Embed Size (px)
Citation preview

Life History Variation of Polymorphic Arctic Charr lvelinus alpinus) in Thingvallavatn,
Directorate for Nature Management, Fkh Research Division, Tungasletta 2, N-7004 Prondheim, Noway
S. SkGlason and S. S. Snorrason institute of Biology, University of Iceland, Grewdsvegur 12, IS- 108 Reykjavik, Iceland
The Roya8 Norwegian Society for Rural Development, P . 0 . Box 7 15, N-20 13 Skjetten, Noway
H. ). Malrnquist and Pa M. J6nasson Freshwater Bislogicab Laboratory, University of Copenhagen, 5 1 Hedsin~orsgade~ DK-3400 Hidberm$ Denmark
Bwstibute of Freshwater Research, 5-170 1 1 Drottingholrn, Sweden
and T. Lindem Institute of Physics, University of Oslo, P.0. Box 7048, Bdindern, N-03 16 Osb 3, Nomay
]onsscsn, B., S. Skljlason, S. S. Snsrrassn, 8. T. Sandlund, H. ). Malrnquist, P. M. Jbnasson, R. Gydemo, and T. Lindern. 1988. Life history variation of polymorphic Arctic charr (SalveBinus alpinus) in Thingvallavatn, Iceland. Can. ). Fish. Aquat. Sci. 45: 1537-1 547.
The Arctic charr (SalveBinus aipinus) in Thingvallavatn, Iceland, exhibits four morpks. Small knthivorous charr grow slowly, have an asymptotic length of 13 cm, and mature at an age of 2 4 yr. Large knthivorous charr grow faster and almost rectilinearly with age. They mature at an age of 3-1 1 yr. Planktivorous and piscivorous charr have even higher initial growth rate, but psanktivorous charr cease growing at 20 cm and piscivorous charr at a fork length of 30 cm. They mature at 3-5 yr and 5-10 yr of age, respectively. The gonadosomatic index and the annual allocation of energy into gonads relative to soma were higher for planktivorous and piscivorous than for knthivorous chars. The results indicate that piscivorous and planktivorous charr belong to the same gene po l ; piscivorous charr are probably recruited from the most fast-growing p!ankivorous individuals and start feeding on fish at a length of 23 cm. The knthivorous rnorphs may belong to different stocks due to significant differences "a several life history variables as well as genetics, spawning time, and feeding habitat. The four morphs seem to be differentiated within the lake through trophic radiation, but should be considered as conspecifics.
Les ombles arctiques QSalveBinus alpinus) du Thingvallavatn dlcelande) presentent quatre types morphologiques. Les petits ombles benthivows ont une croissance lente, atteignent une longueur asymgtotique de 13 cm, et maturent quand i9s ont entre 2 et 4 a. Les gros ombles benthivotes croissent plus rapidement et de fason psesque rectiiigne en fsnction de l3ge. 11s atteignent la maturite h un age se situant entre 3 et 1 1 a. Les ambles planctivores et piscivoses presentent un taux de croissance initial encore plus t?lev$, mais les individus planctivores cessent de crottre quand its ont atteint une longueur la folirche de 20 cm et les individus piscivores A une longueur de 30 cm. tes gamrnes d'ages de maturation sont, respectivement, de 3 A 5 a et de 5 A 10 a. L'indice gonadoso- matique et !e rapport annuel de ('4nergie repartie entre les gonades et Be soma 4taient plus 4Iev4~ chez les ombles planctivores et piscivores que chez les ombles benthivores. hes r4saeltats obtenus indiquent que les ombles pis- civores et planctivores appartiennent au meme r6servoir gbnique, les individus piscivores paovenant probable- ment des individus planctivores A croissawce !a plus rapide et commensant a se nourrir de poissons lorsqu'ibs atteignent 23 cm de longueur. Les types benthivores pourraient appartenir Zi d'autres stocks car divers pararn&tres de leur cycle vital de m$me qlie leurs caracteristiques g&n4tiques, leur p6riode de frai et Beur habitat d'alimentation diff$rent & facon marque%. Au sein du lac, les quatre types semblent stetre diffkaenciks par rayonnement tro- phique mais devraient Gtre consid6r4s cons+cifiques.
Received ApriB 38, I987 Accepted May 19, 1988 (J9256)
lConm~bution h r n the ~ g v d l a v ~ Rojmt.
Can. 3. Fish. &mt. Sci., &Pol. 45, 1988
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.

ife histories of orgznisms appear to be adapted to the envi- roment though natural selection, similar to what is L, own for mo~hologicd characters (Fisher 1930; Schd-
fer 1874; Stems 1976, 1980). For exmple, experiments with msquitsfish (Gambusia aflais) (S t m s 1883, 19841, cut- throat trout (Sa&mo chrki), a d Dolly Vmden c h m QSalvelinus dm) (Jonsson et d. 1984) indicate that age and size at sexual maturity change in response to chmges in the environment. Schdfer and Elson (1975) gave circumstantial evidence that changes in age at sexual maturity of Atlantic salmon ('&Elmo saIar) in North America in recent yeas were due to envhn- mentally induced changes in genetics.
Within lakes, Arctic c h m (Swrlvelinw alpinus) exhibit up to four variants (Beh&e 1980, 1984). Different morphs may be distinguished by a number of traits, such as choice of fwd md habitat, individud growth rate, age md size at sexual maturity, time and place of spawning, body pmprtions , coloration, and parasites (Bidon 1980; Johnson and Bums 1984). kct ic chm9 like many other fish s p i e s , develop rnoqhs through trophic differentiation (or ecological polymorphism) within breeding populations (Skeslet 1973; Sage and Selmder 1975; Capbell 1979; T m e r a d Grosse 1980; H i n h and Jonsson 1982; Komfield et d. 1982; Gmdzien and Turner 1984). Under such conditions, a single population may split into several geneti- cally different l w d stocks (Kirkpat~ck and Selmder 1979; Rymm et d. 1979; Ferguson a d Mason 198 1; Rymm 1981; Hind= et al. 19861, which at least in salmonids may be based on their well-d(46:urnented reproductive homing behavior (Stuart 1953, 1957; Frost 1965; Beh&e 1972). From the perspective of evolutiowuy genetics, the discontinuous trophic variation found in kc t ic c h m may well be the type of niche-specific plymovhism that, for exmple, Maynard Smith (1%) md Turner and Grosse (1980) have regarded as a fundamental pre- requisite for s y m p a ~ c speciation.
kct ic c h m in ningvdlavab (the n m e means lake by the Thingvellir, the original site of the lOOO-yr-oId Icelandic Pa- Iiment) exhibit four phenotypicd and ecological variants (Samundsson 1904; Snomson 1 982; Skfikson 1983; Mdrnqu- ist et d. 1985, 1986). These rnorphs are small md large ben- thivorous charr and pBanktivorous and piscivorous charr (Table I). The benthivorous morphs live in the littoral zone. Smdl knthivorous c h m stay in smdl crevices and under lava
stones in the substrahm during the day, but at least some of &em mcur in the epibenthic zone at night. Large benthivorous c h m are epibenthic. Both %Peed mainly on the snail Lymmea peregra. PId~vorc~~ms md piscivoraus char occur in both pelagic md epiknthic habitats. P l d ~ v o r o u s cham feed m i d y on crustacean zmplmkton md chironomid pupae. Piscivomus c h m prey mainly on threespine sticklebacks (Gasterosfeus acukatus), but may also take small cham (Mdmquist et d. 1985; Sandlund et d. 1987).
Bbs%r&ions on the spawning grounds have shown that phe- notypically similar fish mate; the rnoqhs overlap only partly in spawning time (Table 1: Skiilason 19831, md no natural crossbredieag between moqhs has been obsewrd (K. Gun- nmson and H. Sigwj6nsdottir, Institute of Biology, University of Iceland, GrensBsvegur 12, IS- 108 Reykjav&, Iceland, p r s . corn . ) .
Moqholsgicdl, the benthivorous rngsrphs have overshot mouths, md pBdtivorous md piscivorms rnorphs have ter- mind mouths. This inherited difference is significant soon after the onset of external feeding (Sk%ason 1986). The number of gill rakers is significantly different among all rnorphs except between piscivorous md p l d t i v o r o ~ s cham (Table 1). These morphological differences mong mophs indicate that they are genetically different. However, based on electrophmeeic and- ysis of the protein products of 39 loci, the morphs are very closely related. The values of Nei9s genetic distance range b r n
to 0.001 26 (Magn6sson and Ferguson 1987). Alhough the genetic differentiation between small benthaivorous c h m a d the other three rnorphs is significant, Magnljisson md Ferguson (1987) concludd that they are conspcifics md do not represent different evolutionary lineages. The small genetic differences between the morphs indicate that thek sepaation, if my, is very recent.
