Embed Size (px)
Citation preview

of March 13, 2016.This information is current as
BLT1-Deficient MiceInhibited Aortic Aneurysm Formation in
and Robert E. GersztenShen, Anna R. Cruz, Masanori Aikawa, Andrew D. LusterPruitt, Thomas J. T. Anderson, Fjoralba Kristo, Dongxiao Neil Ahluwalia, Alexander Y. Lin, Andrew M. Tager, Ivy E.
http://www.jimmunol.org/content/179/1/691doi: 10.4049/jimmunol.179.1.691
2007; 179:691-697; ;J Immunol
Referenceshttp://www.jimmunol.org/content/179/1/691.full#ref-list-1
, 17 of which you can access for free at: cites 34 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on March 13, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on March 13, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

Inhibited Aortic Aneurysm Formation in BLT1-Deficient Mice1
Neil Ahluwalia,*†‡ Alexander Y. Lin,*†‡ Andrew M. Tager,*† Ivy E. Pruitt,*†
Thomas J. T. Anderson,*† Fjoralba Kristo,*† Dongxiao Shen,*† Anna R. Cruz,*†
Masanori Aikawa,†§ Andrew D. Luster,*† and Robert E. Gerszten2*†‡�
Leukotriene B4 is a proinflammatory lipid mediator generated by the enzymes 5-lipoxygenase and leukotriene A4 hydrolase.Leukotriene B4 signals primarily through its high-affinity G protein-coupled receptor, BLT1, which is highly expressed on specificleukocyte subsets. Recent genetic studies in humans as well as knockout studies in mice have implicated the leukotriene synthesispathway in several vascular pathologies. In this study, we tested the hypothesis that BLT1 is necessary for abdominal aorticaneurysm (AAA) formation, a major complication of atherosclerotic vascular disease. Chow-fed Apoe�/� and Apoe�/�/Blt1�/�
mice were treated with a 4-wk infusion of angiotensin II (1000 ng/min/kg) beginning at 20 wk of age, in a well-established murineAAA model. We found a reduced incidence of AAA formation as well as concordant reductions in the maximum suprarenal/infrarenal diameter and total suprarenal/infrarenal area in the angiotensin II-treated Apoe�/�/Blt1�/� mice as compared with theApoe�/� controls. Diminished AAA formation in BLT1-deficient mice was associated with significant reductions in mononuclearcell chemoattractants and leukocyte accumulation in the vessel wall, as well as striking reductions in the production of matrixmetalloproteinases-2 and -9. Thus, we have shown that BLT1 contributes to the frequency and size of abdominal aortic aneurysmsin mice and that BLT1 deletion in turn inhibits proinflammatory circuits and enzymes that modulate vessel wall integrity. Thesefindings extend the role of BLT1 to a critical complication of vascular disease and underscore its potential as a target forintervention in modulating multiple pathologies related to atherosclerosis. The Journal of Immunology, 2007, 179: 691–697.
A mounting body of correlative evidence has linked theleukotriene synthesis pathway to several chronic inflam-matory conditions, including atherosclerosis. 5-Lipoxy-
genase (5-LO)3 and its derivatives are highly expressed withinhuman carotid, aortic, and coronary artery plaques (1–3). Further-more, genetic studies have associated particular variants of 5-LO,its accessory protein 5-LO-activating protein, as well as the en-zyme leukotriene A4 (LTA4) hydrolase with stroke and myocardialinfarction in humans (4–6).
Functional studies in murine models have begun to address therole of bioactive lipid products in atherogenesis and related com-plications, including abdominal aortic aneurysms (AAA; Ref. 7).AAAs are caused by pathological changes in the aortic wall that
lead to segmental weakening, progressive dilation, and spontane-ous rupture (8). Chronic inflammation of the aortic wall and pro-teinase-mediated degradation of structural matrix proteins are be-lieved to contribute to the development of AAA (9), although theprecise molecular mechanisms that underlie vessel wall changesremain poorly understood. Zhao et al. (10) recently demonstratedthat 5-LO plays a striking role in aortic aneurysm formation in acholate-induced model. Interestingly, although 5-LO deficiencyhad a pronounced effect on aortic dilation, no effect was observedon stenotic atherosclerotic lesions, suggesting that the pathologicalprocesses contributing to aneurysm formation vs plaque formationmay be different. 5-LO generates both pro- and anti-inflammatorypathways (11); consequently, the cumulative effects of its inhibi-tion might vary in different pathological contexts. Furthermore, theauthors demonstrated that the cysteinyl leukotriene, LTD4, wassufficient to induce the expression of the chemokines MIP-1� byMM6 monocytic cells and MIP-2 by HUVECs, suggesting that thecysteinyl leukotriene pathway downstream of 5-LO may play afunctional role in aneurysm formation by potentiating inflamma-tory circuits. However, the bioactive lipid(s) downstream of 5-LOnecessary for aortic dilation remain to be definitively identified.
Leukotriene B4 (LTB4) is an eicosanoid lipid mediator gener-ated by a pathway downstream of 5-LO involving LTA4 hydrolase.LTB4 is a proinflammatory mediator that activates multiple leu-kocyte subsets leading to cell recruitment, production of reactiveoxygen species, and induction of gene expression (12–14). LTB4
signals primarily through its high-affinity G protein-coupled recep-tor, BLT1, which is highly expressed in specific subsets of circu-lating peripheral blood leukocytes, as well as on nonleukocytesincluding endothelial cells and smooth muscle cells (SMC; Refs.15 and 16). A low affinity receptor, BLT2, is also ubiquitouslyexpressed at low levels in many human tissues (17, 18). Twogroups have recently demonstrated as much as a 2-fold reductionin atherosclerotic plaques in Apoe�/�/Blt1�/� compound-deficient
*Center for Immunology and Inflammatory Diseases, Massachusetts General Hospi-tal, Charlestown, MA 02129; †Harvard Medical School, Boston, MA 02115; ‡Car-diovascular Research Center, Massachusetts General Hospital, Charlestown, MA02129; §Vascular Medicine and Atherosclerosis Unit, Cardiovascular Division,Brigham and Women’s Hospital, Boston, MA 02115; and �Donald W. Reynolds Car-diovascular Clinical Research Center on Atherosclerosis at Harvard Medical School,Boston, MA 02115
Received for publication December 6, 2006. Accepted for publication April 24, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the National Institutes of Health (to R.E.G. andA.D.L.), the American Heart Association (Grant-in-Aid to R.E.G.), the Donald W.Reynolds Foundation (to R.E.G. and M.A.), and a predoctoral award from the SarnoffCardiovascular Research Foundation (to N.A.).2 Address correspondence and reprint requests to Dr. Robert E. Gerszten at Cardiol-ogy Division and Center for Immunology and Inflammatory Diseases, MassachusettsGeneral Hospital East-8307, 149 13th Street, Charlestown, MA 02129. E-mail ad-dress: [email protected] Abbreviations used in this paper: 5-LO, 5-lipoxygenase; LTA4, leukotriene A4;LTB4, leukotriene B4; LTD4, leukotriene D4; AAA, abdominal aortic aneurysm; AngII, angiotensin II; qPCR, quantitative RT-PCR; MMP, matrix metalloproteinase;SMC, smooth muscle cell.
