Embed Size (px)
Citation preview

IDE�TIFYI�G GE�ES THAT REGULATE SECO�DARY
GROWTH I� POPLAR
A corky mutant reveals a novel regulator of secondary growth and
development in Populus
and
Do shoot apical meristem identity proteins regulate the vascular
cambium? Evidence for a role of CLAVATA1 in the regulation of
secondary growth in Arabidopsis and Populus
by
Michael John Bush
A thesis submitted to the Department of Biology
in conformity with the requirements for
the degree of Master of Science
Queen’s University
Kingston, Ontario, Canada
(September, 2008)
Copyright © Michael John Bush, 2008

ii
Abstract
Plant growth and development is largely controlled in regions of totipotential cells around
the plant body called meristems. The well characterized shoot and root apical meristems
are responsible for vertical growth, in which many key players have been well studied.
Lateral (secondary) growth is controlled by the vascular and cork cambiums, which are
much less understood. A rapid growth of interest in a new model angiosperm tree, poplar,
has facilitated the study of the two cambiums, specifically into their regulation at the
genomic and proteomic levels. This study describes recent work carried out to explore the
genetic regulation of secondary growth in poplar. Two genes have been identified that
were previously not associated with the process of secondary growth. The first, a gene
lacking annotation (FM#2), was identified through the investigation of a mutant from an
activation-tagged population of poplar. This mutant showed aberrant secondary growth,
with an increase in the phloem:xylem ratio. It also developed a thick, rough bark, and was
subsequently named corky. Constructs to recapitulate this phenotype have been produced
to allow the link between the gene FM#2 and the corky phenotype to be firmly
established. The second gene was identified through a reverse genetics strategy to test if
the Arabidopsis shoot apical meristem regulator, CLAVATA1 also played a role in the
regulation of the vascular cambium. When it was downregulated in Arabidopsis, a
significant increase in secondary growth was observed. Antisense and hairpin-RNAi
constructs were produced to attempt downregulation of the gene in poplar using both
traditional Agrobacterium-mediated transformations, and the recently developed strategy
of induced somatic sector analysis.

iii
Co-Authorship
Chapter 2
Preliminary work on the corky project was carried out during my undergraduate honours
thesis project. The mutant was originally identified during this period, as was the
insertion site of the activation tag. Full stem qRT-PCR was also initiated at this time.
Localization of the activation tag was repeated at the onset of my Master's degree to
verify the accuracy of the results. Full stem qRT-PCR data was not used in the production
of this thesis, but rather, all qRT-PCR was repeated using tissue specific isolates to
dissect which tissues were showing any misregulation.
Chapter 3
I hereby acknowledge co-authorship on Chapter 3 of this thesis. Measurements of the
original clv1-1 mutant were carried out by Martha Mullaly. Production of the CLV1-
antisense lines of Arabidopsis and experiments carried out upon them were by Shuyou
Han, as was a first draft of the paper regarding those results. Antisense construct
production and the organization of the first attempt at downregulating PtCLV1 was by
Shuyou Han. Genetic and anatomical analysis of the trees produced during the first
attempt at PtCLV1 downregulation was by Michael Bush. Design, production and
transformation of the antisense and hairpin-RNAi ISSA constructs to downregulate
PtCLV1, PtBAM1, PtBAM1-LIKE and PtBAM3, were carried out by Michael Bush. Final
drafts of the full paper were written by Michael Bush.

iv
Acknowledgements
There are many to thank, but I'll try to be brief:
Thanks to everybody who have helped me out in the Regan Lab. All in all, it has been a
great three years. I'd also like to extend thanks to those who have helped me out in
Queen's Biology in general, especially those in the Snedden and Plaxton labs. The basis
of my academic life has been more affected in the rooms and halls of the third and fifth
floors, than it ever was by time spent in a classroom. I'd like to extend special thanks to
Mr. Jeremy Duguay for patiently answering my millions of questions and reintroducing
me to the chip truck, Mr. Edward Harrison (a.k.a Lex Wily) for being the Go-To-Guy and
Mrs. Barb Vanderbeld for teaching a country music loving 20 year old how to do science.
I'd like to thank my committee members Drs. Wayne Snedden and Bill Plaxton, whose
advice and guidance was always appreciated. It was working in your labs that I had my
first exposure to molecular biology and biochemistry and which has made a great
impression on my professional aspirations.
To my supervisor Dr. Sharon Regan, I'd like to thank you for the chance to work in your
lab with a team of great people and on such interesting projects. The knowledge that I've
gained over the past three years will be instrumental in my future endeavors.
Thanks to my parents and family who have ridden along on the rollercoaster that is my
life. Your support has always meant a lot to me.
Finally, to my wife - who lives with a husband aspiring to be a professional geek - I'd like
to thank you most of all (and also to apologize; jk.).

v
Table of Contents
Abstract ............................................................................................................................... ii
Co-Authorship.................................................................................................................... iii
Acknowledgements............................................................................................................ iv
Table of Contents................................................................................................................ v
List of Figures .................................................................................................................... ix
List of Tables ...................................................................................................................... x
List of Abbreviations ......................................................................................................... xi
Chapter 1 Introduction to the regulation of the vascular cambium .................................... 1
Chapter 2 A corky mutant reveals a novel regulator of secondary growth and
development in Populus...................................................................................................... 7
2.1 Abstract ..................................................................................................................... 7
2.2 Introduction ............................................................................................................... 8
2.3 Materials and Methods ............................................................................................ 13
2.3.1 Plant growth and propagation........................................................................... 13
2.3.2 Anatomical characterization of the corky phenotype ....................................... 14
2.3.3 Localization of the activation tag insertion site................................................ 15
2.3.4 Testing local genes for altered expression using qRT-PCR............................. 15
2.3.5 Sequence confirmation of FM#2 cDNA........................................................... 17
2.3.6 Cloning of poplar FM#2 and Arabidopsis homologs for misexpression studies
................................................................................................................................... 17
2.3.7 SALK lines corresponding to FM#2’s potential Arabidopsis homologs ......... 20

vi
2.3.8 Induced somatic sector analysis as a recapitulation strategy............................ 20
2.4 Results ..................................................................................................................... 21
2.4.1 corky mutant identified in activation-tagged population showing altered
morphology of secondary vasculature. ...................................................................... 21
2.4.2 corky mutant’s single insertion site near three unknown genes ....................... 25
2.4.3 qRT-PCR shows single gene drastically upregulated in corky vascular tissue 27
2.4.4 FM#2 represents a novel gene of unknown function ....................................... 27
2.4.5 Production of misexpression constructs for FM#2 and its Arabidopsis
homologs ................................................................................................................... 33
2.5 Discussion ............................................................................................................... 34
2.5.1 corky mutant identified showing altered morphology in secondary vasculature
................................................................................................................................... 34
2.5.2 FM#2 represents a novel gene of unknown function ....................................... 38
2.5.3 Recapitulation of the corky phenotype to establish gene-phenotype link ........ 40
2.6 Conclusion............................................................................................................... 43
Chapter 3 Do shoot apical meristem identity proteins regulate the vascular cambium?
Evidence for a role of CLAVATA1 in the regulation of secondary growth in Arabidopsis
and Populus....................................................................................................................... 44
3.1 Abstract ................................................................................................................... 44
3.2 Introduction ............................................................................................................. 45
3.3 Materials and Methods ............................................................................................ 50
3.3.1 Plant material, growth and transformation ....................................................... 50

vii
3.3.2 Plasmid construction for production of antisense-CLV1 lines in Arabidopsis . 51
3.3.3 Northern blot analysis....................................................................................... 51
3.3.4 Preparation of the hand section of hypocotyls and stems................................. 52
3.3.5 RNAi construct to downregulate PtCLV1 in poplar ......................................... 52
3.3.6 Induced somatic sector analysis (ISSA) as a transformation strategy.............. 52
3.4 Results ..................................................................................................................... 53
3.4.1 Comparison of the secondary xylem and phloem in the hypocotyl of wildtype
and clv1-1 plants in Arabidopsis ............................................................................... 53
3.4.2 Detection of clv1-1 phenotypes in antisense-CLV1 lines of Arabidopsis ........ 54
3.4.3 Antisense-CLV1 construct inhibits endogenous CLV1 expression ................... 58
3.4.4 Secondary growth in Arabidopsis hypocotyl is elevated by antisense inhibition
of CLV1...................................................................................................................... 59
3.4.5 Comparison of stem radius values between wildtype and antisense lines ....... 62
3.4.6 RNAi to downregulate PtCLV1 in Populus...................................................... 62
3.4.7 Arabidopsis CLV1 has two potential homologs in Populus ............................. 63
3.5 Discussion ............................................................................................................... 66
3.5.1 Identifying clv1-like phenotypes in CLV1-antisense lines in Arabidopsis ....... 67
3.5.2 The vascular cambium of Arabidopsis is negatively regulated by CLV1........ 68
3.5.3 Arabidopsis CLV1 has multiple potential homologs in Populus ...................... 69
3.6 Conclusions ............................................................................................................. 71
Chapter 4 General Discussion........................................................................................... 73
Bibliography ..................................................................................................................... 76

viii
Appendix A Primers used for activation tag localization, RACE and SALK line screening
in corky project. ................................................................................................................ 88
Appendix B Primers used for cloning purposes in corky project ..................................... 89
Appendix C qRT-PCR primers to assay expression of FM#1, FM#2 and FM#3 in the
corky mutant...................................................................................................................... 90
Appendix D Schematic of selected primer locations used in corky project ..................... 91
Appendix E Vector map for ISSA overexpression and antisense constructs. .................. 92
Appendix F Vector map for ISSA hairpin-RNAi constructs ............................................ 93
Appendix G Further anatomical study of the corky mutant.............................................. 94
Appendix H Summary of filtered models within proximity to the corky insertion site. .. 97
Appendix I Reference list of bioinformatic tools utilized in Chapters 2 and 3 ................ 98
Appendix J Athena promoter analysis on Arabidopsis family members of FM#2......... 100
Appendix K Prediction of secondary structure for FM#2. ............................................. 101
Appendix L Miscellaneous primers used in the clavata project..................................... 102
Appendix M Primers used for ISSA construct production in clavata project ................ 103
Appendix N A diagrammatic representation of ISSA amplicons. .................................. 104
Appendix O The radius of xylem and phloem/cortex in the hypocotyls of wildtype (WT)
and CLV1-antisense lines................................................................................................ 105
Appendix P The radius of stems of wildtype (WT) and CLV1-antisense lines. ............. 106
Appendix Q Pileup of Arabidopsis and Populus CLV1 and BAM family members....... 107
Appendix R Model for FM#2 and CLAVATA1 function in secondary growth ............ 111
Appendix S Construct flowchart for corky and clavata projects .................................... 112

ix
List of Figures
Figure 1. The anatomy of wildtype and corky. ................................................................. 24
Figure 2. Graphic representation of the corky insertion site............................................. 26
Figure 3. qRT-PCR results from tissue specific samples showing upregulation of FM2 in
corky xylem, phloem and cortex tissues. ................................................................... 28
Figure 4. Analysis of the Arabidopsis and Populus genes identified as belonging to the
FM#2 gene family. .................................................................................................... 30
Figure 5. Anatomy of original Arabidopsis clv1-1 mutant.. ............................................. 55
Figure 6. Floral, stem and hypocotyl variation between wildtype and CLV1-antisense
lines............................................................................................................................ 57
Figure 7. Comparison of the radii and cross-sectional area of xylem and phloem/cortex
amongst wildtype and CLV1-antisense lines ............................................................. 61
Figure 8. Cladogram showing the relationship between the CLV1 and BAM genes in
Arabidopsis and poplar. ............................................................................................. 65

x
List of Tables
Table 1. Primers used in the identification and characterization of the corky mutant...... 16
Table 2. Cycling conditions qRT-PCR testing of FM#1, FM#2 and FM#3 ..................... 18
Table 3. FM#2 and its potential family members in Arabidopsis and Populus................ 32
Table 4. Shoot apical meristem proteins known to be involved with CLV1, their potential
homologs in poplar and their stem tissue expression patterns................................... 64

xi
List of Abbreviations
717 Populus alba x P. tremula hybrid
α32
P dCTP P32
labelled deoxycytosine triphosphate
ACRE180 Avvr9/Cf9 Rapidly Elicited Protein 180
AGI Arabidopsis Gene Identication Number
apl altered phloem development
ATHB15 Arabidopsis thaliana homeobox 15
ATHB-8 Arabidopsis thaliana homeobox 8
BAM1 BIG APICAL MERISTEM 1
BAM2 BIG APICAL MERISTEM 2
BAM3 BIG APICAL MERISTEM 3
BAR Botany Array Resource
bp base pair
bri1 brassinosteroid-insensitive1
brl1 brassinosteroid-insensitive1-like protein 1
brl3 brassinosteroid-insensitive1-like protein 3
CDD conserved domain database
cDNA complementary DNA
CLE CLAVATA3/ESR RELATED
CLV1 CLAVATA 1
CLV2 CLAVATA 2

xii
CLV3 CLATATA 3
Col Columbia ecotype
cov1 continuous vascular ring 1
cpd constituitive photomorphogenesis and dwarfism
CR5 COR5YE
CTAB cetyl trimethylammonium bromide
DAPI 4',6-diamidino-2-phenylindole
DAS dense alignment surface
DDBJ DNA Data Bank of Japan
DNA deoxyribonucleic acid
dwf7 dwarfy 7
eld1 elogation defective 1
EMBL-EBI European Molecular Biology Lab - European
Bioinformatics Institute
ESR EMBRYO SURROU5DI5G REGIO5
ESTs expressed sequence tag
FM#1 Filtered Model # 1
FM#2 Filtered Model # 2
FM#3 Filtered Model # 3
FQA fiber quality analyzer
GFP GREEN FLUORESCENT PROTEIN
GPAT5 GLYCEROL-3-PHOSPHATE

xiii
ACYLTRA5SFERASE 5
GUS β-glucuronidase
HD-ZIP III Homeodomain leucin zipper class 3 family
hr hour
IBIVU Centre for Integrative Bioinformatics Vrige
University
IFL1 I5TRAFASCICULAR FIBERLESS 1
ISSA induced somatic sector analysis
JGI Joint Genomics Institute
KanR kanamycin resistant
KanS kanamycin sensitive
kb kilobase pairs
KEGG Kyoto Encylopedia of Genes and Genomes
L Linnaeus
LEP LEAFY PETIOLE
Ler Landsberg ecotype
LRR-RLK Leucine-rich repeat receptor-like kinase
mins minutes
MIR116 MicroR5A-116
mm millimeter
mRNA messenger RNA
MS Murashige and Skoog Media

xiv
NCBI National Centre of Bioinformatic Information
ND not detected
ng nanograms
nm nanometer
NV non-vascular
ºC degrees Celsius
ORF open reading frame
P. Populus
PC phloem/cambium
PCR polymerase chain reaction
PhGA2OX2 Phaseolus coccineus GIBBERELLIC ACID 2-
OXIDASE 2
PI Plastochron Index
POL POLTERGEIST
populusDB populus database
PtBAM1 Populus BIG APICAL MERISTEM 1
PtBAM1-LIKE Populus BIG APICAL MERISTEM 1 - LIKE
PtBAM3 Populus BIG APICAL MERISTEM 3
PtCLV1 Poplulus trichocarpa CLAVATA 1
PtCLV1a Populus trichocarpa CLAVATA1a (hypothetical)
PtCLV1b Populus trichocarpa CLAVATA1b (hypothetical)
qRT-PCR quantitative real-time polymerase chain reaction

xv
RACE rapid amplification of cDNA ends
RAM root apical meristem
RNA ribonucleic acid
RNAi RNA-interference
s second
S.E. standard error
SAM shoot apical meristem
SBC Stolkholm Bioinformatics Centre
SIGnAL Salk Institute Genomic Analysis Laboratory
STM SHOOTMERISTEMLESS
TAE tris-acetate ethylenediaminetetraacetic acid
TAIR The Arabidopsis Information Resource
TBO Toluidene Blue O
T-DNA Transfer-DNA
TIGR The Institute for Genomic Research
UBC University of British Columbia
UTR untranslated region
UV ultraviolet
v/v volume per volume
VAS VASCULAR TISSUE SIZE
WOL WOODE5 LEG
WSU Washington State University

xvi
WUS WUSCHEL
X xylem tissue
λ wavelength
µm micrometer

1
Chapter 1
Introduction to the regulation of the vascular cambium
Plant biology now has three different genomic models, as a result of recent sequencing
initiatives. These three models, all angiosperms, include an herbaceous dicot, a monocot
and a woody dicot; Arabidopsis (Airabidopsis thaliana ssp. columbia; (The Arabidopsis
Genome Initiative, 2000)), rice (Oryza sativa (L.) ssp. japonica; (Goff, et al., 2002)) and
poplar (Populus trichocarpa; (Tuskan, et al., 2006)) respectively. Each of the three
models has its own strengths and weaknesses for use in molecular biology. Being the
most widely used system for plant molecular biology, Arabidopsis is an excellent model
of plant biology for many reasons,p including its rapid generation time, small genome,
minimal growth requirements, high seed yield and the variety of tools and techniques
which have been designed specifically for its use. However, as it is herbaceous, it is a less
useful model for traits such as wood production, maturation, perennial growth, and bark
formation, amongst others (Jansson and Douglas, 2007). While Arabidopsis does produce
secondary xylem that is physiologically similar to the wood of poplar (Chaffey, 1999), it
produces it in small quantities. Though its small size and rapid generation time makes it
amenable to molecular biology, there is perhaps a better model for these traits now
available. Due to its sequenced genome, poplar has been recently brought into
mainstream molecular biology. New tools are now available (and continue to be
developed) to allow in depth study of this angiosperm tree. These include large libraries

2
of ESTs (Kohler, et al., 2003; Lee, et al., 2005; Sterky, et al., 1998; Sterky, et al., 2004),
the production of full-length cDNA clones (Nanjo, et al., 2007; Ralph, et al., 2008), DNA
microarray capabilities (Tuskan, et al., 2006), as well as a recent flood of adaptations to
traditional protocols including those for transformations (Cseke, et al., 2007;
Spokevicius, et al., 2006; Tzfira, et al., 1998), genetics (Brunner, et al., 2004; Ralph, et
al., 2008; Regan, et al., 1999), and proteomics (Plomion, et al., 2006) (for review see
Jansson and Douglas, 2007). While poplar research does have its drawbacks, (chiefly its
large physical size and long generation time), its relatively small, diploid genome, fast
growth, and vegetative propagation make it amenable to genetic study (Chaffey, 2002).
Due to the global importance of wood, there are many industries related to, and
applications of, forest tree genetic research (Cooke and Rood, 2007). These involve the
bioenergy, biofuel and pulp and paper industries (Pan, et al., 2006; Yemshanov and
McKenney, 2008), carbon sequestration and environmental remediation efforts
(Balatinecz and Kretschmann, 2001), insight into arboreal domestication, and finally an
overall increase to our understanding of fundamental plant biology. Because of the great
importance of wood and forestry byproducts in our society, the knowledge to develop and
produce these commodities is critical to maximizing and sustaining their yields.
As do all living things, plants grow via cell division, elongation and expansion. However,
in a relatively unique manner, the process of organogenesis is constantly ongoing
throughout the plant’s lifespan (Clark, et al., 1997). This phenomenon is achieved
through controlled differentiation in a cache of undifferentiated cells close to the apical

3
ends of the shoot and root, referred to as the shoot apical meristem (SAM) and root apical
meristem (RAM) respectively. These meristems are indeterminate; with their cells
remaining undifferentiated throughout the plant's lifespan, yet the daughter cells
produced near their exterior boundaries will eventually differentiate and undergo
organogenesis. The SAM and RAM therefore require a balance between the processes of
differentiation and proliferative maintenance. Without this balance, the meristem can
exhaust its supply of undifferentiated cells and cease to function (i.e. similar to the
determinate meristems such as the floral meristem) (Clark, et al., 1997). The SAM and
RAM are responsible for growth in the vertical plane, with the SAM specifically
responsible for production of leaves, inflorescences, and primary vasculature and the
RAM responsible for the production of the roots. Besides primary growth meristems,
most plants also have secondary meristems which lead to growth in the lateral plane.
These meristems can be cylinders of undifferentiated cells, which when active, add to the
diameter of the stem or root, through the process of secondary growth. This growth is due
to periclinal divisions (along the radius of the stem) which create radial files, and
anticlinal divisions (perpendicular to the radius of the stem) which also add to the girth of
the stem or root. Radial files show distinct visible lineages which can be traced back to
the precursor meristematic cells. Differentiation occurs on both the interior and exterior
borders of secondary meristems. This bidirectional differentiation is important as it leads
to different tissues being produced on either side of the meristem. There are two
secondary meristems; the cork cambium and the vascular cambium. The cork cambium
produces cork to its exterior and ground tissues to its interior. The vascular cambium

4
produces two types of vasculature; phloem on its exterior and xylem on its interior. These
tissues will eventually mature into secondary vascular tissues (inner bark and wood). The
cork consists of dead and heavily waxed cells. It is produced to prevent desiccation and
herbivory, while still allowing gas exchange through tiny air holes. The secondary
vasculature is developed to increase the ability of water and gases (via the xylem), as well
as solutes (via the phloem), to be transported throughout the plant body. The secondary
vasculature is also responsible for mechanical support of the plant.
Wood is the end product in the maturation of secondary xylem. Briefly, secondary xylem
produced by the vascular cambium undergoes a period of cellular expansion and
elongation. This is followed by the gradual establishment of a secondary cell wall,
composed of lignin, hemicellulose and cellulose (Mellerowicz and Sundberg, 2008).
Following the establishment of the secondary cell wall, the cell undergoes programmed
cell death, the last stage of xylem cell maturation.
One of the drawbacks of plant-science research outside the realm of Arabidopsis is the
lack of mutants for phenotype-based gene discovery. In poplar research, this is one of the
largest limitations. Poplar, with its long generation time, is only amenable to dominant
mutation strategies, as recessive mutations would likely need to be selfed to demonstrate
phenotypes, requiring long production periods. Of the mutant production strategies
currently available, the most promising for high-throughput productions is activation-
tagging (Busov, et al., 2003; Weigel, et al., 2000). This technique has recently been

5
utilized to produce a population of 1800 mutants, the largest of its kind in the world
(Harrison, et al., 2007). These mutants carry an activation tag consisting of a tetrameric
repeat of enhancer elements from the cauliflower mosaic virus 35S promoter. The tag
causes the increased endogenous expression of genes local to the insertion point rather
than the ectopic expression found with typical overexpression constructs (Weigel, et al.,
2000). While a great strategy to help understand gene-function relationships, activation
tagging is not the only route to mutant production and gene identification in poplar.
When describing mutants, Arabidopsis researchers historically have not looked for
variation in secondary growth characteristics, as they are most apparent under strict
growth conditions. As such, many genes may have been identified in Arabidopsis which
would also produce aberrations in secondary growth, but haven’t yet been screened for.
Novel gene-function relationships could be identified by screening genes that are known
to have developmental regulatory properties in some meristematic regions, by looking at
their resulting phenotypes following misexpression in others.
Over the past few years, two mutants showing modified secondary growth have been
identified in the Regan Lab. The first, clavata1 (clv1), is well known for its namesake's
role as a negative regulator of SAM and floral meristem maintenance. When CLAVATA1
was downregulated in Arabidopsis, lines also showed an increase in secondary growth.
This exemplifies how even well studied mutants can be tested for further phenotypes. The
second mutant, which was identified as part of the poplar activation tagging project, is

6
referred to as corky. This mutant has an altered distribution of cells in its secondary
vasculature, as well as in the outer bark.
This thesis outlines two different strategies to identify gene-function relationships
involved with secondary growth, and more specifically, with the regulation of the
vascular cambium. The first strategy will test the hypothesis that if a mutant showing
aberrant secondary vasculature is identified in a population of activation-tagged poplar
trees, the activated gene is likely a regulator of the vascular cambium. The second
strategy will address the hypothesis that if shoot apical meristem regulatory genes are
also naturally expressed in the vascular cambium, then they likely have a role in its’
regulation as well. Using these two strategies two predicted regulators of the vascular
cambium will be tested to determine if they are involved in secondary growth. This will
hopefully open new avenues of research to help identify more regulators of this important
tissue.

7
Chapter 2
A corky mutant reveals a novel regulator of secondary growth and
development in Populus
2.1 Abstract
As a result of recent advances in genomics and other molecular techniques, we are
beginning to better understand wood formation, especially the genes involved in
construction of the cell wall. However, the molecular mechanisms that regulate the “stem
cells” responsible for wood formation, the vascular cambium, remain relatively unknown.
In particular, we understand very little about how the cambium partitions resources to
produce the appropriate amount of xylem and phloem. In a recently produced population
of activation-tagged poplar, a mutant has been identified with an altered distribution of
xylem and phloem cells such that the phloem is significantly wider than usual, and the
xylem remains very small. In addition to this, the mutant also appears to have an altered
activity of the cork cambium, as the thickness of the bark tissue far exceeds that of wild
type. These altered trunk morphologies coincide with an alteration in overall tree growth
and development; the tree grows much slower than wildtype and is unable to grow
upright. Identification of the gene responsible for this dramatic phenotype is underway
and here we will report on our progress in understanding the mutant at the anatomical and
molecular levels.

8
2.2 Introduction
Often considered paramount to the study of plant biology are the control mechanisms that
coordinate growth and development throughout the plant. Critical to these mechanisms
are regions around the plant body which maintain clusters of totipotential stem cells,
referred to as meristems. These regions are responsible for balancing the differentiation
of new cells with the maintenance of their undifferentiated population, allowing growth
and development. While vertical growth is controlled via the shoot and root apical
meristems (SAM and RAM respectively), secondary or lateral growth is achieved
through lateral meristems. These meristems are narrow cylinders of undifferentiated cells
found throughout the stem and root which when active, divide and differentiate along
both their interior and exterior boundaries, resulting in different tissues being produced
on either side of each meristem. There are two lateral meristems; the cork and the
vascular cambiums. The cork cambium produces phellum (cork) to its exterior and
phelloderm (cortex) to its interior. The vascular cambium produces phloem to its exterior
and xylem to its interior. These tissues will eventually mature into secondary vascular
tissues, inner bark and wood respectively. Secondary growth facilitates many processes
important for the maturation of various plants, especially trees. These include the
production of cork and ground tissues (used in storage, protection and support), as well as
the production, and subsequent maturation, of new phloem and xylem (responsible for the
translocation of solutes/solvents and physical support).

