Embed Size (px)
Citation preview

Host Genetic Risk Factors for Community-acquired Pneumonia
Lyubov E. Salnikova, Dr. Sci. (Biol.)a,b,*, Tamara V. Smelaya, Dr. Sci. (Med.)b, Viktor V.
Moroz, Corresponding. Member of Russian Acad. Med. Sci.b, Arkady M. Golubev, Dr. Sci.
(Med.)b, Alexander V. Rubanovich, Dr. Sci. (Biol.)a
aN.I. Vavilov Institute of General Genetics, Russian Academy of Sciences, 3 Gubkin Street,
Moscow 117971, Russia
bV. A. Negovsky Research Institute of General Reanimatology, Russian Academy of Medical
Sciences, 25 Petrovka Street, Build.2, Moscow 107031, Russia
* Correspondence: Lyubov E. Salnikova, Institute of General Genetics, Russian Academy of
Sciences, 3 Gubkin Street, Moscow 117971, Russia, phone 74991328958, fax 4991328962
E-mail: [email protected]
Abstract
This study was conducted to establish the contribution of genetic host factors in the
susceptibility to community acquired pneumonia (CAP) in Russian population. Patients with
CAP (n = 334), volunteers without a previous history of CAP, constantly exposed to
infectious agents, control A group (n=141) and a second control group B consisted of healthy
persons (n=314) were included in the study. All subjects were genotyped for 13 polymorphic
variants in the genes of xenobiotics detoxification CYP1A1 (rs2606345, rs4646903,
rs1048943), GSTM1 (Ins/del), GSTT1 (Ins/del), ABCB1 rs1045642); immune and
inflammation response IL-6 (rs1800795), TNF-a (rs1800629), MBL2 (rs7096206), CCR5
(rs333), NOS3 (rs1799983), angiotensin-converting enzyme ACE (rs4340), and occlusive
vascular disease/hyperhomocysteinemia MTHFR (rs1801133). Seven polymorphic variants in
genes CYP1A1, GSTM1, ABCB1, NOS3, IL6, CCR5 and ACE were associated with CAP. For
two genes CYP1A1 and GSTM1 associations remained significant after correction for
multiple comparisons. Multiple analysis by the number of all risk genotypes showed a highly

significant association with CAP (P=2.4 x 10-7, OR=3.03, 95% CI 1.98-4.64) with the
threshold for three risk genotypes. Using the ROC-analysis, the AUC value for multi-locus
model was estimated as 68.38.
Highlight:
To establish the contribution of genetic host factors in the susceptibility to community
acquired pneumonia (CAP) in Russian population. Rs2606345 (CYP1A1) is strongly
associated with the risk of CAP. Genetic predisposition to CAP is attributed to cumulative
contribution of polymorphisms at a number of genes involved in xenobiotics detoxification,
immune and inflammation response and renin-angiotensin system.
Key words:
Community-acquired pneumonia, gene polymorphism, multi-locus model, ROC analysis,
CYP1A1
Abbreviations:
AIC, Akaike information criterion; AUC, area under curve, CAP, community acquired
pneumonia; CI, confidence interval; CNV, copy number variations, Del (D), deletion; FDR,
false discovery rate; HWE, Hardy-Weinberg equilibrium; HWP, Hardy-Weinberg
probability; Ins (I), insertion; LD, linkage disequilibrium; OR, odds ratio; ROC, receiver
operating characteristic; SE, standard error; Se, sensitivity; SNP, single-nucleotide
polymorphism, Sp, specificity.

1. Introduction
An estimated 1500000 cases and 44400 deaths occur annually from community
acquired pneumonia in Russia. The analysis of genetic and environmental risk factors
predisposing to CAP is clearly warranted. Risk of CAP has been associated with pathogen
virulence, host susceptibility and epidemiologic factors. It has been shown that polymorphic
variants at some host genes can modify risk of CAP, including those critical for the host
response to CAP - innate immune system (Endeman et al., 2009; Endeman et al., 2008;
Endeman et al., 2011; Gallagher et al., 2003; Garcia-Laorden et al., 2008; Gentile et al.,
2003; Gomi et al., 2004; Herpers et al., 2009; Lingappa et al., 2011; Martín-Loeches et al.,
2012; Mukamal et al., 2010; Schaaf et al., 2005; Solé-Violán et al., 2010; Solé-Violán et al.,
2011; Waterer et al., 2001; Wunderink et al., 2002; Yuan et al., 2008; Zúñiga et al., 2012),
the lung's defense against inhaled microorganisms (García-Laorden et al., 2011; Lingappa et
al., 2011), inhibition of fibrinolysis (Yende et al., 2007) and renin-angiotensin system (de
Garde et al., 2008; Morimoto et al., 2002; Takahashi et al., 2005).
Given the results of the abovementioned studies, we have selected 13 polymorphic
variants which were assigned to 11 different genes based on their potential association with
molecular pathways implicated in CAP pathogenesis, including both previously unexplored
variations and variations reported to be associated with susceptibility to CAP, and/or its
severity and outcome for the latter to reveal significance of previously identified associations
in Russian population (IL-6 rs1800795, TNF-a rs1800629, MBL2 rs7096206, ACE rs4340).
Xenobiotics detoxification genes (CYP1A1 rs2606345, rs4646903, rs1048943, GSTM1
Ins/del, GSTT1 Ins/del, ABCB1 rs1045642) were also included in our study as they encode
enzymes involved in the detoxification and excretion of a broad range of exogenous and
endogenous compounds, thereby participating in general resistance to occupational hazards
and endogenous harmful metabolites as well as in biotransformation of drugs. Two more

genes involved in immune and inflammation response (CCR5 rs333, NOS3 rs1799983) were
selected as the receptor obliteration (CCR5) and nitric oxide production (NOS3) might
influence CAP initiation and progression. The MTHFR locus (rs1801133) was chosen as
gene-candidate for occlusive vascular disease and hyperhomocysteinemia, the both
syndromes possibly extending inflammation processes.
2. Materials and methods
2.1. Patients and controls
334 patients with CAP (age 26.93 ± 0.82 years, 91.9% males), hospitalized at the
clinical bases of V. A. Negovsky Research Institute of General Reanimatology, Moscow,
Russia, were included in the study. Confirmed diagnosis of pneumonia was based on the
presence of acute symptoms resulted from lower respiratory tract infection, confirmed by
clinical, radiological and laboratory data.
Initially control group was composed of 188 unrelated healthy volunteers without a
previous history of relevant infectious diseases. Volunteers were medical workers, the staff of
the auxiliaries (drivers, technicians, mechanics, electricians, security officers). The subjects
from the control group were in contact with patients and therefore they were constantly
exposed to infectious agents. All volunteers started to work in 2005 year or later and were
under medical supervision in 2007-2012 years. To date, 141 subjects from this group have
not contracted CAP and were therefore included in Control group A (age 21.06 ± 0.42 years,
92.5% males). A second control group (control B) consisted of 314 healthy persons (41.65 ±
1.03 years, 91.1% males).
All patients and control subjects were Russian and were residents of the Central Federal
District of European Russia. Investigated CAP and control A groups were rather
homogeneous as to the age of those under study (Figure 1) and their sex (91.9% and 92.2%
male in control A and CAP groups correspondingly).

The study was approved by the Inter-Institutional Ethical Committee and was conducted
according to the principles of the Declaration of Helsinki. All participants provided written
informed consent.
2.2. Genotyping
DNA was isolated from 200 μl of blood using gDNA purification kit Diatom DNA
Prep 200 (Isogene laboratory, Moscow, Russia). The genotyping was performed using an
allele-specific tetraprimer PCR developed to genotype a relatively large number of samples in
a cost-effective and time-saving manner. In this method, allele-specific DNA products are
amplified by means of applying appropriately designed two-pair primers (four primers) into
an ordinary PCR tube (Hamajima, 2001). Amplification was carried out on an ABI thermal
cycler using two external and two internal sequence-specific primers (Table 2) and tubes
PCR MasterMix (Isogene laboratory, Moscow, Russia) as previously described (Moroz et al.,
2011). For each SNP, 10% of randomly taken DNA samples (cases and controls) were
genotyped twice and no discrepancies were observed. The PCR data were validated by
sequencing (Evrogen, Moscow, Russia).
2.3. Statistical analysis
Exact Hardy Weinberg Equilibrium (HWE) tests were performed for each SNP
independently using the goodness-of-fit χ2 test to compare the observed and expected
genotype frequencies for CAP and control groups. Single SNP statistical analysis was
performed using WinSTAT software (Robert K. Fitch Software, Germany). Two-tailed
Fisher’s exact test was also performed to evaluate association between SNP and disease.
Odds ratios were estimated using WINPEPI software (Abramson, 2011). The distributions of
genotypes in cases and controls were compared using a logistic regression analysis.