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on March 13, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

mice as compared with Apoe�/� controls (19), although a role forBLT1 in aneurysm formation has not been previously addressed.
In this study, we tested the hypothesis that BLT1 contributes notonly to atherosclerosis generally, but specifically to AAA forma-tion. This study defines key participants downstream of 5-LO inAAA formation and reveals novel interactions between BLT1 andenzymes that modulate vessel wall stability. In contrast to studiesmodulating the upstream enzyme 5-LO, inhibition of BLT1 hasconcordant effects on both AAA and stenotic plaque formation.These findings extend the role of BLT1 to a critical complicationof vascular disease and underscore its potential as a target forintervention in modulating multiple pathologies related toatherosclerosis.
Materials and MethodsAnimals
Blt1�/� mice (12) were backcrossed 10 times into a C57BL/6J backgroundand subsequently bred with Apoe�/� mice (20) (The Jackson Laboratory).Intercrosses of Apoe�/�/Blt1�/� mice yielded offspring that entered thestudy. Genotyping for BLT1 (12) and Apoe (21) was performed by PCR.All mice were fed a normal chow diet. Institutional approval was obtainedfor all experiments described in this manuscript.
Blood and plasma analysis
A total of 0.5–1.0 ml of blood was obtained from experimental mice byright ventricular puncture upon euthanasia. A small aliquot of blood wasanalyzed for complete blood count (Drew Scientific Group), and the re-mainder was used for enzymatic colorimetric analysis of total cholesteroland triglyceride levels (Roche Diagnostics).
Angiotensin II (Ang II) infusion
Apoe�/� and Apoe�/�/Blt1�/� mice were treated with Ang II (1000 ng/min/kg; Sigma-Aldrich) or vehicle control via ALZET model 2004 osmoticpumps (ALZA Corp.) beginning at 20 wk of age as previously described(8). After 4 wk of infusion, mice were sacrificed for blood collection, andaortas were dissected.
Blood pressure analysis
Systolic blood pressure was measured serially in conscious mice using atail-cuff system (Kent Scientific model XBP1000) during three trainingsessions at baseline and then 1 wk after the placement of the Ang II pump.
Morphometric analysis
Aortas were dissected from mice of the indicated age and pinned on a blackwax board; images were obtained with a Nikon Coolpix camera (Nikon)attached to an inverted microscope as previously described (15, 22). Aorticdiameter and area were determined by IPLab image analysis software (BDBiosciences). Measurements were conducted by a single observer blindedto the genotype and treatment of the mice.
Murine aortic immunohistochemistry
Suprarenal aortas were isolated after perfusion of mice with paraformal-dehyde (30 ml) under physiological pressure and then embedded in OCT.Serial 5-�m sections were cut, and every fifth section was stained with
H&E. Immunohistochemistry was performed with Abs to identify SMCs(�-actin, 1/100; Abcam), macrophages (Mac3, 1/20; BD Pharmingen), andT cells (CD4, 1/50; BD Pharmingen) as previously described (15). 5-LOand LTA4 hydrolase were detected by goat anti-mouse Ab (Santa CruzBiotechnology; dilution, 1/30). MMP2 and MMP9 were detected by rabbitanti-mouse Ab (Chemicon; dilution, 1/25). Negative controls were pre-pared with substitution with an isotype control Ab or nonimmune sera.Stained sections were digitally captured, and quantitative analysis was per-formed with IPLab image analysis software (BD Biosciences). Thresholdcolor analysis by a single observer quantified areas stained for a particularAg and total lesion area. We divided areas stained for a particular Ag bytotal lesion area to account for variability in lesion size.
RNA isolation and quantitative RT-PCR (qPCR)
Total RNA was isolated from suprarenal aortas (between the subclavianartery and the takeoff of the right renal artery) from mice perfused withbuffer RLT (Applied Biosystems) using mechanical homogenization witha rotostator and RNeasy (Qiagen) columns (22). Briefly, after DNase Idigestion, equivalent amounts of RNA from each sample were reversetranscribed using TaqMan reverse-transcription reagents, including oli-go(dT)15, random hexamers, and Multiscribe reverse transcriptase(Applied Biosystems). Quantitative RT-PCR (qPCR) reactions wereconducted with the Multiplex QPCR system as described (22). Ampli-fication plots were analyzed with MX4000 software, version 3.0. Geneexpression was normalized to GAPDH or �2-microglobulin as aninternal control.
LTB4 measurement in aortas
Whole aortas were homogenized in ethanol and centrifuged, and superna-tants were passed over C18 Sep-Pak columns (Waters). After drying withnitrogen gas, the residue was dissolved in an extraction buffer for ELISAassay as per the manufacturer’s recommendations (Neogen).
Isolation and treatment of primary human monocytes
Human monocytes were purified from healthy donors by Ficoll-Hypaquedensity gradient centrifugation at 15°C (lymphocyte separation medium;Organon Teknika) followed by negative selection magnetic bead purifica-tion (Miltenyi Biotech) to �92% purity as previously described (23).Monocytes were stimulated at 37.5°C with vehicle control, 100 nmol/LLTB4 in RPMI 1640, 100 nmol/L LTD4 in RPMI 1640, or LPS (100 ng/ml)in RPMI 1640 for 6 h. Reactions were stopped on ice. RNA extraction fromcells was performed using RNeasy columns, and qPCR reactions per-formed as described above. Secreted proteins in the supernatant were mea-sured using human ELISA kits for MCP-1 (R&D Systems), IL-8 (R&DSystems), and MIP-2 (PeproTech).
Statistical analysis
Data were analyzed with SigmaStat (Systat Software) using Student’s t testand two-way ANOVA as indicated. Aneurysm incidence was analyzed byFisher’s exact test. For the RT-PCR studies, transcript levels were com-pared using one- or two-way ANOVA. When significant differences werefound, Tukey’s post hoc test (honestly significant difference) was used toperform pairwise comparisons of group means. A probability value of�0.05 was considered statistically significant. All data are reported asmean � SEM or SD as indicated.
FIGURE 1. A, BLT1 mRNA expression in suprare-nal aortas as assessed by qPCR. After 4 wk of Ang IItreatment, mice were sacrificed, and aortas were per-fused with buffer RLT and dissected. RNA from su-prarenal aortas was prepared, and qPCR was per-formed as per Materials and Methods. Data reflectn � 3 mice for each genotype and treatment condi-tion. �, p � 0.01. B, LTB4 quantitation in wholemouse aortas of the indicated genotype and treatmentas assessed by tissue ELISA as detailed in Materialsand Methods. Data reflect n � 3 mice for each ge-notype and treatment condition. �, p � 0.05.
