Embed Size (px)
Citation preview

Journal of Asian Earth Sciences 83 (2014) 80–90
Contents lists available at ScienceDirect
Journal of Asian Earth Sciences
journal homepage: www.elsevier .com/locate / jseaes
Hooked: Habits of the Chinese Permian gigantopterid Gigantonoclea
http://dx.doi.org/10.1016/j.jseaes.2014.01.0201367-9120/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding author. Present address: Courant Research Centre Geobiology,Georg-August-Universität Göttingen, Goldschmidtstraße 3, 37077 Göttingen,Germany. Tel.: +49 (0)551 3991296.
E-mail address: [email protected] (L.J. Seyfullah).
Leyla J. Seyfullah a,⇑, Ian J. Glasspool b, Jason Hilton a
a School of Geography, Earth and Environmental Sciences, University of Birmingham, Edgbaston, Birmingham B15 2TT, UKb Science and Education, The Field Museum of Natural History, 1400 S. Lake Shore Drive, Chicago, IL 60605, USA
a r t i c l e i n f o
Article history:Received 1 October 2013Received in revised form 12 December 2013Accepted 17 January 2014Available online 28 January 2014
Keywords:Cathaysian floraGigantonocleaGigantopteridGrowth architectureShanxi ProvinceShihhotse formation
a b s t r a c t
Based upon anatomical evidence, Permian aged gigantopterid fossils are in general reconstructed asclimbing or scrambling plants. Gigantonoclea, a genus of adpressed gigantopterid foliage from thePermian of northern China, has been reported to co-occur with hook-like organs that were interpretedas indicating a scrambling/climbing habit. We reinvestigated these hook-like structures and re-evaluatedthe nature of the co-occurrences in context with the flora preserved in each plant-bearing fossil ‘bed’ inthe North China sedimentary succession. New findings show that the species Gigantonoclea hallei proba-bly climbed using specially adapted clusters of compound grappling hook-like shoots borne on the stems.This structural arrangement comprising shoots of hooks is new to the scrambling/climbing concept ingigantopterids. However, a key figured specimen previously reported as showing intermediatehook-tipped leaf morphology on a sole pinnule tip is discounted as such and is reinterpreted as a ‘normal’pinnule partially hidden under sediment that results in an unusual appearance to this pinnule tip.Adaptations for climbing or scrambling based upon ‘hooked leaves’ observed in Gigantonoclea lagreliiare no longer supported and are reinterpreted as incompletely expanded leaves where the vernationprocess was interrupted. These data weaken prior interpretations of G. lagrelii as a climber/scramblerand raise doubts about the ubiquity of hooks amongst the gigantopterids as structures enabling themto climb or scramble their way through the Permian world.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction detail to determine their botanical affinity (Li and Yao, 1983;
Gigantopterids are an enigmatic group of plants that aregrouped by their megaphyllous leaves with distinctive reticulatevenation and their distribution in North and South America andSoutheast Asia through the Permian (Glasspool et al., 2004; DiMicheleet al., 2011). Although they are unlikely to be monophyletic(Glasspool et al., 2004; DiMichele et al., 2011), the gigantopteridsrepresent intriguing examples of convergent evolution in the plantkingdom as they possess anatomical and morphological similari-ties to modern angiosperms (flowering plants, Halle, 1927; Li andTaylor, 1998, 1999), while occurring in the Late Palaeozoic over100 Ma before these plants are thought to have first evolved (Bellet al., 2010). Gigantopterids are generally thought to have seedplant affinities (Glasspool et al., 2004; Taylor et al., 2006; DiMicheleet al., 2005, 2011), although their higher-order relationships re-main unknown: few reproductive organs have been describedand those that are known have been characterised with insufficient
Mamay, 1989; Wang, 1999; Glasspool et al., 2004; DiMicheleet al., 2011). DiMichele et al. (2005) have suggested a peltaspermous(seed plant) affinity for some American gigantopterids based onassociations of the leaves and peltaspermous reproductive organs.
Although various species and genera have been assigned to thegigantopterids (e.g. Appendix 1 of Li et al. (1994)), the gigantopte-rid concept was restricted by Glasspool et al. (2004) to includeonly the genera Gigantopteris Schenk ex. Potonié emend Glasspoolet al. and Gigantonoclea Koidzumi that are characterised by thepresence of megaphylls with continuous laminae, eucamptodr-omous venation, and higher-order veins, third order or above,that anastomose to form complex meshes while lacking suturalveins (for summary of excluded genera see DiMichele et al.,2011). However, the venation of Gigantopteris is far more complexand angiosperm-like than that of Gigantonoclea and on this basisthese two genera appear to be quite distinct within this concept.Both Gigantopteris and Gigantonoclea have been reported fromChina, Southeast Asia (Halle, 1927, 1929; Asama, 1974) and Oman(Berthelin et al., 2003), with further reports of Gigantopteris fromTurkey (Wagner, 1962) and Mexico (Carrillo Bravo, 1965) andGigantonoclea from Papua New Guinea and Thailand (Li and Shen,1996). However, Gigantopteris does not occur in North America

L.J. Seyfullah et al. / Journal of Asian Earth Sciences 83 (2014) 80–90 81
and the occurrence of Gigantonoclea reported by Mamay (1988)remains uncertain (DiMichele et al., 2011).
Our current understanding of gigantopterid growth habit is pri-marily based on evidence from anatomically preserved leaves andstems from the Late Permian of southern China that preserve evi-dence of adaptations for climbing that suggest these plants werevine- or liana-like. Aculeovinea Li et Taylor (Li and Taylor, 1998)has spines on its stem and Vasovinea Li et Taylor (Li and Taylor,1999) has spines and glandular trichomes on its stem that areinterpreted as adaptations for climbing. This interpretation ofgrowth habit in Vasovinea is further supported by its co-occurrencewith isolated compound hook structures assumed to belong to thesame plant (Li and Taylor, 1999).
Several other Palaeozoic pteridosperms have been interpretedas climbing plants, vines or lianas including PseudomariopterisDanzé-Corsin (Krings and Kerp, 2000) and Karinopteris Boersma(DiMichele et al., 1984), from which cuticular preparations showprolongations of the pinna axes with recurved tips. Cuticular prep-arations show various tendril type appendages in the pterido-sperms Dicksonites pluckenetii (Schlotheim ex Sternberg) Sterzel(Krings et al., 2003), Blanzyopteris praedentata (Gothan) Krings etKerp (Krings et al., 2003) and Lescuropteris genuina (Grand’Eury)Remy et Remy (Krings and Kerp, 1997). Unfortunately cuticularpreservation is unknown in Gigantopteris and Gigantonoclea fromnorthern China, and therefore other evidence is required to assesstheir growth habits.
Adaptations for climbing in Gigantopteris nicotianaefolia fromnorthern China have not been previously identified (Glasspoolet al., 2004), but foliage with apparently hooked tips has beeninterpreted as evidence of a climbing habit for Gigantonoclea lagre-lii (Halle, 1929). Also, a single pinnule tip assigned to Gigantonocleahallei was interpreted by Halle (1929) as having a ‘hooked’ apex,while various hook-like structures have been associated with G.hallei and have been interpreted as indicative of a climbing habit(Halle, 1927, 1929). This interpretation of Gigantonoclea fromnorthern China (Halle, 1929) led Yao (1983) to interpret the ecol-ogy of tropical southern Chinese foliage remains attributed toGigantonoclea that co-occurred with similar hook-like structuresas scrambling woody plants lacking specialised organs of attach-ment. In our view, such interpretations remain tentative.
Here we evaluate evidence for the climbing habit of Gigantonoc-lea from the Lower and Upper Shihhotse formations of northernChina, with specimens held in the Halle Collection in Sweden. Thisis one of the largest collections of compression/impression plantfossils from the Permian of Shanxi Province and includes speci-mens on which many Cathaysian species circumscriptions arebased including Gigantonoclea hallei and G. lagrelii. Halle’s historicalcollection is unique since the localities are no longer available andcomparable collections do not exist. This collection also permitsreinvestigation of the specimens Halle (1927, 1929) figured asbearing hook-like structures in association with Gigantonocleafoliage fragments to determine their structure and affinity.