Using SNPStats package, five genetic models were considered (codominant,
dominant, recessive, over-dominant and log-additive), and the best genetic model was
selected using AIC value. The lowest AIC value was considered the best-fitting model for the
fitted variant. For genotypes with minor allele frequencies <5% only dominant genetic
models were considered (minor homozygotes+heterozygotes vs. major homozygotes).
Bonferroni correction and FDR approach were used for adjustment for multiple
comparisons. FDR approach has been well-documented in the literature as a means to control
type I error while maximizing power of analysis (Storey, 2002; Benjamini and Yekutieli,
2002). In contrast to Bonferroni correction, false discovery rate controls the expected
proportion of false discoveries among all rejected hypotheses. WINPEPI test power and
sample size calculators were used to evaluate type II error.
Pairwise LDs were measured as Lewontin’s D’-values and estimated from genotype
data using the expectation-maximization algorithm implemented in SNPStats software (Sole
et al., 2006). The SNPStats software was also used to estimate haplotype frequencies in both
case and control groups as well as for evaluation of haplotype association with CAP by
individual haplotype P-values. To estimate the cumulative effect, variables were coded as 0
and for the protective and risk genotypes, respectively.
3. Results
In total, 304 patients with pneumonia and 455 control subjects were included in the
study. The main demographic and clinical variables of the patients and control subjects are
shown in Table 1. Patients and controls were predominantly male. Bilateral radiographic
opacities were
observed in 98 (29.3%) patients. In 108 (32.3%) cases a causative agent could be identified.
Streptococcus pneumoniae was the most common etiologic agent (70, 64.8% of the patients
with known causative microorganism).

V. A. Negovsky Research Institute of General Reanimatology specializes in the
general critical care. The clinical departments of the Institute include pulmonary divisions
and intensive care units of several hospitals. CAP patients with severe pneumonia and
pulmonary complications hospitalized at the clinical bases of the Institute often required more
intensive treatment, admission to an ICU and prolonged hospital stay. The overall median
duration of hospital stay was 27.40±10.16 days and 178 patients (53.3%) were admitted to
the intensive care ward.
3.1. Individual SNP analysis
13 polymorphic variants (11 SNPs and 2 CNV - GSTM1, GSTT1) were genotyped in
three groups from Russian population: 334 patients with community-acquired pneumonia
(CAP), 141 control subjects from Control group A (without CAP) and 314 healthy
individuals from general population (control B). The data are presented in Table 3. SNPs
were analyzed for Hardy-Weinberg equilibrium (HWE). The majority of SNPs were in HWE,
with just few showing a marginally-significant departure from HWE. Four of the 13 SNPs
showed a significant association with CAP. A comparison of distributions between cases and
Control group A revealed significant effects of SNPs – rs2606345T (CYP1A1), rs333Ins
(CCR5), rs4340Del (ACE), with the highest association detected for the CYP1A1 locus (SNP
rs2606345, P= 3.9 x 10-5). The frequency of deletion at the GSTM1 gene in control group A
slightly exceed those in cases and control group B. CAP susceptibility was also associated
with the GSTM1 gene insertion (homozygotes and heterozygotes) in case-control A study. It
should be noted that the effect of SNP rs2606345 is determined also in cases in comparison
with Control B group (P=1.4x10-5).
We additionally applied the logistic regression analysis and the Akaike information
criterion (AIC) to establish whether the effects of the alleles are best described by any of five
genetic models (codominant, dominant, overdominant, recessive, and log-additive). In case-

control A study the effects of rs1045642T (ABCB1) were best described by the log-additive
model (P=0.049, OR=1.34, 95% CI 1.01-1.78, AIC=577); the effects of rs1800795 (IL-6) by
the over-dominant model (G/G-C/C vs. G/C; P=0.033, OR=1.55, 95% CI 1.04-2.32,
AIC=563.7); and the effects of rs1799983 (NOS3) by the over-dominant model (G/G-T/T vs.
G/T; P=0.031, OR=1.56, 95% CI 1.04-2.32, AIC 567.8). Interestingly, for the both genes
regulating inflammation and immune response NOS3 and IL6 the protective effect for
heterozygotes was found.
To exclude the false-positive associations with CAP, we used Bonferroni correction
for multiple comparisons (13 polymorphic variants in cases and two control groups; 39
comparisons). The P-value after Bonferroni correction remained significant for CYP1A1 gene
(case-control A, P=0.01; case-control B, P=0.0036). The association between GSTM1 and
CAP was also confirmed in the case-control A using the 5% FDR test (P<0.0058).
3.2. Haplotype-based association analysis
Haplotype-based association analysis evaluates the joint effects of closely linked
genetic markers on a trait of interest. When compared to its single-marker counterparts, this
multi-marker approach can be more powerful to detect associations when the causal variants
are not genotyped (de Bakker et al., 2005; Zaitlen et al., 2007), have low frequency (de
Bakker et al., 2005, Schaid, 2004), or exhibit cis-acting effects (Clark, 2004; Schaid, 2004).
To evaluate the association between the haplotype CYP1A1 (rs2606345 and two other
CYP1A1 SNPs rs4646903 and rs1048943) and CAP, we estimated the standardized measure
of LD, D' (Table 4).
The values of LD did not significantly differ between the two control groups. The
analysis of CYP1A1 haplotype association with CAP was performed for case-control A and
case-control B studies. Protective effect of rs2606345G minor allele was pronounced for its
combination with two major alleles of rs4646903T and rs1048943A (haplotype N2), but

increased significantly when combined with variant allele rs4646903C (haplotype N3). We
therefore conclude that the interaction between these two SNPs can further increase the
protective effects of CYP1A1.
3.3. Multiple SNP analysis
We next established the cumulative effects of multiple SNPs at the analyzed genes on
CAP. A highly significant association between the number of risk genotypes (CYP1A1
rs2606345 T/T, GSTM1 I/*, ABCB1 T/T, IL-6 CC-GG, NOS3 TT-GG, CCR5 I/I , ACE D/D)
and CAP was found. Thus 85 and 111 subjects from control A and CAP group possessed 0-3
risk genotypes, respectively, whereas the incidence of subjects carrying from 4 to 7 risk
genotypes in the CAP group substantially exceed that in control (180 vs. 47; OR=3.03, 95%
CI 1.98-4.64, P=2.4 x 10-7). These effects were further confirmed by the comparison of
relative frequency of risk genotypes in the CAP group and controls (Figure 2).
We used area under the curve (AUC) statistics of the receiver operating characteristic
(ROC) to assess the overall performance of estimated risk in discriminating CAP cases and
controls. ROC is a plot of the sensitivity vs. (100-specificity) of classifying cases at various
thresholds. AUC quantifies the overall ability to discriminate between those who have the
disease and those who do not have the disease and ranges from 0.5 (no effect) to 1 (complete
effect). AUC for our panel, consisting of 7 genetic variations is 68.38 (Figure 3), which is
rather high for genetic markers.
After correction for multiple comparisons, 5 genes (ABCB1, NOS3, IL6, CCR5, ACE)
in case-control A study proved to be insignificantly associated with CAP. To assay if the two
associated in single gene analysis markers (CYP1A1 and GSTM1) are responsible for the
whole significance of the association effect, multiple gene analysis was carried out for either
these or five remaining SNPs. Two-gene analysis revealed a less pronounced association

