Embed Size (px)
Citation preview

lable at ScienceDirect
European Journal of Medicinal Chemistry 73 (2014) 126e134
Contents lists avai
European Journal of Medicinal Chemistry
journal homepage: http: / /www.elsevier .com/locate/ejmech
Original article
Insights on pregnane-X-receptor modulation. Natural andsemisynthetic steroids from Theonella marine sponges
Valentina Sepe a,*, Claudio D’Amore b, Raffella Ummarino a, Barbara Renga b,Maria Valeria D’Auria a, Ettore Novellino a, Annamaria Sinisi a, Orazio Taglialatela-Scafati a,Yoichi Nakao c, Vittorio Limongelli a, Angela Zampella a, Stefano Fiorucci b
aDipartimento di Farmacia, Università di Napoli “Federico II”, 80131 Napoli, ItalybDipartimento di Medicina Clinica e Sperimentale, Università degli Studi di Perugia, 06132 Perugia, ItalycDepartment of Chemistry and Biochemistry, Waseda University, 3-4-1 Okubo, Shinjuku-ku, Tokyo 169-8555, Japan
a r t i c l e i n f o
Article history:Received 2 August 2013Received in revised form28 November 2013Accepted 2 December 2013Available online 15 December 2013
Keywords:Theonella marine sponges4-MethylenesteroidsConicasterolPregnane-X-receptorStructureeactivity relationshipMolecular docking
Abbreviations: ABC transporters, ATP-binding cassvation function 2; COSY, correlation spectroscopy; CYPDBD, DNA binding domain; FXR, farnesoid-X-receptoferases; HepG2, human hepatoma cell line; HMBC, hHPLC, high performance liquid chromatography; HRtrospray ionization mass spectrometry; HSQC, hecoherence; LBD, ligand binding domain; MPLC, medtography; NMR, nuclear magnetic resonance; NRs,idinium chlorochromate; PDB, Protein Data Bank; PPCR, real time polymerase chain reaction; SULTs, sulfchromatography; UGTs, UDP-glucuronyltransferases.* Corresponding author. Tel.: þ39 081 678526; fax:
E-mail address: [email protected] (V. Sepe).
0223-5234/$ e see front matter � 2013 Elsevier Mashttp://dx.doi.org/10.1016/j.ejmech.2013.12.005
a b s t r a c t
Pregnane-X-receptor (PXR) is a member of nuclear receptors superfamily that activates gene tran-scription by binding to responsive elements in the promoter of target genes. PXR is a master geneorchestrating the expression/activity of genes involved in the metabolism of endobiotics includingbilirubin, bile acids, glucose and lipid. In addition PXR oversights the metabolism of the large majority ofxenobiotics including a large amount of prescribing drugs. Thus, developing PXR ligands represents agreat opportunity for a therapeutic intervention on human diseases including diabetes, obesity, dysli-pidemias and liver disorders. To this end, natural compounds represent an arsenal of new chemicalscaffolds useful for the identification of novel PXR ligands. Here, we report a series of 4-methylenesteroidderivatives isolated from Theonella marine sponges as novel PXR modulators. In addition, combiningmedicinal chemistry, pharmacological experiments and computational studies, we have investigated theeffects of different modifications on ring A and on the side chain of 4-methylenesteroid derivatives to-ward PXR modulation. This study provides the molecular bases of ligand/PXR interaction useful fordesigning novel PXR modulators.
� 2013 Elsevier Masson SAS. All rights reserved.
1. Introduction
Pregnane-X-receptor (PXR) is a member of nuclear receptors(NRs) super-family. PXR orchestrates the complex and intricatenetwork of xenobiotic metabolism. After ligand binding, PXRtranslocates to nucleus and binds to its promoter responsive ele-ments as obligatory heterodimer with retinoid-X-receptor. PXRshares with other NRs several structural features such as a highly
ette transporters; AF-2, acti-s, cytochrome P superfamily;r; GSTs, glutathione-S-trans-etero-multi bond correlation;ESIMS, high-resolution elec-teronuclear single-quantumium pressure liquid chroma-nuclear receptors; PCC, pyr-XR, pregnane-X-receptor; RTotransferases; TLC, thin-layer
þ39 081 678552.
son SAS. All rights reserved.
conserved N-terminal DNA-binding domain (DBD), a lessconserved ligand binding domain (LBD), and one transactivationdomain, the activation function 2 (AF-2). The key difference withthe other NRs lies in the unique conformational flexibility and thelarge volume of the LBD that is able to accommodate an extraor-dinary variety of exogenous compounds [1]. Consequently, in theliver and in the gastro-intestinal tract, PXR regulates the expressionof genes for Phase I and II drug metabolizing enzymes, includingCYPs, carboxylesterases, alcohol and aldehyde dehydrogenases,glucuronyltransferases (UGTs), sulfotransferases (SULTs) andglutathione transferases (GSTs) [2], and numerous efflux trans-porters [3] (ABC drug efflux transporters, breast cancer resistanceprotein, multidrug resistance-associated proteins and P-glycoprotein).
Apart from the afore mentioned pivotal role in regulating drugdetoxification, PXR plays also a pleiotropic role in endobioticmetabolism. PXR regulates bile acids homeostasis [4,5], glucose andlipid metabolism [6,7], energy homeostasis, immune responses [8]and, notably, drugedrug interactions [9]. As a result, PXR ligands(agonists and antagonists) are recognized opportunity for thepharmacotherapy of several human diseases including diabetes,

Fig. 1. Conicasterol and theonellasterol as biomarkers of Theonella conica and Theonellaswinhoei, respectively.
V. Sepe et al. / European Journal of Medicinal Chemistry 73 (2014) 126e134 127
obesity, dyslipidemia, liver disorders [10], immune mediate dys-functions and cancer. Expectedly, in the last decades a considerablenumber of chemicals have been show to function as PXR ligands,including widely prescribed drugs such as statins, antibiotics,anticancer compounds as well as environmental toxicants, plasti-cizers and pesticides. In addition, several natural compounds iso-lated from herbal medicines and marine organisms havehighlighted the potential of novel chemical scaffolds useful for thedevelopment of PXR ligands [6,11]. In this context and in the frameof our interest in the discovery of nuclear receptor modulators frommarine sources [12e17], we have described several compoundsisolated from marine sponges endowed with agonistic activity onPXR including solomonsterols, side chain truncated steroids [18],and the large family of 4-methylenesteroids from Theonella swin-hoei [19e22]. Within these new chemotypes of PXR agonists,solomonsterol A was effective in reducing the development ofclinical signs and symptoms of colitis in a PXR dependent mecha-nism [23] and therefore represents the first example of marine leadin the treatment of inflammatory bowel disease. The rare 4-methylenesteroids represent the exclusive elements of the steroidbiogenetic class in Theonella sponges with conicasterol (1) andtheonellasterol (2) (Fig. 1) as ideal biomarkers of Theonella conicaand T. swinhoei, respectively [24].
These molecules share substituents and unsaturations on thetetracyclic core but differ in the side chainwith a 24S-ethyl group intheonellasterol and a 24R-methyl group in conicasterol. Recentlywedemonstrated that 4-methylenesteroids are endowedwith peculiarpharmacological profiles, ranging fromselective antagonismon FXR[25]of theonellasterol (2) todualmodulationonFXR/PXR [19e22]ofothers 4-methylenesteroids, thus affirming Theonella sponges genusas an invaluable source of nuclear receptor ligands [12].
In this paper we report the results of chemical analysis onthe apolar extracts from a Solomon collection of T. swinhoei andfrom a T. conica specimen collected off Kakeroma Island,Kagoshima pref., leading to the isolation of three new side chainmodified 4-methylenesteroids (Fig. 2), endowed with interestingactivity on PXR. Moreover the isolation of large amount of theparent compound conicasterol (1), obtained from the Japanesecollection of T. conica, allowed to speculate on the effect ofpunctual modifications on ring A and on the side chain towardPXR modulation and notably, to identify new chemotype of PXRmodulators.
