Embed Size (px)
Citation preview

Quaternary Research 72 (2009) 164–173
Contents lists available at ScienceDirect
Quaternary Research
j ourna l homepage: www.e lsev ie r.com/ locate /yqres
Holocene vegetation history in Romanian Subcarpathians
Ioan Tantau a,⁎, Maurice Reille b, Jacques-Louis de Beaulieu b, Sorina Farcas c, Simon Brewer d
a Babeş-Bolyai University, Department of Geology, M. Kogălniceanu Street 1, 400084 Cluj-Napoca, Romaniab IMEP, Université Paul Cézanne (Aix-Marseille III), Europôle Méditerranéen de l'Arbois, B.P. 80, F-13545 Aix-en-Provence cedex 04, Francec Institute of Biological Researches, Republicii Street 48, 400015 Cluj-Napoca, Romaniad Institut d'Astrophysique et de Géophysique, Université de Liège, Bat. B5C, 17 Allée du Six Août, B-4000, Liège, Belgium
⁎ Corresponding author. Fax: +40 264 591 906.E-mail address: [email protected] (I. Tantau)
0033-5894/$ – see front matter © 2009 University of Wdoi:10.1016/j.yqres.2009.05.002
a b s t r a c t
a r t i c l e i n f oArticle history:Received 7 April 2008Available online 5 June 2009
Keywords:Pollen analysisRadiocarbon datingVegetation historyForest dynamicsHuman influencesHoloceneSubcarpathiansRomania
Pollen analysis from a peat core 7.0 m in length, taken from a bog near Bisoca, in a mid-altitude area of theBuzăului Subcarpathian mountains, is used to reconstruct the postglacial vegetation history of the region. Thevegetation record, which is supported by twelve 14C dates, starts at the end of the Late Glacial period. At theLate Glacial/Holocene transition, open vegetation was replaced by forest, suggesting a fast response toclimatic warming. The Holocene began with the expansion of Betula, Pinus and Ulmus, followed, after11,000 cal yr BP, by Fraxinus, Quercus, Tilia and Picea. The rapid expansion of these taxa may be due to theirexistence in the area during the Late Glacial period. At ca. 9200 cal yr BP, Corylus expanded, reaching amaximum after 7600 cal yr BP. The establishment of Carpinus occurred at ca. 7200 cal yr BP, with a maximumat ca. 5700 cal yr BP. Fagus pollen is regularly recorded after 7800 cal yr BP and became dominant at ca.2000 cal yr BP. The first indications of human activities appear around 3800 cal yr BP.
© 2009 University of Washington. Published by Elsevier Inc. All rights reserved.
Introduction
In Romania, the forest succession during the Holocene Epoch iswell known, largely due to the work of the Cluj palynological school(Pop, 1929, 1932, 1942; Pop and Ciobanu 1957; Ciobanu, 1959, 1960;Boscaiu and Lupsa, 1982; Diaconeasa and Farcas, 1995–1996, 1998).The Holocene forest dynamics was subdivided into different forestphases (Pop, 1929, 1932): Pinus (Pinus montana) in Preboreal; Pinus–Picea with few Corylus and Quercetum mixtum, including Ulmus,Quercus and Tilia (early Boreal); Picea–Corylus–Quercetum mixtum(Boreal–Atlantic); Picea–Carpinus (Subboreal) and Picea–Fagus (Sub-atlantic). However, the absence of numerical dates has meant that nochronological framework exists for this succession.
The present study is a newcontributionwithin a larger programmeof systematic reinvestigation of key sites in Romania, which aims atestablishing a numerical chronology of the vegetation history sincethe Late Glacial period. Several sites in the Romanian Carpathians(Farcas et al., 1999; Tantau, 2003, 2006; Tantau et al., 2003) and inTransylvania (Tantau et al., 2006) have already been studied as part ofthis project or other project (Rösch and Fischer, 2000; Feurdean et al.,2001, 2007a; Bodnariuc et al., 2002; Björkman et al., 2002, 2003;Feurdean, 2004, 2005; Feurdean and Bennike, 2004; Magyari et al.,2006; Feurdean and Willis, 2008). These have shown that pastvegetation dynamics are not uniform in the Romanian Carpathians, as
.
ashington. Published by Elsevier I
initially believed: differences exist in the timing of the expansion ofseveral forest taxa (Feurdean, 2004, 2005; Tantau, 2003; Tantau et al.,2006). In order to better understand this dynamic and its possiblecauses, new and well dated sequences are needed from the differentregions of the Carpathians.
The current study presents the results from the Bisoca sequence(890 m a.s.l.) from Romania. The investigated site is situated in theBuzăului Subcarpathians, in the “elbow of the Carpathians.” Threesequences published by Pop and Ciobanu (1957) from the lakes ofBisoca spanned 4.34, 2.60 and 7.20 m. In his paper describing thepeat bogs of Romania, Pop (1960) indicated a maximum depth of4.50 m (in the centre) for the peat bog situated on Lacul cuMuşchi (The Lake with Sphagnum) and 7.20 m (in the centre) forthe peat bog situated on Lacul Negru (Black Lake). The peatdeposits are formed on the Eocene sandstone (Pop and Ciobanu,1957).
The Buzăului Subcarpathians are a hilly region where the activesubcrustal movements produce frequent earthquakes. This region isrecognized as being one of the regions that are the most affected byintense geomorphologic hazards in Europe (Balteanu, 1976). It is theyoungest orogenic unit in Romania built of folded and faulted Neogenemolasse. The slopes are sandstones, sands, marls and clays and areaffected by intense mass movements and gully erosion (Constantin,1999).
The region is characterized by a temperate continental climatewith moderate winters and cool summers. The annual precipitationis N600 mm. The mean annual temperature is 9°C and the meansummer temperature is 20°C (Posea and Ielenicz, 1977).
nc. All rights reserved.

165I. Tantau et al. / Quaternary Research 72 (2009) 164–173
The study area
The Bisoca peat bogs complex (45°32′N, 26°49′E) is located inthe Buzăului Subcarpathians, at an altitude ranging between 850and 900 m, above the Bisoca locality (Figs. 1 and 2). Prior inspectionof the area led to the selection of two peat bogs for coring: Lacul cuMuschi and Lacul Negru. On the basis of the Pop (1960) description,Lacul Negru (altitude 890 m) was selected for coring andpalynological analysis. This Sphagnum peat bog occupies an area ofabout 1.5 ha and it has developed above Eocene sandstone.
From a phytogeographical point of view, the peat bogs are locatedin the Carpathian province (Subcarpathian subprovince) of theCentro-European region (Ciocârlan, 2000). The peat bogs are locatedin the beech belt but are surrounded by an “island” of Pinus sylvestris(6–7 km in diameter), which is characteristic of the area (Pop andCiobanu,1957). Some small islands of Alnus glutinosa also occur in thiszone. Abies and Picea become increasingly frequent at higher altitudes(ca. 1000 m) and towards the north and northeast, while oak grovesoccur towards the south and southeast. The local vegetation type isassigned to the “Eriophoro vaginati-Sphagnetum recurvi” and “Erio-phoro vaginati-Pinetum sylvestris“ plant communities, while thespecific vegetation stage for this altitude (900 m) is typicallycharacterized by the “Hieracio rotundati-Piceetum” and “Symphytocordati-Fagetum” plant community (Coldea et al., 1997).
The peat bog is entirely covered by Pinus sylvestris to which somedifferent trees are added: Picea excelsa, Populus tremula, Betulaverrucosa, B. pubescens, Alnus glutinosa, Prunus avium. Notableamong the herbaceous taxa are Molinia coerulea, Sphagnum sp., Poly-trichium sp., Eriophorum vaginatum, Carex canescens, C. rostrata,Drosera rotundifolia, Vaccinium myrtillus, V. vitis idaea, Menyanthes
Figure 1. Locationmap of the studied zone near Bisoca, in the elbow of the Romanian CarpathFeurdean, 2004, 2005); 2, Rodnei Mts. (Tantau, 2006); 3, Calimani Mts. (Farcas et al., 1999);Retezat Mts. (Farcas et al., 1999); 8, Semenic Mts. (Rösch and Fischer, 2000); 9, Apuseni M(Cârciumaru, 1987).
trifoliata, but oligotrophic taxa, such as Vaccinium oxycoccos, Carexpaucifolia, Andromeda polifolia, Scheuzcheria palustris are missing. Thisfloristic composition distinguishes the Bisoca peat bogs from otherpeat bogs of Eastern Romanian Carpathians (Pop, 1960).
