Embed Size (px)
Citation preview

ORIGINAL ARTICLE
doi:10.1111/j.1558-5646.2007.00091.x
HAWTHORN-INFESTING POPULATIONS OFRHAGOLETIS POMONELLA IN MEXICO ANDSPECIATION MODE PLURALITYXianfa Xie,1 Juan Rull,2 Andrew P. Michel,1 Sebastian Velez,1,3 Andrew A. Forbes,1 Neil F. Lobo,1
Martin Aluja,2 and Jeffrey L. Feder1,4,5
1Department of Biological Sciences, University of Notre Dame, Notre Dame, Indiana 46556-03692Instituto de Ecologıa, Asociacion Civil, Km 2.5 Antigua Carretera a Coatepec No 351, 91070 Xalapa, Veracruz, Mexico
5E-mail: [email protected]
Received July 13, 2006
Accepted January 11, 2007
Categorizing speciation into dichotomous allopatric versus nonallopatric modes may not always adequately describe the geographic
context of divergence for taxa. If some of the genetic changes generating inherent barriers to gene flow between populations
evolved in geographic isolation, whereas others arose in sympatry, then the mode of divergence would be mixed. The apple maggot
fly, Rhagoletis pomonella, has contributed to this emerging concept of a mixed speciation mode “plurality.” Genetic studies have
implied that a source of diapause life-history variation associated with inversions and contributing to sympatric host race formation
and speciation for R. pomonella in the United States may have introgressed from the Eje Volcanico Trans Mexicano (EVTM; a.k.a.
the Altiplano) in the past. A critical unresolved issue concerning the introgression hypothesis is how past gene flow occurred given
the current 1200-km disjunction in the ranges of hawthorn-infesting flies in the EVTM region of Mexico and the southern extreme
of the U.S. population in Texas. Here, we report the discovery of a hawthorn-infesting population of R. pomonella in the Sierra
Madre Oriental Mountains (SMO) of Mexico. Sequence data from 15 nuclear loci and mitochondrial DNA imply that the SMO flies
are related to, but still different from, U.S. and EVTM flies. The host affiliations, diapause characteristics, and phylogeography of
the SMO population are consistent with it having served as a conduit for gene flow between Mexico and the United States. We
also present evidence suggesting greater permeability of collinear versus rearranged regions of the genome to introgression, in
accord with recent models of chromosomal speciation. We discuss the implications of the results in the context of speciation mode
plurality. We do not argue for abandoning the terms sympatry or allopatry, but caution that categorizing divergence into either/or
geographic modes may not describe the genetic origins of all species. For R. pomonella in the United States, the proximate selection
pressures triggering race formation and speciation stem from sympatric host shifts. However, some of the phenological variation
contributing to host-related ecological adaptation and reproductive isolation in sympatry at the present time appears to have an
older history, having originated and become packaged into inversion polymorphism in allopatry.
KEY WORDS: Chromosomal inversions, differential introgression, host races, Sierra Madre Oriental, sympatric speciation.
One of the most contested issues in speciation theory has centered
on biogeography: Is the complete geographic isolation of popula-
tions a prerequisite for speciation or can divergence be initiated in
3Present address: Museum of Comparative Zoology, Harvard Uni-
versity, 26 Oxford St, Cambridge, Massachusetts 02138.4Corresponding author.
the face of gene flow? Recently, however, there has been a grow-
ing awareness that dichotomizing speciation into strictly allopatric
versus nonallopatric modes may not always adequately describe
the geographic context of divergence for taxa. As noted by Mallet
(2005), “If some genetic changes leading to reproductive isola-
tion occur in allopatry, and others in sympatry, then what is the
geographic mode of speciation? An obvious answer would be a
1091C© 2007 The Author(s). Journal compilation C© 2007 The Society for the Study of Evolution.Evolution 61-5: 1091–1105

XIE ET AL.
mixture.” In this regard, Coyne and Orr (2004) have proposed sev-
eral new terms describing situations of mixed geographic mode
including “allo-parapatric” speciation (e.g., speciation by rein-
forcement), “para-allopatric” speciation (when populations ini-
tially differentiate while still in partial contact, but accumulate
substantial reproductive isolation only after a subsequent period
of geographic isolation), and “allo-sympatric” speciation. These
terms are not without their practical difficulties. For example, if
populations underwent repeated episodes of isolation and contact
during divergence, with varying degrees of spatial overlap occur-
ring during different contact periods, then we could have specia-
tion modes such as allo-para-sym-para-allo-parapatric. Neverthe-
less, the endeavor heralds the development of a more pluralistic
view of speciation mode, advancing the field beyond the mindset
of strict allopatric versus nonallopatric exclusivity.
Distinguishing the signature of a mixed, pluralistic mode of
speciation from a single mode of divergence can be a difficult
task, however. Identifying key traits responsible for the reproduc-
tive isolation of taxa and establishing their involvement in the
speciation process is daunting enough. To assess a mixed mode
hypothesis one must further integrate these data with historical in-
formation to show that the genetic bases for key traits separating
taxa originated in different geographic contexts.
Despite the complexity of the undertaking, one might still
expect to see certain patterns of genetic differentiation charac-
teristic of a mixed speciation mode. For example, taxa experi-
encing episodes of population subdivision interspersed with pe-
riods of contact and differential gene flow may display a mosaic
genome structure, with genes associated with different isolating
barriers varying in their coalescence times in a manner reflect-
ing the chronology of their respective evolutionary origins. Thus,
coalescence times for certain genes underlying reproductive isola-
tion may be congruent with periods of past geographic separation
between taxa when the trait differences they encode arose. In con-
trast, coalescence times for other genes may converge on different
periods of ecological radiation or reinforcement when populations
were in contact. Moreover, genomic regions not associated with
reproductive isolation may show little or no divergence if pop-
ulations recently or currently overlap and hybridize (Rieseberg,
et al. 1999; Noor et al. 2001a, b; Machado et al. 2002; Feder
et al. 2003a, 2005; Stump et al. 2005; Turner et al. 2005; Rogers
and Bernatchez 2005; Payseur and Nachman 2005; Osada and
Wu 2005).
The apple maggot fly, Rhagoletis pomonella (Diptera:
Tephritidae), has contributed to the emerging concept of a mixed
speciation mode plurality in which the genetic seeds for reproduc-
tive isolation that germinate in speciation may be planted in differ-
ent geographic contexts. Rhagoletis pomonella has long served as
a model for sympatric speciation via host plant shifting and spe-
cialization for phytophagous insects (Bush 1966, 1969). Indeed,
Bush (1966, 1969) has argued that the four described and sev-
eral undescribed taxa comprising the R. pomonella sibling species
complex in North America, including the apple and hawthorn
races of R. pomonella, the snowberry maggot (R. zephyria), the
blueberry maggot (R. mendax), the shrubby dogwood fly (R. cor-
nivora), and the undescribed flowering dogwood (Cornus florida)
fly, all arose via sympatric host shifts. Molecular sequence data,
however, have added a pluralistic twist to the sympatric story for
Rhagoletis: a geographic source of genetic variation for diapause
life-history may have contributed to the adaptive radiation of the
R. pomonella species group (Feder et al. 2003a). Based on gene
trees constructed for three anonymous nuclear loci and mtDNA,
it was inferred that an ancestral, hawthorn-infesting fly popula-
tion became geographically subdivided into Mexican and U.S.
isolates about 1.57 million years ago (Feder et al. 2003a). During
this period of separation, inversions appeared to have arisen and
fixed in the isolated fly population located in the Eje Volcanico
Trans Mexicano (EVTM; a.k.a. Altiplano) for three different ge-
nomic regions on chromosomes 1, 2, and 3. These inversions may
have fixed in the EVTM isolate due to the capture of favorable
combinations of genes affecting diapause life history in accord
with recent theory supporting the local adaptive significance of
rearrangements (Kirkpatrick and Barton 2006). Following subse-
quent secondary contact, introgression from the EVTM fly pop-
ulation into the United States established inversion clines in the
United States for diapause traits adapting flies to latitudinal and
local variation in host phenology influenced by seasonal climatic
conditions. Sometime later, the prestanding diapause variation in
the latitudinal clines appears to have aided the U.S. population
in sympatrically shifting and adapting to a variety of new plants
with differing fruiting times, including introduced apple. There-
fore, a portion of the genetic variation contributing to the adaptive
radiation of the R. pomonella complex in the United States may
have originated in an earlier time and a different place than the
proximate behavioral and ecological factors triggering sympatric
host shifts and divergence in more recent times.
Four points should be emphasized concerning this mixed,
pluralistic model of divergence hypothesized for R. pomonella.
First, we are not arguing that allopatry was necessary at some
stage of divergence for R. pomonella flies to speciate or that host
race formation in the apple maggot could not have happened with-
out it. Rather, we are contending that some of the quantitative
genetic variation facilitating diapause life-history adaptation for
R. pomonella in the United States and its packaging into chro-
mosomal inversions had geographic roots predating host shift-
ing in the complex. Subsequently, the divergent ecological selec-
tion that transformed within-population diapause variation into
between-race and among-species differences in the United States
occurred in sympatry and was associated with shifts to new host
plants with differing fruiting times.