Fig. 2. New side chain modified 4-methyle
2. Results and discussion
2.1. Structural characterization
Preconicasterol (3) was isolated as an amorphous solid withmolecular formula C28H44O by HR-ESIMS. This compound waseasily identified as a member of the class of 4-methylenesteroids,however its molecular formula suggested that it should not containany branching on the side chain. Analysis of 1D NMR data (Table 1),guided by inspection of COSY, HSQC and HMBC spectra, allowed thecomplete assignment of 1H and 13C NMR resonances. All the 1H/13Cresonances of the tetracyclic core were practically identical to thoseof 1 and 2, while the side chain was demonstrated to include a D24
(dH 5.28) double bond, as indicated also by the downfield shift ofMe-26 and Me-27 (dH 1.62 and 1.72, respectively).
To our knowledge, compound 3 represents the first example ofTheonella 4-methylenesteroids to lack any ramification in the sidechain. On the basis of the current knowledge about the biogenesis ofbranched side chain sterols, preconicasterol (3) should be a likelybiogenetic precursor of the branched derivatives 1 and 2 as shownin Fig. 3.
As for compound 3, the structural elucidation of compounds 4and 5 was also greatly helped by comparison of spectroscopic datawith those of known analogs. Indeed, both 4 and 5 showed NMRsignals of the tetracyclic core superimposable to those of alreadypublished derivatives [19e21,24].
In particular, 24-dehydroconicasterol D (4) shares the ring sys-tem and the two additional hydroxyl groups at C-9 and C-14 withconicasterol D [19], while 25-dehydrotheonellasterol (5) proved topossess the same ring system of compounds 1e3.
COSY analysis of the 1H NMR spectrum of 24-dehydroconicasterol D (4) (Table 1), C29H46O3 by HR-ESIMS, indicated that itsside chain should include an sp2 methylene, three methyl groups(all doublets at dH 0.99, 1.09 and 1.10), two sp3methines and two sp3
methylenes. The HMBC correlations of the sp2 methylene with C-23, C-24 and C-25 indicated its attachment at C-24, thus definingthe structure of compound 4 as a new sterol differing from con-icasterol D by the presence of an additional double bond betweenC-24 and C-28.
The 1H NMR spectrum of compound 5 (Table 1), C30H48O by HR-ESIMS, revealed that its side chain should include an ethyl group(dH 1.35, m and 0.90, t), an sp2 methylene (dH 4.83 and 4.89, br s),two sp3 methylenes, two sp3methines and two methyl groups (dH1.02, d; 1.58, s).
The COSY spectrum of 5 organized all the multiplets of the sidechain within a single spin system starting from Me-21 and termi-nating with Me-29, further connected to ring D spin systemthrough H-20. The HMBC cross-peaks of H2-26 with C-24, C-25 andC-27 completely defined the structure of compound 5 as 25-dehydrotheonellasterol.
On the basis of NMR data, the configuration at C-24 has beenassumed the same as that of the biogenetically related theo-nellasterol and invariably found for 24-ethyl steroids fromTheonella.
nesteroids from Theonella collections.

Table 11H and 13C NMR data (700 MHz, C6D6) for preconicasterol (3), 24-dehydroconicasterol D (4) and 25-dehydrotheonellasterol (5).
Pos. 3 4 5
dHa dC dH
a dC dHa dC
1 1.05 ovlb
1.54 m37.2 1.35 m
2.07 m30.1 1.06 ovl
1.55 ovl37.2
2 1.32 ovl1.83 m
33.8 1.37 m1.90 m
33.4 1.32 m1.83 m
33.8
3 3.81 m 73.5 3.88 m 73.0 3.82 dd(4.8, 11.4)
73.3
4 e 153.8 e 154.2 e 153.85 1.64 ovl 49.7 2.70 br d
(12.51)41.7 1.64 m 50.0
6 1.39 m1.86 m
27.4 1.39 m1.59 m
24.8 1.39 m1.86 m
27.8
7 1.73 m2.47 ddd(2.2, 4.3, 13.7)
29.8 2.37 ddd(1.8, 4.7, 14.6)2.54 m
27.1 1.75 m2.47 ddd(2.1,4.2,13.8)
30.1
8 e 126.6 e 135.3 e 126.69 1.69 ovl 49.5 e 74.9 1.69 m 49.910 e 40.4 e 43.8 e 40.311 1.43 ovl
1.55 m20.9 1.55 m
1.81 m29.2 1.44 ovl
1.56 m21.8
12 1.20 ovl1.98 ovl
37.9 1.41 m1.76 m
34.4 1.21 ovl1.99 m
38.1
13 e 43.5 e 44.2 e 43.514 e 143.0 e 150.3 e 142.015 1.56 ovl
1.38 m25.0 4.47 br d 69.6 1.39 ovl
1.56 ovl25.3
16 2.26 m 26.2 1.54 m1.82 m
39.4 2.27 m 26.4
17 1.19 ovl 57.4 1.62 m 53.7 1.18 m 57.418 0.91 s 18.5 0.71 s 18.3 0.93 s 18.819 0.62 s 13.4 0.68 s 17.0 0.63 s 13.720 1.56 ovl 34.7 1.50 ovl 34.9 1.53 m 34.921 1.04 d (6.6) 19.3 0.99 d (6.7) 19.2 1.02 d (6.8) 19.622 1.23 m
1.59 ovl36.6 1.69 ovl
1.34 ovl33.8 1.17 ovl
1.43 ovl34.0
23 2.02 m2.16 ovl
25.3 2.01 m2.24 m
30.6 1.28 ovl1.42
30.1
24 5.28 t (6.9) 126.3 e 39.2 1.92 ovl 50.325 e 131.1 2.27 hep (6.8) 32.7 e 147.826 1.62 s 17.7 1.09 (6.8) 18.4 4.83 br s
4.89 br s112.0
27 1.72 s 26.0 1.10 (6.8) 20.3 1.58 s 18.328 4.71 br s
5.30 br s103.7 4.90 br s
4.93 br s105.8 1.35 m 27.2
29 4.74 br s5.35 br s
103.7 0.90 t (7.4) 12.8
30 4.71 br s5.30 br s
103.8
a Coupling constants are in parentheses and given in Hertz. 1H and 13C assign-ments aided by COSY, HSQC and HMBC experiments.
b Ovl: overlapped with other signals.
V. Sepe et al. / European Journal of Medicinal Chemistry 73 (2014) 126e134128
2.2. Chemical modification on conicasterol scaffold
As previously reported [24], conicasterol (1) is the majorcomponent of the apolar extract of T. conica. Even if conicasterol, aswell as several derivatives, was proved to be a potent PXR agonist[21], conicasterol was observed to be cytotoxic toward HepG2 cellswhen tested at 50 mM in combination with rifaximin. Nevertheless,we reasoned that the large amount available in our laboratory aswell as the presence on its chemical structure of functional groupsthat could be easily modified make conicasterol a suitable templateto provide insights on the key structural motifs responsible of PXRmodulation. In particular, conicasterol (1) possesses rare structuralfeatures, somewhat difficult to obtain through synthetic routes,especially the exomethylene functionality at C-4. Thus, a simplifi-cation in this part would be particularly precious in the perspectiveof a total synthesis of PXR modulators inspired to conicasterolscaffold and with a better safety profile. As depicted in Scheme 1,
and already reported for theonellasterol [26], hydrogenation ofexocyclic double-bond on conicasterol ring A produced exclusivelythe 4b-methyl derivative 6, as a result of the steric influence playedby b Me-19 in orienting the access of H2 molecule to the doublebond. Structural and stereochemical characterization was based oncareful analysis of 1H NMR spectrum. Up-field shift exhibited by H-3 (from 3.82 in 1 to 3.53 in 6) and the presence of an additionalmethyl doublet at dH 0.93 clearly suggested the presence of anadditional methyl group at C-4. Small coupling constant H-3/H-4(3.2 Hz) was consistent with their cis relationship and thereforewith the b-orientation of methyl group at position-4.