Materials and methods
Our cores were taken in summer 1998 with a hand-operated“Russian sampler” providing cores of 8 cm in diameter. The core fromthe centre of the peat bog reached basal sediments at 7.00 m. A waterpocket was found between 5.85 and 6.30 m depth. The cores weredescribed in the field, thereafter placed in half PVC tubes, andwrapped in plastic films. Before laboratory sub-sampling, all coreswere carefully cleaned and described again.
The cores were regularly sub-sampled (1 cm3) at 5 cm intervals forpollen analysis. The sample preparation followed a standard proce-dure: acetolysis in the case of peat, and gyttja samples and flotationwith Thoulet liquid (Goeury and de Beaulieu, 1979) for clay samples.Microscope thin sections were prepared from the residue and scoredfor pollen. A minimum of 350–400 grains of tree pollen were countedfor each sub-sample, except when pollen concentration was low. Thenomenclature for vascular plants follows Flora Europaea (Tutin et al.,1964–80).
Pollen diagrams were prepared using the GpalWin software(Goeury,1997). The frequencies of pollen for each taxonwere calculatedas percentages of the total sum (AP+NAP). For ecological reasonsthe spores were excluded from the pollen sum. In the pollen diagram(Figs. 4A, B) pollen values lower than 0.5% are represented by dots.
Eleven samples of bulk peat were dated with the classical methodat the Laboratory of the Radiocarbon in Poznan (Poland). An
ians and sitesmentioned in text or references: 1, Gutai Mts. (Björkman et al., 2002, 2003;4–5, Harghita Mts. (Tantau et al., 2003; Tantau, 2006); 6, Avrig (Tantau et al., 2006); 7,ts. (Bodnariuc et al., 2002); archeological sites: 10, Cârlomăneşti; 11, Sărata Monteoru
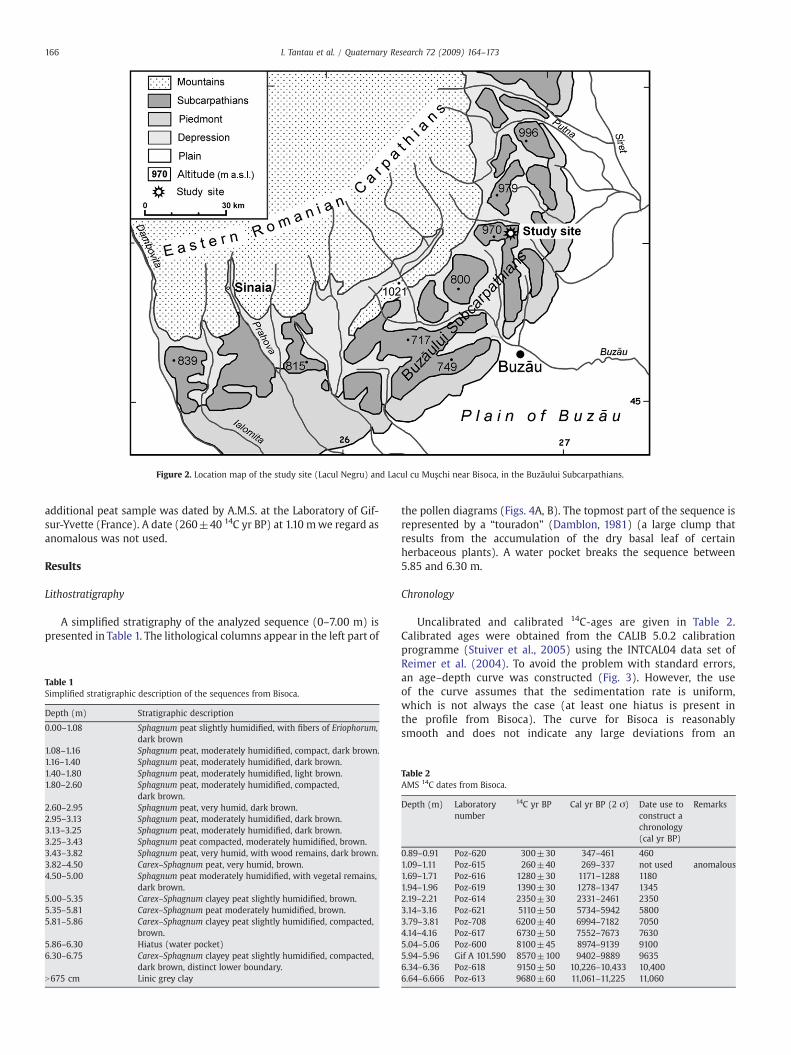
Figure 2. Location map of the study site (Lacul Negru) and Lacul cu Muşchi near Bisoca, in the Buzăului Subcarpathians.
166 I. Tantau et al. / Quaternary Research 72 (2009) 164–173
additional peat sample was dated by A.M.S. at the Laboratory of Gif-sur-Yvette (France). A date (260±40 14C yr BP) at 1.10 mwe regard asanomalous was not used.
Results
Lithostratigraphy
A simplified stratigraphy of the analyzed sequence (0–7.00 m) ispresented in Table 1. The lithological columns appear in the left part of
Table 1Simplified stratigraphic description of the sequences from Bisoca.
Depth (m) Stratigraphic description
0.00–1.08 Sphagnum peat slightly humidified, with fibers of Eriophorum,dark brown
1.08–1.16 Sphagnum peat, moderately humidified, compact, dark brown.1.16–1.40 Sphagnum peat, moderately humidified, dark brown.1.40–1.80 Sphagnum peat, moderately humidified, light brown.1.80–2.60 Sphagnum peat, moderately humidified, compacted,
dark brown.2.60–2.95 Sphagnum peat, very humid, dark brown.2.95–3.13 Sphagnum peat, moderately humidified, dark brown.3.13–3.25 Sphagnum peat, moderately humidified, dark brown.3.25–3.43 Sphagnum peat compacted, moderately humidified, brown.3.43–3.82 Sphagnum peat, very humid, with wood remains, dark brown.3.82–4.50 Carex–Sphagnum peat, very humid, brown.4.50–5.00 Sphagnum peat moderately humidified, with vegetal remains,
dark brown.5.00–5.35 Carex–Sphagnum clayey peat slightly humidified, brown.5.35–5.81 Carex–Sphagnum peat moderately humidified, brown.5.81–5.86 Carex–Sphagnum clayey peat slightly humidified, compacted,
brown.5.86–6.30 Hiatus (water pocket)6.30–6.75 Carex–Sphagnum clayey peat slightly humidified, compacted,
dark brown, distinct lower boundary.N675 cm Linic grey clay
the pollen diagrams (Figs. 4A, B). The topmost part of the sequence isrepresented by a “touradon” (Damblon, 1981) (a large clump thatresults from the accumulation of the dry basal leaf of certainherbaceous plants). A water pocket breaks the sequence between5.85 and 6.30 m.
Chronology
Uncalibrated and calibrated 14C-ages are given in Table 2.Calibrated ages were obtained from the CALIB 5.0.2 calibrationprogramme (Stuiver et al., 2005) using the INTCAL04 data set ofReimer et al. (2004). To avoid the problem with standard errors,an age–depth curve was constructed (Fig. 3). However, the useof the curve assumes that the sedimentation rate is uniform,which is not always the case (at least one hiatus is present inthe profile from Bisoca). The curve for Bisoca is reasonablysmooth and does not indicate any large deviations from an
Table 2AMS 14C dates from Bisoca.