692 BLT1 IN AORTIC ANEURYSM FORMATION
by guest on March 13, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

ResultsBLT1 and LTB4 expression in AAAs
All mice were placed on a normal chow diet for 20 wk and thentreated with Ang II (1000 ng/min/kg) or vehicle control via ALZETosmotic pumps for 4 wk, which generates AAA in a reproduciblemanner, mimicking many of the important features of the human dis-ease (8, 24). We first examined the expression of BLT1 in this modelby qPCR of suprarenal mouse aortas, given that Abs to the murineBLT1 receptor are not available (Fig. 1A). Ang II does not typicallygenerate AAA in control C57BL/6J mice, and BLT1 expression waslow in both the untreated and C57BL/6J treated cohorts. In contrast,we found significant up-regulation of BLT1 expression in the aortasof hyperlipidemic Apoe�/� mice, particularly after Ang II treatment.We similarly documented the up-regulation of the BLT1 ligand,LTB4, in the Ang II-treated, hyperlipidemic mice (Fig. 1B).
Aneurysm formation in BLT1-deficient mice
To directly assess the functional role of BLT1 in AAA formation, westudied Blt1�/� mice that were first backcrossed 10 times into aC57BL/6J background and subsequently bred with Apoe�/� mice.Intercrosses of Apoe�/�/Blt1�/� mice yielded offspring that enteredthe study. After Ang II infusion, Apoe�/� and Apoe�/�/Blt1�/� micehad comparable lipid levels, white blood cell counts and blood pres-sures, thus excluding several potential confounding variables (TableI). No aneurysms were detected in our prior studies of Apoe�/� andApoe�/�/Blt1�/� mice without Ang II infusion (15).
Multiple metrics were used to quantify aneurysm formation inApoe�/� and Apoe�/�/Blt1�/� mice. We first assessed the inci-dence of aneurysms in the animal cohorts, defining an aneurysm asa dilation of the suprarenal aorta �1.5-fold the diameter of theinfrarenal aorta, which is entirely spared from enlargement in thismodel (8, 24, 25). When present, aneurysms were located imme-diately above the takeoff of the right renal artery as previouslyreported (8, 24, 25). Representative aortas from each genotype andtreatment arm are shown in Fig. 2. As expected, the vehicle controlhad no effect on the aortas of either genotype. In contrast, weobserved a significant reduction in aneurysm incidence in AngII-treated Apoe�/�/Blt1�/� mice (13 of 27 positive; 48%) as com-pared with Apoe�/� mice (25 of 29 positive; 86%; p � 0.004; Fig.3A). The 86% incidence of Ang II-triggered aneurysm formation inApoe�/� mice in this study was entirely consistent with previousreports (8, 24, 25).
To comprehensively assess the extent of aortic dilation in Apoe�/�
and Apoe�/�/Blt1�/� mice, we also measured the ratio of maximumsuprarenal diameter/infrarenal diameter, as well as the ratio of thesuprarenal area (from the takeoff of the left subclavian artery to the leftrenal artery)/infrarenal area (from the left renal artery to the iliac bi-furcation). As noted peviously, because Ang II infusion does not typ-ically affect the infrarenal aorta, dividing through by the infrarenaldiameter and area standardized for natural variation between mice.We found significant reductions in both the maximum suprarenal/infrarenal diameter (Fig. 3B) and suprarenal/infrarenal area (Fig. 3C)in the Ang II-treated Apoe�/�/Blt1�/� mice as compared with theApoe�/� controls (diameter ratio, 1.75 � 0.22 vs 2.55 � 0.23, n � 12for each genotype, p � 0.02; area ratio, 4.27 � 0.16 vs 5.67 � 0.29,n � 12 for each genotype, p � 0.001). Finally, we also calculated thetwo-dimensional area of aneurysms (when present) in all cohorts ofmice. Even though fewer Apoe�/�/Blt1�/� mouse aortas developedaneurysms, when aneurysms did develop in this genotype they weresmaller than those in the single knockout Apoe�/� control mice (an-eurysm area to infrarenal area, 0.94 � 0.28 vs 2.57 � 0.43, p � 0.01;Fig. 3D). Taken together, these data from multiple assessments sug-gest that BLT1 deficiency protects against AAA formation in thismurine model.
Table I. Characterization of Ang II-treated Apoe�/� andApoe�/�/Blt1�/� mice
Apoe�/� Apoe�/�/Blt1�/�
Weight (g) 27.0 � 2.4 (32)a 25.3 � 1.7 (33)WBC (1000/ml) 7.5 � 0.7 (23) 8.5 � 0.8 (29)Total cholesterol (mg/dl) 460 � 51 (23) 511 � 58 (23)Triglycerides (mg/dl) 110 � 9 (22) 110 � 9 (25)Systolic blood pressure
(mm Hg)127.2 � 5.2 (5) 132.2 � 5.8 (5)
a Numbers in parentheses, number of mice.
FIGURE 2. En face AAA analysis. Apoe�/� and Apoe�/�/Blt1�/� mice were treated with saline vehicle control or Ang II for 4 wk, and then aortas weredissected. Representative data from each genotype and treatment condition are displayed. Apoe�/�� saline (n � 9); Apoe�/�� Ang II (n � 29);Apoe�/�/Blt1�/� � saline (n � 9); Apoe�/�/Blt1�/� � Ang II (n � 27). No aneurysms were previously detected in our prior studies of Apoe�/� andApoe�/�/Blt1�/� mice without Ang II infusion (15).