1092 EVOLUTION MAY 2007

MEXICAN RHAGOLETIS
Second, the inversions and their component genes do not
display fixed differences between the apple and hawthorn host
races or among the various R. pomonella group sibling species in
the United States, but rather differ in allele/haplotype frequency.
Nonetheless, host races and species differ greatly and even dis-
cretely in their eclosion phenotypes in a manner generating sub-
stantial reproductive isolation. Thus, it is possible to have quanti-
tative genetic differences between host races and species.
Third, the inversion polymorphisms are just one factor con-
tributing to race formation and speciation for R. pomonella flies.
Other, as yet uncharacterized, genes may also affect the dia-
pause differences between the taxa. In addition, host discrimi-
nation traits are also important for causing reproductive isolation
in R. pomonella (Feder et al. 1994; Linn et al. 2003). These other
genes and traits may have more recent, nonallopatric genetic his-
tories that do not involve inversions.
Finally, analysis of an additional set of 12 nuclear loci has im-
plied that differential introgression is occurring between Mexican
EVTM and U.S. populations (Feder et al. 2005). In particular,
the inverted regions of chromosomes 1–3 that introgressed in the
more distant past now appear to be less permeable to gene flow
than loci in putative collinear regions of the genome on other
chromosomes (Feder et al. 2005). Six of the 12 additional loci
analyzed in Feder et al. (2005) map to chromosomes other than
1–3, outside of the rearrangements. These six loci displayed less
divergence (shallower coalescence times) between Mexican and
U.S. populations than genes associated with the inversions (Feder
et al. 2005). The pattern is consistent with new models of chro-
mosomal speciation (Rieseberg 2001; Noor et al. 2001a; Navarro
and Barton 2003; Kirkpatrick and Barton 2006) that hypothesize
that reduced recombination associated with rearrangements facil-
itates the retention of linked blocks of genes conferring differ-
ential adaptation between hybridizing taxa. In contrast, collinear
portions of the genome tend to introgress because of uninhib-
ited recombination. Studies in sunflowers (Rieseberg et al. 1999),
the Drosophila pseudoobscura subgroup (Wang et al. 1997; Noor
et al. 2001a, b; Machado et al. 2002), and Anopheles mosquitoes
(Stump et al. 2005; Turner et al. 2005) have found evidence for
greater introgression in collinear segments of the genome. Simi-
larly, the molecular data for R. pomonella suggest repeated cycles
(n ≥ 2) of population isolation and contact between Mexico and
the United States, with differential gene flow occurring across the
genome (Feder et al. 2005).
There are alternative scenarios to differential introgression
of inversion polymorphism from the EVTM fly population into
the United States, however, that could potentially explain the pat-
tern of genetic differentiation in R. pomonella. For example, it is
always possible that the inversion polymorphism initially arose
in the absence of geographic isolation internally within the U.S.
fly population and formed primary clines, perhaps as a result of
adaptational stress during range expansion from a southern refuge
after a glaciation period about 1.57 million years ago. Under this
hypothesis, the simultaneity of chromosomal origins would be ex-
plained as a pervasive adaptive challenge occurring during range
spread rather than due to a combination of ecology and allopatry.
Later gene flow between the United States and Mexico of unin-
verted chromosomes would be responsible for the greater simi-
larity of these sequences. However, as enumerated in Feder et al.
(2003a), a primary cline scenario would appear less likely than
secondary contact. Even if selective challenges can be great dur-
ing range expansions, this does not insure that co-adapted sets
of alleles will necessarily be captured in an inversion or that
different inversions will arise congruently across the genome at
roughly the same time. Moreover, although genetic exchange is
reduced between rearranged chromosomal sequences, recombi-
nation and gene conversion often occur, albeit at a reduced level
(Charlesworth 1974). Given a 1.57 million years ago timeframe
for the origin and continued co-existence of the inversions in sym-
patry under the primary cline hypothesis, we may therefore expect
to see significant shuffling of sequences within loci on chromo-
somes 1–3 degrading haplotype differences, but we do not. Finally,
the EVTM still displays a mtDNA signature of past geographic
subdivision 1.57 million years ago. If gene flow of uninverted se-
quences occurred in the EVTM, then it would have had to have
been extensive and obliterated all traces of past subdivision that
would have been present in these nuclear haplotypes, as well,
which seems unlikely.
Several critical unresolved issues still remain, however, con-
cerning the differential introgression hypothesis for R. pomonella.
Perhaps the foremost question centers on the current ranges of
hawthorn-infesting flies in Mexico and the United States. Feder
et al. (2005) argued that relatively recent gene flow between the
Mexican population in the EVTM and the United States is re-
sponsible for the limited population structure displayed by the six
putative collinear loci. However, EVTM and U.S. populations are
currently separated by a distance of at least 1200 km, implying
the need for large range shifts in the relatively recent past to ac-
count for the hypothesized gene flow. Occasional long-distance
dispersal of flies could also be invoked to explain the data (e.g., by
hurricanes). But given the extensive gene flow for collinear regions
implied by the molecular data and the likely rarity of long-range
migration given the distances involved, this hypothesis seems un-
likely. Obviously, the current distribution of R. pomonella does not
reflect its past range, which certainly must have changed dramat-
ically along with its hawthorn hosts during cooling and warming
periods associated with Pleistocene glaciations. Despite the po-
tential for substantial fluctuations in fly distribution, however, it
still seems unlikely that the ranges of Mexican EVTM and U.S.
populations changed so drastically in the recent past to bridge the
1200-km gap.
EVOLUTION MAY 2007 1093

XIE ET AL.
The distribution of hawthorn-infesting flies in Mexico is not
fully resolved, however. Bush (1966) first documented the exis-
tence of R. pomonella in the central EVTM highlands of Mexico,
but he reported that the exact limits of the species’ distribution
were unknown. Based on four collecting sites, Bush (1966) con-
sidered the distribution of the fly to likely correspond to the range
of Crataegus mexicana, a native hawthorn host of R. pomonella
in Mexico. This distribution roughly matches the transvolcanic
biogeographic area of endemism in Mexico (Marshall and Lieb-
herr 2000). Moreover, initial molecular genetic studies of Mexican
flies by Feder et al. (2003a) were based on U.S. Department of
Agriculture intercepts of infested C. mexicana fruit from Mexico
City, located on the EVTM plateau.
DISTINCT HAWTHORN FLY POPULATIONS IN MEXICO?
To better understand the biogeography of R. pomonella in Mexico,
Rull et al. (2006) initiated a survey of Rhagoletis on eight of the
13 endemic hawthorn species throughout the country. Three ma-
jor findings emerged from the study. First, the study expanded
the range of “R. pomonella-like” flies to cover much of the geo-
graphic distribution of Crataegus in Mexico, including the Sierra
Madre Oriental Mountain range (SMO) and the Sierra de las Altas
de Chiapas (Fig. 1). The two major exceptions were the apparent
absence of the fly from northwestern Mexico and perhaps parts of
the Sierra Madre del Sur Mountains. Second, the study enlarged
the host range of R. pomonella, documenting infestation for five
of the eight native hawthorn species surveyed. Third, the results
Figure 1. Collection sites for pomonella populations from Mexico
and the United States genetically analyzed in the current study.
See Table 1 for site information and designations. Sites MC and CJ
are located in the Eje Volcanico Trans Mexicano (EVTM) of Mexico,
sites SJ, CB, PL in the Sierra Madre Oriental Mountains (SMO) of
Mexico, site CP in the Sierra de las Altas de Chiapas, and sites NY,
MI, and TX in the United States.
revealed a significant difference in pupal body mass between flies
from the EVTM (mean = 11.17 ± 0.10 mg, n = 5 sites, 30 pu-
pae/site) and those from elsewhere in Mexico (mean = 7.86 ±0.45 mg, n = 6 sites; Fig. 2).
The difference in pupal body mass could either be due to pop-
ulation subdivision in Mexico or reflect environmental rather than
genetic variation. Flies from the EVTM primarily infest C. mex-
icana and C. rosei rosei (Rull et al. 2006). These two hawthorns
fruit late in the field season in the central EVTM highlands (infes-
tation period ranges from mid October to early January; Table 1).
In addition, C. mexicana bears large-sized hawthorn fruit, which
could help account for the larger pupal body size of EVTM flies.
In the SMO, C. rosei parrayana, C. gracilor, C. greggiana, and C.
cuprina are the predominant hosts (Rull et al. 2006). Infestation
occurs considerably earlier in the season in the SMO (late August
to early October; Table 1), suggesting possible allochronic isola-
tion between EVTM and SMO flies attacking different hawthorn
hosts (Rull et al. 2006). (Note: In the United States, R. pomonella
infests a variety of different hawthorn species mainly from mid
August to late October; Berlocher 2000.) However, in the transi-
tion zone between the EVTM and the SMO in the state of Veracruz
(Fig. 1), C. mexicana and C. rosei rosei co-occur with C. rosei
parrayana and C. gracilor, and all four hawthorns are infested by
Figure 2. Mean pupal mass for pomonella flies from Mexican
EVTM and SMO sites plotted against population altitude (n = 30
pupa measured per site) based on data from Rull et al. (2006). Re-
gression coefficient (r2) between altitude and pupal body mass =
0.16, P > 0.22, 10 df. Annual rainfall was also not significantly re-
lated to mean pupal mass (r2 = 0.01, P > 0.77, 10 df). The EVTM
sites in Mexico with increasing elevation from left to right along
the X-axis are Tancitaro, Michoacan (MC); Texcoco, state of Mexico;
Santa Marta, District Federal; Perote, Veracruz; and Coajomulco,
Morelos (CJ). The SMO sites in Mexico from left to right along the
X-axis are Piletas, Veracruz (PL); Los Pinos, Veracruz; San Cistobal,
Chiapas (CP); Casa Blanca, Veracruz (CB); Cerro el Potosı, Nuevo
Leon; and San Joaquin, Queretaro (SJ). Two letter abbreviations
given in parentheses for the sites above designate fly populations
genetically analyzed in the current study.