9
Despite the importance of these processes, our understanding of the molecular regulation
of the cork and vascular cambiums lag far behind that of the SAM (Chaffey, 2002). As a
result of recent advances in genomics and other molecular techniques, researchers are
beginning to better understand wood formation, especially the genes involved in
construction of the cell wall. However, the molecular mechanisms that regulate the
vascular cambium remain relatively unknown. In particular, we understand very little
about how the cambium partitions resources to produce the appropriate amount of xylem
and phloem. Nevertheless, there have been multiple upstream signalling inputs identified
to date, suggesting a complex coordination. Most of the new molecular information
regarding secondary growth is gleaned from mutant analysis. And though the molecular
and genetic mechanisms of secondary growth are only recently beginning to come to
light, details of the anatomy and physiology involved in this type of development are
better understood and have in fact been studied for more than a century (Gregory, 1888).
One only has to look briefly at the non-taxonomic grouping of ‘trees’ to see that outer
bark can be highly variable. It can be smooth, rough, coloured, mosaic-like or anything in
between. However, there have been few mutants reported in which, from one generation
to the next, there have been heritable changes in the outer bark. Examples include the
‘corky’ mutant in cotton (Stephens and Phillips, 1972), ‘corky-stem’ in pigeonpea
(Saxena, et al., 1988) and the ‘granthami’ rubber tree (Bartlett, 1927). Cotton
(Gossypium sp.) normally has smooth shoots and stems. However, a mutant identified by
Stephens showed rough, cork-like layers on the stems and shoots. Back-crossings

10
suggested a three allelic system in which only one combination resulted in the corky
phenotype (Stephens and Phillips, 1972). This is similar to the findings in the pigeonpea
(Cajanus cajan (L.) Millsp.) mutant, in which two alleles were predicted to form a
dominant/recessive system, with the mutant phenotype only expressed in plants
homozygous for the recessive allele (Saxena, et al., 1988). The mutant variety developed
a periderm layer which was not normally present in pigeonpea, resulting in the cork layer.
Rubber trees (Hevea brasiliensis) also produce a relatively smooth bark. Nevertheless, in
1927 a mutant was identified (mut. granthami) which developed a very rough cork layer
(Bartlett, 1927). The mutation was also hypothesized to be recessive, as only through
self-propagation was the phenotype reproduced. All three of these mutant phenotypes are
also very similar to the pathology of Vitis vinifera when exposed to Grapevine Virus B
and showing symptoms of the resulting ‘Corky Bark Disease’ (Schneider, 1973) wherein
a phenotypic rough bark develops. These mutants have likely developed due to altered
regulation of the cork cambium, either directly or indirectly.
In Arabidopsis, fewer mutants have been identified with abnormalities in the regulation
of the cork cambium. However, there are two mutants which have been found to have
altered characteristics of the phellum cells themselves. Specifically, they are suberin
mutants, which is one of the key constituents of the cork secondary cell wall (Soler, et al.,
2007), and show altered suberin deposition. eld1 (elongation defective 1) mutants show
ectopic deposition of suberin, while the knock-down of GLYCEROL-3-PHOSPHATE
ACYLTRA5SFERASE 5 (GPAT5) showed a decrease in suberin production in bark

11
tissues (Beisson, et al., 2007; Cheng, et al., 2000) The low frequency of mutants
identified is hypothesized to be due to the fact that mutants of the cork and cork cambium
are hard to visually screen for in Arabidopsis (Yephremov and Schreiber, 2005).
Contrary to the frequency of mutants of the outer bark, in the secondary vasculature of
Arabidopsis, there have been many mutants identified to date. These involve changes to
the positioning, qualities and quantities of the xylem and phloem cells, derived from a
variety of signalling mechanisms. Some of these arise from hormonal irregularities. For
example, in the shoot, low levels of auxins only allow for the differentiation of phloem
tissues, whereas high auxin levels allow for the differentiation of both phloem and xylem
tissues (Aloni, 1987). Aloni (1987) also suggests a positive regulatory role for ethylene in
xylem differentiation as well. Treatment of the stem with ethrel, an ethylene releasing
chemical, resulted in drastic changes to the vasculature (Junghans, et al., 2004). Stem
diameter increased when 5% ethrel was applied for an experimental period of one month.
This increase was made up of a two-fold increase in the amount of phloem tissue, as well
as a five-fold increase in the amount of xylem present. Also, cells in the secondary xylem
were found to be shorter than those in untreated samples, while regions of sclerified
tissues were found in the secondary phloem. As is often the case in complex signalling
pathways, few hormones seem to act alone. As such, the brassinosteroids have also been
implicated in xylem development. Defective brassinolide receptors (bri1, brl1 and brl3)
all showed a decrease in xylem production, while also showing an increase in that of the
phloem (Cano-Delgado, et al., 2004). Other proteins involved in the production of

12
brassinosteroids also result in vascular phenotypic mutants when disturbed. For example
the constitutive photomorphogenesis and dwarfism (cpd) and dwarfy7 (dwf7) mutants are
both defective in brassinosteroid synthesis and show a marked increase in the
phloem:xylem ratio (Choe, et al., 1999; Szekeres, et al., 1996). This is recapitulated when
using chemical brassinosteroid biosynthesis inhibitors such as brassinazole (Nagata, et
al., 2001). Unfortunately, plant hormones form a highly interconnected system, making it
very difficult to discern the effects of each hormone separately.
While the secondary vasculature tissues are clearly affected by various hormonal signals,
they are also regulated by numerous effectors. In the mutant altered phloem development
(apl), a MYB-coiled-coil transcription factor was misregulated resulting in a loss-of-
function phenotype which included xylem development on both sides of the vascular
cambium and a lack of phloem tissue (Bonke, et al., 2003). The protein was later found to
promote phloem development, suggesting that xylem tissue develops if phloem identity
gene products or spatial signals are not present. Studies on the continuous vascular ring
(cov1) mutant found that changes to the vasculature could be completely independent of
auxin (Parker, et al., 2003). Instead, they showed that the downregulation of COV1
resulted in an increase in both xylem and phloem tissues by an unknown mechanism.
MicroRNAs also play a large regulatory role in vascular development via the vascular
cambium. MIR116 was found to negatively regulate an HD-ZIP III gene, ATHB15,
resulting in an increase in xylem cell differentiation (Kim, et al., 2005). These represent
only a sampling of the stem vasculature mutants identified to date. For further reviews

13
please be directed to the following: (Carlsbecker and Helariutta, 2005; Scarpella and
Meijer, 2004; Scarpella and Meijer, 2004; Ye, 2002).
While there have been many mutants identified in Arabidopsis, there is currently a great
surge of interest and new work being carried out in forest trees to study changes in the
secondary vasculature. Recently we reported on the creation of a new population of
activation-tagged poplars (Harrison, et al., 2007). From this population a mutant showing
aberrations in its secondary vasculature was identified. These include an apparent
increase in the amounts of cork and phloem, as well as a decrease in the amount of xylem
produced. A gene of previously unknown function is proposed to be responsible for the
phenotype. Here we describe both the anatomy of this mutant, called corky and the
molecular characterization of the gene hypothesized to be responsible.
2.3 Materials and Methods
2.3.1 Plant growth and propagation
Trees were maintained in tissue culture and in the greenhouse with previously reported
conditions (Harrison, et al., 2007). corky was reintroduced from soil to tissue culture
using 2 inch shoot apical cuttings, which were then bleached (10 % bleach (v/v) and 0.01
% (v/v) Tween-20) for 17 mins, and rinsed for 1 hr in water. Cuttings were then placed
upright into Maintenance Media (Harrison, et al., 2007) and left to root on a light rack

14
with 16 hr/8 hr (day/night) light schedule. At this point, the line was propagated as
previously described.
2.3.2 Anatomical characterization of the corky phenotype
Both cross and tangential sections of corky were obtained either by hand or by the use of
a Vibratome Series 1000 Sectioning System (Soquelec International; Montreal, QC).
Analysis of the vasculature and secondary growth were carried out on stained and
unstained samples. Those stained were done so with TBO (0.05 % (w/v) Toluidene Blue
O, 1.0 % (w/v) Borax) or 1.3 % phloroglucinol-HCl and mounted in water. Unstained
samples were sectioned and mounted in water. Samples were analyzed with a Zeiss Axio
Imager Z1 microscope (Göttingen, Germany) using a Zeiss AxioCam HRC camera.
Images were taken using no greater than 20x magnification with brightfield conditions.
Sections were also analyzed using an ultraviolet light source, taking advantage of the
natural fluorescence of secondary cell walls. These images were obtained using a
combination of brightfield, a DAPI filter (excitation λ: 359 nm, emission λ: 461 nm), a
green fluorescent protein (GFP) filter (excitation λ: 470 nm, emission λ: 509 nm) and
were then false coloured. Wood fiber analysis was carried out using a maceration
procedure previously reported (Chaffey, et al., 2002). Briefly, samples were boiled in a
solution of 50 % (v/v) glacial acetic acid and 3 % (v/v) hydrogen peroxide for 10 hours.
After boiling, samples were rinsed carefully in water twice and then neutralized with
sodium bicarbonate. Samples were then subjected to vigorous vortexing and multiple
passages through an 18 gauge needle. After initial analysis on the Zeiss Axio Imager Z1
using brightfield, samples were sent to Dr. Shawn Mansfield at UBC to be measured in a

15
Fiber Quality Analyzer (FQA) (OpTest Equipment Inc., Hawksbury, Ontario). The FQA
measures roughly 10 000 cells for a given sample, thus increasing the reliability of the
data.
2.3.3 Localization of the activation tag insertion site
The insertion site of the activation tag in corky was localized using Clontech’s Genome
Walker Universal Kit (Mountainview, CA). Using cetyl trimethylammonium bromide
(CTAB) purified DNA (Murray and Thompson, 1980), the four libraries were created as
directed. The primers AP1 and AP2 were provided, corresponding to the adapter and
nested adapter primers respectively. Gene specific primers were developed from both the
left and right borders of the activation tag T-DNA. The left border gene specific primers
(LBGSP1 and 2) and the right border gene specific primers (RBGSP1 and 2) are listed
below in Table 1 and shown in Appendix D. Amplified bands were cloned using the
pGEM-T Easy Vector System (Promega, Madison, WI) and XL1-Blue competent E. coli
using Chung’s transformation protocol (Chung, et al., 1989). Plasmids were then purified
using Invitrogen’s (Carlsbad, CA) PureLink Quick Plasmid Miniprep Kit, and sequenced.
2.3.4 Testing local genes for altered expression using qRT-PCR
Primers were developed to test the expression levels of genes within 12 kb upstream and
downstream of the insertion site (Appendix C). This was carried out using Primer3
software (Untergasser, et al., 2007) and following the guidelines found in the QuantiTect
SYBR Green qRT-PCR kit manual (Qiagen; Mississauga, Ontario). All reactions
included 25 ng of total RNA and primers at 1 mM. Reactions were run using the

16
Table 1. Primers used in the identification and characterization of the corky mutant.
Miscellaneous and cloning primer sequences can be found in Appendices A and B
respectively.
Task Primer �ame Role
AP1
AP2
Genome walking adaptor
primer 1 & 2
LBGSP1
LBGSP2 T-DNA left border primers
RBGSP1
Genome Walking
RBGSP2 T-DNA right border primers
RAP1
RAP2 Race adapter primers 1 & 2
RACE
GRAILFW-L Gene specific RACE Primer
MBGrailORFF1
MBGrailORFR1
Sense orientation,
Overexpression construct
MBGrailORFASF1
Cloning of FM#2
(Sense and
Antisense) MBGrailORFASR1
Antisense orientation,
Antisense construct
At5g49525Fw2
At5g49525Rv
Arabidopsis homolog #1,
Overexpression construct
At5g26790Fw2
Cloning of FM#2
Arabidopsis
Homologs At5g26790Rv
Arabidopsis homolog #2,
Overexpression construct
LBa1-SALK T-DNA specific primer
SALK93568-LP
SALK93568-RP At5g49525 genomic border primers
SALK27013-LP
Screening for
homozygous
Arabidopsis
SALK Lines SALK27013-RP
At5g26790 genomic border primers
MBP717GRAIL-S
MBGrailORFR1 FM#2 open reading frame amplification
MBGrailRNAi_F
Induced Somatic
Sector Analysis
FM#2 Constructs MBGrailRNAi_R
FM#2 RNAi (Truncated hairpin-RNAi)

17
SmartCycler Platform from Cepheid (Sunnyvale, CA). Cycling conditions were kept
consistent, with only annealing temperatures adjusted as shown in Table 2. Each program
included a 30 minute reverse transcription step at 50 °C, followed by 15 minutes at 95°C.
The three-step cycle was repeated 45 times (94 °C x 15 s, [TAnneal] x 30 s and 72 °C x 30 s
with optics on).
2.3.5 Sequence confirmation of FM#2 cD�A
Total RNA was isolated with CTAB using established protocols (Friedmann, et al., 2007)
from the shoot apical meristem of a wildtype poplar. From this, polyadenylated mRNA
was purified using the DynaBeads mRNA Direct Kit from Dynal Biotech (Oslo, Norway)
following their protocol 2.1 (Sections A and C). The beads were regenerated using their
protocol 3.1. This process was repeated until the 18S and 26S ribosomal bands were no
longer visible on a 1.5 % TAE gel. This purified mRNA was then used to create a RACE
(Rapid Amplification of cDNA Ends) library using the Marathon cDNA Amplification
Kit (Clontech; Mountainview, CA). Using supplied primers AP1 and AP2 (herein called
RAP1 and RAP2 to distinguish them from Genome Walking AP1 and AP2), as well as
GRAILFW-L (Table 1; Appendix D), bands were amplified and afterwards sequenced to
determine the transcript sequence of FM#2.
2.3.6 Cloning of poplar FM#2 and Arabidopsis homologs for misexpression studies
The open reading frame of FM#2 was cloned using Platinum PFx DNA Polymerase
(Invitrogen; Carlsbad, CA) from corky cDNA with MBGrailORFF1/MBGrailORFR1 and
MBGrailASF1/MBGrailASR1 primer combinations (Table 1; Appendix D) and cloned
18
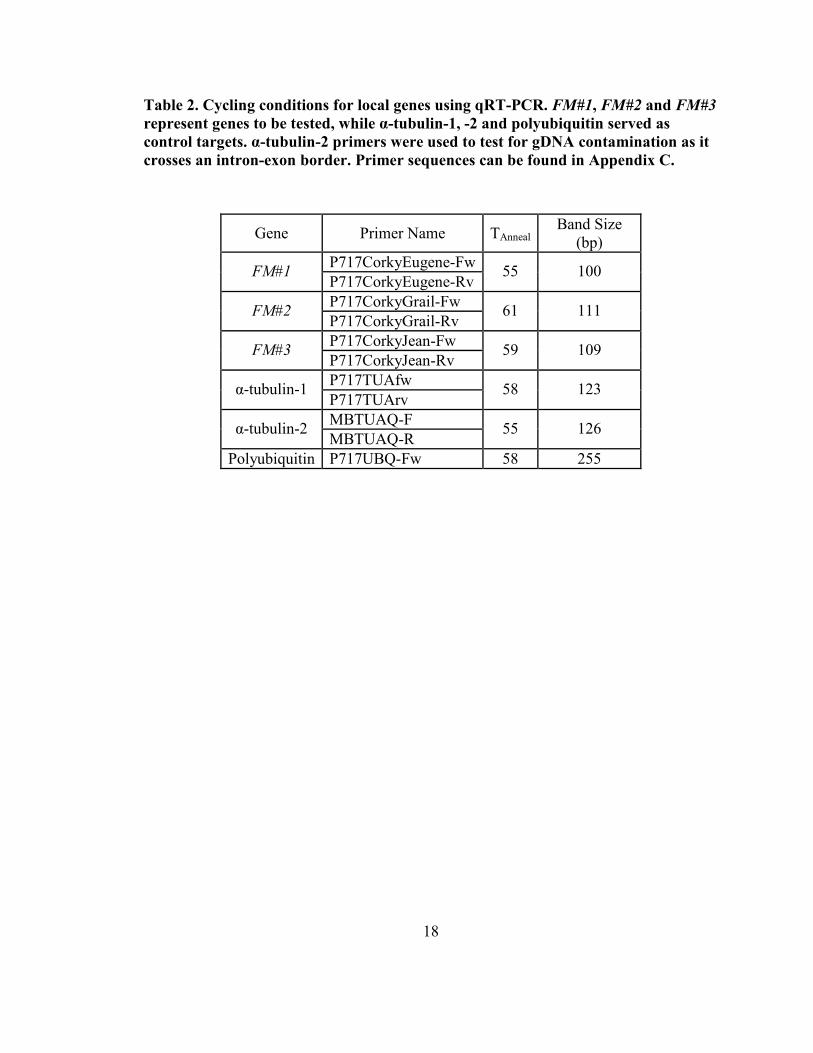
Table 2. Cycling conditions for local genes using qRT-PCR. FM#1, FM#2 and FM#3
represent genes to be tested, while α-tubulin-1, -2 and polyubiquitin served as
control targets. α-tubulin-2 primers were used to test for gD�A contamination as it
crosses an intron-exon border. Primer sequences can be found in Appendix C.
Gene Primer Name TAnneal Band Size
(bp)
P717CorkyEugene-Fw FM#1
P717CorkyEugene-Rv 55 100
P717CorkyGrail-Fw FM#2
P717CorkyGrail-Rv 61 111
P717CorkyJean-Fw FM#3
P717CorkyJean-Rv 59 109
P717TUAfw α-tubulin-1
P717TUArv 58 123
MBTUAQ-F α-tubulin-2
MBTUAQ-R 55 126
Polyubiquitin P717UBQ-Fw 58 255

19
into pGEMT-Easy (Invitrogen; Carslbad, CA). The former product corresponded to the
sense orientation of the open reading frame, while the later product, corresponded to the
antisense orientation. They were excised from pGEM-T Easy using 5coI and BsteII and
then ligated into pCAMBIA 1305.1 (-GUSPLUS) using the 5coI and BsteII sites,
securing their directionality. These sense and antisense constructs were transformed into
Agrobacterium (Strain C58 (pGV3850)) and used in standard transformation poplar
protocols (Harrison, et al., 2007). A third construct was also created using the 5coI/BsteII
excised ORF, where a further excision with FatI was carried out prior to ligation into
pCAMBIA 1305.1 (5coI/BsteII cut). The open reading frame of FM#2 was also cloned
using an alternate set of primers for use in the ISSA protocol (see below), which uses
gateway enabled vectors (Invitrogen; Carlsbad, CA). Amplification was carried out as
described below using the MBP717Grail-S and GrailORFR1 primers (Table 1; Appendix
D).
The potential Arabidopsis homologs of FM#2, At5g49525 and At5g26790, were cloned
using the primers pairs At5g49525Fw2/At5g49525Rv and At5g26790Fw2/At5g26790Rv
(Table 1; Appendix D). Both were cloned into pGEMT-Easy and subsequently excised
with 5coI and BsteII. Following ligation into pCAMBIA 1305.1 (-GUSPLUS), the
plasmid was subcloned into Agrobacterium. Finally, Arabidopsis thaliana (Col) were
transfected using the floral dip methodology (Clough and Bent, 1998).

20
2.3.7 SALK lines corresponding to FM#2’s potential Arabidopsis homologs
SALK T-DNA knock-out lines were obtained for the two homologs of FM#2, At5g49525
and At5g26790 (SALK_093568.16.30 and SALK_027013.31.00 respectively) (Alonso,
et al., 2003). PCR screening for homozygous knock-out lines was done via a multiplexing
protocol as found on the SALK website (http://signal.salk.edu/tdnaprimers.2.html).
Primers were designed using the aforementioned website (Table 1). After screening for
homozygosity in the T3 generation, T4 seeds were planted in soil where they were grown
under long day [16 hr/ 8 hr (day/night)] conditions, and will be analyzed anatomically for
histological variation in the hypocotyl. Staining with TBO and phloroglucinol will be
carried out as previously described above. Measurements of the phloem and xylem
regions of cross sections will be made using the Zeiss AxioVision 4.6 software suite.
2.3.8 Induced somatic sector analysis as a recapitulation strategy
A total of four constructs were created to use with Induced Somatic Sector Analysis
(ISSA) (Van Beveren, et al., 2006). These included (as mentioned above) a sense, an
antisense, a hairpin-RNAi (all based on the open reading frame of FM#2) and a truncated
hairpin-RNAi (a 22 nucleotide fragment). The fragments were originally amplified using
Platinum PFx DNA Polymerase (Invitrogen; Carlsbad, CA) and cloned into an entry
vector using the pCR8/GW/TOPO TA Cloning Kit (Invitrogen; Carlsbad, CA). Using the
gateway technology (Invitrogen; Carlsbad, CA), expression vectors were created using
pre-established single or double cassette, gateway-enabled pCAMBIA1305.1 vectors (for
vector maps, see Appendices E and F). “Cambial windows” were created in greenhouse
trees by making two vertical cuts in the bark with a scalpel. A horizontal cut was made

21
between the two vertical cuts, joining them at their basal end. The flap of bark was then
lifted and the trees were directly inoculated with the Agrobacterium strain AGL1,
harbouring one of the four constructs, or one of the two empty vectors. Following
inoculation, cambial windows were shut and sealed with parafilm to promote healing of
the wound and subsequent growth. Trees were transformed in the summer and left to
grow for 4 months before sampling took place.
2.4 Results
2.4.1 corky mutant identified in activation-tagged population showing altered
morphology of secondary vasculature.
The corky mutant was visually identified in the activation-tagged population of Populus
alba x P. tremula as it demonstrates gross morphological changes including what
appears to be an inability to support itself. The entire tree can not stand upright but
instead has a vine-like appearance (Figure 1; A). corky also displays slower growth, with
new shoots often arresting development unexpectedly. This is in stark contrast to
wildtype trees which grow very rapidly and rarely require structural support. The vine-
like growth habit was not obvious for some time, and as such corky was originally
identified due to its extremely rough outer bark (Figure 1; B,C). Cross sections of the
stem reveal that the corky mutant also has an unusually high amount of cork tissue
(Figure 1; D, E) implying the cork cambium is similarly affected. However, upon further
examination, there proved to be variation in the secondary vasculature as well. In the
same cross sections of the corky stem, there appears to be an increase in phloem and a

22
Figure 1. The anatomy of wildtype (A, B, D, F, H, J, L) and corky (A, C, E, G, I, K,
M). Trees in profile (A) showing the typical wildtype growth pattern of primarily
apical dominance, and the stunted, vine-like growth of corky (left to right: wildtype,
wildtype, corky, corky). Stems in profile (B, C) as well as unstained cuttings (D, E)
show the rough bark of corky and and the increased amount of bark present in the
mutant. In cross sections stained in 1.3 % phloroglucinol-HCl (F, G) in which
lignified tissues appear pink, differing ratios between xylem and phloem tissues are
apparent. Upon higher magnification (5x) and staining with TBO, xylem cell size
appears to be different between wildtype and the mutant (H, I) (scale bars represent
200 µm). UV illuminated cross-sections (J, K) at 10x magnification, showing
variation in the orientation of phloem fibers (scale bars represent 200 µm).
Longitudinal sections (L, M) of bark and phloem tissues showing regular files in
wildtype and a larger, unorganized bundle in corky (5x) (scale bars represent
200µm).

23
BB
A
KJ
C
D EE
FF G
IH
ML MLL M
BB
A
KJ
C
D EE
FF G
IH
ML MLL M

24
decrease in xylem when compared to wildtype controls. This is more easily observed
when cross sections are either stained with phloroglucinol - where lignified tissues such
as wood stain a dark pink colour (Figure 1; F,G) or with Toluidene Blue O (TBO), a
polychromatic stain, under 5x magnification (Figure 1; H,I). This phenotype suggests that
there is an alteration in either the activity of the vascular cambium or in the maturation of
the xylem and phloem cells themselves. Upon closer examination, the cells in the xylem
of corky appeared to be smaller in diameter. This was assessed by macerating various
xylem samples and sending them to be analyzed in the Fiber Quality Analyzer at UBC.
The cross sections (Figure 1; F, G) also seem to show more phloroglucinol staining in the
phloem of corky than in wildtype, indicating an increased amount of lignified tissue may
be present. UV fluorescence of secondary cell walls (Figure 1; J, K) show that the
phloem fibers in corky appear to be established in a different orientation than those of
wildtype. While wildtype phloem fibers are typically found in anticlinal bunches or
bands, the phloem fibers of corky appear to be in almost radial bands. The phloem fibers
were examined using longtitudinal sections and are shown in Appendix G. It was
observed that while small differences in the amount of phloem were present before the
first fully expanded leaf, more dramatic changes were observed at plastochron indices
(PI) 10 and 20. At PI 10, a second row of phloem fibers was visible in corky while
wildtype had only one row until PI 20. The largest difference was observed between PI
15 and PI 20 where corky showed a vast increase in the amount of phloem, cortex and
outer bark. This difference appears to coincide with the appearance of the outer bark
morphology. The cells constituting the phloem fibers seem very large and bulky,
suggesting that they are in fact different than those of wildtype (Figure 1; L, M). Samples

25
were taken to macerate these fibers to analyze their dimensions. Measurements of the
macerated wood fibers of corky suggest that the corky cells (426.10 µm) are shorter than
those of wildtype (528.62 µm) (Appendix G). They were found to be roughly 80 % of the
length of wildtype fibers. Preliminary analysis of fiber length, through a collaboration
with Dr. Shawn Mansfield at UBC’s Department of Wood Science, has potentially
confirmed this finding bringing reliability to this observation by analyzing 10 000 fibers
(Appendix G). corky fibers were still roughly 85% the length of wildtype's. However,
samples at various developmental stages remain to be tested to increase confidence in this
finding. Samples of the phloem along the developing shoot have been macerated and sent
to Dr. Mansfield to also determine if they are statistically different, and if so, at what
stage of growth the change has occurred at. In older samples of corky cross-sections of
the stem suggest that there may be fewer vessel elements in the xylem, a finding that will
also be investigated by Dr. Mansfield.
2.4.2 corky mutant’s single insertion site near three unknown genes
Southern blot analysis was carried out on corky, as well as a number of other mutants,
using DIG-labeled probes to various positions throughout the T-DNA (Harrison, et al.,
2007). corky was shown to have one insertion site. Using ‘genome walking’, which is a
modified TAIL-PCR technique, the location of this insertion site was identified. A 25 kb
region surrounding the insertion site contains three predicted genes or filtered models
(FMs), with each having associated ESTs. The activation tag falls in an intergenic region,
roughly 4 kb away from the ends of both of the first two genes (Figure 2), and
approximately 9 kb from the start site of the remaining gene. All three of the genes,
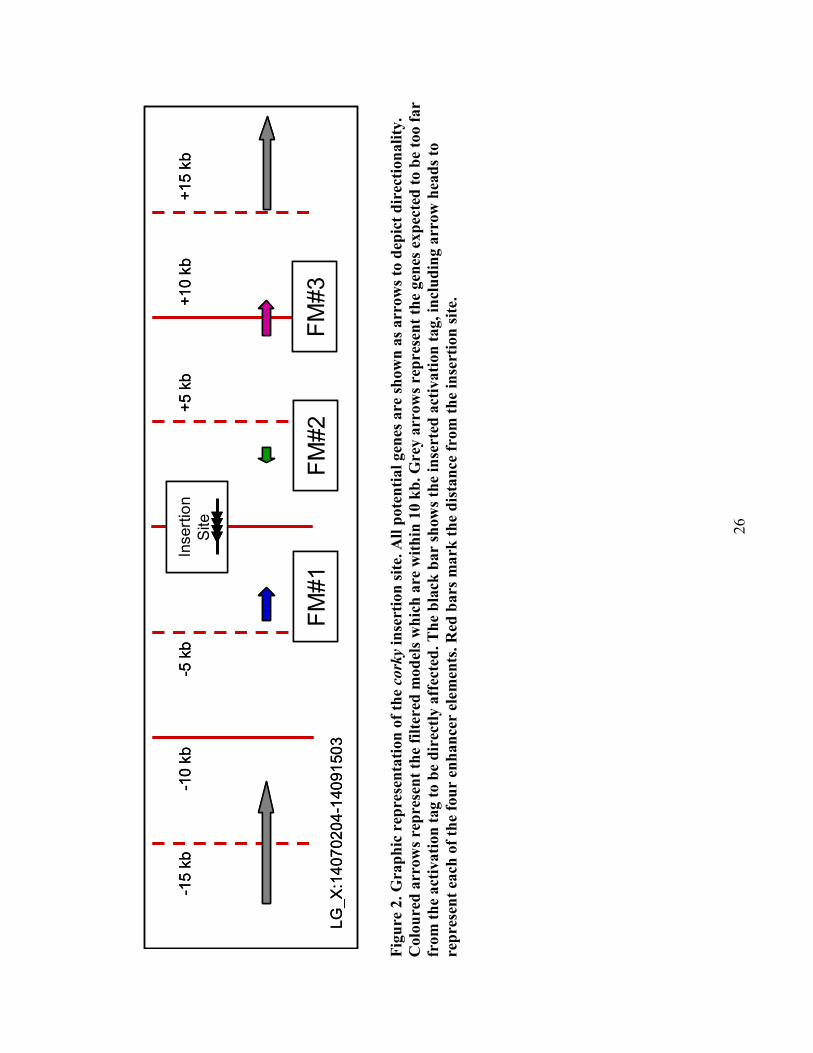
26
LG_X:14070204-14091503
-15 kb
+15 kb
+10 kb
+5 kb
-5 kb
-10 kb
FM#1
FM#2
FM#3
Insertion
Site
LG_X:14070204-14091503
-15 kb
+15 kb
+10 kb
+5 kb
-5 kb
-10 kb
FM#1
FM#2
FM#3
Insertion
Site
Fig
ure
2.
Gra
ph
ic r
epre
sen
tati
on
of
the co
rky
inse
rtio
n s
ite.
All
pote
nti
al
gen
es a
re s
how
n a
s arr
ow
s to
dep
ict
dir
ecti
on
ali
ty.
Colo
ure
d a
rrow
s re
pre
sen
t th
e fi
lter
ed m
od
els
wh
ich
are
wit
hin
10 k
b. G
rey
arr
ow
s re
pres
ent
the
gen
es e
xp
ecte
d t
o b
e t
oo f
ar
from
th
e a
ctiv
ati
on
tag t
o b
e d
irec
tly a
ffec
ted
. T
he
bla
ck b
ar
show
s th
e in
sert
ed a
ctiv
ati
on
tag, in
clu
din
g a
rrow
hea
ds
to
rep
rese
nt
each
of
the
fou
r en
han
cer
elem
ents
. R
ed b
ars
ma
rk t
he
dis
tan
ce f
rom
th
e in
sert
ion
sit
e.

