Embed Size (px)
Citation preview

I
SD
a
ARAA
KRPPOFS
1
sfsyofwtfso
bt
0d
Agricultural Water Management 105 (2012) 38– 47
Contents lists available at SciVerse ScienceDirect
Agricultural Water Management
jo u r n al hom ep age: www.elsev ier .com/ locate /agwat
nfluence of artificially restricted rooting depth on soybean yield and seed quality
hardendu K. Singh, Valerio Hoyos-Villegas, James H. Houx III, Felix B. Fritschi ∗
ivision of Plant Sciences, University of Missouri, 1-31 Agriculture Building, Columbia, MO 65211, United States
r t i c l e i n f o
rticle history:eceived 24 May 2011ccepted 28 December 2011vailable online 21 January 2012
eywords:estricted rooting depthslastic linersroteinilatty acideed yield
a b s t r a c t
The amount of plant available soil water is strongly influenced by soil type and rooting depth. This studywas conducted to investigate the influence of limited plant available soil water on soybean (Glycine max(L.) Merr.) yield and seed composition. Six soybean cultivars were grown in 2007, 2008, and 2009 ina field with plastic liners inserted at depths of 0.30, 0.45, 0.60, 0.75, and 0.90 m to limit the rootingdepth and thus the amount of available water. Compared to the long term mean (508 mm), distinctdistribution patterns and amounts of rainfall among the three growing seasons (290, 675, and 440 mmin 2007, 2008 and 2009, respectively) resulted in significant differences in yield and seed compositionamong years. The overall yield, seed weight (g seed−1), oil concentration, linoleic acid and linolenic acidwere the lowest and protein concentration, palmitic acid, stearic acid and oleic acid were the highestin 2007 compared to the other two years. These differences were greater in plants grown under severerooting depth restrictions. Restricted rooting depth affected soybean seed quality such as protein and oilconcentration and fatty acid composition, not only when rainfall was below average, but also when itwas above average. The amount of rainfall received from beginning of pod development through full pod(R3–R4) stages was highly correlated with yield, seed weight, oil and protein. Yield and seed weight werenegatively correlated with protein and positively with oil, and protein and oil were strongly negatively
correlated. Linoleic and linolenic acids were negatively correlated with palmitic, stearic and oleic acids.Under non-limiting moisture conditions (2008), a rooting depth of 0.30 m appeared to provide ampleresources for plant growth, indicating that effects observed in drier years were largely a function ofwater availability. Results presented in this study illustrate that artificially limiting rooting depth underfield conditions may serve as means to manipulate plant-available soil water to study plant responses towater deficit stress without modifying the above-ground environment.. Introduction
Water availability is a major determinant of crop yield. Waterupply is often suboptimal relative to crop water demand, thusrequently limiting crop growth and yield. In fact, water deficittress is considered the most important abiotic stress limiting cropield world-wide (Boyer, 1982). Agriculture is the largest userf fresh water (70%, rivers and ground water) (FAO, 2003), andaces increasing competition from a growing population. Combinedith predicted environmental changes such as altered precipita-
ion patterns (Longenberger et al., 2006), increased future demandor water and food associated with population growth will pose aignificant challenge, thus providing strong impetus for the devel-pment of more drought tolerant crop plants.
Soybean (Glycine max (L.) Merr.) seeds contain high levels ofoth protein (≈42%) and oil (≈23%) and are the primary source ofhe world’s supply of protein and vegetable oil (Conner et al., 2004;
∗ Corresponding author. Tel.: +1 573 882 3023; fax: +1 573 882 1469.E-mail address: [email protected] (F.B. Fritschi).
378-3774/$ – see front matter © 2012 Elsevier B.V. All rights reserved.oi:10.1016/j.agwat.2011.12.025
© 2012 Elsevier B.V. All rights reserved.
Dornbos and Mullen, 1992). Environmental factors such as wateravailability and temperature can drastically influence seed compo-sition (Conner et al., 2004; Dornbos and Mullen, 1992; Rotundo andWestgate, 2009). The reproductive stages (Fehr et al., 1971) frombeginning of flowering (R1) to seed filling (R6) are particularly sen-sitive to water stress (Dogan et al., 2007; Eck et al., 1987; Sweeneyet al., 2003). Eck et al. (1987) reported that water deficit stress fromearly flowering (R1) until the beginning of pod development (R3)reduced seed yields by 9–13%, but when water stress was extendeduntil after full pod stage (R4.5), yields were reduced by 46%. In addi-tion to seed yield, seed composition is also influenced by waterdeficit stress. Rotundo and Westgate (2009) conducted a meta anal-ysis of published information and reported water deficit induceddecreases in total (mg seed−1) protein, oil, and carbohydrate con-tents. These reductions were associated with greater reduction inoil and carbohydrate accumulation in seed as compared to the pro-tein, resulting in an increase in the final protein concentration (%
seed dry weight).Crop plants adjust to water availability by altering their veg-etative and reproductive growth which ultimately affects yieldand seed quality such as concentration and nature of protein, oil

S.K. Singh et al. / Agricultural Water Management 105 (2012) 38– 47 39
F of thr
asftudpebuyatiad2fitigdndavs
aosawmssbpabibseia
ig. 1. Layout of the field site including organization of the replications, orientationestricted rooting depths treatments.
nd fatty acids (Rotundo and Westgate, 2010). Under water deficittress, selection for physiological and biochemical traits that con-er adaptation to drought could complement breeding programshat are mainly based on direct selection for grain yield. Detailednderstanding of the mechanisms underlying tolerance to watereficit stress is necessary for trait based selection in breedingrograms aimed at developing drought tolerant genotypes. How-ver, a major difficulty and cost for physiological research andreeding of drought tolerant cultivars in humid regions is thenpredictability of drought stress, and the imposition of stressear after year to allow direct selection for yield. Because of this,s well as the complexity of drought research under field condi-ions, much of the research on plant adaptation to drought stresss conducted in controlled environments. While such studies havedvanced our understanding of the fundamental principles, resultso not always translate directly to field situations (Samarah et al.,004). In environments where drought is a predictable occurrence,eld experiments can be conducted using supplemental irrigationo ensure a desired range of water stress conditions. However,n regions where the climate is either mostly favorable for croprowth or unpredictable, field research on water deficit stress isifficult. Fields with limited water holding capacity, soil uniformity,on-limiting nutrient availability and with a reasonable chance of arought are required, but not readily available to many researchersnd breeders (Pathan et al., 2007). Therefore, field facilities that pro-ide additional control and/or increased likelihood of water deficittress can prove useful and cost effective.
