Embed Size (px)
Citation preview

This article was downloaded by: [Purdue University]On: 20 July 2015, At: 07:57Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: 5 Howick Place, London, SW1P 1WG
Journal of Applied AnimalResearchPublication details, including instructions forauthors and subscription information:http://www.tandfonline.com/loi/taar20
Inhibition of StructuralCarbohydrate Fermentation byCellulase Filtrates of AlfalfaD. J. R. Cherney a , J. H. Cherney b & J. J. Voleneca Departments of Animal Science , CornellUniversity , Ithaca, NY, 14853-4801, USAb Crop, Soil and Atmospheric Science , CornellUniversity , Ithaca, NY, 14853-4801, USAPublished online: 11 Nov 2011.
To cite this article: D. J. R. Cherney , J. H. Cherney & J. J. Volenec (1993) Inhibitionof Structural Carbohydrate Fermentation by Cellulase Filtrates of Alfalfa, Journal ofApplied Animal Research, 3:1, 19-30, DOI: 10.1080/09712119.1993.9705949
To link to this article: http://dx.doi.org/10.1080/09712119.1993.9705949
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified withprimary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.

This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015

J. Appl. Anim. Res 3 (1993) 19-30
Inhibition of Structural Carbohydrate Fermentation by Cellulase Filtrates
of Alfalfa D. J. R. Cherney', J. H. Cherney2, J. J. Volenee.
Departments of Animal Science'; Crop, Soil and Atmospheric Science' Cornell University, Ithaca, NY, USA, 14853-4801
(Received November 4,1992; accepted January 1, 1993)
Abstract Cherney, D. J. R., Cherney, J. H. andvolenec, J. J. 1993. Inhibition of structural carbohydrate fermentation by cellulase filtrates of alfalfa. J. Appl. A n i i . Res., 3 : 19-30
Objectives were to investigate the influence of isolated phenol ic-ca rbohyd ra te complexes from a1 fa 1 f a (Med icago sa tiva L.) on structural carbohydmte fermentation. Neutral detergent residues of leaf and stem alfalfa tissues were incubated in purifed cellulase and filtrates fractionated using size-exclusion chromatography on Sepharose CL-6B. One major absorbance peak (A320) was observed from these fractions. The fraction containing theabsorbancepeak (A320) inhibited in vitrodigestion ofstructuml carbohydrates by rumen microorganisms. Contents of this CL-6B peak were further fractionated on Sephadex LH-20. In vitro incubations with the one major peak from LH-20 also inhibited digestion of structural carbohydrates. It is not known whether phenolic-portions of the complexes were responsible for fiber digestion inhibition or i f carbohydrate-portions were responsible for the observed effect.
*Department of Amnomy, Rtrdue University, West'hfayette, IN 47907.
19
J. Appi. Anii. Res. 0971-2119193/$05.00 0 GSP, India
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015

20 D. J. R. Cherney and coworkers
Key words : Phenolic-carbohydrate complexes, fiber digestion, neutral detergent fiber, alkali-labile phenolics.
In trod uc tion Phenolic-carbohydrate complexes may be a factor limiting structural carbohydrate digestion in ruminants (Akin et al., 1988). Several studies have investigated the inhibitory potential of cellulase-extracted or rumen microorganism-extracted plant filtrates on structural carbohydrate fermentation. Jung (1988) indicated that complexes released during in uitm ruminal fermentation of alfalfa (Medicago satiua L.) hay, smooth bromegrass (Bromus inermus Leyss.) hay and maize (Zea mays L.) silage restricted fiber fermentation. Cherney et al. (1992) reported inhibition of fiber digestion when cellulase filtrates of sorghum stem [Sorghum bicolor (L.) Moench] were added to in v i m fermentations of sorghum leaf blade. Cellulase filtrates separated by size exclusion chromatography (Sepharose CL-6B or Sephadex LH-20) indicated that only one fraction inhibited structural carbohydrate digestion. These plant filtrates were phenolic-carbohydrate complexes that appeared to be high in ferulic acid(FA) and had highxylose : arabinose ratios. Legumesmay not behave similarly. Objectives of this study were to isolate phenolic-carbohydrate complexes from alfalfa cell walls and to investigate the influence of isolated complexes on structural carbohydrate fermentation.