693The Journal of Immunology
by guest on March 13, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
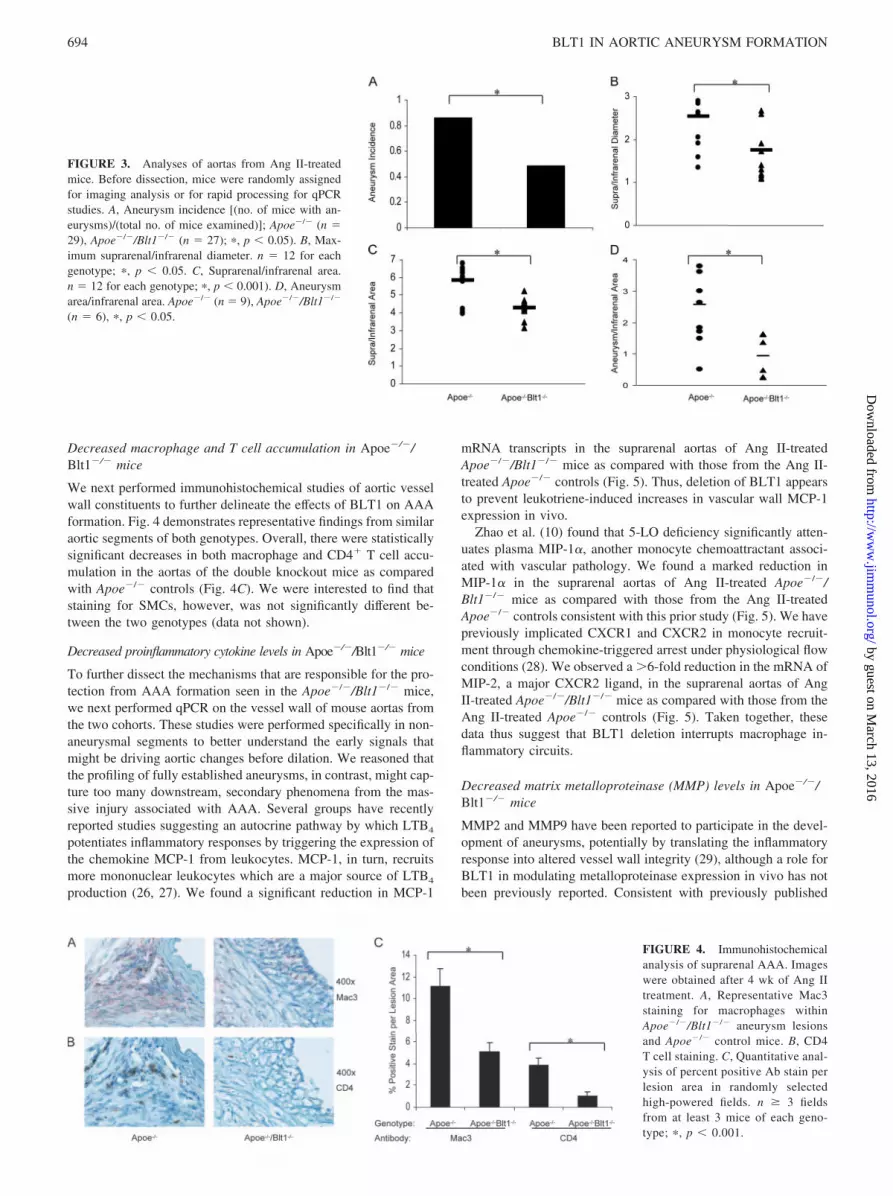
Decreased macrophage and T cell accumulation in Apoe�/�/Blt1�/� mice
We next performed immunohistochemical studies of aortic vesselwall constituents to further delineate the effects of BLT1 on AAAformation. Fig. 4 demonstrates representative findings from similaraortic segments of both genotypes. Overall, there were statisticallysignificant decreases in both macrophage and CD4� T cell accu-mulation in the aortas of the double knockout mice as comparedwith Apoe�/� controls (Fig. 4C). We were interested to find thatstaining for SMCs, however, was not significantly different be-tween the two genotypes (data not shown).
Decreased proinflammatory cytokine levels in Apoe�/�/Blt1�/� mice
To further dissect the mechanisms that are responsible for the pro-tection from AAA formation seen in the Apoe�/�/Blt1�/� mice,we next performed qPCR on the vessel wall of mouse aortas fromthe two cohorts. These studies were performed specifically in non-aneurysmal segments to better understand the early signals thatmight be driving aortic changes before dilation. We reasoned thatthe profiling of fully established aneurysms, in contrast, might cap-ture too many downstream, secondary phenomena from the mas-sive injury associated with AAA. Several groups have recentlyreported studies suggesting an autocrine pathway by which LTB4
potentiates inflammatory responses by triggering the expression ofthe chemokine MCP-1 from leukocytes. MCP-1, in turn, recruitsmore mononuclear leukocytes which are a major source of LTB4
production (26, 27). We found a significant reduction in MCP-1
mRNA transcripts in the suprarenal aortas of Ang II-treatedApoe�/�/Blt1�/� mice as compared with those from the Ang II-treated Apoe�/� controls (Fig. 5). Thus, deletion of BLT1 appearsto prevent leukotriene-induced increases in vascular wall MCP-1expression in vivo.
Zhao et al. (10) found that 5-LO deficiency significantly atten-uates plasma MIP-1�, another monocyte chemoattractant associ-ated with vascular pathology. We found a marked reduction inMIP-1� in the suprarenal aortas of Ang II-treated Apoe�/�/Blt1�/� mice as compared with those from the Ang II-treatedApoe�/� controls consistent with this prior study (Fig. 5). We havepreviously implicated CXCR1 and CXCR2 in monocyte recruit-ment through chemokine-triggered arrest under physiological flowconditions (28). We observed a �6-fold reduction in the mRNA ofMIP-2, a major CXCR2 ligand, in the suprarenal aortas of AngII-treated Apoe�/�/Blt1�/� mice as compared with those from theAng II-treated Apoe�/� controls (Fig. 5). Taken together, thesedata thus suggest that BLT1 deletion interrupts macrophage in-flammatory circuits.
Decreased matrix metalloproteinase (MMP) levels in Apoe�/�/Blt1�/� mice
MMP2 and MMP9 have been reported to participate in the devel-opment of aneurysms, potentially by translating the inflammatoryresponse into altered vessel wall integrity (29), although a role forBLT1 in modulating metalloproteinase expression in vivo has notbeen previously reported. Consistent with previously published
FIGURE 4. Immunohistochemicalanalysis of suprarenal AAA. Imageswere obtained after 4 wk of Ang IItreatment. A, Representative Mac3staining for macrophages withinApoe�/�/Blt1�/� aneurysm lesionsand Apoe�/� control mice. B, CD4T cell staining. C, Quantitative anal-ysis of percent positive Ab stain perlesion area in randomly selectedhigh-powered fields. n � 3 fieldsfrom at least 3 mice of each geno-type; �, p � 0.001.
FIGURE 3. Analyses of aortas from Ang II-treatedmice. Before dissection, mice were randomly assignedfor imaging analysis or for rapid processing for qPCRstudies. A, Aneurysm incidence [(no. of mice with an-eurysms)/(total no. of mice examined)]; Apoe�/� (n �29), Apoe�/�/Blt1�/� (n � 27); �, p � 0.05). B, Max-imum suprarenal/infrarenal diameter. n � 12 for eachgenotype; �, p � 0.05. C, Suprarenal/infrarenal area.n � 12 for each genotype; �, p � 0.001). D, Aneurysmarea/infrarenal area. Apoe�/� (n � 9), Apoe�/�/Blt1�/�
(n � 6), �, p � 0.05.