On the other hand, double bond reduction on conicasteroneintermediate [27], obtained through PCC oxidation of the oxy-methine function at C-3, afforded a mixture of diasteroisomericketone derivatives differing in the orientation of the methyl groupat C-4. Sodium borohydride treatment on the crude mixture, fol-lowed by HPLC separation, furnished again alcohol 6 along with thediastereoisomers 7 and 8, with different stereochemical arrange-ments of the substituents at C-3 and C-4.
The stereochemical assignment reported in Scheme 1 wassubstantiated by the almost complete superimposition of protonresonances of all nuclei belonging to the tetracyclic nucleus withthose reported for the corresponding derivatives of theonellasterol[26].
Pursuing on ring A modification, also the effect of the intro-duction of an additional polar groupwas investigated preparing the4-ketone derivative 9 through ozonolysis of conicasterol (1) fol-lowed by dimethylsulfide work-up (O3, CH2Cl2, �78 �C, 5 min, thenMeOH and Me2S in excess).
Finally the role of the stereochemistry at C-24 was also exam-ined by treatment of a dehydroconicasterol sample [21] withhydrogen in presence of platinum oxide as catalyst (Scheme 2).
The reaction proceeded smoothly through the concomitantreduction of the exomethylene at C-4 and C-24 affording derivative6 and 10, efficiently separated on a C-18 ISIS HPLC column. Havingassigned the 24R-methyl side chain in 6, careful comparison of 1HNMR spectra of derivatives 6 and 10, differing exclusively in theregion of methyl resonances (0.8e1.1 ppm), clearly inferred 10 asthe C-24 epimer of 6.
2.3. Pharmacological evaluation and structureeactivityrelationships
Natural compounds 3e5 and semi-synthetic conicasterol ana-logs 6e10were evaluated on PXR in a luciferase reporter assay on ahuman hepatocyte cell line (HepG2 cells) transiently transfectedwith pSG5-PXR, pSG5-RXR, pCMV-bgalactosidase, and p(CYP3A4)-TK-Luc vectors (Fig. 4).
HepG2 cells were stimulated with compounds 3e10 in thepresence or in absence of rifaximin (10 mM), a well characterizedPXR agonist. As shown in Fig. 4A, compounds 3 and 8, whenadministered alone, transactivate PXR with a potency comparablewith that of rifaximin. Even if partial inhibitions of PXR trans-activation caused by rifaximin are shown in Fig. 4B, none com-pound of this series could be judged an effective antagonist. Indetails, when cells were co-exposed to compounds 3 and 8 andrifaximin, the two compounds caused a slightly reversion of thepharmacological effect of rifaximin. This finding is commonwhen afull agonist (rifaximin) is mixed with partial (less potent) agonistssuch as compounds 3 and 8. As concern compound 6, even if aslight antagonistic effect was demonstrated at 50 mM, this behaviorwas not confirmed when 6 was co-administered with rifaximin atdifferent concentration [28].
To further investigate the structureeactivity relationship, wefirst performed the substitution of the exomethylene at C-4 with a
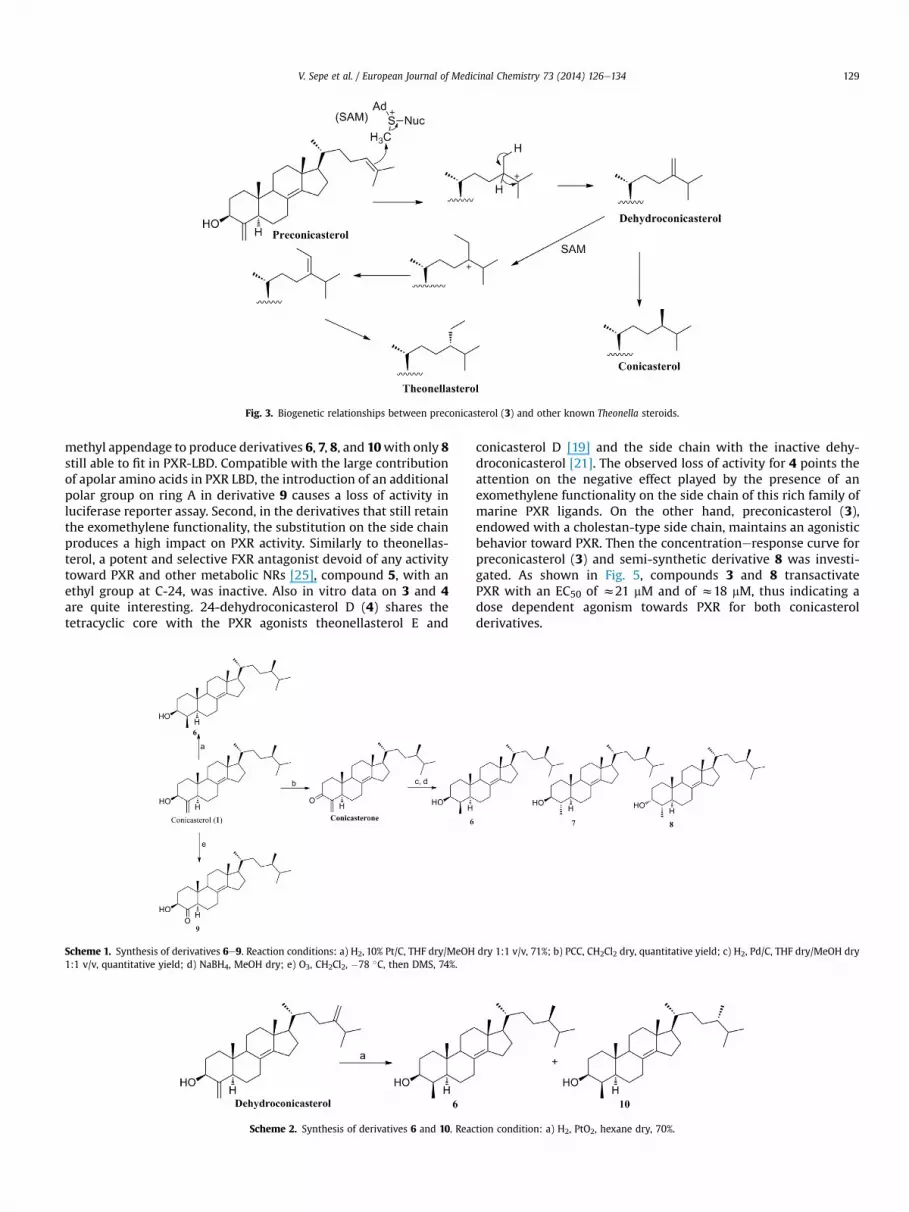
Fig. 3. Biogenetic relationships between preconicasterol (3) and other known Theonella steroids.