Depth (m) Laboratorynumber
14C yr BP Cal yr BP (2 σ) Date use toconstruct achronology(cal yr BP)
Remarks
0.89–0.91 Poz-620 300±30 347–461 4601.09–1.11 Poz-615 260±40 269–337 not used anomalous1.69–1.71 Poz-616 1280±30 1171–1288 11801.94–1.96 Poz-619 1390±30 1278–1347 13452.19–2.21 Poz-614 2350±30 2331–2461 23503.14–3.16 Poz-621 5110±50 5734–5942 58003.79–3.81 Poz-708 6200±40 6994–7182 70504.14–4.16 Poz-617 6730±50 7552–7673 76305.04–5.06 Poz-600 8100±45 8974–9139 91005.94–5.96 Gif A 101.590 8570±100 9402–9889 96356.34–6.36 Poz-618 9150±50 10,226–10,433 10,4006.64–6.666 Poz-613 9680±60 11,061–11,225 11,060
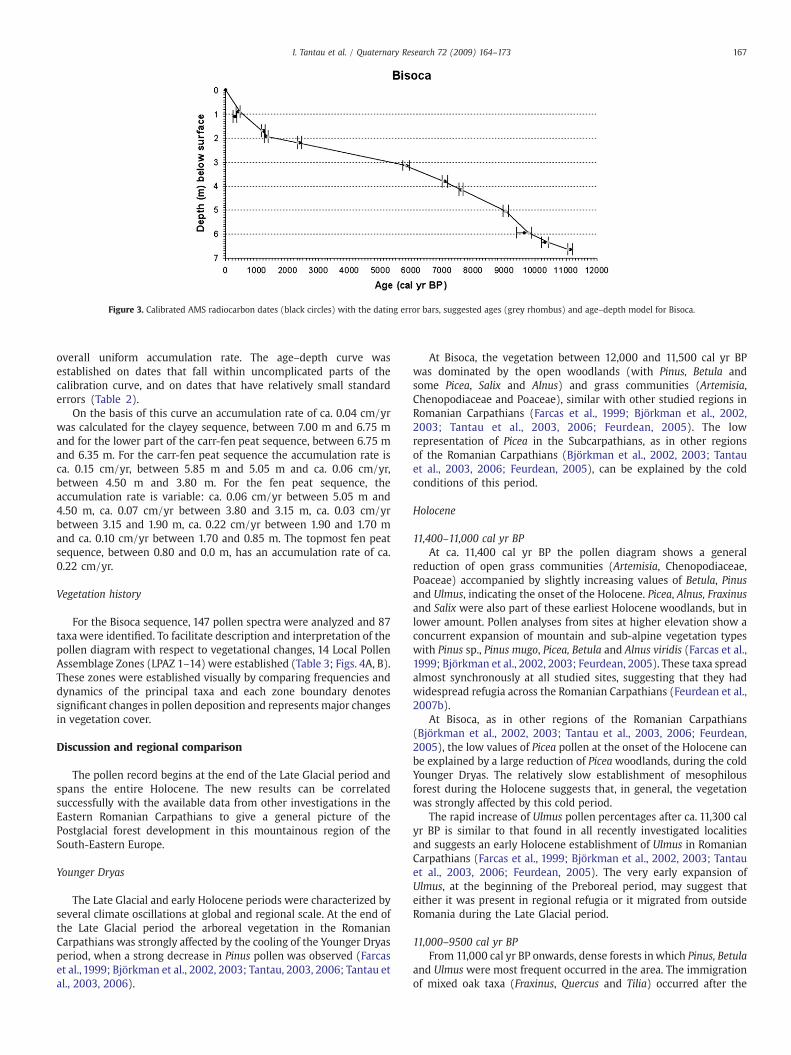
Figure 3. Calibrated AMS radiocarbon dates (black circles) with the dating error bars, suggested ages (grey rhombus) and age–depth model for Bisoca.
167I. Tantau et al. / Quaternary Research 72 (2009) 164–173
overall uniform accumulation rate. The age–depth curve wasestablished on dates that fall within uncomplicated parts of thecalibration curve, and on dates that have relatively small standarderrors (Table 2).
On the basis of this curve an accumulation rate of ca. 0.04 cm/yrwas calculated for the clayey sequence, between 7.00 m and 6.75 mand for the lower part of the carr-fen peat sequence, between 6.75 mand 6.35 m. For the carr-fen peat sequence the accumulation rate isca. 0.15 cm/yr, between 5.85 m and 5.05 m and ca. 0.06 cm/yr,between 4.50 m and 3.80 m. For the fen peat sequence, theaccumulation rate is variable: ca. 0.06 cm/yr between 5.05 m and4.50 m, ca. 0.07 cm/yr between 3.80 and 3.15 m, ca. 0.03 cm/yrbetween 3.15 and 1.90 m, ca. 0.22 cm/yr between 1.90 and 1.70 mand ca. 0.10 cm/yr between 1.70 and 0.85 m. The topmost fen peatsequence, between 0.80 and 0.0 m, has an accumulation rate of ca.0.22 cm/yr.
Vegetation history
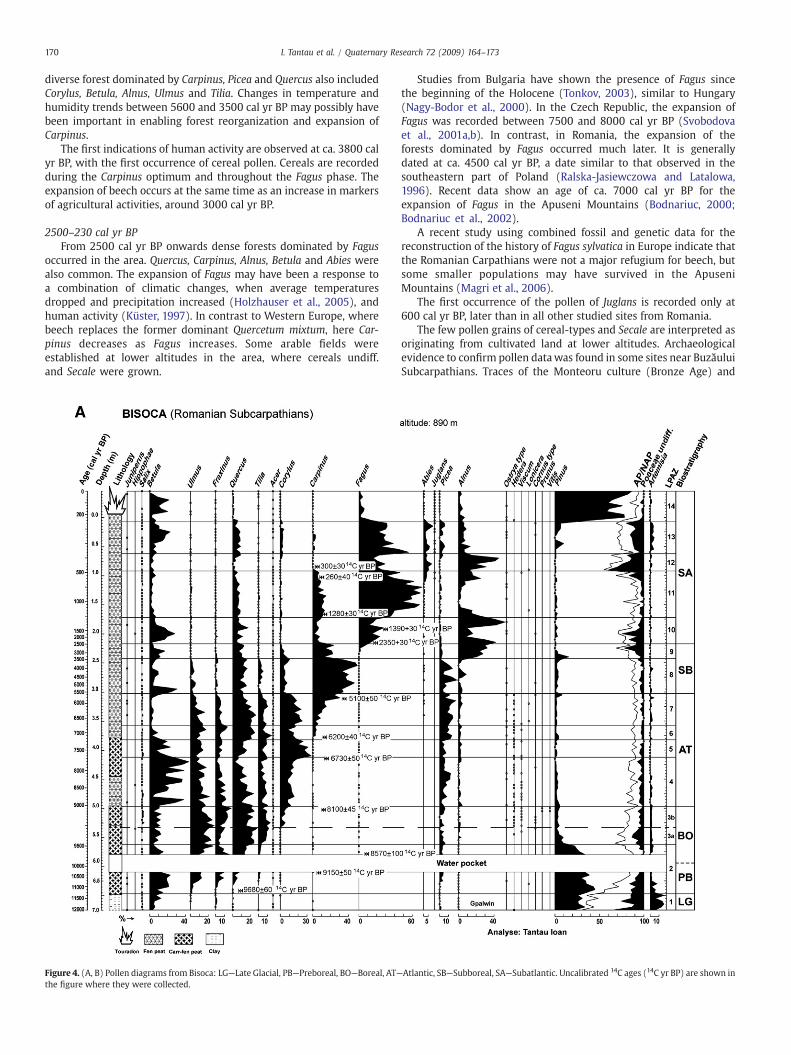
For the Bisoca sequence, 147 pollen spectra were analyzed and 87taxa were identified. To facilitate description and interpretation of thepollen diagram with respect to vegetational changes, 14 Local PollenAssemblage Zones (LPAZ 1–14) were established (Table 3; Figs. 4A, B).These zones were established visually by comparing frequencies anddynamics of the principal taxa and each zone boundary denotessignificant changes in pollen deposition and represents major changesin vegetation cover.
Discussion and regional comparison
The pollen record begins at the end of the Late Glacial period andspans the entire Holocene. The new results can be correlatedsuccessfully with the available data from other investigations in theEastern Romanian Carpathians to give a general picture of thePostglacial forest development in this mountainous region of theSouth-Eastern Europe.
Younger Dryas
The Late Glacial and early Holocene periods were characterized byseveral climate oscillations at global and regional scale. At the end ofthe Late Glacial period the arboreal vegetation in the RomanianCarpathians was strongly affected by the cooling of the Younger Dryasperiod, when a strong decrease in Pinus pollen was observed (Farcaset al., 1999; Björkman et al., 2002, 2003; Tantau, 2003, 2006; Tantau etal., 2003, 2006).
At Bisoca, the vegetation between 12,000 and 11,500 cal yr BPwas dominated by the open woodlands (with Pinus, Betula andsome Picea, Salix and Alnus) and grass communities (Artemisia,Chenopodiaceae and Poaceae), similar with other studied regions inRomanian Carpathians (Farcas et al., 1999; Björkman et al., 2002,2003; Tantau et al., 2003, 2006; Feurdean, 2005). The lowrepresentation of Picea in the Subcarpathians, as in other regionsof the Romanian Carpathians (Björkman et al., 2002, 2003; Tantauet al., 2003, 2006; Feurdean, 2005), can be explained by the coldconditions of this period.