between number of risk genotypes and CAP susceptibility. For no risk genotype (N=47 in
control, N=56 in CAP group) versus 1-2 risk genotypes (N=86 in control, N=174 in CAP
group) P-value was 3.8 x 10-5 (OR=2.67, 95% CI 1.70-4.22). At the same time, the remaining
SNPs (ABCB1, NOS3, IL6, CCR5, ACE) showed the cumulative effects of these genotypes on
CAP susceptibility. For 0-2 risk genotypes (N=76 in control, N=121 in CAP) versus 3-5 risk
genotypes (N=121 in control, N=176 in CAP) association was significant P=1.6 x 10-3,
OR=1.97, 95% CI 1.30-2.99. These results suggest that the five risk genotypes ABCB1 T/T,
NOS3 T/T-G/G, IL-6 C/C-G/G, CCR5 I/I and ACE D/D contribute to the whole association
enhancing its significance and OR.
3.4. Test power analysis for case-control association study
The statistical power of association studies strongly depends on the sample size,
which is particularly true for minor alleles with frequencies below 0.25. As a number of
minor alleles with the frequencies, p, ranging between 0.06 and 0.31 (CYP1A1 rs1048943G,
p=0.06, rs4646903C, p=0.25, GSTT1 D/D, p=0.16, TNF-α rs1800629A, p= 0.14, and MBL2
rs7096206G, p=0.31) did not show significant risk associations with CAP, we estimated the
statistical power of our study. To detect OR=1.5 in our sample size for the whole case-control
A group, the power for ranges from 11.9% to 42.91% for minor allele frequencies 0.06 and
0.31, respectively (two-sided Fisher’s test). The effects of minor alleles can therefore be
detected by profiling much larger number of control and CAP-affected subjects. On the other
hand, the results of Bonferroni correction and FDR test showed that in our study the type 1,
statistical error was avoided.
4. Discussion
The results of our study demonstrate that a number of SNPs show a significant
association with CAP. We found that three xenobiotics detoxification genes (CYP1A1 - first

stage of detoxification - activation, GSTM1 - second stage - actually detoxification and
ABCB1 - P-glycoprotein - efflux transporter) are apparently (CYP1A1 and GSTM1) or
nominally (ABCB1) associated with CAP in Russian population.
CYP1A1 is by far the most widely studied human pulmonary CYP gene regulating
enzyme, expressed mainly in the epithelium of the peripheral airways, i.e. bronchiolar,
terminal bronchiolar and alveolar epithelium. CYP1A1 is highly inducible by exposure to
PAH, polychlorinated biphenyls (PCBs) and other chemicals capable of binding to and
activating the Ah receptor. The CYP1A1 SNPs included in our study are functional. Thus, the
intron-located rs2606345G/T determines enhanced gene expression in the presence of
specific substrates (allele G) or in their absence (allele T) (Rotunno et al., 2009; Wang et al.
2008). The minor allele at the CYP1A1 SNP rs4646903C, located near Gene-3, also shows an
increased inducibilty (Meletiadis et al., 2006), and, according to our data, enhances the
protective effect of rs2606345G in haplotype G-C (Table 2). Another CYP1A1 SNP
rs1048943A/G (Ile/Val), resulted in a missense amino acid substitution, is characterized by
the substrate-specific increased activity for minor allele G and is not associated with CAP
susceptibility.
Resolution of inflammation depends on the removal of apoptotic cells as on active
suppression of inflammatory mediator production. The elimination of apoptotic cells and cell
bodies by phagocytes prevent exposure of surrounding tissue to potentially cytotoxic,
immunogenic, or inflammatory cellular content (Kang et al., 2010). Proinflammatory
cytokines and endogenous and exogenous xenobiotics (for example endogenous harmful
metabolites and drugs intermediates) may interact to potentiate both DNA damage and the
inflammatory response. This interaction is compounded by the fact that TNF-α inhibits
apoptotic cell clearance in the lung, even more exacerbating acute inflammation (Borges et

al., 2009). Genetically determined variability in CYP1A1 activity may contribute to
augmentation of inflammation.
According to our data, the GSTM1 insertion represents another important component
of genetic factors predisposing to CAP. However, in contrast to the CYP1A1 effects, there is
no evidence for the interaction with occupational exposure. As the GSTM1 enzyme has a
broad substrate specificity, at the beginning of the treatment the GSTM1(+) subjects can more
efficiently metabolize a number of drugs, thus belonging to a group of “fast metabolizers”.
Another hypothesis might deal with “proapoptotic” GSTM1 activity. It was shown that
during exposure to oxidative stress and other stress stimuli, the GSTM1 protein directly
regulates Ask1 (apoptosis signal-regulating kinase 1), an upstream activator of the stress-
activated protein kinase p38 (Cho et al., 2001). It was also shown that the Ask1 activation
involves the heat shock-induced dissociation of its inhibitor, glutathione S-transferase Mu1-1
GSTM1-1 (Dorion et al., 2002). Heat shock at the beginning of inflammation might be
attributed to the specific stress stimuli running GSTM1 mediated apoptosis extending
inflammation processes.
The ABCB1 rs1045642T/T genotype was nominally associated with CAP in our
study, though the results were insignificant after correction for multiple comparisons.
Nevertheless, this risk genotype can make a certain contribution to the overall cumulative
effect of CAP-associated risk genotypes (Figure 1). The ABCB1 gene encodes a membrane-
bound transporter that actively effluxes a wide range of compounds from cells. ABCB1 is
expressed in various tissues to protect them from the adverse effect of toxins. The
pharmacokinetics of drugs that represent substrates for the ABCB1 enzyme may also
influence disease outcome and treatment efficacy. Although ABCB1 is a well conserved gene,
there is increasing evidence that its polymorphisms affect substrate specificity and may be the
expression level and/or stability of its mRNA (Fung and Gottesman, 2009). For example, it

has been shown that under experimental acute inflammation ABCB1 mRNA expression is
significantly down-regulated in PBMCs (peripheral blood mononuclear cells) obtained from
the subjects harboring 3435C in comparison to those without it (Markova et al., 2006). We
therefore suggest that during inflammation the fast efflux of drugs in the subjects carrying
3435T/T genotype may contribute to the progression of the disease.
Three polymorphic variants of genes regulating immune response and inflammation
development were associated (though insignificantly after correction for multiple
comparisons) with CAP.
It should be noted that to date the results of a number of studies on the effects of IL-6
and NOS3 genes polymorphism on inflammation remain highly controversial. Thus according
to Terry and co-authors, the IL-6 − rs1800795G/C polymorphism affects transcription by
altering the serum levels of IL-6, with the C allele associated with significantly lower levels
of plasma IL-6 (Terry et al., 2000). However, the results of the association studies are indeed
controversial as they show the deleterious effects of the minor allele IL-6 174C (Martín-
Loeches et al., 2012; Schaaf et al., 2005), the absence of association (Chauhan and McGuire,
2008; Schlüter et al., 2002), and even suggest its protective role (Marshall et al., 2002).
Functional studies of NOS3 rs1799983G/T polymorphism have demonstrated that minor T
allele codes a protein with decreased enzymatic activity (Wang et al., 2000), leading to a
reduction in nitric oxide production (Veldman et al., 2002). For NOS3 polymorphism the
deleterious effect for rs1799983T allele was demonstrated (Ma et al., 2011) as well as the
protective effect of this minor allele which may be attributed to decreased inflammatory
signaling (Hildebrandt et al., 2010).
The immune response may vary depending on the causative pathogen, clinical and
ethnic heterogeneity of studied groups and some other factors (Cooper et al., 2002; Rosseau
et al., 2007). One of them might be molecular heterosis which is common in humans and may

occur in up to 50% of all gene associations (Comings and MacMurray, 2000). Molecular
heterosis is attributed to the phenomenon of overdominance, where the phenotype of
heterozygotes is superior to that of homozygotes. Heterozygosity may facilitate the dynamic
balance between proinflammatoty and antiinflammatory factors, thus providing better defense
against exposure to a wider range of causative agents, exogenous and endogenous pathogens.
It therefore appears that the abovementioned protective effects for heterozygotes at the NOS3
and IL-6 loci may be attributed to this phenomenon.
The CCR5 chemokine receptor is exploited by HIV-1 to gain entry into the CD4+ T
cells. A deletion mutation (del32) confers resistance against HIV by obliterating the
expression of the receptor on the cell surface. The allele exists at appreciable frequencies
only in Europe, and within Europe, the frequency is higher in the north. Several studies have
demonstrated that polymorphism at the CCR5 locus in both the coding and regulatory regions
affects susceptibility to HIV infection. The much-studied CCR5-del32 allele is a 32-bp
deletion that introduces a premature stop codon into the CCR5 chemokine-receptor locus and
thus obliterates the receptor. CCR5- del32 is at average allele frequency of 10% across
Europe, translating into a homozygote frequency of about 1% (Galvani and Novembre,
2005). Bubonic plague as well as smallpox could have posed sufficient selection to account
for the rise of the CCR5-del32 allele in Europe. The central role that chemokine receptors
play in the inflammatory immune response makes it likely that the obliteration of CCR5
would have negative fitness repercussions in the absence of a compensatory protective effect.
However, to date no effects associated with this deletion have been documented in clinical
studies. Moreover, nonfunctional CCR5- del32 increases the likelihood of recovery from
hepatitis B in humans (Thio et al., 2007), reduces the risk of acute rejection in liver
transplantation (Heidenhain et al., 2009), lowers the risk of developing acute graft-versus-
host disease after allogeneic hematopoietic stem cell transplantation (Bogunia-Kubik et al.,