Life history variables of the four rnoqhs may be adapted separately, though natural selection, to their respective habi- tats, assuming that they are ~eprductively isolated. If so, we predict that smdl kn$gaivorom cham should be more slow gowing &an large benthivorous chm. Small benthivorous c h m should be able to move freely within the stone matrix of the lava bottom. Large benthivorous chm9 living on the ht- tom, face a higher m m k r sf potentid predators. 'Though high growth rates, predation risk is decreased (Nikolsky 1963).
TABLE 1. ~ o ~ h 0 ~ f B g ~ ~ d a d 62~0b@~d Ceh~86kfish~~ 8% &t3 f8l.E kCtk ~ h m W@S h niwfldla~ab. D8Q f~d~m Mdmqui$t t2et d. (1985), Smdlund et d. (1987), and S. S. Snorrassn et d. (unpbl. data).
Mean number
Morph Phenotypic chaacteiza~on of giII r&ers Spawning period Habitat Main f d -- - -
Small benthivorous Overshot mouth, adult fiih length -
25.2 7 - 38 cm, pm marks along the flanks, dark coloration
Large knhivorous Overshot mouth, adult fish length 25.b 9 - 55 cm, no p m marks, dark eoloratisn
Pkmktivoroaas IPemind mouth, adult fish length 27.4 13 - 26 cm, no p m m&s, silvery coloration
Rscivoms Terminal math, adult fish length 27. B 23 - 65 cm, wo p m IEU~S,
light to silvery ccolomtiasn
July - August Littoral L y m e a peregm
September - October Whole lake Cmsta~ezara zwplimkton, c l i r o n o ~ d pupae
September - 0c%0kr Whole lake espiwe seicMeback (Gasterostem aculeam)
1538 Cm. J. Fish. Aqmf. Sci., Val. 45, 1988
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.

RG. I . mngvdlavatn with 20-m depth contour. Lacations of benthic CB) and pelagic (P) gillnet fishing are indicated.
Pldtivorous and piscivorous c h m should be even more fast growing, as the risk of predation is even higher in the 1 zone than in the structured littoral zone (Crowder and Cooper 1982). The age at maturity of the various morphs should be close to the inflection points of their respective growth curves, so that theis net reproductive rates are r n a h = i i z d (Stems 1976, 1980; Jonsson et d. 1984; Sensen 1985). Furthemore, we pre- dict that the m u d re investment. among sexually mature fish is higher for vorous and piscivorous c h m than for the benthivomus . The latter prediction is based on differentid mortality expectations in the various habitats as indicated above. Therefore, the following null hypothesis was teskd: life history traits such as rate, age at sexual matu- rity, sex ratios, and size-related reproductive investment do not differ mong morphs.
Thingvdlavatn (64'6 'EF, 2 B08'W, 10% rn a. s. 1.) is the largest M e in Iceland (Fig. I), with an area of 84 b2, mean depth of 34.1 m, and mimaam depth of 114 rn (Rist 1975). 'khe Bake is situated in a fault valley in an area of tectonic and geothemd activity. Apart from a large lava flow from the northeast,
yr ago, sporadic tectonic activity and minor local volcanic eruptions have disturbed parts of the lake basin, especially the northern basin. The major disturbances have occurred at intervals of several thousand years (Samundsson
1965). The southern basin has most probably remained unaltered since the glacial retreat about 1 1 500 yr ago (Fig. 1).
Approximately 90% of the inflow to the lake is subterranean through the pornus lava rocks in the catchment area (Rist and 6 ~ s s o n 1986). The nature of the water inflow dong with the Bake rnorphometry and rather windy climate (Einasson 1988) stabilize both the mnud water temperature amplitudes and the chemical composition of the lake (Snonason 1982). The temperature varies between 30 and 11°C during the year, with a maximum in the uppermost 25 ma in July-August. Fpecific conductivity (K,,) is approximately 65 pS *cm - ~(Olafsson 1980), and Secchi disc transparency is 6-15 rn in summer. The
d in 1959, and since then the water Bevel has fluctuated less than 50 crn during the yea.
ThingvdaBIavatn has a Iarge pelagic zone and a benthic zone consisting of t h e main strata: (1) a littoral zone consisting of Bava md hard bottom often with loose boulders md rocks at & 10 m depth, (2) a littoral soft bottom covered by dense stands of Nitelk opaca at 10-20 rn depth, and (3) a profmdd soft bottom of diatomic gyttja (Liradegaad 1980; Lastein 1983) at depths below 24% m. The zoobenthos is dominated by chirowomid larvae, tubificids, and molluscs. Mmimaam densities of chironomids and molluscs are found in the stony littoral zone (0-1 4% m) (Eindegmd 1980; Snomason 1982). The zoopBmkton are predominantly Daphnia iongispins, Cyclops abyssorurn medimus, md Leptodtaptomus minutus (Antonsson 1977). The lake supports a dense population of kc t ic cham as well as
cklebacks and a few brown trout ('galmo trutta). s annually fish between 7 and 70 metric tons of
p l~ t ivo rous c h m on the spawning grounds, which amounts to approximately 330% of the biomass of adult planktivorous c h m (T. Lindern et al., unpubl. data). Large benthivorous md piscivoroaas c h m are also fished to some degree (Kristjhsson and Adalsteinsssn 1984). Among small knthivomaas chm, only the very ddest and largest are fished as a by-product of the planktivorous cha r fisheries. The ecosystem of Thingvdlavatn thus appears to be very stable for the subarctic region (Spaomson 1982; 96nason and Ehdegaard 1988).
Data Collection a d Analysis
kc t ic c h m were collected (N = 6332) from Thingvallavatn during thee periods: August 1983, May-June 1984, and August-December 1984. Fish were caught with benthic and pelagic nets. The benthic nets were I .$ m and the pelagic nets 6 m in vertical dimension. The nets were gangs of panels with 12 different bar mesh sizes: 10, 12.5, 15, 18.5,22,25,29,33, 38, 43, 50, and 60 m. Benthic nets were used in thee loca- tions (Fig. l ) during at least two 24-h periods in each sampling period on the following substrata and depth zones: (1) stony bottom, 0-10 m; (2) Nitelk bottom, 10-20 m; and (3) soft bottom, 20-35 m. Pelagic nets were set in two locations (Fig. 1) at 0-4, 6-12, 263-26, 35-41, 50-56, and 66-72 m for at least two 24-41 periods at each depth zone and sampling series. In the Nitella zone at Midfell, pelagic nets were set for one 24-h period in each sampling series in depth zones 0-4 and on the bottom at 7-13 rn (Fig. 1). Pelagic nets were lowered along ropes anchored on the bottom and kept at the correct depths by a combined use of weights a d floats. More details about sam- pling are given in Smdlund et al. (1987). The nets used caught fish from 7 to 4 em in length,
Arctic char were classified into morphs on the basis of body length, upper and lower jaw lengths, head shape, and color
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.
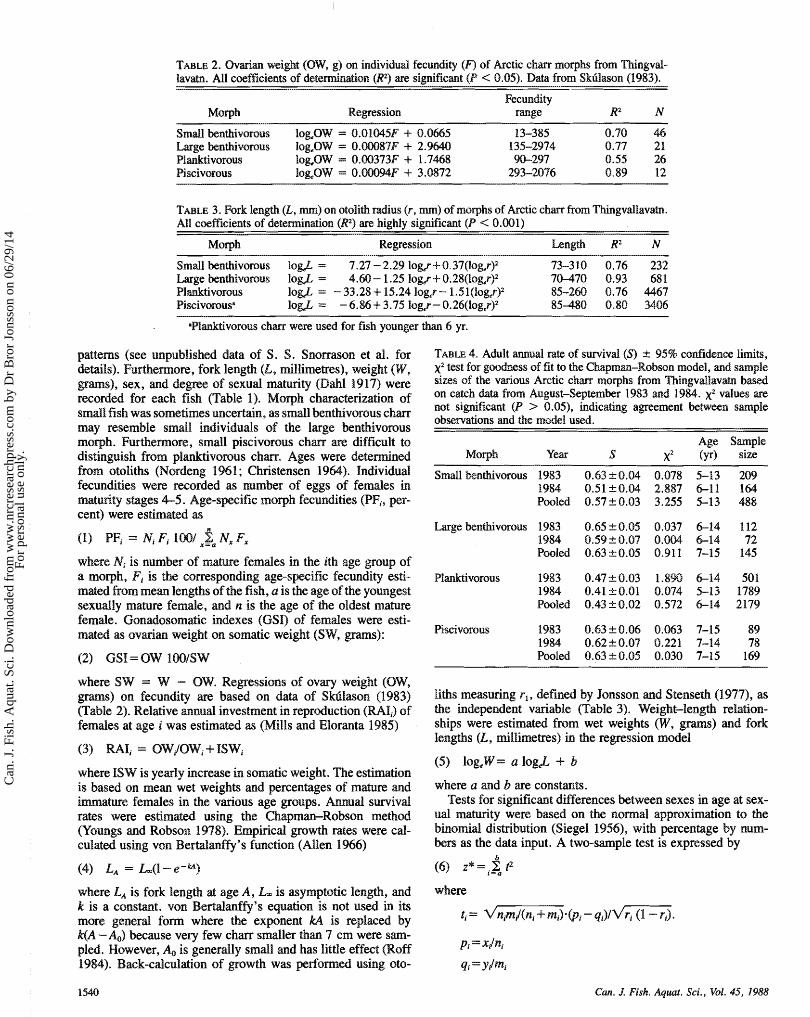
TABLE 2. Ovarim weight (OW9 g) on individual fecundity (F) of Arctic c h m morghs from Thiwgvd- llavatn. All cmfficknts of deteminat.iow (R2) are significant (P < 0.05). Dah from Skfilason (1983).