694 BLT1 IN AORTIC ANEURYSM FORMATION
by guest on March 13, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
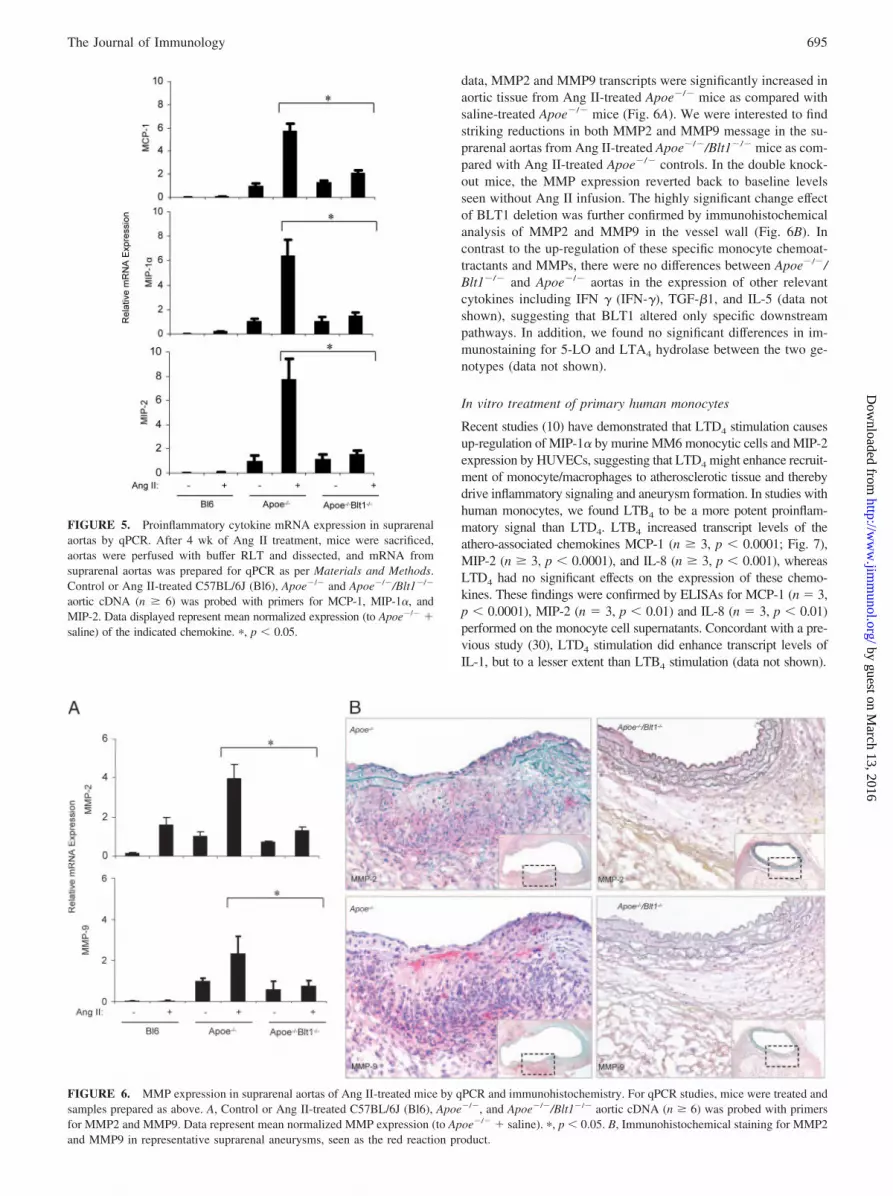
data, MMP2 and MMP9 transcripts were significantly increased inaortic tissue from Ang II-treated Apoe�/� mice as compared withsaline-treated Apoe�/� mice (Fig. 6A). We were interested to findstriking reductions in both MMP2 and MMP9 message in the su-prarenal aortas from Ang II-treated Apoe�/�/Blt1�/� mice as com-pared with Ang II-treated Apoe�/� controls. In the double knock-out mice, the MMP expression reverted back to baseline levelsseen without Ang II infusion. The highly significant change effectof BLT1 deletion was further confirmed by immunohistochemicalanalysis of MMP2 and MMP9 in the vessel wall (Fig. 6B). Incontrast to the up-regulation of these specific monocyte chemoat-tractants and MMPs, there were no differences between Apoe�/�/Blt1�/� and Apoe�/� aortas in the expression of other relevantcytokines including IFN � (IFN-�), TGF-�1, and IL-5 (data notshown), suggesting that BLT1 altered only specific downstreampathways. In addition, we found no significant differences in im-munostaining for 5-LO and LTA4 hydrolase between the two ge-notypes (data not shown).
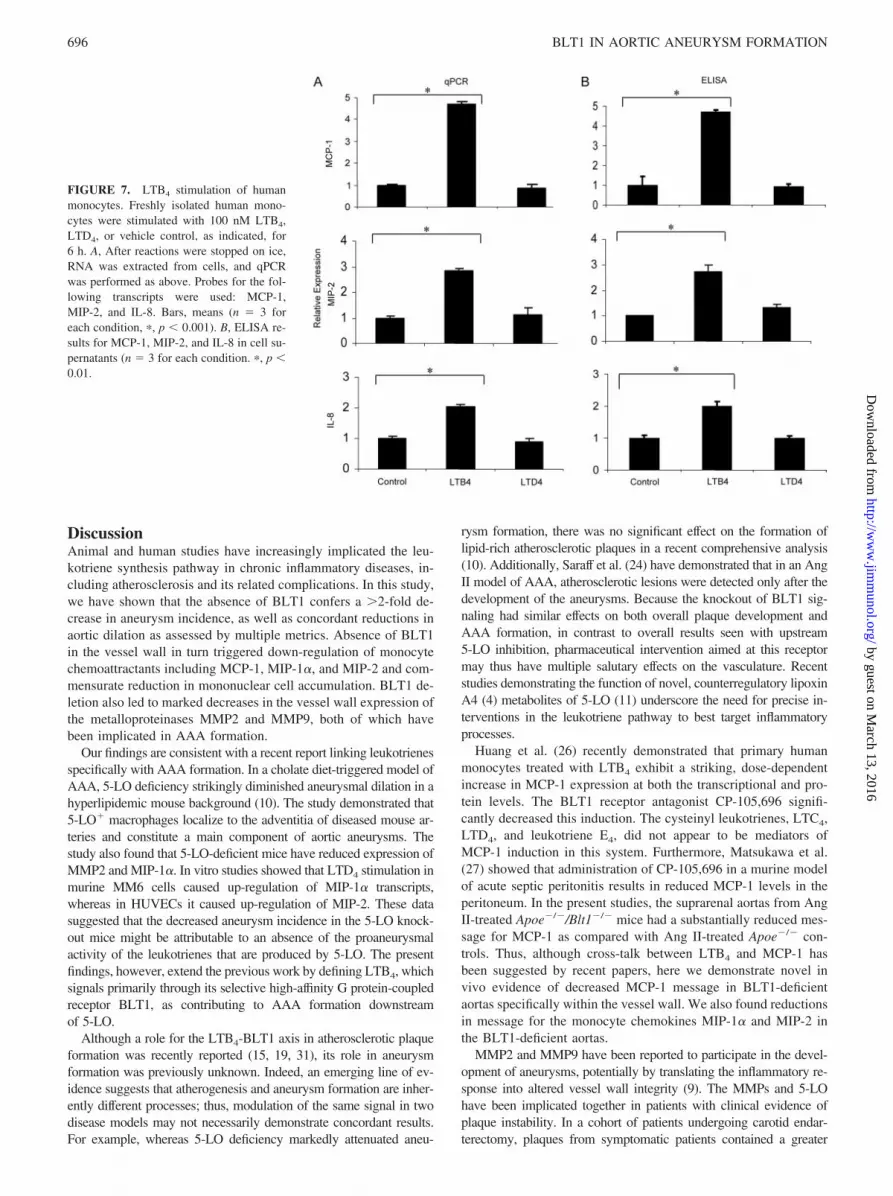
In vitro treatment of primary human monocytes
Recent studies (10) have demonstrated that LTD4 stimulation causesup-regulation of MIP-1� by murine MM6 monocytic cells and MIP-2expression by HUVECs, suggesting that LTD4 might enhance recruit-ment of monocyte/macrophages to atherosclerotic tissue and therebydrive inflammatory signaling and aneurysm formation. In studies withhuman monocytes, we found LTB4 to be a more potent proinflam-matory signal than LTD4. LTB4 increased transcript levels of theathero-associated chemokines MCP-1 (n � 3, p � 0.0001; Fig. 7),MIP-2 (n � 3, p � 0.0001), and IL-8 (n � 3, p � 0.001), whereasLTD4 had no significant effects on the expression of these chemo-kines. These findings were confirmed by ELISAs for MCP-1 (n � 3,p � 0.0001), MIP-2 (n � 3, p � 0.01) and IL-8 (n � 3, p � 0.01)performed on the monocyte cell supernatants. Concordant with a pre-vious study (30), LTD4 stimulation did enhance transcript levels ofIL-1, but to a lesser extent than LTB4 stimulation (data not shown).