2. Geological setting
The plant fossils examined are from the Permian of ShanxiProvince, China that were collected from specific ‘beds’ withinthe sedimentary succession exposed in the Eastern and WesternHills section of the Tai-Yuan-Fu (now Taiyuan) area as docu-mented by Norin (1922) and Halle (1927). These ‘beds’ are notequivalent to geological ‘beds’ but rather comprise tens ofmetres of strata that are equivalent to lithostratigraphic mem-bers within a formation (see Glasspool et al., 2004). Althoughnot from individual beds, the ‘beds’ are useful for stratigraphicsub-division and enable localised correlations between Eastern
and Western Hills sections (see Norin, 1922; Halle, 1927). Thefossils are from the Lower and Upper Shihhotse formations(Norin, 1922; Halle, 1927; Shen, 1995; Stevens et al., 2011) thatprogress from fluvial/deltaic into fluvial, alluvial and lagoonalfacies with frequent coal seams (Liu, 1990), the flora of whichis generally interpreted as representing riparian plant communi-ties (e.g. Stevens et al., 2011).
The Lower Shihhotse Formation is assigned to the upper part ofthe Lower Permian corresponding to the upper part of the Kungu-rian Stage of the Cisuralian and the lower part of the Roadian Stageof the Guadalupian. The Lower Shihhotse Formation grades up-wards into the Upper Shihhotse Formation is considered to be mid-dle to late Permian age. The age range of the Upper ShihhotseFormation is uncertain but probably from the latest Wordian orearliest Capitanian to either the end of the Capitanian (c. 5 Maduration Stevens et al., 2011), or to the middle of the Wuchiapin-gian Stage of the Lopingian (c. 10 Ma duration, Stevens et al.,2011). Stratigraphic correlation is based on regional scale lithostra-tigraphy and plant fossil biostratigraphy for which further strati-graphic resolution is unavailable (see Shen, 1995; Glasspoolet al., 2004; Hilton and Cleal, 2007; Wang, 2010; Stevens et al.,2011).
3. Materials and methods
Specimens are deposited in the Department of Palaeobotany,Swedish Museum of Natural History, (NRM) Stockholm. Each slabin each ‘bed’ was checked for any plant macro fossil remains and,where feasible, all were identified. Gigantopterid foliage and/orhook-like structures, along with any additional evidence for a climb-ing habit (e.g. sinuous and rarely branching stems) were noted. Thefive ‘beds’ sensu Norin (1922) and Halle (1927) that contain gigant-opterid foliage and also hook-like structures (Table 1) were sepa-rated and reconstructed as much as possible. As previously notedby Hilton and Li (2003) and Seyfullah and Hilton (2009), in some in-stances individual slabs from this collection contain different partsof the same fossil plant specimen and can allow the reconstructionof larger articulated fossil plant specimens by physically matchingsurfaces of each slab with other slabs and identifying adjacent slabsor parts and counter-parts of the same slabs where present. Re-assembly of the fossil plant parts allows the most complete speci-mens to be investigated rather just than basing observations andconclusions on the isolated slabs alone.
There are three species of gigantopterid sensu lato present in the Hallecollection: Gigantonoclea hallei (Asama 1959) ‘Gu and Zhi’ 1974,Gigantonoclea lagrelii (Halle) Koidzumi 1936 and Cathaysiopteris whitei(Halle) Koidzumi 1936. Specimens of Cathaysiopteris were very fragmen-tary and restricted to ‘beds’ 13, 14 and 17, and no climbing-related or-gans were found in these beds, except for a portion of undeterminablestem, thus Cathaysiopteris and ‘beds’ 13, 14 and 17 were excluded fromthis study. Foliage assignable to either of the two Gigantonoclea speciesoccurs as fragments of different sizes across seven ‘beds’ (Table 1). In‘bed’ 26, seven small fragments of G. lagrelii occur, but no other organsof interest were found besides liana-like stem fragments. A separatesmall fragment of ?G. lagrelii also occurs in ‘bed’ 33, but this speciesassignmentcouldnotbeconfirmed andnoclimbing-related organswerefound in this ‘bed’, and so this was also excluded from the study. Leafmorphology was carefully checked to exclude any damage caused byherbivory that might affect any interpretations since Glasspool et al.(2003) documented herbivory to be fairly common in Gigantonocleafrom this collection. For each remaining ‘bed’’, all plant remains wereidentified to ensure that the floral composition was known (Table 2).Some specimens had historically been prepared by dégagement to re-move overlying sediment (see Fairon-Demaret et al., 1999). No fossilswith cuticle or cuticular remains were recoverable.
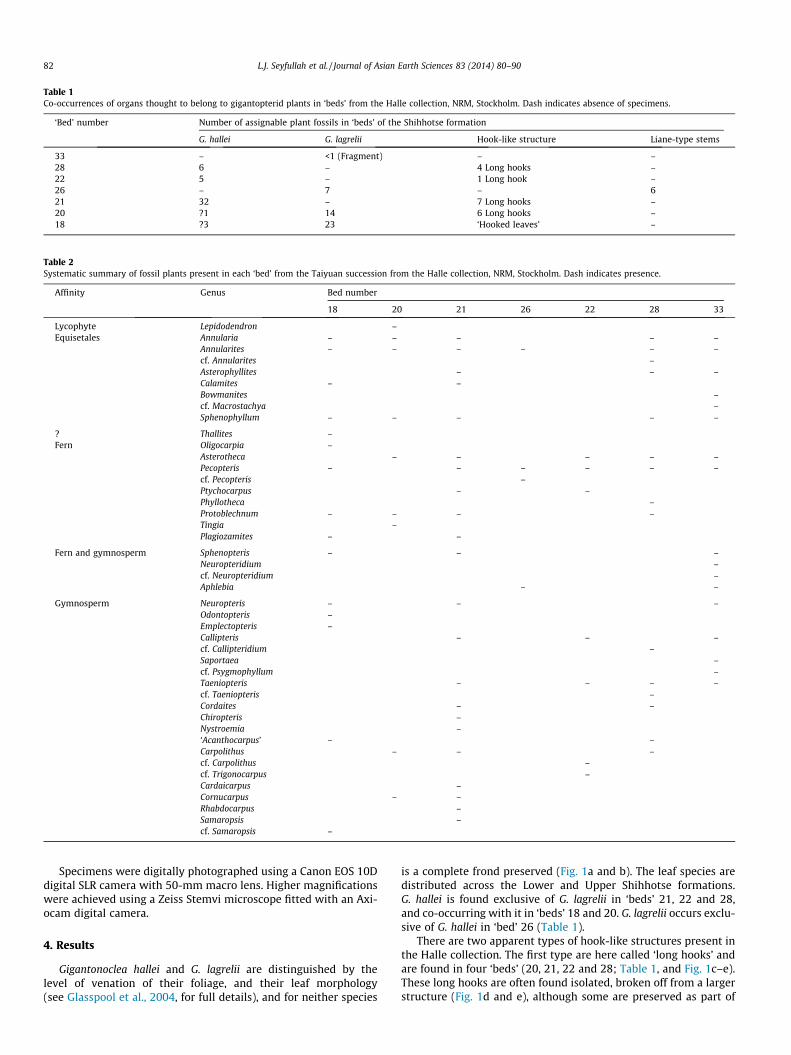
Table 1Co-occurrences of organs thought to belong to gigantopterid plants in ‘beds’ from the Halle collection, NRM, Stockholm. Dash indicates absence of specimens.
‘Bed’ number Number of assignable plant fossils in ‘beds’ of the Shihhotse formation
G. hallei G. lagrelii Hook-like structure Liane-type stems
33 – <1 (Fragment) – –28 6 – 4 Long hooks –22 5 – 1 Long hook –26 – 7 – 621 32 – 7 Long hooks –20 ?1 14 6 Long hooks –18 ?3 23 ‘Hooked leaves’ –
Table 2Systematic summary of fossil plants present in each ‘bed’ from the Taiyuan succession from the Halle collection, NRM, Stockholm. Dash indicates presence.
