Embed Size (px)
Citation preview

Original article
Invasion of epithelial mammalian cells by Paracoccidioides brasiliensisleads to cytoskeletal rearrangement and apoptosis of the host cell
Maria José Soares Mendes-Giannini a,*, Samira Abdallah Hanna a,Juliana Leal Monteiro da Silva a, Patricia Ferrari Andreotti a, Luciana Raquel Vincenzi a,
Gil Benard b, Henrique Leonel Lenzi c, Christiane Pienna Soares a
a Departamento de Análises Clínicas, Faculdade de Ciências Farmacêuticas-UNESP, Rua Expedicionários do Brasil,1621-CEP, 14801-902 Araraquara, SP, Brazil
b Laboratório de Alergia e Imunologia Clínica e Experimental e Clínica de Doenças Infecciosas e Parasitárias,Faculdade de Medicina, USP, São Paulo, SP, Brazil
c Departamento de Patologia, Instituto Oswaldo Cruz-Fiocruz, Rio de Janeiro, RJ, Brazil
Received 7 November 2003; accepted 11 May 2004
Available online 02 July 2004
Abstract
Paracoccidioides brasiliensis (Pb) yeast cells can enter mammalian cells and probably manipulate the host cell environment to favor theirown growth and survival. We studied the uptake of strain Pb 18 into A549 lung and Vero epithelial cells, with an emphasis on the repercussionsin the cytoskeleton and the apoptosis of host cells. Cytoskeleton components of the host cells, such as actin and tubulin, were involved in theP. brasiliensis invasion process. Cytochalasin D and colchicine treatment substantially reduced invasion, indicating the functional participa-tion of microfilaments (MFs) and microtubules (MTs) in this mechanism. Cytokeratin could also play a role in the P. brasiliensis interactionwith the host. Gp43 was recognized by anti-actin and anti-cytokeratin antibodies, but not by anti-tubulin. The apoptosis induced by this fungusin infected epithelial cells was demonstrated by various techniques: TUNEL, DNA fragmentation and Bak and Bcl-2 immunocytochemicalexpression. DNA fragmentation was observed in infected cells but not in uninfected ones, by both TUNEL and gel electrophoresis methods.Moreover, Bcl-2 and Bak did not show any differences until 24 h after infection of cells, suggesting a competitive mechanism that allowspersistence of infection. Overexpression of Bak was observed after 48 h, indicating the loss of competition between death and survival signals.In conclusion, the mechanisms of invasion of host cells, persistence within them, and the subsequent induction of apoptosis of such cells mayexplain the efficient dissemination of P. brasiliensis.© 2004 Published by Elsevier SAS.
Keywords: Paracoccidioides brasiliensis; Cytoskeleton; Invasion; Apoptosis; Host–cell biology
1. Introduction
The thermal dimorphic fungus Paracoccidioides brasil-iensis causes paracoccidioidomycosis (PCM), the leadingendemic deep mycosis in Latin America. The disease ischaracterized by a chronic granulomatous inflammation, andpatients may present a spectrum of clinical manifestations,ranging from benign and localized to severe and dissemi-nated forms [1]. The adhesion of the infectious propagules to
host cells is a crucial step to further invasion, colonizationand growth and, during the infection, P. brasiliensis candevelop various phenotypes (growth, invasion and metasta-sis), depending on the fungal strain, the host and other factors[2,3].
Although not traditionally considered a typical intracellu-lar pathogen, P. brasiliensis can enter epithelial cells in vitro[4,5] and in vivo [6,7]. The ability of P. brasiliensis to adhereto and invade non-professional phagocytes has been recog-nized in previous studies [3–5]. We observed adhesion ofP. brasiliensis to the cell membrane, followed by its engulf-ment by cellular extensions, inclusion in phagosomal vacu-
* Corresponding author. Tel.: +55-16-201-6556; fax: +55-16-201-6547.E-mail address: [email protected] (M.J. Soares Mendes-
Giannini).
Microbes and Infection 6 (2004) 882–891
www.elsevier.com/locate/micinf
1286-4579/$ - see front matter © 2004 Published by Elsevier SAS.doi:10.1016/j.micinf.2004.05.005

oles and the tendency of the fungus to be located close to thenuclear membrane [5].
Pathogenic microorganisms utilize a variety of molecularstrategies that subvert host cell mechanisms and enable thesepathogens to invade susceptible host cells [8–11]. Somemicroorganisms do not require the overt involvement ofmicrotubules (MTs) or microfilaments (MFs), whereas oth-ers induce a host actin-based cytoskeletal reorganization,which leads to MF-dependent macropinocytotic membraneengulfment of these pathogens [8–10]. However, the recruit-ment of MT, MF and intermediary filament proteins, follow-ing adherence to the cell surface, has not yet been investi-gated in P. brasiliensis–epithelial cell interaction.
Another consequence of invasion by intracellular patho-gens is apoptosis or programmed cell death of the host cell,which is observed in professional or non-professional phago-cytic cells [12]. Recently, Cacere et al. [13] suggested that inPCM patients, apoptosis plays a role in the antigen-specifichyporesponsiveness of T cells to the main antigen of P. bra-siliensis, the 43-kDa glycoprotein. The ability of pathogensto induce apoptosis of phagocytes might be an importantvirulence factor, for it would curtail the host’s defensemechanisms. P. brasiliensis and other fungi can exploit ph-agocyte apoptosis to their own advantage, and their intracel-lular residence in epithelial cells could potentially elicit thistype of cell death response. Therefore, we studied the inter-action between P. brasiliensis and epithelial cells, with par-ticular emphasis on the cytoskeleton changes and the mo-lecular events leading to apoptosis.
2. Materials and methods
2.1. Organism
The yeast form of P. brasiliensis strain 18 was maintainedin peptone–yeast extract–glucose (PYG) medium at 37 °Cfor 3 days and suspended in phosphate-buffered saline (PBS)at 1.0 × 105 cells per ml for use in the adherence andinternalization assays.
2.2. Antisera and reagents
The cell-free antigen, a rich solution of cell-wall-associated antigens, was prepared as described elsewhere[14]. The polyclonal antibody raised against cell-free antigenof P. brasiliensis was prepared in rabbits by intradermalinjections of 1.0 ml of antigen mixed with 1.0 ml of completeFreund’s adjuvant. Subsequent injections of antigen withincomplete Freund’s adjuvant were given weekly for a periodof 4 weeks and then monthly for a period of 3 months. Therabbits were bled 7 days after the last dose. The immunoglo-bulin fractions of the antiserum were obtained by precipita-tion with ammonium sulfate.
Monoclonal antibody (Mab) anti-human actin, anti-tubulin, anti-pan cytokeratin, fluorescein isothiocyanate
(FITC)-labeled phalloidin, anti-rabbit IgG FITC conjugate,anti-mouse IgG FITC conjugate, anti-mouse/rabbit IgG per-oxidase conjugate, anti-rabbit/mouse IgG tetramethyl-rhodamine isothiocyanate (TRITC) conjugate and anti-rabbitIgG R-phycoerythrin conjugate were purchased from Sigma,St. Louis, MO, USA. Monoclonal anti-human Bcl-2 onco-protein and rabbit polyclonal anti-human Bak were pur-chased from Dako Corporation, Glostrup, Denmark.
2.3. Epithelial cells
The type II pneumocyte cell line A549 and Vero cells(African green monkey kidney) were obtained from theAmerican Type Culture Collection (Manassas, VA). The cellswere seeded overnight in Dulbecco’s modified Eagle’s me-dium (DMEM) (InstitutoAdolfo Lutz, Brazil), supplementedwith 10% heat-inactivated fetal calf serum (Cultilab, Brazil).