FIGURE 5. Proinflammatory cytokine mRNA expression in suprarenalaortas by qPCR. After 4 wk of Ang II treatment, mice were sacrificed,aortas were perfused with buffer RLT and dissected, and mRNA fromsuprarenal aortas was prepared for qPCR as per Materials and Methods.Control or Ang II-treated C57BL/6J (Bl6), Apoe�/� and Apoe�/�/Blt1�/�
aortic cDNA (n � 6) was probed with primers for MCP-1, MIP-1�, andMIP-2. Data displayed represent mean normalized expression (to Apoe�/� �saline) of the indicated chemokine. �, p � 0.05.
FIGURE 6. MMP expression in suprarenal aortas of Ang II-treated mice by qPCR and immunohistochemistry. For qPCR studies, mice were treated andsamples prepared as above. A, Control or Ang II-treated C57BL/6J (Bl6), Apoe�/�, and Apoe�/�/Blt1�/� aortic cDNA (n � 6) was probed with primersfor MMP2 and MMP9. Data represent mean normalized MMP expression (to Apoe�/� � saline). �, p � 0.05. B, Immunohistochemical staining for MMP2and MMP9 in representative suprarenal aneurysms, seen as the red reaction product.
695The Journal of Immunology
by guest on March 13, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
DiscussionAnimal and human studies have increasingly implicated the leu-kotriene synthesis pathway in chronic inflammatory diseases, in-cluding atherosclerosis and its related complications. In this study,we have shown that the absence of BLT1 confers a �2-fold de-crease in aneurysm incidence, as well as concordant reductions inaortic dilation as assessed by multiple metrics. Absence of BLT1in the vessel wall in turn triggered down-regulation of monocytechemoattractants including MCP-1, MIP-1�, and MIP-2 and com-mensurate reduction in mononuclear cell accumulation. BLT1 de-letion also led to marked decreases in the vessel wall expression ofthe metalloproteinases MMP2 and MMP9, both of which havebeen implicated in AAA formation.
Our findings are consistent with a recent report linking leukotrienesspecifically with AAA formation. In a cholate diet-triggered model ofAAA, 5-LO deficiency strikingly diminished aneurysmal dilation in ahyperlipidemic mouse background (10). The study demonstrated that5-LO� macrophages localize to the adventitia of diseased mouse ar-teries and constitute a main component of aortic aneurysms. Thestudy also found that 5-LO-deficient mice have reduced expression ofMMP2 and MIP-1�. In vitro studies showed that LTD4 stimulation inmurine MM6 cells caused up-regulation of MIP-1� transcripts,whereas in HUVECs it caused up-regulation of MIP-2. These datasuggested that the decreased aneurysm incidence in the 5-LO knock-out mice might be attributable to an absence of the proaneurysmalactivity of the leukotrienes that are produced by 5-LO. The presentfindings, however, extend the previous work by defining LTB4, whichsignals primarily through its selective high-affinity G protein-coupledreceptor BLT1, as contributing to AAA formation downstreamof 5-LO.
Although a role for the LTB4-BLT1 axis in atherosclerotic plaqueformation was recently reported (15, 19, 31), its role in aneurysmformation was previously unknown. Indeed, an emerging line of ev-idence suggests that atherogenesis and aneurysm formation are inher-ently different processes; thus, modulation of the same signal in twodisease models may not necessarily demonstrate concordant results.For example, whereas 5-LO deficiency markedly attenuated aneu-
rysm formation, there was no significant effect on the formation oflipid-rich atherosclerotic plaques in a recent comprehensive analysis(10). Additionally, Saraff et al. (24) have demonstrated that in an AngII model of AAA, atherosclerotic lesions were detected only after thedevelopment of the aneurysms. Because the knockout of BLT1 sig-naling had similar effects on both overall plaque development andAAA formation, in contrast to overall results seen with upstream5-LO inhibition, pharmaceutical intervention aimed at this receptormay thus have multiple salutary effects on the vasculature. Recentstudies demonstrating the function of novel, counterregulatory lipoxinA4 (4) metabolites of 5-LO (11) underscore the need for precise in-terventions in the leukotriene pathway to best target inflammatoryprocesses.
Huang et al. (26) recently demonstrated that primary humanmonocytes treated with LTB4 exhibit a striking, dose-dependentincrease in MCP-1 expression at both the transcriptional and pro-tein levels. The BLT1 receptor antagonist CP-105,696 signifi-cantly decreased this induction. The cysteinyl leukotrienes, LTC4,LTD4, and leukotriene E4, did not appear to be mediators ofMCP-1 induction in this system. Furthermore, Matsukawa et al.(27) showed that administration of CP-105,696 in a murine modelof acute septic peritonitis results in reduced MCP-1 levels in theperitoneum. In the present studies, the suprarenal aortas from AngII-treated Apoe�/�/Blt1�/� mice had a substantially reduced mes-sage for MCP-1 as compared with Ang II-treated Apoe�/� con-trols. Thus, although cross-talk between LTB4 and MCP-1 hasbeen suggested by recent papers, here we demonstrate novel invivo evidence of decreased MCP-1 message in BLT1-deficientaortas specifically within the vessel wall. We also found reductionsin message for the monocyte chemokines MIP-1� and MIP-2 inthe BLT1-deficient aortas.