Affinity Genus Bed number
18 20 21 26 22 28 33
Lycophyte Lepidodendron –Equisetales Annularia – – – – –
Annularites – – – – – –cf. Annularites –Asterophyllites – – –Calamites – –Bowmanites –cf. Macrostachya –Sphenophyllum – – – – –
? Thallites –Fern Oligocarpia –
Asterotheca – – – – –Pecopteris – – – – – –cf. Pecopteris –Ptychocarpus – –Phyllotheca –Protoblechnum – – – –Tingia –Plagiozamites – –
Fern and gymnosperm Sphenopteris – – –Neuropteridium –cf. Neuropteridium –Aphlebia – –
Gymnosperm Neuropteris – – –Odontopteris –Emplectopteris –Callipteris – – –cf. Callipteridium –Saportaea –cf. Psygmophyllum –Taeniopteris – – – –cf. Taeniopteris –Cordaites – –Chiropteris –Nystroemia –‘Acanthocarpus’ – –Carpolithus – – –cf. Carpolithus –cf. Trigonocarpus –Cardaicarpus –Cornucarpus – –Rhabdocarpus –Samaropsis –cf. Samaropsis –
82 L.J. Seyfullah et al. / Journal of Asian Earth Sciences 83 (2014) 80–90
Specimens were digitally photographed using a Canon EOS 10Ddigital SLR camera with 50-mm macro lens. Higher magnificationswere achieved using a Zeiss Stemvi microscope fitted with an Axi-ocam digital camera.
4. Results
Gigantonoclea hallei and G. lagrelii are distinguished by thelevel of venation of their foliage, and their leaf morphology(see Glasspool et al., 2004, for full details), and for neither species
is a complete frond preserved (Fig. 1a and b). The leaf species aredistributed across the Lower and Upper Shihhotse formations.G. hallei is found exclusive of G. lagrelii in ‘beds’ 21, 22 and 28,and co-occurring with it in ‘beds’ 18 and 20. G. lagrelii occurs exclu-sive of G. hallei in ‘bed’ 26 (Table 1).
There are two apparent types of hook-like structures present inthe Halle collection. The first type are here called ‘long hooks’ andare found in four ‘beds’ (20, 21, 22 and 28; Table 1, and Fig. 1c–e).These long hooks are often found isolated, broken off from a largerstructure (Fig. 1d and e), although some are preserved as part of
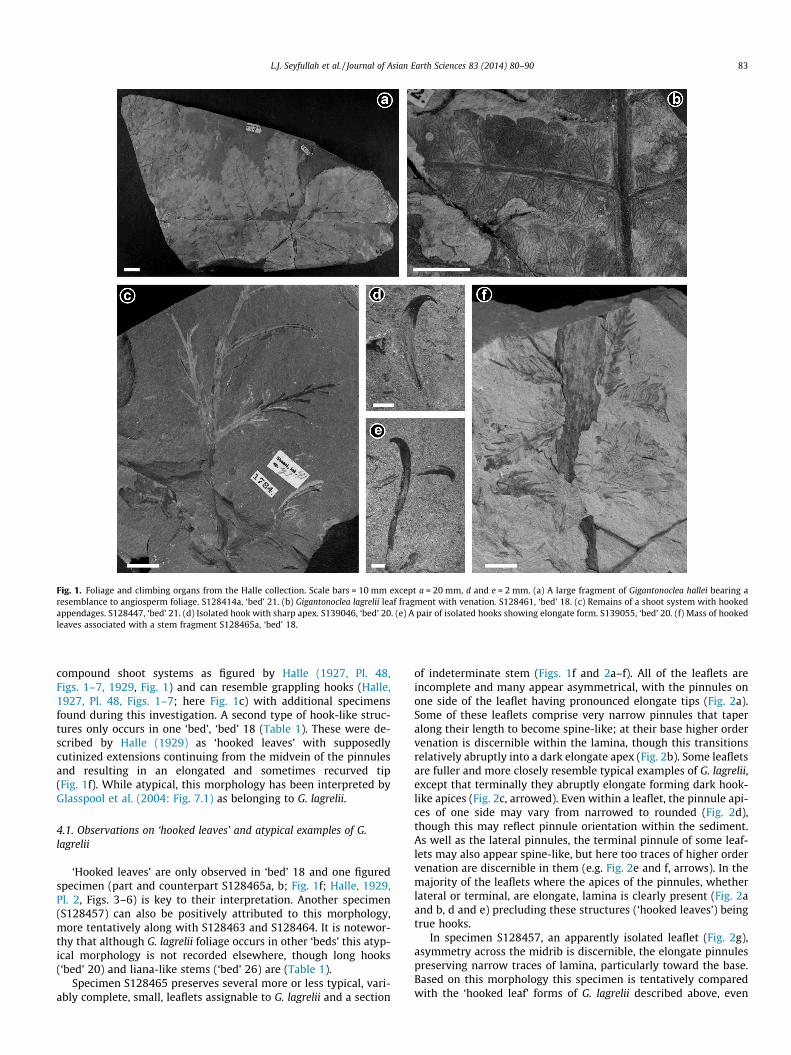
Fig. 1. Foliage and climbing organs from the Halle collection. Scale bars = 10 mm except a = 20 mm, d and e = 2 mm. (a) A large fragment of Gigantonoclea hallei bearing aresemblance to angiosperm foliage. S128414a, ‘bed’ 21. (b) Gigantonoclea lagrelii leaf fragment with venation. S128461, ‘bed’ 18. (c) Remains of a shoot system with hookedappendages. S128447, ‘bed’ 21. (d) Isolated hook with sharp apex. S139046, ‘bed’ 20. (e) A pair of isolated hooks showing elongate form. S139055, ‘bed’ 20. (f) Mass of hookedleaves associated with a stem fragment S128465a, ‘bed’ 18.
L.J. Seyfullah et al. / Journal of Asian Earth Sciences 83 (2014) 80–90 83
compound shoot systems as figured by Halle (1927, Pl. 48,Figs. 1–7, 1929, Fig. 1) and can resemble grappling hooks (Halle,1927, Pl. 48, Figs. 1–7; here Fig. 1c) with additional specimensfound during this investigation. A second type of hook-like struc-tures only occurs in one ‘bed’, ‘bed’ 18 (Table 1). These were de-scribed by Halle (1929) as ‘hooked leaves’ with supposedlycutinized extensions continuing from the midvein of the pinnulesand resulting in an elongated and sometimes recurved tip(Fig. 1f). While atypical, this morphology has been interpreted byGlasspool et al. (2004: Fig. 7.1) as belonging to G. lagrelii.
4.1. Observations on ‘hooked leaves’ and atypical examples of G.lagrelii
‘Hooked leaves’ are only observed in ‘bed’ 18 and one figuredspecimen (part and counterpart S128465a, b; Fig. 1f; Halle, 1929,Pl. 2, Figs. 3–6) is key to their interpretation. Another specimen(S128457) can also be positively attributed to this morphology,more tentatively along with S128463 and S128464. It is notewor-thy that although G. lagrelii foliage occurs in other ‘beds’ this atyp-ical morphology is not recorded elsewhere, though long hooks(‘bed’ 20) and liana-like stems (‘bed’ 26) are (Table 1).
Specimen S128465 preserves several more or less typical, vari-ably complete, small, leaflets assignable to G. lagrelii and a section
of indeterminate stem (Figs. 1f and 2a–f). All of the leaflets areincomplete and many appear asymmetrical, with the pinnules onone side of the leaflet having pronounced elongate tips (Fig. 2a).Some of these leaflets comprise very narrow pinnules that taperalong their length to become spine-like; at their base higher ordervenation is discernible within the lamina, though this transitionsrelatively abruptly into a dark elongate apex (Fig. 2b). Some leafletsare fuller and more closely resemble typical examples of G. lagrelii,except that terminally they abruptly elongate forming dark hook-like apices (Fig. 2c, arrowed). Even within a leaflet, the pinnule api-ces of one side may vary from narrowed to rounded (Fig. 2d),though this may reflect pinnule orientation within the sediment.As well as the lateral pinnules, the terminal pinnule of some leaf-lets may also appear spine-like, but here too traces of higher ordervenation are discernible in them (e.g. Fig. 2e and f, arrows). In themajority of the leaflets where the apices of the pinnules, whetherlateral or terminal, are elongate, lamina is clearly present (Fig. 2aand b, d and e) precluding these structures (‘hooked leaves’) beingtrue hooks.