Regression
small benthivorous l%OW = 0.01045F + 0.0665 13-385 0.70 46 Large knt%nivorous IogeOW = O.W87F + 2.9640 1 35-2974 0.77 21 Plmktivorous log,OW = 0.00373%" + 1.7468 90-297 0.55 26 Rscivorous log,OW = 0.W94F + 3.0872 293-2076 0.89 12
TABLE 3, Fork length (L, nun) on otolith radius (r , m) of morphs of Arctic c h from Thingvdlavatn. AH coefficients of dekmination (R2) are highly significant (P < 0.001)
Moqh Regression Length R2 M
- -
aPimktivorous c h m were used for fish younger than 6 yr.
patterns (see unpublished data of S. S. Snonason et d. for details). Furtherno=, fork length (L, niIBimetres), weight (W,
s), sex, a d degree of sexual. maturity (Dh l 31917) were recorded for each fish (Table 1). Moqh chmacterization of smdl fish was sometimes uncertain, as small benthivorous char may resemble s d l individuals of the large benthivorous moqh. Furthemore, smdl piscivorous char are difficult to distinguish from pHa&tivomus chm. Ages were detedned from otolith% (Nordeng 196 1 ; Ch-istensen 1964). Individual fecundities were recorded as number of eggs of females in maturity stages 4-5. Age-specific morph fecundities (PK, per- cent) were estimated as
where %$", is number of mawe females in the ith age group of a msph, F, is the corresponding age-specific fecundity esti- mated from mean lengths of the fish, a is the age ofthe youngest sexually mature female, and n is the age of the oldest mature female. Gonadosomatic indexes (GSI) of femdes were esti- mated as ovarian weight on somatic weight (SWB grams):
where SIN = W - OW. Regressions of ovary weight (OW, gyms) on fecundity are based on data of Skiilason (1983) (Table 2). Relative m u d investment in reproduction (MIi) of females at age i was estimated as (Mills and Elormta 1985)
where ISW is yearly increase in somatic weight. The estimation is based on mean wet weights and percentages of mature a d
atwe femdes in the various age groups. Anmd s w i v d rates were estimated using the Chapm-Robson method (Youngs and Robson 1978). Empirical growth rates were cal- culated using vow Bertdmffy's function (Allen 19%)
T ~ L E 4. Adult m u d rate of survival (S) 2 95% confidence limits, >g2 test fm goodness of fit to the Chapman-Robson model, and sample sizes of the various Arctic char morphs f m Thingvdlavatn based ow catch data from August-September 1983 and 2984. x2 values are not significant (P > 0.051, indicating agreement between sample observations md the model used.
Age Sample Morph Year S xZ (yr) size
Small benthivorous 1983 1984 Pooled
Luge benfhivorous 1983 1984 Pooled
P ~ ~ i v o ~ o u s 1983 1984 Pooled
Rscivomus 1983 1984 Pooled
hths measuring r,, defined by Jonssow md Stenseth (1977), as the inkpendent variable (Table 3). Weight-length relation- ships were estimated from wet weights (W, grams) md fork lengths (L, millimetres) in the regression model
where LA is fork length at age A, Em is asymptotic length, a d where
where a and b are constmts. Tests for significant differences between sexes in age at sex-
u d maturity were based on the noma1 approximation to the binomial distribution (Siegel 1956), with percentage by num- bas as the data input. A two-sample test is expressed by
k is a constant. voi~ertd&ffy 's equation i s not used in its more general form where the exponent kA is replaced by ti = Unimnnj + m~.@, - 4i)/m. k(A -A,) because very few char smaller than 7 em were sun- pled. However, A, is generally smdl a d has little effect (Rcsff pi = n]nj
1984). Back-calculation of growth was performed using oto- 4i = ~ d m i
Can. 9. Fish. Aquab. Sci., V01. 45, 1988
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.

ri = (xi 9 yi)l(ni i- mJ almost parallel between 1 md 3 yr sf age, but plmktivorous c h m ceased growing in length at a considerably earlier age than
where xi and yi are the numbers of males and piscivorous chm. The back-calculated mean lengths of small at age i? ni mi the numbers of a" females at bn&ivorous and large benthvomus c h m were age i , ti is disbibuted with (ni - ') degrees of different at all ages (&tests, P < 0.00 1). There was no and '* is x2 with (b - a) degrees of significant difference in growth rate between sexes or sexually
Survival, Growth, m d Size
Annual survivd rate of adults was lowest for pldtivorous c h m (8.41-0.47) (Table 4). For smdl benthivoroams, Iarge knthivorous, and piscivomams ehm, it was 0.5 14.69. Adult age groups which yielded a significant (P < 0.05) fit to the Chapman-Robson model were used for the analysis. The data indicated that the survival rate was higher in 1983 than in 1984 mong small knthivomus chm, large bewthivsrous chm, md pldtivorous cha r (ttests, P < 0.05).
The mean size of mature individuals increased from small knthivsr6pus cham through plawktivorous and large knthivomus c h m to piscivorous c h m (Table 5). The range in length of adult large benthivorous c h m was Iarger than that of the &her adult moqhs.
The small benthivorous mopgh was the slowest growing, md gmwth rates were successively higher in Ixge benthivorous, plmktivorous, and piscivorous chm. The growth rates levelled off at approximately 13, 20, md 30 cm in small knakivorous, pldtivorous , a d piscivorous ckm, respectively. Piscivorous cham, feeding on planktivorous chwr (cannibals), were sometimes caught in the lake, md such individuals may become larger than 60 crn in length (Mdmquist et dal. 1989). In the present samples, however, here were H ~ C B piscivorous c h m larger than 48 cm (Table 5). Large benthivorous c h a r continued to increase in length (L,,, millimetres) with increasing age (A, years) at an dmost rectilinear rate:
Asymptotic length (Lm) md Brody's growth coefficient (k) of VOW Bertdmffy's growth model (Equation (4)) differed significantly between morphs (Table 5). Fish that were classifid as intermediate between small knthivorous and large knthivorous cham had L, 6- SD = 200 + 47 Those &at on the basis sf external morphology were classified as intemediate between g l ~ t i v o r o u s md piscivorous char had La = 229 zk 9 nun (N = 15).
Back-caHculation of growth using otoliths (Fig. 2) cornesponded well w mpiricd growth rates (Table 5). The growth curves of pl ivorous md piscivorous c h m were
immature md mature fish within moqhs.