2006), and also attenuates the adverse effects of inflammation on overall and cardiovascular
mortality in ESRD (end-stage renal disease) (Muntinghe et al., 2009). Our data on the
protective effects del32 in CAP group are therefore in line with these results.
We also detected the protective effect of ACE insertion in CAP-control A study. The
insertion/deletion (I/D) polymorphism at this gene substantially affects the serum level of the
ACE protein. The I/D polymorphism has been reported to account for 47% of the variance in
serum ACE level, whereas the DD genotype is associated with the highest levels of serum
ACE (Rigat et al., 1990). It has also been shown that the effects of I/D polymorphism on
serum ACE level may not be directly related to the presence of a deletion per se, but
attributed to its strong linkage with the G2350A ACE allele (Zhu et al., 2001). Given that the
DD homozygotes have a lower cough reflex compared with the carries of II and ID genotypes
(Takahashi et al., 2001) and the decreased serum level of the proinflammatory angiotensin-II
(Reyes-Engel et al., 2006), the ACE I/D polymorphism can also predispose to pneumonia.
The study presented some limitations that should be addressed. Our case and control
A groups were well-stratified, but patients were rather young predominantly between the ages
of 16 and 30 (271, 81%). Our results may not be generalizable to older women or men.
In summary, we have provided the first experimental evidence for the associations of
allelic variation at genes coding detoxification enzymes with the risk of CAP. Our results
also demonstrate that predisposition to CAP is strongly attributed to the effects of a number
of genes with low penetrance and therefore imply that inter-locus interactions may be
regarded as an important component of polygenic and multifactorial factors of susceptibility
to CAP. Finally, our data on the effects of heterozygosity at immune and inflammation genes
can potentially partly explain the existing inconsistency of the results of some recent
association studies.

Acknowledgements
This work was supported by a Grant of Presidium of the Russian Academy of Sciences
program “Fundamental sciences for medicine”-2012; Fundamental Research Programs of the
Russian Academy of Sciences "Biodiversity and dynamics of gene pools" (2006-2010);
"Biological Diversity", subdivision "Gene Pools and Genetic Diversity" (2011-2012).
References
1. Abramson, J.H., 2011. WINPEPI updated: computer programs for epidemiologists,
and their teaching potential. Epidemiol Perspect Innov. 8: 1. doi:10.1186/1742-5573-
8-1.
2. de Bakker, P.I., Yelensky, R., Pe'er, I., Gabriel, S.B., Daly, M.J., Altshuler, D., 2005.
Efficiency and power in genetic association studies. Nat Genet. 37(11), 1217-1223.
3. Benjamini, Y., Yekutieli, D., 2001. The control of the false discovery rate in multiple
testing under dependency. Ann Stat. 29 (4), 1165–1188.
4. Bogunia-Kubik, K., Duda, D., Suchnicki, K., Lange A., 2006. CCR5 deletion
mutation and its association with the risk of developing acute graft-versus-host
disease after allogeneic hematopoietic stem cell transplantation. Haematologica. 91
(12), 1628-1634.
5. Borges, V.M., Vandivier, R.W., McPhillips, K.A., Kench, J.A., Morimoto, K.,
Groshong, S.D., Richens, T.R., Graham, B.B., Muldrow, A.M., Heule, L.V., Henson,
P.M., Janssen, W.J., 2009. TNFα inhibits apoptotic cell clearance in the lung,
exacerbating acute inflammation. Am J Physiol Lung Cell Mol Physiol. 297 (4),
L586–L595.
6. Chauhan, M., McGuire, W., 2008. Interleukin-6 (-174C) polymorphism and the risk

of sepsis in very low birth weight infants: meta-analysis. Arch Dis Child Fetal
Neonatal Ed. 93 (6), F427-429.
7. Cho, S.G., Lee, Y.H., Park, H.S., Ryoo, K., Kang, K.W., Park, J., Eom, S.J., Kim,
M.J., Chang, T.S., Choi, S.Y., Shim, J., Kim, Y., Dong, M.S., Lee, M.J., Kim, S.G.,
Ichijo, H., Choi, E.J., 2011. Glutathione S-transferase mu modulates the stress-
activated signals by suppressing apoptosis signal-regulating kinase 1. J Biol Chem.
276 (16), 12749-12755.
8. Clark, A.G., 2004. The role of haplotypes in candidate gene studies. Genet Epidemiol.
27(4), 321-333.
9. Comings, D.E., MacMurray, J.P., 2000. Molecular heterosis: a review. Mol Genet
Metab. 71 (1-2), 19-31.
10. Cooper, D.N., Nussbaum, R.L., Krawczak, M. 2002. Proposed guidelines for papers
describing DNA polymorphism-disease associations. Hum Genet. 110, 207–208.
DOI: 10.1007/s00439-001-0672-4
11. Dorion, S., Lambert, H., Landry, J., 2002. Activation of the 38 signaling pathway by
heat shock involves the dissociation of glutathione S-transferase Mu from Ask1. The
Journal of Biological Chemistry. 277 (34), 30792-30797.
12. Endeman, H., Cornips, M.C.A., Grutters, J.C., van den Bosch, J.M., Ruven, H.J.T.,
van Velzen-Blad, H., Rijkers, G.T., Biesma, D.H., 2009. The Fcγ Receptor IIA-
R/R131 Genotype Is Associated with Severe Sepsis in Community-Acquired
Pneumonia. Clin Vaccine Immunol. 16 (7), 1087–1090. doi: 10.1128/CVI.00037-09.
13. Endeman, H., Herpers, B.L., de Jong, B.A., Voorn, G.P., Grutters, J.C., van Velzen-
Blad, H., Biesma, D.H., 2008. Mannose-binding lectin genotypes in susceptibility to
community-acquired pneumonia. Chest. 134 (6), 1135-1140.

14. Endeman, H., Meijvis, S.C.A., Rijkers, G.T., van Velzen–Blad, H., van Moorsel,
C.H.M., Grutters, J.C., Biesma, D.H., 2011. Systemic cytokine response in patients
with community-acquired pneumonia. Eur Respir J. 37, 1431–1438.
15. Fung, K.L., Gottesman, M.M., 2009. A synonymous polymorphism in a common
MDR1 (ABCB1) haplotype shapes protein function. Biochim Biophys Acta. 1794 (5),
860–871.
16. Gallagher, P.M., Lowe, G., Fitzgerald, T., Bella, A., Greene, C.M., McElvaney, N.G.,
O'Neill, S.J., 2003. Association of IL-10 polymorphism with severity of illness in
community acquired pneumonia. Thorax. 58 (2), 154-156.
17. Galvani, A.P., Novembre J., 2005. The evolutionary history of the CCR5-Delta32
HIV-resistance mutation. Microbes Infect. 7(2), 302-309.
18. García-Laorden, M.I., Rodríguez de Castro, F., Solé-Violán, J., Rajas, O., Blanquer,
J., Borderías, L., Aspa, J., Briones, M.L., Saavedra, P., Marcos-Ramos, J.A.,
González-Quevedo, N., Sologuren, I., Herrera-Ramos, E., Ferrer, J.M., Rello, J.,
Rodríguez-Gallego, C., 2011. Influence of genetic variability at the surfactant proteins
A and D in community-acquired pneumonia: a prospective, observational, genetic
study. Crit Care. 15 (1), R57. doi: 10.1186/cc10030.
19. Garcia-Laorden, M.I., Sole-Violan, J., Rodriguez de Castro, F., Aspa, J., Briones,
M.L., Garcia-Saavedra, A., Rajas, O., Blanquer, J., Caballero-Hidalgo, A., Marcos-
Ramos, J.A., Hernandez-Lopez, J., Rodriguez-Gallego, C., 2008. Mannose-binding
lectin and mannose-binding lectin-associated serine protease 2 in susceptibility,
severity, and outcome of pneumonia in adults. J Allergy Clin Immunol. 122 (2), 368-
374, e 2. doi:10.1016/j.jaci.2008.05.037.
20. Van de Garde, E.M., Endeman, H., Deneer, V.H., Biesma, D.H., Sayed-Tabatabaei,