Various field techniques have been developed to restrict plantccess to water. Movable rainout shelters have been used to carryut drought experiments (Arkin et al., 1976). Factors such as con-truction costs along with maintenance expenses and restrictedrea covered are significant drawbacks. Other methods such asater table management (Mejia et al., 2000) and managed soiloisture systems (Reetz et al., 1979) aim either to restrict the
oil water from reaching the rooting zone or restrict root access tooil water by imposing mechanical impedance (Young et al., 1997),uried mesh layers in the soil profile (McKenzie et al., 2009) or rootruning (Ma et al., 2008). Each system tends to have limitationsssociated with its design, including aspects such as stress causedy root pruning, soil physical strength, and root system plastic-
ty leading to selective penetration of roots through mechanicalarriers, thus causing inconsistencies in response to drought. The
ystem employed in this study consists of impermeable plastic lin-rs buried horizontally at different depths in the soil to completelympede roots from accessing water deeper in soil profile, and maylso be fraught with similar limitations, thus restricting its utility toe rows, arrangement and dimensions of the undisturbed plots and buffers, and the
study plant adaptations to water deficit stress. The main objectivesof this study were to test the utility of this system with varying soilprofile depths imposed by artificial restricted rooting depth (RRD),and to examine the influence of the series of RRDs on soybean yieldand seed quality.
2. Materials and methods
A field study was conducted at the Bradford Research and Exten-sion Center, Columbia, Missouri USA (38◦ 53′N, 92◦ 12′W) in 2007,2008 and 2009.
2.1. Field description
The system used for this study was designed to alter the amountof plant available soil water by confining the root system to spe-cific depths, and was constructed in 1978 as described by Griffin(1980). In brief, a series of five parallel channels 61 m in length and7.3 m wide were excavated to depths of 0.30, 0.45, 0.60, 0.75 and0.90 m (hereafter referred to as rooting depths or treatments) inthe direction of the slope (≈1.2%, East to West) of the field (Fig. 1).The bottom of each channel was lined with plastic, and drain tiles(2′′ Turf Flow pipe, Hancor Inc., Findlay, OH, USA) were installedon top of the plastic liner parallel to the slope to avoid water log-ging. Channels were filled with thoroughly mixed top soil (MexicoSilt Loam [fine, smectitic, mesic, Vertic Epiaqualf]) that was care-fully repacked as the soil was added. To prevent interference, 2.5 mwide strips were left undisturbed between treatments. In addition,areas of 61 by 11 m remained undisturbed as check plots (con-trol treatment, thus there were six treatments) along the sides ofthe shallowest and deepest channels. These check plots allow forcomparisons between crops grown on an undisturbed, natural soilprofile and the channels whose upper soil horizons were excavatedand then used to refill the plot thus creating a disturbed profile.
2.2. Crop management
Six cultivars ranging in maturity group (MG) from 3.0 to 3.9were planted in 2007, 2008 and 2009. Cultivars ‘Pioneer 93M11′
(MG 3.1), ‘Asgrow 3705′ (MG 3.7), ‘Asgrow 3905′ (MG 3.9), and‘Pioneer 93M90′ (MG 3.9) hereafter referred to as C1, C2, C3 and
C4, respectively, were planted in all three years. Cultivar ‘DeKalb36–52’ (MG 3.6) was planted in 2007 and 2008 and replaced with‘DeKalb 38–52’ (MG 3.8) in 2009, hereafter referred to as C5. In 2008and 2009, ‘Merschman Kennedy 836RR’ (MG 3.6) that was grown in
40 S.K. Singh et al. / Agricultural Water M
Fig. 2. Weather data including mean temperature (a), rainfall (b), evapotranspira-tion (c), and vapor pressure deficit (d) for 2007, 2008, 2009 and long-term averages(Cd
2t
2sWwflrlmmef
2
osto(cas
1971–2002) for the growing seasons at Bradford Research and Extension Center,olumbia, Missouri USA (38◦ 53′N, 92◦ 12′W). Soybean growth stages (M, maturity),ays after planting (DAP) and the amount of the rainfall received are also shown (b).
007, was replaced with ‘Asgrow 3005’ (MG 3.0), hereafter referredo as C6.
Soybean seeds were no-till sown to a depth of 25 mm on May2 of 2007, May 21 of 2008 and May 23 of 2009. Each cultivar wasown in 6 rows spaced 0.38 m apart at a density of 45 seeds m−2.
ithin each rooting depth, four replications of the six cultivarsere completely randomized. The randomization was the same
or all rooting depths since planting was conducted perpendicu-arly across all channels and control plots at the same time. Theesulting individual experimental units were 6 rows wide and 7.3 mong. Weed control was performed chemically (Roundup®) and
anually each year as needed. No pest control was required. Toinimize differences as a result of differential availability of nutri-
nts between rooting depths, soil analyses were conducted and soilertility remained high during the study.
.3. Weather data
For each growing season, rainfall, temperature, and evap-transpiration (ET) data were obtained from a weathertation located within 500 m of the field (Fig. 2). The longerm record between 1971 and 2000 for this region wasbtained from the High Plains Regional Climate Center
HPRCC) website (http://www.hprcc.unl.edu/cgi-bin/cli perl lib/liMAIN.pl?mo1791; Columbia WSO AP, MO station # 231791,ccessed on May 20, 2011). The data for temperature and ET weremoothed by running average, a local smoothing technique thatanagement 105 (2012) 38– 47
averages values at neighboring points, using statistical softwareSigmaPlot (v. 11, Systat Software inc., Chicago, IL, USA).