V. Sepe et al. / European Journal of Medicinal Chemistry 73 (2014) 126e134 129
methyl appendage to produce derivatives 6, 7, 8, and 10with only 8still able to fit in PXR-LBD. Compatible with the large contributionof apolar amino acids in PXR LBD, the introduction of an additionalpolar group on ring A in derivative 9 causes a loss of activity inluciferase reporter assay. Second, in the derivatives that still retainthe exomethylene functionality, the substitution on the side chainproduces a high impact on PXR activity. Similarly to theonellas-terol, a potent and selective FXR antagonist devoid of any activitytoward PXR and other metabolic NRs [25], compound 5, with anethyl group at C-24, was inactive. Also in vitro data on 3 and 4are quite interesting. 24-dehydroconicasterol D (4) shares thetetracyclic core with the PXR agonists theonellasterol E and
Scheme 1. Synthesis of derivatives 6e9. Reaction conditions: a) H2, 10% Pt/C, THF dry/MeOH1:1 v/v, quantitative yield; d) NaBH4, MeOH dry; e) O3, CH2Cl2, �78 �C, then DMS, 74%.
Scheme 2. Synthesis of derivatives 6 and 10. Reac
conicasterol D [19] and the side chain with the inactive dehy-droconicasterol [21]. The observed loss of activity for 4 points theattention on the negative effect played by the presence of anexomethylene functionality on the side chain of this rich family ofmarine PXR ligands. On the other hand, preconicasterol (3),endowed with a cholestan-type side chain, maintains an agonisticbehavior toward PXR. Then the concentrationeresponse curve forpreconicasterol (3) and semi-synthetic derivative 8 was investi-gated. As shown in Fig. 5, compounds 3 and 8 transactivatePXR with an EC50 of z21 mM and of z18 mM, thus indicating adose dependent agonism towards PXR for both conicasterolderivatives.
dry 1:1 v/v, 71%; b) PCC, CH2Cl2 dry, quantitative yield; c) H2, Pd/C, THF dry/MeOH dry
tion condition: a) H2, PtO2, hexane dry, 70%.

Fig. 4. Transactivation assay performed on HepG2 cells transiently transfected with full-length PXR, RXR, bgal and the canonical PXRE containing 3 inverted repeats (IR1). At 24 hpost-transfection, cells were primed with (A) rifaximin (R) and compounds 3e10 (10 mM) or with (B) compounds 3e10 (50 mM) plus rifaximin (10 mM) for 18 h (*: p < 0.05 vs nottreated cells; #: p < 0.05 vs rifaximin).
Fig. 5. Concentration response-curves for 3 and 8. HepG2 cells were transfected for PXR transactivation assay and stimulated with rising concentrations of 3 and 8 (1, 10 and 50 mM).Rifaximin (1, 10 and 50 mM) was used as a positive control to evaluate the PXR transactivation. RLU: Luciferase Relative light Units, RRU: Renilla Relative light Units. Results areexpressed as mean � standard error; *p < 0.05 vs not treated cells.
V. Sepe et al. / European Journal of Medicinal Chemistry 73 (2014) 126e134130
To further characterize these derivatives, the effect of the twomodulators, 3 and 8, on the expression of CyP3A4 (cytochrome P)and on Multidrug Resistance Proteins (MDR1 and MPR3), threecanonical PXR target genes, was examined by RT-PCR. As shown in
Fig. 6. Real-Time PCR analysis of mRNA relative expression of the PXR target genes CYP3A4,normalized relatively to GAPDH mRNA and are expressed relative to those of not treated c
Fig. 6, the exposure of HepG2 cells to 3 and 8 increased theexpression of mRNA for the above PXR targeted genes, therebyconfirming at molecular level that these compounds are PXRagonists.
MDR1, MRP3 in HepG2 cells treated with rifaximin (10 mM), 3 and 8 (10 mM). Values areells (NT), which are arbitrarily set to 1. (*: p < 0.05 vs not treated cells).

Fig. 7. Representation of the binding mode of the agonists, preconicasterol (3) (violet sticks in A) and derivative 8 (cyan sticks in B), predicted by docking calculations in the PXR LBD(PBD code 3HVL). PXR is shown as green cartoon, while AF-2 helix is colored in orange. Amino acids involved in ligand binding are shown as green and orange sticks. Residues fromPro268 to Arg287, from Ser350 to Arg360, and all hydrogens are omitted for clarity. (For interpretation of the references to color in this figure legend, the reader is referred to theweb version of this article.)
V. Sepe et al. / European Journal of Medicinal Chemistry 73 (2014) 126e134 131
2.4. Molecular docking studies
To elucidate at atomic level the interaction between PXR and thederivatives generated in this study, docking calculations on themost active compounds of the series have been performed. Inparticular, three different X-ray structures of the PXR LBD (pdbcodes: 3hvl, 1nrl and 1m13) have been used to take into account theconformational plasticity of the binding site. In fact, docking soft-ware treats protein as a rigid body, thus the use of multiple con-formations of the target, if available, is advisable to increase thechances of success of the calculation. In the present case, theagonist compounds, preconicasterol (3) and derivative 8, wereundergone to docking calculations that predicted a very similarbinding behavior for both agonists in PXR-LBD. In particular,considering the docking results obtained using the three LBD X-raystructures, the most occurring binding mode of preconicasterol (3)and derivative 8 is that shown in Fig. 7.
In this state, the hydroxyl group on the ring A of preconicasterol(3) and derivative 8 engages H-bond interactions with the back-bone carbonyl oxygens of His407 and Thr408. These interactionsrepresent the anchor point of the ligands in the LBDwith the rest ofthe molecule oriented in the binding pocket forming furtherfavorable interactions such as the hydrophobic contacts betweenthe Me-19 and the ring A with Met243, Phe251, Met425 andPhe429. In particular, the last two residues are located in the acti-vation function 2 region (AF-2) on a small flexible alpha helix (AF-2helix) responsible for the binding of the co-activator and the co-repressor peptides. This part of the protein is supposed to un-dergo conformational motion responsible for the transition of thenuclear receptor from the agonist to the antagonist form. A similarmechanism of action has been very recently suggested by us andother authors for other nuclear receptors [13,29e31]. On thisbackground, it is reasonable to think that the hydrophobic packageestablished at the AF-2 region by agonist compounds, pre-conicasterol (3) and derivative 8, stabilizes PXR in the agonistconformation.
Finally, in the LBD, the steroidal scaffold of 3 and 8 engagessimilar interactions with Leu209, while the flexible side chains aresettled in a very hydrophobic pocket. Here, preconicasterol (3) andderivative 8 establish a number of favorable contacts with residuessuch as Phe288, Trp299, Tyr306 and Met246 (Fig. 7A and B).
Of relevance, despite the different stereochemistry at C-3 onring A and the presence of a methylene group in preconicasterol (3)
instead of the methyl one in derivative 8, the binding pattern ofthese compounds is very similar, supporting the reproducibilityand the reliability of the docking calculations.
3. Conclusion
NRs play a key role in cellular homeostasis, controlling a widerange of reactions through the regulation of the transcription ofspecific target genes. This receptor family shares common struc-tural features such as the presence of the LBD responsible for thebinding of ligands and co-activator peptides necessary to activatethe gene transcription. Despite these similarities, each nuclear re-ceptor shows a specific pattern of interaction with its ligands that,togetherwith the conformational plasticity of the LBD, hampers therational drug design of new active molecules. In this scenario,elucidating the molecular requisites for the binding to a specificnuclear receptor represents an important advance to understand itsfunctional mechanism and to provide the bases for an exogenouscontrol of its activity.
In this framework, we have developed a series of natural andsemisynthetic steroids from Theonella marine sponges, as novelmodulators of the nuclear receptor PXR. We have assessed theiractivity profiles through a number of pharmacological experimentsand, with the aid of computational studies, we have investigatedthe effects of differentmodifications on ring A and on the side chainof the 4-methylenesteroid derivatives toward PXR. This studyreaffirms the role of natural products as essential chemical probesin the today’s research arsenal to shed light on complex biologicalprocesses and biochemical pathways.