F.A., Ruven, H.J., Leufkens, H.G., van den Bosch, J.M., 2008. Angiotensin-
converting enzyme insertion/deletion polymorphism and risk and outcome of
pneumonia. Chest. 133 (1), 220-225.
21. Gentile, D.A., Doyle, W.J., Zeevi, A., Howe-Adams, J., Kapadia, S., Trecki, J.,
Skoner, D.P., 2003. Cytokine gene polymorphisms moderate illness severity in infants
with respiratory syncytial virus infection. Hum Immunol. 64 (3), 338-344.
22. Gomi, K., Tokue, Y., Kobayashi, T., Takahashi, H., Watanabe, A., Fujita, T., Nukiwa,
T., 2004. Mannose-binding lectin gene polymorphism is a modulating factor in
repeated respiratory infections. Chest. 126 (1), 95-99.
23. Hamajima, N., 2001. PCR–CTPP: a new genotyping technique in the era of genetic
epidemiology. Experimental Reviews in Molecular Diagnosis. 1 (1), 119–123.
24. Heidenhain, C., Puhl, G., Moench, C., Lautem, A., Neuhaus, P., 2009. Chemokine
receptor 5Delta32 mutation reduces the risk of acute rejection in liver transplantation.
Ann Transplant. 14 (3), 36-44.
25. Herpers, B.L., Endeman, H., de Jong, B.A., de Jongh, B.M., Grutters, J.C., Biesma,
D.H., van Velzen-Blad, H., 2009. Acute-phase responsiveness of mannose-binding
lectin in community-acquired pneumonia is highly dependent upon MBL2 genotypes.
Clin Exp Immunol. 156 (3), 488-494.
26. Hildebrandt, M.A., Komaki, R., Liao, Z., Gu, J., Chang, J.Y., Ye, Y., Lu, C., Stewart,
D.J., Minna, J.D., Roth, J.A., Lippman, S.M., Cox, J.D., Hong, W.K., Spitz, M.R.,
Wu, X., 2010. Genetic variants in inflammation-related genes are associated with
radiation-induced toxicity following treatment for non-small cell lung cancer. PLoS
One. 5 (8):e12402. doi: 10.1371/journal.pone.0012402.
27. Kang, X., Kim, H.J., Ramirez, M., Salameh, S., Ma, X., 2010. The septic shock-

associated IL-10 -1082 A > G polymorphism mediates allele-specific transcription via
poly(ADP-Ribose) polymerase 1 in macrophages engulfing apoptotic cells. J
Immunol. 184 (7), 3718-3724.
28. Lingappa, J.R., Dumitrescu, L., Zimmer, S.M., Lynfield, R., McNicholl, J.M.,
Messonnier, N.E., Whitney, C.G., Crawford, D.C., 2011. Identifying host genetic risk
factors in the context of public health surveillance for invasive pneumococcal disease.
PLoS One. 6 (8):e23413. doi: 10.1371/journal.pone.0023413.
29. Ma, P., Zhu, Y., Qiu, H., Liu, J., Wang, Y., Zeng, L., 2011. Endothelial nitric oxide
synthase 894G→T but not -786T→C gene polymorphism is associated with organ
dysfunction and increased mortality in patients with severe sepsis. J Trauma. 71 (4),
872-877.
30. Markova, S., Nakamura, T., Sakaeda, T., Makimoto, H., Uchiyama, H., Okamura, N.,
Okumura, K., 2006. Genotype-dependent down-regulation of gene expression and
function of MDR1 in human peripheral blood mononuclear cells under acute
inflammation. Drug Metab Pharmacokinet. 21 (3), 194-200.
31. Marshall, R.P., Webb, S., Hill, M.R., Humphries, S.E., Laurent, G.J., 2002. Genetic
polymorphisms associated with susceptibility and outcome in ARDS. Chest. 121 (3),
suppl 68S-69S. doi: 10.1378/chest.121.3_suppl.68S.
32. Martín-Loeches, I., Solé-Violán, J., Rodríguez de Castro, F., García-Laorden, M.I.,
Borderías, L., Blanquer, J., Rajas, O., Briones, M.L., Aspa, J., Herrera-Ramos, E.,
Marcos-Ramos, J.A., Sologuren, I., González-Quevedo, N., Ferrer-Agüero, J.M.,
Noda, J., Rodríguez-Gallego, C., 2012. Variants at the promoter of the interleukin-6
gene are associated with severity and outcome of pneumococcal community-acquired
pneumonia. Intensive Care Med. 38 (2), 256-262.

33. Meletiadis, J., Chanock, S., Walsh, T.J., 2006. Human pharmacogenomic variations
and their implications for antifungal efficacy. Clin Microbiol Rev. 19(4), 763–787.
34. Morimoto, S., Okaishi, K., Onishi, M., Katsuya, T., Yang, J., Okuro, M., Sakurai, S.,
Onishi, T., Ogihara, T., 2002. Deletion allele of the angiotensin-converting enzyme
gene as a risk factor for pneumonia in elderly patients. Am J Med. 112 (2), 89-94.
35. Moroz, V.V., Smelaia, T.V., Salnikova, L.E., Golubev, A.M., Rubanovich, A.V.,
2011. Genetic study of predisposition to community-acquired pneumonia. Vestn Ross
Akad Med Nauk. 11, 12-16.
36. Mukamal, K.J., Pai, J.K., O'Meara, E.S., Tracy, R.P., Psaty, B.M., Kuller, L.H.,
Newman, A.B., Yende, S., Curhan, G.C., Siscovick, D.S., Rimm, E.B., 2010. CRP
gene variation and risk of community-acquired pneumonia. Respirology. 15(1), 160-
164.
37. Muntinghe, F.L., Verduijn, M., Zuurman, M.W., Grootendorst, D.C., Carrero, J.J.,
Qureshi, A.R., Luttropp, K., Nordfors, L., Lindholm, B., Brandenburg, V., Schalling,
M., Stenvinkel, P., Boeschoten, E.W., Krediet, R.T., Navis, G., Dekker, F.W., 2009.
CCR5 deletion protects against inflammation-associated mortality in dialysis patients.
J Am Soc Nephrol. 20 (7), 1641-1649.
38. Reyes-Engel, A., Morcillo, L., Aranda, F.J., Ruiz, M., Gaitan, M.J., Mayor-Olea, A.,
Aranda, P., Ferrario, C,M., 2006. Influence of gender and genetic variability on
plasma angiotensin peptides. J Renin Angiotensin Aldosterone Syst. 7 (2), 92-97. doi:
10.3317/jraas.2006.015.
39. Rigat, B., Hubert, C., Alhenc-Gelas, F., Cambien, F., Corvol, P., Soubrier, F., 1990.
An insertion/deletion polymorphism in the angiotensin I-converting enzyme gene
accounting for half the variance of serum enzyme levels. J Clin Invest. 86 (4), 1343-