In specimen S128457, an apparently isolated leaflet (Fig. 2g),asymmetry across the midrib is discernible, the elongate pinnulespreserving narrow traces of lamina, particularly toward the base.Based on this morphology this specimen is tentatively comparedwith the ‘hooked leaf’ forms of G. lagrelii described above, even
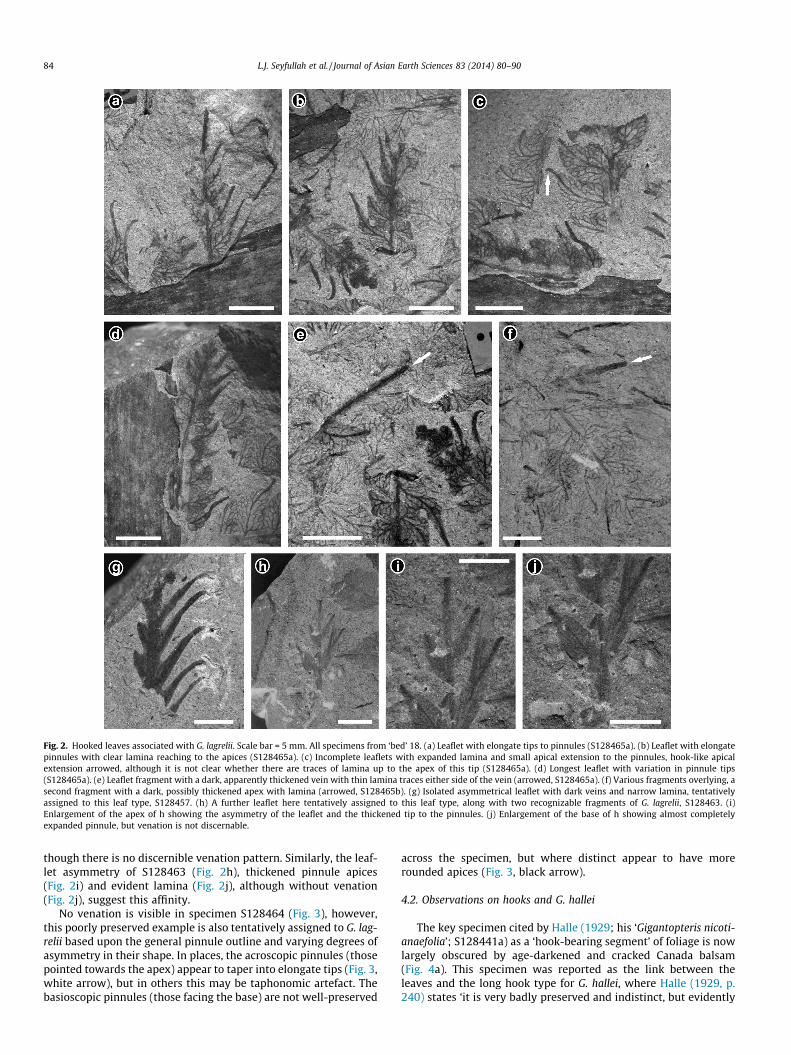
Fig. 2. Hooked leaves associated with G. lagrelii. Scale bar = 5 mm. All specimens from ‘bed’ 18. (a) Leaflet with elongate tips to pinnules (S128465a). (b) Leaflet with elongatepinnules with clear lamina reaching to the apices (S128465a). (c) Incomplete leaflets with expanded lamina and small apical extension to the pinnules, hook-like apicalextension arrowed, although it is not clear whether there are traces of lamina up to the apex of this tip (S128465a). (d) Longest leaflet with variation in pinnule tips(S128465a). (e) Leaflet fragment with a dark, apparently thickened vein with thin lamina traces either side of the vein (arrowed, S128465a). (f) Various fragments overlying, asecond fragment with a dark, possibly thickened apex with lamina (arrowed, S128465b). (g) Isolated asymmetrical leaflet with dark veins and narrow lamina, tentativelyassigned to this leaf type, S128457. (h) A further leaflet here tentatively assigned to this leaf type, along with two recognizable fragments of G. lagrelii, S128463. (i)Enlargement of the apex of h showing the asymmetry of the leaflet and the thickened tip to the pinnules. (j) Enlargement of the base of h showing almost completelyexpanded pinnule, but venation is not discernable.
84 L.J. Seyfullah et al. / Journal of Asian Earth Sciences 83 (2014) 80–90
though there is no discernible venation pattern. Similarly, the leaf-let asymmetry of S128463 (Fig. 2h), thickened pinnule apices(Fig. 2i) and evident lamina (Fig. 2j), although without venation(Fig. 2j), suggest this affinity.
No venation is visible in specimen S128464 (Fig. 3), however,this poorly preserved example is also tentatively assigned to G. lag-relii based upon the general pinnule outline and varying degrees ofasymmetry in their shape. In places, the acroscopic pinnules (thosepointed towards the apex) appear to taper into elongate tips (Fig. 3,white arrow), but in others this may be taphonomic artefact. Thebasioscopic pinnules (those facing the base) are not well-preserved
across the specimen, but where distinct appear to have morerounded apices (Fig. 3, black arrow).
4.2. Observations on hooks and G. hallei
The key specimen cited by Halle (1929; his ‘Gigantopteris nicoti-anaefolia’; S128441a) as a ‘hook-bearing segment’ of foliage is nowlargely obscured by age-darkened and cracked Canada balsam(Fig. 4a). This specimen was reported as the link between theleaves and the long hook type for G. hallei, where Halle (1929, p.240) states ‘it is very badly preserved and indistinct, but evidently

Fig. 3. Leaf fragment with asymmetrical pinnule morphology, here tentativelyassigned to the ‘hooked-leaf’ type associated with G. lagrelii. S128464, ‘bed’ 18, scalebar = 5 mm. Acroscopic pinnules taper into elongate tips occasionally. Some pinnuletips are better preserved and do show an elongation (white arrow). Basioscopicpinnules appear to have more rounded apices (black arrow).
L.J. Seyfullah et al. / Journal of Asian Earth Sciences 83 (2014) 80–90 85
represents a part of a leaf of G. nicotianaefolia with lateral leaflets ofnormal shape, but rather deeply dentate or lobed. The lowermostpinna on the left, of which only a small basal fragment is preserved,is more dissected than the others, with long and narrow segments.The contour of these segments is indistinct, but the first one on thedistal side is seen to be protracted into a narrow terminal partconsisting chiefly of the thick secondary vein. This terminal partis curved at the end to form a sharp hook. . .. The specimen thusshows a transition between the two sets of impressions’. On closeinspection, the ‘hook-like tip’ of the foliage is probably an artefactof a slight covering of sediment over the left hand side of thepinnule apex, reducing it to a hook-like shape (Fig. 4a and b, arrowat b). The Canada balsam covering of the specimen (S128441a)prohibits further preparation or more detailed imaging (Fig. 4b).However, investigation of the counterpart (S128441b) clearlyshows that the tips of the foliage do not terminate in a hook, butare elongate (Fig. 4c and d) and despite being just an impressionappear almost thickened (Fig. 4c), partly due to the way thepinnules are lying in the sediment. The precise ‘segment’ orpinnule supposed to show this hooked-tip morphology is incom-plete apically in the counterpart (Fig. 4c and d black arrow), butone of the other tips is complete (Fig. 4d, white arrow 4); however,organic connection between this foliage fragment and the stem towhich foliage assignable to G. hallei is attached could not be shown(Fig. 4c and d).