27
designated FM#1, FM#2 and FM#3, are identified as hypothetical proteins, lack user
annotation and have potential Arabidopsis homologs which are also annotated only as
expressed proteins (Appendix H).
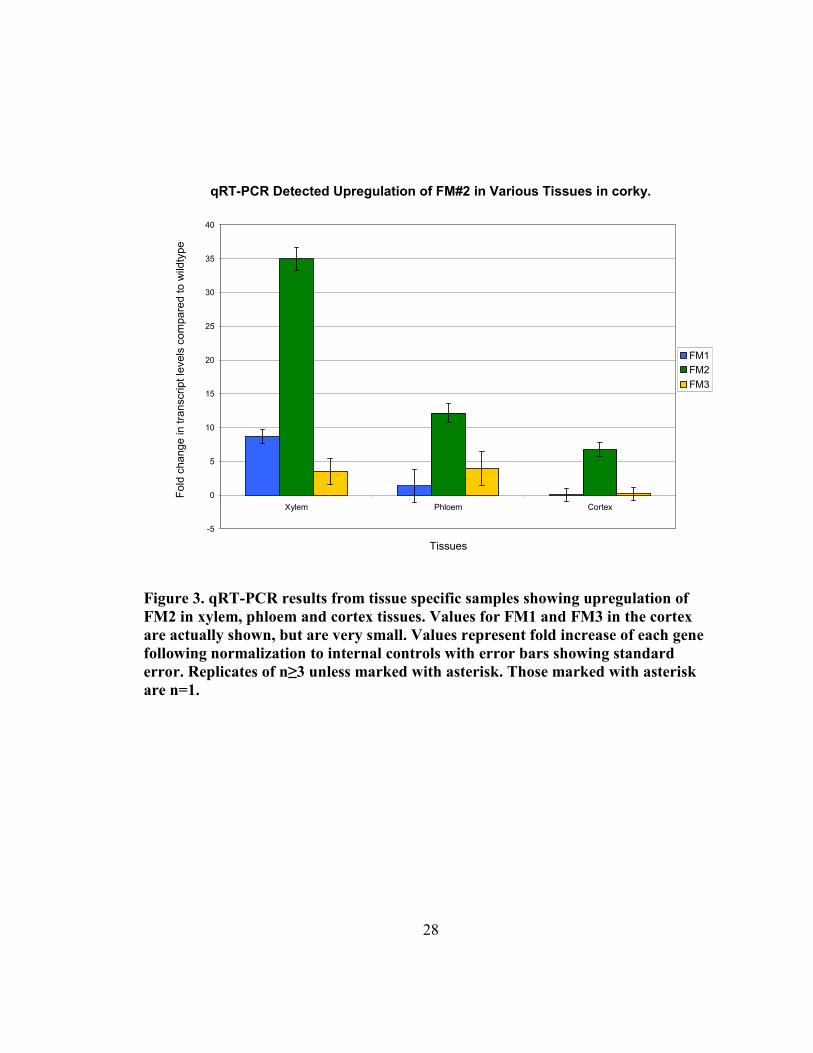
2.4.3 qRT-PCR shows single gene drastically upregulated in corky vascular tissue
Quantitative RT-PCR has been carried out to test the expression of the three closest gene
models to the corky insertion site. Primers were designed for each of the gene models, as
well as for housekeeping genes, including a polyubiquitin and an α-tubulin. As the
activation-tag is expected to have heightened endogenous expression of local genes,
rather than ectopic expression, qRT-PCR screening was concentrated to secondary
vascular tissues (Figure 3). Using these tissues, one of the local genes, FM#1, was
upregulated 8.7 fold (± 1.1 (S.E)) and 1.3 (± 2.4 (S.E)) in xylem and phloem/cambium
tissues respectively. It showed no upregulation in cortical tissues. FM#3, while furthest of
the three from the activation tag, showed more consistent values of 3.5 (± 1.9 (S.E)) and
3.9 (± 2.5 (S.E)) in xylem and phloem/cambium, and again was not upregulated in
cortical tissues. FM#2 showed higher fold increases than either of the other two filtered
models. In xylem tissues, FM#2 was upregulated 35.0 (± 1.7 (S.E)), while a more modest
increase was found in phloem/cambium and cortical tissues, 12.1 (± 1.4 (S.E)) and 6.7
respectively. The pattern of upregulation is consistent with the tissues showing the corky
phenotype.
2.4.4 FM#2 represents a novel gene of unknown function
A comparison of the nucleotide sequence of the Populus trichocarpa FM#2 and the
28
qRT-PCR Detected Upregulation of FM#2 in Various Tissues in corky.
-5
0
5
10
15
20
25
30
35
40
Xylem Phloem Cortex
Tissues
Fold change in transcript levels compared to wild
type
FM1
FM2
FM3
Figure 3. qRT-PCR results from tissue specific samples showing upregulation of
FM2 in xylem, phloem and cortex tissues. Values for FM1 and FM3 in the cortex
are actually shown, but are very small. Values represent fold increase of each gene
following normalization to internal controls with error bars showing standard
error. Replicates of n≥3 unless marked with asterisk. Those marked with asterisk
are n=1.

29
corresponding EST sequences revealed a discrepancy in the overall length of the
anticipated transcript for FM#2. More specifically, one of the ESTs associated with
FM#2 was actually 600 bp longer (in the 3' UTR) than the filtered model and other ESTs
predicted. RACE (Rapid Amplification of cDNA Ends) was carried out to determine the
sequence of the transcript's 3' end. The full 3' UTR sequence which was obtained, aligns
well with the filtered model, suggesting that the 'extended' filtered model is an artifact.
This was later confirmed through comparison to published full-length cDNA clones, the
sequence of which were recently made available (Nanjo, et al., 2007). As such the filtered
model was used for all cloning purposes. After various bioinformatics strategies, very
little functional data was obtained regarding this gene (information regarding the various
bioinformatics tools, including hosts, references and links can be found in Appendix G).
The gene itself appears to be very short, with the transcript being 529 bp, and the open
reading frame coding only 82 amino acids. ExPASy's 'ProtParam' tool describes FM#2 as
a 9.8 kDa protein with a hypothetical pI of 4.51. At NCBI's reference sequence protein
database, FM#2 has high homology to two proteins in Vitis vinifera, one in Medicago
trunculata and two in Arabidopsis. There are also similar proteins in both rice and
tobacco. These are all listed as expressed or hypothetical proteins of unknown function,
with the exception of the tobacco gene. It was identified as Avr9/Cf9 Rapidly Elicited
protein 180 (ACRE180) (Rowland, et al., 2005), but is still without a known function. In
all cases, the carboxy-terminal region is well conserved (Figure 4). There also appears to
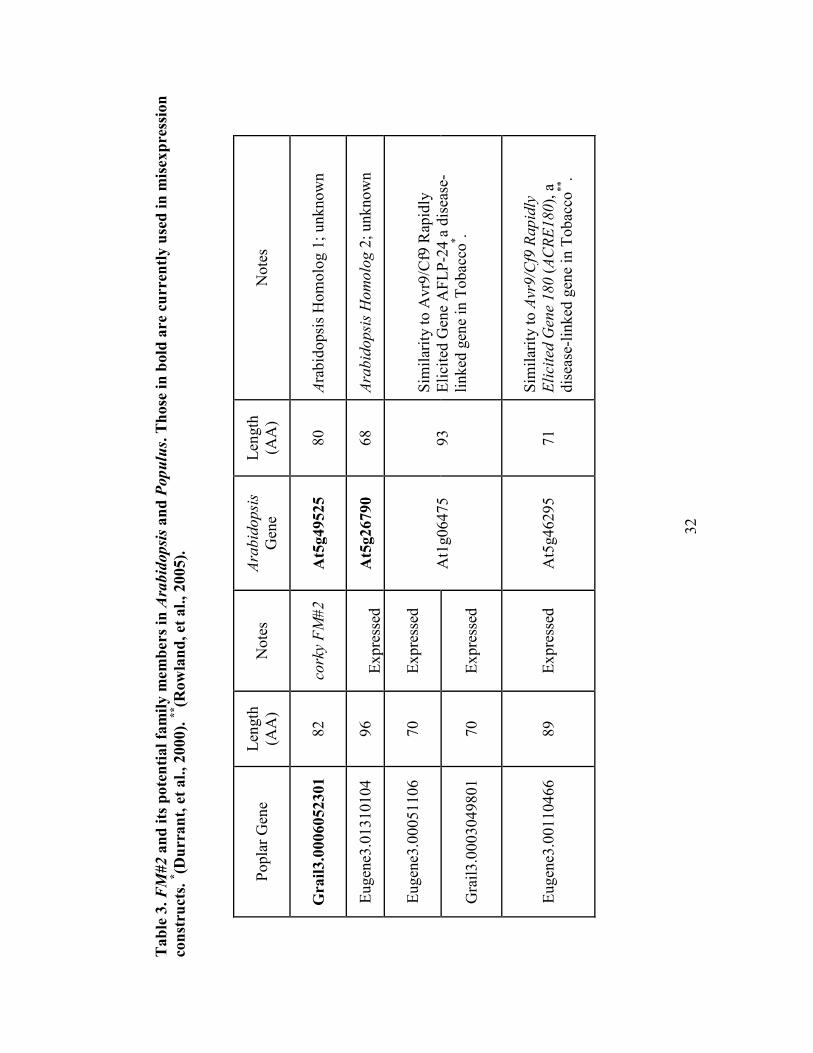
be a small gene family of which FM#2 is possibly involved. Four genes in Populus are

30
Figure 4. Analysis of the Arabidopsis and Populus genes identified by Plant Tribes as
belonging to the FM#2 gene family. (A) Alignment produced using PRALI�E with
conservation rated using the heat map in the top left corner. (B) Cladogram showing
the potential relationship between the genes (Larkin, et al., 2007). Arabdidopsis
genes are labeled with their AGI numbers, while poplar genes are listed by JGI
model numbers. A summary of nomenclature is found in Table 3. Here FM#2 is
listed as grail3.00060523 (in A) and grail 3.0006052301 (in B).
A
B

31
identified as being familial by Plant Tribes (Wall, et al., 2008) and have corresponding
genes in Arabidopsis, which are also suggested to be in a family by PARALOG (Table 3,
Figure 4). When the Arabidopsis genes were submitted to the Athena (Arabidopsis
thaliana expression network analysis) promoter element analysis software, many
potential elements were identified (Appendix J). However, of 23 different elements
identified, only 4 were found in all of the three genes (At5g26790, At5g46295,
At5g49525; At1g06475 was identified as an invalid accession number). These included a
W-box promoter motif, a TATA-box motif, and sites for both MYB1 and MYB4 binding.
FM#2 had no other known motifs detected by the CDD (Conserved Domain Database) at
NCBI (National Centre of Bioinformatic Information), nor were there any found in other
databases such as EMBL (European Molecular Biology Laboratory) and KEGG (Kyoto
Encyclopedia of Genes and Genomes). Some evidence suggests that the protein’s C-
terminus could fold into a single pass transmembrane helix domain, with the N-terminus
being a non-cytoplasmic domain (Interpro Scan at EMBL, and both DAS (Dense
Alignment Surface) and Phobius at SBC (Stockholm Bioinformatics Centre) (Cserzo, et
al., 1997; Käll, et al., 2004; Zdobnov and Apweiler, 2001)) as depicted in Appendix K.
However, it remains unclear as to which cellular membrane it would be targeted to
pTARGET predicts with 93% confidence that FM#2 is localized to the plasma
membrane, but other prediction data was less conclusive. According to TIGR, At5g49525
and At5g26790 are also both predicted to have a transmembrane domain and a prenyl
32
Tab
le 3
. FM
#2 a
nd
its
pote
nti
al
fam
ily m
em
bers
in A
rabidopsis
an
d Populus.
Th
ose
in
bold
are
cu
rren
tly u
sed
in
mis
exp
ress
ion
con
stru
cts.
*(D
urr
an
t, e
t al.
, 2000).
**(R
ow
lan
d, et
al.
, 2005).
Popla
r G
ene
Len
gth
(AA
)
Note
s
Ara
bid
opsi
s
Gen
e
Len
gth
(AA
)
Note
s
Gra
il3.0
006052301
82
cork
y F
M#
2
At5
g49525
80
Ara
bid
opsi
s H
om
olo
g 1
; unknow
n
Eugen
e3.0
1310104
96
Ex
pre
ssed
A
t5g26790
68
Ara
bid
opsi
s H
om
olo
g 2
; unknow
n
Eugen
e3.0
0051106
70
Ex
pre
ssed
Gra
il3.0
003049801
70
Ex
pre
ssed
At1
g06475
93
Sim
ilar
ity t
o A
vr9
/Cf9
Rap
idly
Eli
cite
d G
ene
AF
LP
-24 a
dis
ease
-
linked
gen
e in
Tobac
co*.
Eugen
e3.0
0110466
89
Ex
pre
ssed
At5
g46295
71
Sim
ilar
ity t
o A
vr9/C
f9 R
apid
ly
Eli
cite
d G
ene
180 (
AC
RE
180),
a
dis
ease
-lin
ked
gen
e in
Tobac
co**.

33
group binding site (due to its CAAX motif), suggesting that they too are membrane
bound. Using SUBA (Arabidopsis Subcellular Database) and the BAR (Botany Array
Resource), they are predicted to be mitochondrial, chloroplastic or perhaps (in the case of
At5g49525) nuclear bound proteins (Heazlewood, et al., 2007).
2.4.5 Production of misexpression constructs for FM#2 and its Arabidopsis
homologs
To test the hypothesis that FM#2 is actually CORKY, various misexpression constructs
have been developed to try to recapitulate the corky phenotype. These include constructs
for stable transformation (traditional tissue culture transformation), as well as for a
recently developed expression system called induced somatic sector analysis (ISSA)
(Spokevicius, et al., 2006; Van Beveren, et al., 2006). A summary of the status of each of
the constructs is found in Appendix S. In both cases, the open reading frame of FM#2
was cloned in both sense and antisense directions, allowing for both sense and antisense
expression lines to be created. In the ISSA system, two hairpin-RNAi lines were also
created, one being the 250 bp ORF in both orientations, while the other is a 22 bp insert,
again in both orientations. These constructs have all been transformed into their
respective Agrobacterium strains. In the case of the stable expression lines, long term
storage stocks were made of the lines to await their entrance in the lab’s tissue culture
pipeline. However, in the case of the ISSA expression lines, the transfection of trees with
the various lines will be completed shortly. These trees will continue to grow for four
months before they can be cut and assayed for phenotypes associated with the expression
of the various constructs. In addition, the two closest homologs of FM#2 in Arabidopsis

34
have been cloned into overexpressor constructs to see if overexpression of these
homologs in Arabidopsis will show a corky-like phenotype. In contrast to this approach,
SALK promoter knock-out lines have been ordered for these two genes as well to see if
there is a detectable phenotype when the expression levels are lowered.
2.5 Discussion
2.5.1 corky mutant identified showing altered morphology in secondary vasculature
The corky phenotype is pleiotropic, affecting the stature, bark and cellular structure of the
tree. corky shows a stunted, vine-like growth pattern unable to stand straight like
wildtype. The tree also has a rough bark that makes it very conspicuous. However, it
wasn't until a cross section was made of corky's stem that the other aspects of the
phenotype were revealed. These included changes at both the tissue and cellular level.
At the tissue level, there are changes to the outer bark, both in quality and quantity.
Variation in the amount of cork tissues produced point to a change in the regulation of the
cork cambium. Furthermore, it is not a continuous stele of enhanced cork production, but
rather broken and discontinuous, again suggesting that a profound disruption of
meristematic function has in fact occurred. Further examination of the cross-sections of
corky resulted in a large amount of phenotypic data being amassed. As shown in Figure
1, staining with Toluidine Blue O (TBO), a polychromatic stain, clearly shows a change
in the amounts of secondary vasculature present in the corky stem. Secondary xylem,

35
which typically stains blue to blue-green, with the typical thick secondary cell walls, was
present in much lower amounts in corky when compared to wildtype stems of equal
diameter. Accordingly, there appeared to be much more phloem and cortical tissues in the
mutant. This variation in secondary vasculature suggests that a change has also occurred
in corky which is affecting the vascular cambium. That both the vascular and cork
cambiums were producing more tissues to their exteriors may be coincidental, but may
also imply that a similar regulatory role has been affected in the activation-tagged line.
Through staining with phloroglucinol-HCl, a polyphenolic-specific stain, the TBO data
was corroborated. There was much less secondary xylem observed in corky, made
obvious by the bright pink staining of the lignified secondary cell walls (Figure 1).
Unexpectedly, the phloroglucinol staining also showed variation in corky's lignified
phloem fibers. Typically phloem fibers are found in well defined bundles which develop
in concentric steles. In contrast to this, the phloem fibers of corky are often found in
jagged, radially-expanding groups. These were in jagged, radially-expanding groups,
rather than being in well defined bundles which develop in concentric steles, as is found
in wildtype. It is not known, whether or not these regions have radially expanded to
support the larger tissues of phloem and bark which corky develops.
A further alteration revealed by phloroglucinol staining was that the cells of the
secondary xylem in corky appeared to be smaller and had either smaller, or less

36
numerous, vessel elements. To test whether or not this was the case, xylem samples were
macerated to break intercellular connections and middle lamella. When observed under
the microscope, xylem fibers appeared to be shorter than those of wildtype (Appendix G).
To further examine this, a xylem sample of corky and wildtype were sent to Dr. Shawn
Mansfield at UBC's Department of Wood Science to assay the lengths and widths of
10,000+ fibers and vessels. The scale of the assay brings reliability to these findings,
although fails to supplement the need for further sampling. Preliminary data has returned
suggesting that our values were correct; corky appears to have shorter xylem fibers. Data
regarding the vessels is still pending. If it can be confirmed that corky has shorter xylem
cells, the difference in xylem fibers (used for vertical support) and vessel elements (used
for water transport) could perhaps shed light on the inability of corky to grow and support
itself in the vertical plane. This could be a finding which links the phenotype on the
morphological level to a phenotype on the cellular scale. Due to the variation in
developmental conditions from tree to tree, we have recently prepared numerous samples
of corky and wildtype to be sent for further analysis. While wildtype trees are regularly
pruned to control their height, corky, a slow growing mutant, isn't. The variable growth
rate makes the task of obtaining 'equivalent' tissues slightly difficult as age,
developmental stage and maturity do not necessarily correlate consistently in corky.
Some clones appear to grow for a period of time before the corky external bark
phenotype manifests, while others show the mottled stem in new tissues. Furthermore, as
sampling the xylem from the tree's trunk effectively requires sacrificing the tree (or at

37
least its growth for the next few months) xylem samples are only taken when corky
clones are being harvested for microscopy purposes.
corky has shown itself to be a complex mutant, with variations in a number of
developmentally crucial tissues. Together they exhibit a very similar phenotype to the
pathology of 'Corky Bark Disease' in grape (Beukman and Gifford, 1969). Both corky
and the infected grapevines show over-proliferation in the phloem tissues, as well as the
externally apparent rough and cracked bark. The corky phenotype is also similar to the
Granthami mutant of Hevea brasiliensis (rubber tree), where the mutant was identified
due to its aberrant bark development, increased latex production and an inability to
maintain vertical growth without physical support (Bartlett, 1927). Granthami also
demonstrates a similar pattern of initial bark formation to that of corky. Unfortunately, in
both the cases of 'Corky Bark Disease' and the Granthami mutant, no gene or process has
been identified to be responsible for these phenotypes. Both mutants are found in
economically important species and show potential for crop improvement, either through
the production of more latex, or through the study of the pathology in grape. These
examples emphasize the purpose of the current study, in which gene identification and
the resulting association with gene function or phenotype can help to open doors in
numerous directions, which are as yet underdeveloped. Therefore, once the gene is linked
to the corky phenotype, it will be the first gene identified for causing such a phenotype.

38
2.5.2 FM#2 represents a novel gene of unknown function
corky’s single insertion site was determined to be located near three unknown genes. To
test which may be misregulated and therefore possibly responsible for the corky
phenotype, qRT-PCR was carried out on these nearby predicted genes. As corky has
much more bark tissue than wildtype, any transcripts specifically expressed in these
tissues would seem to be upregulated in full stem RNA isolates. However, it was
expected that the transcript of the causative gene would be upregulated in the tissues that
demonstrate the altered phenotype, and as such, RNA was isolated from xylem, phloem-
cambium and cortex. This allows direct examination of the altered tissues, while still
being comparable to wildtype control, thus lowering the rate of false positive detection of
misregulation. Upon investigation, FM#2 was substantially upregulated in xylem, phloem
and outer bark tissues. The polypeptide encoded by FM#2 contained no known motifs,
nor similarity to other previously identified proteins. However, while it shares no
homology to identified proteins, it does show conservation across plant species,
especially in the C-terminus, which is predicted to form a transmembrane helix. FM#2 is
predicted to be a member of a small gene family with four other poplar genes, a finding
corroborated by the prediction of a similar gene family when looking at their respective
Arabidopsis homologs. A related gene identified in tobacco gave the only functional
information to be obtained through homology studies. While still of unknown function,
this gene was identified as 'ACRE180' (Avr/Cf9 Rapidly Elicited 180) a gene which was
upregulated during a pathogenic response screen (Rowland, et al., 2005). The closest

39
homolog of this gene in Arabidopsis was found to be At1g46295, a member of the
Arabidopsis FM#2 gene family. Upon closer examination, another member of the FM#2
Arabidopsis gene family, At1g06475, was also implicated in the Avr/Cf9 screen
(Durrant, et al., 2000). The majority of the information returned by bioinformatics tools
(ex. SUBA and Athena) was below their respective levels of significance. Physical
parameters of the predicted protein (i.e. its size and hypothetical isoelectric point) suggest
a small acidic protein. It has been previously documented that various small basic
proteins were linked to leucine-rich repeat receptor-like kinase (LRR-RLK) signalling -
which is hypothesized to be related to growth and development (Beveridge, et al., 2007).
CLAVATA1 (CLV1) and BIG APICAL MERISTEM 1 (BAM1), two of the more than
200 identified LRR-RKs, have been shown to modulate developmental signals in
Arabdiopsis (DeYoung, et al., 2006; Dievart, et al., 2003). Specifically CLV1 initiates a
signalling cascade when it interacts with the secreted protein CLAVATA3 (CLV3)
(Ogawa, et al., 2008). CLV3 is a short (96 amino acid), basic protein that is cleaved into
an active 12 amino acid peptide prior to interaction with CLV1. Knock out mutants of
CLV3 show gross malformation of the shoot apical meristem, showing that this small
protein is very important to the integrity of meristematic development (Clark, et al.,
1995). CLV3 is a member of a large family of genes which show sequence similarity at
the C-terminus, dubbed the CLE (CLAVATA3/ESR-related) family (Cock and
McCormick, 2001; Opsahl-Ferstad, et al., 1997). They generally include an N-terminal
hydrophobic region and are often overlooked by most gene annotation programs due to

40
their small size, suggesting that there could be many more than previously identified.
FM#2 is similar to these genes in that its' predicted to encode a protein which is very
small, charged and has a terminal hydrophobic region. If it can be shown that FM#2 is
actually the gene responsible for the corky phenotype, it could prove to be the first
member of a family of meristematic- signalling ligands. However, this is purely
speculative at this point. Before FM#2 can be linked to the phenotype, however, the
phenotype must be recapitulated by overexpressing FM#2.
2.5.3 Recapitulation of the corky phenotype to establish gene-phenotype link
While the upregulation of FM#2 in the phenotypically-mutant tissues would suggest that
the two are linked, it is still necessary to prove that overexpression of the single gene is
sufficient to cause the phenotype. Recapitulation can be a complicated task for many
reasons and there are pitfalls to each recapitulation strategy. The best way to overcome
this is to use multiple strategies. In this study three types of recapitulation are currently in
progress. The first is a 'stable' transformation, being the same type of transformation as
was used to produce the activation-tagged population in the first place. The second is a
new technique from collaborators in Melbourne, Australia, known as induced somatic
sector analysis (ISSA). The third is a more traditional heterologous system
transformation, in which poplar genes (or their Arabidopsis homologs) can be
transformed into Arabidopsis to see if phenotypes are as predicted. To date, there has
only been one gene identified from activation tagging efforts in poplar (Busov, et al.,
2003). In this study however, the recapitulation was carried out using a giberellic acid 2-

41
oxidase gene (PhGA2ox2) from bean (Phaseolus coccineus) rather than from poplar.
Thus, to date there have been no reported successful recapitulations of poplar activation-
tagged genes in the literature.
Stable transformations involve the transformation of plant cells with Agrobacterium in
sterile tissue culture conditions. After the transformation phase, tissue cuttings are
screened on antibiotic-containing media, which also include auxins to induce callus
formation. All cells which are not transformed die off, and at the end of the
transformation, each plant is fully transgenic, having completely developed from the
single cell which initiated the callus. This recapitulation strategy is by far the most likely
to result in phenotypic transgenic lines. It is pliable in that both wildtype and mutant lines
can be used as the initial tissue source with overexpressor and antisense constructs to
produce or ameliorate the mutant phenotype respectively. However, a draw back to this
strategy is the amount of time it takes to go from callus to regenerated plant. It takes
roughly nine months in tissue culture before poplar samples are placed in soil. In the case
of corky this strategy will also require at least six to twelve months of growth in soil
before the phenotype will be readily visible, as cork production is not obvious until the
tree reaches considerable size.
The ISSA strategy contrasts the stable transformation method. ISSA involves inoculating
Agrobacterium harbouring the constructs directly onto the exposed vascular cambium of

42
tree species while in soil. After the wound response and the healing of the 'cambial
window', regular lateral growth is reinitiated. During this time, any cells which were
transformed will divide as per usual. If cells in the cambium are transformed, xylem and
phloem cells will be produced which will also be transgenic, thus radial files of cells will
develop expressing the construct. This technique has only a four month growing period
and doesn't involve direct hormone and antibiotic treatments as does the stable
transformation method. While it may be possible to see an altered phenotype in the
transformed window, drawbacks to this strategy include that it is limited to genes and
proteins that function in the stem, and that at best, only specific files of cells are
transformed, making phenotypic analysis more difficult in some cases.
Finally, in the heterologous system, poplar genes (or their homologs) can be expressed in
different fast-growing species, such as Arabidopsis. This leads to transgenic lines more
quickly than either of the other two methods. However, not every gene in poplar will be
present in Arabidopsis (and vice versa) which could lead to false negative results.
Additionally, the closest homolog may serve a different function in another species.
As all of these methods have clear benefits, all three have been initiated in this study. For
the stable method, both an overexpression construct and an antisense construct have been
created and are currently ready to begin transformations into both wildtype and corky
backgrounds. As mentioned previously, this is to both recreate the corky mutant

43
(overexpression in wildtype) and to ameliorate the corky phenotype (antisense in mutant
background). Using the ISSA method, 4 constructs (a sense, antisense, hairpin-RNAi and
truncated hairpin-RNAi) have been produced and were transformed into poplar as
summarized in Appendix S. Finally, the two Arabidopsis homologs have been cloned and
used to create overexpression constructs. SALK knockout lines have previously been
ordered and screened for homozygosity and only need to be grown out to see the effect of
knocking out these two genes as well.
While these transformations are currently in various stages of completion, the molecular
aspects of the work are nearly completed. As the transformations reach phenotypic
developmental stages, the outcomes of the various strategies will be determined.
However, as the specific results from each strategy are not available yet, it remains to be
seen if FM#2 is actually the causative agent for the corky phenotype.
2.6 Conclusion
The identification and anatomical characterization of a novel mutant of poplar was
described. The line, named corky, for its abundant cork production was found to show
aberrations at the whole plant, tissue and cellular levels of the stem. Also, the
hypothesized causal agent for the phenotype was described. This previously unknown
gene, currently named FM#2, is possibly linked (indirectly) to pathogen response, but
shows no conserved domains, nor homology to previously identified genes. Various
longterm recapitulation strategies are in progress to confirm FM#2's role in corky.

44
Chapter 3
Do shoot apical meristem identity proteins regulate the vascular
cambium? Evidence for a role of CLAVATA1 in the regulation of
secondary growth in Arabidopsis and Populus.
3.1 Abstract
The shoot apical meristem (SAM) produces all above-ground post-embryonic
organs in higher plants, such as leaves and stems, including the primary vasculature. The
secondary vasculature (secondary xylem and phloem) is subsequently produced by
another meristematic tissue known as the vascular cambium. It has previously been
shown that the growth of the shoot and the floral meristems are negatively regulated by
the CLV1 protein, such that the dominant negative clv1 mutant of Arabidopsis has
unusually large floral and shoot apical meristems. In this study, we have investigated
whether CLV1 also plays a role in the regulation of the vascular cambium. The clv1
mutant produced more secondary xylem than wildtype, indicating that CLV1 may also
negatively regulate the vascular cambium. This finding is particularly significant to forest
biotechnology since increasing xylem production is synonymous with a greater yield of
wood. As a first step towards increasing xylem production in a tree, we show here that
downregulation of CLV1 via antisense in Arabidopsis is sufficient to produce the
increased secondary xylem phenotype seen in the original missense clv1 mutant. Out of
41 transgenic lines, 8 lines showed distinctive clv1-like phenotypes related to the shoot

45
apical and floral meristems. Transgenic lines with strong floral phenotypes also had
increased amounts of secondary xylem, indicating that the antisense strategy is sufficient
to produce the desired clv1-like phenotype. In Populus, a recently adapted model for
studying various traits, including secondary growth, CLV1 has two predicted homologs.
This suggests a slightly more complex regulation than is found in Arabidopsis. The CLV1
homologs, as well as a group of CLV1-like genes were downregulated in poplar using a
recently reported method of transformation to determine if these genes play similar roles
in trees.
3.2 Introduction
The vascular cambium is a cylindrical meristem that produces secondary xylem and
phloem. In angiosperm and gymnosperm trees, the secondary xylem matures to become
wood, an important natural resource. Like other meristems, the control of cell
proliferation within the vascular tissue is believed to be largely under genetic regulation
(Jenik and Irish, 2000; Meyerowitz, 1997), but is less understood since the meristem is
difficult to isolate due to several technical challenges (Chaffey, 2002).
Arabidopsis undergos secondary growth (Gendreau, et al., 1997); (Dolan and Roberts,
1995); (Lev-Yadun, 1995) and has a vascular cambium (Busse and Evert, 1999). The
utility of Arabidopsis as a model system to study secondary growth and the vascular
cambium has been demonstrated (Chaffey, et al., 2002). Chaffey et al. (2002) found that

46
Arabidopsis undergoes secondary growth in the hypocotyl, especially under short day
conditions. Also, they found that the secondary xylem of Arabidopsis closely resembles
that of the wood of an angiosperm tree, and thus, can be used to address basic questions
about wood formation. This finding offers new prospects for the study of cambial growth
and secondary xylem development. However, in using Arabidopsis there are numerous
drawbacks, including amongst others, the small amount of secondary tissues which are
actually produced as well as the technical difficulties which emerge from working with
such small samples.
Therefore, to study the vascular cambium, the larger process of secondary growth as well
as the various other traits which are more easily observed in trees, poplar has emerged as
a key model. This is due to a number of large-scale EST identification projects (Kohler,
et al., 2003; Sterky, et al., 1998; Sterky, et al., 2004), the full sequencing of the Populus
genome (Tuskan, et al., 2006), its' relatively fast growth and the ability of many poplars
to be clonally propagated. To date, studies have focused on identifying genes responsible
for cell wall constituents such as lignin (Rogers and Campbell, 2004), cellulose (Taylor,
2008) and hemicellulose (Mellerowicz and Sundberg, 2008), and only recently on the
regulation of the vascular cambium (Druart, et al., 2007; Hertzberg, et al., 2001; Oh, et
al., 2003; Schrader, et al., 2004a; Schrader, et al., 2004b; Zhao, et al., 2005).