Materials and Methods Alfalfa (variety Hi-Phy) samples were field harvested, immediately frozen, lyophylized, and ground to pass through a cyclone mill (UDY Corporation, Fort Collins, CO; 0.5-mm screen). Maturity, determined using the mean stage by count procedure of Kalu and Fick (1981) was 2.79, which corresponds to an early bud stage. Samples were separated into leaf and stem fractions.
Sample processing is essentially that described previously (Cherney et al., 1992) and is outlined in Fig. 1. Neutral detergent residues were prepared from dried, ground plant samples refluxed in neutral detergent (Robertson and Van Soest, 1981; as modified by Cherney et al., 1989 a) for 1 h and dried at 60 C. Residues were incubated in purified cellulase (Worthington Biochemical Corp., Freehold, NJ) for 48 h. Filtrates were lyophylized, reconstituted, and separated into fractions using size exclusion chromatography. Filtrate from 8 g of neutral detergent residues was injected onto the chromatography column. The column (5 x 100 cm) contained Sepharose CL-6B (Pharmacia Co., Piscataway, NJ); fractions were eluted using 0.02 M NaCl which had been degassed under vacuum. Fractions were collected in test tubesusing a flow rate of approximately
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015

Alfalfa cellulase filtrates-fiber digestibility 21
Alfalfa leaf and stem tissues refluxed in ND (lh) and dried (60y)
chromatographically-purified cellulase (1050 units mgl fiber) (0.5 M acetate buffer, pH 4.8)
1
1
ND residue (8 g)
1 Cellulase extract (200 mL) 1 lyophilized
Reconstituted to concentrated volume (5 mL) size exclusion chromatography (Sepharose CLdB, 5x100 cm column) (0.02 M NaCl at 1.8 mL, min-l)
17 CL-6B Fractions (void to inclusion volumes)
Influence of stem fractions on in vitro structural CHO digestion
Phenolic acid analysis
Stem fraction exhibiting effect
Size exclusion chromatography (Sephadex LH-20,2.5 x 100 cm column) (0.5% acetic acid at 0.5 mL min-1) I
10 LH-20 Fractions (void to inclusion volumes) + Influence of stem fractions on in vitro structural CHO digestion
Fig. 1 : Scheme of analysis [chemical and in vitro structural carbohydrates (CHO)] of cellulase filtrates of neutral detergent (NO) extracted alfalfa tissues.
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015

22 D. J. R. Cherney and coworkers
1.8 ml min-'. Absorbance at 280 and 320 nm was used to determine location of peaks. These are primary wavelengths for absorbance by many phenolic compounds Vanner and Morrison, 1983).
Isolated fractions of interest from the Sepharose CL-6B separation were further size fractionated using a column (2.5 x 100 cm) containing Sephadex LH-20 (Pharmacia Co., Piscataway, NJ). Fractions were eluted using 0.5% acetic acid which had been degassed under vacuum. Fractions were collected in test tubes using a flow rate of approximately 0.5 ml min-l.
For phenolic analyses, isolated peaks from chromatographic separations were hydrolyzed in 2 ml of 1 M NaOH for 24 h under Nz at room temperature in the absence of light to yield alkali-labile phenolics. Samples were acidified with 0.36 ml of 6 M HCl, vacuum filtered and brought to 12.5 ml volume with water. Phenolics were determined using high-performance liquid chromatography (Cherney et al., 1989 b). Aqueous acetic acid (0.7%; solvent A) and 50% aqueous acetonitrile (Solvent B) were the solvents w e d under gradient conditions. The gradient went from 10 to 20% B over the first 10 min, 20 to 50% B over the next 15 min, and 50% B for 5 min, followed by re-equilibration.