Weight-Length Relationships
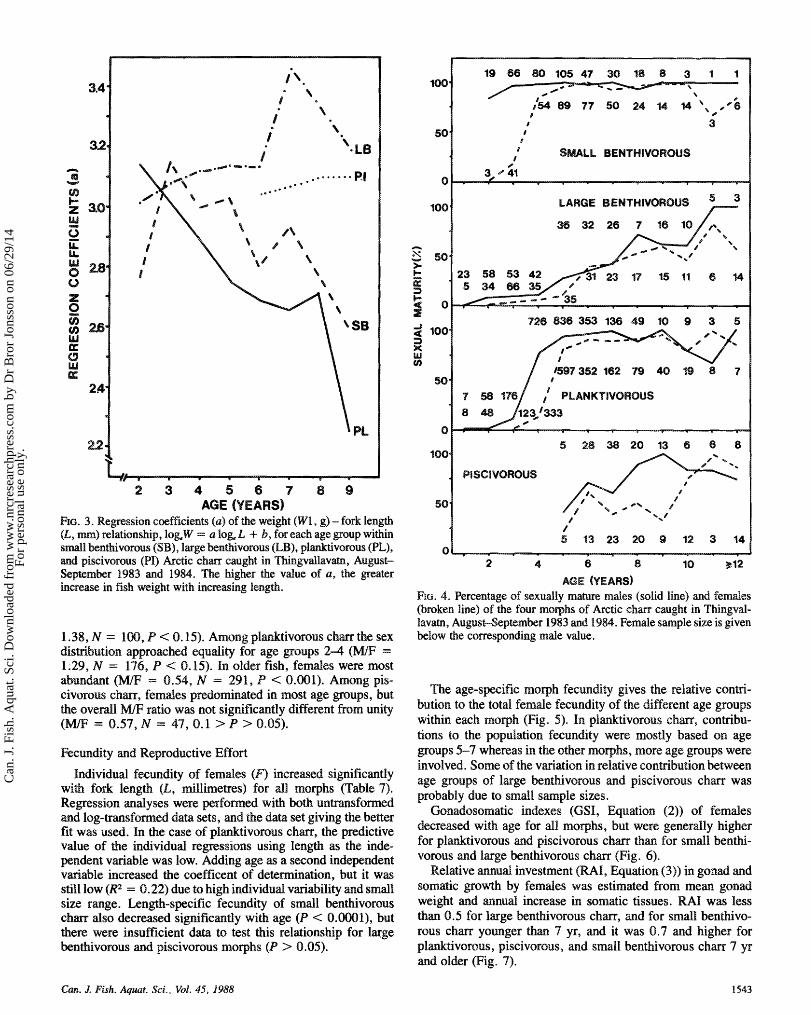
Weight-leng~ relationships show the increase in weight rel- ative to length; the higher the regression coefficient (a, Equa- tion (511, the quicker weight will increase with length. This relationship differed significantly among the morphs in both spring a d autumn (test of homogeneity of regression coeffi- cients, June: F ,,,,, = 64.68, P < 0.0001; August: F,,,,, -- 8.685, P < 0.05). Significantly different weight-leng th ~ l a - tionships were found between the sexes within small benthi- vorous a d planktivorous c h m md spring and autumn samples within pldt ivomus a d piscivomus c h m (Table 6). Among smdl benthivorous mQ pldtivorous ehm, intrmorph regres- sion coefficients decreased with age within age groups 2-9 yr, whereas in large benthivorous md piscivomus chm, it did m t (Fig. 3). A significant heterogeneity in regression coefficients mong age groups was found only for pHanktivorous c h m (F3,2721 = 7.49, P < 6.001). This means that weight increase with increasing length is smaller among older age groups of small benthivorous md p%mktivorous cham than in large ben- thivorous and piscivorous chm.
Age md Sex
Age at sexual maturity differed between morphs (Fig. 4). Age at maturity of each of the four moqhs did not differ between 1983 and 1984 (P > 0.05), and they were therefore pooled. Smdl benthivorous (2-17 yr of age) and pldt ivomus c h m (1-14 yr of age) boh matured at am early age and over a few age p u p s . Among small knthivorous chm, more than 50% of the males in age group 2 were matwe, and 50% of the females in age group 4 were matwe. The difference between sexes was significant (z* = 105.34,4 df, B < 0.000 I). Among p l ~ t i v o r o u s chm, m r e than 56% of the males in age group 4 and females in age group 5 were mature- This difference between sexes was d s s highly significant (z* = 588.26, 5 df, P < 0.0001). Large benthivorous (2-18 yr of age) and pisci- vorous c h m (5-18 yr sf age) matwed later a d over a wider range of years. Among large bentbivorous chm, 50% maturity of both sexes was attained at age 8 md mong piscivorous cham at age 6. Age at maturity differed between sexes for piscivorous cham (z* = 24.69, 5 df, P < 0.001), but not for luge benthi-
TABLE 5 . Empirical sizes (m) md the parameters (f 95% confidence limits) L (nun) md & of the exponeratid growth model LA = L(1- C-&) where A is age (yr) of' Arctic char from Thiaagvdlavatn, based on mBed data from Aueaast-Se~kmkr in 1983 md 1984.
Adult fork Ben&
Mdes Females
M w ~ M e Range Mean Range La k N
Can. J. Fish. P$CBwf. Sci., VoIOE. 45, 1988
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.
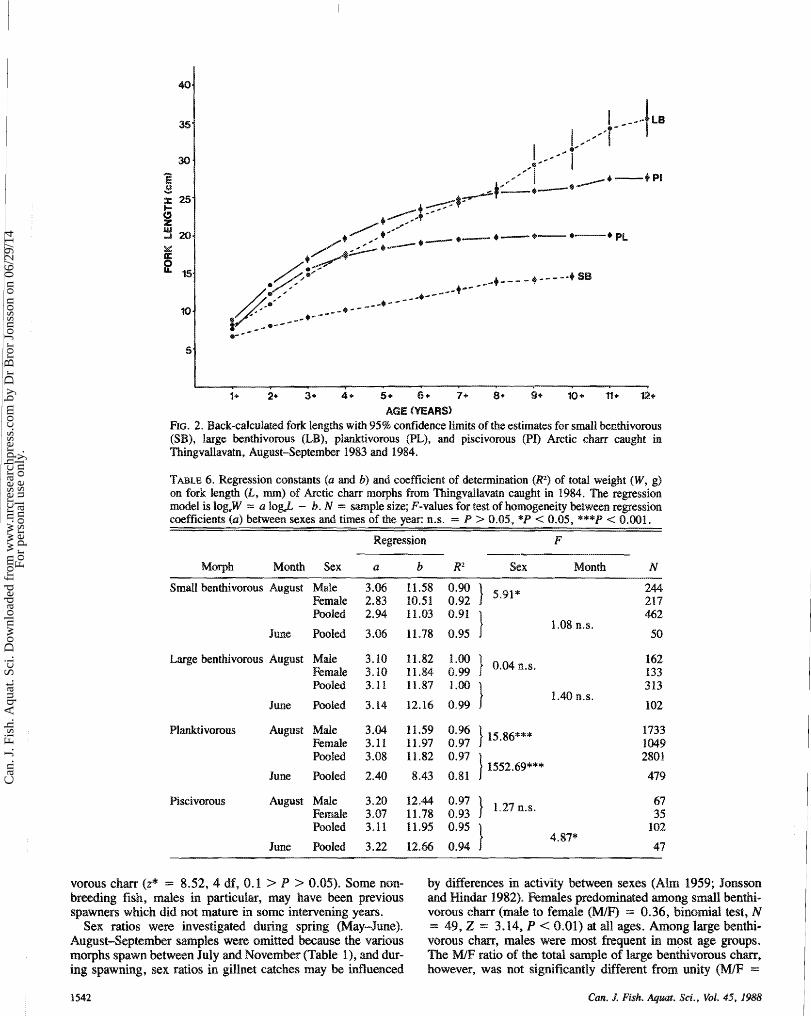
AGE (YEARS) FIG. 2. Back-cdeulated fork lengths with 95% confidence limits of the estimates for small benthivorous (SB), large kntkiivosoaas (EB), pHanktivorous (PE), md pi~~ivorous (PI) Arctic chm caught in Thing%rdlavatn9 AugustSeptember 1 983 and E 984.
TABLE 6. Regression constants (a and b) and coefficient of determination (R2) of total weight (W9 g) on fork length (L, m) of Arctic cham mophs from Thingvd%avatn caught in 1984. The regression model is hg,W = a IogJ - b. N = sample size; F-values for test of homogeneity between regression coefficients (a) between sexes and times sf the year: n.s. = P > 0.05, *P < 0.05, ***P < 0.001.
Regression F
M w ~ Month Sea at & RP Sex Month N Small benthivorous August Male
Female Pooled
June Pooled
Emge benthivorous August Male Female Pooled
June holed
Pldtivomns August Mde Female Poo1d
June Pooled
Piscivorous August Male Female Pooled
vorous clam (z* = 8.52, 4 df, 0.1 > B > 0.05). Some non- breeding fish, males in particular, may have been previous spawners which did not mature in some intervening years.
Sex ratios were investigated during spring (May-June). August-September samples were omitted k a m e the various morphs spawn between July md November (Table I), and dug- ing spawning, sex ratios in gillnet catches may be influenced
by differences in activity between sexes (Alm 1959; Jonsson and Hindax 1982). Females p r e d o ~ n a e d among smdl benthi- vorous cham (male to female (WF) = 0.36, binomial test, N = 49, Z = 3.14, P < 0.01) at dI ages. Among large benthi- vorous ehm, males were most frequent in most age goups. The WF ratio of the total sample of large knthivsmus cham9 however, was not significantly different from unity (MI? =
Can. J. Fish. Aqw. Sci., VoE, 45, 1988
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.
2 3 4 5 6 7 8 9 AGE (YEARS)
FIG. 3. Regression coefficients (a) of the wei@t (W1, g) - fork length (L, m) re%ationship, bg,W = a l og k + b, far each age goup within s d l bentPivornus (SB), large knthivorous (LB), pldaivmsus (PL), and piscivssroaas (PI) Arctic cham caught in Thingvdlavatn, August- Skptewnkr 1983 and 1984. The higher the vdue of a, the greater increase in fish weight with increasing length.