47
One of the major strategies to identify potential developmental regulators of the vascular
cambium in trees is the analysis of microarray data from across the phloem, cambium and
wood forming tissues (Hertzberg, et al., 2001; Schrader, et al., 2004b). Included in these
studies are those which compare the expression values temporally, utilizing the active
and dormant cycles of secondary growth in a temperate climate (Druart, et al., 2007).
This is further augmented with the bioinformatical analysis of ESTs obtained from these
same tissues (Sterky, et al., 1998). Another alternative strategy used to identify new
regulators of the vascular cambium, is to look at previously identified developmental
mutants and to assay them again for vascular phenotypes which can be easily missed if
not grown under the proper conditions. Through all of these strategies, data that was
previously collected is reviewed to help shed light on the current questions of regulatory
identity. And while an alternative forward genetics approach could also be attempted,
here we report on a case where a known regulator of the SAM was assayed for
phenotypes of the vascular cambium.
The regulation of the shoot apical and floral meristems in Arabidopsis have been well
characterized, and both have been shown to be negatively regulated by the leucine-rich
repeat receptor like kinase (LRR-RLK) CLAVATA1 (CLV1) (Clark, et al., 1993). CLV1 is
expressed specifically on the periphery of the central region in the SAM and in a similar
region of the early floral meristem. Furthermore, CLV1 expression in the floral meristem
changes preceding organ initiation (Smyth, et al., 1990). In wildtype plants, the

48
proliferation and differentiation of the SAM and floral meristem are balanced, so that the
structures and sizes of the meristems are maintained (Clark, et al., 1997). However,
Arabidopsis plants homozygous for loss of function mutations at the CLV1 locus
accumulate excess undifferentiated cells, suggesting that CLV1 plays an important role in
regulating the growth of these meristems (Clark, et al., 1997). However, while there have
been phenotypes described for the shoot and floral meristems, no literature has been
found which describes possible changes in the lateral meristems (vascular and cork
cambiums), nor the root apical meristem.
While, CLV1 is a known negative regulator of the SAM, there have been many other
factors implicated in controlling meristem growth and differentiation. Those regulating
meristem growth include (amongst others) WUSCHEL (WUS) (Laux, et al., 1996),
SHOOTMERISTEMLESS (STM) (Lenhard, et al., 2002), SHOOTLESS1 and 2 (Satoh,
et al., 2003), ZWILLE/PINHEAD (Lynn, et al., 1999; Moussian, et al., 1998), BIG
APICAL MERISTEM1, 2 and 3 (BAM) (DeYoung, et al., 2006; Hord, et al., 2006), and
POLTERGEIST (POL) (Pogany, et al., 1998; Yu, et al., 2000). These all work
antagonistically with CLV1, and attempt to increase or maintain meristem size. There are
various negative regulators of the SAM as well, including ULTAPETELA1 (Carles, et
al., 2005), CLAVATA2, CLAVATA3 (Clark, et al., 1997) and CORYNE (CRN)
(Muller, et al., 2008), which function in a similar manner as CLV1. A signalling pathway
has been predicted in which CLAVATA 3 (CLV3) binds a CLAVAT1/CLAVATA2

49
heterodimer, initiating a signalling cascade resulting in the repression of WUSCHEL and
leading to stem cell differentiation (Clark, 2001). Recently it has been reported that
CLAVATA2 (CLV2) may instead interact with CORYNE. However, as mutants for any
of the CLAVATAs are able to affect SAM regulation, it is likely that they play a
dominant role in the signalling cascade. The BIG APICAL MERISTEM or BAM family
is represented in Arabidopsis by three members, BAM1, BAM2 and BAM3 (DeYoung,
et al., 2006). The BAM proteins are distinct from many other meristematic regulators in
that they are expressed in numerous tissues (DeYoung, et al., 2006). They also appear to
have an opposing function to that of the CLAVATAs, even though they actually have
very homologous sequences, sharing 55, and 49% identity with CLV1 over the length of
the entire protein (DeYoung, et al., 2006). It is hypothesized by DeYoung (2006) that it is
due to their expression pattern that they have a different role to that of CLV1, as it was
found that partial complementation between bam1, bam2 and clv1 was achievable.
Furthermore BAM genes are expressed in regions surrounding the central zone where
CLV1 exists on the periphery (DeYoung, et al., 2006), but are not found within.
As mentioned previously, we know much less about the genes regulating the vascular
cambium than about those of the SAM and floral meristem. As CLV1 is known to affect
the development of shoot apical and floral meristems, and since the vascular cambium is
derived from the SAM, it was hypothesized that CLV1 may also play a regulatory role in
the vascular cambium. However, the systems are likely not completely analogous, as it

50
was recently shown that while CLV1 transcript was identified in the vascular cambium
where CLV3 and WUS, two of its SAM signalling partners, were not identified (van
Raemdonck, et al., 2005). This suggests that perhaps a modified signalling pathway
might exist. We show here that the diameter of the secondary vascular tissue is greater in
clv1-1 lines when compared to wildtype, suggesting that CLV1 does regulate the vascular
cambium. As the ultimate goal of this research is to understand the regulation of wood
formation in trees, we test here whether an antisense approach is sufficient to cause the
increase in secondary vascular development that is seen in the clv1-1 mutant. The success
of this approach in Arabidopsis shows promise for the application to forest trees such as
poplar.
3.3 Materials and Methods
3.3.1 Plant material, growth and transformation
Arabidopsis thaliana (L.) seeds were surface sterilized and grown at 20-22 oC in either
long day (16 hr light/8 hr dark) or short day (8 hr light /16 hr dark) conditions.
Arabidopsis (Col) was transformed via the floral dip methodology (Clough and Bent,
1998) using Agrobacterium tumefaciens (strain LBA4404). Poplar (Populus alba x P.
tremula) were grown in the greenhouse under previously reported conditions (Harrison,
et al., 2007) and transformations were carried out using the 'induced somatic sector
analysis' (ISSA) methodology described below (Van Beveren, et al., 2006).

51
3.3.2 Plasmid construction for production of antisense-CLV1 lines in Arabidopsis
Total RNA was extracted from whole 10 day-old plants grown in the short day conditions
and cDNA was subsequently synthesized. A 605 bp fragment, which corresponds to
nucleotides 482 to 1087 of the CLV1 (At1g75820) mRNA, was amplified with primers
AtCLV1anti-FW and AtCLV1anti-RV (see Appendix L). The PCR products were TA-
cloned into the pGEM-T Easy vector (Invitrogen, Carlsbad, CA), excised with KpnI and
BamHI and subcloned into pSOL2. The cauliflower mosaic virus
35S:A5TISE5SECLV1:NOS cassette was then excised using the two 5ot1 sites and
inserted into the single 5otI site of pSOL6 to create the plasmid 'pSH1'. Transformations
were carried out as described above using Agrobacterium harbouring the pSH1 plasmid.
3.3.3 �orthern blot analysis
Total RNA was isolated from floral tissues of wildtype (Col) and CLV1-antisense lines of
Arabidopsis. RNA was also obtained from hypocotyl tissue in the wildtype control. A
Northern blot was prepared using standard protocols. The RNA gel blot was hybridized
with a 605 bp probe, created from a region of the CLV1 gene using the AtCLV1probe-
FW and AtCLV1probe-RV primers (see Appendix L). The probe was produced using the
Rediprime II DNA labeling system (Amersham-Biosciences now GE, Mississauga, ON),
and labeled with α-32
P dCTP as per manufacturer's instructions. Hybridization and
detection were as per Church's methodology (Church and Gilbert, 1984).

52
3.3.4 Preparation of the hand section of hypocotyls and stems
Cross section of hypocotyls of both transgenic and wildtype plants were made by hand
and stained with Toluidine Blue O pH 4.4 as per typical methodologies. Measurements of
hypocotyl diameters were made below the rosette using calipers. Measurements of xylem
and phloem/cortex diameters were obtained from magnified tissues.
3.3.5 R�Ai construct to downregulate PtCLV1 in poplar
An RNAi strategy was utilized to try to downregulate PtCLV1 in poplar. Prior to the
publication of the complete poplar genome, the 179 bp construct was produced based on
CLV1-like ESTs, using the primers CLV1pop01 and CLV1pop02 (Appendix L) and then
transformed via typical Agrobacterium-mediated transformation into the Populus alba, x
P. tremula hybrid (called 717) (Harrison, et al., 2007). Trees were grown for more than
two years in the greenhouse prior to phenotypic and molecular investigation.
3.3.6 Induced somatic sector analysis (ISSA) as a transformation strategy
A total of eight constructs were created to use in ISSA transformations (Van Beveren, et
al., 2006) to determine the impact of downregulating CLAVATA1/CLAVATA1-LIKE
genes in the vascular cambium of poplar. For each of the four target genes (PtCLV1b,
PtBAM1, PtBAM1-LIKE and PtBAM3), both an antisense and a hairpin-RNAi construct
were created. Primers used in the amplification of the fragments are shown in Appendix
M. The gene fragments (see Appendix N) were each originally amplified using Platinum
PFx DNA Polymerase (Invitrogen; Carlsbad, CA) and cloned into an entry vector using

53
the pCR8/GW/TOPO TA Cloning Kit (Invitrogen; Carlsbad, CA). Using Gateway®
technology (Invitrogen; Carlsbad, CA), the eight expression vectors were created using
pre-established single or double cassette (antisense or hairpin-RNAi respectively),
gateway-enabled pCAMBIA1305.1 vectors (for vector maps, see Appendices E and F).
The plasmids were transformed into Agrobacterium (strain AGL1). Poplars growing in
the greenhouse were prepared for transformation by creating “cambial windows”. This
was accomplished by making two vertical cuts in the bark with a scalpel. A horizontal cut
was made between the two vertical cuts, joining them at their basal end. The flap of bark
was then lifted and the trees were directly inoculated with Agrobacterium harbouring one
of the eight constructs, or one of the two empty vectors. Following inoculation, cambial
windows were shut and sealed with parafilm to promote healing of the wound and
subsequence growth. Trees were transformed in the summer and left to grow for 4
months before sampling took place.
3.4 Results
3.4.1 Comparison of the secondary xylem and phloem in the hypocotyl of wildtype
and clv1-1 plants in Arabidopsis
The link between CLV1 and secondary growth was first observed while looking at
hypocotyl cross sections of the Arabidopsis clv1-1 mutant. This line contains an EMS
derived point mutation, which results in a G856D missense mutation (Clark, et al., 1997).
Wildtype plants produced much less secondary growth than plants of the clv1-1

54
background (Figure 5). Measurements of radii were obtained to quantify the change in
secondary growth and the averages of these measurements are also shown in Figure 5.
From this data, it was hypothesized that disruption of CLV1 expression levels,will result
in the production of more secondary growth in tree species. One goal of this study was to
determine if the phenotype could also be observed in poplar, but as creating a similar
point mutation in poplar CLV1 (PtCLV1) isn't feasible, a dominant mutation strategy was
needed. Antisense downregulation was attempted first in Arabidopsis to determine if this
technique would be able to phenocopy the EMS and insertional-mutagenesis techniques
previously used to create the original clv1mutants. Following the successful test, this
strategy was then attempted in poplar.
3.4.2 Detection of clv1-1 phenotypes in antisense-CLV1 lines of Arabidopsis
To test whether it is possible to mimic the clv1-1 phenotype by a transgenic approach, a
fragment of the wildtype CLV1 was expressed under the cauliflower mosaic virus 35S
promoter in an antisense orientation. A total of 41 antisense transformed transgenic plants
were obtained, and all were analyzed for phenotypes indicative of a clv1-1-like mutant
phenotype. Eight antisense lines, which accounted for 20% of total antisense transformed
plants, mimicked the phenotype of the clv1-1 mutant and were designated as
Anti_L(number). This suggests that expression of the antisense construct, whose
sequence is complimentary to a region of the leucine rich repeat (LRR) motif of the clv1
gene, results in a phenotype similar to the strongest CLV1 mutant alleles, clv1-4 and clv1-
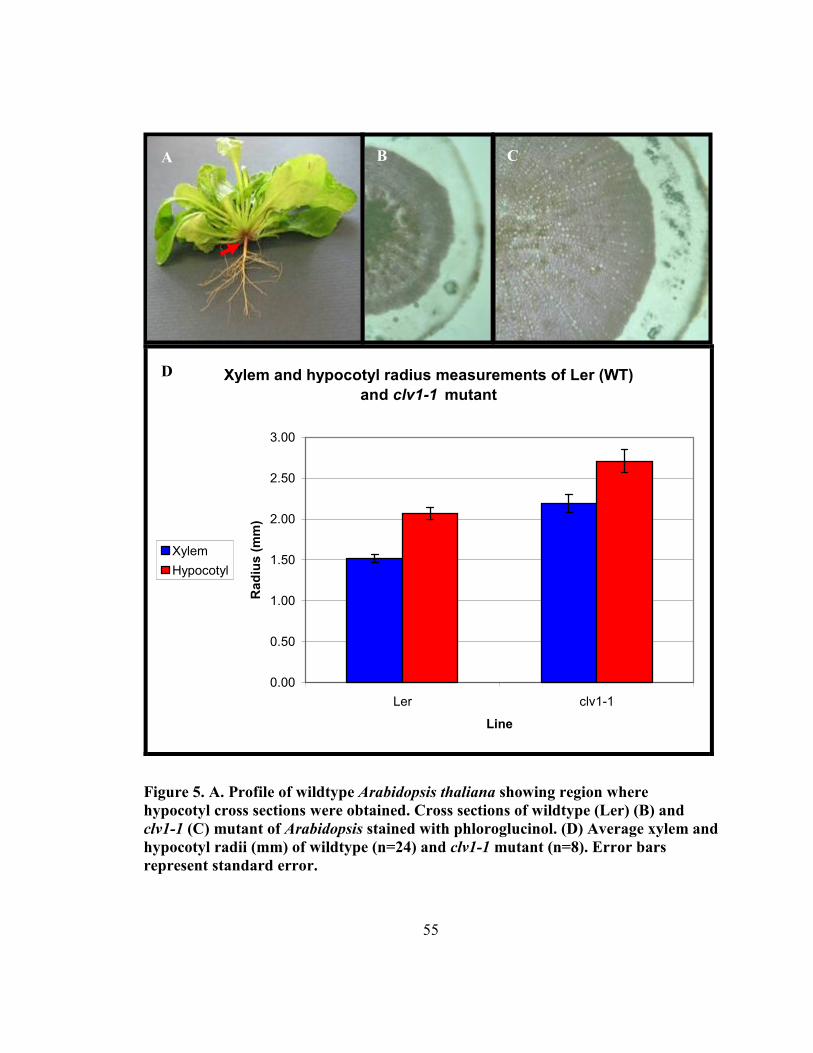
55
Xylem and hypocotyl radius measurements of Ler (WT)
and clv1-1 mutant
0.00
0.50
1.00
1.50
2.00
2.50
3.00
Ler clv1-1
Line
Ra
diu
s (
mm
)
Xylem
Hypocotyl
Figure 5. A. Profile of wildtype Arabidopsis thaliana showing region where
hypocotyl cross sections were obtained. Cross sections of wildtype (Ler) (B) and
clv1-1 (C) mutant of Arabidopsis stained with phloroglucinol. (D) Average xylem and
hypocotyl radii (mm) of wildtype (n=24) and clv1-1 mutant (n=8). Error bars
represent standard error.
A B C
D

56
8. These alleles both contain missense mutations in the LRR domain (Clark et al., 1997).
Typically, Anti_L8, 9, 16, 18, 23, 27, 30, and L32 had extra petals, thus mimicking the
clv1-1 floral phenotype (Figure 6). Two lines of interest included Anti_L22 which had 3
petals, and Anti_L38 which had deformed petals. More specifically, the angle between
the petals of line Anti_L38 was changed, while the angle between the petals of wildtype
plant is always 90o.
In the T1 generation, the plant height (cm), silique length (cm), and stem diameter (mm)
were compared between wildtype plant (n=16) and antisense transformed plants (n= 41).
The results showed that transgenic plants (49.68cm) were approximately 6 cm higher
than wildtype plants (43.56 cm), and this difference was significant (P<0.01); in terms of
the length of silique, antisense transformed plants (1.16cm) were significantly shorter
than that of wildtype plants (1.33cm) (P<0.01), but antisense transformed plants did not
display the club-shaped phenotype that is found in the clv1-1 mutant in the Landsberg-
erecta background. And the diameter of the stem (1.29 mm) of transgenic plants was
bigger than wildtype plants (0.99 mm) (P<0.01), indicating that transgenic plants
produced more secondary growth than wildtype plants did.
Furthermore, the floral meristem of Anti_L41 was chosen to be compared with that of
wildtype, and it was found that it was enlarged compared to that of wildtype (data not
shown). This is consistent with the clv1 floral meristem, which can be twice as tall as the
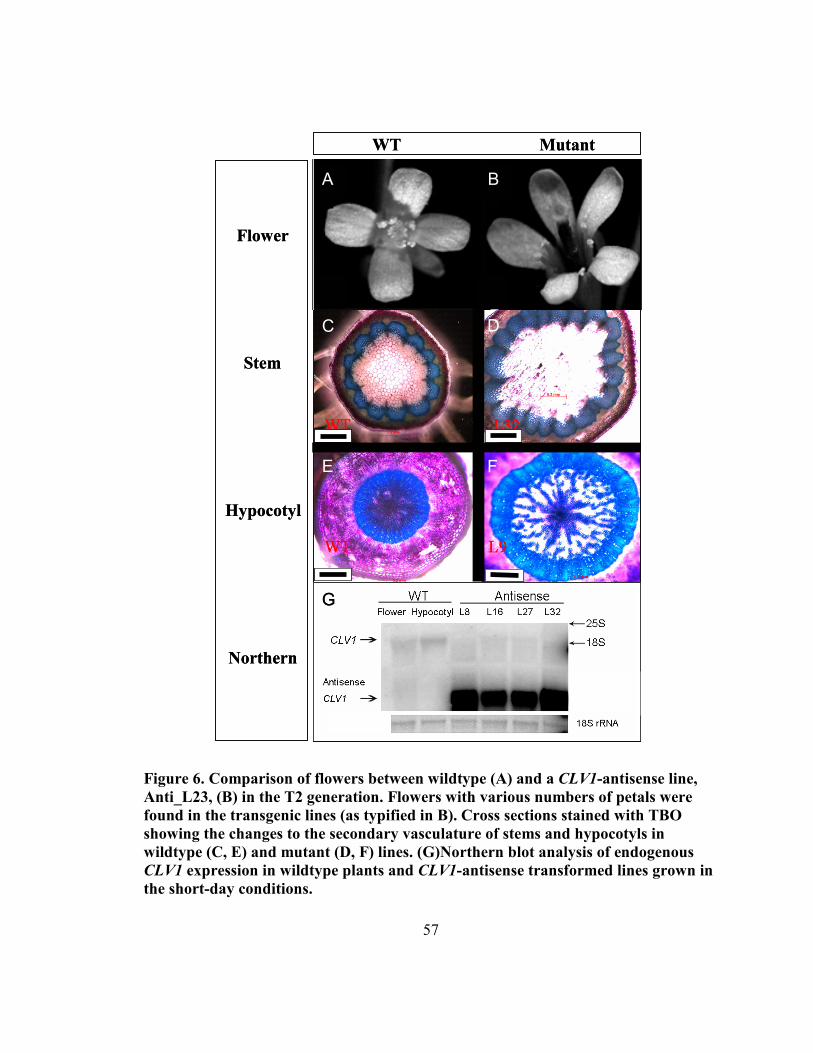
57
Figure 6. Comparison of flowers between wildtype (A) and a CLV1-antisense line,
Anti_L23, (B) in the T2 generation. Flowers with various numbers of petals were
found in the transgenic lines (as typified in B). Cross sections stained with TBO
showing the changes to the secondary vasculature of stems and hypocotyls in
wildtype (C, E) and mutant (D, F) lines. (G)�orthern blot analysis of endogenous
CLV1 expression in wildtype plants and CLV1-antisense transformed lines grown in
the short-day conditions.
WT Mutant
C D
E F
Flower
Stem
Hypocotyl
�orthern
WT Mutant
B A
B A
B A
B A
C D
E F
Flower
Stem
Hypocotyl
�orthern
G G

58
wildtype floral meristem at the earliest stage of organ initiation (Clark et al., 1997).
Analysis of maturing flowers revealed that the clv1 floral meristem continued to
proliferate and gave rise to a large mass of undifferentiated cells in the center of the
flower, instead of terminating like the wildtype floral meristem.
The segregation pattern of T2 plants were examined by screening the seeds on MS solid
media augmented with kanamycin. It was found that of the progeny of lines Anti-L2, 3,
9, 10, 11, 16, 19, 20, 22, 24, 30, 31, 37, 38, 39, 41, roughly 25% were sensitive to the
kanamycin (KanS), following typical Mendelian inheritance patterns. From this it was
predicted that these plants contain a single copy of the transgene. On the other hand, the
proportion of KanS plants in other transgenic lines, for instance, Anti-L1, 5, 8, 32, were
less than 5%. These transformants displayed non-Mendelian segregation ratios with a
very high proportion of KanR plants among the progeny, suggesting these transgenic lines
contain multiple, unlinked copies of the antisense construct. These results were
confirmed by southern blot (data not shown). All of the aforementioned lines were
advanced to T3 generation in order to get homozygous mutants.
3.4.3 Antisense-CLV1 construct inhibits endogenous CLV1 expression
Under normal growth conditions, the flower is the only major organ of the plant in which
CLV1 mRNA is abundant (Smyth, et al., 1990). For this reason, flowers were selected as
the organ used to visually indicate alterations in CLV1 expression levels. To determine
CLV1 expression patterns in other tissues, total RNA was isolated from Arabidopsis root,
hypocotyl, stem, leaf, SAM, and flower. RT-PCR results showed that that CLV1 was

59
expressed in the flower and the shoot meristem, while no PCR products were found in the
root, hypocotyl, stem or leaf. To further explore if the CLV1 gene is expressed in the
vascular meristem of hypocotyl, total hypocotyl RNA was isolated from two-month-old
plant grown under the short day conditions. CLV1 was expressed in 2-month-old
hypocotyls (data not shown), indicating that the CLV1 gene may be involved in the
activity of the vascular meristem in the hypocotyl.
To determine the expression levels of clv1 in the antisense transformants, a Northern blot
was performed using a 605 bp CLV1 fragment probe (Figure 6G). In the control lanes, no
transgene signal was found in wildtype flower and hypocotyl. More importantly, the
CLV1 gene was more highly expressed higher in the wildtype flower and hypocotyl than
that of transformed lines. This suggests that the endogenous CLV1 transcript in
transformed lines was downregulated by the antisense construct.
3.4.4 Secondary growth in Arabidopsis hypocotyl is elevated by antisense
inhibition of CLV1
The xylem and phloem/cortex are easily distinguished in a hand-section stained with
Toluidine Blue O (TBO) (Figure 6). The secondary xylem stains blue as mature vessels
and fibers have thick secondary cell walls, while the phloem/cortex regions stain a deeper
pink shade due to the polychromatic nature of TBO. Wildtype and CLV1-antisense lines
were analyzed in cross section to see if they would show variation in their secondary
growth patterns. In the stem, an increase in the number of vascular bundles was

60
detectable. In the hypocotyls, there was more secondary growth than in wildtype. Ten
antisense lines and one wildtype (Col) line were chosen for measuring the total xylem
and hypocotyls radii with each line represented by at least 6 plants. The phloem/cortex
radius was determined by subtracting the xylem radius from the radius of the hypocotyl.
The results are shown in Appendix O and graphically represented in Figure 7 (A). As the
radius is actually a linear representation of the stem, simple calculations were carried out
to determine the change to the overall area of the xylem and phloem tissues in the cross
sections. These are represented in Figure 7 (B). Both the radii of the xylem and the
phloem/cortex were greater than those found in wildtype plants. For instance, xylem
radius of Anti_L10 line has been increased by 59% compared to wildtype line. The
average xylem radius of 10 antisense transformed lines was increased by 40% compared
to wildtype controls, and the average phloem/cortex radius was increased by 39%. T-test
results show that xylem radius of all antisense transformed lines except for Anti_L18 and
L30 are significant greater than that wildtype plants (P<0.01), and phloem/cortex radius
of Anti_L9, 27, 30, and 32 are bigger than that of wildtype plants (P<0.05). These results
indicate that disruption in the CLV1 locus causes an increase in the radius of xylem and
phloem, leading to a significant increase of wood formation. Thus the control of both
secondary xylem and phloem formation is likely negatively regulated by CLV1.
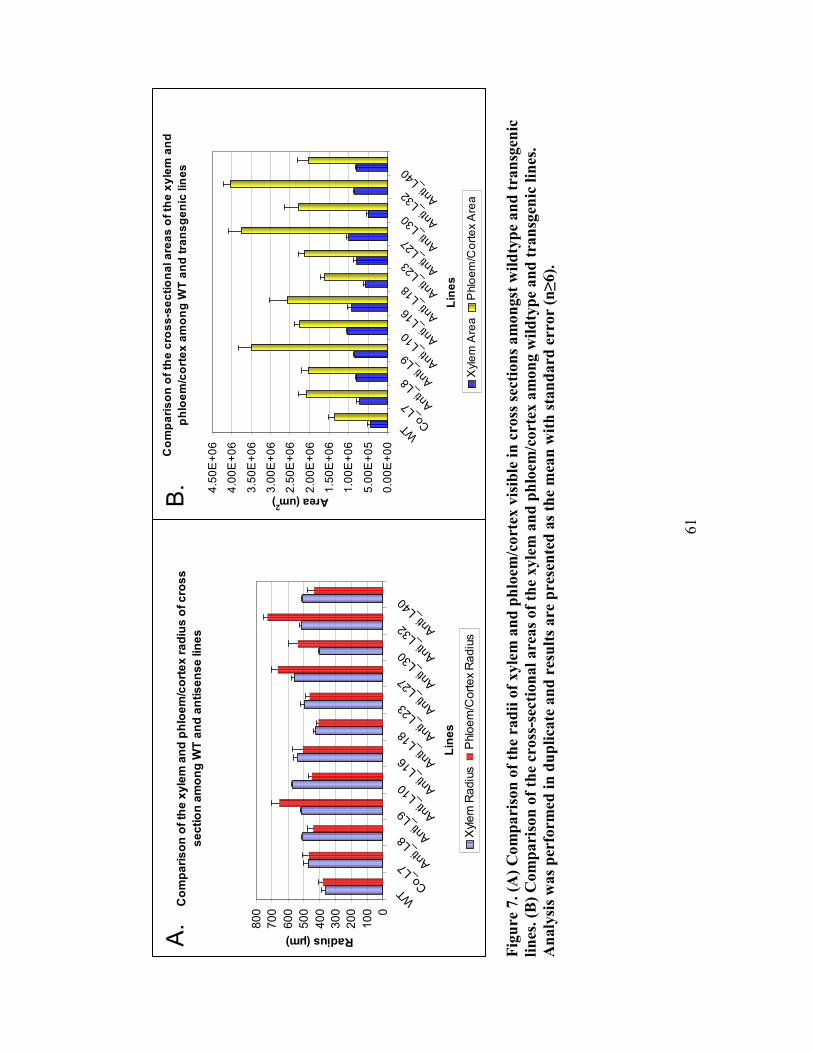
61
Fig
ure
7.
(A)
Co
mp
ari
son
of
the
rad
ii o
f xyle
m a
nd
ph
loem
/co
rtex
vis
ible
in
cro
ss s
ecti
on
s a
mo
ngst
wil
dty
pe
an
d t
ran
sgen
ic
lin
es. (B
) C
om
pari
son
of
the
cross
-sec
tion
al
are
as
of
the
xyle
m a
nd
ph
loem
/corte
x a
mon
g w
ild
typ
e an
d t
ran
sgen
ic l
ines
.
An
aly
sis
was
per
form
ed
in
du
pli
cate
an
d r
esu
lts
are
pre
sen
ted
as
the
mea
n w
ith
sta
nd
ard
err
or
(n≥
6).
Co
mp
ari
so
n o
f th
e x
yle
m a
nd
ph
loe
m/c
ort
ex
ra
diu
s o
f c
ros
s
se
cti
on
am
on
g W
T a
nd
an
tis
en
se lin
es
0
100
200
300
400
500
600
700
800
WT
Co_
L7Ant
i_L8
Ant
i_L9 A
nti_
L10 A
nti_
L16 A
nti_
L18 A
nti_
L23 A
nti_
L27 A
nti_
L30 A
nti_
L32 A
nti_
L40
Lin
es
Radius (µm)
Xylem Radius
Phloem/C
ortex Radius
Co
mp
ari
so
n o
f th
e c
ros
s-s
ec
tio
na
l a
rea
s o
f th
e x
yle
m a
nd
ph
loe
m/c
ort
ex
am
on
g W
T a
nd
tra
ns
ge
nic
lin
es
0.00E+00
5.00E+05
1.00E+06
1.50E+06
2.00E+06
2.50E+06
3.00E+06
3.50E+06
4.00E+06
4.50E+06
WT C
o_L7
Ant
i_L8 A
nti_
L9 Ant
i_L1
0 Ant
i_L1
6 Ant
i_L1
8 Ant
i_L2
3 Ant
i_L2
7 Ant
i_L3
0 Ant
i_L3
2 Ant
i_L4
0
Lin
es
Area (um2)
Xylem Area
Phloem/C
ortex Area
A.
B.