2.4. Assay of P. brasiliensis–cell interaction
Cells were cultured at 36.5 °C in 24-well plates, with thewell-bottoms covered with coverslips, and adjusted to 3.0 ×104 cells per well. Then, 1.0 × 105 yeast forms per ml ofP. brasiliensis were added to the cells, reaching ayeast/epithelial cell ratio of 30:1, and incubated for 2–5 h at36.5 °C, to observe adhesion and invasion processes. Aftereach period of incubation, the medium was discarded, thecells were washed with PBS and the coverslips bearing thecultured cells were fixed with 2% paraformaldehyde andanalyzed by conventional fluorescence and scanning confo-cal laser (LSM-410, Carl Zeiss, Jena, Göttingen) micros-copy.
2.5. Ketoconazole protection assay
The adhesion and invasion was also evaluated by keto-conazole protection assay as previously described [5]. Ac-cording to previous data [4] ketoconazole does not kill theintracellular fungi, but only the extracellular. The objectiveof this assay is to evaluate quantitatively the P. brasiliensisinvasion of epithelial cells. The epithelial cells growing inmicrotiter plates were incubated with a suspension of P. bra-siliensis 18 (1.0 × 105 yeasts per ml) for 1, 2, 3, 5, 8 and 24 hat 36.5 °C. Thereafter, the plates were washed three timeswith PBS to remove non-adherent yeast forms and subse-quently overlaid with 15 µg/ml of ketoconazole solution inMEM for 1 h at 36.5 °C, to kill the cells that had not beeninternalized. The infected mammalian cells were lysed with100 µl of MilliQ-water and the suspension was plated onmodified synthetic medium. The ketoconazole concentrationwas chosen in previous titration experiments for its ability tokill the fungi but not the cells. This experiment was per-formed by the plating assay and vital dye staining (trypanblue). Epithelial cells infected with fungi, not treated withketoconazole were plated, as a positive control of infection.The infected epithelial cells treated with ketoconazole were
883M.J. Soares Mendes-Giannini et al. / Microbes and Infection 6 (2004) 882–891

lysed and plated, and the observed fungal growth corre-sponded to the internalized, live fungal cells. After 5 days,colonies were counted independently to determine the totaland internalized P. brasiliensis yeast cells. With the countsfrom the positive control and the ketoconazole-treated fungalcells, the percentage of adhering yeasts was calculated. As anadditional control, experiments were run in parallel with anon-pathogenic strain of Saccharomyces cerevisiae to checkfor the specific features of the P. brasiliensis invasion pro-cess.
2.6. Immunofluorescence assays for determinationof cytoskeleton assembly
To study cytoskeletal changes that occur upon P. brasil-iensis infection, infected cell monolayers were fixed asabove, washed in PBS, and permeabilized in 0.5% TritonX-100 for 15 min. After the washing step, primary anti-cytoskeleton (actin, tubulin and cytokeratin) or anti-P. bra-siliensis (cell-free) were added for 1 h, unbound antibodieswere removed by washing in PBS, and then secondary con-jugated antibodies (FITC, TRITC or phycoerythrin labeled)were added for 1 h. The cytoskeleton components and P. bra-siliensis were observed simultaneously using double filtersets, fluorescein, tetramethylrhodamine or phycoerythrin.Following infection, the coverslips were washed three timeswith PBS and fixed with 2.0% paraformaldehyde, thenwashed extensively, and the cells were permeabilized. Tostain F-actin we also used FITC-labeled phalloidin. It wasdiluted in PBS, suspended in Evans blue and used at aconcentration of 10 µg/ml at 37 °C for 1 h, after which thecells were washed three times with PBS containing 0.2%Tween-20 (PBS-T), mounted in PBS-glycerin and viewed byfluorescence microscopy (Leica).
2.7. Cytoskeleton involvement: effects of cytochalasin Dand colchicine
Compounds that cause depolymerization of MFs or MTswere individually used to pretreat A549 or Vero-cell mono-layers before and during the 5-h invasion period. In allexperiments, epithelial cells were pretreated with cytochala-sin D at 1.0 and 10 µg/ml for 30 min [15] or colchicine (1.0,5.0 and 25 µg/ml) in appropriate tissue culture medium usinga stock solution of 1.0 mg/ml in absolute ethanol at –20 °C.Results of preliminary experiments indicated that 5.0 µg/mlwas the optimal concentration to modulate the internalizationof P. brasiliensis. Concomitantly, P. brasiliensis yeast cellswere also submitted to a preliminary treatment with cytocha-lasin D and colchicine. We also included a control withfungal cells killed by treatment with ketoconazole, followedby an experiment with epithelial cells treated with colchicineand cytochalasin D. The number of fungi internalized withinepithelial cells was quantified in the cultures. The degree ofinternalization of P. brasiliensis was determined after keto-conazole treatment, as described above. Intracellular fungi,
pretreated or not with cytochalasin D and colchicine, werecounted, and data for all binding and invasion assays wereexpressed as mean ± S.D. (n = 3).
2.8. Influence of gp43 on cytoskeleton
2.8.1. Purification of gp43Gp43 was purified from crude exoantigen of P. brasilien-
sis, as previously described [16]. Briefly, the exoantigen wasfractionated by affinity chromatography on columns of pro-tein A-purified rabbit anti-gp43 IgG coupled to CNBr-Sepharose (Pharmacia-Biotech), and further purification wasachieved by gel filtration. Analysis by sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) andsilver staining showed a single band with a molecular mass of43 kDa.
2.8.2. Gp43 treatment of Vero cellsVero cells were cultured at 106 per ml in 100 µl per well in
a 24-well plate at 37 °C. Various concentrations (6.25,12.5 and 25.0 µg/ml) of purified gp43 were added at thebeginning of incubation. We used a control with the gp43denatured at a high temperature before being added to thecell monolayers. After 5 h, the cells were washed and theF-actin and cytokeratin filaments were observed by FITC-phalloidin and pan-cytokeratin antibody, respectively, usingimmunofluorescence assays. The percentage of viable cellswas estimated with trypan blue dye and untreated epithelialcells were used as a control.
2.9. Protein blot analysis
P. brasiliensis cell-free antigen was separated by SDS-PAGE and transferred to a nitrocellulose membrane. Immu-noblot analysis was carried out as previously described [17].Briefly, blots were prepared, blocked with 1% bovine serumalbumin (BSA) in PBS for 4 h, and then probed with mono-clonal anti-actin, anti-tubulin and anti-pan-cytokeratin anti-bodies diluted in immunoblot-blocking solution (PBS con-taining 5% nonfat dry milk and 0.1% Tween-20) overnight at4 °C. The blots were washed briefly and then incubated for1 h at 4 °C with a goat anti-mouse immunoglobulin–rabbitperoxidase conjugate diluted 1:500 in blocking solution. Theblots were washed and then developed with diaminobenzi-dine (Sigma).
2.10. Apoptosis assay
2.10.1. TUNEL techniqueThe terminal D-transferase-mediated dUTP-fluorescein
nick-end labeling (TUNEL) assay was done essentially asdescribed by Zychlinsky et al. [18]. In brief, epithelial cellswere grown on glass coverslips and incubated for 1, 2, 3, 5, 8,24 and 30 h with P. brasiliensis, washed and fixed withparaformaldehyde, as described in Section 2.6. The cover-slips were blocked with BSA, permeabilized for 15 min with
884 M.J. Soares Mendes-Giannini et al. / Microbes and Infection 6 (2004) 882–891
0.2% Triton X-100 and incubated with terminalD-transferase and fluorescent dUTP (Boehringer Man-nheim). Then, the coverslips were incubated in a humidchamber with anti-cell-free P. brasiliensis serum. After60 min of incubation, the coverslips were washed and incu-bated with secondary antibody conjugated with TRITC.Apo-ptotic bodies and P. brasiliensis were visualized simulta-neously using double filter sets for FITC and TRITC.