1.38, N = 188, P < 8.15). Among plmktivorous e h m the sex distxibution approached equality for age groups 2 4 (WF = 1.29, N = 176, P < 8.15). In older fish, femdes were most abundant = 0.54, M = 291, P < 0.001). Among pis- civorasus femdes predominated in most age goups, but
erdl WF ratio was not significantly different from unity = 0.57, N = 47, 0.1 > P > 0.05).
Fecundity and Reproductive E f h t
Individual fecundity of females (F) increased significantly with fork length (L, dllimetres) for all rnorphs (Table 7). Regression mdyses were performed with both untransfomed md log-trmsfmed data sets, and the data set giving the better fit was used. In the case of plmktiv~rous chm, the predictive vdue of the indiavidud regressions using length as the inde- pendent variable was low. Adding age sas a second independent variable increased the cwfficent of d e t e ~ n a ~ o n , but it was still Bow (IS" 0.22) due to high individual variability and small size range. Length-specific fecundity of smdl benthivorous c h m dso decreased significantly with age (P < O.OOOll), but there were insufficient data to test this relationship for large knthivorous md piscivorous moqhs (P ) 0.05).
19 86 80 105 47 30 18 8 3 1 1
8 I 3
50 I d
P P SMALL BENTHIVOROUS
LARGE BENTHIVOWOUS /--
2 4 6 8 10 312
AGE (YEARS) FIG. 4. Percentage of sexually rnabre mdes (solid line) md females (broken line) of the four morphs s f Arctic c h m caught in Thinpal- Iavatn, August-September 5 983 and 1984. Female sample size is given k l s w the corresponding mde vdue.
The age-specific moqh fecundity gives the relative contri- bution to the total female fecundity of the different age groups within each morph (Fig. 5). In plamktivorous chm, contribu- tions to the population fecundity were mostly based on age groups 5-7 whereas in the other moqhs, more age groups were involved. Some of the variation in relative contribution between age groups of large knthivsrous and piscivorsu~ char was probably due to small sample sizes.
Gonadcpsomatic indexes (GSI, Equation (2)) of females decreased with age for dl morphs, but were generally higher
ivorous and piscivorous c h m than for small benthi- vomtss and large benthivorous e h m (Fig. 6).
Relative mnud investment (RAh, Equation (3)) in gonad and somatic growth by females was estimated from mean gonad weight md mnud increase in somatic tissues. RAH was kss than 0.5 for large benthivorous chm, and for small benthivo-
k m younger than 7 yr, and it was 0.7 a d higher for vorous, piscivorasus, and small benthivorous c h m 7 yr
md older (Fig. 7).
Can. .I. Fish. Aquot. Sci., Vol. 45, 1988
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.
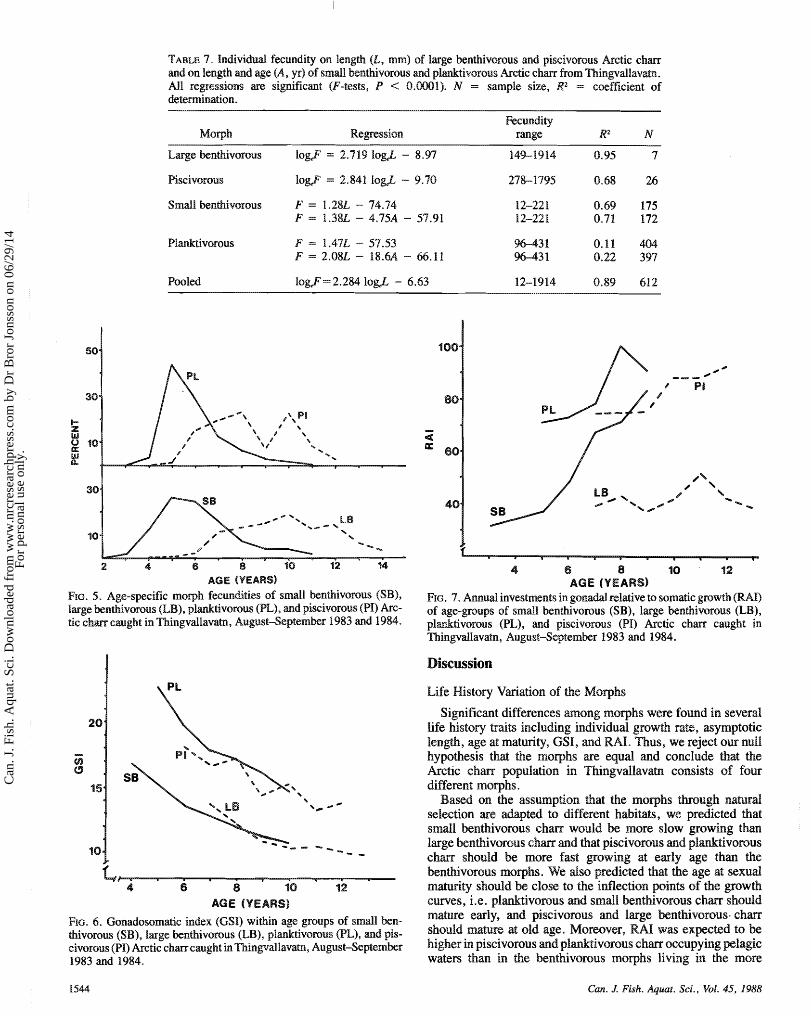
TABLE 7. Individual fecundity on length (L, m) of large benthivorous and piseivorous Arctic c h m md on length and age (A, yr) sf smd1 benthivorou~ and pl&tiv~~rous kc t ie e h m from Thingvdlavatn. All regressions m significant (F-tests, P < 0.0001). N = sample size, R2 = coefficient of
Fecundity range Ra N
Large benthivorous logF = 2.7 19 logJ - 8.97 149-1914 8.95 7
Pooled logF=2.2M log& - 6.63 12-1914 0.89 61%
AGE (YEARS)
86. 5. Age-specific moph fecundities of small bewthivorous (SB), large knthi~orous (LB), p%&tiv~mans (PL), and giscivorous (PI) k c - tic e h m caught in Thhsrgvdlavatn, August-Septemkr 1983 md 1984.
4 6 8 10 12 AGE (YEARS)
Rc. 7. Annual investments in gasnadd relative to somatic growth (RAI) of age-groups of small benthivorous (SB), large benthivorous (EB), p l d ~ v o r o u s (PL), and piscivorous (PI) kc t i c c h m caught in Tl~iingvdlavatn~ August-Septemkr 1983 md 1984.
Discussion
i, . 4 6 8 10 12
AGE (YEARS)
Re. 6 . Gonadosomzsaic index (GSI) within age groups of small k n - thivoroaas (SB), large benthivomus (LB), plmktivomm (PL), and pis- eivorous (PI) kc t i e e h m caught %tin Thingvdlavatn, August-September 1983 md 1984.
Life History Variation of the Moqhs
Significant differences among moqhs were found in several Eife history traits including individual growth rate, asymptotic length, age at maturity7 GSE, a d RAI. Thus, we reject our null hypothesis that the rnorphs are equd and conclude that the k c t i c cha r pspuIation in Thitngvallavab consists s f four different msqhs .
Based on the assumption that the moqhs though natural selectism are adapted to different habitats, we predicted haat small benthivorous c h m would be more slow growing than large ben~ivorous ehm md that ppiscivorous md plmktivomus c h m should be more fast growing at early age than the benthivorous moqhs. We dso predicted that the age at sexual maturity ssfaoukl be close to the inflection points of the growth curves, i . e . pl&tivarous md smdl benthivorous c h m should mature early, md piscivorous m d large benthivorous. c h m should mature at old age. Moreover, was expected to be higher in piseivoro~ss m d pB&tivorous ehm occupying pelagic waters than in the benthivorous morphs living in the more
H 544 Can. J. Fish. Aqua. Sci., Val. 4.5, I988
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.

sheltered littoral zone. This prediction was based on differences in expected mortalities of fish living in structured and unstructured habitats (Chmov et d . 1977; Crswder and Cooper 1982). Our results are in accordmce with these expectations. However, we were not able to measure juvenile mortality. Adult mortality rate was higher for the p l~ t ivo rous morph than for the other morphs. One of the reasons for this may be the high fishing rate in the lake.
Differences in age at maturity cause some of the variation in adult sizes mong the morphs, as maturity iw1:luences the Individual growth rate of the fish. The growth rate decreases when maturity is attained because the energy expenditure to gonadal development a d reproductive activity reduces somatic growth (Cdow 1979; Reznick and Endler 1982; Roff 1983). Thus, early-maturing fish become smd1 md late-maturing ones may grow large. Therefore, differences in age at sexual maturity dso partly explain the observed differences in adult body size between planktivoroms , piscivorous , and small and large benthivoroaas chm.
