 274–281.
Bhutani, N., Brady, J.J., Damian, M., Sacco, A., Corbel, S.Y., and Blau, H.M.
(2010). Reprogramming towards pluripotency requires AID-dependent DNA
demethylation. Nature 463, 1042–1047.
Cortazar, D., Kunz, C., Selfridge, J., Lettieri, T., Saito, Y., MacDougall, E., Wirz,
A., Schuermann, D., Jacobs, A.L., Siegrist, F., et al. (2011). Embryonic lethal
phenotype reveals a function of TDG in maintaining epigenetic stability.
Nature 470, 419–423.
Cortellino, S., Xu, J., Sannai, M., Moore, R., Caretti, E., Cigliano, A., Le Coz, M.,
Devarajan, K., Wessels, A., Soprano, D., et al. (2011). Thymine DNA glycosy-
lase is essential for active DNA demethylation by linked deamination-base
excision repair. Cell 146, 67–79.
Cui, S., Ross, A., Stallings, N., Parker, K.L., Capel, B., and Quaggin, S.E.
(2004). Disrupted gonadogenesis and male-to-female sex reversal in Pod1
knockout mice. Development 131, 4095–4105.
Ehrich, M., Nelson, M.R., Stanssens, P., Zabeau, M., Liloglou, T., Xinarianos,
G., Cantor, C.R., Field, J.K., and van den Boom, D. (2005). Quantitative
high-throughput analysis of DNA methylation patterns by base-specific
cleavage and mass spectrometry. Proc. Natl. Acad. Sci. USA 102, 15785–
15790.
Fickett, J.W. (1982). Recognition of protein coding regions in DNA sequences.
Nucleic Acids Res. 10, 5303–5318.
Mo
Frohman, M.A. (1993). Rapid amplification of complementary DNA ends for
generation of full-length complementary DNAs: thermal RACE. Methods
Enzymol. 218, 340–356.
Ginno, P.A., Lott, P.L., Christensen, H.C., Korf, I., andChedin, F. (2012). R-loop
formation is a distinctive characteristic of unmethylated human CpG island
promoters. Mol. Cell 45, 814–825.
Guo, J.U., Su, Y., Zhong, C., Ming, G.L., and Song, H. (2011). Hydroxylation of
5-methylcytosine by TET1 promotes active DNA demethylation in the adult
brain. Cell 145, 423–434.
Gupta, R.A., Shah, N., Wang, K.C., Kim, J., Horlings, H.M., Wong, D.J., Tsai,
M.C., Hung, T., Argani, P., Rinn, J.L., et al. (2010). Long non-coding RNA
HOTAIR reprograms chromatin state to promote cancer metastasis. Nature
464, 1071–1076.
He, Y.F., Li, B.Z., Li, Z., Liu, P., Wang, Y., Tang, Q., Ding, J., Jia, Y., Chen, Z., Li,
L., et al. (2011). Tet-mediated formation of 5-carboxylcytosine and its excision
by TDG in mammalian DNA. Science 333, 1303–1307.
Ito, S., D’Alessio, A.C., Taranova, O.V., Hong, K., Sowers, L.C., and Zhang, Y.
(2010). Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal
and inner cell mass specification. Nature 466, 1129–1133.
Kangaspeska, S., Stride, B., Metivier, R., Polycarpou-Schwarz, M., Ibberson,
D., Carmouche, R.P., Benes, V., Gannon, F., and Reid, G. (2008). Transient
cyclical methylation of promoter DNA. Nature 452, 112–115.
Kinney, S.M., Chin, H.G., Vaisvila, R., Bitinaite, J., Zheng, Y., Esteve, P.O.,
Feng, S., Stroud, H., Jacobsen, S.E., and Pradhan, S. (2011). Tissue-specific
distribution and dynamic changes of 5-hydroxymethylcytosine in mammalian
genomes. J. Biol. Chem. 286, 24685–24693.
Kriaucionis, S., and Heintz, N. (2009). The nuclear DNA base 5-hydroxymethyl-
cytosine is present in Purkinje neurons and the brain. Science 324, 929–930.
Li, W., Notani, D., Ma, Q., Tanasa, B., Nunez, E., Chen, A.Y., Merkurjev, D.,
Zhang, J., Ohgi, K., Song, X., et al. (2013). Functional roles of enhancer
RNAs for oestrogen-dependent transcriptional activation. Nature 498,
516–520.
Lu, J.R., Bassel-Duby, R., Hawkins, A., Chang, P., Valdez, R., Wu, H., Gan, L.,
Shelton, J.M., Richardson, J.A., and Olson, E.N. (2002). Control of facial mus-
cle development by MyoR and capsulin. Science 298, 2378–2381.
Maiti, A., and Drohat, A.C. (2011). Thymine DNA glycosylase can rapidly excise
5-formylcytosine and 5-carboxylcytosine: potential implications for active de-
methylation of CpG sites. J. Biol. Chem. 286, 35334–35338.
Metivier, R., Gallais, R., Tiffoche, C., Le Peron, C., Jurkowska, R.Z.,
Carmouche, R.P., Ibberson, D., Barath, P., Demay, F., Reid, G., et al. (2008).