2.4. Yield and yield components
Grain harvests were performed on end-trimmed plots with asmall plot combine (Wintersteiger America Inc., Lincoln, NE, USA)when all plots had reached harvest maturity. Seed weight and seedmoisture content were measured (M-20 P Grain Moisture Tester,Dickey-John Corp., Auburn, IL, USA) and final seed weight wasadjusted to 13% moisture. Individual seed weight was determinedby randomly selecting 500 seeds from each replication.
2.5. Seed protein, oil and fatty acid analysis
A subsample of 25 g of seed was randomly withdrawn fromeach replication for analysis of seed composition. Protein andoil concentrations were determined by non-destructive infraredspectroscopy at the National Center for Agricultural UtilizationResearch (NCAUR) USDA-ARS facility, Peoria, IL. Fatty acid concen-trations (C16:0 Palmitic, C18:0 Stearic, C18:1 Oleic, C18:2 Linoleic& C18:3 Linolenic) of the total seed oil from three seeds sam-pled randomly from each 25-g subsample were determined by gaschromatography (GC) at the same facility. Seeds were hammer-cracked manually (not pulverized) and the oil was extracted bysoaking overnight in closed vials containing 1.5 ml of a mixtureof CHCl3:Hexane:MeOH (8:5:2, v:v:v). Then, 0.1 ml of sodiummethoxide (∼0.35 M) solution and 0.5–1.0 ml of the extraction fluidwere added and vials placed into the GC auto-sampler. Sampleswere analyzed on a GC (Agilent 6890 GC with an Agilent J&WGC column, Quantum Analytics, Inc., Foster City, CA) equippedwith a flame ionization detector (320 ◦C, H2 @ 35 mL/min, air@ 400 mL/min, makeup gas of N2 @ 25 mL/min). The area per-centage of the five main peaks (Palmitic acid, Stearic acid, Oleicacid, Linoleic acid and Linolenic acid) was determined and thefive peaks were normalized so that the area percentage added upto 100.
2.6. Statistical analysis
The experimental design was a completely randomized blockwith four replications of the six cultivars in each rooting depth.All statistical analyses were performed using SAS as describedbelow (SAS-Institute-Inc., 2004). Although cultivars were repli-cated four times within depth, each rooting depth was onlyrepresented by a single, non-replicated channel. Therefore, due tothe field setup, a true replication of rooting depth is lacking, thusrestricting the application of conventional statistical tests. Hence,two-tailed t-tests were performed using the pdiff = control option inthe LSMEANS statement of PROC MIXED. This procedure examinesif any treatments are significantly different from the control, anddetermines the magnitude and direction of these differences for allmain effect means (Mejia et al., 2000; SAS-Institute-Inc., 2004). Theresponse of genotypes across years and their interactions with eachrooting depth was tested using PROC MIXED. Treatment and geno-type were treated as fixed effects and replications nested withinyears as random effect. Regression and correlation analyses wereperformed between rainfall across years and measured variablesusing PROC REG and PROC CORR.
3. Results
3.1. Weather conditions
Weather conditions during the crop cycle varied consid-erably among the years (Fig. 2). In comparison to the long

S.K. Singh et al. / Agricultural Water Management 105 (2012) 38– 47 41
Fig. 3. Seed yield (left) and seed weight (right) of soybean cultivars grown at different rooting depths for three years. Bars indicate mean ± standard error from four replications.T ing dit 5, ress ltivar
t(strepsadat(aaR(
3
(raaarMip
he two-tailed t-test mean comparisons of all rooting depths against control showhe figures with ***, **, * and NS representing P ≤ 0.001, P ≤ 0.01, P ≤ 0.05 and P > 0.0hown at the bottom of the figure. The Y and C represent the effects of years and cu
erm (1971–2000) mean temperature (20.9 ◦C) and total rainfall508 mm), the mean temperatures during the soybean growingeasons of 2007, 2008 and 2009 were 22.2, 20.4, and 19.6 ◦C, respec-ively, and the total rainfall amounts were 290, 675, and 440 mm,espectively. Among the three years, the 2007 season had the low-st rainfall amounts and highest evapotranspiration and vaporressure deficits, which lead to the most pronounced water deficittress conditions during the study duration. In 2008, above aver-ge precipitation did not allow development of significant watereficits stress. While the 2007 rainfall was below long term aver-ges throughout the season, precipitation in 2009 was below longerm conditions only during late reproductive (R6 and later) stagesFig. 2b). The total rainfall received between seven unfolded trifoli-te leaves (V7) and R6 in 2009 was slightly higher than in 2007 (162s compared to 155 mm), while the total rainfall received between1 and R6 in 2009 was lower (73 mm) as compared to 2007 (82 mm)Fig. 2b).
.2. Grain yield
Distinct environmental conditions resulted in significantP < 0.01) differences in grain yield among the three years at allooting depths (Fig. 3). The differences in environmental conditionsmong the three years are clearly revealed by the average yieldscross treatments of 0.84 Mg ha−1 in 2007, 4.59 Mg ha−1 in 2008,nd 3.57 Mg ha−1 in 2009. However, cultivar grain yields within
ooting depth did not consistently differ across all depths and years.oreover, reduced rooting depths tended to reduce soybean yieldn 2007 and 2009 averaged over cultivars. In 2007, the mean com-arison test of all rooting depths against the control (two-tailed
fferences of means against control along with the significance levels are shown inpectively. Results from within rooting depth analysis of variance (Anova) are alsos, respectively within each rooting depth analysis of variance.
test) were highly significant (P < 0.001) and showed marked reduc-tion in yield as rooting depth decreased from 0.90 m (27%) to 0.30 m(76%), averaged over cultivars. In 2008, the yields in all rootingdepth treatments were greater (P < 0.001) than the control exceptat 0.30 m. However, in 2009, yields in the 0.90, 0.45, and 0.30 mtreatments were lower (P < 0.001; 18%, 9%, and 38%, respectively)than those of the control whereas yields 0.75 and 0.60 m treatmentswere not different (P > 0.05) from the control.