1094 EVOLUTION MAY 2007
MEXICAN RHAGOLETIS
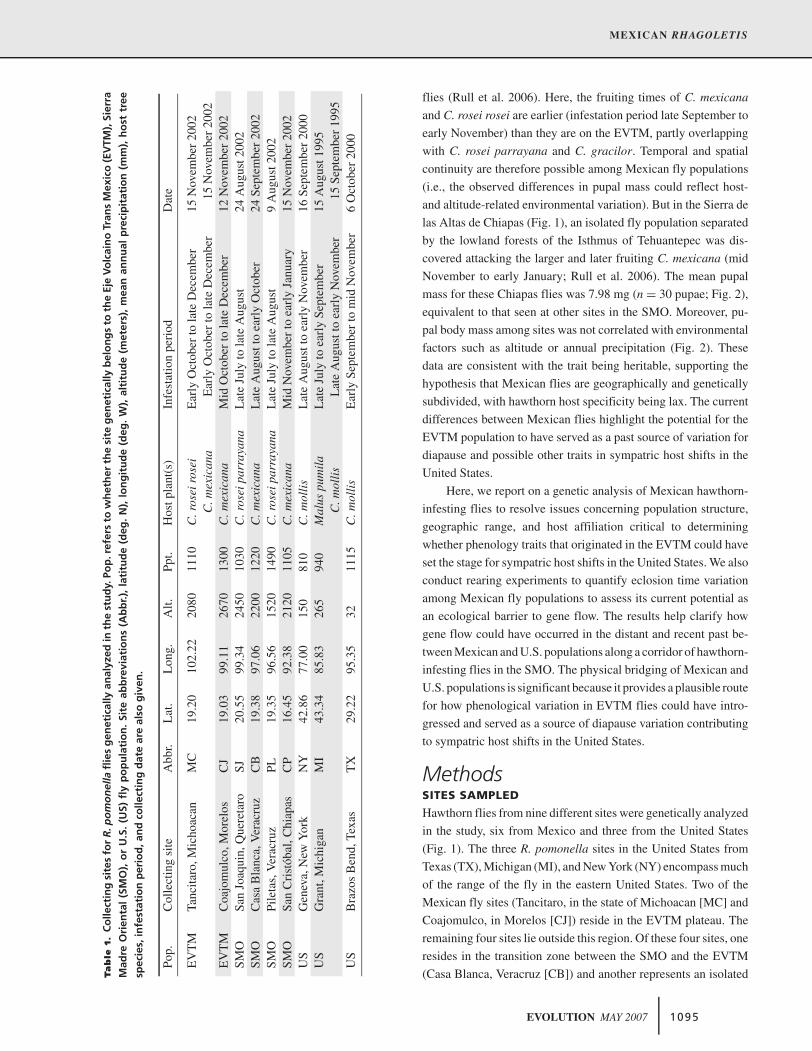
Tab
le1.
Co
llect
ing
site
sfo
rR
.po
mo
nel
lafl
ies
gen
etic
ally
anal
yzed
inth
est
ud
y.Po
p.r
efer
sto
wh
eth
erth
esi
teg
enet
ical
lyb
elo
ng
sto
the
Eje
Vo
lcai
no
Tran
sM
exic
o(E
VTM
),Si
erra
Mad
reO
rien
tal
(SM
O),
or
U.S
.(U
S)fl
yp
op
ula
tio
n.
Site
abb
revi
atio
ns
(Ab
br.)
,la
titu
de
(deg
.N
),lo
ng
itu
de
(deg
.W
),al
titu
de
(met
ers)
,m
ean
ann
ual
pre
cip
itat
ion
(mm
),h
ost
tree
spec
ies,
infe
stat
ion
per
iod
,an
dco
llect
ing
dat
ear
eal
sog
iven
.
Pop.
Col
lect
ing
site
Abb
r.L
at.
Lon
g.A
lt.Pp
t.H
ostp
lant
(s)
Infe
stat
ion
peri
odD
ate
EV
TM
Tanc
itaro
,Mic
hoac
anM
C19
.20
102.
2220
8011
10C
.ros
eiro
sei
Ear
lyO
ctob
erto
late
Dec
embe
r15
Nov
embe
r20
02C
.mex
ican
aE
arly
Oct
ober
tola
teD
ecem
ber
15N
ovem
ber
2002
EV
TM
Coa
jom
ulco
,Mor
elos
CJ
19.0
399
.11
2670
1300
C.m
exic
ana
Mid
Oct
ober
tola
teD
ecem
ber
12N
ovem
ber
2002
SMO
San
Joaq
uin,
Que
reta
roSJ
20.5
599
.34
2450
1030
C.r
osei
parr
ayan
aL
ate
July
tola
teA
ugus
t24
Aug
ust2
002
SMO
Cas
aB
lanc
a,V
erac
ruz
CB
19.3
897
.06
2200
1220
C.m
exic
ana
Lat
eA
ugus
tto
earl
yO
ctob
er24
Sept
embe
r20
02SM
OPi
leta
s,V
erac
ruz
PL19
.35
96.5
615
2014
90C
.ros
eipa
rray
ana
Lat
eJu
lyto
late
Aug
ust
9A
ugus
t200
2SM
OSa
nC
rist
obal
,Chi
apas
CP
16.4
592
.38
2120
1105
C.m
exic
ana
Mid
Nov
embe
rto
earl
yJa
nuar
y15
Nov
embe
r20
02U
SG
enev
a,N
ewY
ork
NY
42.8
677
.00
150
810
C.m
olli
sL
ate
Aug
ustt
oea
rly
Nov
embe
r16
Sept
embe
r20
00U
SG
rant
,Mic
higa
nM
I43
.34
85.8
326
594
0M
alus
pum
ila
Lat
eJu
lyto
earl
ySe
ptem
ber
15A
ugus
t199
5C
.mol
lis
Lat
eA
ugus
tto
earl
yN
ovem
ber
15Se
ptem
ber
1995
US
Bra
zos
Ben
d,Te
xas
TX
29.2
295
.35
3211
15C
.mol
lis
Ear
lySe
ptem
ber
tom
idN
ovem
ber
6O
ctob
er20
00
flies (Rull et al. 2006). Here, the fruiting times of C. mexicana
and C. rosei rosei are earlier (infestation period late September to
early November) than they are on the EVTM, partly overlapping
with C. rosei parrayana and C. gracilor. Temporal and spatial
continuity are therefore possible among Mexican fly populations
(i.e., the observed differences in pupal mass could reflect host-
and altitude-related environmental variation). But in the Sierra de
las Altas de Chiapas (Fig. 1), an isolated fly population separated
by the lowland forests of the Isthmus of Tehuantepec was dis-
covered attacking the larger and later fruiting C. mexicana (mid
November to early January; Rull et al. 2006). The mean pupal
mass for these Chiapas flies was 7.98 mg (n = 30 pupae; Fig. 2),
equivalent to that seen at other sites in the SMO. Moreover, pu-
pal body mass among sites was not correlated with environmental
factors such as altitude or annual precipitation (Fig. 2). These
data are consistent with the trait being heritable, supporting the
hypothesis that Mexican flies are geographically and genetically
subdivided, with hawthorn host specificity being lax. The current
differences between Mexican flies highlight the potential for the
EVTM population to have served as a past source of variation for
diapause and possible other traits in sympatric host shifts in the
United States.
Here, we report on a genetic analysis of Mexican hawthorn-
infesting flies to resolve issues concerning population structure,
geographic range, and host affiliation critical to determining
whether phenology traits that originated in the EVTM could have
set the stage for sympatric host shifts in the United States. We also
conduct rearing experiments to quantify eclosion time variation
among Mexican fly populations to assess its current potential as
an ecological barrier to gene flow. The results help clarify how
gene flow could have occurred in the distant and recent past be-
tween Mexican and U.S. populations along a corridor of hawthorn-
infesting flies in the SMO. The physical bridging of Mexican and
U.S. populations is significant because it provides a plausible route
for how phenological variation in EVTM flies could have intro-
gressed and served as a source of diapause variation contributing
to sympatric host shifts in the United States.