MMP2 and MMP9 have been reported to participate in the devel-opment of aneurysms, potentially by translating the inflammatory re-sponse into altered vessel wall integrity (9). The MMPs and 5-LOhave been implicated together in patients with clinical evidence ofplaque instability. In a cohort of patients undergoing carotid endar-terectomy, plaques from symptomatic patients contained a greater
FIGURE 7. LTB4 stimulation of humanmonocytes. Freshly isolated human mono-cytes were stimulated with 100 nM LTB4,LTD4, or vehicle control, as indicated, for6 h. A, After reactions were stopped on ice,RNA was extracted from cells, and qPCRwas performed as above. Probes for the fol-lowing transcripts were used: MCP-1,MIP-2, and IL-8. Bars, means (n � 3 foreach condition, �, p � 0.001). B, ELISA re-sults for MCP-1, MIP-2, and IL-8 in cell su-pernatants (n � 3 for each condition. �, p �0.01.
696 BLT1 IN AORTIC ANEURYSM FORMATION
by guest on March 13, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

amount of 5-LO, MMP2, and MMP9 than those from asymptomaticpatients (32). Furthermore, immunofluorescence showed that 5-LOand MMPs colocalized in activated macrophages. Our data thereforeextend prior associative studies by demonstrating novel, striking ef-fects of BLT1 deletion on MMPs in the vessel wall in vivo.
Recent studies by our group (15, 33) and others (16) have dem-onstrated the inducible expression of BLT1 on previously unan-ticipated cell types, including SMC. Indeed, in a prior study usinga Western-style diet, there was a significant decrease in SMC ac-cumulation in the atherosclerotic plaques of Apoe�/�/Blt1�/�
mice as compared with the single-knockout Apoe�/� controls (15).In the present study, however, we observed no significant differ-ences in SMC distribution in the AAA. The different findings be-tween the two studies may be due to the different diets or the verydifferent pathology generated by the Ang II infusion and also pro-vide further evidence for differential signals modulating plaqueformation vs AAA formation.
The use of knockout mice in the present studies of course lim-ited the scope of our investigation to interventions that precede theonset of the disease model. Future studies must also addresswhether pharmacological blockade of BLT1 can actually reversethe formation of AAA once present. Indeed, pharmacological in-hibition of the JNK pathway conferred a striking reversal of AAAformation in an animal model (34); analogous studies with BLT1antagonism would further enhance clinical relevance.
In conclusion, our data have extended the role of the LTB4-BLT1 axis as critical to aneurysm formation in mice. As withclassic peptide chemokines, the pleiotropic effects of lipid che-moattractants in disease contexts are increasingly appreciated.These findings are clinically relevant because small-molecule an-tagonists to BLT1 are under development and would thus targetmultiple vessel wall cell types and multiple vascular pathologies.
DisclosuresThe authors have no financial conflict of interest.
References1. Spanbroek, R., R. Grabner, K. Lotzer, M. Hildner, A. Urbach, K. Ruhling,
M. P. Moos, B. Kaiser, T. U. Cohnert, T. Wahlers, et al. 2003. Expanding ex-pression of the 5-lipoxygenase pathway within the arterial wall during humanatherogenesis. Proc. Natl. Acad. Sci. USA 100: 1238–1243.
2. De Caterina, R., A. Mazzone, D. Giannessi, R. Sicari, W. Pelosi, G. Lazzerini,A. Azzara, R. Forder, F. Carey, D. Caruso, et al. 1988. Leukotriene B4 productionin human atherosclerotic plaques. Biomed. Biochim. Acta 47: S182–S185.
3. Qiu, H., A. Gabrielsen, H. E. Agardh, M. Wan, A. Wetterholm, C. H. Wong,U. Hedin, J. Swedenborg, G. K. Hansson, B. Samuelsson, G. Paulsson-Berne, andJ. Z. Haeggstrom. 2006. Expression of 5-lipoxygenase and leukotriene A4 hy-drolase in human atherosclerotic lesions correlates with symptoms of plaque in-stability. Proc. Natl. Acad. Sci. USA 103: 8161–8166.
4. Dwyer, J. H., H. Allayee, K. M. Dwyer, J. Fan, H. Wu, R. Mar, A. J. Lusis, andM. Mehrabian. 2004. Arachidonate 5-lipoxygenase promoter genotype, dietaryarachidonic acid, and atherosclerosis. N. Engl. J. Med. 350: 29–37.
5. Helgadottir, A., A. Manolescu, A. Helgason, G. Thorleifsson, U. Thorsteinsdottir,D. F. Gudbjartsson, S. Gretarsdottir, K. P. Magnusson, G. Gudmundsson,A. Hicks, et al. 2006. A variant of the gene encoding leukotriene A4 hydrolaseconfers ethnicity-specific risk of myocardial infarction. Nat. Genet. 38: 68–74.
6. Helgadottir, A., A. Manolescu, G. Thorleifsson, S. Gretarsdottir, H. Jonsdottir,U. Thorsteinsdottir, N. J. Samani, G. Gudmundsson, S. F. Grant, G. Thorgeirsson,et al. 2004. The gene encoding 5-lipoxygenase activating protein confers risk ofmyocardial infarction and stroke. Nat. Genet. 36: 233–239.
7. Gitlin, J. M., D. B. Trivedi, R. Langenbach, and C. D. Loftin. 2007. Geneticdeficiency of cyclooxygenase-2 attenuates abdominal aortic aneurysm formationin mice. Cardiovasc. Res. 73: 227–236.
8. Daugherty, A., M. W. Manning, and L. A. Cassis. 2000. Angiotensin II promotesatherosclerotic lesions and aneurysms in apolipoprotein E-deficient mice. J. Clin.Invest. 105: 1605–1612.
9. Thompson, R. W., P. J. Geraghty, and J. K. Lee. 2002. Abdominal aortic aneu-rysms: basic mechanisms and clinical implications. Curr. Probl. Surg. 39:110–230.
10. Zhao, L., M. P. Moos, R. Grabner, F. Pedrono, J. Fan, B. Kaiser, N. John,S. Schmidt, R. Spanbroek, K. Lotzer, et al. 2004. The 5-lipoxygenase pathwaypromotes pathogenesis of hyperlipidemia-dependent aortic aneurysm. Nat. Med.10: 966–973.
11. Levy, B. D., G. T. De Sanctis, P. R. Devchand, E. Kim, K. Ackerman,B. A. Schmidt, W. Szczeklik, J. M. Drazen, and C. N. Serhan. 2002. Multi-pronged inhibition of airway hyper-responsiveness and inflammation by lipoxinA4. Nat. Med. 8: 1018–1023.
12. Tager, A. M., J. H. Dufour, K. Goodarzi, S. D. Bercury, U. H. von Andrian, andA. D. Luster. 2000. BLTR mediates leukotriene B4-induced chemotaxis and ad-hesion and plays a dominant role in eosinophil accumulation in a murine modelof peritonitis. J. Exp. Med. 192: 439–446.
13. Borgeat, P., et. al. 1979. Transformation of arachadonic acid by rabbit polymor-phonuclear lymphocytes. J. Biol. Chem. 254: 2643–2646.