3.3. Seed weight
Seed weight measured as mass per seed was strongly influenced(P ≤ 0.01) by the growing seasons and cultivars at all rooting depths.Further, significant year × cultivar interactions were observed ateach rooting depth (Fig. 3). Seed weight generally increased as yielddecreased within a given year. The overall individual seed weightin 2007 was approximately 45% lower compared to 2008 and 2009.In 2007, averaged over cultivars, seed weight was reduced 9–18%(P < 0.001) for rooting depths of 0.90, 0.75, and 0.60 m compared tothe control. In contrast, seed weight was 30% greater in the 0.30 mrooting depth than the control across all cultivars. In 2008, seedweight increased 6–10% for all rooting depths compared to the con-trol, following a similar trend as was observed for grain yield. In2009, no effect of rooting depth on seed weight was observed.
3.4. Protein concentration
Seed protein concentration was strongly influenced by year, cul-tivar, and their interaction (P < 0.01) at each rooting depth (Fig. 4).Across all treatments, seed protein concentration was highest in

42 S.K. Singh et al. / Agricultural Water Management 105 (2012) 38– 47
F wn ati
2rtaeca
3
ieodttiprci
3
yoiaao
ig. 4. Seed protein (left) and oil concentrations (right) of soybean cultivars gronformation.
007, averaging 43% as compared to 39% and 40% in 2008 and 2009,espectively. Rooting depth treatments did not influence seed pro-ein concentration in the same manner across all years. In 2007nd 2008, the two shallowest rooting depths (0.30 and 0.45 m)xhibited lower seed protein concentrations (P < 0.001) than theontrol. In contrast, in 2009, seed protein concentration was greatert 0.30 m rooting depth than the control (P < 0.001).
.5. Oil concentration
As for protein concentration, seed oil concentration was alsonfluenced by year, and cultivar, and their interaction (P < 0.01) atach rooting depth (Fig. 4). However, compared to protein, seedil concentration responded conversely across years and rootingepths. The overall oil concentration was lower in 2007 (16.7%)han in 2008 (20.4%) and 2009 (20.5%). In 2007, seed oil concen-ration decreased as the rooting depth decreased until 0.60 m andncreased thereafter as compared to the control. In 2008, com-ared to the control, higher oil concentration was observed at everyooting depth except 0.90 m. In contrast to 2007 and 2008, oil con-entration in the 0.30 m rooting depth was lower than the controln 2009.
.6. Fatty acid composition
Results from fatty acid analyses are presented in Fig. 5.ear × cultivar interactions were not significant (P > 0.05) for mostf the rooting depths. However, fatty acid composition was strongly
nfluenced by year (generally P < 0.01) and was often also differentmong cultivars. The overall concentrations of palmitic acid (12.6%)nd stearic acid (4.3%) were highest in 2007 as compared to thether two years. Rooting depth had either no effect (P > 0.05) ordifferent rooting depths for three years. Please refer to the Fig. 3 for additional
tended to decrease palmitic acid or increase stearic acid percentageacross years with few exceptions in 2007.
3.7. Relationship between the amount of rainfall at differentsoybean growth stages and, yield, seed weight, and protein and oilconcentrations
Linear regression analyses of rainfall amounts for defined devel-opmental stages and all measured parameters showed that thecoefficients of determination (R2) were highest for the pod devel-opment stages (R3–R4) as compared to other reproductive and latevegetative stages (Fig. 6). These results were consistent across allthe rooting depths, except at 0.30 m for protein and oil (Fig. 6).However, protein and oil exhibited the highest R2 at 0.30 m depthfor all developmental stages except R3–R4. Averaged across rootingdepths, the R2 values at the R3–R4 stage were 0.84 for yield, 0.63for seed weight, 0.57 for protein, and 0.49 for oil when regressedwith the amount of rainfall during the corresponding period.
3.8. Relationships among the measured parameters
The Pearson’s correlation matrix (r) developed using three yearsof data from six rooting depths and six cultivars indicated signifi-cant positive or negative correlations among many of the measuredparameters (Table 1). Protein was inversely correlated with yield(r = −0.75; P < 0.001), seed weight (r = −0.83; P < 0.001) and oil(r = −0.87; P < 0.001). In contrast, oil showed a positive correlationwith yield (r = 0.72; P < 0.001) and seed weight (r = 0.88; P < 0.001).
Polyunsaturated fatty acids (linoleic and linolenic) were positivelycorrelated with yield, seed weight and oil, and were negativelycorrelated with protein. However, palmitic, stearic, and oleic acidsshowed the opposite trend.
S.K. Singh et al. / Agricultural Water Management 105 (2012) 38– 47 43
F rs groi
afttt
w(id
4
4
4
irftjwc0rmdrA
ig. 5. Seed palmitic acid (16:0) and stearic acid (18:0) percentage of soybean cultivanformation.
Linear regression analyses between protein and oil evaluated forll rooting depths revealed coefficients of determination rangingrom 0.49 to 0.84 (Fig. 7). Slopes of the curves were steepest at thewo shallowest rooting depths and flattest at the 0.75 m depth. Also,he range in the data points was smaller in the shallower comparedo the deeper rooting depths.
The changes in seed weight across years and rooting depthsere exponentially related to seed oil and protein concentrations
Fig. 8). The oil concentration showed an exponential rise to max-mum (Fig. 8a; r2 = 0.84) whereas protein showed an exponentialecay (Fig. 8b; r2 = 0.72) relationship with seed weight.