1346. doi: 10.1172/JCI114844.
40. Rosseau, S., Hocke, A., Mollenkopf, H., Schmeck, B., Suttorp, N., Kaufmann, S.H.E.,
Zerrahn, J., 2007. Comparative transcriptional profiling of the lung reveals shared and
distinct features of Streptococcus pneumoniae and influenza A virus infection.
Immunology. 120 (3), 380–391.
41. Rotunno, M., Yu, K., Lubin, J.H., Consonni, D., Pesatori, A.C., Goldstein, A.M.,
Goldin, L,R., Wacholder, S., Welch, R., Burdette, L., Chanock, S.J., Bertazzi, P.A.,
Tucker, M.A., Caporaso, N.E., Chatterjee, N., Bergen, A.W., Landi, M.T., 2009.
Phase I metabolic genes and risk of lung cancer: multiple polymorphisms and mRNA
expression. PLoS One. 4 (5):e5652. doi:10.1371/journal.pone.0005652.
42. Schaaf, B., Rupp, J., Müller-Steinhardt, M., Kruse, J., Boehmke, F., Maass, M., Zabel,
P., Dalhoff, K., 2005. The interleukin-6 -174 promoter polymorphism is associated
with extrapulmonary bacterial dissemination in Streptococcus pneumoniae infection.
Cytokine. 31 (4), 324-328.
43. Schaid, D.J., 2004. Evaluating associations of haplotypes with traits. Genet
Epidemiol. 27 (4), 348-364.
44. Schlüter, B., Raufhake, C., Erren, M., Schotte, H., Kipp, F., Rust, S., Van Aken, H.,
Assmann, G., Berendes, E., 2002. Effect of the interleukin-6 promoter polymorphism
(-174 G/C) on the incidence and outcome of sepsis. Crit Care Med. 30 (1), 32-37.
45. Sole X, Guino E, Valls J, Iniesta, R., Moreno, V., 2006. SNPStats: a web tool for the
analysis of association studies. Bioinformatics. 22 (15), 1928-1929. doi:
10.1093/bioinformatics/btl268.
46. Solé-Violán J, de Castro F, García-Laorden MI, Blanquer J, Aspa J, Borderías L,
Briones ML, Rajas O, Carrondo IM, Marcos-Ramos JA, Ferrer Agüero JM, Garcia-

Saavedra A, Fiuza MD, Caballero-Hidalgo A, Rodriguez-Gallego C. Genetic
variability in the severity and outcome of community-acquired pneumonia.// Respir
Med. 2010 Mar;104(3):440-7. Epub 2009 Nov 8.
47. Solé-Violán, J., García-Laorden, M.I., Marcos-Ramos, J.A., de Castro, F.R., Rajas,
O., Borderías, L., Briones, M.L., Herrera-Ramos, E., Blanquer, J., Aspa, J., Florido,
Y., García-Bello, M.A., Ferrer-Agüero, J.M., Sologuren, I., Rodriguez-Gallego, C.,
2011. The Fcγ receptor IIA-H/H131 genotype is associated with bacteremia in
pneumococcal community-acquired pneumonia. Crit Care Med. 39 (6), 1388-1393.
48. Storey, J.D., 2002. A direct approach to false discovery rates. J R Stat Soc B Stat
Methodol. 64, 479-498.
49. Takahashi, T., Morimoto, S., Okaishi, K., Kanda, T., Nakahashi, T., Okuro, M.,
Murai, H., Nishimura, Y., Iwai, K., Matsumoto, M., 2005. Reduction of pneumonia
risk by an angiotensin I-converting enzyme inhibitor in elderly Japanese inpatients
according to insertion/deletion polymorphism of the angiotensin I-converting enzyme
gene. Am J Hypertens. 18 (10), 1353-1359.
50. Takahashi, T., Yamaguchi, E., Furuya, K., Kawakami, Y., 2001. The ACE gene
polymorphism and cough threshold for capsaicin after cilazapril usage. Respir Med.
95 (2), 130-135.
51. Terry, C.F., Loukaci, V., Green, F.R., 2000. Cooperative influence of genetic
polymorphisms on interleukin 6 transcriptional regulation. J Biol Chem. 275 (24),
18138–18144. doi: 10.1074/jbc.M000379200.
52. Thio, C.L., Astemborski, J., Bashirova, A., Mosbruger, T., Greer, S., Witt, M.D.,
Goedert, J.J., Hilgartner, M., Majeske, A., O'Brien, S.J., Thomas, D.L., Carrington,
M., 2007. Genetic protection against hepatitis B virus conferred by CCR5Delta32:

Evidence that CCR5 contributes to viral persistence. J Virol. 81 (2), 441-445.
53. Veldman, B.A., Spiering, W., Doevendans, P.A., Vervoort, G., Kroon, A.A.,
Vervoort, G., Kroon, A.A., de Leeuw, P.W., Smits, P., 2002. The Glu298Asp
polymorphism of the NOS 3 gene as a determinant of the baseline production of nitric
oxide. J Hypertens 20 (10), 2023–2027.
54. Wang, S., Chanock, S., Tang, D., Li, Z., Jedrychowski, W., Perera, F.P., 2008.
Assessment of interactions between pah exposure and genetic polymorphisms on
PAH- DNA adducts in African American, Dominican and Caucasian mothers and
newborns Cancer Epidemiol Biomarkers Prev. 17(2), 405-413. doi: 10.1158/1055-
9965.EPI-07-0695
55. Wang, X.L., Sim, A.S., Wang, M.X., Murrell, G.A., Trudinger, B., Wang, J., 2000.
Genotype dependent and cigarette specific effects on endothelial nitric oxide synthase
gene expression and enzyme activity. FEBS Lett 471 (1), 45–50.
56. Waterer, G.W., Quasney, M.W., Cantor, R.M., Wunderink, R.G., 2001. Septic shock
and respiratory failure in community-acquired pneumonia have different TNF
polymorphism associations. Am J Respir Crit Care Med. 163(7), 1599-1604.
57. Wunderink, R.G., Waterer, G.W., Cantor, R.M., Quasney, M.W., 2002. Tumor
necrosis factor gene polymorphisms and the variable presentation and outcome of
community-acquired pneumonia. Chest. 121 (3 Suppl), 87S. doi:
10.1378/chest.121.3_suppl.87S.
58. Yende, S., Angus, D.C., Ding, J., Newman, A.B., Kellum, J.A., Li, R.., Ferrell, R.E.,
Zmuda, J., Kritchevsky, S.B., Harris, T.B., Garcia, M., Yaffe, K., Wunderink, R.G.,
for the Health ABC Study, 2007. 4G/5G plasminogen activator inhibitor-1
polymorphisms and haplotypes are associated with pneumonia. Am J Respir Crit Care

Med. 176 (11), 1129-1137. doi:10.1164/rccm.200605-644OC.
59. Yuan, F.F., Marks, K., Wong, M., Watson, S., De Leon, E., McIntyre, P.B., Sullivan,
J.S., 2008. Clinical relevance of TLR2, TLR4, CD14 and FcgammaRIIA gene
polymorphisms in Streptococcus pneumoniae infection. Immunology and Cell
Biology. 86, 268–270.
60. Zaitlen, N., Kang, H.M., Eskin, E., Halperin, E., 2007. Leveraging the HapMap
correlation structure in association studies. Am J Hum Genet. 80 (4), 683-691.
61. Zhu, X., Bouzekri, N., Southam, L., Cooper, R.S., Adeyemo, A., McKenzie C.A.,
Luke, A., Chen, G., Elston, R.C., Ward, R.., 2001. Linkage and association analysis of
angiotensin–converting enzyme (ACE)–gene polymorphisms with ACE concentration
and blood pressure. Am J Hum Genet. 68 (5), 1139–1148.
62. Zúñiga, J., Buendía-Roldán, I., Zhao, Y., Jiménez, L., Torres, D., Romo, J., Ramírez,
G., Cruz, A., Vargas-Alarcon, G., Sheu, C.C., Chen, F., Su, L., Tager, A.M., Pardo,
A., Selman, M., Christiani, D.C., 2012. Genetic variants associated with severe
pneumonia in A/H1N1 influenza infection. Eur Respir J. 39 (3), 604-610

Table 1. Charateristics of the sample and control groups involved in the study.
Feature Control A n (%) Control B n (%) CAP n (%) Total number 141 314 334 Age 21.06 ± 0.42 41.65 ± 1.03 26.93±0.82 Sex Male Female
130 (92.2) 11 (7.8)
286 (91.1) 28 (8.9)
307 (91.9) 27 (8.1)
Progression Bilateral Unilateral
- - 98 (29.34) 236 (70.66)
Background diseases: No Yesa
- - 326 (97.6) 8 (2.4)
Co-morbidity: No Yesb
- - 303(90.72) 31(9.28)
ICU admission No Yes
- - 156 (46.71) 178 (53.29)
Infectious pathogens Streptococcus pneumoniae Monoflora S. pneumoniae with other infectious pathogens Other infectious pathogens
- - 108 (32.34)c 70 (39.33) 12 (17.14) 58 (82.86) 38 (21.35)
Duration of hospitalization - - 27.40±10.16 (days) Hospital mortality - - 12 (3.59) a Ischemic cardiopathy, diabetes, obesity. b The most frequently purulent antritis (51.61%).
cData available for 108 cases only.