2.10.2. Analysis of DNA fragmentationEpithelial cells (3.0 × 104 cells per well) were infected
with P. brasiliensis for 30 h. The cells were harvested forDNA isolation, and detached epithelial cells were centri-fuged at 1000 × g for 5 min at 4 °C, and the pellet was washedwith ice-cold PBS and resuspended in lysis buffer. DNA wasisolated from the lysates with the GFX™ Pharmacia Kit,according to the manufacturers. Electrophoresis was per-formed with a 2% agarose gel and DNA was stained withethidium bromide. As a negative control, uninfected cellswere treated by the same procedure as the infected cells.
2.10.3. In situ immunoperoxidase—Bcl-2 and BakEpithelial cells were grown in 24-well plates to confluence
and incubated with P. brasiliensis for 1, 2, 3, 5, 8, 24 and48 h, to study the pattern and distribution of Bcl-2 and Bak bythe in situ immunoperoxidase reaction. The reaction wasdeveloped using anti-Bak and anti-Bcl-2 antibodies diluted1:200 and 1:80 in PBS, respectively. As a positive control,epithelial cells were treated with methotrexate at 1 µM for24 h [19].
2.10.4. StatisticsData from repeated experiments were analyzed by un-
paired Student’s t-test.
3. Results
3.1. Invasion assay using ketoconazole
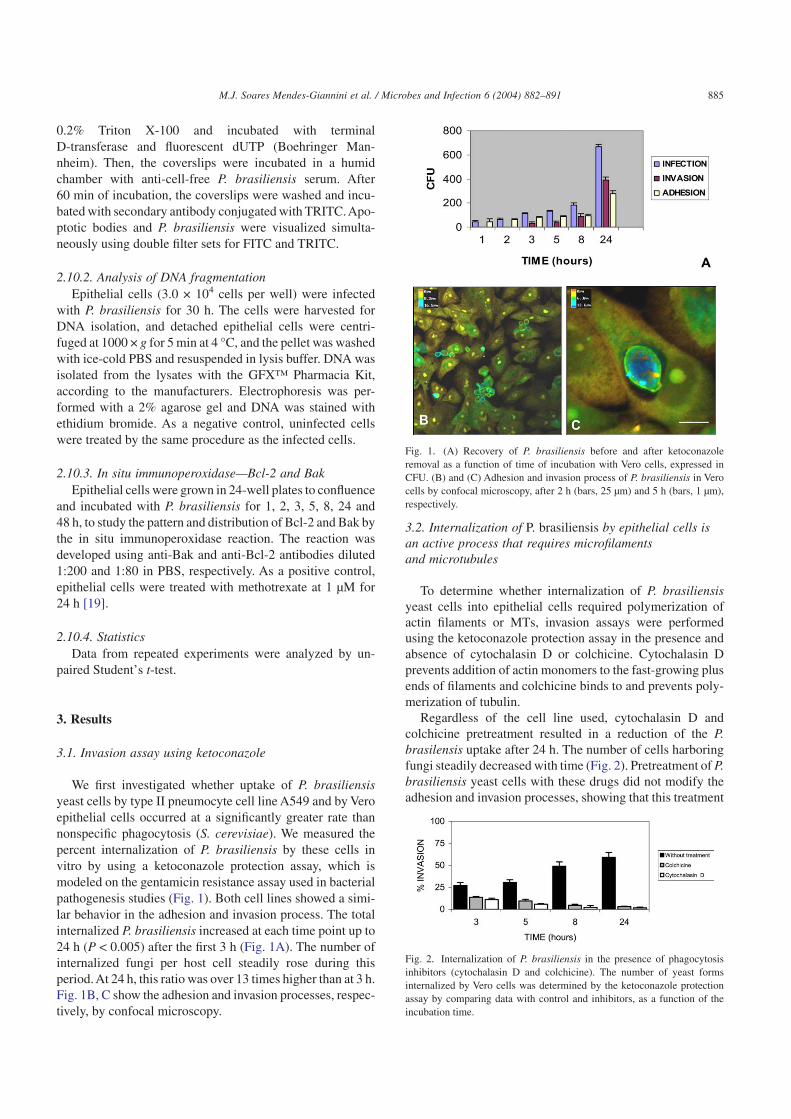
We first investigated whether uptake of P. brasiliensisyeast cells by type II pneumocyte cell line A549 and by Veroepithelial cells occurred at a significantly greater rate thannonspecific phagocytosis (S. cerevisiae). We measured thepercent internalization of P. brasiliensis by these cells invitro by using a ketoconazole protection assay, which ismodeled on the gentamicin resistance assay used in bacterialpathogenesis studies (Fig. 1). Both cell lines showed a simi-lar behavior in the adhesion and invasion process. The totalinternalized P. brasiliensis increased at each time point up to24 h (P < 0.005) after the first 3 h (Fig. 1A). The number ofinternalized fungi per host cell steadily rose during thisperiod. At 24 h, this ratio was over 13 times higher than at 3 h.Fig. 1B, C show the adhesion and invasion processes, respec-tively, by confocal microscopy.
3.2. Internalization of P. brasiliensis by epithelial cells isan active process that requires microfilamentsand microtubules
To determine whether internalization of P. brasiliensisyeast cells into epithelial cells required polymerization ofactin filaments or MTs, invasion assays were performedusing the ketoconazole protection assay in the presence andabsence of cytochalasin D or colchicine. Cytochalasin Dprevents addition of actin monomers to the fast-growing plusends of filaments and colchicine binds to and prevents poly-merization of tubulin.
Regardless of the cell line used, cytochalasin D andcolchicine pretreatment resulted in a reduction of the P.brasilensis uptake after 24 h. The number of cells harboringfungi steadily decreased with time (Fig. 2). Pretreatment of P.brasiliensis yeast cells with these drugs did not modify theadhesion and invasion processes, showing that this treatment
Fig. 1. (A) Recovery of P. brasiliensis before and after ketoconazoleremoval as a function of time of incubation with Vero cells, expressed inCFU. (B) and (C) Adhesion and invasion process of P. brasiliensis in Verocells by confocal microscopy, after 2 h (bars, 25 µm) and 5 h (bars, 1 µm),respectively.
Fig. 2. Internalization of P. brasiliensis in the presence of phagocytosisinhibitors (cytochalasin D and colchicine). The number of yeast formsinternalized by Vero cells was determined by the ketoconazole protectionassay by comparing data with control and inhibitors, as a function of theincubation time.
885M.J. Soares Mendes-Giannini et al. / Microbes and Infection 6 (2004) 882–891
does not directly affect fungal viability. The controls withfungi killed with ketoconazole showed no fungi adheringeither to the cells treated with the drugs (cytochalasin andcolchicine) or to the untreated epithelial cells.