A series of studies using standard in uitro digestions using filtered rumen fluid (Marten and Barnes, 1980) were carried out in sealed serum bottles to investigate the influence of isolated fractions on neutral detergent fiber (NDF) digestion. After digestion, samples were extracted with neutral detergent solution (Robertson and Van Soest, 1981, as modified by Cherney et al., 1989 a), filtered, rinsed and dried at 100 C. In uitro incubations were conductedusing either 0.25 g dry matter of alfalfa or orchardgrass (Dactylis glomerata L.) as substrate. In all studies, filtrates were lyophylized and reconstituted such that filtrates added to incubations did not exceed 4% of the total volume. Equal treatment volumes were added to all incubations. Concentrated solutions of isolated peaks were added to serum bottles 2 h after the start of the incubation with rumen fluid. Samples were corrected for bacterial and residual dry matter by use of rumen fluid blanks.
Rate of digestion of alfalfa, as influenced by addition of isolated peaks at quantities of 0 and 1.5 g alfalfa stem NDF, was determined. T i e s used for determination of rate were 0,12,24,48 and 72 h. Limited amounts of cellulase extracts precluded the inclusion of more sample times. Estimates for kinetic parameters of digestion were determined using a direct non-linear least squares estimating procedure (SAS, 1982). The model for kinetics of digestion was a simple f i t -order kinetic equation with a discrete lag time (Mertens and Loften, 1980). All coefficients of determination (r2) were greater than 0.99. Statistical
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015

Alfalfa cellulase filtmtes-fiber digestibility 23
design was a randomized complete block with a 2 x 2 x 2 factorial arrangement of substrate, rate and replication.
To determine the influence of Sepharose CL-6B isolated fractions on in ui tro dry matter digestibility of orchardgrass, 17 sections of approximately 100 ml each; including the void and inclusion volumes, were isolated and lyophylized. Concentrated solutions of isolated sections were added to in ui tro incubations at quantities equivalent to 0, 51 , and 2 g of alfalfa stem NDF. Statistical design was a randomized complete block with a 4 x 17 x 2 factorial arrangement of rate, fraction and replication.
Further studies were conducted to determine the influence of Sephadex LH-20 isolated sections on fiber digestion, again using 0.25 g of orchardgrass as a substrate. Ten groups of fractions, including the void and inclusion volumes, were isolated and lyophylized. Concentrated solutions of isolated fractions were added to in uitm incubations at quantities equivalent to 0,1.75 and 5.0 g alfalfa NDF.