Differences in adult sizes of pIan.ktivorons and ~ ~ S C I V O ~ O U S c h m are probably also due to differences in egg size and diet. Piseivomus cham have lager eggs than p l~ t ivo rous e h m (Sk6lasow 19831, and their offspring have consequently a growth and size advantage during the first weeks (Kaakov 198 1 ; Kmler a d Kato 1983; Skfilason 1986). Growth rates of these two rnsrphs are pad%el during the 3 first yr of life, and piscivorous c h m develop probably from the fastest growing individuals that when young are classified as p l~ t ivo rous cham. Piscivorous c h m switch usually from plankton to fish feeding at a fork length of approximately 23 cm (Malmquist B988), and p l ~ t i v o r o u s fish that reach this threshold length m y become piscivorous. Most planktivorous cham, however, never p w that Barge and remain plankton feeders.
Size differences between smdl and lage knthivorous c h m may also partly originate from differences in egg size, as the smdle~ rnorph has smaller eggs than the large one. Diet differences, on the other hand, should not account for differences in adult size, as both rnorphs largely fwd on Lymmea peregra. However, they do occupy different feeding habitats (Smdlund et al. 1987). Smdl benthivorous c h m live sheltered within the stone matrix of the lava bottom whereas the large knthivomus c h m live more exposed upon the substratum. A small body size m y be advantageous for fish living within the substratum because the interstitial spaces are limited and smdl individuals are able to maneuver more freely among the stones than large fish. We do not know any c h m predator feeding on c h m within the bottom substratum. For cham jiving upon the bottom substratum, a large body size may be advantageous because foraging efficiency is probably positively comBated with size, and the risk of predation decreases with size (Nikolsky 1963). Medium-sized fish may do relatively poorly in both habitats. Under such conditions, fish experiencing different growth rates and body sizes may select different habitats, md natural selection may favor early age at maturity among slow-growers and old age among fast- growers. In this way, adaptation to life in two different habitats may influence the growth and size differences between the two knthivorous morphs in Thingvallavata. Thus, diet, habitat choice, and egg size may be i m p t for mmph differentiation of the Arctic c h m in Thingvallavatn.
Individual fecundity varied among moqhs; it is lowest in small benthivorous cham a d highest mong large benthivorous cham and piscivorous chm. The main rewon is that fecundity
increases with fish size. The benthivorous fish invested less in gonadal than somatic growth compared with the pladctivoroars md piscivorous moqhs. Furthermore, the length-specific fecundity decreased with age in pldtivorous and s d l benthivorsus chm, even though the relathe mount of energy allocated to gonadal development increased with age. Reduction in length-specific fecundity with increasing age has also been found in other popuiations of Arctic c h m (Jonswn a d Mind= 1982) and in other salmonids as brown trout (lornsson 1985).
The relative amount of energy allocated to gonadal production increased with age in all moqhs. Increased investment in reproductive relative to somatic growth with age may be a general trend among adult iteroparous fish s p i e s until they reach senility (N&olsky 1963). One reason may be that the chance of surviving mother yea decreases with age. On the other hand, young fish may show restraint in gonadal production, thus, pstspawning mortality decreases md future growth and fecundity increase.
Sex ratios sf piscivorous, adult pla&tivorous, md smdl benthivomus cham showed a female excess whereas males were more abundant in most age group of large benthivorous chm. Female excess in the thee former mophs may result from earlier age at maturity of males than of females. Natural mortality of fish that me sexually mature is higher than that of immature individuals at the same age (Sviirdson 1951; Cdow 1979; Jonsson et d. 1984). Among large benthivorous c h m the sexes attained maturity at the same age.
The variation in the adult sizes and ages at maturity of the c h m wrorphs in Thiragvallavatn embraces a large part of the totd variation observed in lake resident stocks sf kc t ic c h m throughout the HoHactic distribution area of the species (e.g. Schmidt-Nielsen 1939; Nilsson 1955; Frost 1 965; Nilsson and Filipsson 197 1 ; AMsteinsson 1979; Jonsssn a d gstli 1979; Johnson 1980; Jonsson and Hindar 1982; Nordeng 1983; Gydemo 1984; Klemetsen et d. 1985; Hindar et al. 1986). Smdl benthivorous cham, on one hand, are of the same size and age as the smallest md youngest adult c h m observed elsewhere (Johnson 1980). The large size and high age at maturity of piscivorous and benthivorons c h m axe not uncommon for subarctic populations (Klernetsen and Grotnes 1980; Nilsen and memetsen 1984), and even higher ages at maturity have been reported from ppulatims in northern Canada (Johnson 1980) and Bear Island (Klemetsen et d. 1985). No single lake, however, is hown to support such a diverse variety of Arctic c h m as found in Thingvallavatn.
Evolution of the Moqhs
The c h m morphs in ThingvdBavatn m y have been devel- oped through trophic differentiation within the lake. The idea of trophic radiation in kc t ic cham populations within Hikes has been advocated by Hindw and Ponsson (1982) and Hindm et id. (1986). In a s m e y of kc t ic c h m in Nomay, Hindar et al. (1986) documented greater genetic distances between stocks of the same rnorph in different lakes than between rnorphs in the s m e lake, supporting the hypothesis that the morphs have evolved with the Mes. Arctic e h m is commonly observed to differentiate into two or three sympatric trophic variants (Jshn- son 1980). Pour sympatric morphs, as in Thingvallavatn, are rare (Be%mke 1984). We consider the possibilities of trophic radiation mong Arctic c h m in this lake as particularly good for the subarctic region for two reasons: (1) eke I&e offers sev- eral different food niches which are relatively stable both within
Can. J- Fish. A ~ U C ~ P . Sci., VoE. 45, 1988
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.

and between yeas (see Study h a ) and (2) there are only two other freshwater resident fish species in Iceland, brown trout and threespine stickleback. Brown trout is rare in Thingvdla- vatn due to very restricted spawning grounds. Threespine stic- kleback is numerous, but mainly restricted to the mamophyte vegetation (Nitelk opeeca) in the 1&2O m depth zone (0. T. SmdIund et d. unpubl. dda).
The small genetic differences between the c h m morphs in minpdlavam (Magnbsson a d Fergumn 1987) make the mul- tiple invasion theory (Sv&dson 1961; Nymm et al. 1981) improbable. This hypothesis claims that ssympatric c h m rnoqhs we sibling species which evolved dlopa~cally during the Pleistocene glaciations and in postglacial time have immigrated into some lakes where they now reside as different species.
We do not b o w to what extent mmrd cross-b~eding occws among the morphs. However, the morphs overlap only parfly in spawning times and areas, md obsewdions of spawning c h m indicate that phenotypicdly similar fish tend to mate with each other (Jonsson a d Hindu 1982; K. Gunnason a d N. Sigmj6nsd6ttirY p r s . corn . ) . Thus the genetic exchange between the rnoqhs may be limited. The genetic difference between the morphs, however, is very small; the vdue of Nei9s genetic distance is only I / 10 of that of Arctic cham populations in eastern and western Noway9 which has been geographically isolated at Beast since the postglacial immigration BO 000 yr ago (Hinadar et d. 1986). Thus, if the mophs in Thingvallavatn are reproductively isolated, this separation probably occurred recent1 y.
The present investigation shows that one lake may support several mophs of Arctic chm. The morphic differentiation seems to have occurred though trophic radiation within the lake. Reproductive isolation between the trophic variants may be maintained though behavioral mchmisms. We regad the c h m morphs as conspecifi~s, equivalent to local, sympatric stocks in other salmonids such as brown trout (Rymm et al, 1979; Rymm and SthI 198 1 ; Ferguson md Mason 198 1).
This work would mot have been possible without the help of the following people: G s t i n n P. Magnusson, Thorleifur Eiriksson, J6hmn J6wsmm and other land owners at Thiaingvallavatn, Nina Jonsson, 9 m Ruud-Hmsen, Robert 9. Behdce, Geoffrey Fryer, Kjeeil Hindar? To&- j8m J b i , Lionel B~hnson, John E. Thorps, and Amy Lightfoot. Fur- h e m o r e , we are indebted to the Nytional Power Plant Company (Lmdsvirkjun), Thingvdla Commissioh of the I c e k n d c Parliament, Banish Natural Science Research Council, Icelandic Natural Scknce Research Council, University of Iceland, Icelandic Foundation for h d Development (ByggdasjMur), Nordic Council for Ecology, Nordic Cultural Foundation, md NATO Science Foundation, d l s f which we gratefully achowledge.