3.3. Actin network and microtubules
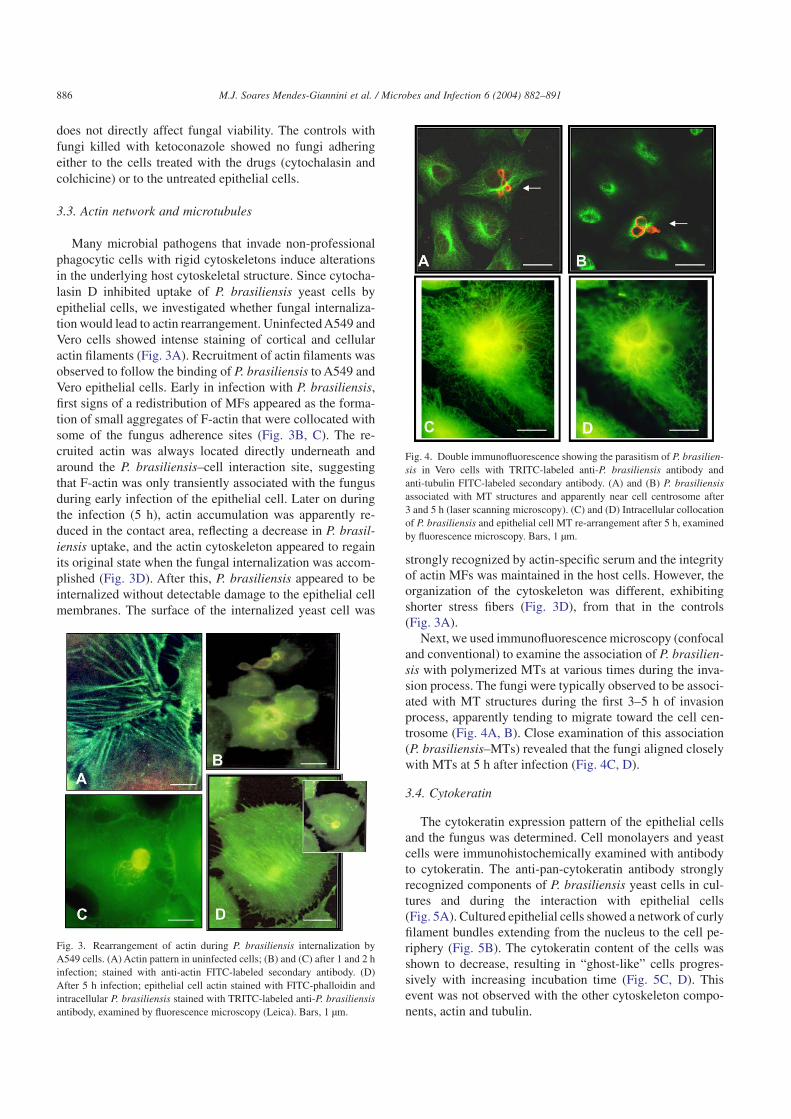
Many microbial pathogens that invade non-professionalphagocytic cells with rigid cytoskeletons induce alterationsin the underlying host cytoskeletal structure. Since cytocha-lasin D inhibited uptake of P. brasiliensis yeast cells byepithelial cells, we investigated whether fungal internaliza-tion would lead to actin rearrangement. Uninfected A549 andVero cells showed intense staining of cortical and cellularactin filaments (Fig. 3A). Recruitment of actin filaments wasobserved to follow the binding of P. brasiliensis to A549 andVero epithelial cells. Early in infection with P. brasiliensis,first signs of a redistribution of MFs appeared as the forma-tion of small aggregates of F-actin that were collocated withsome of the fungus adherence sites (Fig. 3B, C). The re-cruited actin was always located directly underneath andaround the P. brasiliensis–cell interaction site, suggestingthat F-actin was only transiently associated with the fungusduring early infection of the epithelial cell. Later on duringthe infection (5 h), actin accumulation was apparently re-duced in the contact area, reflecting a decrease in P. brasil-iensis uptake, and the actin cytoskeleton appeared to regainits original state when the fungal internalization was accom-plished (Fig. 3D). After this, P. brasiliensis appeared to beinternalized without detectable damage to the epithelial cellmembranes. The surface of the internalized yeast cell was
strongly recognized by actin-specific serum and the integrityof actin MFs was maintained in the host cells. However, theorganization of the cytoskeleton was different, exhibitingshorter stress fibers (Fig. 3D), from that in the controls(Fig. 3A).
Next, we used immunofluorescence microscopy (confocaland conventional) to examine the association of P. brasilien-sis with polymerized MTs at various times during the inva-sion process. The fungi were typically observed to be associ-ated with MT structures during the first 3–5 h of invasionprocess, apparently tending to migrate toward the cell cen-trosome (Fig. 4A, B). Close examination of this association(P. brasiliensis–MTs) revealed that the fungi aligned closelywith MTs at 5 h after infection (Fig. 4C, D).
3.4. Cytokeratin
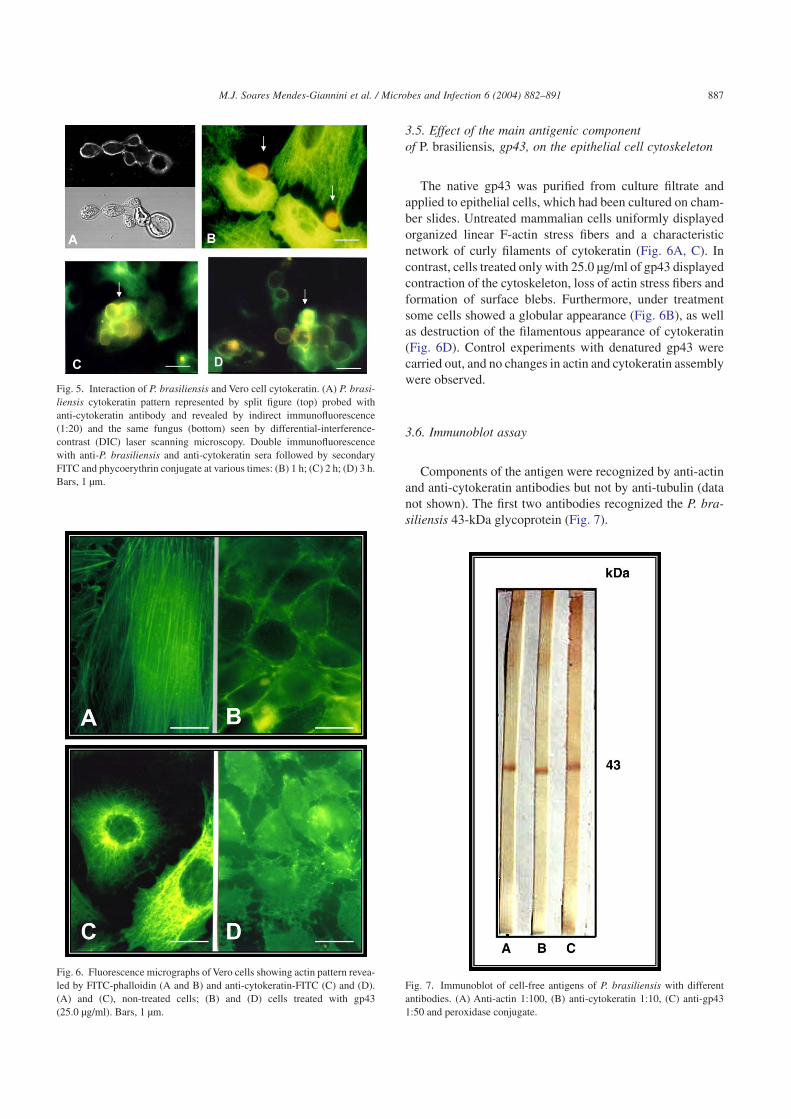
The cytokeratin expression pattern of the epithelial cellsand the fungus was determined. Cell monolayers and yeastcells were immunohistochemically examined with antibodyto cytokeratin. The anti-pan-cytokeratin antibody stronglyrecognized components of P. brasiliensis yeast cells in cul-tures and during the interaction with epithelial cells(Fig. 5A). Cultured epithelial cells showed a network of curlyfilament bundles extending from the nucleus to the cell pe-riphery (Fig. 5B). The cytokeratin content of the cells wasshown to decrease, resulting in “ghost-like” cells progres-sively with increasing incubation time (Fig. 5C, D). Thisevent was not observed with the other cytoskeleton compo-nents, actin and tubulin.