. Discussion
.1. Influence of rooting depth
.1.1. Grain yieldThe different amounts and patterns of rainfall received dur-
ng the three growing seasons allowed for the assessment of theestricted rooting depths under a broad range of moisture regimes,rom below average (2007) to above average (2008). Among thehree years, the lowest seed yields were observed in 2007 in con-unction with the lowest rainfall amounts. Particularly in 2007, the
ater deficit stress was magnified by restricted rooting depths,ausing a decline in yield in each restricted rooting depth from.90 m to 0.30 m. In 2008, the season with the highest rainfall andeduced vapor pressure deficits, crop water needs were largely
et by rainfall and thus, plants did not require access to water ineeper soil horizons. This is illustrated by yield data from the 0.30 mooting depth treatment that were not different from the control.verage yields in 2009 were intermediate between 2007 and 2008.
wn at different rooting depths for three years. Please refer to the Fig. 3 for additional
This was likely caused in large part by differences in rainfall pat-terns among the years. While the total rainfall between V7 and R6was similar between 2009 (162 mm) and 2007 (155 mm), approxi-mately 3-fold greater amounts were received in 2009 (60 mm) thanin 2007(22 mm) during R3–R4 stages (Fig. 2b). Considering that thehighest coefficients of determination between the amount of rain-fall and yield were observed at the R3–R4 stage, differences in waterdeficit stress during this period likely contributed to the discrep-ancy in yield between 2007 and 2009. In general, maximum yieldloss occurs when water stress coincides with reproductive stages ofsoybean development (Dogan et al., 2007; Eck et al., 1987; Stegmanet al., 1990). Our results support previous reports indicating thatwater stress during R3–R4 stages is particularly detrimental to soy-bean yield (Eck et al., 1987; Korte et al., 1983; Stegman et al., 1990).For instance, Korte et al. (1983) reported that a single irrigation atthe R1 stage had little effect on seed yield (+20 kg ha−1), whereasa single irrigation at R3–R4 stage drastically increased seed yield(+379 kg ha−1) relative to the non-irrigated field, irrespective ofcultivar or year. The lack of significant yield reduction at the 0.75and 0.60 m rooting depths, as compared to the control treatment in2009, indicates that the plants were able to access stored soil mois-ture as a result of above average rainfall early in the growing season.In contrast, the restricted soil volume in the 0.45 and 0.30 m root-ing depths limited soil moisture reserves and, hence, caused loweryields.
Yield data from 2008 indicate that the artificial restriction of therooting depth to 0.30 m did not reduce yield in comparison to thecontrol treatment. Therefore, a rooting depth of 0.30 m appeared
to provide ample water for plant growth. Gardner (1983) reportedon soil water uptake patterns of different crops and concluded thatsoil water is initially mostly extracted from the near soil surfaceregions where plant roots are most prolific, and that the zone of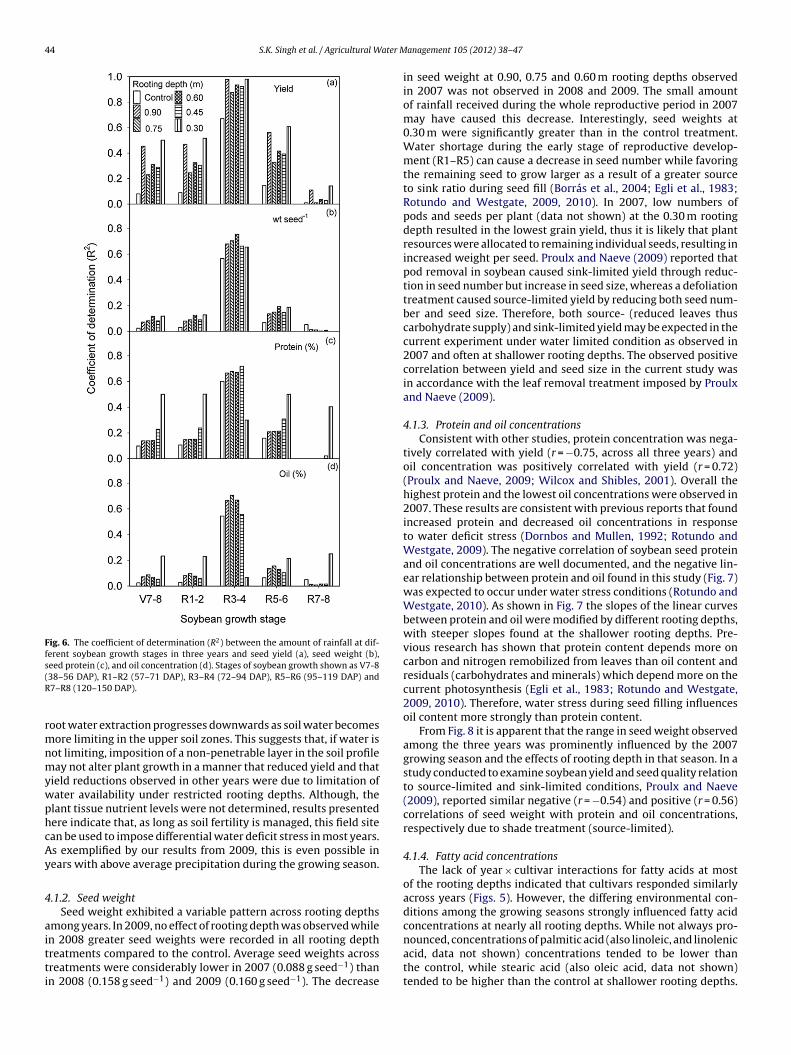
44 S.K. Singh et al. / Agricultural Water M
Fig. 6. The coefficient of determination (R2) between the amount of rainfall at dif-ferent soybean growth stages in three years and seed yield (a), seed weight (b),seed protein (c), and oil concentration (d). Stages of soybean growth shown as V7-8(R
rmnmywphcAy
4
aitti
38–56 DAP), R1–R2 (57–71 DAP), R3–R4 (72–94 DAP), R5–R6 (95–119 DAP) and7–R8 (120–150 DAP).
oot water extraction progresses downwards as soil water becomesore limiting in the upper soil zones. This suggests that, if water is
ot limiting, imposition of a non-penetrable layer in the soil profileay not alter plant growth in a manner that reduced yield and that
ield reductions observed in other years were due to limitation ofater availability under restricted rooting depths. Although, thelant tissue nutrient levels were not determined, results presentedere indicate that, as long as soil fertility is managed, this field sitean be used to impose differential water deficit stress in most years.s exemplified by our results from 2009, this is even possible inears with above average precipitation during the growing season.