Long hooks can be found in a variety of sizes and states of pres-ervation across ‘beds’ 20, 21, 22 and 28 (Fig. 5a–k). The majority ofthese specimens (Fig. 5a–i and k) were previously un-described/un-figured. These objects occur as single hooks (Fig. 5a), as clusters
(Fig. 5b and c) and rarely in groups still attached in a shoot-likestructure (Fig. 5d). The hooks are all smooth and elongate, some re-tain some organic tissue fragments (Fig. 5a–k), but these could notbe recovered. The hooks occur associated with other plant frag-ments (Fig. 5a and e) and in isolation (Fig. 5d and f). The hooktip varies in size and shape, from blunt (Fig. 5d) to very sharp(Fig. 5g and h) although at least some of this is taphonomic and re-lates to their position on top of (Fig. 5g) or partially within the sur-rounding sediment (Fig. 5i). Isolated, single hooks often appearshorter (Fig. 5g and h), as they have likely broken off from some-where above their base within the shoots of hooks. Rare specimenspreserve long hooks borne on branching shoots (Fig. 5i–k). Com-parison of the lengths of the larger more complete disarticulatedhooks (e.g. Figs. 1e and 5c; 15–17 mm) correspond well with thoseof the longer shoot-borne hooks (e.g. Fig. 5i–k, 15–21 mm). No evi-dence for abscission for either hooks or shoots of hooks was ob-served, mainly due to the fragmentary nature of the specimens.
Comparing the branching morphology of both Gigantonocleahallei (there are three averaged modal values, probably dependingon relative position on the branch: 43�, 61�, and 89�) and G. lagrelii(majority branch around 60�) and that of the hooked shoots(branching around 60� and 88�; compare Fig. 1g with Fig. 5i–k)there is a strong similarity between the branching angles and sizesof the fragments of the different organs. However, this relationshipcannot be confirmed by organic connection between the two setsof organs.
4.3. Co-occurrence of Gigantonoclea foliage with hook-like structuresand putative liana-like stems
There are five ‘beds’ where confirmable foliage of Gigantonocleaand hook-like structures co-occur (beds 18, 20, 21, 22 and 28,Table 1). Putative liana-like (woody stems rather than herbaceousvines sensu Burnham, 2009) stem remains (Fig. 6a) were rare andhighly fragmentary and only found in ‘bed’ 26, along with foliageof G. lagrelii (Table 1). The stems comprise six fragments, nonemore than 30 cm in length or 4 cm in width and are interpretedas woody, based on the longitudinal striations indicating woodycompression remains (Fig. 6a). These stems could not be positivelycorrelated with any plant present within ‘bed’ 26 (Table 2) wherethese stems occur since there are few other plant remains in this‘bed’, with just the foliar remains of plants with equisetalean(Annularites) or fern affinities (Pecopteris, Aphlebia) present. Thesestems could also not be positively correlated with any plant re-mains from any of the other ‘beds’ (see Table 2) since these stemsare only preserved as compressions without any foliage or fructif-ications attached, although evidence of axial budding was noted(Fig. 6a). However, the branching angles of these stems (average87�) correspond closely to one of the values (89�) found forbranched examples of G. hallei (Fig. 6b).
5. Interpretation and discussion
Ideally analysis of the habit of the plants documented hereshould be based on in situ assemblages in which the fossil plantsare preserved in life position such as the forests preserved undervolcanic ash documented by Wang et al. (2012). Although fromthe earliest Permian, this ash-fall preserved flora is dominated bymarattialean ferns and lacks gigantopterids. In the present paperwe have analysed fossil plants preserved in a historically importantcollection that represents the most complete and representativecollection available from the Permian of North China. We have alsoundertaken fieldwork across North China and have not been able tocollect comparable materials on which to base our results and toovercome any potential collecting biases evident from the Halle
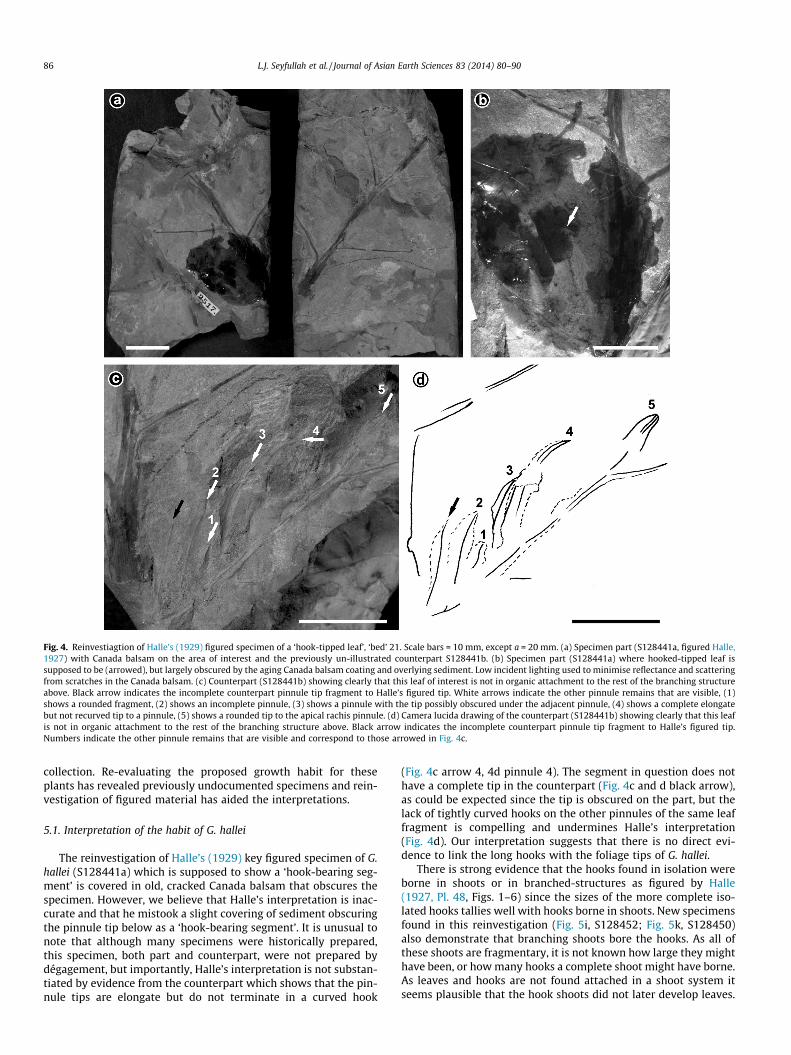
Fig. 4. Reinvestiagtion of Halle’s (1929) figured specimen of a ‘hook-tipped leaf’, ‘bed’ 21. Scale bars = 10 mm, except a = 20 mm. (a) Specimen part (S128441a, figured Halle,1927) with Canada balsam on the area of interest and the previously un-illustrated counterpart S128441b. (b) Specimen part (S128441a) where hooked-tipped leaf issupposed to be (arrowed), but largely obscured by the aging Canada balsam coating and overlying sediment. Low incident lighting used to minimise reflectance and scatteringfrom scratches in the Canada balsam. (c) Counterpart (S128441b) showing clearly that this leaf of interest is not in organic attachment to the rest of the branching structureabove. Black arrow indicates the incomplete counterpart pinnule tip fragment to Halle’s figured tip. White arrows indicate the other pinnule remains that are visible, (1)shows a rounded fragment, (2) shows an incomplete pinnule, (3) shows a pinnule with the tip possibly obscured under the adjacent pinnule, (4) shows a complete elongatebut not recurved tip to a pinnule, (5) shows a rounded tip to the apical rachis pinnule. (d) Camera lucida drawing of the counterpart (S128441b) showing clearly that this leafis not in organic attachment to the rest of the branching structure above. Black arrow indicates the incomplete counterpart pinnule tip fragment to Halle’s figured tip.Numbers indicate the other pinnule remains that are visible and correspond to those arrowed in Fig. 4c.
86 L.J. Seyfullah et al. / Journal of Asian Earth Sciences 83 (2014) 80–90
collection. Re-evaluating the proposed growth habit for theseplants has revealed previously undocumented specimens and rein-vestigation of figured material has aided the interpretations.