Holocene
11,400–11,000 cal yr BPAt ca. 11,400 cal yr BP the pollen diagram shows a general
reduction of open grass communities (Artemisia, Chenopodiaceae,Poaceae) accompanied by slightly increasing values of Betula, Pinusand Ulmus, indicating the onset of the Holocene. Picea, Alnus, Fraxinusand Salix were also part of these earliest Holocene woodlands, but inlower amount. Pollen analyses from sites at higher elevation show aconcurrent expansion of mountain and sub-alpine vegetation typeswith Pinus sp., Pinus mugo, Picea, Betula and Alnus viridis (Farcas et al.,1999; Björkman et al., 2002, 2003; Feurdean, 2005). These taxa spreadalmost synchronously at all studied sites, suggesting that they hadwidespread refugia across the Romanian Carpathians (Feurdean et al.,2007b).
At Bisoca, as in other regions of the Romanian Carpathians(Björkman et al., 2002, 2003; Tantau et al., 2003, 2006; Feurdean,2005), the low values of Picea pollen at the onset of the Holocene canbe explained by a large reduction of Piceawoodlands, during the coldYounger Dryas. The relatively slow establishment of mesophilousforest during the Holocene suggests that, in general, the vegetationwas strongly affected by this cold period.
The rapid increase of Ulmus pollen percentages after ca. 11,300 calyr BP is similar to that found in all recently investigated localitiesand suggests an early Holocene establishment of Ulmus in RomanianCarpathians (Farcas et al., 1999; Björkman et al., 2002, 2003; Tantauet al., 2003, 2006; Feurdean, 2005). The very early expansion ofUlmus, at the beginning of the Preboreal period, may suggest thateither it was present in regional refugia or it migrated from outsideRomania during the Late Glacial period.
11,000–9500 cal yr BPFrom 11,000 cal yr BP onwards, dense forests inwhich Pinus, Betula
and Ulmus were most frequent occurred in the area. The immigrationof mixed oak taxa (Fraxinus, Quercus and Tilia) occurred after the
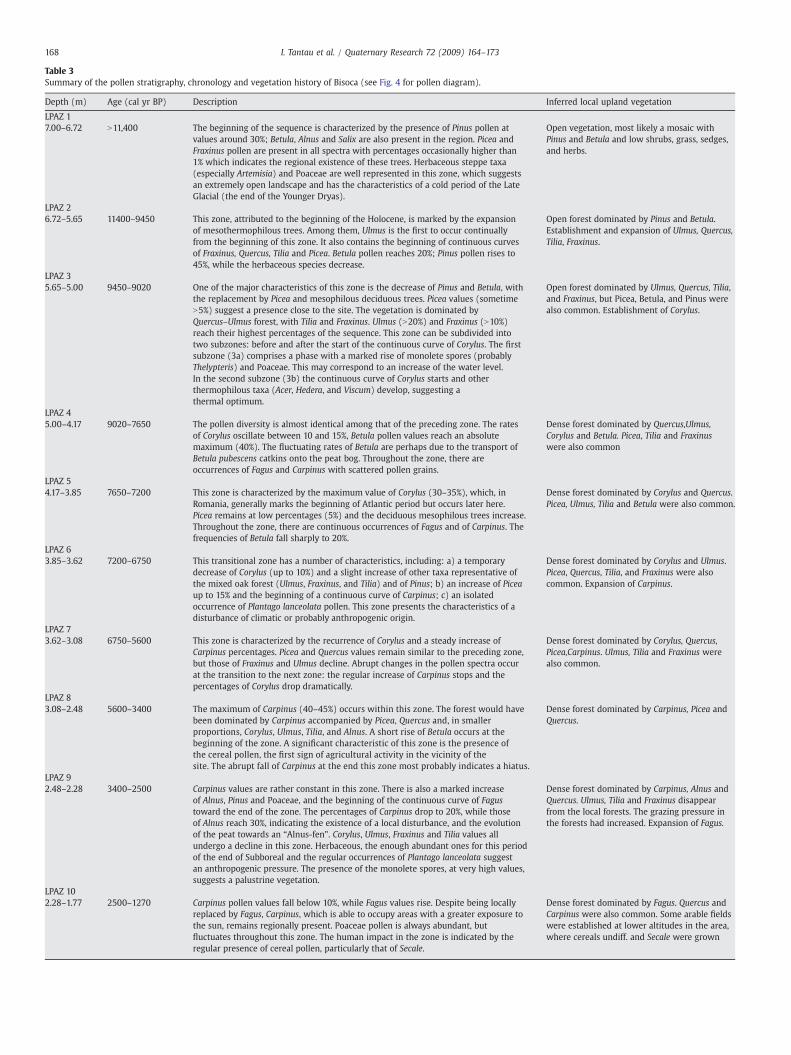
Table 3Summary of the pollen stratigraphy, chronology and vegetation history of Bisoca (see Fig. 4 for pollen diagram).
Depth (m) Age (cal yr BP) Description Inferred local upland vegetation
LPAZ 17.00–6.72 N11,400 The beginning of the sequence is characterized by the presence of Pinus pollen at
values around 30%; Betula, Alnus and Salix are also present in the region. Picea andFraxinus pollen are present in all spectra with percentages occasionally higher than1% which indicates the regional existence of these trees. Herbaceous steppe taxa(especially Artemisia) and Poaceae are well represented in this zone, which suggestsan extremely open landscape and has the characteristics of a cold period of the LateGlacial (the end of the Younger Dryas).
Open vegetation, most likely a mosaic withPinus and Betula and low shrubs, grass, sedges,and herbs.
LPAZ 26.72–5.65 11400–9450 This zone, attributed to the beginning of the Holocene, is marked by the expansion
of mesothermophilous trees. Among them, Ulmus is the first to occur continuallyfrom the beginning of this zone. It also contains the beginning of continuous curvesof Fraxinus, Quercus, Tilia and Picea. Betula pollen reaches 20%; Pinus pollen rises to45%, while the herbaceous species decrease.
Open forest dominated by Pinus and Betula.Establishment and expansion of Ulmus, Quercus,Tilia, Fraxinus.
LPAZ 35.65–5.00 9450–9020 One of the major characteristics of this zone is the decrease of Pinus and Betula, with
the replacement by Picea and mesophilous deciduous trees. Picea values (sometimeN5%) suggest a presence close to the site. The vegetation is dominated byQuercus–Ulmus forest, with Tilia and Fraxinus. Ulmus (N20%) and Fraxinus (N10%)reach their highest percentages of the sequence. This zone can be subdivided intotwo subzones: before and after the start of the continuous curve of Corylus. The firstsubzone (3a) comprises a phase with a marked rise of monolete spores (probablyThelypteris) and Poaceae. This may correspond to an increase of the water level.In the second subzone (3b) the continuous curve of Corylus starts and otherthermophilous taxa (Acer, Hedera, and Viscum) develop, suggesting athermal optimum.
Open forest dominated by Ulmus, Quercus, Tilia,and Fraxinus, but Picea, Betula, and Pinus werealso common. Establishment of Corylus.
LPAZ 45.00–4.17 9020–7650 The pollen diversity is almost identical among that of the preceding zone. The rates
of Corylus oscillate between 10 and 15%, Betula pollen values reach an absolutemaximum (40%). The fluctuating rates of Betula are perhaps due to the transport ofBetula pubescens catkins onto the peat bog. Throughout the zone, there areoccurrences of Fagus and Carpinus with scattered pollen grains.
Dense forest dominated by Quercus,Ulmus,Corylus and Betula. Picea, Tilia and Fraxinuswere also common
LPAZ 54.17–3.85 7650–7200 This zone is characterized by the maximum value of Corylus (30–35%), which, in
Romania, generally marks the beginning of Atlantic period but occurs later here.Picea remains at low percentages (5%) and the deciduous mesophilous trees increase.Throughout the zone, there are continuous occurrences of Fagus and of Carpinus. Thefrequencies of Betula fall sharply to 20%.
Dense forest dominated by Corylus and Quercus.Picea, Ulmus, Tilia and Betula were also common.
LPAZ 63.85–3.62 7200–6750 This transitional zone has a number of characteristics, including: a) a temporary
decrease of Corylus (up to 10%) and a slight increase of other taxa representative ofthe mixed oak forest (Ulmus, Fraxinus, and Tilia) and of Pinus; b) an increase of Piceaup to 15% and the beginning of a continuous curve of Carpinus; c) an isolatedoccurrence of Plantago lanceolata pollen. This zone presents the characteristics of adisturbance of climatic or probably anthropogenic origin.
Dense forest dominated by Corylus and Ulmus.Picea, Quercus, Tilia, and Fraxinus were alsocommon. Expansion of Carpinus.
LPAZ 73.62–3.08 6750–5600 This zone is characterized by the recurrence of Corylus and a steady increase of
Carpinus percentages. Picea and Quercus values remain similar to the preceding zone,but those of Fraxinus and Ulmus decline. Abrupt changes in the pollen spectra occurat the transition to the next zone: the regular increase of Carpinus stops and thepercentages of Corylus drop dramatically.