.1.2. Seed weightSeed weight exhibited a variable pattern across rooting depths
mong years. In 2009, no effect of rooting depth was observed while
n 2008 greater seed weights were recorded in all rooting depthreatments compared to the control. Average seed weights acrossreatments were considerably lower in 2007 (0.088 g seed−1) thann 2008 (0.158 g seed−1) and 2009 (0.160 g seed−1). The decreaseanagement 105 (2012) 38– 47
in seed weight at 0.90, 0.75 and 0.60 m rooting depths observedin 2007 was not observed in 2008 and 2009. The small amountof rainfall received during the whole reproductive period in 2007may have caused this decrease. Interestingly, seed weights at0.30 m were significantly greater than in the control treatment.Water shortage during the early stage of reproductive develop-ment (R1–R5) can cause a decrease in seed number while favoringthe remaining seed to grow larger as a result of a greater sourceto sink ratio during seed fill (Borrás et al., 2004; Egli et al., 1983;Rotundo and Westgate, 2009, 2010). In 2007, low numbers ofpods and seeds per plant (data not shown) at the 0.30 m rootingdepth resulted in the lowest grain yield, thus it is likely that plantresources were allocated to remaining individual seeds, resulting inincreased weight per seed. Proulx and Naeve (2009) reported thatpod removal in soybean caused sink-limited yield through reduc-tion in seed number but increase in seed size, whereas a defoliationtreatment caused source-limited yield by reducing both seed num-ber and seed size. Therefore, both source- (reduced leaves thuscarbohydrate supply) and sink-limited yield may be expected in thecurrent experiment under water limited condition as observed in2007 and often at shallower rooting depths. The observed positivecorrelation between yield and seed size in the current study wasin accordance with the leaf removal treatment imposed by Proulxand Naeve (2009).
4.1.3. Protein and oil concentrationsConsistent with other studies, protein concentration was nega-
tively correlated with yield (r = −0.75, across all three years) andoil concentration was positively correlated with yield (r = 0.72)(Proulx and Naeve, 2009; Wilcox and Shibles, 2001). Overall thehighest protein and the lowest oil concentrations were observed in2007. These results are consistent with previous reports that foundincreased protein and decreased oil concentrations in responseto water deficit stress (Dornbos and Mullen, 1992; Rotundo andWestgate, 2009). The negative correlation of soybean seed proteinand oil concentrations are well documented, and the negative lin-ear relationship between protein and oil found in this study (Fig. 7)was expected to occur under water stress conditions (Rotundo andWestgate, 2010). As shown in Fig. 7 the slopes of the linear curvesbetween protein and oil were modified by different rooting depths,with steeper slopes found at the shallower rooting depths. Pre-vious research has shown that protein content depends more oncarbon and nitrogen remobilized from leaves than oil content andresiduals (carbohydrates and minerals) which depend more on thecurrent photosynthesis (Egli et al., 1983; Rotundo and Westgate,2009, 2010). Therefore, water stress during seed filling influencesoil content more strongly than protein content.
From Fig. 8 it is apparent that the range in seed weight observedamong the three years was prominently influenced by the 2007growing season and the effects of rooting depth in that season. In astudy conducted to examine soybean yield and seed quality relationto source-limited and sink-limited conditions, Proulx and Naeve(2009), reported similar negative (r = −0.54) and positive (r = 0.56)correlations of seed weight with protein and oil concentrations,respectively due to shade treatment (source-limited).
4.1.4. Fatty acid concentrationsThe lack of year × cultivar interactions for fatty acids at most
of the rooting depths indicated that cultivars responded similarlyacross years (Figs. 5). However, the differing environmental con-ditions among the growing seasons strongly influenced fatty acidconcentrations at nearly all rooting depths. While not always pro-
nounced, concentrations of palmitic acid (also linoleic, and linolenicacid, data not shown) concentrations tended to be lower thanthe control, while stearic acid (also oleic acid, data not shown)tended to be higher than the control at shallower rooting depths.
S.K. Singh et al. / Agricultural Water Management 105 (2012) 38– 47 45
Fig. 7. Relationship between seed protein and oil concentration across three years at different rooting depths: control (a), 0.90 m (b), 0.75 m (c), 0.60 m (d), 0.45 m (e) and0.30 m (f). Mean of four replications for each cultivar in each year are presented.
Table 1Pearson’s correlation coefficients (r) and their level of significance showing the relationships among the measured parameters. Correlation analyses were conducted usingmeans for each rooting depth and cultivar (3 years, 6 rooting depths and 6 cultivar = 108 data points for each parameter). The percentage of fatty acid concentrations areshown as Palmitic (16:0), Stearic (18:0), Oleic (18:1), Linoleic (18:2) and Linolenic (18:3) acid.
Yield Wt seed−1 Protein (%) Oil (%) 16:0 18:0 18:1 18:2
Wt seed−1 0.83***
Protein (%) −0.75*** −0.83***
Oil (%) 0.72*** 0.88*** −0.87***
16:0 −0.55*** −0.65*** 0.55*** −0.71***
18:0 −0.56*** −0.36*** 0.48*** −0.44*** 0.15 NS18:1 −0.31*** −0.24* 0.32*** −0.22* −0.24* 0.23*
18:2 0.38*** 0.33*** −0.44*** 0.39*** 0.04 NS −0.33*** −0.95***
18:3 0.84*** 0.74*** −0.68*** 0.65*** −0.44*** −0.59*** −0.48*** 0.48***
N
Ed(Dcprdo
4s
rtstir
S, not significant (P > 0.05).* Significant level of P < 0.05.