14. Ford-Hutchinson, A. W., e. al. 1980. Leukotriene B4, a potent chemokinetic andaggregating substance released from polymorphonuclear lymphocytes. Nature286: 264–265.
15. Heller, E. A., E. Liu, A. M. Tager, S. Sinha, J. D. Roberts, S. L. Koehn, P. Libby,E. R. Aikawa, J. Q. Chen, P. Huang, et al. 2005. Inhibition of atherogenesis inBLT1-deficient mice reveals a role for LTB4 and BLT1 in smooth muscle cellrecruitment. Circulation 112: 578–586.
16. Back, M., D. X. Bu, R. Branstrom, Y. Sheikine, Z. Q. Yan, and G. K. Hansson.2005. Leukotriene B4 signaling through NF-�B-dependent BLT1 receptors onvascular smooth muscle cells in atherosclerosis and intimal hyperplasia. Proc.Natl. Acad. Sci. USA 102: 17501–17506.
17. Yokomizo, T., K. Kato, K. Terawaki, T. Izumi, and T. Shimizu. 2000. A secondleukotriene B4 receptor, BLT2: a new therapeutic target in inflammation andimmunological disorders. J. Exp. Med. 192: 421–432.
18. Tager, A. M., and A. D. Luster. 2003. BLT1 and BLT2: the leukotriene B4
receptors. Prostaglandins Leukot. Essent Fatty Acids 69: 123–134.19. Subbarao, K., V. R. Jala, S. Mathis, J. Suttles, W. Zacharias, J. Ahamed, H. Ali,
M. T. Tseng, and B. Haribabu. 2004. Role of leukotriene B4 receptors in thedevelopment of atherosclerosis: potential mechanisms. Arterioscler. Thromb.Vasc. Biol. 24: 369–375.
20. Plump, A. S., J. D. Smith, T. Hayek, K. Aalto-Setala, A. Walsh, J. G. Verstuyft,E. M. Rubin, and J. L. Breslow. 1992. Severe hypercholesterolemia and athero-sclerosis in apolipoprotein E-deficient mice created by homologous recombina-tion in ES cells. Cell 71: 343–353.
21. Kuhlencordt, P. J., J. Chen, F. Han, J. Astern, and P. L. Huang. 2001. Geneticdeficiency of inducible nitric oxide synthase reduces atherosclerosis and lowersplasma lipid peroxides in apolipoprotein E-knockout mice. Circulation 103:3099–3104.
22. Bjorkbacka, H., V. V. Kunjathoor, K. J. Moore, S. Koehn, C. M. Ordija,M. A. Lee, T. Means, K. Halmen, A. D. Luster, D. T. Golenbock, andM. W. Freeman. 2004. Reduced atherosclerosis in MyD88null mice links elevatedserum cholesterol levels to activation of innate immunity signaling pathways.Nat. Med. 10: 416–421.
23. Gerszten, R. E., E. B. Friedrich, T. Matsui, R. R. Hung, L. Li, T. Force, andA. Rosenzweig. 2001. Role of phosphoinositide 3-kinase in monocyte recruit-ment under flow conditions. J. Biol. Chem. 276: 26846–26851.
24. Saraff, K., F. Babamusta, L. A. Cassis, and A. Daugherty. 2003. Aortic dissectionprecedes formation of aneurysms and atherosclerosis in angiotensin II-in-fused, apolipoprotein E-deficient mice. Arterioscler. Thromb. Vasc. Biol. 23:1621–1626.
25. Bruemmer, D., A. R. Collins, G. Noh, W. Wang, M. Territo, S. Arias-Magallona,M. C. Fishbein, F. Blaschke, U. Kintscher, K. Graf, R. E. Law, and W. A. Hsueh.2003. Angiotensin II-accelerated atherosclerosis and aneurysm formation is at-tenuated in osteopontin-deficient mice. J. Clin. Invest. 112: 1318–1331.
26. Huang, L., A. Zhao, F. Wong, J. M. Ayala, M. Struthers, F. Ujjainwalla,S. D. Wright, M. S. Springer, J. Evans, and J. Cui. 2004. Leukotriene B4 stronglyincreases monocyte chemoattractant protein-1 in human monocytes. Arterioscler.Thromb. Vasc. Biol. 24: 1783–1788.
27. Matsukawa, A., C. M. Hogaboam, N. W. Lukacs, P. M. Lincoln, R. M. Strieter,and S. L. Kunkel. 1999. Endogenous monocyte chemoattractant protein-1(MCP-1) protects mice in a model of acute septic peritonitis: cross-talk betweenMCP-1 and leukotriene B4. J. Immunol. 163: 6148–6154.
28. Gerszten, R. E., E. A. Garcia-Zepeda, Y. C. Lim, M. Yoshida, H. A. Ding,M. A. Gimbrone, Jr., A. D. Luster, F. W. Luscinskas, and A. Rosenzweig. 1999.MCP-1 and IL-8 trigger firm adhesion of monocytes to vascular endotheliumunder flow conditions. Nature 398: 718–723.
29. Longo, G. M., W. Xiong, T. C. Greiner, Y. Zhao, N. Fiotti, and B. T. Baxter.2002. Matrix metalloproteinases 2 and 9 work in concert to produce aortic an-eurysms. J. Clin. Invest. 110: 625–632.
30. Rola-Pleszczynski, M., and I. Lemaire. 1985. Leukotrienes augment interleukin1 production by human monocytes. J. Immunol. 135: 3958–3961.
31. Aiello, R. J., P. A. Bourassa, S. Lindsey, W. Weng, A. Freeman, and H. J.Showell. 2002. Leukotriene B4 receptor antagonism reduces monocytic foamcells in mice. Arterioscler. Thromb. Vasc. Biol. 22: 443–449.
32. Cipollone, F., A. Mezzetti, M. L. Fazia, C. Cuccurullo, A. Iezzi, S. Ucchino,F. Spigonardo, M. Bucci, F. Cuccurullo, S. M. Prescott, and D. M. Stafforini.2005. Association between 5-lipoxygenase expression and plaque instability inhumans. Arterioscler. Thromb. Vasc. Biol. 25: 1665–1670.
33. Tager, A. M., S. K. Bromley, B. D. Medoff, S. A. Islam, S. D. Bercury,E. B. Friedrich, A. D. Carafone, R. E. Gerszten, and A. D. Luster. 2003. Leu-kotriene B4 receptor BLT1 mediates early effector T cell recruitment. Nat Im-munol. 4: 982–990.
34. Yoshimura, K., H. Aoki, Y. Ikeda, K. Fujii, N. Akiyama, A. Furutani, Y. Hoshii,N. Tanaka, R. Ricci, T. Ishihara, et al. 2005. Regression of abdominal aorticaneurysm by inhibition of c-Jun N-terminal kinase. Nat. Med. 11: 1330–1338.
697The Journal of Immunology
by guest on March 13, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from





