Dense forest dominated by Corylus, Quercus,Picea,Carpinus. Ulmus, Tilia and Fraxinus werealso common.
LPAZ 83.08–2.48 5600–3400 The maximum of Carpinus (40–45%) occurs within this zone. The forest would have
been dominated by Carpinus accompanied by Picea, Quercus and, in smallerproportions, Corylus, Ulmus, Tilia, and Alnus. A short rise of Betula occurs at thebeginning of the zone. A significant characteristic of this zone is the presence ofthe cereal pollen, the first sign of agricultural activity in the vicinity of thesite. The abrupt fall of Carpinus at the end this zone most probably indicates a hiatus.
Dense forest dominated by Carpinus, Picea andQuercus.
LPAZ 92.48–2.28 3400–2500 Carpinus values are rather constant in this zone. There is also a marked increase
of Alnus, Pinus and Poaceae, and the beginning of the continuous curve of Fagustoward the end of the zone. The percentages of Carpinus drop to 20%, while thoseof Alnus reach 30%, indicating the existence of a local disturbance, and the evolutionof the peat towards an “Alnus-fen”. Corylus, Ulmus, Fraxinus and Tilia values allundergo a decline in this zone. Herbaceous, the enough abundant ones for this periodof the end of Subboreal and the regular occurrences of Plantago lanceolata suggestan anthropogenic pressure. The presence of the monolete spores, at very high values,suggests a palustrine vegetation.
Dense forest dominated by Carpinus, Alnus andQuercus. Ulmus, Tilia and Fraxinus disappearfrom the local forests. The grazing pressure inthe forests had increased. Expansion of Fagus.
LPAZ 102.28–1.77 2500–1270 Carpinus pollen values fall below 10%, while Fagus values rise. Despite being locally
replaced by Fagus, Carpinus, which is able to occupy areas with a greater exposure tothe sun, remains regionally present. Poaceae pollen is always abundant, butfluctuates throughout this zone. The human impact in the zone is indicated by theregular presence of cereal pollen, particularly that of Secale.
Dense forest dominated by Fagus. Quercus andCarpinus were also common. Some arable fieldswere established at lower altitudes in the area,where cereals undiff. and Secale were grown
168 I. Tantau et al. / Quaternary Research 72 (2009) 164–173
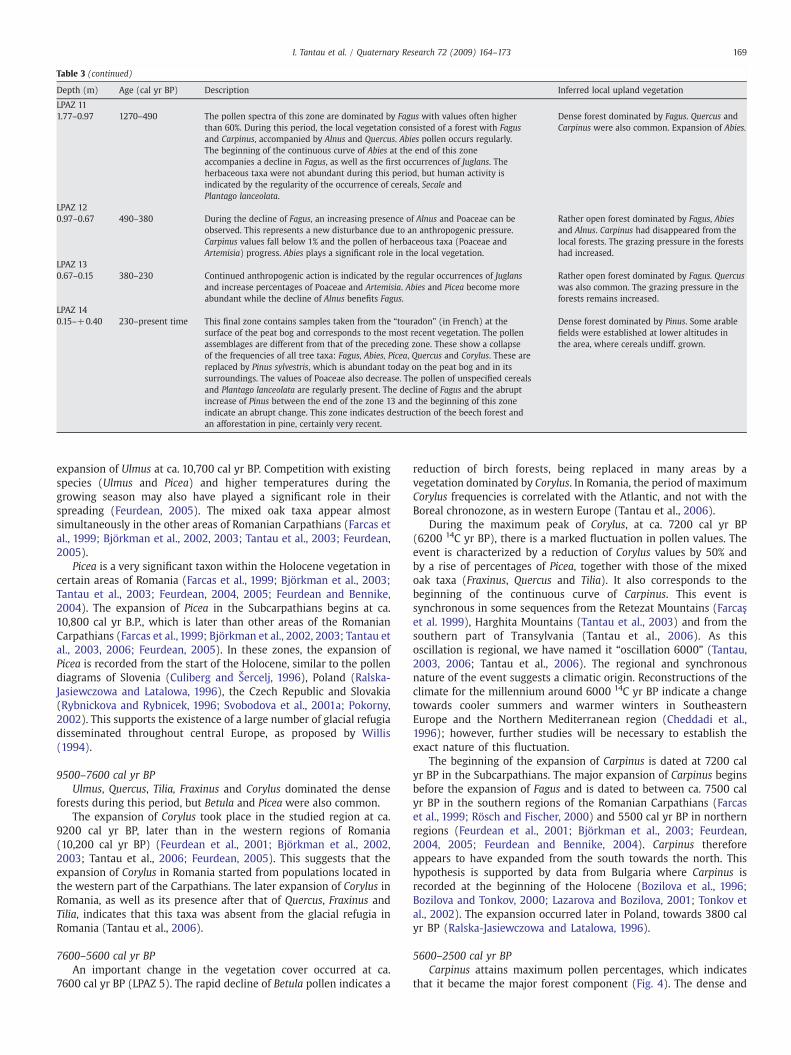
Table 3 (continued)
Depth (m) Age (cal yr BP) Description Inferred local upland vegetation
LPAZ 111.77–0.97 1270–490 The pollen spectra of this zone are dominated by Fagus with values often higher
than 60%. During this period, the local vegetation consisted of a forest with Fagusand Carpinus, accompanied by Alnus and Quercus. Abies pollen occurs regularly.The beginning of the continuous curve of Abies at the end of this zoneaccompanies a decline in Fagus, as well as the first occurrences of Juglans. Theherbaceous taxa were not abundant during this period, but human activity isindicated by the regularity of the occurrence of cereals, Secale andPlantago lanceolata.
Dense forest dominated by Fagus. Quercus andCarpinus were also common. Expansion of Abies.
LPAZ 120.97–0.67 490–380 During the decline of Fagus, an increasing presence of Alnus and Poaceae can be
observed. This represents a new disturbance due to an anthropogenic pressure.Carpinus values fall below 1% and the pollen of herbaceous taxa (Poaceae andArtemisia) progress. Abies plays a significant role in the local vegetation.
Rather open forest dominated by Fagus, Abiesand Alnus. Carpinus had disappeared from thelocal forests. The grazing pressure in the forestshad increased.
LPAZ 130.67–0.15 380–230 Continued anthropogenic action is indicated by the regular occurrences of Juglans
and increase percentages of Poaceae and Artemisia. Abies and Picea become moreabundant while the decline of Alnus benefits Fagus.
Rather open forest dominated by Fagus. Quercuswas also common. The grazing pressure in theforests remains increased.
LPAZ 140.15–+0.40 230–present time This final zone contains samples taken from the “touradon” (in French) at the
surface of the peat bog and corresponds to the most recent vegetation. The pollenassemblages are different from that of the preceding zone. These show a collapseof the frequencies of all tree taxa: Fagus, Abies, Picea, Quercus and Corylus. These arereplaced by Pinus sylvestris, which is abundant today on the peat bog and in itssurroundings. The values of Poaceae also decrease. The pollen of unspecified cerealsand Plantago lanceolata are regularly present. The decline of Fagus and the abruptincrease of Pinus between the end of the zone 13 and the beginning of this zoneindicate an abrupt change. This zone indicates destruction of the beech forest andan afforestation in pine, certainly very recent.
Dense forest dominated by Pinus. Some arablefields were established at lower altitudes inthe area, where cereals undiff. grown.
169I. Tantau et al. / Quaternary Research 72 (2009) 164–173
expansion of Ulmus at ca. 10,700 cal yr BP. Competition with existingspecies (Ulmus and Picea) and higher temperatures during thegrowing season may also have played a significant role in theirspreading (Feurdean, 2005). The mixed oak taxa appear almostsimultaneously in the other areas of Romanian Carpathians (Farcas etal., 1999; Björkman et al., 2002, 2003; Tantau et al., 2003; Feurdean,2005).
Picea is a very significant taxon within the Holocene vegetation incertain areas of Romania (Farcas et al., 1999; Björkman et al., 2003;Tantau et al., 2003; Feurdean, 2004, 2005; Feurdean and Bennike,2004). The expansion of Picea in the Subcarpathians begins at ca.10,800 cal yr B.P., which is later than other areas of the RomanianCarpathians (Farcas et al., 1999; Björkman et al., 2002, 2003; Tantau etal., 2003, 2006; Feurdean, 2005). In these zones, the expansion ofPicea is recorded from the start of the Holocene, similar to the pollendiagrams of Slovenia (Culiberg and Šercelj, 1996), Poland (Ralska-Jasiewczowa and Latalowa, 1996), the Czech Republic and Slovakia(Rybnickova and Rybnicek, 1996; Svobodova et al., 2001a; Pokorny,2002). This supports the existence of a large number of glacial refugiadisseminated throughout central Europe, as proposed by Willis(1994).