62
3.4.5 Comparison of stem radius values between wildtype and antisense lines
To test if there was also a change in the activity of the vascular cambium of the stem, the
radius of stem and the number of bundles was measured at the base and number of
bundles was counted based on the cross sections as seen in Appendix P. It was found that
there was an increase in the number of vascular bundles between wildtype and antisense
lines. Also, there was an increase in the radius and subsequent area of secondary xylem in
the antisense lines. On average, a 38 % increase in radius was observed in antisense lines
when compared to wildtype. T-test results show that the stem radii of Anti_L8, 9, 16, 18,
23, and 30 were significantly larger than that of wildtype plants (P<0.01).These results,
when combined with northern blot analysis suggest that CLV1 has been successfully
downregulated at the transcript level in transgenic plants, and that the resulting phenotype
is similar to the strong alleles of the original clv1 mutants. As a result of the decrease in
CLV1 expression, the floral meristems were affected as was shown by the change in the
floral architecture. Downregulation also resulted in an increase in cambial meristem
activity, which caused an increase in the production of both secondary xylem and phloem
in the hypocotyl, indicating that CLV1 is a negative regulator of the vascular cambium.
3.4.6 R�Ai to downregulate PtCLV1 in Populus
To see if CLV1 could also affect the amount of secondary xylem produced in trees, a
construct was produced to downregulate the gene in poplar. At the time, the poplar
genome was not yet available, and as such, an EST with high identity to AtCLV1 was
chosen to design an RNAi construct. The resulting 8 RNAi lines were tested for

63
downregulation of PtCLV1 via qRT-PCR, however, no change was detected (data not
shown). The lines were also screened for phenotypes in the stem, however no mutant
phenotypes were observed (data not shown). Concurrent with these studies was an effort
to link the EST to the recently released poplar genomic sequence. It was determined that
the "PtCLV1" RNAi construct actually showed higher identity to a PtCLV1-like gene,
PtBAM1-LIKE. As such, a new attempt was made to downregulate PtCLV1.
3.4.7 Arabidopsis CLV1 has two potential homologs in Populus
To see if the antisense strategy in Arabidopsis could also alter the expression of CLV1
and the subsequent change in vascular cambium regulation, a similar strategy was
developed for use in Populus. The first stage of this work was the determination of which
gene(s) in poplar would be targeted for downregulation. Shown in Table 4 are various
genes related to CLV1 signalling in the SAM. Also included are their potential homologs
in poplar, as predicted by the populusDB. Expression data from Arabidopsis
shoot/hypocotyl and for the vascular cambium of poplar were obtained from recent
papers. It appears that there are in fact two homologs to CLV1 in poplar. There are also
many highly related CLV1-like sequences. Of these, two genes homologous to
Arabidopsis BAM1, which is very closely related to CLV1, are also conserved and highly
consistent with PtCLV1 (Figure 8). A protein sequence ClustalW/PRALINE is available
for the Arabidopsis and Populus homologs of CLV1, BAM1 and BAM3 in Appendix Q.
As such, PtCLV1, PtBAM1, PtBAM1-LIKE and PtBAM3 were all targeted for
downregulation using ISSA. These constructs were designed to be roughly 200-350 bp
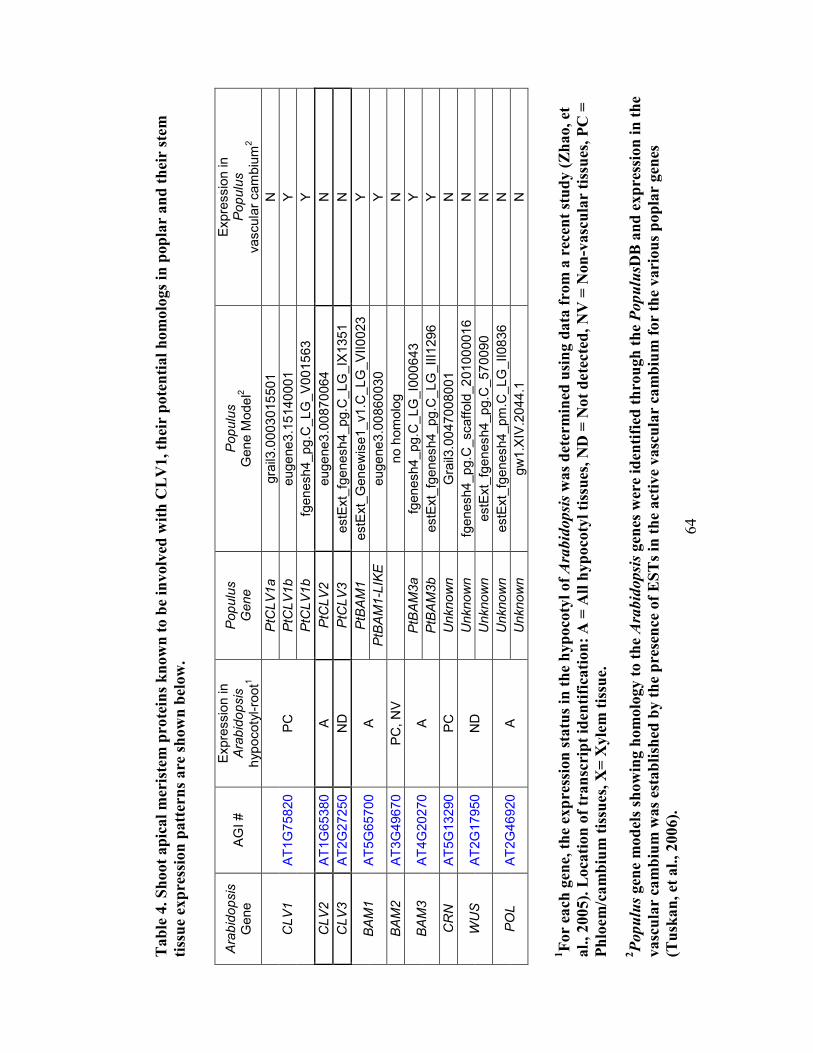
64
Tab
le 4
. S
hoot
ap
ical
mer
iste
m p
rote
ins
kn
ow
n t
o b
e in
volv
ed w
ith
CL
V1, th
eir
pote
nti
al
hom
olo
gs
in p
op
lar
an
d t
hei
r st
em
tiss
ue
exp
ress
ion
patt
ern
s are
sh
ow
n b
elow
.
Arabidopsis
Gene
AGI #
Expression in
Arabidopsis
hypocotyl-root1
Populus
Gene
Populus
Gene M
odel2
Expression in
Populus
vascular cambium
2
PtCLV1a
grail3.0003015501
N
PtCLV1b
eugene3.15140001
Y
CLV1
AT1G75820
PC
PtCLV1b
fgenesh4_pg.C
_LG_V001563
Y
CLV2
AT1G65380
A
PtCLV2
eugene3.00870064
N
CLV3
AT2G27250
ND
PtCLV3
estExt_fgenesh4_pg.C
_LG_IX1351
N
PtBAM1
estExt_Genewise1_v1.C
_LG_VII0023
Y
BAM1
AT5G65700
A
PtBAM1-LIKE
eugene3.00860030
Y
BAM2
AT3G49670
PC, NV
no homolog
N
PtBAM3a
fgenesh4_pg.C
_LG_I000643
Y
BAM3
AT4G20270
A
PtBAM3b
estExt_fgenesh4_pg.C
_LG_III1296
Y
CRN
AT5G13290
PC
Unknown
Grail3.0047008001
N
Unknown
fgenesh4_pg.C
_scaffold_201000016
N
WUS
AT2G17950
ND
Unknown
estExt_fgenesh4_pg.C
_570090
N
Unknown
estExt_fgenesh4_pm.C_LG_II0836
N
POL
AT2G46920
A
Unknown
gw1.XIV.2044.1
N
1F
or
each
gen
e, t
he
exp
ress
ion
sta
tus
in t
he
hyp
oco
tyl
of Ara
bidopsis
was
det
erm
ined
usi
ng d
ata
fro
m a
rec
ent
stu
dy (
Zh
ao, et
al.
, 2005).
Loca
tion
of
tran
scri
pt
iden
tifi
cati
on
: A
= A
ll h
yp
oco
tyl
tiss
ues
, �
D =
�ot
dete
cte
d,
�V
= �
on
-vasc
ula
r ti
ssu
es, P
C =
Ph
loem
/ca
mb
ium
tis
sues,
X=
Xyle
m t
issu
e.
2Populus
gen
e m
od
els
show
ing h
om
olo
gy t
o t
he A
rabidopsis
gen
es w
ere
iden
tifi
ed t
hro
ugh
th
e PopulusD
B a
nd
exp
ress
ion
in
th
e
vasc
ula
r ca
mb
ium
was
esta
bli
shed
by t
he
pre
sen
ce o
f E
ST
s in
th
e act
ive
vasc
ula
r ca
mb
ium
for
the
vari
ou
s p
op
lar
gen
es
(Tu
skan
, et
al.
, 2006).
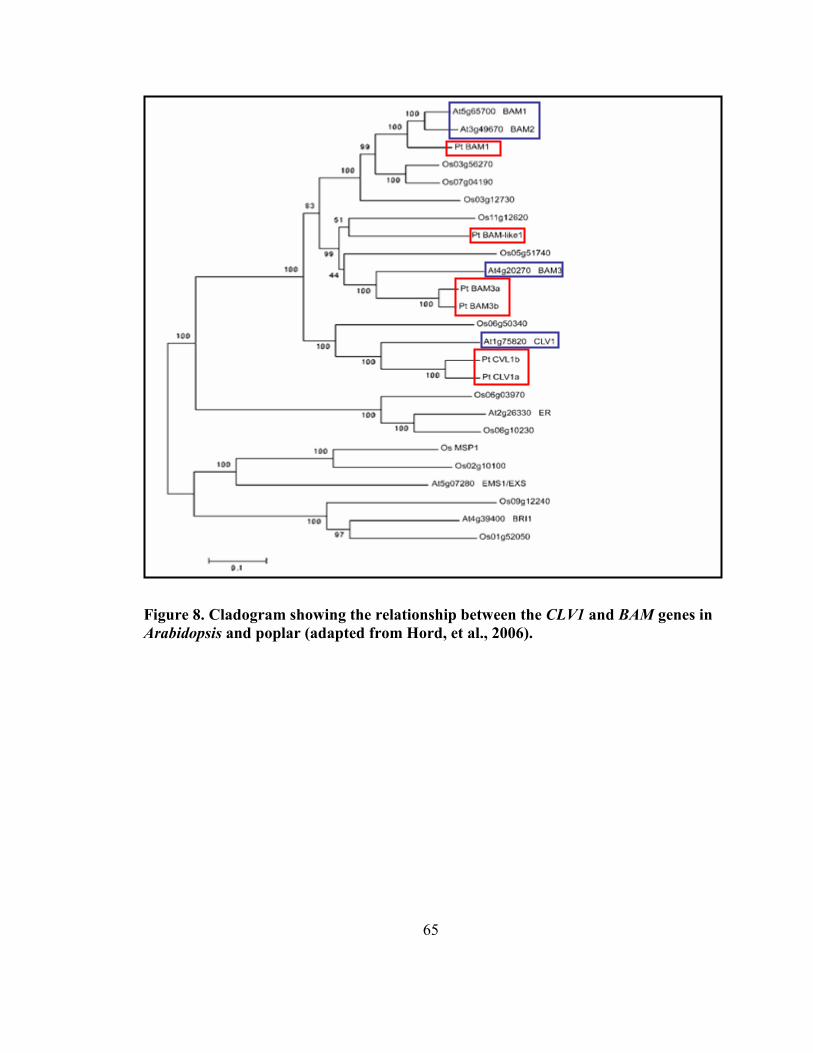
65
Figure 8. Cladogram showing the relationship between the CLV1 and BAM genes in
Arabidopsis and poplar (adapted from Hord, et al., 2006).

66
long, in regions which will ideally show a low level of non-specific complimentarity to
other transcripts. These are depicted in Appendix N.
3.5 Discussion
Our results have shown that antisense is a suitable technique to downregulate CLV1
expression. Eight out of 41 transformed Arabidopsis lines had extra petals in the flower,
which is typical of a strong CLV1-like phenotype. Northern blot analysis of the transgenic
lines showed that the transgene transcript was highly expressed in the flowers of
transformed lines while the levels of endogenous CLV1 transcripts were greatly reduced.
In northern blots of wild-type plants, the CLV1 transcript was found in the 2 month-old
hypocotyl. Since this is a tissue that is undergoing extensive secondary growth, we
hypothesized that CLV1 may be directly involved in the regulation of the vascular
cambium. We investigated the role of CLV1 in the development of the vascular
cambium through the downregulation of CLV1 expression by antisense. Consistent with
the clv1 mutant, the CLV1-antisense plants had significantly more secondary growth,
suggesting that CLV1 is a negative regulator of the vascular cambium. Based on this
work, it is reasonable to hypothesize that a similar strategy to downregulate CLV1
expression in the stem of a tree would also result in an increase in xylem formation. The
potential 11application of this strategy to trees is being explored.

67
3.5.1 Identifying clv1-like phenotypes in CLV1-antisense lines in Arabidopsis
The clv1 floral meristem can be twice as tall as the wildtype floral meristem at the earliest
stage of organ initiation (Clark et al., 1997). However, dissecting the meristem out of an
Arabidopsis plant and measuring its size by scanning electron microscopy is far too time
consuming and labor intensive to be used as a screen for analyzing transgenic plants.
Luckily, clv1 mutants have an additional phenotype in the development of the floral
meristem that is much easier to detect than its effect on the development of the SAM; the
formation of extra petals on the flower. This phenotype was chosen as a preliminary
indicator of a clv1-like phenotype. Based on this initial screen it is possible that some
transgenic lines with weaker floral phenotypes, but equally strong vascular cambium
phenotypes, were missed, but because of the large number of plants to be analyzed, it was
essential to restrict the phenotypic analysis to the most promising lines. We have shown
that the number of petals in the flower is a good indicator of a strong clv1-like phenotype
in the transgenic plants.
In our experiments, CLV1-antisense lines had phenotypes similar to the original mutant
thus indicating that the antisense strategy was effective in interfering with endogenous
CLV1 levels. All transgenic lines were advanced to the T3 generation and homozygous
lines were chosen for analysis. Only 20% of the antisense transformed plants displayed a
strong clv1 mutant phenotype. Because we were able to mimic strong clv1-like
phenotypes in these transgenic plants, it appears that the antisense transcript was

68
produced in the correct tissues and was able to successfully disrupt typical expression
patterns of CLV1.
The transgenic lines showed the broad range of flower phenotypes. For example,
Anti_L22 showed 3 petals, while Anti_L9, 16, 18, 23, and 32 showed extra petals.
Several other lines had no change in petal number, instead had flower shape phenotypes
such that the angle of the petals when compared to the rest of the flower were atypical.
This variation in phenotype from one line to the next is not unexpected, since transgene
expression is known to be affected by copy number and insertion location. Both of these
factors can alter the level of expression of the transgene and ultimately affect its impact
on endogenous CLV1 expression levels (Palauqui and Vaucheret, 1998).
3.5.2 The vascular cambium of Arabidopsis is negatively regulated by CLV1
Our results have shown that the growth of vascular cambium and the vascular meristem
are both negatively regulated by CLV1. In the primary stem, wildtype plants have 13
vascular bundles while the Anti_L32 has 20 bundles. This also resulted in an increase in
stem radius by 38 % when antisense lines were compared to wildtype plants. This is
consistent with previous findings that showed that bundle number is proportional to
meristem size. The increase of bundle number and stem radius could result from a direct
consequence of the increase in meristem size or indirect consequence of the increase in
organ number associated with larger meristem. Another distinct feature of antisense

69
transformed plants is that they have sturdy stems compared to wildtype plants, likely
resulting from the increased number of lignified vascular bundles.
The secondary growth in the hypocotyl of Arabidopsis made it possible to also
investigate the effect of CLV1 on the vascular cambium. Because of the similarity in
structure of the secondarily-thickened hypocotyl and the stem of trees, it is expected that
these results will be the most applicable to wood formation (Chaffey, et al., 2002). Our
results have shown that both xylem radius and phloem/cortex radius are found to be
greater than that of wildtype plants. Over all, the average xylem radius of 10 antisense
transformed lines is increased by 40 % compared to wildtype, and the average
phloem/cortex radius is increased by 39 %. Accordingly, the area of xylem and
phloem/cortex are increased by 86 % and 93 % respectively.
3.5.3 Arabidopsis CLV1 has multiple potential homologs in Populus
Our work has shown that CLV1 is negatively regulating the growth of the vascular
cambium in Arabidopsis. We have shown that CLV1 transcripts accumulate in 2-month
old hypocotyls and downregulating CLV1 results in an increase of both xylem and
phloem. Our results have also shown that the clv1-like phenotypes can be duplicated by
antisense expression, making this strategy possible for use in other plants, such as poplar
trees. It is reasonable to expect that CLV1 is also active in the vascular cambium of trees,
and in fact, EST sequencing has shown that one of the homologs of AtCLV1 is present in
the vascular cambium of poplar during both active and dormant periods (Tuskan, et al.,

70
2006). These same EST initiatives, combined with microarray analysis in both
Arabidopsis and poplar have indicated that other genes that are found in the SAM appear
to be present in the vascular cambium. These include, amongst others, the CLV1-like
gene, BAM1, and its closest variants AtBAM2 and AtBAM3. AtBAM1 and AtBAM3 appear
to have direct homologs in poplar, identified as PtBAM1 and PtBAM3. AtBAM1 also
shows very high homology to another gene in poplar, annotated as PtBAM1-LIKE (Hord,
et al., 2006). Unexpectedly, the predicted homolog of AtBAM2 was also PtBAM1. If
AtBAM2 had a direct homolog expressed in the vascular cambium it may have suggested
a completely conserved role of the BAMs in meristem development. However, it is
unknown what role BAM1Like will have in the stem. While exploring the possibility of
downregulating CLV1 in poplar, it was decided to also downregulate the PtCLV1-like
genes PtBAM1, PtBAM1-LIKE and the associated PtBAM3 to see if they would also show
phenotypes.
While full stable transformations of these genes could be carried out in tissue culture, a
recently reported technique was used instead to carry out the downregulation
experiments. Induced somatic sector analysis (ISSA) is a new transformation technique
which transforms a number of cells in a tree that is already soil based (Spokevicius, et al.,
2006). After transformation, these cells will proceed to divide and develop, carrying the
transgene.. The desired targets of the transformation are the cells of the vascular
cambium. If a cell in the vascular cambium is transformed, all of its progeny will also

71
bear the transgene. This is also the case in all other dividing cells, however, cambial cell
transformations are desirable due to the totipotential nature of the meristematic cells.
These cells will divide bidirectionally, producing transgenic phloem and xylem in radial
files. The transformed files can then be compared against the neighbouring wildtype files
to identify any visible changes in cell type or growth patterns. As both the antisense and
hairpin-RNAi constructs also harbour a GUS construct, transformed files can be
identified through GUS staining (Van Beveren, et al., 2006). This allows the
identification of which cell files are and aren't transformed before trying to screen for
phenotypes. Once the trees are transformed, a growth period of 3 to 4 months is required
to allow ample time for the healing of the inoculation wound, but mainly to allow ample
growth and development, making any phenotypes more easily detectable.
3.6 Conclusions
CLV1 is the first example of a negative regulator of the vascular meristem, and it appears
to affect xylem and phloem equally. There have been several positive regulators of
vascular meristems identified. The first regulator that was identified was WOL
(WOODE5 LEG), in which a recessive mutation resulted in reduced proliferation of
procambial cells, deformed xylem organization, and absence of phloem cells within root
vascular tissue (Scheres, et al., 1995). The second and third are two tissue specific genes
regulating the growth of phloem and xylem separately (van der Graaff, et al., 2002). VAS
(VASCULAR TISSUE SIZE) positively regulates growth of phloem cambial and

72
pericycle cells while LEP (LEAFY PETIOLE) can cause a specific increase in the
number of xylem cells. The third positive regulator ATHB-8 has been found to be
involved with the growth of vascular meristem (Baima, et al., 2001). Originally, ATHB-8
was found to participate in a positive feedback loop of auxin signalling transduction,
which in turn then positively regulated the activity of procambial and cambial cells
resulting in differentiation. The final regulator is IFL1 (I5TERFASCICULAR
FIBERLESS1), which is necessary for lateral meristem initiation and normal organ
development (Zhong, et al., 1997).
Future work will look specifically at the expression of CLV1 in poplar, at a variety of
growth regimes that are known to up and down regulate the activity of the vascular
cambium. Assuming that the vascular cambium is as complicated as the SAM, we can
expect that there are many other genes performing regulatory roles in the vascular
cambium. Some candidates might be the other genes that regulate the SAM, such as STM,
CLV2, CLV3, WUS, POL, but it is also possible that there are other proteins involved.
Through the study of these (and other) previously identified meristematic regulators, this
list will likely grow in the near future as the phenotypic markers and techniques are
available. As the influence of bioremediation and biofuel industries become more and
more pronounced, there will be more attention paid to the production of their raw
materials and more of the regulatory network will be made clear.

73
Chapter 4
General Discussion
This study describes recent work carried out to explore the genetic regulation of
secondary growth in poplar. Two genes, previously not associated with this process,
have been proposed to be involved with the regulation of secondary growth,.The
predicted roles of each of the genes are modeled in Appendix R These proposals was
arrived at through the use of both forward and reverse genetic strategies.
Firstly, a forward genetics approach was taken through the investigation of a mutant
identified in a recently described population of activation-tagged poplars (Harrison, et al.,
2007). The mutant showed aberrant secondary growth and was analyzed to determine
which gene had been misregulated to cause the resulting phenotype. This previously
undescribed gene was then partially characterized.
The second strategy that was used was a reverse genetics approach. Here, a previously
described regulator of the shoot apical and floral meristems was assayed to determine if it
also had a role in regulating the vascular cambium. It was determined that in Arabidopsis,
it did, as a significant increase in secondary growth was observed upon downregulating
the gene. To see if the effect was limited to the herbaceous dicot Arabidopsis, antisense
and hairpin-RNAi constructs were produced to downregulate the gene in poplar trees.

74
In this study the induced somatic sector analysis (ISSA) technique has been extensively
utilized to initiate an exploration of gene function. ISSA was chosen (amongst other
transformation strategies) for numerous reasons. Firstly, it is a system designed
specifically to look at transgenes which may alter the stem and its vasculature. Secondly,
it is a faster transformation strategy than traditional stable transformations. In the case of
some developmental mutants, the up to two year transformation period found with some
methods of transformation, can be a formidable barrier to academic study. Thirdly, it
transforms relatively few cells per transformation. While this could be considered a
drawback of the ISSA technique, the fact that the experiments only transform a few cells
or cell files actually creates a unique situation in developmental mutagenesis - an internal
control. Altering developmental regulators can result in transgenic plants which have
altered growth characteristics with pleotropic effects. These can include changes to the
rate or patterns of growth. In mutants such as these, it becomes very difficult to establish
comparable controls. This is typified in the corky mutant, in which it is hard to determine
which plants are directly comparable; whether it is those rooted at the same time, those
with a similar diameter shoot or those producing cork (i.e. similarity in age, development,
or maturity). The use of ISSA helps to solve this problem. As only a few files in each
stem segment will be transformed, the majority of the tree is still phenotypically
wildtype. Thus, the tree should still be functioning normally making a distinct phenotype
more easily observed. Finally, ISSA may also allow the specific modulation of one

75
meristem and not another. In full transformations using the cauliflower mosaic virus 35S
promoter, when a gene is misexpressed, it is done so in all of the plant tissues including
the multiple meristematic regions. This quickly creates a very complex situation when the
gene being misexpressed is a developmental or meristematic regulator. As the
transformed plants grow and develop, it becomes difficult to discern the effects on
development from one meristem versus another. ISSA circumvents this by only
transforming cells in the stem, and thus, for most transgenes, localizing the effects of the
transformation to the vascular and cork cambiums. This proves very useful in the clavata
project, as a role has already been identified for CLV1 in the shoot apical and floral
meristems (Clark, et al., 1997).
The work presented in this thesis has described a novel mutant in poplar, as well as
identified a previously undescribed gene which may be involved in the corky phenotype.
It has also shown that even in previously well described mutants, such as clv1-1 in
Arabidopsis, there are still new tangents to be studied. It is hoped that this will open the
doors to the review of other meristem regulating genes to see if they too have previously
unknown roles. This further exemplifies the importance of the study of plant anatomy and
physiology concurrently with genetics and biochemistry to gain a more holistic model of
plant growth and development.

76
Bibliography
Literature Cited
Aloni R (1987) Differentiation of vascular tissues. Annu Rev Plant Physiol 38: 179-204
Alonso JM, Stepanova A�, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK,
Zimmerman J, Barajas P, Cheuk R, Gadrinab C, Heller C, Jeske A, Koesema E,
Meyers CC, Parker H, Prednis L, Ansari Y, Choy �, Deen H, Geralt M, Hazari �,
Hom E, Karnes M, Mulholland C, �dubaku R, Schmidt I, Guzman P, Aguilar-
Henonin L, Schmid M, Weigel D, Carter DE, Marchand T, Risseeuw E, Brogden D,
Zeko A, Crosby WL, Berry CC, Ecker JR (2003) Genome-wide insertional
mutagenesis of Arabidopsis thaliana. Science 301: 653-657
Altschul S, Madden T, Schaffer A, Zhang J, Zhang Z, Miller W, Lipman D (1997)
Gapped BLAST and PSI-BLAST: a new generation of protein database search programs.
Nucl Acids Res 25: 3389-3402
Baima S, Possenti M, Matteucci A, Wisman E, Altamura MM, Ruberti I, Morelli G
(2001) The Arabidopsis ATHB-8 HD-Zip protein acts as a differentiation-promoting
transcription factor of the vascular meristems. Plant Physiol 126: 643-655
Balatinecz JJ, Kretschmann DE (2001) Properties and utilization of poplar wood. In DI
Dickmann, JG Isebrands, JE Eckenwalder, J Richardson, eds, Poplar culture in North
America. NRC Research Press, Ottawa, ON, pp 277-291
Bartlett HH (1927) A corky-barked mutation of Hevea brasiliensis. Botanical Gazette
84: 200-207
Beisson F, Li Y, Bonaventure G, Pollard M, Ohlrogge JB (2007) The acyltransferase
GPAT5 is required for the synthesis of suberin in seed coat and root of Arabidopsis. Plant
Cell 19: 351-368
Beukman EF, Gifford EM,Jr. (1969) Anatomic effects of Corky Bark Virus in Vitis.
Hilgardia 40: 73-103

77
Beveridge CA, Mathesius U, Rose RJ, Gresshoff PM (2007) Common regulatory
themes in meristem development and whole-plant homeostasis. Curr Opin Plant Biol 10:
44-51
Bonke M, Thitamadee S, Mähönen AP, Hauser M, Helariutta Y (2003) APL
regulates vascular tissue identity in Arabidopsis. Nature 426: 181-186
Brunner A, Yakovlev I, Strauss S (2004) Validating internal controls for quantitative
plant gene expression studies. BMC Plant Biol 4: 14
Busov VB, Meilan R, Pearce DW, Ma C, Rood SB, Strauss SH (2003) Activation
tagging of a dominant gibberellin catabolism gene (GA 2-oxidase) from poplar that
regulates tree stature. Plant Physiol 132: 1283-1291
Busse JS, Evert RF (1999) Vascular differentiation and transition in the seedling of
Arabidopsis thaliana (Brassicaceae). Int J Plant Sci 160: 241(1)
Cano-Delgado A, Yin Y, Yu C, Vafeados D, Mora-Garcia S, Cheng J, �am KH, Li
J, Chory J (2004) BRL1 and BRL3 are novel brassinosteroid receptors that function in
vascular differentiation in Arabidopsis. Development 131: 5341-5351
Carles CC, Choffnes-Inada D, Reville K, Lertpiriyapong K, Fletcher JC (2005)
ULTRAPETALA1 encodes a SAND domain putative transcriptional regulator that
controls shoot and floral meristem activity in Arabidopsis. Development 132: 897-911
Carlsbecker A, Helariutta Y (2005) Phloem and xylem specification: pieces of the
puzzle emerge. Curr Opin in Plant Biol, 8: 512-517
Chaffey � (2002) Why is there so little research into the cell biology of the secondary
vascular system of trees? New Phytol 153: 213-223
Chaffey � (1999) Wood formation in forest trees: from Arabidopsis to Zinnia . Trends
Plant Sci 4: 203-204
Chaffey �, Cholewa E, Regan S, Sundberg B (2002) Secondary xylem development in
Arabidopsis: a model for wood formation. Physiol Plantarum 114: 594-600
Cheng J, Lertpiriyapong K, Wang S, Sung ZR (2000) The role of the Arabidopsis
ELD1 gene in cell development and photomorphogenesis in darkness. Plant Physiol 123:
509-520