~ A L S ~ M S S B N , H. 1979. Size md f d sf &tic @him Salvefinus aipireus and stickleback (Gasterssteus acrakeaaus in Lake Myvatn. Bikos 32: 228-4231.
&LEN, #. R. 1966. A method of fitting growth curves of the von Bertalanffy type to observed data. B. Fish. Res. Board Can, 23: 163-179.
A m , G. 1959. Connection between maturity, size md age in fishes. Rep. Inst. Freshwater Res. Brottningholm 40: 5-145.
ANTONSSOF~, 0. 1977. Zwplmktsn of Lake ningvdlavatn 1972-75. Pat- graduate thesis, University of Iceland, Reykjav&, Iceland. (In Icelandic)
BWON, B. K. Em.] 1980. Chams, sdmonid Fashe of the genus Salvehi~s. Dr. W. Junk Publisher, The Hague, The Nethalands.
B E P H ~ , R. J. 9972. The systematics of salmnid fishes of recently glaciated Iakes. 5. Fish. Res. Board Cm. 29: 639471.
1986. A systemtic review of the genus Salvelims, p. 4 1 4 N . Bn E. Ka Bdon [ed.] Chms, sdmowid fishes of the gems Salvedinus. Dr. W. Junk Publisher, The Hague, The Netherlands.
1984. Ogmizing the diversity of the Arctic c h m complex, p. 3-21. Hn L. Johnson and B. B m s [ed.] Biobgy of the kc t i c chm. km. Int. Symp. Arctic Chm. University of Manitoba Press, Winnipeg, Man.
CALOW, P. 1979. The cost of ~prduckion - a physiologicd appro~ch. Bid. Rev. 54: 2 3 4 .
CAMPBELL, R. N. 1979. Ferox trout, SaHrno trufta L., and chm, SalveHinus alpinus (L.), in Scottish lochs. J. Fish Biol. 14: 1-29.
CWOY, E. L., E. H. OIPHANS, AND K. HYATT. 1977. kological implications of resource depression. Am. Nat. 100: 345-357.
C W S ~ S E N , J. Ad. 1964. Burning of otolith, a technique for age determi- nation of soles md other fishes. J. Cons. h t . Explor. Mer 29: 73-81.
CR~WDER, E. B. AND W. E. C ~ E R . 1982. Habitat structural c o q k x i w md the interaction between bluegills and their prey. Ecology 63: 1802-18 13.
DAHL, K. 1917. Studier og fow@k over $met og @rretvimd. Centdtrykkeriet, Kristimia. Trout md trout waters in Norway). (English s u m q in Sdmon sand Trout Magazine Nos. 17 md 18, December 1918 and April 1919.
EMARSSON, M. A. 1988. Climatic conditions of the Lake Thingvdlavatn m a . Oikos. (In press)
Fmouso~ , A., AND Fo MM. MASON, 1981. Allozyme evidence fmreprductively isolated sympatric ppuhtisns of brown bout Salmo lrutta L. in b u g h Melvin, Ireland. J. Hish Biol. 18: 629-4542.
Frsmw, R. A. 1930. The genetical theory of natural selection. Clxendon Press, Oxford, England. 272 p.
FROST, W. E. 1965. Breeding habits of Windmere chm, Salvelinus wblkughbii (Giknther), and their bearing on speciation of these fish. RQC. R. Sm. Esnd. Ser. B 163: 232-284.
G R W ~ N , T. A., AND B. J. TURNER. 1984. D k c t evidence that the Blyoden wophs are single biological species. Evolution 38: 4 0 2 4 7 .
GYDEMO, R. 1984. Preliminary survey results of the distribution of the Arctic c h species complex in Iceland, p. 91-107. Hn L. Johnson anand B. L. Bums [ed.] Biology of the Arctic chm. Roc. Int. Symp. Arctic University sf Manitoba Press, Winnipeg, Man.
H~mm, K., AND B. JONSSON. 1982. Habitat mQ food segregation of dwarf and mmd kc t i c char (Salvekinus alpinus) from Vangsvatnet Lake, west- ern Nomay. Cm. J. Hsh. Aquat. Sci. 39: 103&1M5.
HINDAR, K., N. RYMAN, AND G. S T ~ L . 1986. Genetic differentiation mong l w d populations and morghotypes of Arctic chm, Sa1veHinus alpinus. Bid. 9. Linn. Soc. 27: 269-285.
JENSEN, A. E. 1985. Relations mong net reproductive rate and Iife history pameters for I& whitefish (Coregonus clupeafomis). Can. J. Fish. Aquat- Sci. 42: 164-168.
JOHNSON, L. 1980. The Arctic c k , p. 15-9$. In E. K. Bdon led.] Chms, salmonid fishes s f the gmus SalveHin~ss. Dr. W. Junk Publisher, The Hague, The Netherlands.
JOHNSON, L., AND B. L. B m s [ED.] 1984. Biology sf the Arctic chm: Roc. Int. Symp. Arctic Chm. University of Manitoba Ress, Winnipeg, Manapa.
~ ~ N A s S O M , P. Ad., AND K. L ~ E G A A ~ . 1988. k0SyStem sbdi%s of N~~H&IP At1antic Ridge lakes. Verh. Ent. Ver* Limo1. 23. (In press)
JONSSON, B. 1985. Life history patterns of freshwater resident md sea-run migrant brown trout in Noway. T m s . Am. Fish. Sm. 1 14: 182-194.
JONSON, B . , AND K. HINLPBR. 1982. Reproductive strategy of dwarf md noma% Arctic char (SaHvelinats alpinus) h m Vangsvatraet M e , western Norway. Gm. 5. Fish. Aqua. Sci. 39: 1404-1413.
J m s s o ~ , B . , K. H m m , AND T. G . NOR~COE. 1 984. Optimal age at sexual mtufity s f sympatric and experimentdIy dlspatric cu t tha t trout md Dolly Vuden chm. &cologia (Berl .) 6 1 : 3 1 !&325.
J s m s o ~ , B., AND T. BZ~STU. 1979. Demographic strategy in char c o m p d with brown trout in Lake Lme, Western Norway. Rep. Inst. Freshwater Res. Drottningholm 58: 45-54.
SONSSON, B . , AND N. C. S m s ~ m . 1977. A method for estimating fish length from otolith s ix . Rep. Enst. Freshwater Res. Bmtbingholrn 56: 81-86.
KAMLER, E., AND T. KATO. 1983. Efficiency of yolk utilization by Salmo gaird- neri in relation to incubtion temperature end egg size. Pol. Arch. Hydro- bid. 30: 271-30636.
K m o v , K. V. 198 1 . The effect ofthe size of Atlantic salmon eggs on embryos a d devins. J. Fish Biol. 19: 353-360.
ATRICK, M., rn W. K. SUNDER. 1979. Genetics of speciation in lake whitefish in the Allegash bash. Evolution 33: 478-485.
K E ~ T S E N , A., AND P. G E P O ~ . 1980. Coexistence and immigration of two syrnpatric Arctic char9 p. 757-763. In. E. K. Bdon [ed.] Chm, sdmonid fishes sf the gems Sakvekinus. Dr. W. Bunk Publisher, The Hague, The Netherlands.
Can. 3. Fish. Aquat. Sci., Val. 45, 1988
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.

Kmm~sm, A., P. G ~ a m s , H. H01.m~ AND K. K I U S T O ~ S E N . 1985. Bear Island chm. Rep. Inst. Freshwater Res. h t t nhgho lm 62: 98-1 19.
~o-, I., B. c. S m , p. S. GAGHON, AND 9. N. TAYLOR. 1982. m e cichlid fish of Cuatro Clknegas, Mexico: direct evidence of cmspecificity among distinct tmphic morphs. Evolution 36: 65
K N S T J ~ S S ~ , J., AND Pi. ~ ~ ~ H N S S O N . 1984. The ecology,and management of the Arctic cham k M e Myvratn, North Icdand, p. 341-347. Ha L. Johson and B. Bums [ed.] Biology of the Arctic cham. Roc. Int. Syhaap. Arctic chm. University of Mamitoba Press, Winnipeg, Man.
L A S ~ , E. 1983. Decomposition and sedimentation processes in oligotrophic subarctic Lake Thingpdla, Icelmd. B i o s 40: 103-1 12.
L m m m , C. 1980. Bathymetric distribution of ChironowEidae (Diptera) in the oligoakophie 1&e ThingvaHlavatn, Iceland, p. 225-232. Pa D. A. Mur- my [ed. ] Chironsmidae, ecologg., systematics, cytology amd physiology. Pergmsn Press, Oxford and New York.
~ G W S S O N . K. P. , AND M. M. $;WGUSON. 1987. Genetic analysis of four sympatric moqhs of Arctic cham* SakQkveHims alpinus, from 'Fhingvdla- vatn, Iceland. Ewviron. Biol. Hshes 20: 67-73.