4. Experimental section
4.1. General experimental procedures
Specific rotations were measured on a PerkineElmer 243 Bpolarimeter. High-resolution ESI-MS spectra were performed witha Micromass QTOF spectrometer. ESI-MS experiments were per-formed on an Applied Biosystem API 2000 triple-quadrupole massspectrometer. NMR spectra on preconicasterol (3), 24-dehydroconicasterol D (4) and 25-dehydrotheonellasterol (5)were obtained on Varian Inova 700 NMR spectrometer (1H at700 MHz, 13C at 175 MHz, respectively) equipped with a Sun

V. Sepe et al. / European Journal of Medicinal Chemistry 73 (2014) 126e134132
hardware, d (ppm), J in Hz, spectra referred to C6HD5 as internalstandards (dH 7.16, dC 128.4).
NMR spectra on all synthetic intermediates (derivatives 6e10)were obtained on Varian Inova 400 NMR spectrometer (1H at400 MHz, 13C at 100 MHz, respectively) and recorded in C6HD5 asinternal standards.
HPLC was performed using a Waters Model 510 pump equippedwith Waters Rheodine injector and a differential refractometer,model 401.
Reaction progress was monitored via thin-layer chromatog-raphy (TLC) on Alugram� silica gel G/UV254 plates. Silica gel MNKieselgel 60 (70e230 mesh) from Macherey-Nagel Company wasused for column chromatography. All chemicals were obtainedfrom SigmaeAldrich, Inc. Solvents and reagents were used assupplied from commercial sources with the following exceptions.Hexane, dichloromethane and tetrahydrofuranewere distilled fromcalcium hydride immediately prior to use. Methanol was driedfrom magnesium methoxide as follow. Magnesium turnings (5 g)and iodine (0.5 g) are refluxed in a small (50e100 mL) quantity ofmethanol until all of the magnesium has reacted. The mixture isdiluted (up to 1 L) with reagent grade methanol, refluxed for 2e3 hthen distilled under nitrogen.
All reactions were carried out under argon atmosphere usingflame-dried glassware.
The purities of compounds were determined to be greater than95% by HPLC.
4.2. Sponge material and separation of individual sterols
4.2.1. T. conicaThe sponge (S07101) was collected off Amami-oshima Is.,
Kagoshima prefecture, Japan, on June 22, 2007 and kept at �25 �Cuntil extracted. The frozen sponge (1020 g) was extracted withMeOH (5�1000mL) and evaporated. The extract was suspended inH2O (1100 mL) and extracted with CHCl3 (2 � 800 mL) and n-BuOH(2 � 700 mL). The CHCl3 and n-BuOH layers were combined,evaporated, and partitioned between 90% MeOH (800 mL) and n-hexane (2 � 800 mL).
The n-hexane extract was chromatographed by silica gel MPLCusing a solvent gradient system from CH2Cl2 to CH2Cl2/MeOH 1:1.
Fractions eluted with CH2Cl2:MeOH 995:5 (40 mg) were furtherpurified by HPLC on a Nucleodur 100-5 C18 (5 mm; 10 mmi.d. � 250 mm) with MeOH/H2O (999.5:0.5) as eluent (flow rate3 mL/min) to give 0.7 mg of preconicasterol (3) (tR ¼ 58.5 min).
4.2.1.1. Preconicasterol 3. White amorphous solid; [a]D25 þ14.6 (c0.07, MeOH); 1H and 13C NMR data in C6D6 given in Table 1; HRMS-ESI m/z 397.3475 [M þ Hþ], C28H45O requires 397.3475.
4.2.2. T. swinhoei (R3170)For general experimental procedure, sponge collection and
Kupchan’s partitioning procedure see Zampella et al. [19]. The n-hexane extract (19.7 g) was chromatographed in two runs by silicagel MPLC using a solvent gradient system from CH2Cl2 to CHCl2/MeOH 1:1.
Fractions eluted with CH2Cl2:MeOH 99:1 (351 mg) were furtherpurified by HPLC on a Nucleodur 100-5 C18 (5 mm; 10 mmi.d. � 250 mm) with MeOH/H2O (998:2) as eluent (flow rate 3 mL/min) to give 1.7 mg of 25-dehydrotheonellasterol (5)(tR ¼ 12.4 min).
The chloroformic extract (4.76 g) was chromatographed by silicagel MPLC using a solvent gradient system from CH2Cl2 toCHCl2:MeOH 1:1.
Fractions eluted with CH2Cl2:MeOH 97:3 (3 mg) were furtherpurified by HPLC on a Luna 5m 100-5 C18 (5 mm; 4.5 mm
i.d. � 250 mm) with MeOH/H2O (95:5) as eluent (flow rate 0.8 mL/min) to give 0.2 mg of 24-dehydroconicasterol D (4) (tR ¼ 9 min).
4.2.2.1. 24-Dehydroconicasterol D 4. White amorphous solid;[a]D25 �67.0 (c 0.02, CHCl3); 1H and 13C NMR data in C6D6 given inTable 1; HRMS-ESI m/z 443.3527 [M þ Hþ], C29H47O3 requires443.3525.
4.2.2.2. 25-Dehydrotheonellasterol 5. White amorphous solid;[a]D25 þ17.9 (c 0.17, MeOH); 1H and 13C NMR data in C6D6 given inTable 1; HRMS-ESI m/z 425.3788 [M þ Hþ], C30H49O requires425.3783.
4.3. Chemical modification
4.3.1. (24R)-24-Methyl-4b-methyl-5a-cholest-8(14)en-3b-ol 6A conicasterol sample (10mg, 0.024mmol) was hydrogenated in
THF dry/MeOH dry 1:1 v/v (5 mL) in presence of Pt/C as catalyst.The reaction was stirred under H2 for 15 min. The mixture wasfiltered through SiO2 and the recovered filtrate was concentrated toafford 7 mg of pure compound 6 (0.017 mmol, 71%).
4.3.2. (24R)-24-Methyl-4b-methyl-5a-cholest-8(14)en-3b-ol 6[a]25D ¼ þ10.1 (c 0.03, CH3OH); selected 1H NMR (400 MHz
C6D6): d 3.53 (1H, m), 2.47 (1H, dd, J ¼ 3.2, 12.1 Hz), 1.03 (3H, d,J ¼ 6.5 Hz), 0.94 (3H, s), 0.93 (3H, d, J ¼ 6.6 Hz), 0.91 (3H, d,J ¼ 6.8 Hz), 0.86 (6H, d, J ¼ 6.8 Hz), 0.75 (3H, s). HRMS-ESI m/z415.3937 [M þ Hþ], C29H51O requires 415.3940.
4.3.3. (24R)-24-Methyl-4a-methyl-5a-cholest-8(14)en-3b-ol 7 and(24R)-24-methyl-4a-methyl-5a-cholest-8(14)en-3a-ol 8
Pyridinium chlorochromate (52 mg, 0.24 mmol) in CH2Cl2 dry(2mL) was added at the solution of conicasterol (50mg, 0.12mmol)in dichloromethane (5 mL). The reaction mixture was stirred atroom temperature for 12 h, then was added water. The aqueousphase was extracted with dichloromethane (3 � 30 mL) and thecombined organic phases were dried with Na2SO4 and evaporatedto dryness. The residue was passed through a short column of silicagel (2 g) and eluted with CH2Cl2 to give conicasterone [27] (50 mg,quantitative yield) as amorphous solid, that was subjected to nextstep without any purification procedure. Conicasterone was hy-drogenated in presence of palladium 5% wt on activated carbon(5 mg) in THF dry/MeOH dry 1:1 v/v (5 mL) in an oven-dried 25 mLflask. The reaction was stirred at room temperature under H2 for1 h. Then the mixture was filtered through SiO2 and the recoveredfiltrate was concentrated. Purification by HPLC on a Nucleodur Isis100-5 C18 (5 mm; 4.5 mm i.d. � 250 mm) with MeOH/H2O(999.5:0.5) as eluent (flow rate 1 mL/min) afforded 33.6 mg (67.3%from conicasterone) of 4b-methyl derivative (tR ¼ 48 min) and16.4 mg (32.7% from conicasterone) of 4a-methyl derivative(tR ¼ 52 min) as amorphous solids. To a solution of 4b-methyl de-rivative (30 mg, 0.07 mmol) in dry methanol (5 mL) was addedNaBH4 (13 mg, 0.35 mmol) at 0 �C. After 1 h, the reaction wasquenched by addition of MeOH (3 mL) and then concentrated un-der vacuo. Ethyl acetate and water were added and the separatedaqueous phase was extracted with ethyl acetate (3 � 30 mL). Thecombined organic phases were washed with water, dried withNa2SO4 and concentrated to obtain compound 6 as a white solid(26 mg, 87%).