MethodsSITES SAMPLED
Hawthorn flies from nine different sites were genetically analyzed
in the study, six from Mexico and three from the United States
(Fig. 1). The three R. pomonella sites in the United States from
Texas (TX), Michigan (MI), and New York (NY) encompass much
of the range of the fly in the eastern United States. Two of the
Mexican fly sites (Tancitaro, in the state of Michoacan [MC] and
Coajomulco, in Morelos [CJ]) reside in the EVTM plateau. The
remaining four sites lie outside this region. Of these four sites, one
resides in the transition zone between the SMO and the EVTM
(Casa Blanca, Veracruz [CB]) and another represents an isolated
EVOLUTION MAY 2007 1095

XIE ET AL.
population in the Sierra de las Altas de Chiapas (San Cristobal
[CP]). The other two sites (Piletas, Veracruz [PL] and San Joaquin,
Queretaro [SJ]) are part of the SMO proper.
HOST PLANTS
Flies were collected as larvae in infested hawthorn fruit at all nine
sites and either immediately dissected from the fruit and frozen
for later genetic analysis or reared to adulthood in the laboratory.
At the Grant, MI, site, flies were collected from both sympatric
apple and hawthorn trees. Hawthorn host species and collecting
dates are given in Table 1. Note that the transitional CB popula-
tion at Casa Blanca, Veracruz, was sampled from C. mexicana, the
primary host for flies in the EVTM. The isolated CP population
from Chiapas was also collected from C. mexicana, a plant that is
not endemic to the region but was likely introduced by Tlaxcal-
tecan Indians accompanying Spanish conquistadors in the 1600s
(Standley and Steyermark 1946).
GENES SEQUENCED
Sequence data were generated for 15 nuclear loci and a 946 bp
fragment of the mitochondrial genome containing the 3′ portion
of COI, leucine tRNA, and the COII genes. The sequences are av
ailable in GenBank (accession numbers AY152477-AY152526,
AY930466-AY931013, and DQ812553-DQ812885). These nu-
clear and mtDNA loci formed the basis for the previous genetic
studies of EVTM and U.S. flies (Feder et al. 2003a, 2005). Nine
Table 2. Loci sequenced in the study. Given are chromosome map
positions (Chr.), probability level (P) for conformation to a molecu-
lar clock, the ML substitution model (Model) determined by Mod-
elTest using the Akaike information criterion (Posada and Cran-
dall, 1998), and the minimum number of recombination events
(Rec.) estimated by the method of Hudson and Kaplan (1985) for
each locus.
Locus Chr. P Model Rec.
P181 1 1.0000 HKY 0P220 1 0.9841 TIM 3P3072 1 0.9488 TIM+G 4P2473 2 0.9920 TrN+I 1P2956 2 0.9810 HKY+G 4P667 2 0.9992 TVM+G 5P8 2 1.0000 TVM 1P22 3 0.8646 TVM+I 0P7 3 0.7385 HKY 3P2963 4 0.9825 TIM+G 3P661 4 0.3171 TVM+G 5P1700 5 0.9999 HKY 4P2620 5 1.0000 K81uf 0P309 5 1.0000 K81uf+I 3P3060 – 1.0000 HKY 2mtDNA – 0.9991 TrN+I 0
of the loci (P220, P181, P3072, P2956, P667, P8, P2473, P7, and
P22) map to chromosomes 1–3 and are subsumed by chromoso-
mal inversions (Roethele et al. 2001; Feder et al. 2003b; Table 2)
(haploid n = 6 for R. pomonella). Allozyme markers within these
three inverted regions of the genome display significant allele fre-
quency differences between the sympatric apple and hawthorn
host races across the northeastern and midwestern United States
(Feder et al. 1988, 1990; McPheron et al. 1988; Feder and Bush
1989; Berlocher 2000). All three of the rearranged regions on
chromosomes 1–3 have been shown to correlate with diapause-
related traits, accounting for up to 30% of the phenotypic variation
for eclosion time within the hawthorn race at the Grant, MI, site
(Feder et al. 1993). The variation in diapause differentially adapts
the fly races to a three- to four-week difference in the peak fruiting
times of apples versus hawthorns, generating allochronic premat-
ing, as well as postzygotic ecological isolation between apple and
hawthorn flies (Feder et al. 1993, 1997a, b; Filchak et al. 2000).
Markers in these regions also display latitudinal allele frequency
clines within the U.S. host races (Feder and Bush 1989; Feder et al.
1990; Berlocher 2000). The clines match geographic variation in
host fruiting time; alleles associated with later adult eclosion time
and reduced propensity for nondiapause development are found in
higher frequencies in southern populations (Feder and Bush 1989;
Feder et al. 1990; Berlocher 2000).
Five other loci analyzed in the study (P661, P2963, P1700,
P2620, and P309) map to chromosomes 4 and 5 in the R. pomonella
genome (Table 2). The exact map position of P3060 is not known
but it is not located on chromosomes 1–3 (Feder, unpubl. data).
Allozymes on chromosomes 4 and 5 do not differ in allele fre-
quency between the host races, do not vary clinally, do not cor-
relate with the timing of eclosion, and do not display high lev-
els of linkage disequilibrium in nature (Feder et al. 1988, 1990,
1993, 2003b). The genetic data imply that P661, P2963, P1700,
P2620, P309, and P3060 are not associated with chromosomal
rearrangements (Feder et al. 2003b). In addition, these six loci ap-
pear to have introgressed more readily in the recent past between
EVTM and U.S. populations, possibly due to uninhibited recom-
bination in combination with a lack of differential selection (Feder
et al. 2005).
DNA CLONING AND SEQUENCING
Genomic DNA were isolated from individual flies and PCR am-
plified for 35 cycles (94◦C, 30 sec.; 52◦C, 1 min., 72◦C, 1.5
min.) using locus-specific primers for the 15 nuclear and mtDNA
fragment as described in Roethele et al. (2001). Products were
TA cloned into pCR II vectors (Invitrogen Corp., Carlsbad, CA).
PCR amplification products were initially cloned separately for
a minimum of two flies from each study site, with four to six
clones sequenced per locus per fly in both the 5′ and 3′ direc-
tions on an ABI 3700 sequencer using the ABI Prism�
BigDyeTM
1096 EVOLUTION MAY 2007

MEXICAN RHAGOLETIS
Terminator (Applied Biosystems Corp., Foster City, CA) version
3.0 system. To increase sample sizes for certain sites, we also sepa-
rately amplified genomic DNA for four to eight flies from the site,
and TA cloned the pooled amplification products for sequencing.
To avoid analysis of identical alleles from the same individual,
sequences generated from the pooled library were not included
unless they differed from each other.
GENE AND GENETIC DISTANCE TREE CONSTRUCTION
Parsimony and maximum likelihood gene trees were constructed
using PAUP∗b10 (Swofford 2002). For the parsimony analysis,
gaps were treated as a fifth base pair, with indels of identical
length and sequence position recoded to count as single muta-
tional steps. Rhagoletis electromorpha, which belongs to the sis-
ter species group (R. tabellaria) to R. pomonella, was used as an
outgroup taxon to root trees. Parsimony and maximum likelihood
trees were very similar and so we report the results for only the
parsimony trees here. Intragenic recombination was statistically
tested using the methods of Hudson and Kaplan (1985). Putative
recombinant alleles and gene regions were identified and these
alleles were excluded from initial parsimony gene tree construc-
tion. Recombinant alleles were then added to the trees by hand to
generate allele networks. The molecular clock was tested for each
locus for R. pomonella and R. electromorpha sequences by com-
paring log-likelihood scores enforcing versus relaxing the clock
hypothesis for the best supported DNA substitution model iden-
tified using ModelTest (Posada and Crandall 1998). To quantify
gene tree topology and genetic divergence, relative node depths
(RNDs) were calculated between the major haplotype classes of
alleles in Mexico and the United States as in Feder et al. (2005) by
dividing the number of substitution differences between a given
pair of Mexican and U.S. alleles by the mean number of sub-
stitutions between each and the outgroup R. electromorpha se-
quence. Assuming a molecular clock (which none of the nuclear
loci or mtDNA violated; Table 2) the mean RND for all pairs of
Mexican and U.S. alleles between two haplotype classes estimates
the age of separation of the haplotypes relative to the divergence
time of R. electromorpha plus the coalescence time in the com-
mon ancestor.
Neighbor-joining trees (Saitou and Nei 1987) summarizing
the overall genetic relatedness of populations were constructed
using PHYLIP, version 3.66 (Felsenstein 1989). Trees were con-
structed separately for loci mapping to chromosomes 1–3 and
those residing elsewhere in the genome. To construct the neighbor-
joining trees, mean pairwise uncorrected genetic distances were
first computed separately for each locus between each of the nine
EVTM, SMO, and U.S. populations, as well as between these
populations and the outgroup R. electromorpha using Mega, ver-
sion 3.1 (Kumar et al. 2004). The pairwise distance between two
populations for a locus was then divided by the average distance
of all R. pomonella populations to R. electromorpha to standard-
ize for sequence length and substitution rate differences among
loci. The standardized distances were averaged across the nine
loci on chromosomes 1–3 and the six loci mapping elsewhere to
give overall pairwise distance measures used for tree construction.
For chromosome 1–3 loci, north (N) and south-north (SN) hap-
lotypes were considered separately in the calculations of genetic
distance for NY and MI sites, generating NYN, NYSN, MIN, and
MISN populations for the analysis.