5.1. Interpretation of the habit of G. hallei
The reinvestigation of Halle’s (1929) key figured specimen of G.hallei (S128441a) which is supposed to show a ‘hook-bearing seg-ment’ is covered in old, cracked Canada balsam that obscures thespecimen. However, we believe that Halle’s interpretation is inac-curate and that he mistook a slight covering of sediment obscuringthe pinnule tip below as a ‘hook-bearing segment’. It is unusual tonote that although many specimens were historically prepared,this specimen, both part and counterpart, were not prepared bydégagement, but importantly, Halle’s interpretation is not substan-tiated by evidence from the counterpart which shows that the pin-nule tips are elongate but do not terminate in a curved hook
(Fig. 4c arrow 4, 4d pinnule 4). The segment in question does nothave a complete tip in the counterpart (Fig. 4c and d black arrow),as could be expected since the tip is obscured on the part, but thelack of tightly curved hooks on the other pinnules of the same leaffragment is compelling and undermines Halle’s interpretation(Fig. 4d). Our interpretation suggests that there is no direct evi-dence to link the long hooks with the foliage tips of G. hallei.
There is strong evidence that the hooks found in isolation wereborne in shoots or in branched-structures as figured by Halle(1927, Pl. 48, Figs. 1–6) since the sizes of the more complete iso-lated hooks tallies well with hooks borne in shoots. New specimensfound in this reinvestigation (Fig. 5i, S128452; Fig. 5k, S128450)also demonstrate that branching shoots bore the hooks. As all ofthese shoots are fragmentary, it is not known how large they mighthave been, or how many hooks a complete shoot might have borne.As leaves and hooks are not found attached in a shoot system itseems plausible that the hook shoots did not later develop leaves.
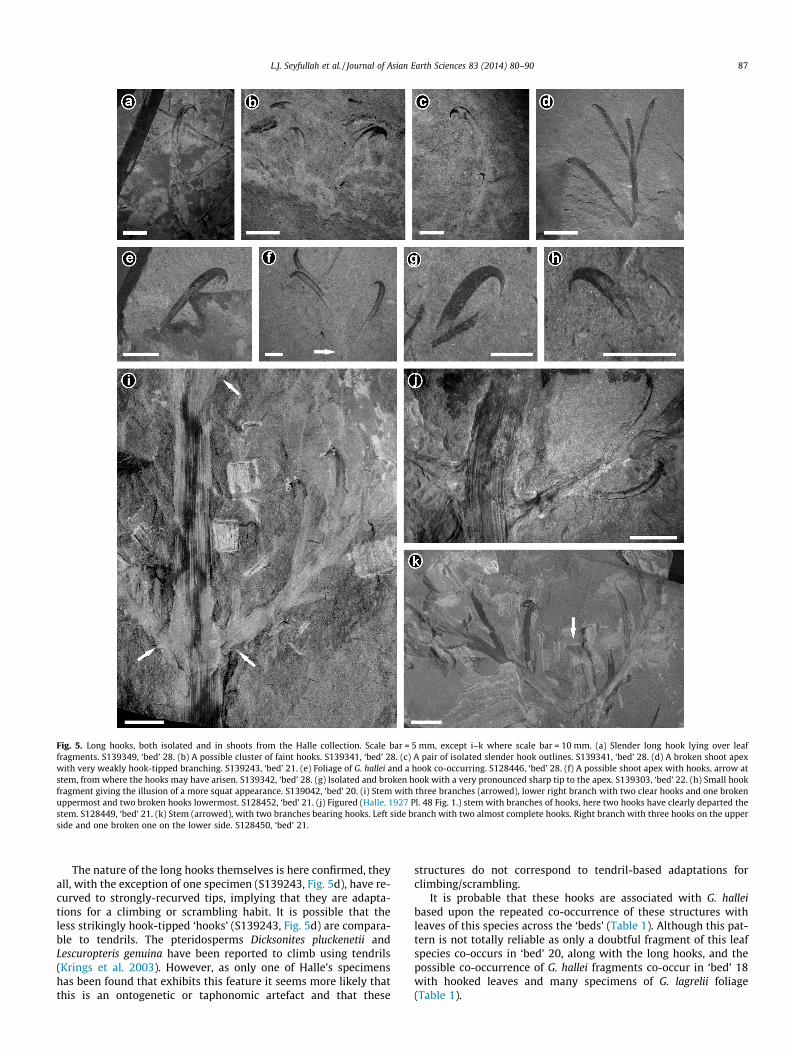
Fig. 5. Long hooks, both isolated and in shoots from the Halle collection. Scale bar = 5 mm, except i–k where scale bar = 10 mm. (a) Slender long hook lying over leaffragments. S139349, ‘bed’ 28. (b) A possible cluster of faint hooks. S139341, ‘bed’ 28. (c) A pair of isolated slender hook outlines. S139341, ‘bed’ 28. (d) A broken shoot apexwith very weakly hook-tipped branching. S139243, ‘bed’ 21. (e) Foliage of G. hallei and a hook co-occurring. S128446, ‘bed’ 28. (f) A possible shoot apex with hooks, arrow atstem, from where the hooks may have arisen. S139342, ‘bed’ 28. (g) Isolated and broken hook with a very pronounced sharp tip to the apex. S139303, ‘bed’ 22. (h) Small hookfragment giving the illusion of a more squat appearance. S139042, ‘bed’ 20. (i) Stem with three branches (arrowed), lower right branch with two clear hooks and one brokenuppermost and two broken hooks lowermost. S128452, ‘bed’ 21. (j) Figured (Halle, 1927 Pl. 48 Fig. 1.) stem with branches of hooks, here two hooks have clearly departed thestem. S128449, ‘bed’ 21. (k) Stem (arrowed), with two branches bearing hooks. Left side branch with two almost complete hooks. Right branch with three hooks on the upperside and one broken one on the lower side. S128450, ‘bed’ 21.
L.J. Seyfullah et al. / Journal of Asian Earth Sciences 83 (2014) 80–90 87
The nature of the long hooks themselves is here confirmed, theyall, with the exception of one specimen (S139243, Fig. 5d), have re-curved to strongly-recurved tips, implying that they are adapta-tions for a climbing or scrambling habit. It is possible that theless strikingly hook-tipped ‘hooks’ (S139243, Fig. 5d) are compara-ble to tendrils. The pteridosperms Dicksonites pluckenetii andLescuropteris genuina have been reported to climb using tendrils(Krings et al. 2003). However, as only one of Halle’s specimenshas been found that exhibits this feature it seems more likely thatthis is an ontogenetic or taphonomic artefact and that these
structures do not correspond to tendril-based adaptations forclimbing/scrambling.
It is probable that these hooks are associated with G. halleibased upon the repeated co-occurrence of these structures withleaves of this species across the ‘beds’ (Table 1). Although this pat-tern is not totally reliable as only a doubtful fragment of this leafspecies co-occurs in ‘bed’ 20, along with the long hooks, and thepossible co-occurrence of G. hallei fragments co-occur in ‘bed’ 18with hooked leaves and many specimens of G. lagrelii foliage(Table 1).

Fig. 6. Stems from the Halle collection. Scale bars = 10 mm. (a) Sinuous but undeterminable stem with axillary bud (arrowed). S130251, ‘bed’ 26. (b) Stem with branching G.hallei foliage. S128418, ‘bed’ 21.
88 L.J. Seyfullah et al. / Journal of Asian Earth Sciences 83 (2014) 80–90
Halle (1929, p. 239) noted ‘the great resemblance in the generalappearance and the manner of branching, the hook-bearing speci-mens exactly corresponding to a part of the vascular skeleton ofthe ordinary leaves’. This is corroborated by this investigation.Looking at the comparative branching morphology (branching an-gles and stem sizes) of both species of Gigantonoclea and thehooked shoots (compare Fig. 1g with Fig. 5i–k), there is circum-stantial evidence that the hooked shoots were associated withthe G. hallei plant. However, this cannot be confirmed by organicconnection between the two sets of organs. For the first time itcan be shown that a new structural arrangement of hooks existed,whereby they were borne in shoots, making up compound clustersof climbing aids. Overall, it can be concluded that the long hooksrelate to the branching shoots, and that these have a very stronglikelihood of relating to the G. hallei plant.
Based upon the striking morphological similarity of the com-pound hook shoots to the pinnate leaf venation pattern seen incompound examples of G. hallei it is suggested that these struc-tures represent modified leaves and not stems.