9500–7600 cal yr BPUlmus, Quercus, Tilia, Fraxinus and Corylus dominated the dense
forests during this period, but Betula and Picea were also common.The expansion of Corylus took place in the studied region at ca.
9200 cal yr BP, later than in the western regions of Romania(10,200 cal yr BP) (Feurdean et al., 2001; Björkman et al., 2002,2003; Tantau et al., 2006; Feurdean, 2005). This suggests that theexpansion of Corylus in Romania started from populations located inthe western part of the Carpathians. The later expansion of Corylus inRomania, as well as its presence after that of Quercus, Fraxinus andTilia, indicates that this taxa was absent from the glacial refugia inRomania (Tantau et al., 2006).
7600–5600 cal yr BPAn important change in the vegetation cover occurred at ca.
7600 cal yr BP (LPAZ 5). The rapid decline of Betula pollen indicates a
reduction of birch forests, being replaced in many areas by avegetation dominated by Corylus. In Romania, the period of maximumCorylus frequencies is correlated with the Atlantic, and not with theBoreal chronozone, as in western Europe (Tantau et al., 2006).
During the maximum peak of Corylus, at ca. 7200 cal yr BP(6200 14C yr BP), there is a marked fluctuation in pollen values. Theevent is characterized by a reduction of Corylus values by 50% andby a rise of percentages of Picea, together with those of the mixedoak taxa (Fraxinus, Quercus and Tilia). It also corresponds to thebeginning of the continuous curve of Carpinus. This event issynchronous in some sequences from the Retezat Mountains (Farcaşet al. 1999), Harghita Mountains (Tantau et al., 2003) and from thesouthern part of Transylvania (Tantau et al., 2006). As thisoscillation is regional, we have named it “oscillation 6000” (Tantau,2003, 2006; Tantau et al., 2006). The regional and synchronousnature of the event suggests a climatic origin. Reconstructions of theclimate for the millennium around 6000 14C yr BP indicate a changetowards cooler summers and warmer winters in SoutheasternEurope and the Northern Mediterranean region (Cheddadi et al.,1996); however, further studies will be necessary to establish theexact nature of this fluctuation.
The beginning of the expansion of Carpinus is dated at 7200 calyr BP in the Subcarpathians. The major expansion of Carpinus beginsbefore the expansion of Fagus and is dated to between ca. 7500 calyr BP in the southern regions of the Romanian Carpathians (Farcaset al., 1999; Rösch and Fischer, 2000) and 5500 cal yr BP in northernregions (Feurdean et al., 2001; Björkman et al., 2003; Feurdean,2004, 2005; Feurdean and Bennike, 2004). Carpinus thereforeappears to have expanded from the south towards the north. Thishypothesis is supported by data from Bulgaria where Carpinus isrecorded at the beginning of the Holocene (Bozilova et al., 1996;Bozilova and Tonkov, 2000; Lazarova and Bozilova, 2001; Tonkov etal., 2002). The expansion occurred later in Poland, towards 3800 calyr BP (Ralska-Jasiewczowa and Latalowa, 1996).
5600–2500 cal yr BPCarpinus attains maximum pollen percentages, which indicates
that it became the major forest component (Fig. 4). The dense and
170 I. Tantau et al. / Quaternary Research 72 (2009) 164–173
diverse forest dominated by Carpinus, Picea and Quercus also includedCorylus, Betula, Alnus, Ulmus and Tilia. Changes in temperature andhumidity trends between 5600 and 3500 cal yr BP may possibly havebeen important in enabling forest reorganization and expansion ofCarpinus.
The first indications of human activity are observed at ca. 3800 calyr BP, with the first occurrence of cereal pollen. Cereals are recordedduring the Carpinus optimum and throughout the Fagus phase. Theexpansion of beech occurs at the same time as an increase in markersof agricultural activities, around 3000 cal yr BP.
2500–230 cal yr BPFrom 2500 cal yr BP onwards dense forests dominated by Fagus
occurred in the area. Quercus, Carpinus, Alnus, Betula and Abies werealso common. The expansion of Fagus may have been a response toa combination of climatic changes, when average temperaturesdropped and precipitation increased (Holzhauser et al., 2005), andhuman activity (Küster, 1997). In contrast to Western Europe, wherebeech replaces the former dominant Quercetum mixtum, here Car-pinus decreases as Fagus increases. Some arable fields wereestablished at lower altitudes in the area, where cereals undiff.and Secale were grown.
Figure 4. (A, B) Pollen diagrams from Bisoca: LG—Late Glacial, PB—Preboreal, BO—Boreal, AT—the figure where they were collected.
Studies from Bulgaria have shown the presence of Fagus sincethe beginning of the Holocene (Tonkov, 2003), similar to Hungary(Nagy-Bodor et al., 2000). In the Czech Republic, the expansion ofFagus was recorded between 7500 and 8000 cal yr BP (Svobodovaet al., 2001a,b). In contrast, in Romania, the expansion of theforests dominated by Fagus occurred much later. It is generallydated at ca. 4500 cal yr BP, a date similar to that observed in thesoutheastern part of Poland (Ralska-Jasiewczowa and Latalowa,1996). Recent data show an age of ca. 7000 cal yr BP for theexpansion of Fagus in the Apuseni Mountains (Bodnariuc, 2000;Bodnariuc et al., 2002).
A recent study using combined fossil and genetic data for thereconstruction of the history of Fagus sylvatica in Europe indicate thatthe Romanian Carpathians were not a major refugium for beech, butsome smaller populations may have survived in the ApuseniMountains (Magri et al., 2006).
The first occurrence of the pollen of Juglans is recorded only at600 cal yr BP, later than in all other studied sites from Romania.
The few pollen grains of cereal-types and Secale are interpreted asoriginating from cultivated land at lower altitudes. Archaeologicalevidence to confirm pollen datawas found in some sites near BuzăuluiSubcarpathians. Traces of the Monteoru culture (Bronze Age) and
Atlantic, SB—Subboreal, SA—Subatlantic. Uncalibrated 14C ages (14C yr BP) are shown in
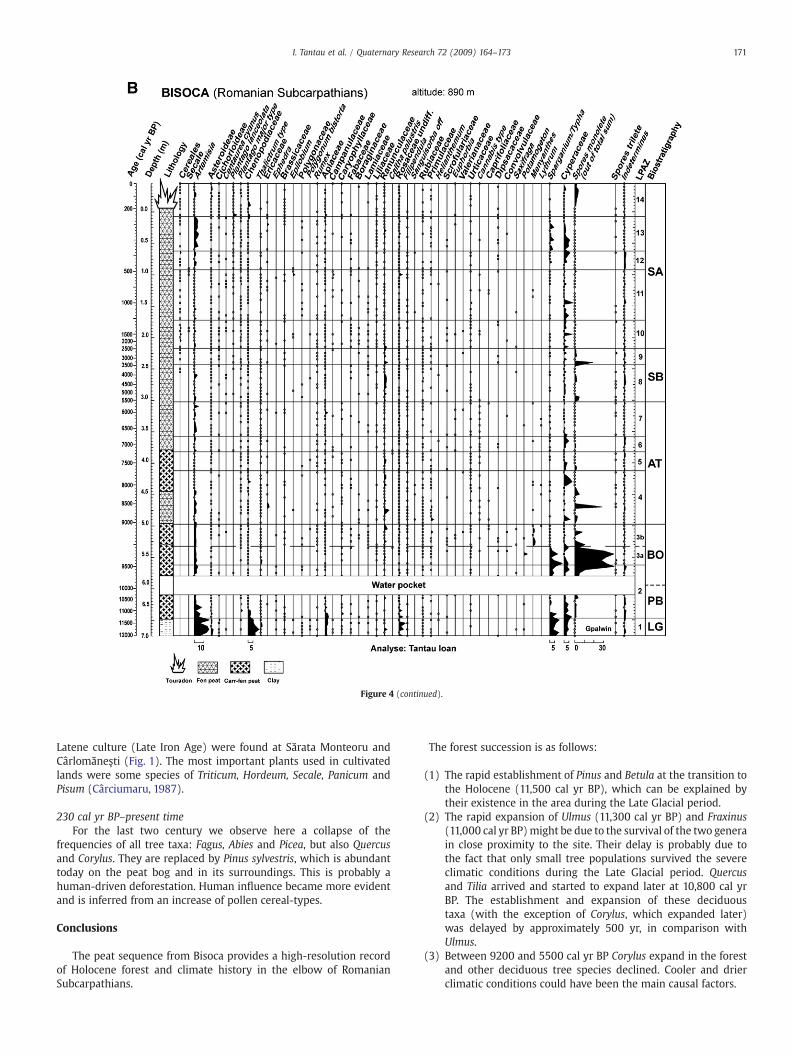
Figure 4 (continued).