*** Significant level of P < 0.001.
nvironmental conditions, particularly air temperature androught, alter soybean seed quality and fatty acid concentrationDornbos and Mullen, 1992; Karki and Scott, 2008). According toornbos and Mullen (1992) drought had little effect on fatty acidomposition while high temperature reduced the proportion ofolyunsaturated fatty acids. The trend of decreased polyunsatu-ated fatty acid concentrations across years suggests that rootingepth restriction may also alter fatty acid composition in waysther than through reduced soil water availability.
.2. Utility and limitations of the restricted rooting depth studyite
The existing field site allows the assessment of differences inooting depth, albeit imposed artificially by the insertion of a plas-ic liner, under identical climatic conditions. Therefore, the site may
erve as a valuable tool to compare and contrast plant responseso various rooting depths within and across species. The compar-son of multiple soybean cultivars in this study revealed similaresponses to the rooting depth treatments and the associateddifferences in soil water availability. Clearly, the rooting depthtreatments revealed environment-dependent influences on soy-bean each year. For instance, in the driest season (2007) all rootingdepth treatments reduced yield compared to the control. In con-trast, no significant yield reductions in any of the rooting depthtreatments were observed in a season with above average rainfall(2008). However, in 2009, a year with near average but below 2008rainfall, reduced yields were observed at the two shallowest rootingdepth treatments (0.45 m and 0.30 m). Results from this three-yearstudy indicate that the differences in plant available water as influ-enced by the rooting depth treatment were of utmost importance.The influences of artificial restriction of rooting depth on variousgrowth and physiological processes were not investigated in thisstudy and thus remain unclear. However, if yield were used as anintegrative measure of growth processes, the absence of differencesin yield between the control and 0.30 m rooting depth treatment in
2008 would indicate that restriction of rooting depth did not havea major unintended impact. Moreover, the noticeable benefits inyield observed in 2008 under restricted rooting depths other than0.3 m, might have been due to improved conditions resulting from
46 S.K. Singh et al. / Agricultural Water M
Fap
scbettow
credsboeamszc(
ustiutewdyiwcl
ig. 8. Relationship of seed weight and seed oil (a) and protein (b) concentrationcross years. Mean of four replications for each cultivar at each rooting depth areresented for each year.
oil excavation and refilling with topsoil. An additional treatmentonsisting of soil disturbance without the installation of a plasticarrier would have provided an avenue to assess the impact of soilxcavation and refilling per se, but was not included when the sys-em was established. Thus, if soil fertility is maintained high in allreatments, it appears that this study site allows the examinationf plant responses to water deficit stress even in growing seasonsith near average precipitation such as 2009.
Field studies of plant responses to water stress pose large diffi-ulties in regions of significant rainfall. Therefore, studies of plantesponse to water deficit stress are often limited to controllednvironments (greenhouse or growth chambers) or shifted to therier regions of the world. Some methodologies including rainouthelters, water table management, mesh with variable density ofiopores, and managed soil moisture systems have been devel-ped and used (McKenzie et al., 2009; Mejia et al., 2000; Reetzt al., 1979). The field site employed for this study may provide onedditional means to conduct such studies, and is the only of theseethods that completely restricts root access to water in the sub-
oil. This method eliminates the flow of capillary water to the rootone which may improve uniformity of a limited water supply andould also improve the accuracy of water balance determinationsReetz et al., 1979).
Installation of the plastic layer at depth requires removal of thepper soil horizons and thus causes complete destruction of theoil physical structure which cannot be fully accounted for by evenhe most careful repacking process. Thus, differences in soil phys-cal structure between control and rooting depth treatments arenavoidable. A control for this disturbance (disturbance withouthe installation of a barrier) could be used to elucidate how thexcavation and refilling of soil affect plant growth. However, thisas overlooked when the treatments were first established. Whileifferences among rooting depth treatments were observed in aear with above-average rainfall, control over water availability
mposed by differences in soil water holding capacity are limited inet years. In contrast, during dry years, rooting depth treatmentsan be complemented by irrigation treatments to impose additionalevels of water availability.
anagement 105 (2012) 38– 47
5. Conclusions
These results indicate that the experimental site can be used tocontrol water availability to plants. In general, the effects of dif-ferent rooting depths on soybean seed yield and seed compositionwere highly pronounced in the driest year (2007), less pronouncedin a year with near-normal precipitation (2009), and limited inthe wettest year (2008). However, rooting depth affected soybeanseed quality such as protein and oil concentration and fatty acidcomposition not only when rainfall was below average, but alsowhen it was above average. The amount of rainfall received atR3–R4 stage of soybean showed the highest correlation with yield,seed weight, and oil and protein concentrations. Shallower root-ing depths resulted in a larger reduction in oil concentration perunit increase in protein concentration as compared to the control.Seed weight exhibited a strong positive exponential relationshipwith oil and strong negative relationship with protein. Under non-limiting moisture conditions (2008), a rooting depth of 0.30 mappeared to provide ample resources for plant growth, indicatingthat effects observed in drier years were largely a function of wateravailability.
The study site provided an economical way to examine theimpact of rooting depth on soybean yield and seed composition.Data from three growing seasons ranging in rainfall from below-to above-average indicated that water holding capacity in the dif-ferent treatments was the dominant factor influencing those traits.Thus, artificial restriction of rooting depth may serve as a tool toexamine certain plant responses to water deficit stress under fieldconditions even in regions with significant rainfall. In addition,elimination of capillary water movement from the subsoil into therooting zone by a plastic layer, may allow more accurate estimationof the soil moisture component when conducting water balancestudies. Further, with the direct measurements of soil and planthydraulic conditions, a close link between treatment effect andplant growth, and yield, can be established in future experiments.