Fig. 3. Rearrangement of actin during P. brasiliensis internalization byA549 cells. (A) Actin pattern in uninfected cells; (B) and (C) after 1 and 2 hinfection; stained with anti-actin FITC-labeled secondary antibody. (D)After 5 h infection; epithelial cell actin stained with FITC-phalloidin andintracellular P. brasiliensis stained with TRITC-labeled anti-P. brasiliensisantibody, examined by fluorescence microscopy (Leica). Bars, 1 µm.
Fig. 4. Double immunofluorescence showing the parasitism of P. brasilien-sis in Vero cells with TRITC-labeled anti-P. brasiliensis antibody andanti-tubulin FITC-labeled secondary antibody. (A) and (B) P. brasiliensisassociated with MT structures and apparently near cell centrosome after3 and 5 h (laser scanning microscopy). (C) and (D) Intracellular collocationof P. brasiliensis and epithelial cell MT re-arrangement after 5 h, examinedby fluorescence microscopy. Bars, 1 µm.
886 M.J. Soares Mendes-Giannini et al. / Microbes and Infection 6 (2004) 882–891
3.5. Effect of the main antigenic componentof P. brasiliensis, gp43, on the epithelial cell cytoskeleton
The native gp43 was purified from culture filtrate andapplied to epithelial cells, which had been cultured on cham-ber slides. Untreated mammalian cells uniformly displayedorganized linear F-actin stress fibers and a characteristicnetwork of curly filaments of cytokeratin (Fig. 6A, C). Incontrast, cells treated only with 25.0 µg/ml of gp43 displayedcontraction of the cytoskeleton, loss of actin stress fibers andformation of surface blebs. Furthermore, under treatmentsome cells showed a globular appearance (Fig. 6B), as wellas destruction of the filamentous appearance of cytokeratin(Fig. 6D). Control experiments with denatured gp43 werecarried out, and no changes in actin and cytokeratin assemblywere observed.
3.6. Immunoblot assay
Components of the antigen were recognized by anti-actinand anti-cytokeratin antibodies but not by anti-tubulin (datanot shown). The first two antibodies recognized the P. bra-siliensis 43-kDa glycoprotein (Fig. 7).
Fig. 5. Interaction of P. brasiliensis and Vero cell cytokeratin. (A) P. brasi-liensis cytokeratin pattern represented by split figure (top) probed withanti-cytokeratin antibody and revealed by indirect immunofluorescence(1:20) and the same fungus (bottom) seen by differential-interference-contrast (DIC) laser scanning microscopy. Double immunofluorescencewith anti-P. brasiliensis and anti-cytokeratin sera followed by secondaryFITC and phycoerythrin conjugate at various times: (B) 1 h; (C) 2 h; (D) 3 h.Bars, 1 µm.
Fig. 6. Fluorescence micrographs of Vero cells showing actin pattern revea-led by FITC-phalloidin (A and B) and anti-cytokeratin-FITC (C) and (D).(A) and (C), non-treated cells; (B) and (D) cells treated with gp43(25.0 µg/ml). Bars, 1 µm.
Fig. 7. Immunoblot of cell-free antigens of P. brasiliensis with differentantibodies. (A) Anti-actin 1:100, (B) anti-cytokeratin 1:10, (C) anti-gp431:50 and peroxidase conjugate.
887M.J. Soares Mendes-Giannini et al. / Microbes and Infection 6 (2004) 882–891
3.7. Apoptosis assay
3.7.1. TUNEL techniqueTo determine whether epithelial cells undergo apoptosis
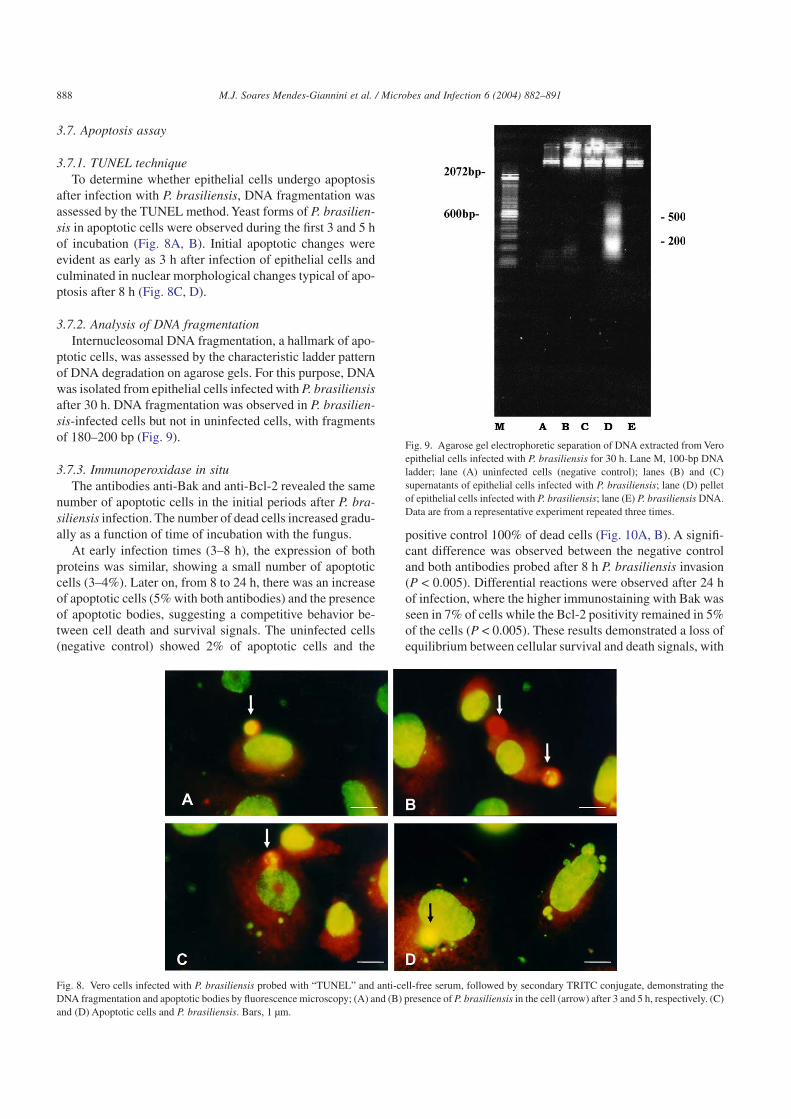
after infection with P. brasiliensis, DNA fragmentation wasassessed by the TUNEL method. Yeast forms of P. brasilien-sis in apoptotic cells were observed during the first 3 and 5 hof incubation (Fig. 8A, B). Initial apoptotic changes wereevident as early as 3 h after infection of epithelial cells andculminated in nuclear morphological changes typical of apo-ptosis after 8 h (Fig. 8C, D).
3.7.2. Analysis of DNA fragmentationInternucleosomal DNA fragmentation, a hallmark of apo-
ptotic cells, was assessed by the characteristic ladder patternof DNA degradation on agarose gels. For this purpose, DNAwas isolated from epithelial cells infected with P. brasiliensisafter 30 h. DNA fragmentation was observed in P. brasilien-sis-infected cells but not in uninfected cells, with fragmentsof 180–200 bp (Fig. 9).
3.7.3. Immunoperoxidase in situThe antibodies anti-Bak and anti-Bcl-2 revealed the same
number of apoptotic cells in the initial periods after P. bra-siliensis infection. The number of dead cells increased gradu-ally as a function of time of incubation with the fungus.