Table 2. Summary on primer sequencesa, amplicons, and PCR cycling conditions for
individual SNPs analyzed in the study.
rs# of SNP Primer sequences 5’-3’, external forward/reverse, internal allele-specific forward/reverse
PCRb Amplicon length (bp)
CYP1A1 rs2606345
F gggagatggatggttcctaccac 94 оC (30s) 65.5 оC (18s) 72 оC (30s) 34x
388 R cctcctaagggtggcttgtcagt F(c) cctgcagttggcaatctgtcac 155 R(a) cctttgctgggagacaatcaggt 277
CYP1A1 rs4646903
F aggaggtagcagtgaagaggtg 94 оC (27s)
63.2 оC (18s) 72 оC (30s) 34x
373 R ggcgtaagtcagcacagtgatt F(c) tcactgtaacctccacctccc 250 R(t) ggagaatcgtgtgagccca 162
CYP1A1 rs1048943
F gagctccactcacttgacacttct 94 оC (30s) 58 оC (18s) 72 оC (30s) 32×
465 R cagtgtctatgagtttcaggctgaatctt F(a) gaagtgtatcggtgagacca 285 R(g) ctcccagcgggcaac 211
GSTM1 Ins/del GSTT1 Ins/Del
F gaactccctgaaaagctaaagc 94 оC (30s) 60 оC (18s) 72 оC (30s) 32x
219 R gttgggctcaaatatacggtgg F ttccttactggtcctcacatctc 459 R tcaccggatcatggccagca
ABCB1 rs1045642
F tgttttcagctgcttgatgg 94 оC (30s) 61.5 оC (20s) 72 оC (30s) 32x
377 R aacccaaacaggaagtgtgg
F(c) gtgttgtcacaggaagagatc 236 R(t) tcctttgctgccctcaca 179
ACE rs4340 CCR5 rs333
F ctgctgcctatacagtcacttt 94 оC (20s) 56 оC (20s) 72 оC (40s) 33x
436 R gtggccatcacattcgtcagat 149 F gtggtgacaagtgtgatcactt 240 R tcgacaccgaagcagagttttta 208
IL-6 rs1800795
F tgcatgacttcagctttactctttg 94 оC (40s) 64.5 оC (18s) 72 оC (40s)
375 R ggggagatagagcttctctttcgtt F(g) ccccctagttgtgtcttgcg 241
R(c) gcaatgtgacgtcctttagcatg 32x 176 TNF-α rs1800629
F cctcaaggactcagctttctgaag 94 оC (40s) 63 оC (20s) 72 оC (40s) 33x
312 R accttctgtctcggtttcttctcc F(g) caataggttttgaggggcatgg 195 R(a) gaggctgaaccccgtcct 156
NOS3 rs1799983
F ggagatgaaggcaggagacagt 94 оC (40s) 63 оC (20s) 72 оC (40s) 33x
465 R cgatctcagtgctcatgtaccag F(g) tgcaggccccagatgag 314 R(t) cagaaggaagagttctggggga 189
MBL2 rs7096206
F gcatgccctctgtcctacaatc 94 оC (40s) 63о C (20s) 72 оC (40s) 33x
349 R ccgaagaggacatggagagaaa
F(g) gctggaagactataaacatgctttcg 152 R(c) ccatttgttctcactgccacg 243
ACE rs4340 CCR5
F ctgctgcctatacagtcacttt 94 оC (20s) 56 оC (20s) 72 оC (40s)
436 R gtggccatcacattcgtcagat 149 F gtggtgacaagtgtgatcactt 240

rs333 R tcgacaccgaagcagagttttta 33x 208 MTHFR rs1801133
F gccactcactgttttagttcaggctgtg 94 оC (40s) 70.2 оC (18s) 72 оC (40s) 30x
390 R tggagggagcttatgggctctcc F(t) gaaggagaaggtgtctgcgggagt 174 R(c) aagctgcgtgatgatgaaatcgg 262
a The Primer3 algorithm was used to design external primer sequences.
b PCR conditions are provided as temperature (duration) of denaturation, annealing and
extension phase, respectively, followed by the number of cycles for a given SNP. Each PCR
reaction was finished by a final extension phase lasting 5 min.