ANALYSIS OF POPULATION DIFFERENTIATION AND
STRUCTURE
Hierarchical analysis of molecular variance (AMOVA) was per-
formed using Arlequin 2.0 (Schneider et al. 2000) to test for
genetic structuring among R. pomonella populations. For the
AMOVA analysis, the nine sites under study were initially divided
into three populations based on the results from the neighbor-
joining genetic distance trees that distinguished (1) EVTM (MC
and CJ sites), (2) SMO (SJ, CB, PL, and CP sites), and (3) the
United States (MI, NY, and TX sites). AMOVA was performed for
the nine chromosome 1–3 loci with N haplotypes both included
and excluded from the analyses. The analysis with N haplotypes
excluded was performed because N haplotypes tend to inflate
intrasite and among site variation within populations relative to
among-population divergence for the nine chromosome 1–3 loci
due to the latitudinal clines they display in the United States. Note
that although the CP site from the Sierra de las Altas de Chiapas
is geographically isolated from the SMO, genetically the Chiapas
flies were found to be very similar to SMO flies (Fig. 3) and so
they were designated as part of the general SMO population in the
statistical analysis of population structure.
ECLOSION EXPERIMENTS
We performed a series of controlled rearing experiments to quan-
tify eclosion time variation among Mexican fly populations. Fly
larvae were collected from infested hawthorn fruit at the six Mex-
ican sites in the fall of 2003 and allowed to pupate in an ambient
temperature room at the Instituto de Ecologıa, A.C., in Xalapa,
Veracruz. Three weeks after puparium formation, pupae were col-
lected and placed in moist vermiculite in petri dishes. The petri
dishes were held at 5◦C in a refrigerator for 14 weeks to simulate
winter. After removal from the cold, pupae were placed in a 26.5◦C
constant temperature chamber and eclosing adults collected on a
daily basis as they emerged.
ResultsNone of the sequenced genes deviated significantly from a molec-
ular clock (Table 2). Twelve of the 15 nuclear loci displayed ev-
idence for possible recombination, as implied by the method of
Hudson and Kaplan (Table 2). Inferred recombination was gener-
ally limited, however, to alleles within the same haplotype class
EVOLUTION MAY 2007 1097
XIE ET AL.
of inversion or geographic population (EVTM, SMO, or United
States; see online Supplementary Figs. S1–S15). There was no
evidence for recombination among mtDNA sequences (Table 2).
GENES ON CHROMOSOMES 1–3
Neighbor-joining genetic distance and parsimony gene trees for
the nine nuclear loci on chromosomes 1–3 implied the existence of
three genetically distinguishable hawthorn-infesting populations
of pomonella flies in North America: (1) EVTM, (2) SMO, and
(3) United States (see Fig. 3A for the neighbor-joining tree and
Fig. 4A–C for representative gene trees for the loci P220, P2956,
and P7; online Supplementary Figs. S1–S9 provide more exten-
sive gene trees of all nine loci sequenced from chromosomes 1–3).
Genetic subdivision was also evident in the hierarchical AMOVA
analysis. FCT values among EVTM, SMO, and U.S. populations
were significant for all nine chromosome 1–3 loci, regardless of
whether N haplotypes were included or excluded from the analysis
(Table 3). Even with N haplotypes included in the AMOVA, the
percentage of variation explained was substantially higher among
EVTM, SMO, and U.S. populations (mean = 43.4 ± 4.27%, SE,
n = 9) than among sites within these populations (mean = 12.4 ±2.80% SE, n = 9). Population structuring was even more pro-
nounced when N haplotypes were removed from the AMOVA
analysis, with the mean percentage of genetic variation explained
by among-population differences rising to 51.2 ± 7.71%. Seven of
the nine genes displayed synapomorphic substitutions or unique
combinations of derived mutations distinguishing EVTM, SMO,
and U.S. flies. As in Feder et al. (2003a, 2005), N haplotypes
were unique to northern U.S. sites (MI and NY), whereas variation
within the southern clade of “SN/M” (south-north and Mexico) al-
leles differentiated among EVTM, SMO, and Texas flies (Fig. 4A–
C; online Supplementary Figs. S1–S9).
The SMO fly population was more closely related, in an over-
all sense, to the U.S. than to EVTM population (Fig. 3A). The
mean genetic distance between SMO and U.S. populations for
SN haplotypes relative to R. electromorpha was less (0.245 ±0.031 SE; n = 8 loci; locus P8 possessed only N haplotypes in the
United States) than that between the SMO and EVTM (0.353 ±0.0591; n = 9 loci). However, the loci P3072, P2473, and P667
deviated from this general trend (see online Supplementary Figs.
S3, S4, and S6), implying that EVTM and SMO flies (or at least
a subset of alleles in these two populations) were most closely
related. In no case, however, were EVTM and U.S. SN alleles
genealogically more closely related than EVTM and SMO haplo-
types (mean relative genetic distance between EVTM and United
States = 0.368 ± 0.052, n = 9 loci; see Fig. 4A–C and online Sup-
plementary Figs. S1–S9). Thus, the SMO population appears to
comprise a composite set of alleles with ties to both the EVTM and
United States.
Figure 3. Neighbor-joining trees based on overall genetic dis-
tances among EVTM, SMO, and U.S. fly populations relative to
the outgroup R. electromorpha for (A) chromosome 1–3 loci, and
(B) loci not residing on chromosomes 1–3. Trees are scaled so that
the lengths of the branch from the R. electromorpha/R. pomonella
node to terminal populations are the same in (A) and (B). Relative
genetic distances are given on branches. For chromosome 1–3 loci,
the N and SN clades of haplotypes present at NY and MI were con-
sidered as separate populations (NYN, NYSN, MIN, and MISN in [A],
respectively).
GENES MAPPING OUTSIDE CHROMOSOMES 1–3
The six loci mapping outside chromosomes 1–3 also showed sig-
nificant genetic structuring among EVTM, SMO, and U.S. flies,
as indicated by hierarchical AMOVA analysis (Table 3). P661,
P2963, P1700, P2620, P309, and P3060 all displayed significant
FCT values (Table 3). Although a significant proportion of the
genetic variation present for P661, P2963, P1700, P2620, P309,
1098 EVOLUTION MAY 2007
MEXICAN RHAGOLETIS
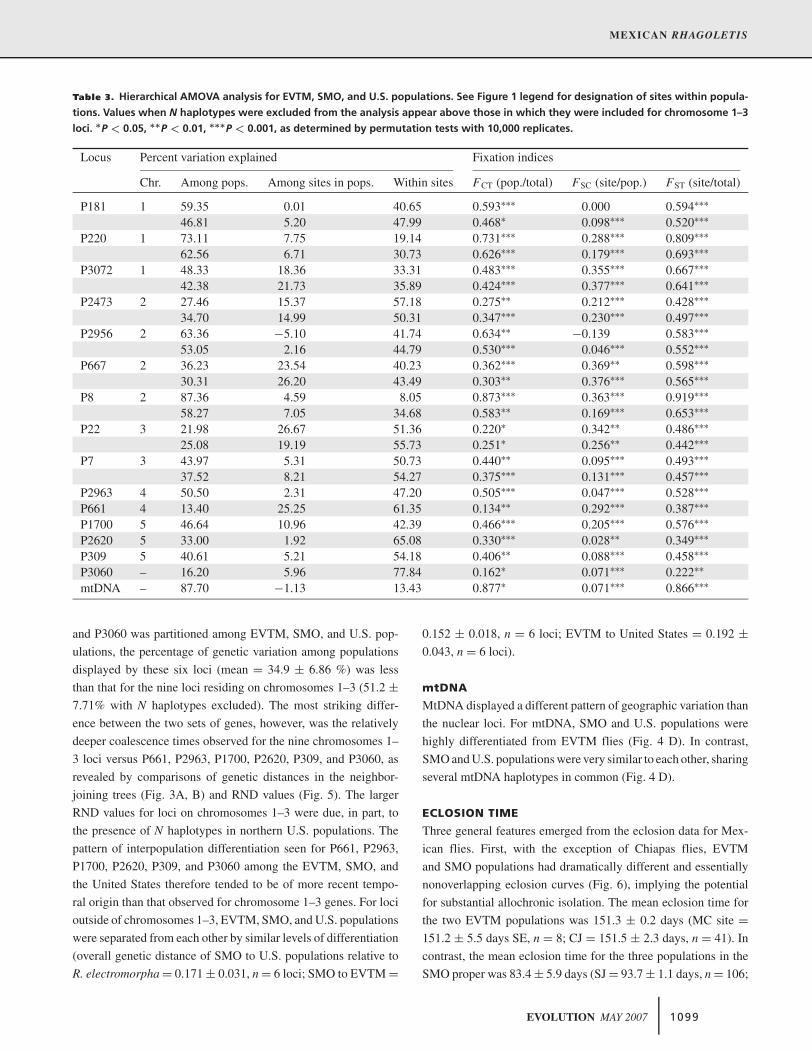
Table 3. Hierarchical AMOVA analysis for EVTM, SMO, and U.S. populations. See Figure 1 legend for designation of sites within popula-
tions. Values when N haplotypes were excluded from the analysis appear above those in which they were included for chromosome 1–3
loci. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001, as determined by permutation tests with 10,000 replicates.