At early infection times (3–8 h), the expression of bothproteins was similar, showing a small number of apoptoticcells (3–4%). Later on, from 8 to 24 h, there was an increaseof apoptotic cells (5% with both antibodies) and the presenceof apoptotic bodies, suggesting a competitive behavior be-tween cell death and survival signals. The uninfected cells(negative control) showed 2% of apoptotic cells and the
positive control 100% of dead cells (Fig. 10A, B). A signifi-cant difference was observed between the negative controland both antibodies probed after 8 h P. brasiliensis invasion(P < 0.005). Differential reactions were observed after 24 hof infection, where the higher immunostaining with Bak wasseen in 7% of cells while the Bcl-2 positivity remained in 5%of the cells (P < 0.005). These results demonstrated a loss ofequilibrium between cellular survival and death signals, with
Fig. 8. Vero cells infected with P. brasiliensis probed with “TUNEL” and anti-cell-free serum, followed by secondary TRITC conjugate, demonstrating theDNA fragmentation and apoptotic bodies by fluorescence microscopy; (A) and (B) presence of P. brasiliensis in the cell (arrow) after 3 and 5 h, respectively. (C)and (D) Apoptotic cells and P. brasiliensis. Bars, 1 µm.
Fig. 9. Agarose gel electrophoretic separation of DNA extracted from Veroepithelial cells infected with P. brasiliensis for 30 h. Lane M, 100-bp DNAladder; lane (A) uninfected cells (negative control); lanes (B) and (C)supernatants of epithelial cells infected with P. brasiliensis; lane (D) pelletof epithelial cells infected with P. brasiliensis; lane (E) P. brasiliensis DNA.Data are from a representative experiment repeated three times.
888 M.J. Soares Mendes-Giannini et al. / Microbes and Infection 6 (2004) 882–891
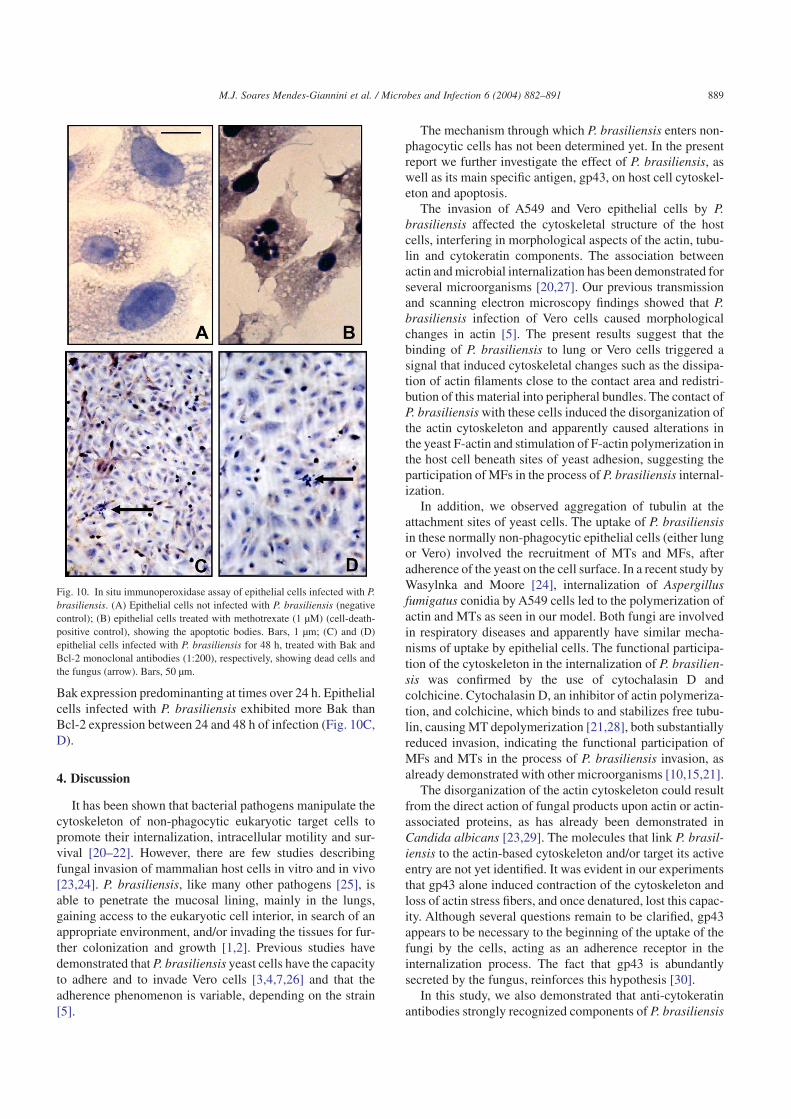
Bak expression predominanting at times over 24 h. Epithelialcells infected with P. brasiliensis exhibited more Bak thanBcl-2 expression between 24 and 48 h of infection (Fig. 10C,D).
4. Discussion
It has been shown that bacterial pathogens manipulate thecytoskeleton of non-phagocytic eukaryotic target cells topromote their internalization, intracellular motility and sur-vival [20–22]. However, there are few studies describingfungal invasion of mammalian host cells in vitro and in vivo[23,24]. P. brasiliensis, like many other pathogens [25], isable to penetrate the mucosal lining, mainly in the lungs,gaining access to the eukaryotic cell interior, in search of anappropriate environment, and/or invading the tissues for fur-ther colonization and growth [1,2]. Previous studies havedemonstrated that P. brasiliensis yeast cells have the capacityto adhere and to invade Vero cells [3,4,7,26] and that theadherence phenomenon is variable, depending on the strain[5].
The mechanism through which P. brasiliensis enters non-phagocytic cells has not been determined yet. In the presentreport we further investigate the effect of P. brasiliensis, aswell as its main specific antigen, gp43, on host cell cytoskel-eton and apoptosis.
The invasion of A549 and Vero epithelial cells by P.brasiliensis affected the cytoskeletal structure of the hostcells, interfering in morphological aspects of the actin, tubu-lin and cytokeratin components. The association betweenactin and microbial internalization has been demonstrated forseveral microorganisms [20,27]. Our previous transmissionand scanning electron microscopy findings showed that P.brasiliensis infection of Vero cells caused morphologicalchanges in actin [5]. The present results suggest that thebinding of P. brasiliensis to lung or Vero cells triggered asignal that induced cytoskeletal changes such as the dissipa-tion of actin filaments close to the contact area and redistri-bution of this material into peripheral bundles. The contact ofP. brasiliensis with these cells induced the disorganization ofthe actin cytoskeleton and apparently caused alterations inthe yeast F-actin and stimulation of F-actin polymerization inthe host cell beneath sites of yeast adhesion, suggesting theparticipation of MFs in the process of P. brasiliensis internal-ization.
In addition, we observed aggregation of tubulin at theattachment sites of yeast cells. The uptake of P. brasiliensisin these normally non-phagocytic epithelial cells (either lungor Vero) involved the recruitment of MTs and MFs, afteradherence of the yeast on the cell surface. In a recent study byWasylnka and Moore [24], internalization of Aspergillusfumigatus conidia by A549 cells led to the polymerization ofactin and MTs as seen in our model. Both fungi are involvedin respiratory diseases and apparently have similar mecha-nisms of uptake by epithelial cells. The functional participa-tion of the cytoskeleton in the internalization of P. brasilien-sis was confirmed by the use of cytochalasin D andcolchicine. Cytochalasin D, an inhibitor of actin polymeriza-tion, and colchicine, which binds to and stabilizes free tubu-lin, causing MT depolymerization [21,28], both substantiallyreduced invasion, indicating the functional participation ofMFs and MTs in the process of P. brasiliensis invasion, asalready demonstrated with other microorganisms [10,15,21].
The disorganization of the actin cytoskeleton could resultfrom the direct action of fungal products upon actin or actin-associated proteins, as has already been demonstrated inCandida albicans [23,29]. The molecules that link P. brasil-iensis to the actin-based cytoskeleton and/or target its activeentry are not yet identified. It was evident in our experimentsthat gp43 alone induced contraction of the cytoskeleton andloss of actin stress fibers, and once denatured, lost this capac-ity. Although several questions remain to be clarified, gp43appears to be necessary to the beginning of the uptake of thefungi by the cells, acting as an adherence receptor in theinternalization process. The fact that gp43 is abundantlysecreted by the fungus, reinforces this hypothesis [30].