Table 3. The distribution of genotypes among CAP and two control groupsa.
Genes and genotypes
Community-acquired pneumonia
Control subjects (main control group – control A)
P-valueb for dominant (dom) or recessive (rec)
modelc, OR, 95% CI (Case-control A)
Healthy control subjects (population control group –
control B)
P-valueb for dominant (dom) or recessive (rec)
modeld, OR, 95% CI (Case-control B)
Number (%) HWP Number (%) HWP Number (%) HWP
Xenobiotics detoxification
n=334 n=134 n=311
CYP1A1 rs2606345
G/G 37 (11.08) P=0.002
25 (18.66) P=0.677
P=3.9x10-5 (rec)e OR=2.40 1.59-3.64
38 (12.22) P=0.311
P=1.4x10-5 (rec)f OR=2.00 1.46-2.74
T/G 111 (33.23) 63 (47.01) 153 (49.20) T/T 186 (55.69) 46 (34.33) 120 (38.59)
n=323 n=131 n=280
CYP1A1 rs4646903
T/T 265 (82.04) P=0.256
98 (74.81)
P=0.099
P=0.093 OR=1.54 0.95-2.50
218 (77.86) P=0.128
P=0.220 OR=1.30 0.87-1.94 T/C 57 (17.65) 33 (25.19) 61 (21.79)
C/C 1 (0.31) 0 (0.00) 1 (0.36) n=323 n=132 n=281
CYP1A1 rs1048943 Ile462Val
A/A 291 (90.09)
P=0.349
124 (93.94)
P=0.720
P=0.207 OR=1.70 0.77-3.79
256 (91.10) P=0.435
P=0.780 OR=1.13 0.65-1.95 A/G 32 (9.91) 8 (6.06) 25 (8.90)
G/G 0 (0.00) 0 (0.00) 0 (0.0) n=331 n=140 n=307 GSTM1 Del(D/D)-Ins (I/*)
D/D 134 (40.48) -
78 (55.71) -
P=0.0032g OR=1.85 1.24-2.75
140 (45.60) -
P=0.20 OR=0.81 0.59-1.11 I/ 197 (59.52) 62 (44.29) 167 (54.40)
n=331 n=140 n=307
GSTT1 Del(D/D)-Ins (I/*)
D/D 63 (19.03) -
22 (15.71) -
P=0.433 OR=1.26 0.74-2.14
42 (13.68) -
P=0.112 OR=1.43 0.94-2.18 I/ 268 (80.97) 118 (84.29) 265 (86.32)
n=332 n=141 n=314
ABCB1 rs1045642 Ile1145=
C/C 53 (15.96) P=0.066
31 (21.99) P=0.261
P=0.081 (dom) OR=1.46 0.97-2.20
54 (17.20) P=0.890
P=0.053 (dom) OR=1.37 1.00-1.88
T/C 139 (41.87) 63 (44.68) 151 (48.09) T/T 140 (42.17) 47 (33.33) 109 (34.71)
Immune response regulation and inflammation
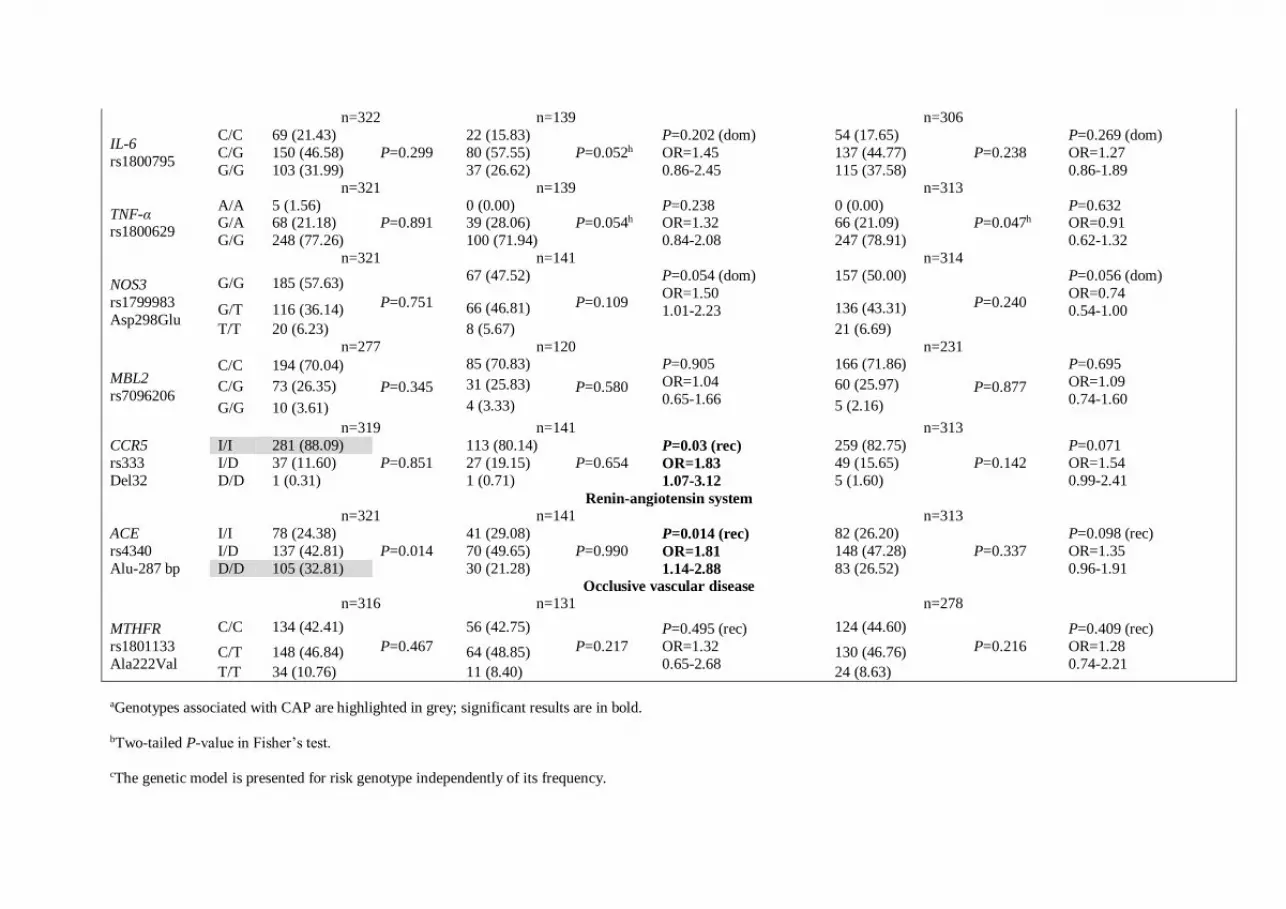
n=322 n=139 n=306
IL-6 rs1800795
C/C 69 (21.43) P=0.299
22 (15.83) P=0.052h
P=0.202 (dom) OR=1.45 0.86-2.45
54 (17.65) P=0.238
P=0.269 (dom) OR=1.27 0.86-1.89
C/G 150 (46.58) 80 (57.55) 137 (44.77) G/G 103 (31.99) 37 (26.62) 115 (37.58)
n=321 n=139 n=313
TNF-α rs1800629
A/A 5 (1.56) P=0.891
0 (0.00) P=0.054h
P=0.238 OR=1.32 0.84-2.08
0 (0.00) P=0.047h
P=0.632 OR=0.91 0.62-1.32
G/A 68 (21.18) 39 (28.06) 66 (21.09) G/G 248 (77.26) 100 (71.94) 247 (78.91)
n=321 n=141 n=314
NOS3 rs1799983 Asp298Glu
G/G 185 (57.63) P=0.751
67 (47.52)
P=0.109
P=0.054 (dom) OR=1.50 1.01-2.23
157 (50.00)
P=0.240
P=0.056 (dom) OR=0.74 0.54-1.00 G/T 116 (36.14) 66 (46.81) 136 (43.31)
T/T 20 (6.23) 8 (5.67) 21 (6.69) n=277 n=120 n=231
MBL2 rs7096206
C/C 194 (70.04)
P=0.345
85 (70.83)
P=0.580
P=0.905 OR=1.04 0.65-1.66
166 (71.86)
P=0.877
P=0.695 OR=1.09 0.74-1.60
C/G 73 (26.35) 31 (25.83) 60 (25.97)
G/G 10 (3.61) 4 (3.33) 5 (2.16)
n=319 n=141 n=313 CCR5 rs333 Del32
I/I 281 (88.09) P=0.851
113 (80.14) P=0.654
P=0.03 (rec) OR=1.83 1.07-3.12
259 (82.75) P=0.142
P=0.071 OR=1.54 0.99-2.41
I/D 37 (11.60) 27 (19.15) 49 (15.65) D/D 1 (0.31) 1 (0.71) 5 (1.60)
Renin-angiotensin system n=321 n=141 n=313 ACE rs4340 Alu-287 bp
I/I 78 (24.38) P=0.014
41 (29.08) P=0.990
P=0.014 (rec) OR=1.81 1.14-2.88
82 (26.20) P=0.337
P=0.098 (rec) OR=1.35 0.96-1.91
I/D 137 (42.81) 70 (49.65) 148 (47.28) D/D 105 (32.81) 30 (21.28) 83 (26.52)
Occlusive vascular disease n=316 n=131 n=278
MTHFR rs1801133 Ala222Val
C/C 134 (42.41) P=0.467
56 (42.75) P=0.217
P=0.495 (rec) OR=1.32 0.65-2.68
124 (44.60) P=0.216
P=0.409 (rec) OR=1.28 0.74-2.21
C/T 148 (46.84) 64 (48.85) 130 (46.76) T/T 34 (10.76) 11 (8.40) 24 (8.63)
aGenotypes associated with CAP are highlighted in grey; significant results are in bold.
bTwo-tailed P-value in Fisher’s test.
cThe genetic model is presented for risk genotype independently of its frequency.

dThe same genetic model as in ‘case-control A’.
e,fSignificant after Bonferroni correction for multiple comparisons: Pe=0.010, Pf=0.00364
gSignificant after 5%FDR correction for multiple comparisons P<0.00577
hBorderline significance for HWE in the control.

Table 4. LD statistics and CYP1A1 haplotype association with CAP
CYP1A1 (15q24.1) D’ statistics (over diagonal), P-value (under diagonal)
CAP Control A Control B rs2606345 rs4646903 rs1048943 rs2606345 rs4646903 rs1048943 rs2606345 rs4646903 rs1048943
rs2606345 0.687 0.729 rs2606345 0.999 0.997 rs2606345 0.966 0.998
rs4646903 0 0.632 rs4646903 0 0.839 rs4646903 0 0.951
rs1048943 8∙10-4 0 rs1048943 0 0 rs1048943 0 0
CYP1A1 haplotype associations rs4646903 rs1048943 rs2606345 Haplotype frequencies Case-Control A Case-Control B
Chr. position 75011641 75012985 75017176 CAP Control A Control B OR (95% CI) P value OR (95% CI) P value
N1 T A T 0.6990 0.5803 0.6295 1a --- 1 ---
N2 T A G 0.1928 0.2892 0.2511 0.58 (0.41-0.81) 0.0014 0.70 (0.54-0.92) 0.01 N3 C A G 0.0390 0.1002 0.0704 0.33 (0.18-0.62) 6∙10-4 0.52 (0.30-0.90) 0.019 rare * * * 0.0692 0.0303 0.049 1.93 (0.87-4.30) 0.11 1.26 (0.74-2.14) 0.39 aReference odds ratio (OR) was accepted as OR=1 for the major haplotype; significant results are in bold.

33
Figure 1. Age distribution of patients and controls.

34
Figure 1. Relative frequency distribution by the number of risk genotypes of the SNPs in genes
associated with CAP in case and control groups (CYP1A1 rs2606345, GSTM1, ABCB1, ACE,
CCR5, NOS3, IL-6). Mean and standard error (SE) of relative frequencies are shown.

35
Figure 3. ROC-curve for the aggregation of genetic markers CYP1A1 T606G, GSTM1, ABCB1,
ACE, CCR5, NOS3, IL-6 associated with CAP.





