Locus Percent variation explained Fixation indices
Chr. Among pops. Among sites in pops. Within sites FCT (pop./total) FSC (site/pop.) FST (site/total)
P181 1 59.35 0.01 40.65 0.593∗∗∗ 0.000 0.594∗∗∗
46.81 5.20 47.99 0.468∗ 0.098∗∗∗ 0.520∗∗∗
P220 1 73.11 7.75 19.14 0.731∗∗∗ 0.288∗∗∗ 0.809∗∗∗
62.56 6.71 30.73 0.626∗∗∗ 0.179∗∗∗ 0.693∗∗∗
P3072 1 48.33 18.36 33.31 0.483∗∗∗ 0.355∗∗∗ 0.667∗∗∗
42.38 21.73 35.89 0.424∗∗∗ 0.377∗∗∗ 0.641∗∗∗
P2473 2 27.46 15.37 57.18 0.275∗∗ 0.212∗∗∗ 0.428∗∗∗
34.70 14.99 50.31 0.347∗∗∗ 0.230∗∗∗ 0.497∗∗∗
P2956 2 63.36 −5.10 41.74 0.634∗∗ −0.139 0.583∗∗∗
53.05 2.16 44.79 0.530∗∗∗ 0.046∗∗∗ 0.552∗∗∗
P667 2 36.23 23.54 40.23 0.362∗∗∗ 0.369∗∗ 0.598∗∗∗
30.31 26.20 43.49 0.303∗∗ 0.376∗∗∗ 0.565∗∗∗
P8 2 87.36 4.59 8.05 0.873∗∗∗ 0.363∗∗∗ 0.919∗∗∗
58.27 7.05 34.68 0.583∗∗ 0.169∗∗∗ 0.653∗∗∗
P22 3 21.98 26.67 51.36 0.220∗ 0.342∗∗ 0.486∗∗∗
25.08 19.19 55.73 0.251∗ 0.256∗∗ 0.442∗∗∗
P7 3 43.97 5.31 50.73 0.440∗∗ 0.095∗∗∗ 0.493∗∗∗
37.52 8.21 54.27 0.375∗∗∗ 0.131∗∗∗ 0.457∗∗∗
P2963 4 50.50 2.31 47.20 0.505∗∗∗ 0.047∗∗∗ 0.528∗∗∗
P661 4 13.40 25.25 61.35 0.134∗∗ 0.292∗∗∗ 0.387∗∗∗
P1700 5 46.64 10.96 42.39 0.466∗∗∗ 0.205∗∗∗ 0.576∗∗∗
P2620 5 33.00 1.92 65.08 0.330∗∗∗ 0.028∗∗ 0.349∗∗∗
P309 5 40.61 5.21 54.18 0.406∗∗ 0.088∗∗∗ 0.458∗∗∗
P3060 – 16.20 5.96 77.84 0.162∗ 0.071∗∗∗ 0.222∗∗
mtDNA – 87.70 −1.13 13.43 0.877∗ 0.071∗∗∗ 0.866∗∗∗
and P3060 was partitioned among EVTM, SMO, and U.S. pop-
ulations, the percentage of genetic variation among populations
displayed by these six loci (mean = 34.9 ± 6.86 %) was less
than that for the nine loci residing on chromosomes 1–3 (51.2 ±7.71% with N haplotypes excluded). The most striking differ-
ence between the two sets of genes, however, was the relatively
deeper coalescence times observed for the nine chromosomes 1–
3 loci versus P661, P2963, P1700, P2620, P309, and P3060, as
revealed by comparisons of genetic distances in the neighbor-
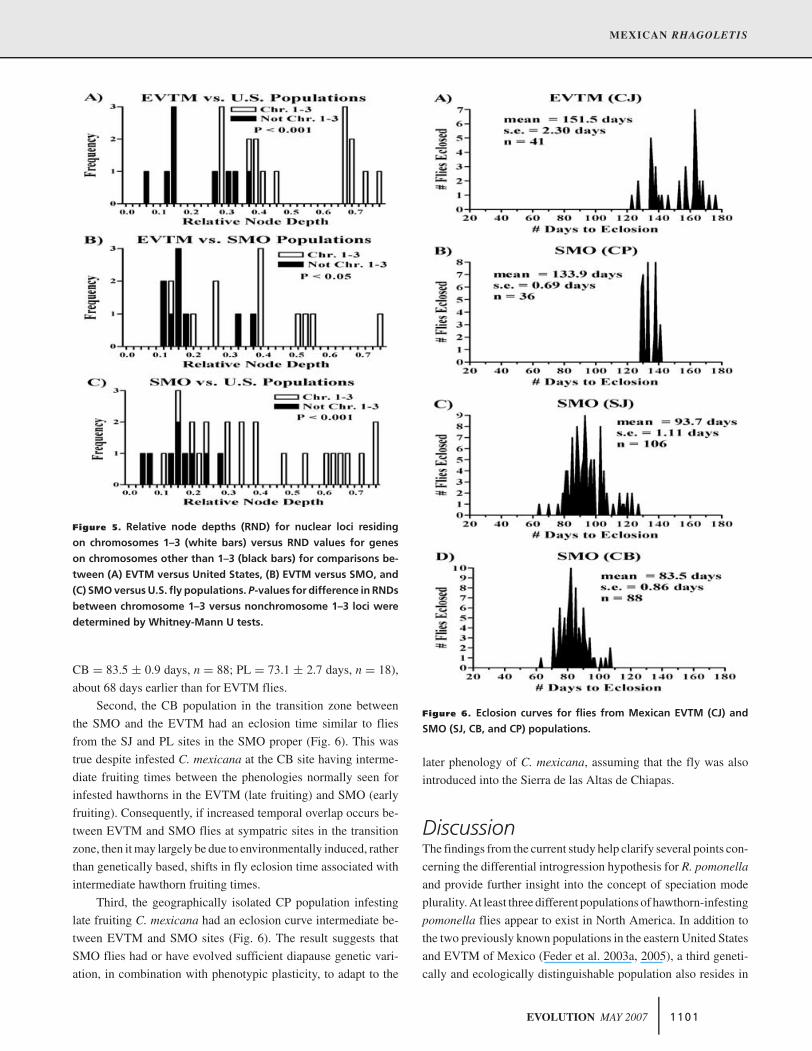
joining trees (Fig. 3A, B) and RND values (Fig. 5). The larger
RND values for loci on chromosomes 1–3 were due, in part, to
the presence of N haplotypes in northern U.S. populations. The
pattern of interpopulation differentiation seen for P661, P2963,
P1700, P2620, P309, and P3060 among the EVTM, SMO, and
the United States therefore tended to be of more recent tempo-
ral origin than that observed for chromosome 1–3 genes. For loci
outside of chromosomes 1–3, EVTM, SMO, and U.S. populations
were separated from each other by similar levels of differentiation
(overall genetic distance of SMO to U.S. populations relative to
R. electromorpha = 0.171 ± 0.031, n = 6 loci; SMO to EVTM =
0.152 ± 0.018, n = 6 loci; EVTM to United States = 0.192 ±0.043, n = 6 loci).
mtDNA
MtDNA displayed a different pattern of geographic variation than
the nuclear loci. For mtDNA, SMO and U.S. populations were
highly differentiated from EVTM flies (Fig. 4 D). In contrast,
SMO and U.S. populations were very similar to each other, sharing
several mtDNA haplotypes in common (Fig. 4 D).
ECLOSION TIME
Three general features emerged from the eclosion data for Mex-
ican flies. First, with the exception of Chiapas flies, EVTM
and SMO populations had dramatically different and essentially
nonoverlapping eclosion curves (Fig. 6), implying the potential
for substantial allochronic isolation. The mean eclosion time for
the two EVTM populations was 151.3 ± 0.2 days (MC site =151.2 ± 5.5 days SE, n = 8; CJ = 151.5 ± 2.3 days, n = 41). In
contrast, the mean eclosion time for the three populations in the
SMO proper was 83.4 ± 5.9 days (SJ = 93.7 ± 1.1 days, n = 106;
EVOLUTION MAY 2007 1099
XIE ET AL.