In this study, we also demonstrated that anti-cytokeratinantibodies strongly recognized components of P. brasiliensis
Fig. 10. In situ immunoperoxidase assay of epithelial cells infected with P.brasiliensis. (A) Epithelial cells not infected with P. brasiliensis (negativecontrol); (B) epithelial cells treated with methotrexate (1 µM) (cell-death-positive control), showing the apoptotic bodies. Bars, 1 µm; (C) and (D)epithelial cells infected with P. brasiliensis for 48 h, treated with Bak andBcl-2 monoclonal antibodies (1:200), respectively, showing dead cells andthe fungus (arrow). Bars, 50 µm.
889M.J. Soares Mendes-Giannini et al. / Microbes and Infection 6 (2004) 882–891

yeast cells, mainly gp43, suggesting that the fungus and thecell cytokeratin filaments possess similar components. Thisnew observation stresses the importance of studying moredeeply the fungal cytoskeleton. It has a role in fungal growth,differentiation and the behavior of fungal zoospores [31] butmany of the components of this system remain poorly char-acterized in fungi. On the other hand, the degradation ofcytokeratin components by P. brasiliensis may be due to theeffect of specific enzymes [32,33] or of gp43, which causedthe loss of its characteristic filament network. The role, if any,of P. brasiliensis adherence to cytokeratin in the pathogen-esis of paracoccidioidomycosis is unclear. Interactions be-tween P. brasiliensis and cytokeratin may take place afterepithelial cells are damaged by gp43 or other factors releasedby P. brasiliensis. Thus, we postulate that adherence ofP. brasiliensis to cytokeratin could play a role in either theinvasion process in vitro or in the process by which P. bra-siliensis crosses epithelial barriers in vivo. P. brasiliensismay take advantage of this system by adhering to cytoskel-etal components such actin MFs, MTs and cytokeratin fila-ments, as demonstrated in other microorganisms [10,27,34,35].
On the other hand, apoptosis has been implicated in themechanism of bacterial pathogenesis [12], and this involves avariety of host–pathogen interactions [36,37]. In this study,we evaluated the induction of apoptosis in epithelial cells byP. brasiliensis and found that the number of apoptotic cellsincreased with time of fungal exposure. This result suggests arequirement of intracellular residence of the fungus, to trig-ger apoptosis of epithelial cells. Other bacterial pathogens,such as Shigella flexneri, need to be internalized to causeapoptosis [18]. Recently, we demonstrated that gp43 inducedhigh levels of apoptosis in peripheral blood mononuclearcells [13]. This molecule may also be responsible for induc-tion of apoptosis in epithelial cells. In this study, we providethe first evidence that P. brasiliensis can induce apoptosis inepithelial cells, demonstrated by the Bak/Bcl-2 immunoper-oxidase reaction, TUNEL assays, and the DNA fragmenta-tion pattern. We observed similar immunostaining patternsfor Bcl-2 and Bak until 24 h of infection, suggesting acompetitive mechanism to promote persistent fungal infec-tion. The mechanism by which P. brasiliensis regulates ex-pression of Bcl-2 and Bak is currently under investigation. Arecent study showed that overexpression of Bcl-2 oncogenewas related to the establishment of persistent viral infectionin a malignant prostate cell line [38]. Infection with Sindbisvirus in certain hamster cell lines resulted in rapid death, aphenomenon that was considerably delayed by stable over-expression of Bcl2 genes [39]. Moreover, another reportshows that a virulent subtype of this virus leads to persistentinfection and increases the expression of Bcl-2, while anavirulent subtype behaves as an apoptosis inducer and in-creases the expression of Bax [40]. Interestingly, our resultsshow a higher number of cells immunostaining with Bak thanBcl-2 after 48 h of infection, indicating a loss of balancebetween death and survival signals. Thus, our results may
suggest that P. brasiliensis remains alive inside the cell, longenough to generate budding cells. Preliminary evidence sug-gests that P. brasiliensis cells more frequently sporulatewhen free within the cytoplasm of the host cell, but less sowhen surrounded in vacuoles (Mendes-Giannini et al., un-published data), and that these unrestrained cytoplasmic fun-gal cells may escape from the host cells. Thus, induction ofapoptosis in the epithelial cells or phagocytes could be ben-eficial to the pathogen, assisting it to evade ingestion and,consequently, favoring further dissemination. Opposing thisview, in chronic granulomatous diseases caused by mycobac-teria, apoptosis of the parasitized host cells has been claimedto be a host protective mechanism [41]. This illustrates thatthe same host response may result in different outcomes ofthe host–parasite interaction. Differently from human patho-genic mycobacterium, P. brasiliensis is a facultative intrac-ellular pathogen, [7] and thus release from the intracellularcompartment may favor dissemination, while in mycobacte-riosis, the bacteria remain within the apoptotic bodies and aresubsequently cleared out [41]. In conclusion, the mecha-nisms of invasion of host cells, persistence within them, andthe subsequent induction of apoptosis of such cells mayexplain the efficient dissemination of P. brasiliensis.
Acknowledgments
This investigation was financially supported by the Brazil-ian Organizations, FAPESP, Conselho Nacional de Desen-volvimento Científico e Tecnológico (CNPq) and PADC-FCF-UNESP.
References
[1] M. Franco, R.P. Mendes, M. Moscardi-Bacchi, M.T. Rezkallah-Iwasso, M.R. Montenegro, Paracoccidioidomycosis, Baillere’s Clin,Trop. Med. Commun. Dis. 4 (1989) 185–196.
[2] M.I. Borges-Walmsley, D. Chen, X. Shu, A.R. Walmsley, The patho-biology of Paracoccidioides brasiliensis, Trends Microbiol. 10(2002) 80–87.
[3] M.J.S. Mendes Giannini, M.L. Taylor, J.B. Bouchara, E. Burger,V.L.G. Calich, E.D. Escalante, S.A. Hanna, H.L. Lenzi,M.P. Machado, M. Miyaji, J.L. Monteiro da Silva, E.M. Mota,A. Restrepo, S. Restrepo, G. Tronchin, L.R. Vincenzi, C.F. Xidieh,E. Zenteno, Pathogenesis II: fungal responses to host responses:interaction of host cells with fungi, Med. Mycol. 38 (2000) 113–123.
[4] M.J.S. Mendes-Giannini, L.C. Ricci, M. Uemura, E. Toscano,C.W. Arns, Infection and apparent invasion of Vero cells by Paracoc-cidioides brasiliensis, J. Med. Vet. Mycol. 32 (1994) 189–195.
[5] S.A. Hanna, J.L. Monteiro da Silva, M.J.S. Mendes-Giannini, Adher-ence and intracellular parasitism of Paracoccidioides brasiliensis inVero cells, Microb. Infect. 2 (2000) 877–884.
[6] T. Brito, J.S. Furtado, R.M. Castro, M. Manini, Intraepithelial parasit-ism as an infection mechanism in human paracoccidioidomycosis,Vir. Arch. Pathol. Anat. 361 (1973) 129–138.
[7] R.M. Tuder, R. El Ibrahim, C.E. Godoy, T. De Brito, Pathology of thehuman pulmonary paracoccidioidomycosis, Mycopathologia 92(1985) 179–188.
890 M.J. Soares Mendes-Giannini et al. / Microbes and Infection 6 (2004) 882–891

[8] P. Clerc, P.J. Sansonetti, Entry of Shigella flexneri into Hela cells:evidence for directed phagocytosis involving actin polymerizationand myosin accumulation, Infect. Immun. 55 (1987) 2681–2688.