Compounds 7 and 8 were obtained by NaBH4 reduction asdescribed before, starting from 4a-methyl derivative (15 mg). Theobtained mixture was purified by HPLC on a Nucleodur Isis 100-5C18 (5 mm; 4.5 mm i.d. � 250 mm) with MeOH/H2O (999.5:0.5) aseluent (flow rate 1 mL/min) to give 6.6 mg (44% from 4a-methyl

V. Sepe et al. / European Journal of Medicinal Chemistry 73 (2014) 126e134 133
derivative) of 7 (tR¼ 47.5 min) and 4.4 mg of 8 (31% from 4a-methylderivative) (tR ¼ 50 min) as amorphous solids.
4.3.4. (24R)-24-Methyl-4a-methyl-5a-cholest-8(14)en-3b-ol 7[a]25D ¼ þ7.2 (c 0.03, CH3OH); selected 1H NMR (400 MHz C6D6):
d 2.99 (1H, m), 2.00 (1H, dt, J ¼ 5.4, 12.6 Hz), 1.03 (3H, d, J ¼ 6.6 Hz),1.01 (3H, d, J¼ 6.5 Hz), 0.94 (3H, s), 0.91 (3H, d, J¼ 6.8 Hz), 0.86 (6H,d, J ¼ 6.6 Hz), 0.70 (3H, s). HRMS-ESI m/z 415.3935 [M þ Hþ],C29H51O requires 415.3940.
4.3.5. (24R)-24-Methyl-4a-methyl-5a-cholest-8(14)en-3a-ol 8[a]25D ¼ �6.6 (c 0.04, CH3OH); selected 1H NMR (400 MHz C6D6):
d 3.55 (1H, br m), 2.48 (1H, dd, J ¼ 3.1, 12.0 Hz), 1.03 (3H, d,J ¼ 7.0 Hz), 0.93 (6H, d, J ¼ 7.3 Hz), 0.91 (3H, s), 0.86 (6H, d,J ¼ 6.6 Hz), 0.75 (3H, s). HRMS-ESIm/z 415.3938 [M þ Hþ], C29H51Orequires 415.3940.
4.3.6. (24R)-24-Methyl-3b-hydroxyl-5a-cholest-8(14)en-4-one 9At a solution of conicasterol (5 mg, 0.012 mmol) in CH2Cl2 dry
kept under argon at�78 �Cwas bubbled a stream of O3 until a blue-colored solution resulted. After stirring for 1 h, excess of ozone wasremoved upon bubbling N2 and the solution was treated withexcess dimethylsulfide (2 mL). After 8 h, the solution was concen-trated under vacuo to remove the solvent and the mixture waspurified by HPLC on a Nucleodur Isis 100-5 C18 (5 mm; 4.5 mmi.d. � 250 mm) with MeOH/H2O (999.5:0.5) as eluent (flow rate1mL/min) to give 3.7mg (74%) of 9 (tR¼ 27.5min) as an amorphoussolid. [a]25D ¼ þ8.6 (c 0.17, CH3OH); selected 1H NMR (400 MHzC6D6): d 3.82 (1H, m), 1.01 (3H, d, J¼ 6.8 Hz), 0.91 (3H, d, J¼ 6.8 Hz),0.86 (6H, d, J ¼ 6.3 Hz), 0.82 (3H, s), 0.45 (3H, s); HRMS-ESI m/z415.3573 [M þ Hþ], C28H47O2 requires 415.3576.
4.3.7. (24R)-24-Methyl-4b-methyl-5a-cholest-8(14)en-3b-ol 6 and(24S)-24-methyl-4b-methyl-5a-cholest-8(14)en-3b-ol 10
At a solution of dehydroconicasterol (10 mg, 0.024 mmol) inhexane dry (5 mL) was added platinum oxide on carbon (5 mg) andthe flask was evacuated and flushed first with argon and then withhydrogen. The reaction was stirred at room temperature under H2for 5 min. The mixture was filtered through Celite, and the recov-ered filtrate was concentrated. The mixture was purified by HPLCon a Nucleodur Isis 100-5 C18 (5 mm; 4.5 mm i.d. � 250 mm) withMeOH/H2O (999.5:0.5) as eluent (flow rate 1 mL/min) to give2.6 mg of 6 (tR ¼ 48 min) and 4.0 mg of 10 (tR ¼ 53 min) asamorphous solids.
4.3.8. (24S)-24-Methyl-4b-methyl-5a-cholest-8(14)en-3b-ol 10[a]25D ¼ �8.0 (c 0.03, CH3OH); selected 1H NMR (400 MHz C6D6):
d 3.53 (1H, m), 2.47 (1H, dd, J¼ 3.2, 12.0 Hz), 1.03 (3H, d, J¼ 6.4 Hz),0.93 (3H,s), 0.92 (3H, d, J¼ 7.0 Hz), 0.91 (3H, d, J¼ 7.0 Hz), 0.86 (3H,d, J ¼ 6.6 Hz), 0.84 (3H, d, J ¼ 7.0 Hz), 0.74 (3H, s). HRMS-ESI m/z415.3942 [M þ Hþ], C29H51O requires 415.3940.
4.4. Luciferase assay
HepG2 cells were cultured at 37 �C in Minimum Essential Me-diumwith Earl’s salts containing10% fetal bovine serum (FBS), 1% L-glutamine and 1% penicillin/streptomycin. Cells were plated in a24-wells plate at 5 � 104 cells/well. The transfection experimentswere performed using Fugene HD (Promega, Milan, Italy) accordingto manufacturer specifications. For PXR mediated transactivation,cells were transfected with 75 ng pSG5-hPXRT1, 75 ng pSG5-RXR,125 ngpCMV-bgal and with 250 ng of the reporter vectorpGL3(henance)PXRE. At 24 h post-transfection, cells were primedwith rifaximin or compounds 3e10 (10 mM in agonism) or withcompounds 3e10 (50 mM plus rifaximin 10 mM, antagonism) for
18 h. Twenty mL of cellular lysates were read using the LuciferaseSubstrate (Promega) and luminescence was measured using theGlomax 20/20 luminometer (Promega). Luciferase activities werenormalized for transfection efficiencies by dividing the relativelight units by b-galactosidase activity expressed from co-transfected pCMV-bgal.
In another experimental setting, HepG2 cells were transfectedwith 75 ng pSG5-hPXRT1, 75 ng pSG5-RXR, 100 ng pGL4.70-Renillaand pGL3(henance)PXRE and treatedwith rising doses of 3 and 8 (1,10 and 50 mM) and rifaximin (1, 10 and 50 mM) as positive control.Twenty mL of cellular lysates were read using Dual-Luciferase Re-porter Assay System (Promega) according manufacturer specifica-tions. Luciferase activities were normalized for transfectionefficiencies by dividing the Luciferase relative light units (RLU) byRenilla relative lights units (RRU) expressed from cells co-transfected with pGL4.70-Renilla.