5.2. Interpretation of the ‘hooked leaves’ of G. lagrelii
While the pinnules of these leaves exhibit a thickened portionof the vein and apex, the lamina below this area is assignable tothe G. lagrelii species (Halle, 1929; Glasspool et al., 2004). Oneinterpretation of these leaves is that they represent an ‘intermedi-ate’ stage between a foliar organ and a specialised climbing organ.This could be similar to the modified leaves used as scrambling/climbing hooks recorded for some pteridosperms that show a mod-ified leaf structure, with laminae present below the hook-likeextensions e.g. Karinopteris Boesma 1972, and PseudomariopterisP. Danzé-Corsin 1953; see summary by Krings et al. (2003). Theseolder pteridosperms (Carboniferous to Early Permian) fromEuamerica exhibit a cutinised hook arising from the vein of thepinnae (Krings et al., 2003), whereas the ‘hooked leaves’ ofG. lagrelii would have to be interpreted as showing a cutinised‘hook’ arising from the vein of the pinnules.
A second possibility is that these ‘hooked leaves’ are dissociatedfragments at an early ontogenetic stage i.e. these are leaves wherethe vernation is incomplete. As the ‘hooked leaf’ structures appearwell-cutinised and therefore thick, this may seem unlikely basedupon comparison with modern plants exhibiting circinate verna-tion where, in juvenile fronds, the young developing pinnules areusually thin and become larger and thicker on maturity. Closeexamination of the ‘hooked leaf’ tips show that lamina is clearlypresent on either side of the elongate pinnule tips in the majorityof the leaflets (Fig. 2a and b, d and e). Therefore, most of these‘hooked leaves’ are not true hooks, since hooks are often describedas cutinized and recurved. Other leaflets with the appearance of avein extending past the apical margin of the pinnules (Fig. 2c and
f), do have an expanded lamina below this vein extension, unfortu-nately, it is not clear whether the lamina also extended up to thetip of this apparently extended vein (Fig. 2c, arrowed).
The inclusion of specimens S128457 (Fig. 2g), S128463 (Fig. 2h)and S128464 (Fig. 3) in this ‘hooked leaf’ group suggests an onto-genetic sequence. The former two specimens exhibit elongate pin-nule apices, as well as asymmetry across the two halves of eachleaflet and both lack venation. S128457 displays traces of narrowlamina, particularly toward the base, but this does extend up tothe ‘hooked’ tips. S128463 preserves expanded lamina below thethickened pinnule tips. Specimen S128464 preserves no venationbut could be interpreted as a nearly mature leaf of G. lagrelii basedon superficial morphological resemblance. Accepting this interpre-tation, the still expanding pinnules would appear to retain a slightapical extension (Fig. 3, white arrow), reminiscent of the less-developed ‘hooked leaves’. The pinnule tips of this specimen arenot well-preserved, but where visible they do not appear well-cuti-nized (Fig. 3). The leaflets of the pinna are asymmetrical, the acro-scopic pinnules are elongated, but the basioscopic pinnules aremore rounded (Fig. 3 black arrow), which also seems true of thedispersed juvenile hooked leaves of G. lagrelii (Fig. 2a–f). Therefore,if these leaflets do belong to G. lagrelii, then an ontogenetic se-quence could be proposed from the most immature S128465a(Fig. 2a–e) and S128457 (Fig. 2g) through S128463 (Fig. 2h) toS128464 (Fig. 3) and finally mature examples of G. lagrelii.
On balance, convincing evidence is lacking to support the firstinterpretation of these ‘hooked leaves’ as an ‘intermediate’ stagebetween a foliar organ and a specialised climbing organ. In partic-ular, in the key figured specimen lamina is apparent along the api-cal extensions of the pinnules (Fig. 2b and d). Therefore, thesestructures seem more likely to represent early ontogenetic stagesin the development of G. lagrelii, though probably not expandingthrough circinate vernation as the pinnule apices may have beenexpected to appear blunter. If these are immature examples of G.lagrelii they would not be the only examples of Gigantonoclea inthese formations exhibiting incomplete vernation; plicate exam-ples of G. hallei (Pl. 43–44, Figs. 2–4) were illustrated by Halle(1927). The interpretation of these structures as ontogenetic,rather than as hooks, leads to the conclusion that a climbing habitfor the G. lagrelii plant cannot be demonstrated.
5.3. Comparison with other gigantopterid plants
Compression material reported from the Permian of southernChina by Yao (1983) illustrates hook-like structures associatedwith the foliage of Gigantonoclea fukienensis Yabe et Oishe (Pl. 1,Figs. 3 and 8–10, Pl. 2, Figs. 7–9), and with the foliage of G. cordataYabe et Oishe (Pl. 1, Figs. 4–7). These hook structures have a similarmorphology to the hooks reported for G. hallei, but there is not en-ough detail for a full comparison.

L.J. Seyfullah et al. / Journal of Asian Earth Sciences 83 (2014) 80–90 89
The foliage most closely resembling G. hallei is the anatomicallypreserved Gigantonoclea guizhouensis Gu et Zhu from the UpperPermian of South China. Morphologically G. guizhouensis is verysimilar to G. hallei, but the venation differs as the latter has shortermeshes and wider accessory meshes. A further difference is thatspines are reported for G. guizhouensis on the abaxial side of themidrib (Li et al., 1994), whereas there was no evidence for this ineither Gigantonoclea species from the Halle collection. Li and Taylor(1998) associated the spinose foliage of G. guizhouensis with theanatomically preserved spinose stem, Aculeovinea yunguiensis Liet Taylor from the Upper Permian of South China. This suggests ascrambling habit for this contemporaneous Gigantonoclea plant.
A second anatomically preserved stem assigned to the gigan-topterids that also has spines and vessels is Vasovinea tianii Li etTaylor, though its foliage type is unknown. This taxon is similarto Aculeovinea but is smaller, with smaller spines and glandulartrichomes. It co-occurs with Gigantopteris-type leaves (Li andTaylor, 1999), compound hooks and isolated permineralised hooks.A separate branching structure of hooks and an isolated hook arealso illustrated from compression material at the same localitywhich correspond to the dimensions recorded for the perminera-lised hooks (Li and Taylor, 1999, Figs. 21 and 22) with a modifiedleaf origin also proposed for these structures based on their vascu-lar anatomy, gross morphology and branching pattern. Thesehooks are similar in size and shape to those described here in asso-ciation with G. hallei.
6. Conclusions
(1) This reinvestigation has shown that G. hallei most probablybore shoots of hooks, as originally suggested by Halle(1927), based on repeated patterns of co-occurrence andthe circumstantial evidence between the branching anglesand stem sizes of the hooked shoots and G. hallei foliage frag-ments; however this could not be confirmed by organic con-nection between the two sets of organs. A leaf origin of thesehooks based on their strikingly similar morphologicalresemblance to the pinnate leaf venation seen in G. halleileaves is postulated, and a scrambling/climbing habit couldtherefore be supported.
(2) ‘Hooked leaves’ of G. lagrelii are shown to have much morelamina present than previously described and precludestheir identification as pinnules terminating in hooks. Thisappearance is postulated to be ontogenetic, the leafletsbeing immature and incompletely opened.
(3) Gigantopterids are a heterogenous group of either para- orpolyphyletic plants that remain poorly defined with manykey details, such as the organic attachment of their supposedreproductive structures (DiMichele et al., 2011; Glasspoolet al., 2004; Mamay, 1988), unknown. Slowly a picture isemerging of this distinctive group, but more careful studiesand more complete specimens, and those with anatomicallypreservation, are needed in order to further clarify ourunderstanding of their structure and affinity.
Acknowledgements
This project was possible through Synthesys funding SE-TAF-3034 ‘‘Reconstructing the habit of the Gigantopteris plant’’ awardedto LJS. We thank the staff at the Swedish Museum of Natural His-tory (Naturhistoriska Riksmuseet, NRM), palaeobotany section,particularly Dr. Stephen McLouglin and Professor Else-Marie Friisfor help through the course of this research, and Professor Shi-
Jun Wang for assistance accessing Chinese literature. We alsothank two reviewers who helped improve this manuscript.