[9] J.D. Clausen, G. Christiansen, H.U. Holst, S. Birkelund, Chlamydiatrachomatis utilizes the host cell microtubule network during earlyevents of infection, Mol. Microbiol. 25 (1997) 441–449.
[10] S. Dramsi, P. Cossart, Intracellular pathogens and the actin cytoskel-eton, Annu. Rev. Cell. Dev. Biol. 14 (1998) 137–166.
[11] B.B. Finlay, S. Falkow, Common themes in microbial pathogenicity,Microbiol. Rev. 53 (1989) 210–230.
[12] A. Zychlinsky, P.J. Sansonetti, Apoptosis in bacterial pathogenesis, J.Clin. Invest. 100 (1997) 63–65.
[13] C.R. Cacere, C.C. Romano, M.J.S. Mendes-Giannini, A.J.S. Duarte,G. Benard, The role of apoptosis in the antigen-specific T cell hypo-responsiveness of paracoccidioidomycosis patients, Clin. Immunol.105 (2002) 215–222.
[14] M.H.S. Blotta, Z.P. Camargo, Immunological response to cell-freeantigens of Paracoccidioides brasiliensis: relationship with clinicalforms, J. Clin. Microbiol. 31 (1993) 671–676.
[15] C.L. Wells, E.M.A. Vande Westerlo, R.P. Jechorek, H.M. Haines,S.L. Erlandsen, Cytochalasin induced actin disruption of polarizedenterocytes can augment internalization of bacteria, Infect. Immun. 66(1998) 2410–2419.
[16] M.J.S. Mendes-Giannini, J.P. Bueno, M.A. Shikanai-Yasuda,A.W. Ferreira, A. Masuda, Detection of the 43 000 molecular weightglycoprotein in sera of patients with paracoccidioidomycosis, J. Clin.Microbiol. 27 (1989) 2842–2845.
[17] H. Towbin, T. Stachelin, J. Gordon, Eletrophoretic transfer of proteinsfrom poliacrylamide gels to nitrocellulose sheets. Procedure and someapplications, Proc. Natl. Acad. Sci. 76 (1979) 4350–4352.
[18] A. Zychlinsky, K. Thirumalai, J. Arondel, R.L. Cantey, A.O. Alipran-tis, P. Sansonetti, In vitro apoptosis in Shigella flexneri infections,Infect. Immun. 64 (1997) 5357–5365.
[19] K. Kobayashi, C. Terada, I. Tsukamoto, Methotrexate-induced apop-tosis in hepatocytes after partial hepatectomy, Eur. J. Pharmacol. 438(2002) 19–24.
[20] M.B. Goldberg, P.J. Sansonetti, Shigella subversion of the cellularcytoskeleton: a strategy for epithelial colonization, Infect. Immun. 61(1993) 4941–4946.
[21] I. Rosenshine, S. Ruschkowski, B.B. Finlay, Inhibitors of cytoskeletalfunction and signal transduction to study bacterial invasion, MethodsEnzymol. 263 (1994) 467–476.
[22] R.D. Hayward, V. Koronabis, Direct nucleation and bundling of actinby the SipC protein of invasive Salmonella, EMBO J. 18 (1999)4926–4934.
[23] I. Tsarfaty, H. Sandovsky-Losica, L. Mittelman, I. Berdicevsky,E. Segal, Cellular actin is affected by interaction with Candida albi-cans, FEMS Microbiol. Lett. 189 (2000) 225–232.
[24] J.A. Wasylnka, M.M. Moore, Uptake of Aspergillus fumigatus conidiaby phagocytic and nonphagocytic cells in vitro: quantification usingstrain expressing green fluorescent protein, Infect. Immun. 70 (2002)3156–3163.
[25] T.S. Sebghati, J.T. Engle, W.E. Goldman, Intracellular parasitism byHistoplasma capsulatum: fungal virulence and calcium dependence,Science 290 (2000) 1368–1372.
[26] J.L. Monteiro da Silva, M.J.S. Mendes-Giannini, M.P. Sandoval,H.L. Lenzi, Adesão e invasão de Paracoccidioides brasiliensis emcultura de células Vero, Rev. Cienc. Farm. 22 (2001) 103–129.
[27] U.S. Sajjan, F.A. Sylvester, J.F. Forstner, Cable-pilited Burkholderiacepacia binds to cytokeratin 13 of epithelial cells, Infect. Immun. 68(2000) 1787–1795.
[28] J.A. Cooper, Effects of cytochalasin and phaloidin on actin, J. Cell.Biol. 105 (1987) 173–178.
[29] H. Sandovsky-Losica, I. Berdicevsky, I. Tsarfaty, E. Segal, Effect ofCandida albicans metabolite(s) on cellular actin, FEMS Microbiol.Lett. 215 (2002) 57–62.
[30] B.U. Stambuck, R. Puccia, M.L.C. Almeida, L.R. Travassos,S. Schenckman, Secretion of the 43 kDa glycoprotein antigen byParacoccidioides brasiliensis, J. Med. Vet. Mycol. 26 (1988) 367.
[31] J.W. Deacon, Modern Mycology, third ed, Blackwell Science, Lon-don, 1997.
[32] M.J.S. Mendes-Giannini, R.A. Moraes, T.A. Ricci, Proteolytic activ-ity of the 43 000 molecular weight antigen secreted by Paracoccidio-ides brasiliensis, Rev. Inst. Med. Trop. 32 (1990) 385–387.
[33] R. Puccia, A.K. Carmona, J.L. Gesztezi, L. Juliano, L.R. Travassos,Exocellular proteolytic activity of Paracoccidioides brasiliensis:cleavage of components associated with the basement membrane,Med. Mycol. 36 (1998) 354–358.
[34] J. Doorbar, S. Ely, J. Sterling, C. Mclean, L. Crawford, Specificinteraction between HPV-16 E1-E4 and cytokeratins result in collapseof epithelial cell intermediate filament network, Nature 352 (1991)824–827.
[35] M. Stewart, Intermediate filament structure and assembly, Curr. Opin.Cell. Biol. 5 (1993) 3–11.
[36] D.L. Vaux, S.J. Korsmeyer, Cell death in development, Cell 96 (1999)245–254.
[37] D. Rotstein, J. Parodo, R. Taneja, J.C. Marshall, Phagocytosis ofCandida albicans induces apoptosis of human neutrophils, Shock 14(2000) 278–283.
[38] B. Levine, Q. Huang, J.T. Isaacs, J.C. Reed, D.E. Griffin, J.M. Hard-wick, Conversion of lytic to persistent alphavirus infection by the bcl2cellular oncogene, Nature 361 (1993) 739–742.
[39] A.J. Mastrangelo, M.J. Hardwick, F. Bex, M.J. Betenbaugh, Bcl-2 andBcl-X limit apoptosis upon infection with alphavirus vectors, Biotech.Bioeng. 67 (2000) 544–554.
[40] E. Appel, A. Katzorf, T. Ben-Moshe, G. Kazimirsky, D. Kobiler,S. Lustig, C. Brodie, Differential regulation of Bcl-2 and Bax expres-sion in cells infected with virulent and nonvirulent strains of Sindbisvirus, Virology 276 (2000) 238–242.
[41] C. Fratazzi, R.D. Arbeit, C. Carini, M.K. Balcewicz-Sablinska,J. Keane, H. Kornfeld, H.G. Remold, Macrophage apoptosis in myco-bacterial infections, J. Leukocyte Biol. 66 (1999) 763–764.
891M.J. Soares Mendes-Giannini et al. / Microbes and Infection 6 (2004) 882–891





