4.5. Real time PCR
HepG2 cells were stimulated 18 h with rifaximin (10 mM) andcompounds 3 and 8 (10 mM). Total RNA was extracted using theTRIzol reagent (Invitrogen), and reverse-transcribed using randomhexamer primers and Super Script-II reverse transcriptase (Invi-trogen). mRNA was quantified by Real-Time quantitative PCR oniCycler apparatus (Biorad) using specific primers: hGAPDH:gaaggtgaaggtcggagt and catgggtggaatcatattggaa; hCYP3A4: caa-gacccctttgtggaaaa and cgaggcgactttctttcatc; hMDR1: gtggggcaagt-cagttcatt and tcttcacctccaggctcagt; hMRP3: cacacggatctgacagacaatgaand acagggcactcagctgtctca. For quantitative RT-PCR, 10 ng of tem-plate was dissolved in a 20 mL solution containing 200 nM of eachprimer and 10 mL of KAPA SYBR FAST Universal qPCR Kit (KAPABIOSYSTEMS). All reactions were performed in triplicate, and thethermal cycling conditions were as follows: 3min at 95 �C, followedby 40 cycles of 95 �C for 15 s, 58 �C for 20 s and 72 �C for 30 s. Therelative mRNA expression was calculated accordingly with the Ctmethod. All PCR primers were designed using the softwarePRIMER3 (http://frodo.wi.mit.edu/primer3/) using publishedsequence data obtained from the NCBI database.
4.6. Computational details
Molecular docking of preconicasterol (3) and derivative 8 in thethree-dimensional X-ray structures of the PXR LBD (PDB codes:3hvl, 1nrl and 1m13) [32e34] without the co-crystallized inhibitorand waters were carried out using the AutoDock software package(version 4.2) [35]. Ligand and receptor structures were converted toAutoDock format files using the ADT software, and the Gesteiger-Marsili partial charges were then assigned. A box around thebinding pocket has defined the docking area and grids points of48 � 40 � 38 with 0.375�A spacing were calculated within this areafor all the ligand atom types using AutoGrid4. For each ligand, 100separate docking calculations were performed. Each docking runconsisted of 25 million energy evaluations using the Lamarckiangenetic algorithm local search (GALS) method. Otherwise defaultdocking parameters were applied. The docking conformations wereclustered on the basis of the root-mean square deviation values(rmsd tolerance ¼ 1.5 �A) between the Cartesian coordinates of theligand atoms and were ranked based on the AutoDock scoringfunction.
Acknowledgments
This work was supported by grants from MAREX-ExploringMarine Resources for Bioactive Compounds: From Discovery toSustainable Production and Industrial Applications (Call FP7-KBBE-

V. Sepe et al. / European Journal of Medicinal Chemistry 73 (2014) 126e134134
2009-3, Project nr. 245137) and fromMIUR, PRIN 2009 Sostanze adattività antitumorale: isolamento da fonti marine and PRIN 2010/2011 (E61J12000210001). NMR spectrawere provided by the CSIAS,Centro Interdipartimentale di Analisi Strumentale, Department ofPharmacy, University of Naples.
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.ejmech.2013.12.005.
References
[1] R.E. Watkins, G.B. Wisely, L.B. Moore, J.L. Collins, M.H. Lambert, S.P. Williams,T.M. Willson, S.A. Kliewer, M.R. Redinbo, The human nuclear xenobiotic re-ceptor PXR: structural determinants of directed promiscuity, Science 292(2001) 2329e2333.
[2] J.M. Rosenfeld, R. Vargas Jr., W. Xie, R.M. Evans, Genetic profiling defines thexenobiotic gene network controlled by the nuclear receptor pregnane X re-ceptor, Mol. Endocrinol. 17 (2003) 1268e1282.
[3] T.W. Synold, I. Dussault, B.M. Forman, The orphan nuclear receptor SXRcoordinately regulates drug metabolism and efflux, Nat. Med. 7 (2001) 584e590.
[4] B. Staudinger, S.A. Goodwin, D. Jones, K.I. Hawkins-Brown, A. MacKenzie,Y. LaTour, C.D. Liu, K.K. Klaassen, J. Brown, T.M. Reinhard, B.H. Willson,S.A. Kliewer, The nuclear receptor PXR is a lithocholic acid sensor that protectsagainst liver toxicity, Proc. Natl. Acad. Sci. U. S. A. 98 (2001) 3369e3374.
[5] S. Fiorucci, S. Cipriani, F. Baldelli, A. Mencarelli, Bile acid-activated receptors inthe treatment of dyslipidemia and related disorders, Prog. Lipid Res. 49 (2010)171e185.
[6] J. Gao, W. Xie, Targeting xenobiotic receptors PXR and CAR for metabolicdiseases, Trends Pharmacol. Sci. 33 (2012) 552e558.
[7] H.I. Swanson, T. Wada, W. Xie, B. Renga, A. Zampella, E. Distrutti, S. Fiorucci,B. Kong, A.M. Thomas, G.L. Guo, R. Narayanan, M. Yepuru, J.T. Dalton,J.Y. Chiang, Role of nuclear receptors in lipid dysfunction and obesity-relateddiseases, Drug Metab. Dispos. 41 (2013) 1e11.
[8] J. Cheng, Y.M. Shah, X. Ma, X. Pang, T. Tanaka, T. Kodama, K.W. Krausz,F.J. Gonzalez, Therapeutic role of rifaximin in inflammatory bowel disease:clinical implication of human pregnane X receptor activation, J. Pharmacol.Exp. Ther. 335 (2010) 32e41.
[9] Y. Chen, Y. Tang, C. Guo, J. Wang, D. Boral, D. Nie, Nuclear receptors in themultidrug resistance through the regulation of drug-metabolizing enzymesand drug transporters, Biochem. Pharmacol. 83 (2012) 1112e1126.
[10] S. Fiorucci, A. Zampella, E. Distrutti, Development of FXR, PXR and CAR ago-nists and antagonists for treatment of liver disorders, Curr. Top. Med. Chem.12 (2012) 605e624.
[11] M.V. D’Auria, V. Sepe, A. Zampella, Natural ligands for nuclear receptors:biology and potential therapeutic applications, Curr. Top. Med. Chem. 12(2012) 637e669.
[12] S. Fiorucci, E. Distrutti, G. Bifulco, M.V. D’Auria, A. Zampella, Marine spongesteroids as nuclear receptor ligands, Trends Pharmacol. Sci. 33 (2012) 591e601.
[13] F.S. Di Leva, C. Festa, C. D’Amore, S. De Marino, B. Renga, M.V. D’Auria,E. Novellino, V. Limongelli, A. Zampella, S. Fiorucci, Binding mechanism of thefarnesoid X receptor marine antagonist suvanine reveals a strategy to forestalldrug modulation on nuclear receptors. Design, synthesis, and biologicalevaluation of novel ligands, J. Med. Chem. 56 (2013) 4701e4717.
[14] C. Festa, G. Lauro, S. De Marino, M.V. D’Auria, M.C. Monti, A. Casapullo,C. D’Amore, B. Renga, A. Mencarelli, S. Petek, G. Bifulco, S. Fiorucci,A. Zampella, Plakilactones from the marine sponge Plakinastrella mamillaris.Discovery of a new class of marine ligands of peroxisome proliferator-activated receptor g, J. Med. Chem. 55 (2012) 8303e8317.
[15] V. Sepe, G. Bifulco, B. Renga, C. D’Amore, S. Fiorucci, A. Zampella, Discovery ofsulfated sterols from marine invertebrates as a new class of marine naturalantagonists of farnesoid-X-receptor, J. Med. Chem. 54 (2012) 1314e1320.