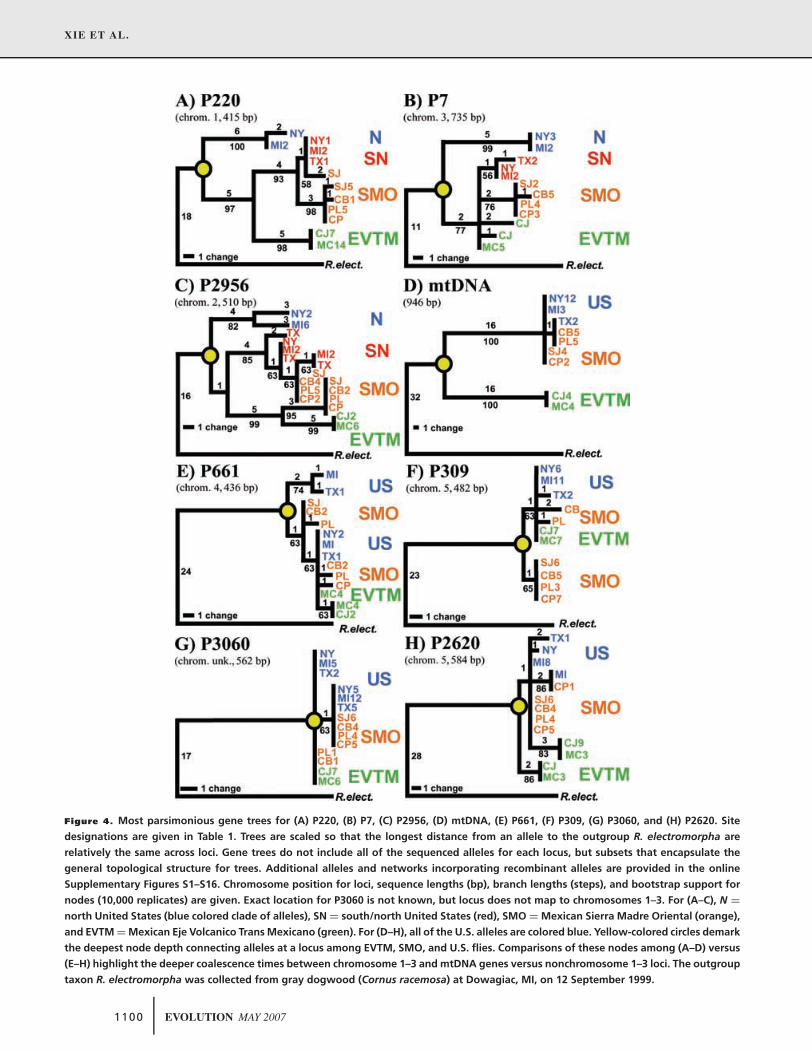
Figure 4. Most parsimonious gene trees for (A) P220, (B) P7, (C) P2956, (D) mtDNA, (E) P661, (F) P309, (G) P3060, and (H) P2620. Site
designations are given in Table 1. Trees are scaled so that the longest distance from an allele to the outgroup R. electromorpha are
relatively the same across loci. Gene trees do not include all of the sequenced alleles for each locus, but subsets that encapsulate the
general topological structure for trees. Additional alleles and networks incorporating recombinant alleles are provided in the online
Supplementary Figures S1–S16. Chromosome position for loci, sequence lengths (bp), branch lengths (steps), and bootstrap support for
nodes (10,000 replicates) are given. Exact location for P3060 is not known, but locus does not map to chromosomes 1–3. For (A–C), N =
north United States (blue colored clade of alleles), SN = south/north United States (red), SMO = Mexican Sierra Madre Oriental (orange),
and EVTM = Mexican Eje Volcanico Trans Mexicano (green). For (D–H), all of the U.S. alleles are colored blue. Yellow-colored circles demark
the deepest node depth connecting alleles at a locus among EVTM, SMO, and U.S. flies. Comparisons of these nodes among (A–D) versus
(E–H) highlight the deeper coalescence times between chromosome 1–3 and mtDNA genes versus nonchromosome 1–3 loci. The outgroup
taxon R. electromorpha was collected from gray dogwood (Cornus racemosa) at Dowagiac, MI, on 12 September 1999.
1100 EVOLUTION MAY 2007
MEXICAN RHAGOLETIS
Figure 5. Relative node depths (RND) for nuclear loci residing
on chromosomes 1–3 (white bars) versus RND values for genes
on chromosomes other than 1–3 (black bars) for comparisons be-
tween (A) EVTM versus United States, (B) EVTM versus SMO, and
(C) SMO versus U.S. fly populations. P-values for difference in RNDs
between chromosome 1–3 versus nonchromosome 1–3 loci were
determined by Whitney-Mann U tests.
CB = 83.5 ± 0.9 days, n = 88; PL = 73.1 ± 2.7 days, n = 18),
about 68 days earlier than for EVTM flies.
Second, the CB population in the transition zone between
the SMO and the EVTM had an eclosion time similar to flies
from the SJ and PL sites in the SMO proper (Fig. 6). This was
true despite infested C. mexicana at the CB site having interme-
diate fruiting times between the phenologies normally seen for
infested hawthorns in the EVTM (late fruiting) and SMO (early
fruiting). Consequently, if increased temporal overlap occurs be-
tween EVTM and SMO flies at sympatric sites in the transition
zone, then it may largely be due to environmentally induced, rather
than genetically based, shifts in fly eclosion time associated with
intermediate hawthorn fruiting times.
Third, the geographically isolated CP population infesting
late fruiting C. mexicana had an eclosion curve intermediate be-
tween EVTM and SMO sites (Fig. 6). The result suggests that
SMO flies had or have evolved sufficient diapause genetic vari-
ation, in combination with phenotypic plasticity, to adapt to the
Figure 6. Eclosion curves for flies from Mexican EVTM (CJ) and
SMO (SJ, CB, and CP) populations.
later phenology of C. mexicana, assuming that the fly was also
introduced into the Sierra de las Altas de Chiapas.
DiscussionThe findings from the current study help clarify several points con-
cerning the differential introgression hypothesis for R. pomonella
and provide further insight into the concept of speciation mode
plurality. At least three different populations of hawthorn-infesting
pomonella flies appear to exist in North America. In addition to
the two previously known populations in the eastern United States
and EVTM of Mexico (Feder et al. 2003a, 2005), a third geneti-
cally and ecologically distinguishable population also resides in
EVOLUTION MAY 2007 1101

XIE ET AL.
the SMO of Mexico. The genetic differences observed between
flies from the EVTM and SMO corresponded to variation in pu-
pal body weight observed among sites (Rull et al. 2006; Fig. 1),
suggesting that morphological as well as genetic differences exist
between these two populations.
The two Mexican populations appear to be primarily differ-
entiated by geography, diapause traits related to hawthorn fruiting
time, and body size, rather than host specificity. This contrasts with
the pattern seen for fly populations in the United States, where
host discrimination and diapause-related adaptations to host phe-
nology have together generated a diversity of host-specific sibling
species and races in sympatry (Berlocher et al. 1993; Berlocher
2000). (Note, however, that the scale of comparison is not equiv-
alent between the countries. In the United States, the host plants
attacked by R. pomonella represent different genera and families,
whereas in Mexico the hosts are different Crataegus species. In the
northeast United States, there is no evidence for different races of
R. pomonella on different hawthorn species, although we cannot
rule out this possibility in the southern United States.) Eclosion ex-
periments conducted in the current study indicate that the diapause
differences between the EVTM and SMO populations correspond
to differences in the fruiting phenologies of their respective host
plants. It is interesting that at one transition zone site between the
EVTM and SMO, flies infesting C. mexicana, one of the two hosts
used in the EVTM, but not found in the SMO proper, genetically
represent the SMO population. The same is also true for the iso-
lated CP population in Chiapas, where C. mexicana and the fly
may have been introduced. These results support the hypothesis
that host specificity among different hawthorn species may not be
strong in Mexico.
The phenology and phylogeography of the SMO population
imply that it may have been a conduit for gene flow between
Mexican EVTM and U.S. flies, bridging the about 1200-km gap
between the two populations. Our current working model is that
Mexican and U.S. fly populations have undergone repeated cycles
(≥ 2) of geographic isolation, contact, and differential introgres-
sion. Consistent with the hypothesis of fragmentation and periodic
contact, the SMO and EVTM populations currently abut through
parts of the Mexican states of Veracruz, Puebla, and Hidalgo (Rull
et al. 2006). Here, C. mexicana and C. rosei rosei co-occur with
C. rosei parrayana. Hawthorns identified as C. rosei rosei have
also been reported through much of the SMO. However, as dis-
cussed in Rull et al. (2006), the plant in this area likely represents
a different subspecies from the C. rosei rosei found on the EVTM,
as they differ in several morphological characters, including fruit
color and size. C. mexicana and C. rosei rosei in the transition
zone fruit earlier than they do on the EVTM and are infested by
fly larvae from late September to early November, with C. ro-
sei parrayana being infested from September to early October
(Rull et al. 2006). Thus, in the EVTM/SMO transition zone, host
species and host fruiting time overlap, creating the potential for
gene flow.
The extent to which the SMO population currently contacts
U.S. flies is not known. However, hawthorns are present in iso-
lated patches in southeastern New Mexico, and possibly the Davis
Mountains of Texas. In north-central Coahuila, the hawthorn C.
greggiana is found concentrated on lower north-facing slopes,
along streamsides, and in other locally mesic spots. Moreover, C.
greggiana is infested by R. pomonella in Mexico (Rull et al. 2006).
Although R. pomonella‘s current distribution does not necessarily
reflect its past geographic distribution, which likely shifted in the
Pleistocene during glacial and interglacial periods, the existence
of hawthorn flies along the SMO provides a possible mechanism
for the bridging of EVTM and U.S. populations at various times
in the past, when hawthorns were pushed further south in their
distribution during periods of northern glaciation. Further sam-
pling is needed in the northern range of the SMO population to
assess the degree of current contact with U.S. flies to determine
if a discontinuous genetic break actually exists or whether U.S.
flies represent the northern extension of SMO flies. If the latter
proves true, a result not inconsistent with the mtDNA data, then the
observed nuclear sequence differences in the current study may
be more accurately interpreted as geographic variation within a
semicontinuous SMO/U.S. population rather than reflecting di-
vergence between differentiated SMO and U.S. fly taxa.