171I. Tantau et al. / Quaternary Research 72 (2009) 164–173
Latene culture (Late Iron Age) were found at Sărata Monteoru andCârlomăneşti (Fig. 1). The most important plants used in cultivatedlands were some species of Triticum, Hordeum, Secale, Panicum andPisum (Cârciumaru, 1987).
230 cal yr BP–present timeFor the last two century we observe here a collapse of the
frequencies of all tree taxa: Fagus, Abies and Picea, but also Quercusand Corylus. They are replaced by Pinus sylvestris, which is abundanttoday on the peat bog and in its surroundings. This is probably ahuman-driven deforestation. Human influence became more evidentand is inferred from an increase of pollen cereal-types.
Conclusions
The peat sequence from Bisoca provides a high-resolution recordof Holocene forest and climate history in the elbow of RomanianSubcarpathians.
The forest succession is as follows:
(1) The rapid establishment of Pinus and Betula at the transition tothe Holocene (11,500 cal yr BP), which can be explained bytheir existence in the area during the Late Glacial period.
(2) The rapid expansion of Ulmus (11,300 cal yr BP) and Fraxinus(11,000 cal yr BP)might be due to the survival of the two generain close proximity to the site. Their delay is probably due tothe fact that only small tree populations survived the severeclimatic conditions during the Late Glacial period. Quercusand Tilia arrived and started to expand later at 10,800 cal yrBP. The establishment and expansion of these deciduoustaxa (with the exception of Corylus, which expanded later)was delayed by approximately 500 yr, in comparison withUlmus.
(3) Between 9200 and 5500 cal yr BP Corylus expand in the forestand other deciduous tree species declined. Cooler and drierclimatic conditions could have been the main causal factors.

172 I. Tantau et al. / Quaternary Research 72 (2009) 164–173
(4) An event that occurs during the maximum of Corylus, at ca.7200 cal yr BP (6200 14C yr BP) can be correlatedwith a climaticmodification. Cooler and moist climatic conditions may havefavored Picea expansion.
(5) Carpinus and Fagus were probably present in the forest afterca. 8000 cal yr BP. However, they expanded only at 7000 calyr BP and at 3000 cal yr BP, respectively, which may beexplained by competition with previously established foresttaxa.
(6) Human impact in the area is mainly related to forestry andforest grazing. In historic times the changes in the natural forestcover were primarily caused by anthropogenic activities,namely animal husbandry and agriculture.
Acknowledgments
The authors thank Florin Farcas for the help during the fieldwork.The comments of the anonymous reviewers and the editorial help ofDr. Alan Gillespie were greatly appreciated. This study was supportedby a grant PNCDI of the Romanian Ministry of Education and Research(CERES 4-205). 14C dates were financed by the FOSSILVA Europeanresearch project.
References
Balteanu, D., 1976. Two case studies of mudflows in the Buzau Subcarpathians.Geografiska Annaler, Series A, Physical Geography 58 (3), 165–171.
Björkman, S., Feurdean, A., Cinthio, K., Wohlfarth, B., Possnert, G., 2002. Late Glacial andearly Holocene vegetation development in the Gutâiului Mountains, northwesternRomania. Quaternary Science Reviews 21, 1039–1059.
Björkman, S, Feurdean, A, Wohlfarth, B, 2003. Late Glacial and Holocene forest dyna-mics at Steregoiu in the Gutâiului Mountains, Northwest Romania. Review ofPalaeobotany and Palynology 124, 79–111.
Bodnariuc, A., 2000. Histoire holocène de la végétation des Monts Apuseni (CarpatesOccidentales, Roumanie)— étude palynologique. Thèse de doctorat, Université PaulSabatier Toulouse III, Laboratoire d'Ecologie Terrestre.
Bodnariuc, A., Bouchette, A., Dedoubat, J.J., Otto, T., Fontugne, M., Jalut, G., 2002.Holocene vegetational history of the Apuseni mountains, central Romania.Quaternary Science Reviews 21, 1465–1488.
Boscaiu, N., Lupsa, V., 1982. Analyse sporo-pollinique de la tourbe eutrophe de la valléede Bahna, près des villages Dersca et Lozna (district Suceava). In: Paleobotany–palynology symposium, Cluj-Napoca, Facultatea de Biologie, Geografie, Geologie.pp. 103–105.
Bozilova, E., Tonkov, S., 2000. Pollen from Lake Sedmo Rilsko reveals southeastEuropean postglacial vegetation in the highest mountain area of the Balkans. NewPhytologist 148, 315–322.
Bozilova, E., Filipova, M., Filipovch, L., Tonkov, S., 1996. Bulgaria. In: Berglund, B.E., Birks,H.J.B., Ralska-Jasiewiczova, M., Wright, H.E. (Eds.), Palaeoecological Events duringthe Last 15000 Years. Regional Syntheses of Palaeoecological Studies of Lakes andMires in Europe. InWiley, Chichester, pp. 701–728.
Cârciumaru,M.,1987. Plante folosite de traco-geto-daci. Thraco-Dacica 8 (1–2),171–176.Cheddadi, R., Yu, G., Guiot, J., Harrison, S.P., Prentice, I.C., 1996. The climate of Europe
6000 years ago. Climate Dynamics 13, 1–9.Ciobanu, I., 1959. Istoria vegetatiei demonstrata in analiza de polen a turbei din RPR.
[Vegetation history demonstrated in the pollen analysis of peat from Romania].Studia Universitatis “Babes-Bolyai”, Seria Biologia 2 (2), 35–45.
Ciobanu, I., 1960. Analiza polinică a turbei mlaştinii Rut din Muntii Harghita [Pollenanalysis of the peat bog Rut fromHarghita Mountains]. Contributii Botanice 231–238.
Ciocârlan, V., 2000. Flora ilustrată a României (Pteridophyta et Spermatophyta)[Illustrated flora from Romania]. Editura Ceres, Bucureşti. 1138 p.
Coldea, G. (Ed.), Sanda, V., Popescu, A., Stefan, N., 1997. Les Associations Végétales deRoumanie 1. Les Associations Herbacées Naturelles. Presse Universitaire Cluj.
Constantin, M., 1999. Some considerations on geomorphologic hazards in Panatau Area(Buzau Subcarpathians). Transactions Japanese Geomorphological Union 20 (3),279–286.
Culiberg, M., Šercelj, A., 1996. Slovenia. In: Berglund, B.E., Birks, H.J.B., Ralska-Jasiewiczova, M., Wright, H.E. (Eds.), Palaeoecological Events during the Last15000 Years. Regional Syntheses of Palaeoecological Studies of Lakes and Mires inEurope. InWiley, Chichester, pp. 687–700.
Damblon, F., 1981. De l'analyse pollinique des touradons de monocotylédones commetémoins de l'histoire récente de la végétation. Comptes Rendus de l'Académie desSciences Paris, série 3 292 (23), 1191–1194.
Diaconeasa, B., Farcas, S., 1995–1996. Stejarisurile amestecate, evolutia si dinamica lor intardiglaciarul si holocenul din Romania [Evolution and dynamics of Quercetummixtum in Romania during the Late Glacial and Holocene]. Contributii Botanice103–115.
Diaconeasa, B., Farcas, S., 1996. L'affirmation subatlantique du charme dans lesstructures sylvestres de Roumanie. Contributii Botanice 2, 245–252.
Farcas, S., de Beaulieu, J.L., Reille, M., Coldea, G., Diaconeasa, B., Goeury, C., Goslar, T., Jull,T., 1999. First 14C datings of Late Glacial and Holocene pollen sequences fromRomanian Carpathes. Comptes Rendus de l'Académie des Sciences de Paris 322,799–807.
Feurdean, A., 2004. Palaeoenvironment in Romania during last 15,000 years. PhD thesisStockholm University, Sweden, 44 pp+4 appendix.
Feurdean, A., 2005. Holocene forest dynamics in northwestern Romania. The Holocene13, 435–446.
Feurdean, A., Bennike, O., 2004. Late Quaternary palaeoecological and paleoclimatolo-gical reconstruction in the Gutaiului Mountains, NW Romania. Journal ofQuaternary Science 19, 809–827.