~~~ALMQUIST, H. 9. 1988. The diet of &tic c h m (Szkvekinus alpinus) in Thi- dlavatn, Icelmd. M.Sc. thesis, University of Copenhagen, &marBc.
QUIST, H. 1, %. S. S N B ~ S O N , ANEB S. SK~LASOH. 1985. P'Bae biology of Arctic c h m (Sealve&ims a8pim.s (L.)) in M e Thingvdlavatn, Icelmd. I. Feeding ecology. N & t & h d i n g 55: 195-217. (In Icelandic, Emg- lish s m q )
1986. The biology of Arctic cham (Scelvelipaess alpinus (L.)) in Lake 'llingvaHlavatn, Iceland. II. Parasites: plerocercoids of the genus Diphyi- Bsbotri&nm (Cestda). N & t ~ ~ d i m n ~ ~ n n 56: 77-87. (In Icelandic, Ewg- lish s u m q )
MAYNARD S M ~ , J. 1966. Sympatric spciation. Am. Nat. 100: 637450. M u , C. A., AND A. ELBWA. 1985. Reproductive strategies in the stone
Imch Noemchei8m barbatrslus. O&os M. 34 1-349. NIKOUKY, G. $I. 19 bgy of fishes. Academic Press, London. NUEN, P., AND A. 1984. Bhna&omous, resident and lmdbcked
Arctic cham on F i rna rk , N. Norway, p. 79-89. In L. John- son and B. L. Burns [d.] Biology of the Arctic chm. Pars@. Int. Symp. Arctic Chm. University of Manitoba Press, Winnipeg, Man.
N ~ S C P N , N.-A. 1955. Studies on the feeding habits of trout md char in north Swedish lakes. Rep. ha. Freshwater Res. Drottninghdm 36: 163-225.
NUSON, N.-A,, AND 0. F ~ ~ s o N . 197 1. C%aracteristics s f two discrete pop- aallPeions of kc t i c char (Salvelims alpinus L.) in a north Swedish lake. Rep. hst . Freshwater Res. Hkottninghslrn 51: 90-108.
NORDENG, H. 1961. On the biology of char in Salamgen, North Norway. I. Age md spawning frequency determined from scales and otoliths. Nytt Mag. h l . 18: 67-123.
1983. Solution to the "char problem" baed on Amtic char (Salvel- ims alpinus) in Norway. Can. J. Rsh. Aquat. Sci. 4: 1372-1387.
N w , L., J. H m , AND R. G ~ E M O . 198 1. The systematics md biology of l a m d ~ ~ k d populations s f Arctic char from northern Europe. Rep. Inst. Freshwater Res. Drottningholm 59: 128-141.
OMSON, J. 1980. Temperature structure and water chemistry of the caldera 6skjuvatn, Iceland. Limol. Ocemog. 25: 77%788.
K, A., m J. A. E ~ L w . 1982. The impact of predation on life history evolution in Trinidadian guppies (Poecila re~icse~atca). Evolution 36: 160- 177.
Wlm, S. 1985. Stoduv~tn. Orkustofnun. OSROD-75 19. (In Icelandic) &ST, S., AND G. OWSSON. 1986. The ice cover of Lake Thingvdhvatm. N6t-
t t h ~ ~ & n g a K i m 56: 239-258. (h Ielandie, English s u m x y ) ROW, D. A. 1983. An dlwation mCBgHe1 of p w t h and reproduction in fish.
Can. J. Fish. Aquat. Sci. 40: 1395-1404. 1984. The evolution of life history parameters in teleosts. Can* J.
Fish. Aquat. Sci. 41: 989-1868. R w , Pa. 1981. Consemation of genetic resources: experiences from the
brown trout (SaJmo fruttce), p. 61-74. In N. R y m [&.I Rsh gene pols . Presemation of genetic resources in relation to wild fish stocks. Ekol. Bull. (StQc&olpan) 34.
R ~ A N , N., F9 Ws ~ L E AND G . ST^. 1979. Reproductive isolation with little g n d c divergence in sympatric populations of brown trout ( S d m trufta). Genetics 92: 247-262.
RYMAN, N., AND 6 . ST&. 1981. Genetic perspectives of the idenMicratisn and consewration of Scandinavian stocks of fish. Can. 9. Fish. Aqwt. Sci. 38: 1562-1575.
S E ~ D S S O N , B. 1904. Fisheries research 1902. Andvari 29: 79-1 19. (In Icelandic)
SZM~DSSQN, K. 1965. Aus der gmlogischen Geschichte &s Thinpallavatns. N&tthfr~~dingaKinn 35: 103-144. (In Icelandic, German summary)
SAGE, R. D., AND R. K. S E M D ~ . 1975. Tmphic radiation W u g h p l y - rnorphism in cichfid fishes. Roc. Natl. Acd. Sci. USA 72: 466!%4673.
S ~ L W D , 0. T., B. JONSSON, H. MKMQUI~, R. Gmmo, P. M. JONASSON, T. LWEM, S. S. S N O ~ W N , AND S. SKI~ASON. 1987. Habitat use of Arctic cham ScdSvelims akpinus in M e Thingvallavatn, Iceland. hviron. Biol. Fishes 28: 263-274.
S c n a m , W. M. 1974. Selection for optimal life histories: the effects of age structure. Ecology 55: 291-383.
SCWWTR, W. M., m P. E EWN. 1975. The daptive significance sf vari- ations in life history among local ppulations of Atlantic sdmon in North America. Ecology 56: 577-590.
SC~DT-NELSEN, K. 1939. Cornparative studies on the food competition between trout and char. K. Norske Vidensk. Wsk, Skr. 4: 1 4 5 .
SEGEL, S. 1956. Nonparametric statistics for the khaviord sciences. McGraw- Hill, New Yak, NY-
S ~ S L E T , S. 1973. The ecosystem of the Arctic lake Nofdhpna, Jan Mayen blaud, IU. Ecology of Arctic char, Salvelimus caipims (L.). Asmte 6: 43- 54.
SKUUSON, S. 1983. Morphology, growth, and reproductive biology mong different morphs of Arctic chm, Sa8velikts dpinus (L.) in Lake Thi- ngvablavatn. Postgraduate thesis, University of Iceland, Reykjavik, Iee- lmd. (In Icelandic, English s u m m y )
1986. Embryo size and early head shape in four sympahc rnorphs of Arctic c h m (SalveIims alpinus) in TFlningvdlavatn, Iceland. M. Se . the- sis, University of Guelph, Guelph, Ont.
SNOWSON, S. S. 1982. The littoral ecosystem md the ecology of Limnea peregra in Lake Thingvallavatn, Iceland, Ph.D. thesis, University of Lh- eqxml, LiverpoB, Englmd.
S ~ A R N S , S, C. 1976. Life-history tactics: a review of the ideas. Q. Rev. Biol. 51: 3-47.
1980. A new view of life-history evolution. Oikos 35: 266-281. 1983. A natural experiment in life-history evolution: field data on the
introduction of mosquitofish (Gambusia cafinis) to Hawaii. Evolution 37: 601-617.
1984. Heritability estimates for age and length at maturity in two populations of mosquitofish that shared ancestors in 1905. Evolution 38: 368-375.
STUART, T. A. 1953. Spawning migation, repduction and young stages of loch trout (Salma mt ta L.). Sci. hvest. Freshwater Salmon Rsh. Res. Scott. Home Dep. 5: 1-39.
1957. The migration and homing khaviow of brown trout (SaBm trubra L.). Sci. Invest. Freshwater Salmon Fish. Res. Scott. Home h p . 18: 1-27.
S V A ~ S O N , 6. 1951. The mregonid problem. IHI. Whitefish f m the Baltic successfuIly introduced into freshwater in the north of Sweden. Rep. Inst. Freshwater Res. Drottningholrn 30: 151-162.
196 1. Young sibling fish in northwest Europe, p. 498-5 13. Ha W. Blair [ed.] Vertebrate speciation. University of Texas Press, Austin, TX.
'IUwm, B. J., AND D. J. GROSSE. 1980. Trophic differentiation in Ilyodon, a gems of stream-dwelling goodeid fishes: speciation versus ecological gslgmorphism. Evolution 34: 259-270.
YOWPSOS, W. D., AND D. S. ROBSON. 1978. Estimation of population number and mortality rates, p. 137-164. PPO IF. Bagenab [ed.] Methods for assess- ment of fish production in f ~ s h waters. IBP Handbook No. 3. 3rd ed. Blackwen Scientific hblications, Oxford, London, Winburgh, and Mellsome.
Can. 9. Fish. &wt. Sci., VoJ. 45, 6988
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
r B
ror
Jons
son
on 0
6/29
/14
For
pers
onal
use
onl
y.


