78
Choe S, �oguchi T, Fujioka S, Takatsuto S, Tissier CP, Gregory BD, Ross AS,
Tanaka A, Yoshida S, Tax FE, Feldmann KA (1999) The Arabidopsis dwf7/ste1
mutant is defective in the ∆7-Sterol C-5 desaturation step leading to brassinosteroid
biosynthesis. Plant Cell 11: 207-222
Chung CT, �iemela SL, Miller RH (1989) One-step preparation of competent
Escherichia coli: transformation and storage of bacterial cells in the same solution. Proc
Natl Acad Sci USA 86: 2172-2175
Church GM, Gilbert W (1984) Genomic sequencing. Proc Natl Acad Sci USA 81:
1991-1995
Clark SE (2001) Meristems: start your signalling. Current Opinion in Plant Biology, 4:
28-32
Clark SE, Running MP, Meyerowitz EM (1995) CLAVATA3 is a specific regulator of
shoot and floral meristem development affecting the same processes as CLAVATA1.
Development 121: 2057-2067
Clark SE, Running MP, Meyerowitz EM (1993) CLAVATA1, a regulator of meristem
and flower development in Arabidopsis. Development 119: 397-418
Clark SE, Williams RW, Meyerowitz EM (1997) The CLAVATA1 gene encodes a
putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell
89: 575-585
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated
transformation of Arabidopsis thaliana . Plant J 16:
Cock JM, McCormick S (2001) A large family of genes that share homology with
CLAVATA3. Plant Physiol 126: 939-942
Cooke JEK, Rood SB (2007) Trees of the people: the growing science of poplars in
Canada and worldwide. Can J Bot 85: 1103-1110
Cseke L, Cseke S, Podila G (2007) High efficiency poplar transformation. Plant Cell
Rep 26: 1529-1538

79
Cserzo M, Wallin E, Simon I, von Heijne G, Elofsson A (1997) Prediction of
transmembrane alpha-helices in prokaryotic membrane proteins: the dense alignment
surface method. Protein Eng 10: 673-676
DeYoung BJ, Bickle KL, Schrage KJ, Muskett P, Patel K, Clark SE (2006) The
CLAVATA1-related BAM1, BAM2 and BAM3 receptor kinase-like proteins are
required for meristem function in Arabidopsis. Plant J 45: 1-16
Dievart A, Dalal M, Tax FE, Lacey AD, Huttly A, Li J, Clark SE (2003) CLAVATA1
dominant-negative alleles reveal functional overlap between multiple receptor kinases
that regulate meristem and organ development. Plant Cell 15: 1198-1211
Dolan L, Roberts K (1995) Secondary thickening in roots of Arabidopsis thaliana:
anatomy and cell surface changes. New Phytol 131: 121-128
Druart �, Johansson A, Baba K, Schrader J, Sjödin A, Bhalerao RR, Resman L,
Trygg J, Moritz T, Bhalerao RP (2007) Environmental and hormonal regulation of the
activity–dormancy cycle in the cambial meristem involves stage-specific modulation of
transcriptional and metabolic networks. Plant J 50: 557-573
Durrant WE, Rowland O, Piedras P, Hammond-Kosack KE, Jones JDG (2000)
cDNA-AFLP reveals a striking overlap in race-specific resistance and wound response
gene expression profiles. Plant Cell 12: 963-977
Friedmann M, Ralph SG, Aeschliman D, Zhuang J, Ritland K, Ellis BE, Bohlmann
J, Douglas CJ (2007) Microarray gene expression profiling of developmental transitions
in Sitka spruce (Picea sitchensis) apical shoots. J Exp Bot 58: 593-614
Gasteiger E, Hoogland C, Gattiker A, Duvaud S, Wilkins MR, Appel RD, Bairoch A
(2005) Protein identification and analysis tools on the ExPASy server. In JM Walker, ed,
The Proteomics Protocols Handbook. Humana Press, Totowa, NJ, pp 571-607
Gendreau E, Traas J, Desnos T, Grandjean O, Caboche M, Hofte H (1997) Cellular
basis of hypocotyl growth in Arabidopsis thaliana. Plant Physiol 114: 295-305
Goff SA, Ricke D, Lan T, Presting G, Wang R, Dunn M, Glazebrook J, Sessions A,
Oeller P, Varma H, Hadley D, Hutchison D, Martin C, Katagiri F, Lange BM,
Moughamer T, Xia Y, Budworth P, Zhong J, Miguel T, Paszkowski U, Zhang S,
Colbert M, Sun W, Chen L, Cooper B, Park S, Wood TC, Mao L, Quail P, Wing R,

80
Dean R, Yu Y, Zharkikh A, Shen R, Sahasrabudhe S, Thomas A, Cannings R, Gutin
A, Pruss D, Reid J, Tavtigian S, Mitchell J, Eldredge G, Scholl T, Miller RM,
Bhatnagar S, Adey �, Rubano T, Tusneem �, Robinson R, Feldhaus J, Macalma T,
Oliphant A, Briggs S (2002) A draft sequence of the rice genome (Oryza sativa (L.) ssp.
japonica). Science 296: 92-100
Gregory EL (1888) Development of cork-wings on certain trees III. Botanical Gazette
13: 312-318
Guda C (2006) pTARGET: a web server for predicting protein subcellular localization.
Nucl Acids Res 34: W210-213
Harrison EJ, Bush M, Plett JM, McPhee DP, Vitez R, O’Malley B, Sharma V,
Bosnich W, Seguin A, MacKay J, Regan S (2007) Diverse developmental mutants
revealed in an activation-tagged population of poplar. Can J Bot 85: 1071-1081
Heazlewood JL, Verboom RE, Tonti-Filippini J, Small I, Millar AH (2007) SUBA:
the Arabidopsis subcellular database. Nucl Acids Res 35: D213-218
Hertzberg M, Aspeborg H, Schrader J, Andersson A, Erlandsson R, Blomqvist K,
Bhalerao R, Uhlén M, Teeri TT, Lundeberg J, Sundberg B, �ilsson P, Sandberg G
(2001) A transcriptional roadmap to wood formation. Proc Natl Acad Sci U S A 98:
14732-14737
Hord CLH, Chen C, DeYoung BJ, Clark SE, Ma H (2006) The BAM1/BAM2
receptor-like kinases are important regulators of Arabidopsis early anther development.
Plant Cell 18: 1667-1680
Jansson S, Douglas CJ (2007) Populus: a model system for plant biology. Annu Rev
Plant Biol 58: 435-458
Jenik PD, Irish VF (2000) Regulation of cell proliferation patterns by homeotic genes
during Arabidopsis floral development. Development 127: 1267-1276
Junghans U, Langenfeld-Heyser R, Polle A, Teichmann T (2004) Effect of auxin
transport inhibitors and ethylene on the wood anatomy of poplar. Plant Biology 6: 22-29
Käll L, Krogh A, Sonnhammer ELL (2004) A combined transmembrane topology and
signal peptide prediction method. J Mol Biol, 338: 1027-1036

81
Kim J, Jung J, Reyes JL, Kim Y, Kim S, Chung K, Kim JA, Lee M, Lee Y, Kim V�,
Chua �, Park C (2005) microRNA-directed cleavage of ATHB15 mRNA regulates
vascular development in Arabidopsis inflorescence stems. Plant J 42: 84-94
Kohler A, Delaruelle C, Martin D, Encelot �, Martin F (2003) The poplar root
transcriptome: analysis of 7000 expressed sequence tags. FEBS Lett 542: 37-41
Larkin MA, Blackshields G, Brown �P, Chenna R, McGettigan PA, McWilliam H,
Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG
(2007) Clustal W and Clustal X version 2.0. Bioinformatics 23: 2947-2948
Laux T, Mayer KF, Berger J, Jurgens G (1996) The WUSCHEL gene is required for
shoot and floral meristem integrity in Arabidopsis. Development 122: 87-96
Lee H, Lee J, �oh E, Bae E, Choi Y, Han M (2005) Generation and analysis of
expressed sequence tags from poplar (Populus alba × P. tremula var. glandulosa)
suspension cells. Plant Science 169: 1118-1124
Lenhard M, Jurgens G, Laux T (2002) The WUSCHEL and SHOOTMERISTEMLESS
genes fulfil complementary roles in Arabidopsis shoot meristem regulation. Development
129: 3195-3206
Lev-Yadun S (1995) Short secondary vessel members in branching regions in roots of
Arabidopsis thaliana. Aust J Bot 43: 435-438
Lynn K, Fernandez A, Aida M, Sedbrook J, Tasaka M, Masson P, Barton MK
(1999) The PI5HEAD/ZWILLE gene acts pleiotropically in Arabidopsis development and
has overlapping functions with the ARGO5AUTE1 gene. Development 126: 469-481
Marchler-Bauer A, Anderson JB, Derbyshire MK, DeWeese-Scott C, Gonzales �R,
Gwadz M, Hao L, He S, Hurwitz DI, Jackson JD, Ke Z, Krylov D, Lanczycki CJ,
Liebert CA, Liu C, Lu F, Lu S, Marchler GH, Mullokandov M, Song JS, Thanki �,
Yamashita RA, Yin JJ, Zhang D, Bryant SH (2007) CDD: a conserved domain
database for interactive domain family analysis. Nucl Acids Res 35: D237-240
Mellerowicz EJ, Sundberg B (2008) Wood cell walls: biosynthesis, developmental
dynamics and their implications for wood properties. Curr Opin Plant Biol 11: 293-300

82
Meyerowitz EM (1997) Genetic control of cell division patterns in developing plants.
Cell 88: 299-308
Moussian B, Schoof H, Haecker A, Jürgens G, Laux T (1998) Role of the ZWILLE
gene in the regulation of central shoot meristem cell fate during Arabidopsis
embryogenesis. The EMBO Journal 17: 1799-1809
Muller R, Bleckmann A, Simon R (2008) The receptor kinase CORYNE of Arabidopsis
transmits the stem cell-limiting signal CLAVATA3 independently of CLAVATA1. Plant
Cell 20: 934-946
Murray MG, Thompson WF (1980) Rapid isolation of high molecular weight plant
DNA. Nucl Acids Res 8: 4321-4325
�agata �, Asami T, Yoshida S (2001) Brassinazole, an inhibitor of brassinosteroid
biosynthesis, inhibits development of secondary xylem in cress plants (Lepidium
sativum). Plant Cell Physiol 42: 1006-1011
�anjo T, Sakurai T, Totoki Y, Toyoda A, �ishiguchi M, Kado T, Igasaki T,
Futamura �, Seki M, Sakaki Y, Shinozaki K, Shinohara K (2007) Functional
annotation of 19,841 Populus nigra full-length enriched cDNA clones. BMC Genomics
8: 448
O'Connor TR, Dyreson C, Wyrick JJ (2005) Athena: a resource for rapid visualization
and systematic analysis of Arabidopsis promoter sequences. Bioinformatics 21: 4411-
4413
Ogata H, Goto S, Sato K, Fujibuchi W, Bono H, Kanehisa M (1999) KEGG: Kyoto
encyclopedia of genes and genomes. Nucl Acids Res 27: 29-34
Ogawa M, Shinohara H, Sakagami Y, Matsubayashi Y (2008) Arabidopsis CLV3
peptide directly binds CLV1 ectodomain. Science 319: 294
Oh S, Park S, Han K (2003) Transcriptional regulation of secondary growth in
Arabidopsis thaliana. J Exp Bot 54: 2709-2722
Opsahl-Ferstad H, Deunff EL, Dumas C, Rogowsky PM (1997) ZmEsr, a novel
endosperm-specific gene expressed in a restricted region around the maize embryo. Plant
J 12: 235-246

83
Palauqui J, Vaucheret H (1998) Transgenes are dispensable for the RNA degradation
step of cosuppression. Proc Natl Acad Sci U S A 95: 9675-9680
Pan X, Gilkes �, Kadla J, Pye K, Saka S, Gregg D, Ehara K, Xie D, Lam D, Saddler
J (2006) Bioconversion of hybrid poplar to ethanol and co-products using an organosolv
fractionation process: Optimization of process yields. Biotechnol Bioeng 94: 851-861
Parker G, Schofield R, Sundberg B, Turner S (2003) Isolation of COV1, a gene
involved in the regulation of vascular patterning in the stem of Arabidopsis. Development
130: 2139-2148
Plomion C, Lalanne C, Claverol S, Meddour H, Kohler A, Bogeat-Triboulot M,
Barre A, Provost GL, Dumazet H, Jacob D, Bastien C, Dreyer E, Daruvar Ad,
Guehl J, Schmitter J, Martin F, Bonneu M (2006) Mapping the proteome of poplar
and application to the discovery of drought-stress responsive proteins. Proteomics 6:
6509-6527
Pogany J, Simon E, Katzman R, De Guzman B, Yu L, Trotochaud A, Clark S (1998)
Identifying novel regulators of shoot meristem development. J Plant Res 111: 307-313
Ralph S, Chun HJ, Cooper D, Kirkpatrick R, Kolosova �, Gunter L, Tuskan G,
Douglas C, Holt R, Jones S, Marra M, Bohlmann J (2008) Analysis of 4,664 high-
quality sequence-finished poplar full-length cDNA clones and their utility for the
discovery of genes responding to insect feeding. BMC Genomics 9: 57
Regan S, Bourquin V, Tuominen H, Sundberg B (1999) Accurate and high resolution
in situ hybridization analysis of gene expression in secondary stem tissues. Plant J 19:
363-369
Rogers LA, Campbell MM (2004) The genetic control of lignin deposition during plant
growth and development. New Phytol 164: 17-30
Rowland O, Ludwig AA, Merrick CJ, Baillieul F, Tracy FE, Durrant WE, Fritz-
Laylin L, �ekrasov V, Sjolander K, Yoshioka H, Jones JDG (2005) Functional
analysis of Avr9/Cf-9 rapidly elicited genes identifies a protein kinase, ACIK1, that is
essential for full Cf-9-dependent disease resistance in tomato. Plant Cell 17: 295-310

84
Satoh �, Itoh J, �agato Y (2003) The SHOOTLESS2 and SHOOTLESS1 genes are
involved in both initiation and maintenance of the shoot apical meristem through
regulating the number of indeterminate cells. Genetics 164: 335-346
Saxena KB, Dundas IS, Faris DG, Gupta SC (1988) Genetics and histology of a corky-
stem mutant in pigeonpea. J Hered 79: 221-222
Scarpella E, Meijer AH (2004) Pattern formation in the vascular system of monocot and
dicot plant species. New Phytol 164: 209-242
Scheres B, Di Laurenzio L, Willemsen V, Hauser MT, Janmaat K, Weisbeek P,
Benfey P� (1995) Mutations affecting the radial organization of the Arabidopsis root
display specific defects throughout the embryonic axis. Development 121: 53-62
Schneider H (1973) Cytological and histological aberrations in woody plants following
infection with viruses, mycoplasmas, rickettsias, and flagellates. Annu Rev Phytopathol
11: 119-146
Schrader J, Moyle R, Bhalerao R, Hertzberg M, Lundeberg J, �ilsson P, Bhalerao
RP (2004a) Cambial meristem dormancy in trees involves extensive remodelling of the
transcriptome. Plant J 40: 173-187
Schrader J, �ilsson J, Mellerowicz E, Berglund A, �ilsson P, Hertzberg M,
Sandberg G (2004b) A high-resolution transcript profile across the wood-forming
meristem of poplar identifies potential regulators of cambial stem cell identity. Plant Cell
16: 2278-2292
Simossis VA, Kleinjung J, Heringa J (2005) Homology-extended sequence alignment.
Nucl Acids Res 33: 816-824
Smyth DR, Bowman JL, Meyerowitz EM (1990) Early flower development in
Arabidopsis. Plant Cell 2: 755-767
Soler M, Serra O, Molinas M, Huguet G, Fluch S, Figueras M (2007) A genomic
approach to suberin biosynthesis and cork differentiation. Plant Physiol 144: 419-431
Spokevicius AV, Van Beveren KS, Bossinger G (2006) Agrobacterium-mediated
transformation of dormant lateral buds in poplar trees reveals developmental patterns in
secondary stem tissues. Funct Plant Biol 33: 133-139

85
Spyropoulos I, Liakopoulos TD, Bagos PG, Hamodrakas SJ (2004) TMRPres2D:
high quality visual representation of transmembrane protein models. Bioinformatics 20:
3258-3260
Stephens SG, Phillips LL (1972) The history and geographical distribution of a
polymorphic system in new world cottons. Biotropica 4: 49-60
Sterky F, Bhalerao R, Unneberg P, Segerman B, �ilsson P, Brunner A, Charbonnel-
Campaa L, Lindvall J, Tandre K, Strauss S, Sundberg B, Gustafsson P, Uhlen M,
Bhalerao R, �ilsson O, Sandberg G, Karlsson J, Lundeberg J, Jansson S (2004) A
Populus EST resource for plant functional genomics. Proc Natl Acad Sci U S A 101:
13951-13956
Sterky F, Regan S, Karlsson J, Hertzberg M, Rohde A, Holmberg A, Amini B,
Bhalerao R, Larsson M, Villarroel R, Van Montagu M, Sandberg G, Olsson O,
Teeri TT, Boerjan W, Gustafsson P, Uhlén M, Sundberg B, Lundeberg J (1998)
Gene discovery in the wood-forming tissues of poplar: Analysis of 5,692 expressed
sequence tags. Proc Natl Acad Sci U S A 95: 13330-13335
Szekeres M, �émeth K, Koncz-Kálmán Z, Mathur J, Kauschmann A, Altmann T,
Rédei GP, �agy F, Schell J, Koncz C (1996) Brassinosteroids rescue the deficiency of
CYP90, a cytochrome P450, controlling cell elongation and de-etiolation in Arabidopsis.
Cell 85: 171-182
Taylor �G (2008) Cellulose biosynthesis and deposition in higher plants. New Phytol
178: 239-252
The Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the
flowering plant Arabidopsis thaliana. Nature 408: 796-815
Tuskan GA, DiFazio S, Jansson S, Bohlmann J, Grigoriev I, Hellsten U, Putnam �,
Ralph S, Rombauts S, Salamov A, Schein J, Sterck L, Aerts A, Bhalerao RR,
Bhalerao RP, Blaudez D, Boerjan W, Brun A, Brunner A, Busov V, Campbell M,
Carlson J, Chalot M, Chapman J, Chen G-, Cooper D, Coutinho PM, Couturier J,
Covert S, Cronk Q, Cunningham R, Davis J, Degroeve S, Dejardin A, dePamphilis
C, Detter J, Dirks B, Dubchak I, Duplessis S, Ehlting J, Ellis B, Gendler K,
Goodstein D, Gribskov M, Grimwood J, Groover A, Gunter L, Hamberger B,
Heinze B, Helariutta Y, Henrissat B, Holligan D, Holt R, Huang W, Islam-Faridi �,

86
Jones S, Jones-Rhoades M, Jorgensen R, Joshi C, Kangasjarvi J, Karlsson J,
Kelleher C, Kirkpatrick R, Kirst M, Kohler A, Kalluri U, Larimer F, Leebens-Mack
J, Leple J-, Locascio P, Lou Y, Lucas S, Martin F, Montanini B, �apoli C, �elson
DR, �elson C, �ieminen K, �ilsson O, Pereda V, Peter G, Philippe R, Pilate G,
Poliakov A, Razumovskaya J, Richardson P, Rinaldi C, Ritland K, Rouze P, Ryaboy
D, Schmutz J, Schrader J, Segerman B, Shin H, Siddiqui A, Sterky F, Terry A, Tsai
C-, Uberbacher E, Unneberg P, Vahala J, Wall K, Wessler S, Yang G, Yin T,
Douglas C, Marra M, Sandberg G, Van de Peer Y, Rokhsar D (2006) The genome of
black cottonwood, Populus trichocarpa (Torr. & Gray). Science 313: 1596-1604
Tzfira T, Zuker A, Altman A (1998) Forest-tree biotechnology: genetic transformation
and its application to future forests. Trends Biotechnol 16: 439-446
Untergasser A, �ijveen H, Rao X, Bisseling T, Geurts R, Leunissen JAM (2007)
Primer3Plus, an enhanced web interface to Primer3. Nucl Acids Res 35: W71-74
Van Beveren KS, Spokevicius AV, Tibbits J, Wang Q, Bossinger G (2006)
Transformation of cambial tissue in vivo provides an efficient means for induced somatic
sector analysis and gene testing in stems of woody plant species. Funct Plant Biol 33:
629-638
van der Graaff E, Hooykaas PJJ, Keller B (2002) Activation tagging of the two closely
linked genes LEP and VAS independently affects vascular cell number. Plant J 32: 819-
830
van Raemdonck D, Pesquet E, Cloquet S, Beeckman H, Boerjan W, Goffner D, El
Jaziri M, Baucher M (2005) Molecular changes associated with the setting up of
secondary growth in aspen. J Exp Bot 56: 2211-2227
Wall PK, Leebens-Mack J, Muller KF, Field D, Altman �S, dePamphilis CW (2008)
PlantTribes: a gene and gene family resource for comparative genomics in plants. Nucl
Acids Res 36: D970-976
Weigel D, Ahn JH, Blazquez MA, Borevitz JO, Christensen SK, Fankhauser C,
Ferrandiz C, Kardailsky I, Malancharuvil EJ, �eff MM, �guyen JT, Sato S, Wang
Z, Xia Y, Dixon RA, Harrison MJ, Lamb CJ, Yanofsky MF, Chory J (2000)
Activation Tagging in Arabidopsis. Plant Physiol 122: 1003-1014

87
Winter D, Vinegar B, �ahal H, Ammar R, Wilson GV, Provart �J (2007) An
“Electronic fluorescent pictograph” browser for exploring and analyzing large-scale
biological data sets. PLoS ONE 2: e718
Ye Z (2002) Vascular tissue differentiation and pattern formation in plants. Annu Rev
Plant Physiol 53: 183-202
Yemshanov D, McKenney D (2008) Fast-growing poplar plantations as a bioenergy
supply source for Canada. Biomass and Bioenergy 32: 185-197
Yephremov A, Schreiber L (2005) The dark side of the cell wall: Molecular genetics of
plant cuticle. Plant Biosystems 139: 74-79
Yu LP, Simon EJ, Trotochaud AE, Clark SE (2000) POLTERGEIST functions to
regulate meristem development downstream of the CLAVATA loci. Development 127:
1661-1670
Zdobnov EM, Apweiler R (2001) InterProScan - an integration platform for the
signature-recognition methods in InterPro. Bioinformatics 17: 847-848
Zhao C, Craig JC, Petzold HE, Dickerman AW, Beers EP (2005) The xylem and
phloem transcriptomes from secondary tissues of the Arabidopsis root-hypocotyl. Plant
Physiol 138: 803-818
Zhong R, Taylor JJ, Ye ZH (1997) Disruption of interfascicular fiber differentiation in
an Arabidopsis mutant. Plant Cell 9: 2159-2170