In all studies, concentrated solutions of isolatedpeaks were added to serum bottles 2 h after the start of the incubation with rumen fluid. Some phenolic acids are very sensitive to air oxidation @hang et al., 1986). Delaying inoculation until 2 h after the start of incubation with rumen fluid helped to assure an anaerobic system. Samples were corrected for bacterial and residual dry matter by use of rumen fluid blanks.
Results and Discussion Filtrates of cellulase digested NDF of alfalfa leaf and stem separated by size exclusion chromatography yielded one major absorbance peak at 320 nm (Fig. 2). This is in contrast to the two major absorbance peaks noted for sorghum leaf blade, leaf sheath and stem filtrates reported by Cherney et al. (1992). Cellulase filtrates of NDF of stem from pearl millet Pennisetum glaucum (L.)], corn, orchardgrass and switchgrass [Panicurn uirgatum &.)I also yielded two major absorbance peaks (data not shown). Profiles at 280 nm were similar to those at 320 nm. Leaf and stem peaks eluted at approximately the same volumes (1800 ml), although peak absorbance profiles for leaf (0.065 f 0.015) were lower than for peak stem absorbance profiles (0.1744 f 0.0239). The second major absorbance peak for sorghum leaf blade and stem filtrates also eluted approximately at 1800 ml volume, and leaf blade absorbances were lower than stem absorbances (Cherney et aE., 1992). However, peak absorbance values were the order of 10 times lower for the alfalfa than for sorghum. Maximum absorbances for orchardgrass, switchgrass, maize and pearl millet were similar
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015
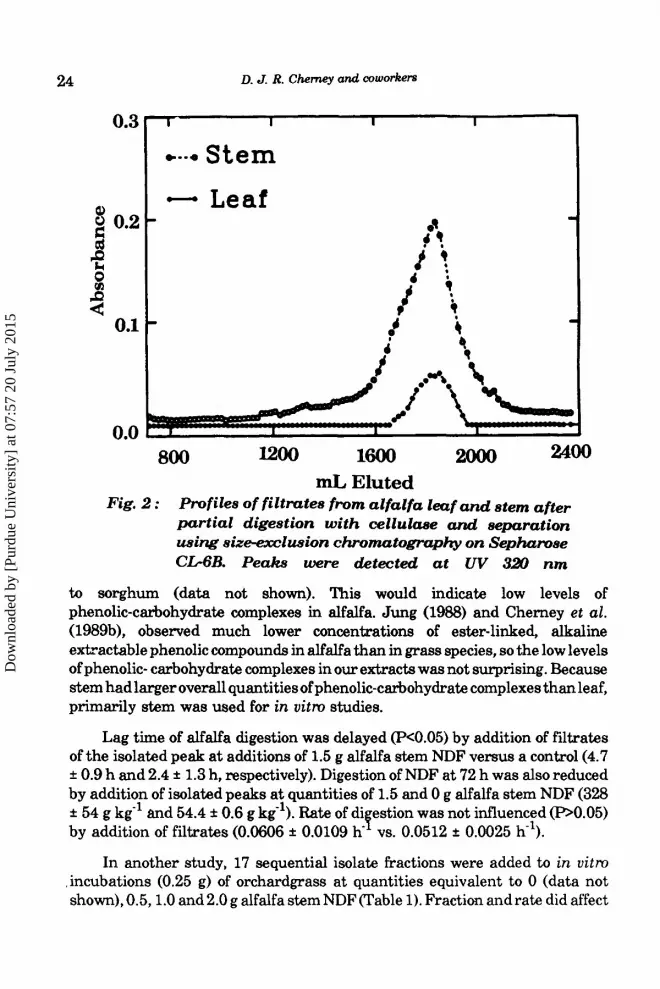
24 D. J. R. Chemey and coworkers
0.3
8 0.2 9 -2
3 0.1
0.0 800 1200 1600 2000 2400
mL Eluted Fig. 2 : Profiles of filtrates from alfalfa leaf a d stem after
partial digestion with cellulase and separation using aize-eazlusion chromatography on Sephamae C M B . Peakt were detected at UV 320 nm
to sorghum (data not shown). This would indicate low levels of phenolic-carbohydrate complexes in alfalfa. Jung (1988) and Cherney et al. (1989b), observed much lower concentrations of ester-linked, alkaline extractable phenolic compounds in alfalfa than in grass species, so the low levels of phenolic- carbohydrate complexes in our extracts was not surprising. Because stem had larger overall quantities of phenolic-carbohydrate complexes than leaf, primarily stem was used for in uitm studies.