Acknowledgements
We thank laboratory personnel at National Center for Agricul-tural Utilization Research (NCAUR) USDA-ARS facility, Peoria, IL forthe analysis of seed composition.
References
Arkin, G.F., Ritchie, J.T., Thompson, M., Chaison, R., 1976. A rainout shelter installationfor studying drought stress. Agron. J. 68, 429–431.
Borrás, L., Slafer, G.A., Otegui, M.E., 2004. Seed dry weight response to source-sinkmanipulations in wheat, maize and soybean: a quantitative reappraisal. FieldCrop Res. 86, 131–146.
Boyer, J.S., 1982. Plant productivity and environment. Science 218, 443–448.Conner, T., Paschal, E.H., Barbero, A., Johnson, E., 2004. The challenges and potential
for future agronomic traits in soybeans. AgBioForum 7, 47–50.Dogan, E., Kirnak, H., Copur, O., 2007. Effect of seasonal water stress on soybean and
site specific evaluation of CROPGRO-Soybean model under semi-arid climaticconditions. Agric. Water Manage. 90, 56–62.
Dornbos, D., Mullen, R., 1992. Soybean seed protein and oil contents and fatty acidcomposition adjustments by drought and temperature. J. Am. Oil Chem. Soc. 69,228–231.
Eck, H.V., Mathers, A.C., Musick, J.T., 1987. Plant water stress at various growth stagesand growth and yield of soybean. Field Crops Res. 17, 1–16.
Egli, D.B., Meckel, L., Phillips, R.E., Radcliffe, D., Leggett, J.E., 1983. Moisture stressand N redistribution in soybean. Agron. J. 75, 1027–1031.
FAO, 2003. Agriculture, food and water: A contribution to the World Water Devel-opment Report.
Fehr, W.R., Caviness, C.E., Burmood, D.T., Pennington, J.S., 1971. Stage of developmentdescriptions for soybeans, Glycine max (L.) Merrill. Crop Sci. 11, 929–931.
Gardner, W.R., 1983. Soil properties and efficient water use: an overview. In: Taylor,
H.M., Jordan, W.R., Sinclair, T.R. (Eds.), Limitations to Efficient Water Use in CropProduction. American Society of Agronomy, Crop Science Society of America/SoilScience Society of America, Madison, WI, USA, pp. 45–64.Griffin, J.L., 1980. Quantification of the effects of water stress on corn growth andyield, M.S. Thesis. University of Missouri, Columbia, MO USA, p. 84.

ater M
K
K
L
M
M
M
P
S.K. Singh et al. / Agricultural W
arki, D., Scott, R.A., 2008. Fluctuation in soybean fatty acids in South Dakota envi-ronments. J. New Seeds 9, 303–320.
orte, L.L., Williams, J.H., Specht, J.E., Sorensen, R.C., 1983. Irrigation of soybeangenotypes during reproductive ontogeny. I. Agronomic responses. Crop Sci. 23,521–527.
ongenberger, P.S., Smith, C.W., Thaxton, P.S., McMichael, B.L., 2006. Developmentof a screening method for drought tolerance in cotton seedlings. Crop Sci. 46,2104–2110.
a, S.-C., Xu, B.-C., Li, F.-M., Liu, W.-Z., Huang, Z.-B., 2008. Effects of root pruning oncompetitive ability and water use efficiency in winter wheat. Field Crops Res.105, 56–63.
cKenzie, B.M., Bengough, A.G., Hallett, P.D., Thomas, W.T.B., Forster, B., McNicol,J.W., 2009. Deep rooting and drought screening of cereal crops: a novel field-based method and its application. Field Crops Res. 112, 165–171.
ejia, M.N., Madramootoo, C.A., Broughton, R.S., 2000. Influence of watertable management on corn and soybean yields. Agric. Water Manage. 46,
73–89.athan, M.S., Lee, J.D., Shannon, J.G., Nguyen, H.T., 2007. Recent advances in breed-ing for drought and salt stress tolerance in soybean. In: Jenks, M.A., Hasegawa,P.M., Jain, S.M. (Eds.), Advances in Molecular Breeding Toward Drought and SaltTolerant Crops. Springer, New York, pp. 739–773.
anagement 105 (2012) 38– 47 47
Proulx, R.A., Naeve, S.L., 2009. Pod removal, shade, and defoliation effects on soybeanyield, protein, and oil. Agron. J. 101, 971–978.
Reetz, H.F., Hodges, H.F., Dale, R.F., 1979. Managed soil moisture system for studyingplant water relation under field condition. Agron. J. 71, 861–865.
Rotundo, J.L., Westgate, M.E., 2009. Meta-analysis of environmental effects on soy-bean seed composition. Field Crops Res. 110, 147–156.
Rotundo, J.L., Westgate, M.E., 2010. Rate and duration of seed component accumu-lation in water-stressed soybean. Crop Sci. 50, 676–684.
Samarah, N., Mullen, R., Cianzio, S., 2004. Size distribution and mineral nutrients ofsoybean seeds in response to drought stress. J. Plant Nutr. 27, 815–835.
SAS-Institute-Inc., 2004. SAS/STAT User’s Guide Version 9. 1. 3. SAS-Institute-Inc.,Cary, NC.
Stegman, E.C., Schatz, B.G., Gardner, J.C., 1990. Yield sensitivities of short seasonsoybeans to irrigation management. Irrig. Sci. 11, 111–119.
Sweeney, D.W., Long, J.H., Kirkham, M.B., 2003. A single irrigation to improve earlymaturing soybean yield and quality. Soil Sci. Soc. Am. J. 67, 235–240.
Wilcox, J.R., Shibles, R.M., 2001. Interrelationships among seed quality attributes insoybean. Crop Sci. 41, 11–14.
Young, I.M., Montagu, K., Conroy, J., Bengough, A.G., 1997. Mechanical impedance ofroot growth directly reduces leaf elongation rates of cereals. New Phytol. 135,613–619.





