88
Appendix A
Primers used for activation tag localization, RACE and SALK line
screening in corky project.
Role Primer �ame Primer Sequence (5' - 3')
AP1 GTAATACGACTCACTATAGGGC
AP2 ACTATAGGGCACGCGTGGT
LBGSP1 CATCCGACATGGCGCGACCGACCTCAACG
LBGSP2 GCTCTGGGGCGTGCAAGGTTCCGGAACG
RBGSP1 GGACCCCCACCCACGAGGAACATCGTGG
Genome
Walking
RBGSP2 GATTGATGTGATATCTAGATCCGAAACTATCAGTG
RAP1 GTAATACGACTCACTATAGGGC
RAP2 ACTATAGGGCACGCGTGGT RACE
GRAILFW-L CAGGTGGCCGGAATTCGGATTCTCTATG
LBa1-SALK TGGTTCACGTAGTGGGCCATCG
SALK93568-LP TATGTTTGTCCGTTTGTCGTG
SALK93568-RP ACGATGGTAGATCCATCAACG
SALK27013-LP TCGTGTCCACACCATTATCAG
Screening for
homozygous
Arabidopsis
SALK lines
SALK27013-RP ACCACATCAATCCTCTCATGC
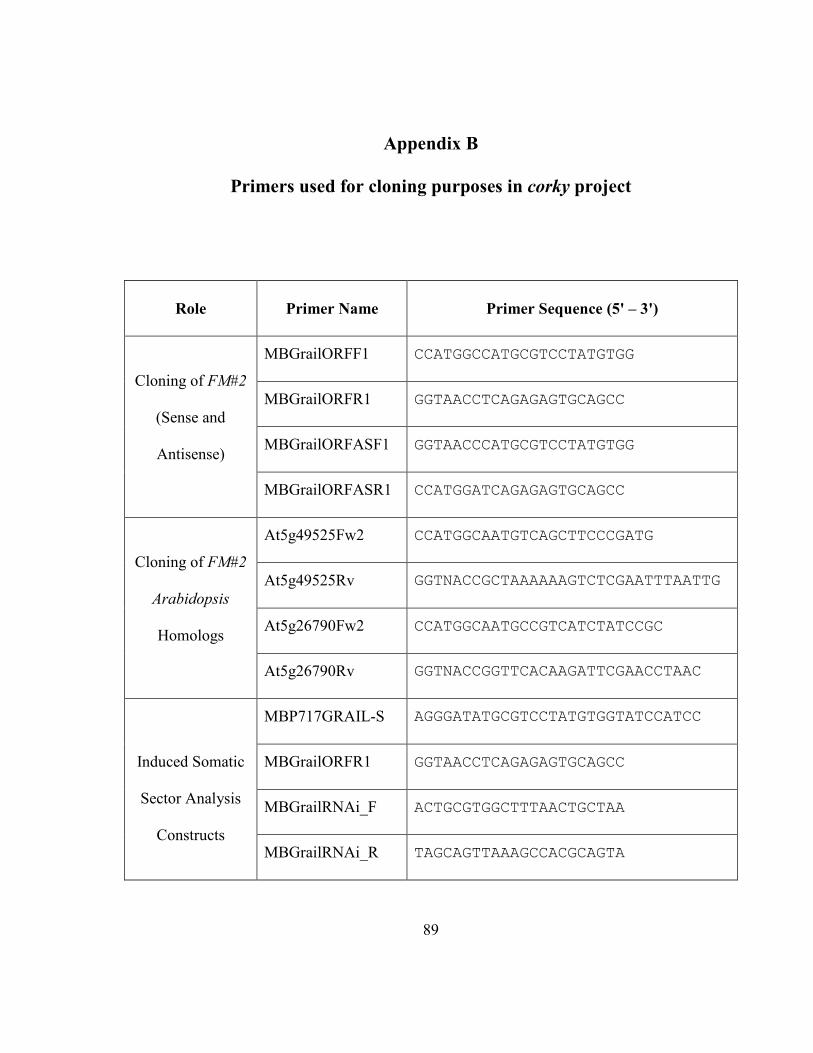
89
Appendix B
Primers used for cloning purposes in corky project
Role
Primer �ame Primer Sequence (5' – 3')
MBGrailORFF1 CCATGGCCATGCGTCCTATGTGG
MBGrailORFR1 GGTAACCTCAGAGAGTGCAGCC
MBGrailORFASF1 GGTAACCCATGCGTCCTATGTGG
Cloning of FM#2
(Sense and
Antisense)
MBGrailORFASR1 CCATGGATCAGAGAGTGCAGCC
At5g49525Fw2 CCATGGCAATGTCAGCTTCCCGATG
At5g49525Rv GGTNACCGCTAAAAAAGTCTCGAATTTAATTG
At5g26790Fw2 CCATGGCAATGCCGTCATCTATCCGC
Cloning of FM#2
Arabidopsis
Homologs
At5g26790Rv GGTNACCGGTTCACAAGATTCGAACCTAAC
MBP717GRAIL-S AGGGATATGCGTCCTATGTGGTATCCATCC
MBGrailORFR1 GGTAACCTCAGAGAGTGCAGCC
MBGrailRNAi_F ACTGCGTGGCTTTAACTGCTAA
Induced Somatic
Sector Analysis
Constructs MBGrailRNAi_R TAGCAGTTAAAGCCACGCAGTA
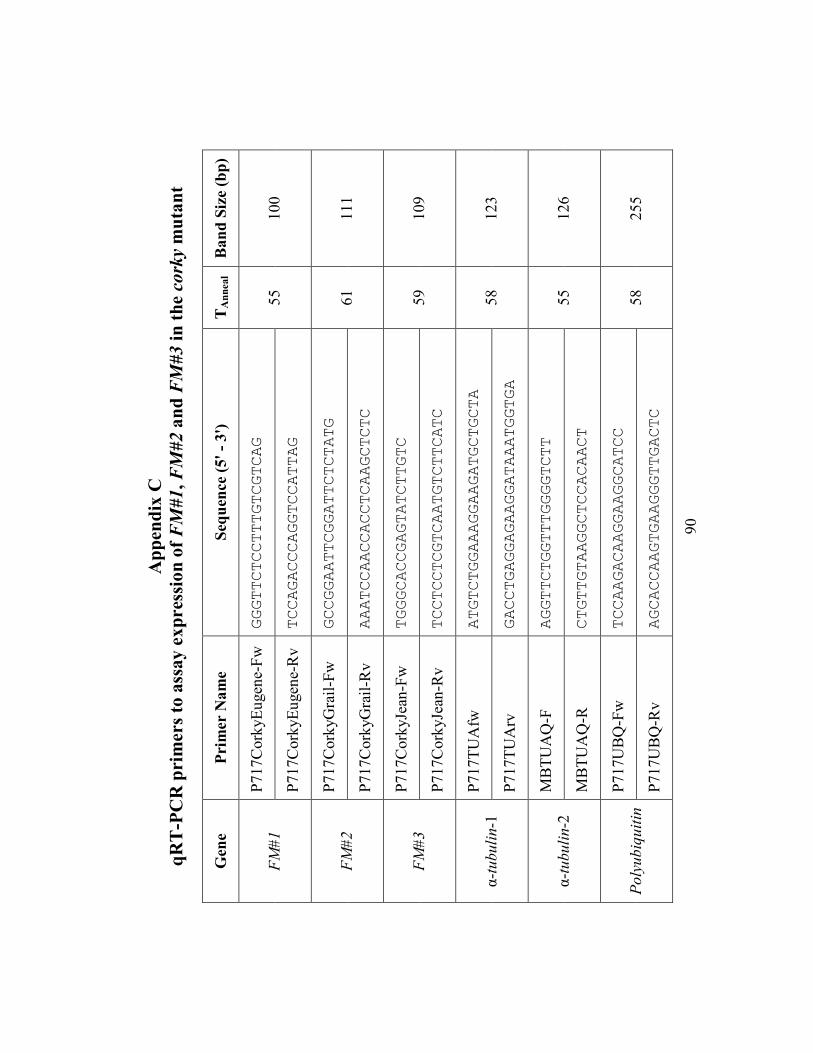
90
Ap
pen
dix
C
qR
T-P
CR
pri
mer
s to
ass
ay
ex
pre
ssio
n o
f FM#1
, FM#2
an
d F
M#3
in
th
e co
rky
mu
tan
t
Gen
e P
rim
er �
am
e S
equ
ence
(5'
- 3')
T
An
nea
l B
an
d S
ize
(bp
)
P717C
ork
yE
ugen
e-F
w GGGTTCTCCTTTGTCGTCAG
FM
#1
P717C
ork
yE
ugen
e-R
v TCCAGACCCAGGTCCATTAG
55
100
P717C
ork
yG
rail
-Fw
GCCGGAATTCGGATTCTCTATG
FM
#2
P717C
ork
yG
rail
-Rv
AAATCCAACCACCTCAAGCTCTC
61
111
P717C
ork
yJe
an-F
w
TGGGCACCGAGTATCTTGTC
FM
#3
P717C
ork
yJe
an-R
v
TCCTCCTCGTCAATGTCTTCATC
59
109
P717T
UA
fw
ATGTCTGGAAAGGAAGATGCTGCTA
α-t
ubuli
n-1
P717T
UA
rv
GACCTGAGGAGAAGGATAAATGGTGA
58
123
MB
TU
AQ
-F
AGGTTCTGGTTTGGGGTCTT
α-t
ubuli
n-2
MB
TU
AQ
-R
CTGTTGTAAGGCTCCACAACT
55
126
P717U
BQ
-Fw
TCCAAGACAAGGAAGGCATCC
Poly
ubiq
uit
in
P717U
BQ
-Rv
AGCACCAAGTGAAGGGTTGACTC
58
255
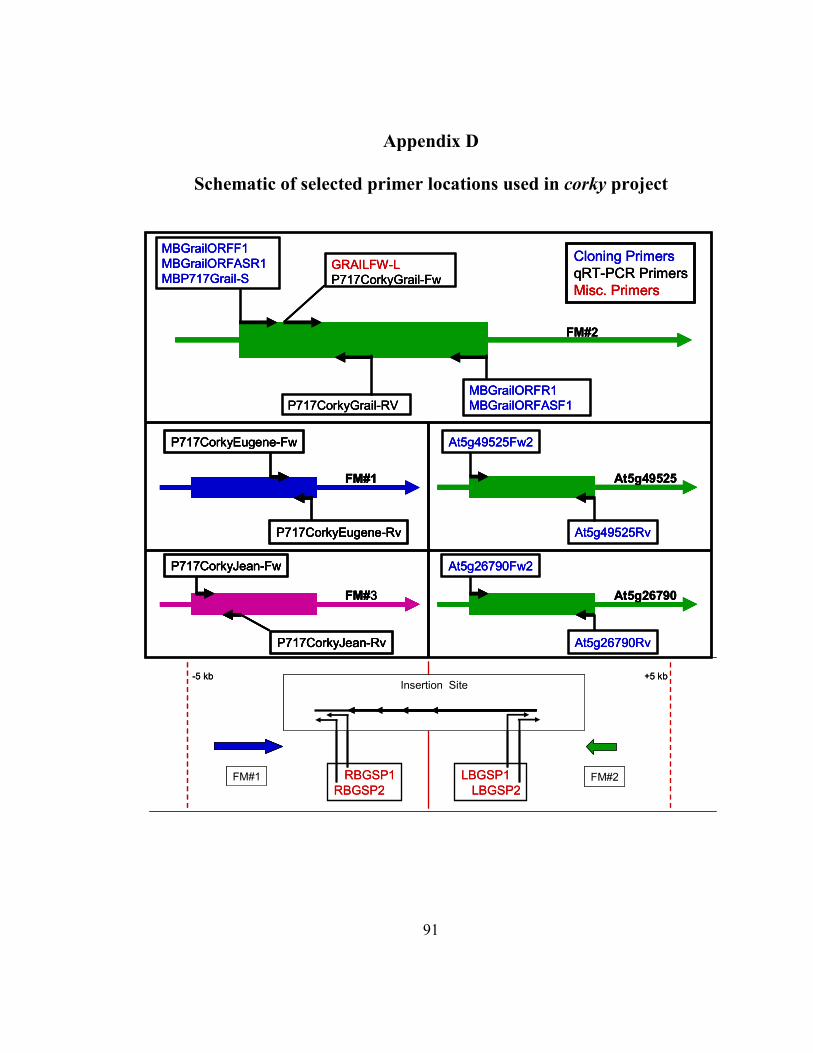
91
Appendix D
Schematic of selected primer locations used in corky project
P717CorkyEugene-Fw
P717CorkyEugene-Rv
FM#1
P717CorkyJean-Fw
P717CorkyJean-Rv
FM#3
MBGrailORFF1
MBGrailORFASR1
MBP717Grail-S
MBGrailORFR1
MBGrailORFASF1
GRAILFW-L
P717CorkyGrail-Fw
P717CorkyGrail-RV
FM#2
At5g49525Fw2
At5g49525Rv
At5g49525
At5g26790Fw2
At5g26790Rv
At5g26790
Cloning Primers
qRT-PCR Primers
Misc. Primers
-5 kb +5 kb
FM#1 FM#2
Insertion Site
LBGSP1
LBGSP2
RBGSP1
RBGSP2
P717CorkyEugene-Fw
P717CorkyEugene-Rv
FM#1
P717CorkyJean-Fw
P717CorkyJean-Rv
FM#3
MBGrailORFF1
MBGrailORFASR1
MBP717Grail-S
MBGrailORFR1
MBGrailORFASF1
GRAILFW-L
P717CorkyGrail-Fw
P717CorkyGrail-RV
FM#2
At5g49525Fw2
At5g49525Rv
At5g49525
At5g26790Fw2
At5g26790Rv
At5g26790
Cloning Primers
qRT-PCR Primers
Misc. Primers
P717CorkyEugene-Fw
P717CorkyEugene-Rv
FM#1
P717CorkyJean-Fw
P717CorkyJean-Rv
FM#3
P717CorkyEugene-Fw
P717CorkyEugene-Rv
FM#1
P717CorkyJean-Fw
P717CorkyJean-Rv
FM#3
MBGrailORFF1
MBGrailORFASR1
MBP717Grail-S
MBGrailORFR1
MBGrailORFASF1
GRAILFW-L
P717CorkyGrail-Fw
P717CorkyGrail-RV
FM#2
At5g49525Fw2
At5g49525Rv
At5g49525
At5g26790Fw2
At5g26790Rv
At5g26790
Cloning Primers
qRT-PCR Primers
Misc. Primers
-5 kb +5 kb
FM#1FM#1 FM#2
Insertion Site
LBGSP1
LBGSP2
RBGSP1
RBGSP2
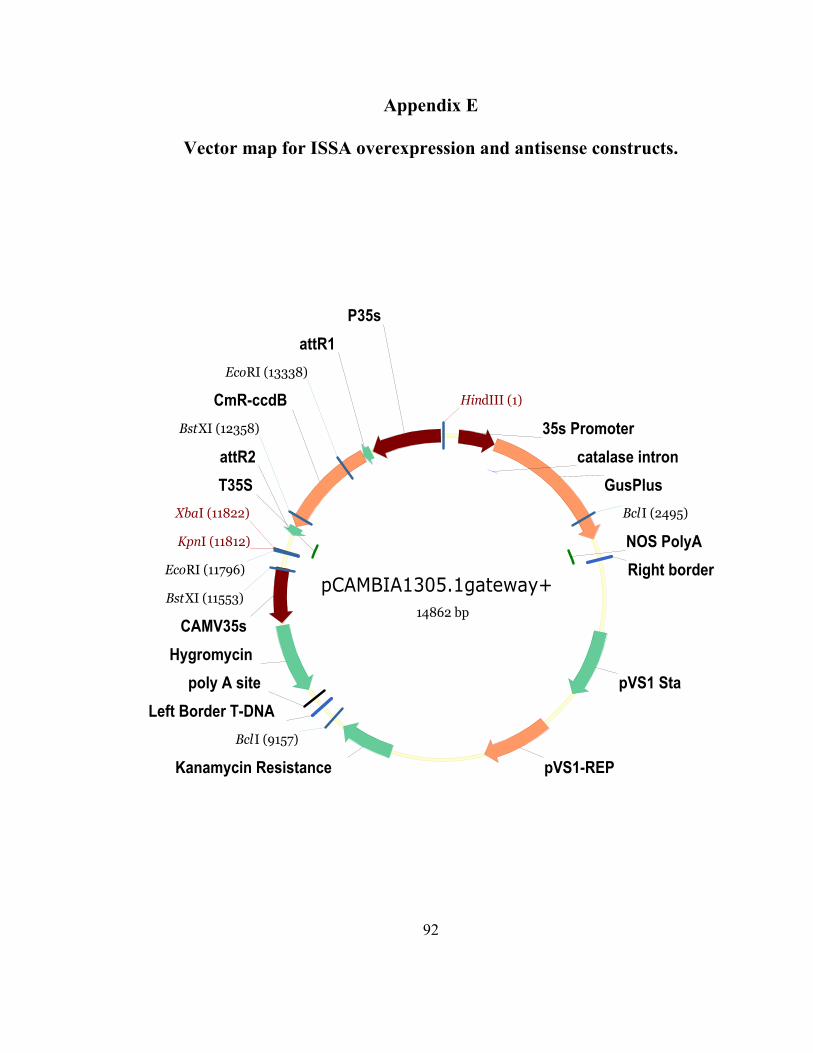
92
Appendix E
Vector map for ISSA overexpression and antisense constructs.
pCAMBIA1305.1gateway+
14862 bp
GusPlus
catalase intron
Kanamycin Resistance
Left Border T-DNA
Hygromycin
Right border T-DNA
pVS1 Sta
T35S
NOS PolyA
poly A site
CAMV35s
P35s
35s Promoter
pVS1-REP
CmR-ccdB
attR2
attR1
HindIII (1)
KpnI (11812)
XbaI (11822)
BstXI (11553)
BstXI (12358)
EcoRI (11796)
EcoRI (13338)
BclI (2495)
BclI (9157)
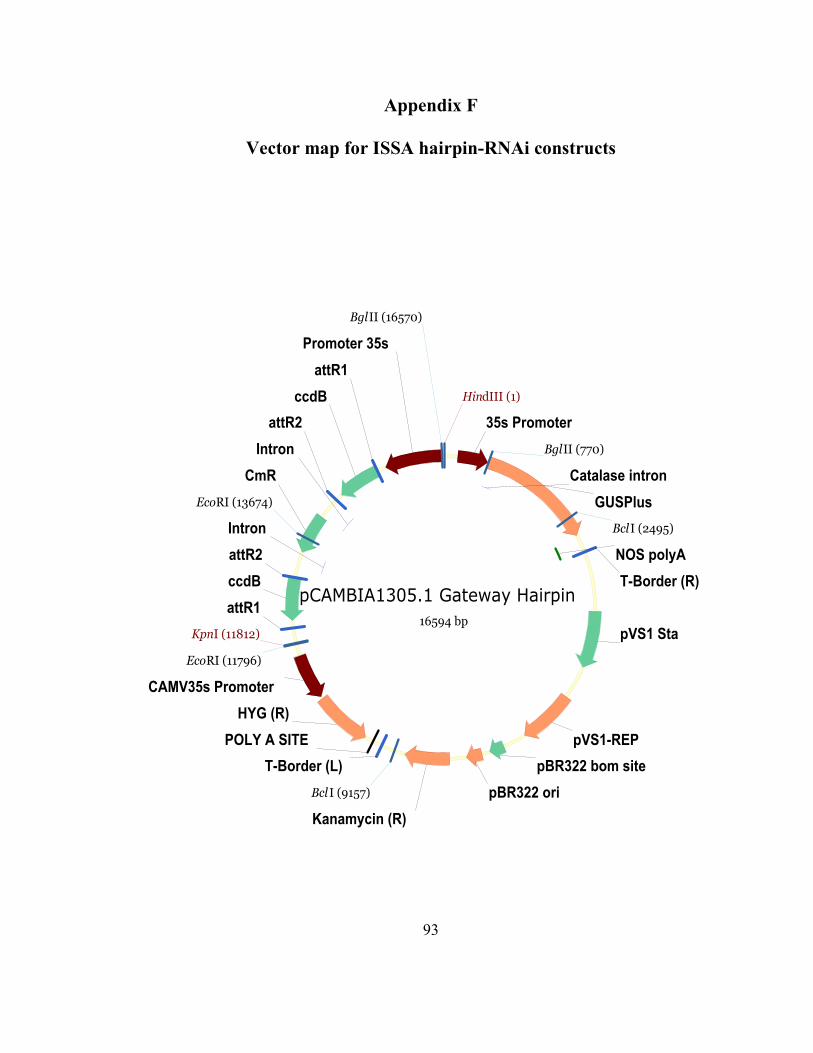
93
Appendix F
Vector map for ISSA hairpin-R�Ai constructs
pCAMBIA1305.1 Gateway Hairpin
16594 bp
GUSPlus
Kanamycin (R)
HYG (R)
Catalase intron
Intron
Intron
pVS1 Sta
T-Border (R)
pBR322 bom siteT-Border (L)
ccdB
ccdB
attR1
attR2
attR2
CmR
NOS polyA
POLY A SITE
35s Promoter
CAMV35s Promoter
Promoter 35s
pVS1-REP
pBR322 ori
attR1
HindIII (1)
KpnI (11812)
EcoRI (11796)
EcoRI (13674)
BclI (2495)
Bcl I (9157)
BglII (770)
BglII (16570)
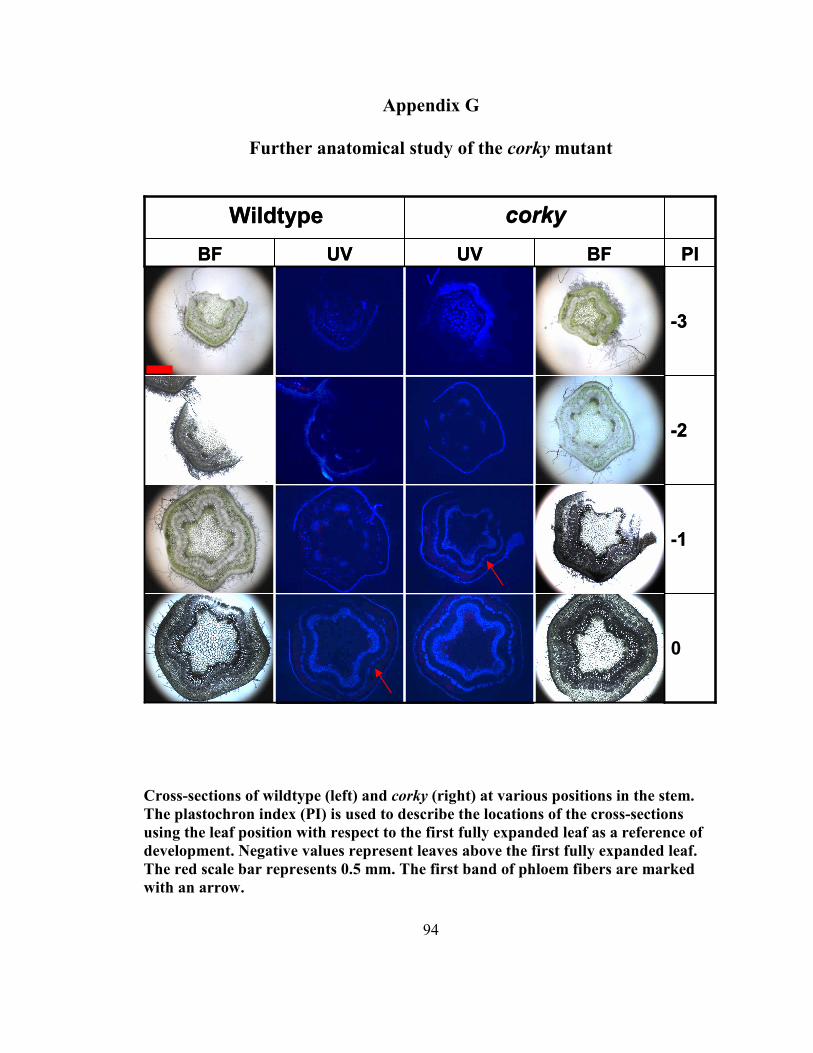
94
Appendix G
Further anatomical study of the corky mutant
Cross-sections of wildtype (left) and corky (right) at various positions in the stem.
The plastochron index (PI) is used to describe the locations of the cross-sections
using the leaf position with respect to the first fully expanded leaf as a reference of
development. �egative values represent leaves above the first fully expanded leaf.
The red scale bar represents 0.5 mm. The first band of phloem fibers are marked
with an arrow.
1
- 2
- 3
PI BF UV UV BF
corky Wildtype
0
- 1
- 2
- 3
PI BF UV UV BF
corky Wildtype
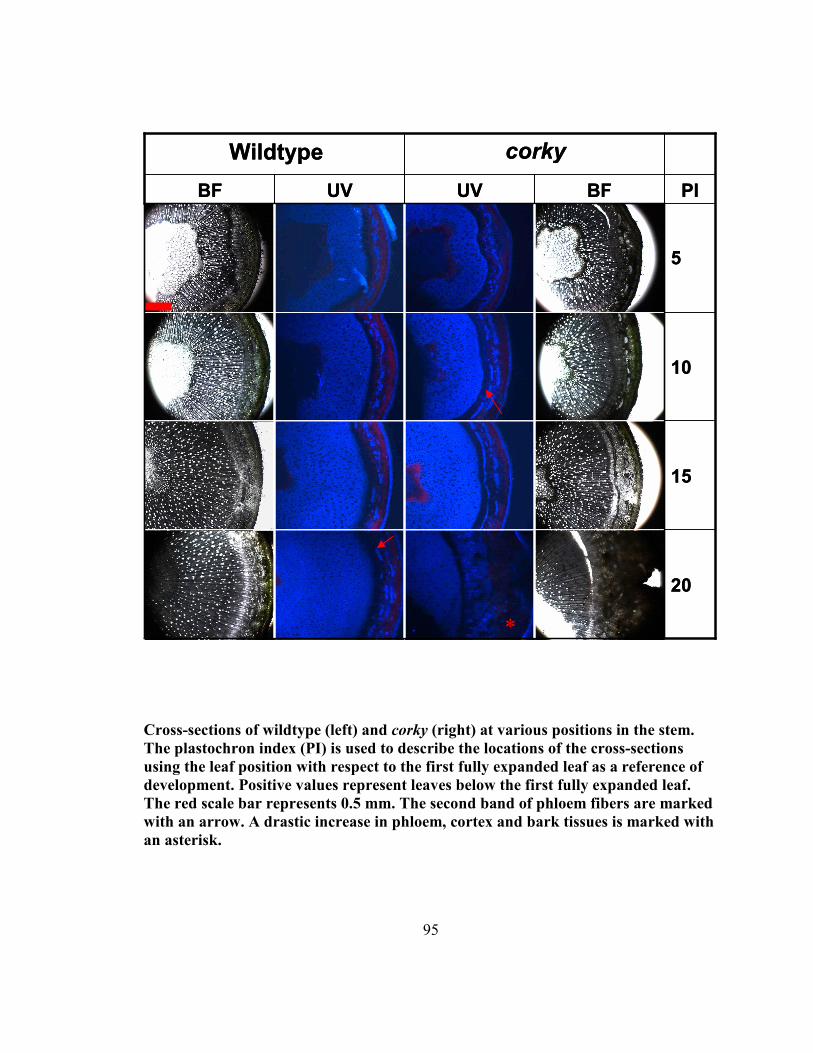
95
Cross-sections of wildtype (left) and corky (right) at various positions in the stem.
The plastochron index (PI) is used to describe the locations of the cross-sections
using the leaf position with respect to the first fully expanded leaf as a reference of
development. Positive values represent leaves below the first fully expanded leaf.
The red scale bar represents 0.5 mm. The second band of phloem fibers are marked
with an arrow. A drastic increase in phloem, cortex and bark tissues is marked with
an asterisk.
20 20
15
10
5
PI BF UV UV BF
corky Wildtype
15
10
5
PI BF UV UV BF
corky Wildtype
*
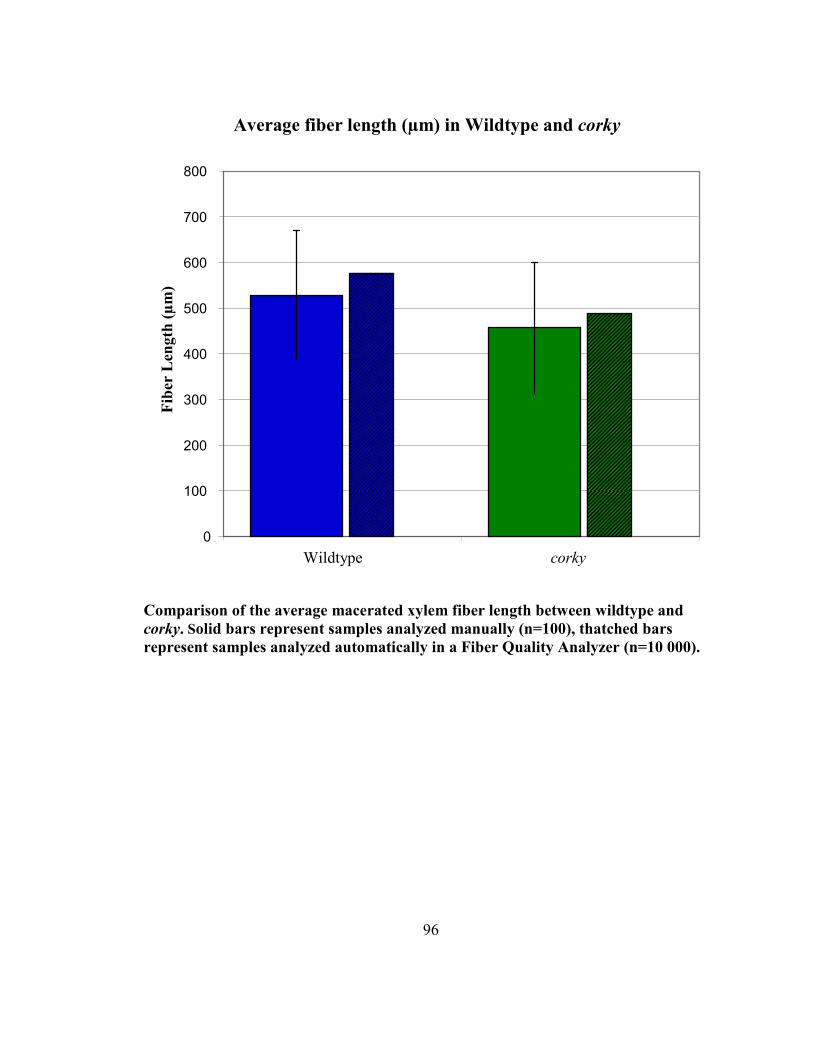
96
Comparison of the average macerated xylem fiber length between wildtype and
corky. Solid bars represent samples analyzed manually (n=100), thatched bars
represent samples analyzed automatically in a Fiber Quality Analyzer (n=10 000).
Average fiber length (µm) in Wildtype and corky
0
100
200
300
400
500
600
700
800
Wildtype corky
Fib
er L
eng
th (
µm
)
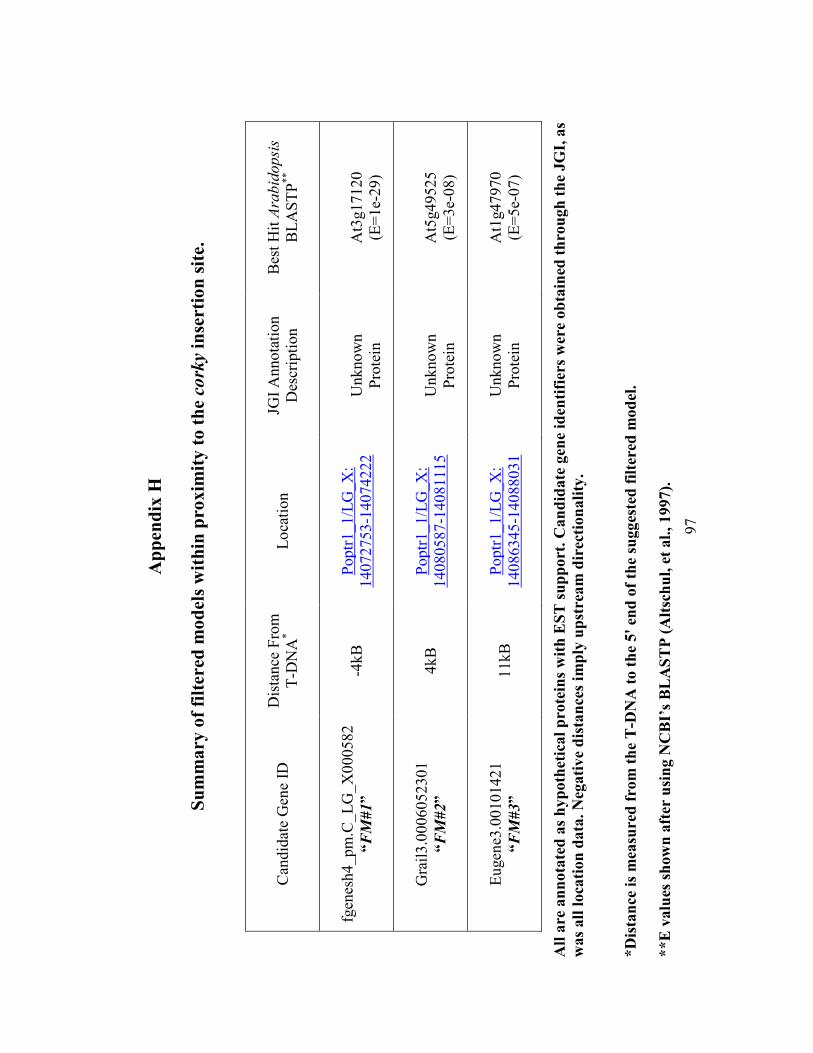
97
Ap
pen
dix
H
Su
mm
ary
of
filt
ered
mo
del
s w
ith
in p
roxim
ity
to
th
e co
rky
inse
rtio
n s
ite.
Can
did
ate
Gen
e ID
Dis
tance
Fro
m
T-D
NA
*
Lo
cati
on
JGI
Annota
tion
Des
crip
tion
Bes
t H
it A
rabid
opsi
s
BL
AS
TP
**
fgen
esh4_pm
.C_
LG
_X
000582
“FM
#1”
-4kB
Poptr
1_1/L
G_X
:
14072753-1
4074222
Unknow
n
Pro
tein
At3
g17120
(E=
1e-
29
)
Gra
il3.0
006052301
“FM
#2”
4kB
Poptr
1_1/L
G_X
:
14080587-1
4081115
Unknow
n
Pro
tein
At5
g49525
(E=
3e-
08
)
Eugen
e3.0
0101421
“FM
#3”
11kB
Poptr
1_1/L
G_X
:
14086345-1
4088031
Unknow
n
Pro
tein
At1
g47970
(E=
5e-
07
)
All
are
an
nota
ted
as
hyp
oth
etic
al
pro
tein
s w
ith
ES
T s
up
port
. C
an
did
ate
gen
e id
enti
fier
s w
ere
ob
tain
ed t
hro
ugh
th
e JG
I, a
s
was
all
loca
tion
data
. �
egati
ve
dis
tan
ces
imp
ly u
pst
rea
m d
irec
tion
ali
ty.
*D
ista
nce
is
mea
sure
d f
rom
th
e T
-D�
A t
o t
he
5'
end
of
the
sugges
ted
fil
tere
d m
od
el.
**E
valu
es s
how
n a
fter
usi
ng �
CB
I’s
BL
AS
TP
(A
ltsc
hu
l, e
t al.
, 1997).
98
Ap
pen
dix
I
Ref
eren
ce l
ist
of
bio
info
rma
tic
too
ls u
tili
zed
in
Ch
ap
ters
2 a
nd
3
Tool
Host
L
ink
R
efer
ence
Fu
nct
ion
Ath
ena
WS
U
ww
w.b
ioin
form
atic
s2.w
su.e
du/A
thena/
(O
'Connor,
et
al., 2
005)
Pro
mote
r el
emen
t an
alysi
s
Cel
l E
FP
Bro
wse
r B
AR
w
ww
.bar
.uto
ronto
.ca/
cell
_efp
/cgi-
bin
/cel
l_ef
p.c
gi
(Win
ter,
et
al., 2
007)
Subce
llula
r lo
cali
zati
on
Clu
stal
W
EM
BL
-EB
I w
ww
.eb
i.ac
.uk/T
oo
ls/c
lust
alw
2/i
nd
ex.h
tml
(Lar
kin
, et
al.
, 2007)
Ali
gnm
ent
gen
erat
or
CD
D -
Conse
rved
Dom
ain D
atab
ase
N
CB
I w
ww
.ncb
i.n
lm.n
ih.g
ov/S
truct
ure
/cd
d/c
dd
.shtm
l
(Mar
chle
r-B
auer
, et
al.
,
2007)
Det
ecti
on o
f co
nse
rved
moti
fs
in p
rote
in s
equen
ces
DA
S -
Den
se
Ali
gnm
ent
Surf
ace
S
BC
w
ww
.sb
c.su
.se/
~m
iklo
s/D
AS
/
(Cse
rzo, et
al.
, 1997)
Tra
nsm
embra
ne
dom
ain a
nd
signal
pep
tide
det
ecti
on
Inte
rPro
Sca
n
EM
BL
-EB
I w
ww
.eb
i.ac
.uk/T
oo
ls/I
nte
rPro
Sca
n/
(Z
dobnov a
nd A
pw
eile
r,
2001)
Tra
nsm
embra
ne
dom
ain a
nd
signal
pep
tide
det
ecti
on
PA
RA
LO
G
KE
GG
w
ww
.geno
me.
jp/k
egg/s
sdb
/
(Ogat
a, e
t al
., 1
999)
Hom
olo
gy d
etec
tion
Phobiu
s S
BC
htt
p:/
/pho
biu
s.sb
c.su
.se/
(K
äll,
et
al., 2
004)
T
ransm
embra
ne
dom
ain a
nd
signal
pep
tide
det
ecti
on
Pla
nt
Tri
bes
Flo
ral
Gen
om
e
Pro
ject
htt
p:/
/fgp
.huck.p
su.e
du/t
rib
edb
/ind
ex.p
l (W
all,
et
al., 2
008)
Iden
tifi
cati
on o
f pote
nti
al
gen
e fa
mil
ies
Populu
sDB
ww
w.p
op
ulu
s.d
b.u
mu.s
e/
(Ste
rky,
et a
l., 2004)
Populu
s ex
pre
ssio
n d
atab
ase
PR
AL
INE
IB
IVU
htt
p:/
/zeus.
cs.
vu.n
l/p
rogra
ms/
pra
linew
ww
/
(Sim
oss
is, et
al.
, 2005)
Ali
gnm
ent
gen
erat
or
PR
OT
PA
RA
M
Ex
pas
y
htt
p:/
/ca.
exp
asy.o
rg/t
oo
ls/p
rotp
aram
.htm
l (G
aste
iger
, et
al.
, 2005)
Pro
tein
ph
ysi
cal
par
amet
er
calc
ula
tor
PT
arget
U
of
Alb
any
htt
p:/
/bio
app
s.ri
t.al
ban
y.e
du/p
TA
RG
ET
(G
uda,
2006)
Subce
llula
r lo
cali
zati
on
TM
RP
res2
D
U o
f A
then
s htt
p:/
/bio
info
rmat
ics.
bio
l.uo
a.gr/
TM
RP
res2
D/
(S
pyro
poulo
s, e
t al
., 2
004)
Sec
ondar
y s
tru
cture
gra
phic
s
99
BA
R
Bota
ny A
rray R
esourc
e h
ttp:/
/ww
w.b
ar.u
toro
nto
.ca
DD
BJ
D
NA
Dat
a B
ank o
f Ja
pan
h
ttp:/
//w
ww
.ddbj.
nig
.ac.
jp/
EM
BL
-EB
I
Euro
pea
n M
ole
cula
r B
iolo
gy L
ab –
E
uro
pea
n
Bio
info
rmat
ics
Inst
itute
h
ttp:/
//w
ww
.ebi.
ac.u
k/
IBIV
U
Cen
tre
for
Inte
gra
tive
Bio
info
rmat
ics
Vri
ge
Univ
ersi
ty
htt
p:/
//w
ww
.ibi.
vu.n
l/
JGI
J
oin
t G
enom
ics
Inst
itute
h
ttp:/
//gen
om
e.jg
i-psf
.org
/Poptr
1_1/P
optr
1_1.h
om
e.htm
l
KE
GG
K
yoto
En
cycl
oped
ia o
f G
enes
and G
enom
es
htt
p:/
//w
ww
.keg
g.c
om
/
NC
BI
N
atio
nal
Cen
tre
of
Bio
info
rmat
ic I
nfo
rmat
ion
htt
p:/
//w
ww
.ncb
i.nlm
.nih
.gov/
SB
C
Sto
ckholm
Bio
info
rmat
ics
Cen
ter
htt
p:/
//w
ww
.sbc.
su.s
e/
SIG
nA
L
Sal
k I
nst
itute
Gen
om
ic A
nal
ysi
s L
abora
tory
h
ttp:/
/sig
nal
.sal
k.e
du/
TA
IR
The
Ara
bid
opsi
s In
form
atio
n R
esourc
e h
ttp:/
//w
ww
.ara
bid
opsi
s.org
/
TIG
R
The
Inst
itute
for
Gen
om
ic R
esea
rch
htt
p:/
//w
ww
.tig
r.org
/tdb/e
2k1/a
th1/
WS
U
Was
hin
gto
n S
tate
Univ
ersi
ty -
Pan
theo
n
htt
p:/
/ww
w.b
ioin
form
atic
s2.w
su.e
du/P
anth
eon/