Lag time of alfalfa digestion was delayed (p<0.05) by addition of filtrates of the isolated peak at additions of 1.5 g alfalfa stem NDF versus a control (4.7 k 0.9 h and 2.4 * 1.3 h, respectively). Digestion of NDF at 72 h was also reduced by addition of isolated peaks at quantities of 1.5 and 0 g alfalfa stem NDF (328 f 54 g kg-' and 54.4 f 0.6 g kg-l). Rate of di estion was not influenced P0.05 ) by addition of filtrates (0.0606 * 0.0109 h- VS. 0.0512 2 0.0025 h-'). f
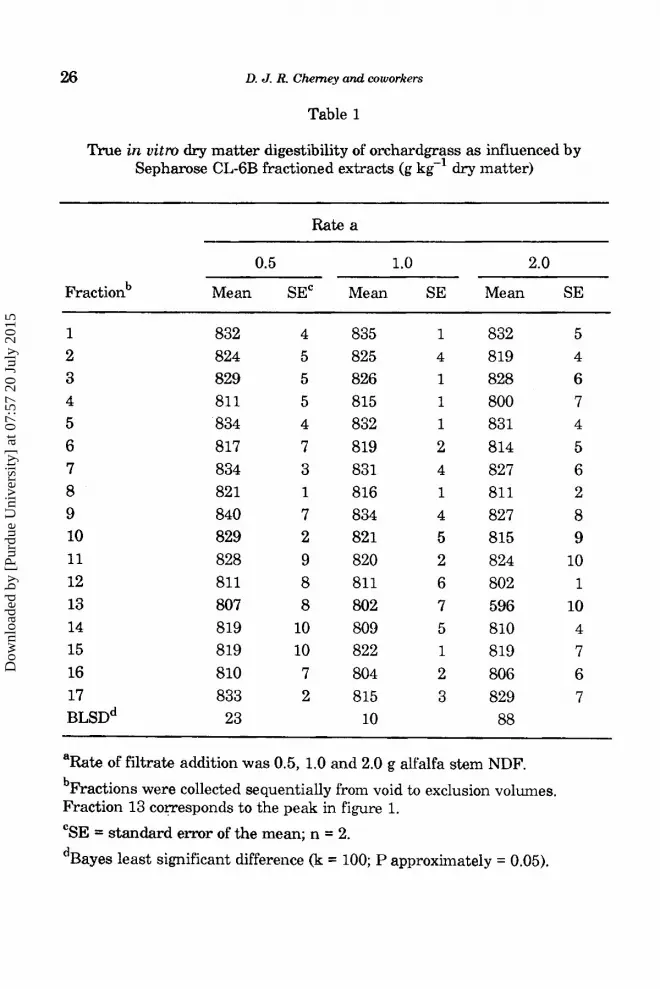
In another study, 17 sequential isolate fractions were added to in vitro ,incubations (0.25 g) of orchardgrass at quantities equivalent to 0 (data not shown), 0.5,l.O and 2.0 g alfalfa stem NDFflable 1). Fraction and rate did affect
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015

Alfalfa celluluse filtrates-fiber digestibility 25
digestibility (P<O.Ol), but there also was a fraction x rate interaction (P<O.Ol). This was primarily due to the small effect of the 0.5 and 1.0 g quantities on digestibility, and the large effect of rate on digestibility at the 2.0 g quanitity. Fraction 13 had the lowest digestibility at all rates, and this fraction corresponded to the absorbance peak observed in Fig. 2. Neutral detergent fiber of orchardgrass was 569 g kg-', indicating that fraction 13 at the 2.0 g quantity almost totally inhibited fiber digestion (596 g kg-l). Control sample digestibility averaged 821 & 3 g kg' in this study, indicating that other fractions at all rates did not greatly affect digestion. Fraction 13 in sorghum also inhibited fiber digestion (Cherney et aZ., 1992), suggesting that compounds of similar molecular weight could be inhibiting digestion in both alfalfa and sorghum.
Sepharose CL-6B has a molecular sieve fractionation range of 1 x lo4 to 1 x lo6 forpolysacharrides. Inclusion volume for this column is at approximately 2000 ml, approximately where the peak eluted. Because all compounds less than 1 x lo4 molecular weight elute at this point, including glucose, further fractionation is necessary to determinethe molecular weight ( s ) of the compound or compounds eliciting the inhibitory effect.
Peak filtrates from CL-6B, further separated by size exclusion chromatography using sephadex LII-20, yielded one major absorbance peak at 320 nm, unlike the absorbance profiles reported for sorghum, which contained several distinct absorbance peaks (Cherney et al., 1992). Absorbance profiles of orchardgrass and switchgrass were similar to those of sorghum in terms of appearance and peak absorbance (datanot shown). The area of peak absorbance for alfalfa stem filtrates eluted at the same volume as reported for sorghum. Like the Sepharose CL-6B fractionation, however, maximum absorbance values for the sorghum were 10 times higher than those reported here for alfalfa.