Cyclical DNA methylation of a transcriptionally active promoter. Nature 452,
45–50.
Niehrs, C., and Schafer, A. (2012). Active DNA demethylation by Gadd45 and
DNA repair. Trends Cell Biol. 22, 220–227.
Pastor, W.A., Aravind, L., and Rao, A. (2013). TETonic shift: biological roles of
TET proteins in DNA demethylation and transcription. Nat. Rev. Mol. Cell Biol.
14, 341–356.
Quaggin, S.E., Schwartz, L., Cui, S., Igarashi, P., Deimling, J., Post, M., and
Rossant, J. (1999). The basic-helix-loop-helix protein pod1 is critically impor-
tant for kidney and lung organogenesis. Development 126, 5771–5783.
Rai, K., Huggins, I.J., James, S.R., Karpf, A.R., Jones, D.A., and Cairns, B.R.
(2008). DNA demethylation in zebrafish involves the coupling of a deaminase,
a glycosylase, and gadd45. Cell 135, 1201–1212.
Richards, K.L., Zhang, B., Sun, M., Dong, W., Churchill, J., Bachinski, L.L.,
Wilson, C.D., Baggerly, K.A., Yin, G., Hayes, D.N., et al. (2011). Methylation
of the candidate biomarker TCF21 is very frequent across a spectrum of
early-stage nonsmall cell lung cancers. Cancer 117, 606–617.
Rinn, J.L., and Chang, H.Y. (2012). Genome regulation by long noncoding
RNAs. Annu. Rev. Biochem. 81, 145–166.
Schafer, A., Karaulanov, E., Stapf, U., Doderlein, G., andNiehrs, C. (2013). Ing1
functions in DNA demethylation by directing Gadd45a to H3K4me3. Genes
Dev. 27, 261–273.
lecular Cell 55, 604–614, August 21, 2014 ª2014 Elsevier Inc. 613

Molecular Cell
LncRNA Demethylates the Tumor Suppressor TCF21
Schmitz, K.M., Schmitt, N., Hoffmann-Rohrer, U., Schafer, A., Grummt, I., and
Mayer, C. (2009). TAF12 recruits Gadd45a and the nucleotide excision repair
complex to the promoter of rRNA genes leading to active DNA demethylation.
Mol. Cell 33, 344–353.
Schmitz, K.M., Mayer, C., Postepska, A., and Grummt, I. (2010). Interaction of
noncoding RNAwith the rDNA promoter mediates recruitment of DNMT3b and
silencing of rRNA genes. Genes Dev. 24, 2264–2269.
Smith, L.T., Lin, M., Brena, R.M., Lang, J.C., Schuller, D.E., Otterson, G.A.,
Morrison, C.D., Smiraglia, D.J., and Plass, C. (2006). Epigenetic regulation of
the tumor suppressor gene TCF21 on 6q23-q24 in lung and head and neck
cancer. Proc. Natl. Acad. Sci. USA 103, 982–987.
Sytnikova, Y.A., Kubarenko, A.V., Schafer, A., Weber, A.N., and Niehrs, C.
(2011). Gadd45a is an RNAbinding protein and is localized in nuclear speckles.
PLoS ONE 6, e14500.
Tahiliani, M., Koh, K.P., Shen, Y., Pastor, W.A., Bandukwala, H., Brudno, Y.,
Agarwal, S., Iyer, L.M., Liu, D.R., Aravind, L., and Rao, A. (2009). Conversion
of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL
partner TET1. Science 324, 930–935.
614 Molecular Cell 55, 604–614, August 21, 2014 ª2014 Elsevier Inc.
Tessema, M., Willink, R., Do, K., Yu, Y.Y., Yu, W., Machida, E.O., Brock, M.,
Van Neste, L., Stidley, C.A., Baylin, S.B., and Belinsky, S.A. (2008). Promoter
methylation of genes in and around the candidate lung cancer susceptibility
locus 6q23-25. Cancer Res. 68, 1707–1714.
Ulitsky, I., and Bartel, D.P. (2013). lincRNAs: genomics, evolution, and mech-
anisms. Cell 154, 26–46.
Wu, H., and Zhang, Y. (2011). Mechanisms and functions of Tet protein-medi-
ated 5-methylcytosine oxidation. Genes Dev. 25, 2436–2452.
Zhao, J., Sun, B.K., Erwin, J.A., Song, J.J., and Lee, J.T. (2008). Polycomb pro-
teins targeted by a short repeat RNA to the mouse X chromosome. Science
322, 750–756.
Zhu, B., Zheng, Y., Hess, D., Angliker, H., Schwarz, S., Siegmann, M., Thiry,
S., and Jost, J.P. (2000). 5-methylcytosine-DNA glycosylase activity is pre-
sent in a cloned G/T mismatch DNA glycosylase associated with the chicken
embryo DNA demethylation complex. Proc. Natl. Acad. Sci. USA 97, 5135–
5139.





