References
Asama, K., 1974. Permian plants from Takakurayama, Japan. Bull. Natl. Sci. Mus.Tokyo 17, 239–250.
Bell, C.D., Soltis, D.E., Soltis, P.S., 2010. The age and the diversification of theangiosperms re-visited. Am. J. Bot. 97, 1296–1130.
Berthelin, M., Broutin, J., Kerp, H., Crasquin-Soleau, S., Platel, J.P., Roger, J., 2003. TheOman Gharif mixed paleoflora: a useful tool for testing Permian Pangeareconstructions. Palaeogeogr. Palaeoclimatol. Palaeoecol. 196, 85–98.
Burnham, R.J., 2009. An overview of the fossil record of climbers: bejucos, sogas,trepadoras, lianas, cipós, and vines. Rev. Bras. Paleontol. 12, 149–160.
Carrillo Bravo, J., 1965. Estudio geologico de una parte del anticlinorio deHuayacocotla. Bull. Mex. Assoc. Petrol. Geol. 17, 73–96.
DiMichele, W.A., Rischbieter, M.O., Eggert, D.L., Gastaldo, R.A., 1984. Stem and leafcuticle of Karinopteris – source of cuticles from the Indiana Paper Coal. Am. J.Bot. 71, 626–637.
DiMichele, W.A., Kerp, H., Krings, M., Chaney, D.S., 2005. The Permian peltaspermradiation: evidence from the southwestern United States. In: Lucas, S.G. (Ed.),The Nonmarine Permian. New Mexico Museum of Natural History and ScienceBulletin, vol. 30. pp. 67–79.
DiMichele, W.A., Looy, C.V., Chaney, D.S., 2011. A new genus of gigantopterid fromthe Middle Permian of the United States and China and its relevance to thegigantopterid concept. Int. J. Plant Sci. 172, 107–119.
Fairon-Demaret, M., Hilton, J., Berry, C.M., 1999. Surface preparation of macrofossils(dégagement). In: Jones, T.P., Rowe, N.P. (Eds.), Fossil Plants and Spores. TheGeological Society of London, pp. 33–35.
Glasspool, I.J., Hilton, J., Collinson, M.E., Wang, S.J., 2003. Foliar herbivory inLate Palaeozoic Cathaysian gigantopterids. Rev. Palaeobot. Palynol. 127,125–13.
Glasspool, I.J., Hilton, J., Collinson, M.E., Wang, S.J., 2004. Defining the gigantopteridconcept: a reinvestigation of Gigantopteris (Megalopteris) nicotianaefolia Schenckand its taxonomic implications. Palaeontology 47, 1339–1361.
Halle, T.G., 1927. Palaeozoic plants from central Shansi. Palaeontol. Sin. Ser. A(Geological Survey China, Peking) 2, 1–316.
Halle, T.G., 1929. On the habit of Gigantopteris. Geol. Foren. I Stock. För. 51, 236–242, Pls. 2–3.
Hilton, J., Cleal, C.J., 2007. The relationship between Euramerican and Cathaysiantropical floras in the Late Palaeozoic: palaeobiogeographical andpalaeogeographical implications. Earth Sci. Rev. 85, 85–116.
Hilton, J., Li, C.S., 2003. Reinvestigation of Nystroemia pectinformis Halle, anenigmatic seed plant from the Late Permian of China. Palaeontology 46, 29–51.
Krings, M., Kerp, H., 1997. Cuticles of Lescuropteris genuina from the Stephanian(Upper Carboniferous) of Central France – evidence for a climbing growth habit.Bot. J. Linn. Soc. 123, 73–89.
Krings, M., Kerp, H., 2000. A contribution to the knowledge of the pteridospermgenera Pseudomariopteris Danzé-Corsin nov. emend. and Helenopteris nov. gen.Rev. Palaeobot. Palynol. 111, 145–196.
Krings, M., Kerp, H., Taylor, T.N., Taylor, E.L., 2003. How Paleozoic vines and lianasgot off the ground: on scrambling and climbing Late Carboniferous-EarlyPermian pteridosperms. Bot. Rev. 69, 204–224.
Li, H.Q., Taylor, D.W., 1998. Aculeovinea yunguiensis gen. et sp. nov.(Gigantopteridales), a new taxon of gigantopterid stem from the UpperPermian of Guizhou Province, China. Int. J. Plant Sci. 159, 1023–1033.
Li, H.Q., Taylor, D.W., 1999. Vessel-bearing stems of Vasovinea tianii gen. et sp. nov.(Gigantopteridales) from the Upper Permian of China. Am. J. Bot. 86, 1563–1575.
Li, H.Q., Tian, B.L., Taylor, E.L., Taylor, T.N., 1994. Foliar anatomy of Gigantonocleaguizhouensis (Gigantopteridales) from the Upper Permian of Guizhou Province,China. Am. J. Bot. 81, 678–689.
Li, X.X., Shen, G.L., 1996. A brief review of the Permian macrofloras in southeast Asiaand their phytogeographical delimitation. J. SE Asian Earth Sci. 13, 161–170.
Li, X.X., Yao, Z.Q., 1983. Fructifications of gigantopterids from South China.Palaeontogr. Abt. B 185, 11–26.
Liu, G.H., 1990. Permo-Carboniferous palaeogeography and coal accumulation andtheir tectonic control in the North and South China continental plates. Int. J.Coal Geol. 16, 73–117.
Mamay, S.H., 1988. Gigantonoclea in the lower Permian of Texas. Phytologia 64,330–332.
Mamay, S.H., 1989. Evolsonia, a new genus of Gigantopteridaceae from the LowerPermian Vale Formation, north-central Texas. Am. J. Bot. 76, 1299–1311.
Norin, E., 1922. The late Palaeozoic and early mesozoic sediments of central Shansi.Bull. Geol. Surv. China 4, 3–80, Pls. 3.
Seyfullah, L.J., Hilton, J., 2009. Re-evaluation of Halle’s fertile pteridospermsfrom the Permian floras of Shanxi Province, China. Plant Syst. Evol. 279,191–218.
Shen, G.L., 1995. Permian floras. In: Li, X.X. (Ed.), Fossil Floras of China through theGeological Ages. Guangdong Science and Technology Press, Guangzhou, pp.127–223.
Stevens, L.G., Hilton, J., Bond, D.P.G., Glasspool, I.J., Jardine, P.E., 2011. Radiation andextinction patterns in Pennsylvanian-Permian floras from North China asindicators for environmental and climate change. J. Geol. Soc. London 168, 607–619.

90 L.J. Seyfullah et al. / Journal of Asian Earth Sciences 83 (2014) 80–90
Taylor, D.W., Li, H.Q., Dahl, J., Fago, F.J., Zinniker, D., Moldowan, J.M., 2006.Biogeochemical evidence for the presence of the angiosperm molecular fossiloleanane in Paleozoic and Mesozoic non-angiospermous fossils. Paleobiology32, 179–190.
Yao, Z.Q., 1983. Ecology and taphonomy of gigantopterids. Bull. Nanjing Inst. Geol.Paleontol., Acad. Sin. 6, 63–84, Pls. 1–4 (in Chinese with English summary).
Wagner, R.H., 1962. On a mixed Cathaysia and Gondwana flora from SE. Anatolia(Turkey). Quatrième Congrès International de Stratigraphie et de Géologie duCarbonifère (Heerlen 1958). Comptes Rendus 3, 745–752.
Wang, J., 2010. Late Paleozoic macrofloral assemblages from Weibei Coalfield, withreference to vegetational change through the Late Paleozoic Ice-age in the NorthChina Block. Int. J. Coal Geol. 83, 292–317.
Wang, J., Pfefferkorn, H.W., Zhang, Z., Feng, Z., 2012. Permian vegetational Pompeiifrom Inner Mongolia and its implications for landscape palaeoecology andpalaeobiogeography of China. Proc. Natl. Acad. Sci. 109, 4927–4932.
Wang, Z.Q., 1999. Gigantonoclea: an enigmatic Permian plant from North China.Palaeontology 42, 329–373.





