Sephadex LH-20 isolates of the one major peak in alfalfa leaf and skin an< corresponding peak in switchgrass were added to in uitro incubations (0.25 g) of orchardgrass at quantities equivalent to 5.0 g of NDF. Fiber digestion was inhibited almost completely by these isolates from alfalfa and switchgrass. In further studies, 10 Sephadex LH-20 sequential isolate fractions were added to in uitro incubations (0.25 g) of orchardgrass at 0, 1.75 and 5.0 g of alfalfa stem NDF to detennine if components of the filtrate that did not absorb at 320 nm also affected digestion (Table 2). It was observed that only fraction 3 reduced digestion over the controls (823 k 1 in this in uitro study). Fraclion 3 corresponded to one of the fractions in sorghum that exhibited fiber digestion inhibition (Cherney et aZ., 1992), indicating that cdmdexes affecting digestion may be similar.
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015
26 D. J. R. Cherney and coworkers
Table 1
True in uitm dry matter digestibility of orchardgrass as influenced by Sepharose CL-6B fractioned extracts (g kg-’ dry matter)
Rate a
0.5 1.0 2.0
Fractionb Mean SEC Mean SE Mean SE
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 BLSD~
832 824 829 81 1 834 817 834 821 840 829 828 811 807 819 819 810 833 23
4 5 5 5 4 7 3 1 7 2 9 8 8
10 10 7 2
835 1 825 4 826 1 815 1 832 1 819 2 831 4 816 1 834 4 821 5 820 2 811 6 802 7 so9 5 822 1 804 2 815 3
10
832 819 828 800 831 814 827 811 827 815 824 802 596 810 819 806 829 88
5 4 6 7 4 5 6 2 8 9
10 1
10 4 7 6 7
aRate of filtrate addition was 0.5, 1.0 and 2.0 g alfalfa stem NDF. bFractions were collected sequentially from void to exclusion volumes. Fraction 13 cozresponds to the peak in figure 1.
“SE = standard error of the mean; n = 2. dBayes least significant difference (k = 100; P approximately = 0.05).
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015

Alfalfa celluIase filtrates-fiber digestibility 27
Table 2
True in uitm dry matter digestibility of orchardgrass as influenced by Sephadex LH-20 fractioned extracts (g kg-1 dry matter)
Fractionb Ratea
1.75 5.0
1 2 3 4 5 6 7 8 9 10
815 838 791 825 839 822 827 836 833 838
811 833 474 837 820 822 812 830 831 828
SE 1 3
aRate of filtrate addition was 1.75, and 5.0 g alfalfa stem NDF. bF'ractions were collected sequentially from void to exclusion volumes. Fraction 3 corresponds to the maximum peak absorbance from this column.
Absorbances at 280 and 320 were similar, indicating that complexes were not protein in nature, as protein absorbs most strongly at 280 and not at 320. In additionmolecular weight of complexes was relatively low (<low), indicating non-protein complexes. Absorbance was possibly due to phenolic-carbohydrate complexes. Any individual phenolic monomers would have eluted late in the LH-20 profiles, as would individual hemicellulosic sugars. phenolic analyses of fraction 3 alfalfa stem isolates indicated that PCA was in lower concentration than FA (ratio of 0.35), which is in agreement with the lower concentraion of
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015

28 D. J. R. Cherney and coworkers
PCA than FA reported in fraction 3 isolates of sorghum (Cherney et al., 1992). I t is not clear from the present study, however, if phenolics in the complexes are responsible for the observed reductions in fiber digestibility or if the carbohydrate complexes were involved in end product inhibition. Alfalfa stem filtrates appeared to have similar effects as sorghum, despite a 10 fold reduction in peak absorbance at 320 nm, which would indicate a lower concentration of phenolics.
While the machanism responsible for reduction in fiber digestibility was not fully elucidated, it did not appear to be a steric effect. If the mechanism of action was one of blocking attachment sites for digestion, then increasing rates of filtrate addition should have resulted in a linear response. This did not appear to happen. Addition of filtrates had little effect on digestion at lower rates. Griggs et aE. (1989) indicated that mixed nuninal bacteria growth was inhibited by addition of cellulase filtrates of corn separated with a CL-6B column (corresponded to our peak) at early times, but that optical densities equalled or exceeded controls at later times. Because optical density was greater than controls a t later times, it was suggested that bacterial populations had been changed (Griggs et aE., 1989). Cherney et al. (1990) indicated that mixedruminal bacteria cultures were able to eventually overcome the addition of chlorogenic acid, a phenolic acid, added at 0,2 or 4% of substrate dry weight, such that by 48 hours, no difference in extent of digestion was observed between treated samples and controls. At the higher rates used in this study, the filtrates appeared to have a sterilization effect. Further research is needed to characterize the mechanism of action of fiber inhibition and to identify which microorganisms are affected.