100
Appendix J
Athena promoter analysis on potential Arabidopsis family members of
FM#2

101
Appendix K
Prediction of secondary structure for FM#2.
Cytoplasmic and extracellular are default labels assigned by the program.The
compartment that FM#2 would be targeted to is unknown.
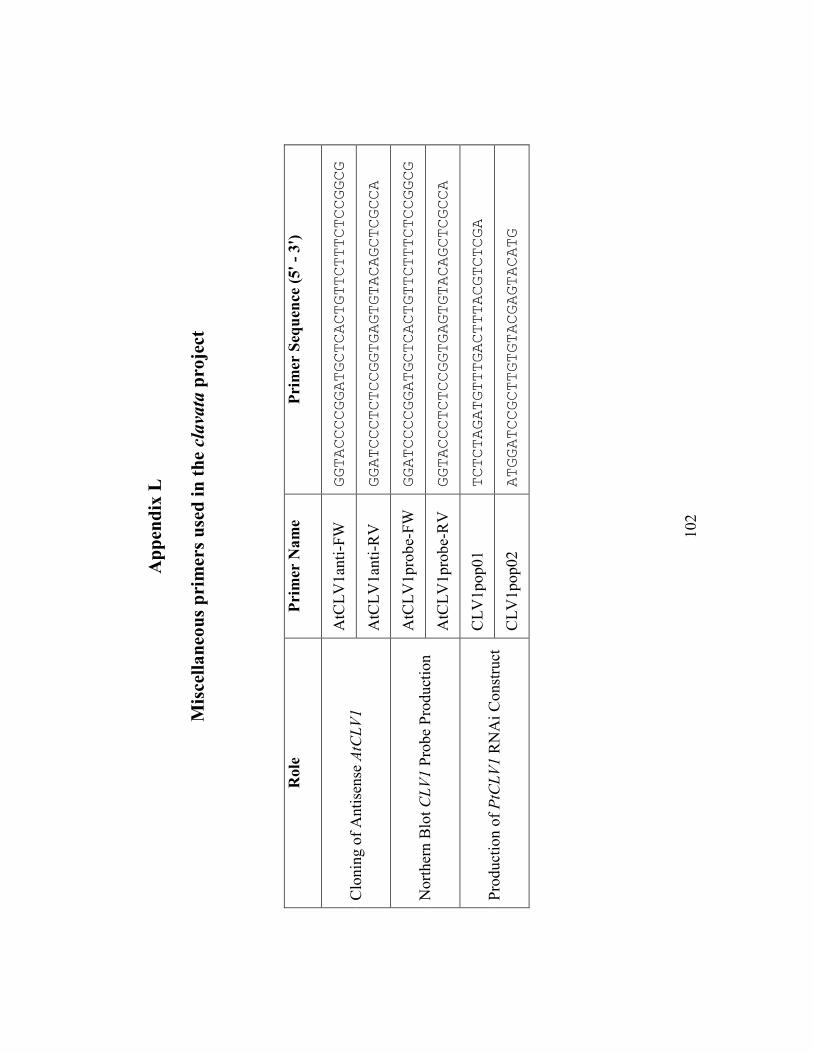
102
Ap
pen
dix
L
Mis
cell
an
eou
s p
rim
ers
use
d i
n t
he clava
ta p
roje
ct
Role
P
rim
er �
am
e P
rim
er S
equ
ence
(5'
- 3
')
AtC
LV
1an
ti-F
W
GGTACCCCGGATGCTCACTGTTCTTTCTCCGGCG
Clo
nin
g o
f A
nti
sense
AtC
LV
1
AtC
LV
1an
ti-R
V
GGATCCCTCTCCGGTGAGTGTACAGCTCGCCA
AtC
LV
1pro
be-
FW
GGATCCCCGGATGCTCACTGTTCTTTCTCCGGCG
Nort
her
n B
lot
CL
V1 P
robe
Pro
duct
ion
AtC
LV
1pro
be-
RV
GGTACCCTCTCCGGTGAGTGTACAGCTCGCCA
CL
V1pop01
TCTCTAGATGTTTGACTTTACGTCTCGA
Pro
duct
ion o
f P
tCL
V1 R
NA
i C
onst
ruct
C
LV
1pop02
ATGGATCCGCTTGTGTACGAGTACATG
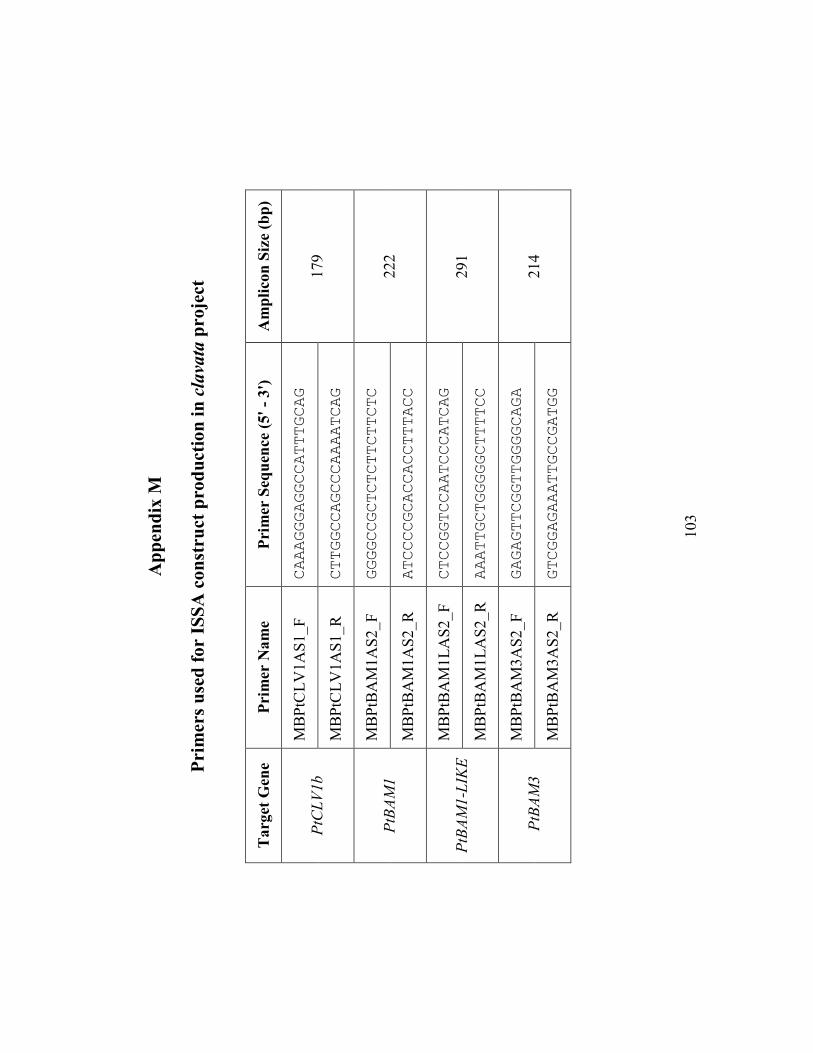
103
Ap
pen
dix
M
Pri
mer
s u
sed
fo
r IS
SA
co
nst
ruct
pro
du
ctio
n i
n clava
ta p
roje
ct
Targ
et G
ene
Pri
mer
�a
me
Pri
mer
Seq
uen
ce (
5'
- 3
')
Am
pli
con
Siz
e (b
p)
MB
PtC
LV
1A
S1_F
CAAAGGGAGGCCATTTGCAG
PtC
LV
1b
MB
PtC
LV
1A
S1_R
CTTGGCCAGCCCAAAATCAG
179
MB
PtB
AM
1A
S2_F
GGGGCCGCTCTCTTCTTCTC
PtB
AM
1
MB
PtB
AM
1A
S2_R
ATCCCCGCACCACCTTTACC
222
MB
PtB
AM
1L
AS
2_F
CTCCGGTCCAATCCCATCAG
PtB
AM
1-L
IKE
MB
PtB
AM
1L
AS
2_R
AAATTGCTGGGGGCTTTTCC
291
MB
PtB
AM
3A
S2_F
GAGAGTTCGGTTGGGGCAGA
PtB
AM
3
MB
PtB
AM
3A
S2_R
GTCGGAGAAATTGCCGATGG
214

104
Appendix �
A diagrammatic representation of ISSA amplicons.
Yellow circle indicates the amplicon location. Red lines represent a BLASTP (via
JGI) result showing conservation between filtered models. A single construct was
designed for PtCLV1, PtBAM1, PtBAM1-LIKE and PtBAM3 to be used in both
antisense and hairpin-R�Ai vectors.
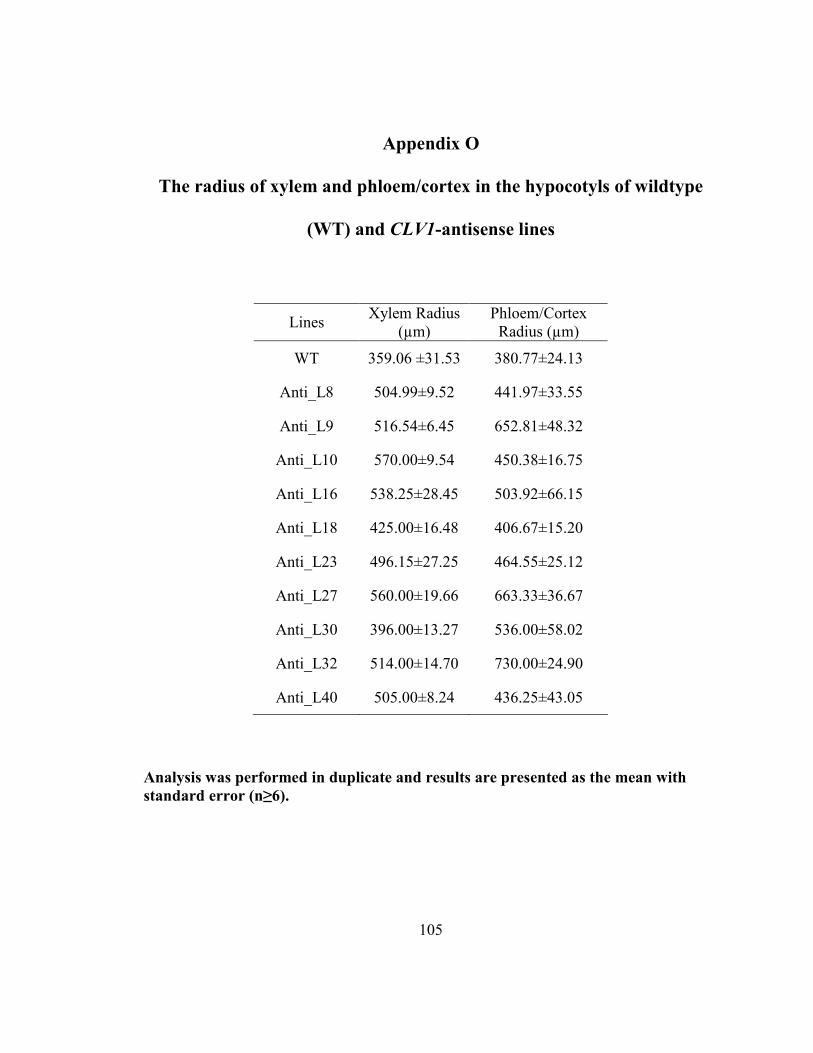
105
Appendix O
The radius of xylem and phloem/cortex in the hypocotyls of wildtype
(WT) and CLV1-antisense lines
Lines Xylem Radius
(µm)
Phloem/Cortex
Radius (µm)
WT 359.06 ±31.53 380.77±24.13
Anti_L8 504.99±9.52 441.97±33.55
Anti_L9 516.54±6.45 652.81±48.32
Anti_L10 570.00±9.54 450.38±16.75
Anti_L16 538.25±28.45 503.92±66.15
Anti_L18 425.00±16.48 406.67±15.20
Anti_L23 496.15±27.25 464.55±25.12
Anti_L27 560.00±19.66 663.33±36.67
Anti_L30 396.00±13.27 536.00±58.02
Anti_L32 514.00±14.70 730.00±24.90
Anti_L40 505.00±8.24 436.25±43.05
Analysis was performed in duplicate and results are presented as the mean with
standard error (n≥6).
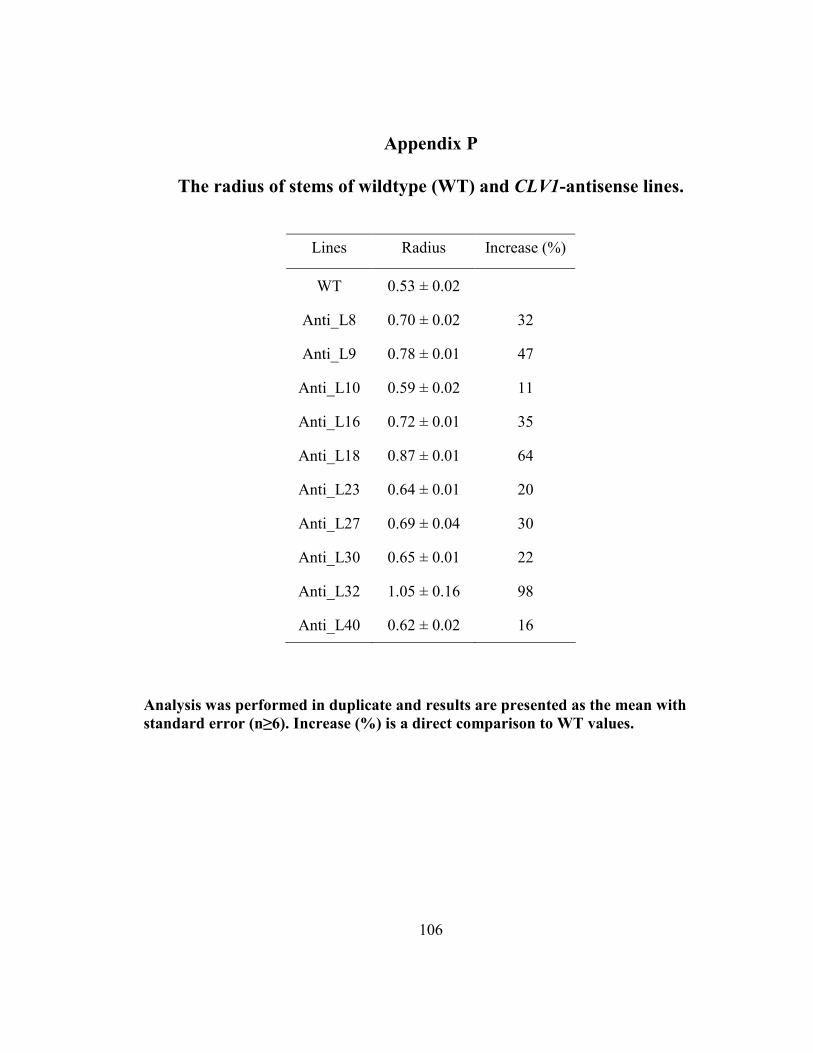
106
Appendix P
The radius of stems of wildtype (WT) and CLV1-antisense lines.
Lines Radius Increase (%)
WT 0.53 ± 0.02
Anti_L8 0.70 ± 0.02 32
Anti_L9 0.78 ± 0.01 47
Anti_L10 0.59 ± 0.02 11
Anti_L16 0.72 ± 0.01 35
Anti_L18 0.87 ± 0.01 64
Anti_L23 0.64 ± 0.01 20
Anti_L27 0.69 ± 0.04 30
Anti_L30 0.65 ± 0.01 22
Anti_L32 1.05 ± 0.16 98
Anti_L40 0.62 ± 0.02 16
Analysis was performed in duplicate and results are presented as the mean with
standard error (n≥6). Increase (%) is a direct comparison to WT values.

107
Appendix Q
Pileup of Arabidopsis and Populus CLV1 and BAM family members
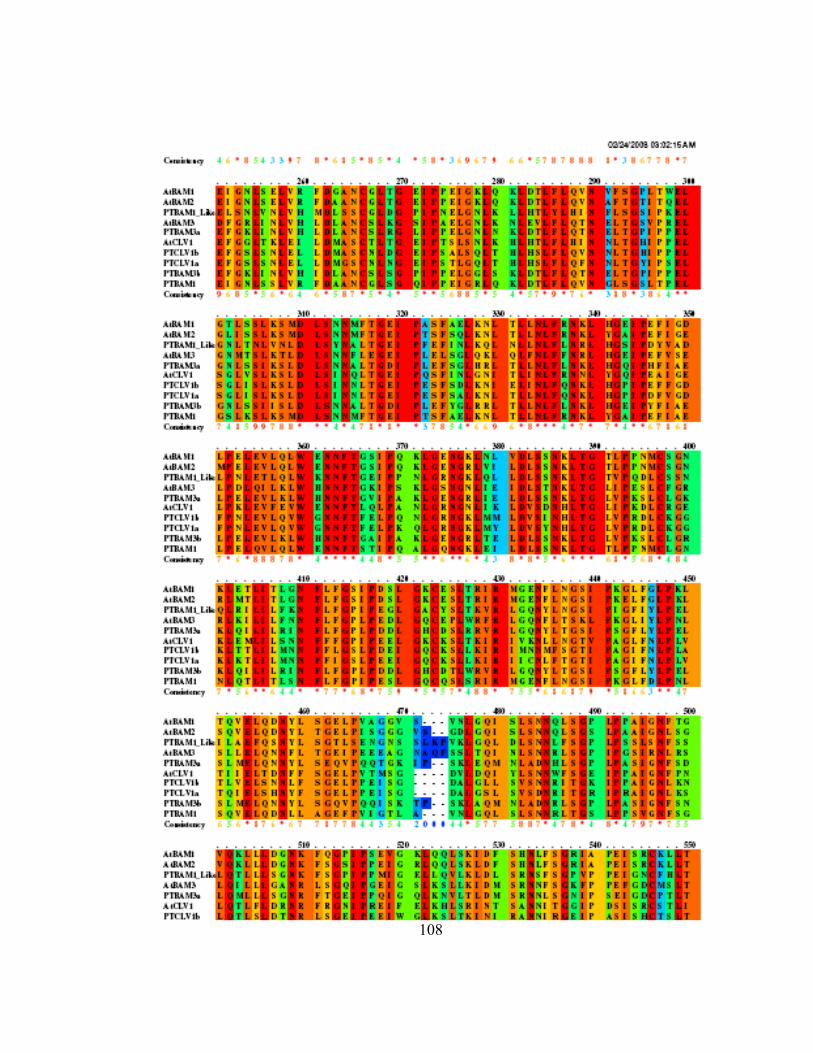
108
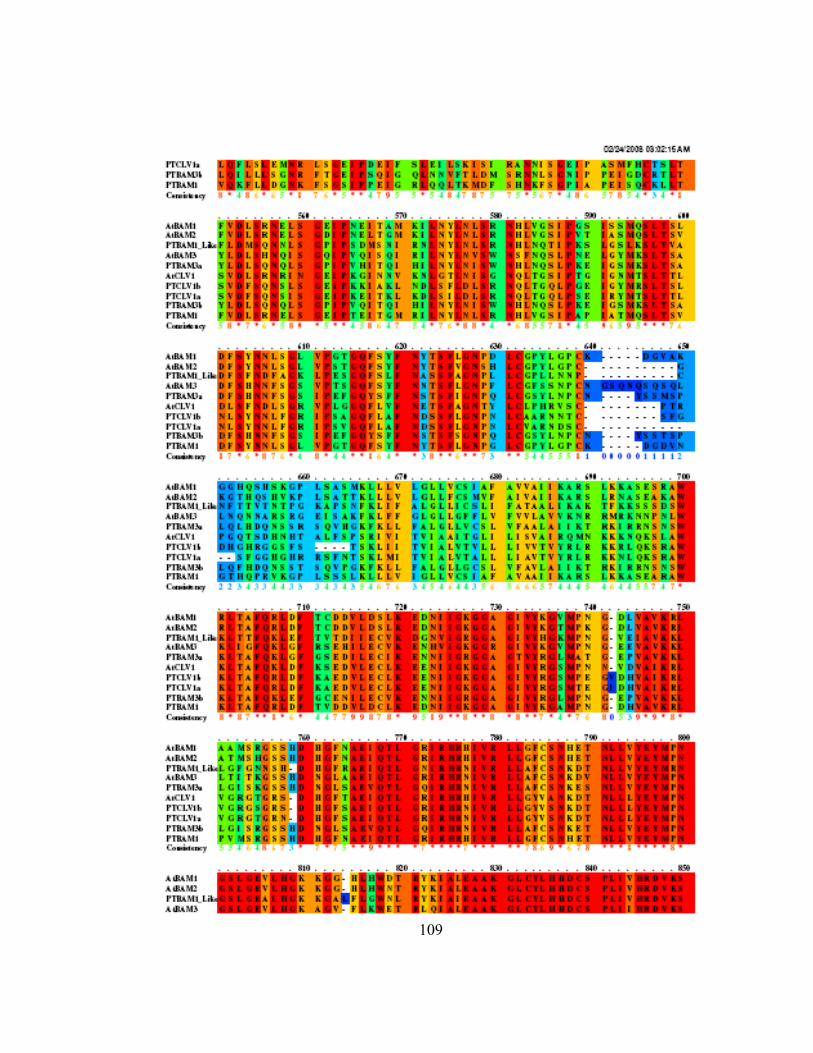
109

110

111
Appendix R
Model for FM#2 and CLAVATA1 function in secondary growth
Vascular CambiumVascular CambiumCork CambiumCork Cambium
Secondary Growth Regulators
corky
WildtypeWildtype
clavataclavata--11
112
Ap
pen
dix
S
Co
nst
ruct
flo
wch
art
fo
r co
rky
an
d clava
ta p
roje
cts
Pro
ject
In
sert
Am
pli
fied
P
GE
MT
EA
SY
Pla
sm
id
Lig
ati
on
pC
AM
BIA
E.c
oli
pC
AM
BIA
A
gro
P
op
lar
Gen
era
tio
n
Sen
se (
MA
-
OR
F)
X
X
X
X
X
α-S
en
se (
FR
O-
AM
) X
X
X
X
X
FA
TI
(OR
F)
X
X
X
X
X
Pro
m1
Pro
m2
Pro
m3
In
sert
Am
pli
fied
P
GE
MT
EA
SY
Pla
sm
id
Lig
ati
on
pC
AM
BIA
E.c
oli
pC
AM
BIA
A
gro
A
ra
bid
op
sis
G
en
era
tio
n
At5
g4
95
25
(Fu
sio
n)
X
X
X
X
X
X
T2
At5
g4
95
25
(MA
-OR
F)
X
X
X
X
X
At5
g2
67
90
(Fu
sio
n)
X
X
X
X
X
X
T2
Gra
il
At5
g2
67
90
(MA
-OR
F)
X
X
X
X
X
113
P
op
lar
Pro
ject
In
sert
T
OP
O
Clo
ned
P
lasm
id
L/R
Rec
om
bin
ase
E.coli
Agro
717
co
rky
Gu
s?
pC
ambia
1305.1
X
X
X
GW
+
X
X
X
Con
trol
Vec
tors
GW
hp
X
X
X
Sen
se
GW
+
X
X
X
X
αS
ense
G
W+
X
X
X
X
X
RN
Ai2
50
X
GW
hp
X
X
X
X
X
G
rail
RN
Ai2
2
X
GW
hp
X
X
X
X
X
GW
+
X
X
X
X
C
AD
AS
X
G
Wh
p
X
X
X
X
GW
+
X
X
X
X
C
LV
1A
S
X
GW
hp
X
X
X
X
GW
+
X
X
X
X
B
AM
1A
S
X
GW
hp
X
X
X
X
GW
+
X
X
X
X
B
AM
LA
S
X
GW
hp
X
X
X
X
GW
+
X
X
X
X
B
AM
3A
S
X
GW
hp
X
X
X
X
CL
V/B
AM
CA
DH
P
X
GW
hp
X
X
X
X





