Feurdean, A., Willis, K.J., 2008. The usefulness of a long-term perspective in assessingcurrent forest conservation management in the Apuseni Natural Park, Romania.Forest Ecology and Management 256, 421–430.
Feurdean, A., Björkman, S., Wohlfarth, B., 2001. A paleoecological reconstruction of theLate Glacial and Holocene based in multidisciplinary studies at Steregoiu site (GutaiMts, NW Romania). Studia Universitatis Babes-Bolyai, Geologia 46, 125–140.
Feurdean, A., Mosbrugger, V., Onac, B.P., Polyak, V., Veres, D., 2007a. Younger Dryas tomid-Holocene environmental history of the lowlands of NW Transylvania,Romania. Quaternary Research 68 (3), 364–378.
Feurdean, A., Wohlfarth, B., Björkman, S., Tantau, I., Bennike, O., Willis, K.J., Farcas, S.,Robertsson, A.M., 2007b. The influence of refugial population on Lateglacial andearly Holocene vegetational changes in Romania. Review of Palaeobotany andPalynology 145, 305–320.
Goeury, C., 1997. GpalWin: gestion, traitement et représentation de la paléoécologie.XV-ème Symposium de l'APLF (Association des Palynologues de Langue Française),Université Claude Bernard, Lyon, Septembre 1997.
Goeury, C., de Beaulieu, J.L., 1979. A propos de la concentration du pollen a l'aide de laliqueur de Thoulet dans les sédiments minéraux. Pollen et Spores 21, 239–251.
Holzhauser, H., Magny, M., Zumbuhl, H.J., 2005. Glacier and lake-level variations inwest-central Europe over the last 3500 years. The Holocene 15 (6), 789–801.
Küster, H., 1997. The role of farming in the postglacial expansion of beech and hornbeamin the oak woodlands of central Europe. The Holocene 7, 239–242.
Lazarova, M., Bozilova, E., 2001. Studies on the Holocene history of vegetation in theregion of lake Srebarna (northeast Bulgaria). Vegetation History and Archae-obotany 10, 87–95.
Magri,D., Vendramin,G., Comps, B., Dupanloup, I.,Geburek, T.,Gömöry,D., Latalowa,M., Litt,T., Paule, L., Roure, J.M., Tantau, Van Der Knaap, W.O., Petit, R., de Beaulieu, J.L., 2006. Anew scenario for the Quaternary history of European beech populations: palaeobota-nical evidence and genetic consequences. New Phytologist 171 (1), 199–223.
Magyari, E.K., Buczkó,, G., Jakab, G., Braun,M., Szántó,, Z., Molnár, M., Pál, Z., Karátson, D.,2006. Holocene palaeohydrology and environmental history in the South HarghitaMountains, Romania. Földtani Közlöny 136 (2), 249–284.
Nagy-Bodor, E., Jarai-Komlodi, M., Medve, A., 2000. Late Glacial and Post-Glacial pollenrecords and inferred climatic changes from Lake Balaton and the Great HungarianPlain. In: Hart, M.B. (Ed.), Climate: Past and Present. Special Publication no. 181.InGeological Society, London, pp. 121–133.
Pokorny, P., 2002. A high-resolution record of Late-Glacial and Early-Holocene climatic andenvironmental change in the Czech Republic. Quaternary International 91, 101–122.
Pop, E., 1929. Analize de polen în turba Carpaţilor Orientali (Dorna-Lucina) [Pollenanalysis in the peat from the Eastern Carpathians]. Buletinul Grădinii Botanice Cluj9 (3–4), 81–210.
Pop, E., 1932. Contributii la istoria padurilor din nordul Transilvaniei [Contributions tothe forest history from the northern Transylvania]. Buletinul Grădinii Botanice Cluj12, 29–102.
Pop, E., 1942. Contributii la istoria padurilor din nordul Transilvaniei [Contributions tothe forest history from the northern Transylvania]. Buletinul Grădinii Botanice Cluj22, 101–177.
Pop, E., 1960. Mlastinile de turba din Republica Populara Română [Peat bogs fromRomania]. Editura Academiei Republici Populare Române, Bucuresti.
Pop, E., Ciobanu, I., 1957. Analize de polen în turba de la Cotul Carpaţilor [Pollen analysisin the peat from the Elbow of Carpathians]. Buletinul Universitatii “V. Babeş” şi“Bolyai” Cluj, Seria Ştiintele Naturii 1 (1–2), 453–471.
Posea, G., Ielenicz, M., 1977. Munţii Buzăului. Ghid turistic. Colecţia Munţii Noştri, 13.Editura Sport-Turism, Sibiu.
Ralska-Jasiewczowa, M., Latalowa, M., 1996. Poland. In: Berglund, B.E., Birks, H.J.B.,Ralska-Jasiewiczova, M., Wright, H.E. (Eds.), Palaeoecological Events during the Last15000 Years. Regional Syntheses of Palaeoecological Studies of Lakes and Mires inEurope. InWiley, Chichester, pp. 404–472.
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Bertrand, C.J.H., Blackwell, P.G.,Buck, C.E., Burr, G.S., Cutler, K.B., Damon, P.E., Edwards, R.L., Fairbanks, R.G., Friedrich,M., Guilderson, T.P., Hogg, A.G., Hughen, K.A., Kromer, B., McCormac, G., Manning, S.,Ramsey, C., Reimer, R.W., Remmele, S., Southon, J.R., Stuiver, M., Talamo, S., Taylor, F.W., van der Plicht, J., Weyhenmeyer, C.E., 2004. IntCal04 atmospheric radiocarbonage calibration, 26–0 cal kyr BP. Radiocarbon 46, 1029–1058.
Rösch, M., Fischer, E., 2000. A radiocarbon dated Holocene pollen profile from the Banatmountains (Southwestern Carpathians, Romania). Flora 195, 277–286.
Rybnickova, E., Rybnicek, K., 1996. Czech Republic. In: Berglund, B.E., Birks, H.J.B.,Ralska-Jasiewiczova, M., Wright, H.E. (Eds.), Palaeoecological Events during the Last15000 Years. Regional Syntheses of Palaeoecological Studies of Lakes and Mires inEurope. InWiley, Chichester, pp. 473–506.
Stuiver, M., Reimer, P.J., Reimer, R.W., 2005. CALIB 5.0. [WWW program anddocumentation].
Svobodova, H., Reille, M., Goeury, C., 2001a. Past vegetation dynamics of Vltavsky luh,upper Vltava river valley in the Sumavamountains, Czech Republic. VegetationHistory and Archaeobotany 10, 185–199.

173I. Tantau et al. / Quaternary Research 72 (2009) 164–173
Svobodova, H., Soukupova, L., Reille, M., 2001b. Diversified development of mountainmires, Bohemian Forest, Central Europe, in the last 13,000 years. QuaternaryInternational 91, 123–135.
Tantau, I., 2003. Recherches pollenanalytiques dans les Carpates Orientales (Roumanie).Histoires de la végétation et de l'impact humain. Thèse de doctorat, Universite'sAix-Marseille III et Babes-Bolyai, Cluj-Napoca.
Tantau, I., 2006. Histoire de la végétation tardiglaciaire et holocène dans les CarpatesOrientales (Roumanie). Presa Universitară Clujeană, Cluj-Napoca. 200 p.
Tantau, I., Reille, M., de Beaulieu, J.L., Farcas, 2006. Late Glacial and Holocenevegetation history in the southern part of Transylvania (Romania): pollenanalysis of two sequences from Avrig. Journal of Quaternary Science 21 (1),49–61.
Tonkov, S., 2003. Holocene palaeovegetation of the Northwestern PirinMountains (Bulgaria)as reconstructed from pollen analysis. Review of Palaeobotany and Palynology 124,1–11.
Tonkov, S., Panovska, H., Possnert, G., Bozilova, E., 2002. The Holocene vegetationhistory of Northern Pirin Mountain, southwestern Bulgaria: pollen analysis andradiocarbon dating of a core from Lake Ribno Banderishko. Holocene 12, 201–210.
Tantau, I., Reille, M., de Beaulieu, J.L., Farcas, S., Goslar, T., Paterne, M., 2003. Vegetationhistory in the eastern Romanian Carpathians: pollen analysis of two sequencesfrom the Mohos crater. Vegetation History and Archaeobotany 12, 113–125.
Tutin, T.G. et al., 1964–80. Flora Europaea, vol 1 (1964); vol 2 (1968); vol 3 (1972); vol 4(1976); vol 5 (1980). Cambridge University Press, Cambridge.
Willis, K.J., 1994. The vegetational history of the Balkans. Quaternary Science Reviews13, 769–788.





