Acknowledgement Contribution of Cornell University and %due University (Agricultural Experiment Station Journal Paper No. 12830).
References Akin, E. E., Rigsby, L. L., Theodorou, M. K. and Hartley, R. D. 1988. Population changes
of fibrolytic rumen bacteria in the presence of phenolic acids and plant extracts. Anim. Feed Sci. Technol., 19 : 261-276.
Cherney, D. J. R., Cherney, J. H., Patterson, J. A. and Axtell, J. D. 1992. In v i m fiber digestion as influenced by phenolic-carbohydrate complexes in sorghum cell walls. Anim. Feed Sci. Technol., 39 : 79-93.
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015

Alfalfa cellulase filtrates-fiber digestibility 29
Cherney, D. J. R., Patterson, J. A. and Cherney, J. H. 1989a. Use of 2-exthoxyethanol and a-amylase in the neutral detergent fiber method of feed analysis. J. Dairy Sci., 71 : 3079-3084.
Cherney, J. H., Anliker, K. S., Albrecht, K. A. and Wood, K. V. 1989b. Soluble phenolic monomers in forage crops. J. Agric. Food Chem., 37 : 345-350.
Cherney, J. H., Cherney, D. J. R., Sollenberger L. E., Patterson, J. A. and Wood, K. V. 1990. Identification of 5-0-caffeoylquinnic acid in limpograss and its influence on fiber digestion. J. Agric. Food Chem., 38 : 2140-2143.
Griggs, T. C., Cherney, J. H., Cherney, D. J. R. and Patterson, J. A. 1989. Phenolic-carbohydrate complexes in maize cell walls and influences on rumen microbes. 1989 Annual Meetings of the American Society of Agronomy, Las Vegas, NV. Agron. Abst., p. 169.
Huang, H. M., Johanning, G. L. and O’Dell, B. L. 1986. Phenolic acid content of food plants and possible nutritional implications. J. Agric. Food Chem., 34 : 48-51.
Jung, H. G. 1988. Inhibitory potential of phenolic-carbohydrate complexes released during nuninal fermentation. J. Agric. Food Cheni., 36 : 782-788.
Kalu, B. A. and Fick, G. W. 1981. Quantifying morphological development of aUalla for studies of herbage quality. Crop Sci., 21 : 267-271.
Marten, G. C. and Barnes, R. F. 1980. Prediction of energy digestibility of forages with in uitro rumen fermentation and fungal enzyme systems. In : Standardization of Analytical Methodology for Feeds. W. J. Pigden, C. (2. Balch and M. Graham (eds). International Development Research Centre, Ottawa, Canada, pp. 61-7 1.
Mertens, D. R. and Loften, J. R. 1980. The effect of starch on forage fiber digestion kinetics in uitro. J. Dairy Sci., 67 : 1437-1446.
Robertson, J. B., and Van Soest, P. J . 1981. The detergent system of analysis and its application to human foods. In : The analysis of dietiiry fiber in food W. P. T. James and 0. Theander (eds) Marcel Dekker Inc, New York. pp. 123-156.
Statistical Analysis System Institute. 1982. SAS User’s Guide : Statistics. SAS Institute, Cary, NC, 584 pp.
Tanner, G. R. and Morrison, I. M. 1983. Phenolic-carbohydrate complexes in the cell wall of Lolium perenne. Phytochem., 22 : 1433-1439.
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015

30 D, J. R. C h e w y and coworkers
Dow
nloa
ded
by [
Purd
ue U
nive
rsity
] at
07:
57 2
0 Ju
ly 2
015





