[16] M.G. Chini, C.R. Jones, A. Zampella, M.V. D’Auria, B. Renga, S. Fiorucci,C.P. Butts, G. Bifulco, Quantitative NMR-derived interproton distances com-bined with quantum mechanical calculations of 13C chemical shifts in thestereochemical determination of conicasterol F, a nuclear receptor ligandfrom Theonella swinhoei, J. Org. Chem. 77 (2012) 1489e1496.
[17] C. Festa, C. D’Amore, B. Renga, G. Lauro, S. De Marino, M.V. D’Auria, G. Bifulco,A. Zampella, S. Fiorucci, Oxygenated polyketides from Plakinastrella mamillarisas a new chemotype of PXR agonists, Mar. Drugs 11 (2013) 2314e2327.
[18] C. Festa, S. De Marino, M.V. D’Auria, G. Bifulco, B. Renga, S. Fiorucci, S. Petek,A. Zampella, Solomonsterols A and B from Theonella swinhoei. The firstexample of C-24 and C-23 sulfated sterols from a marine source endowedwith a PXR agonistic activity, J. Med. Chem. 54 (2011) 401e405.
[19] S. De Marino, R. Ummarino, M.V. D’Auria, M.G. Chini, G. Bifulco, B. Renga,C. D’Amore, S. Fiorucci, C. Debitus, A. Zampella, Theonellasterols and con-icasterols from Theonella swinhoei. Novel marine natural ligands for humannuclear receptors, J. Med. Chem. 54 (2011) 3065e3075.
[20] S. De Marino, V. Sepe, M.V. D’Auria, G. Bifulco, B. Renga, S. Petek, S. Fiorucci,A. Zampella, Towards new ligands of nuclear receptors. Discovery of malai-tasterol A, an unique bis-secosterol from marine sponge Theonella swinhoei,Org. Biomol. Chem. 9 (2011) 4856e4862.
[21] S. De Marino, R. Ummarino, M.V. D’Auria, M.G. Chini, G. Bifulco, C. D’Amore,B. Renga, A. Mencarelli, S. Petek, S. Fiorucci, A. Zampella, 4-Methylenesterolsfrom Theonella swinhoei sponge are natural pregnane-X-receptor agonistsand farnesoid-X-receptor antagonists that modulate innate immunity, Ste-roids 77 (2012) 484e495.
[22] V. Sepe, R. Ummarino, M.V. D’Auria, M.G. Chini, G. Bifulco, B. Renga,C. D’Amore, C. Debitus, S. Fiorucci, A. Zampella, Conicasterol E, a small het-erodimer partner sparing farnesoid X receptor modulator endowed with apregnane X receptor agonistic activity, from the marine sponge Theonellaswinhoei, J. Med. Chem. 55 (2012) 84e93.
[23] V. Sepe, R. Ummarino, M.V. D’Auria, A. Mencarelli, C. D’Amore, B. Renga,A. Zampella, S. Fiorucci, Total synthesis and pharmacological characterizationof solomonsterol A, a potent marine pregnane-X-receptor agonist endowedwith anti-inflammatory activity, J. Med. Chem. 54 (2011) 4590e4599.
[24] E. Kho, D.K. Imagawa, M. Rohmer, Y. Kashman, C. Djerassi, Sterols in marineinvertebrates. 22. Isolation and structure elucidation of conicasterol andtheonellasterol, two new 4-methylene sterols from the Red Sea spongesTheonella conica and Theonella swinhoei, J. Org. Chem. 46 (1981) 1836e1839.
[25] B. Renga, A. Mencarelli, C. D’Amore, S. Cipriani, M.V. D’Auria, V. Sepe,M.G. Chini, M.C. Monti, G. Bifulco, A. Zampella, S. Fiorucci, Discovery thattheonellasterol a marine sponge sterol is a highly selective FXR antagonistthat protects against liver injury in cholestasis, PLoS One 7 (2012) e30443.
[26] V. Sepe, R. Ummarino, M.V. D’Auria, O. Taglialatela-Scafati, S. De Marino,C. D’Amore, B. Renga, M.G. Chini, G. Bifulco, Y. Nakao, N. Fusetani, S. Fiorucci,A. Zampella, Preliminary structureeactivity relationship on theonellasterol, anew chemotype of FXR antagonist, from the marine sponge Theonella swin-hoei, Mar. Drugs 10 (2012) 2448e2466.
[27] M. Kobayashi, K. Kawazoe, T. Katori, I. Kitagawa, Marine natural products XXX.Two new 3-keto-4-methylene steroids, theonellasterone and conicasterone,and a DielseAlder type dimeric steroid bistheonellasterone, from the Okina-wan marine sponge Theonella swinhoei, Chem. Pharm. Bull. 40 (1992) 1773e1778.
[28] Compound 6 was tested, in combination with rifaximin (10 mM), in a dose-responsive manner (10, 25, 50 mM) in a luciferase reporter assay. No effectwas observed at 10 and 25 mM doses and the slightly antagonistic effectrespect to rifaximin at 50 mM resulted not enough significant from a statisticpoint of view, thus preventing from the chance to calculate a formal IC50.
[29] S. Ekins, C. Chang, S. Mani, M.D. Krasowski, E.J. Reschly, M. Iyer,V. Kholodovych, N. Ai, W.J. Welsh, M. Sinz, P.W. Swaan, R. Patel, K. Bachmann,Human pregnane X receptor antagonists and agonists define molecular re-quirements for different binding sites, Mol. Pharmacol. 72 (2007) 592e603.
[30] B.L. Wagner, G. Pollio, P. Giangrande, J.C. Webster, M. Breslin, D.E. Mais,C.E. Cook, W.V. Vedeckis, J.A. Cidlowski, D.P. McDonnell, The novel proges-terone receptor antagonists RTI 3021-012 and RTI 3021-022 exhibit complexglucocorticoid receptor antagonist activities: implications for the develop-ment of dissociated antiprogestins, Endocrinology 140 (1999) 1449e1458.
[31] J.T. Link, B. Sorensen, J. Patel, M. Grynfarb, A. Goos-Nilsson, J. Wang, S. Fung,D. Wilcox, B. Zinker, P. Nguyen, B. Hickman, J.M. Schmidt, S. Swanson, Z. Tian,T.J. Reisch, G. Rotert, J. Du, B. Lane, T.W. von Geldern, P.B. Jacobson, Antidia-betic activity of passive nonsteroidal glucocorticoid receptor modulators,J. Med. Chem. 48 (2005) 5295e5304.
[32] W. Wang, W.W. Prosise, J. Chen, S.S. Taremi, H.V. Le, V. Madison, X. Cui,A. Thomas, K.C. Cheng, C.A. Lesburg, Construction and characterization of afully active PXR/SRC-1 tethered protein with increased stability, Protein Eng.Des. Sel. 21 (2008) 425e433.
[33] R.E. Watkins, P.R. Davis-Searles, M.H. Lambert, M.R. Redinbo, Coactivatorbinding promotes the specific interaction between ligand and the pregnane Xreceptor, J. Mol. Biol. 331 (2003) 815e828.
[34] R.E. Watkins, J.M. Maglich, L.B. Moore, G.B. Wisely, S.M. Noble, P.R. Davis-Searles, M.H. Lambert, S.A. Kliewer, M.R. Redinbo, A crystal structure of hu-man PXR in complex with the St. John’s wort compound hyperforin,Biochemistry 42 (2003) 1430e1438.
[35] G.M. Morris, R. Huey, W. Lindstrom, M.F. Sanner, R.K. Belew, D.S. Goodsell,A.J. Olson, Autodock4 and AutoDockTools4: automated docking with selectivereceptor flexibility, J. Comput. Chem. 30 (2009) 2785e2791.





