A geographically disjunct population of hawthorn-infesting
pomonella flies now exists in the Sierra de las Altas de Chiapas
(represented by collection site CP in the current study), isolated
from other Mexican flies by the lowland forests of the Isthmus
of Tehuantepec. This population is genetically very similar to
SMO flies found to the north (SJ, CB, and PL sites), but the CP
population from the highlands of Chiapas has a different phenol-
ogy from the other SMO flies. In Chiapas, R. pomonella infests
C. mexicana from mid November through December and has an
eclosion curve intermediate between those of EVTM and SMO
flies from elsewhere. Hawthorns are not native to the Sierra de
las Altas de Chiapas, however. C. mexicana was likely introduced
into the region by Tlaxcaltecan Indians accompanying Spanish
conquistadors in the 1600s (Standley and Steyermark 1946). The
genetic similarity of flies from the SMO sites and CP flies sug-
gests that introductions of the plant and fly may have occurred
simultaneously, perhaps from the transition region between the
EVTM plateau and SMO where both occur. If true, then follow-
ing the introduction, the fruiting time of C. mexicana may have
been shifted later in the season in response to local conditions,
becoming similar in phenology to the plant on the EVTM. The
fly subsequently adapted by evolving a later eclosion life history
to match. This implies that the ancestral Chiapas fly population
possessed adequate diapause variation to track the changing phe-
nology of its host. Interestingly, this life-history shift does not
1102 EVOLUTION MAY 2007

MEXICAN RHAGOLETIS
appear to be reflected in significant genetic differentiation for the
CP population from the highlands of Chiapas for any of the 15
nuclear loci or mtDNA genes analyzed in the current study. It is
always possible that R. pomonella from Chiapas do not represent
an introduction, but an endemic population of more distant origin
in Central America that has experienced recent population bot-
tlenecks. Although we consider this scenario unlikely, additional
sampling of sites are needed to define the southern boundaries
of hawthorn-infesting flies in Central America and discount the
demographic hypothesis for Chiapas.
A DUAL ROLE FOR THE INVERSIONS IN SPECIATION?
In addition to their roles in facilitating sympatric host shifts, the in-
versions may also be serving as seeds fostering ongoing allopatric
divergence between Mexican and U.S. flies. The pattern of dif-
ferentiation across the nuclear genome among EVTM, SMO, and
U.S. populations varies in a manner consistent with reduced gene
flow for loci residing in the inverted regions of chromosomes
1–3. These three regions of the genome correlate with diapause
traits differentially adapting R. pomonella in the United States
to geographic and host-related variation in apple and hawthorn
fruiting time (Feder and Bush 1989; Feder et al. 1993, 1997a, b,
2003b; Filchak et al. 2000). These genes display greater diver-
gence among EVTM, SMO, and U.S. populations than loci in
putative collinear regions of the genome on chromosomes other
than 1–3. The reduced differentiation for collinear loci is not due to
an inherently slower substitution rate for these genes. Nucleotide
substitution rates between R. pomonella flies and the outgroup
species R. electromorpha are as great as or greater for these loci
as they are for loci on chromosomes 1–3 (mean Tamura-Nei ge-
netic distance as calculated by Mega, version 3.1, for chromo-
somes 1–3 loci = 0.051 ± 0.007, n = 9, range 0.0212–0.0895;
for nonchromosome 1–3 loci = 0.056 ± 0.006, n = 6, range
0.0331–0.718). Moreover, no locus deviated significantly from
a molecular clock. Rather, the lack of population differentiation
suggests more extensive and recent gene flow for collinear versus
rearranged loci among EVTM, SMO, and U.S. flies. Under this
scenario, past genetic introgression between Mexican and U.S. fly
populations generated the adaptive latitudinal inversion clines for
chromosomes 1–3 seen in the United States. However, subsequent
genetic changes that accumulated within the inversions, as well as
possible additional rearrangements involving these regions, have
since reduced their potential to introgress between Mexico and the
United States, consistent with models of chromosomal speciation
proposed by Rieseberg (2001), Noor et al. (2001a), Navarro and
Barton (2003), and Kirkpatrick and Barton (2006). Thus, even
when considering just the SN class of haplotypes, mean overall
genetic distances still tended to be greater among EVTM, SMO,
and U.S. populations for chromosome 1–3 than nonchromosome
1–3 loci (EVTM to SMO = 0.353 ± 0.0591 vs. 0.152 ± 0.018;
EVTM to United States = 0.368 ± 0.0520 vs. 0.192 ± 0.043;
SMO to United States = 0.245 ± 0.031 vs. 0.171 ± 0.032). The
reduced recombination associated with the rearrangements may
therefore accentuate the isolating effects of the inversions above
the genic consequences of their component loci considered alone.
Moreover, in addition to their role in adaptively differentiating R.
pomonella complex flies in the United States, the inversions may
also be facilitating population divergence on a broader geographic
scale among EVTM, SMO, and U.S. flies.
MtDNA displays a different pattern of genetic differentiation
than either the putative collinear or inverted regions of the nuclear
genome. For mtDNA, the SMO and U.S. populations are very
similar and differ from the EVTM. The pattern implies that mi-
tochondrial introgression has been inhibited between the EVTM
and the other populations since the original isolation of EVTM
flies about 1.57 million years ago (Feder et al. 2003a). However,
mtDNA gene flow appears to be extensive between SMO and U.S.
flies, suggesting that the populations may have only recently sep-
arated or that gene flow is ongoing. The cause for the apparent
impermeability of mtDNA to gene flow between EVTM versus
SMO and U.S. flies remains to be determined, with possible ex-
planations including differential migration of males, cytonuclear
gene interactions, and cytoplasmic incompatibilities due to en-
dosymbionts, such as Wolbachia.
ConclusionsThe discovery of the SMO fly population in Mexico supports
an emerging view of speciation mode plurality. For R. pomonella,
and perhaps many other organisms, allopatric versus nonallopatric
speciation modes may not be as dichotomous and mutually exclu-
sive categories of population divergence as they seem. Rather,
our results suggest that speciation is simultaneously unfolding
across different spatial and host-related axes in R. pomonella,
interconnected by considerations of life-history adaptation, host
specificity, genetic architecture, and gene flow.
In Mexico, geography appears to have contributed to initial
population divergence in a similar manner that host-specific mat-
ing plays in the United States. However, the physical isolation
of Mexican fly populations has not always been complete. In the
past, a portion of the diapause variation that evolved in the EVTM
and was associated with inversion polymorphism appears to have
introgressed via the SMO fly population into what is now the U.S.
population. The inversion polymorphism subsequently played a
complimentary role, when it became coupled with host discrim-
ination traits and ecologically different plant niches, in fostering
shifts to novel hosts and sympatric race formation/speciation in the
United States. Moreover, since the period of initial introgression,
rearranged regions of the genome appear to have evolved addi-
tional changes making them less permeable to gene flow. As a
EVOLUTION MAY 2007 1103

XIE ET AL.
result, Mexican and U.S. fly populations display a mosaic pattern
of differentiation across the genome, with inverted regions on
chromosomes 1–3 showing higher levels of divergence than pu-
tative collinear loci mapping elsewhere. Under this scenario, the
inversions served as focal points for the accumulation and reten-
tion of host and nonhost-related traits contributing to isolation (“is-
lands of speciation”; Turner et al. 2005). Reduced rates of recombi-
nation for rearrangements decreased the permeability of the result-
ing linked blocks of interacting genes contained within them to in-
trogress. Hence, these barriers to gene flow are maintained despite
genetic exchange, forming a foundation for possible speciation.
Several questions still need to be resolved concerning the
transvolcanic EVTM, SMO, and U.S. hawthorn flies and the dif-
ferential introgression hypothesis. Reciprocal crosses are needed
to determine whether EVTM, SMO, and U.S. flies can currently
hybridize (an indicator of the potential for gene flow past and
present), whether any nonhost related barriers to gene flow now
exist among the flies, and whether these populations represent
different species. The pattern of genetic variation suggests that if
such nonhost reproductive barriers exist, then they are likely to
be most pronounced between EVTM versus SMO and U.S. flies.
It would be interesting to determine whether the genes respon-
sible for any observed prezygotic isolation or inherent genomic
incompatibilities disproportionately map within the inversions on
chromosomes 1–3, as predicted by chromosomal speciation mod-
els. The paleobiology of Mexico and the United States must also
be further investigated to determine whether the distributions of
co-occurring fauna and flora, as well as environmental conditions,
are consistent with the historical hypothesis for introgression pro-
posed for R. pomonella.
In conclusion, our study underscores the need for a fully
resolved biogeography to study speciation, even in cases when
the proximate causes of divergence appear to be sympatric and
ecological in nature. We are not arguing to abandon the terms
sympatry or allopatry. Rather, we are advocating thinking beyond a
simple sympatric versus allopatric dichotomy for speciation mode.
The factors contributing to divergence may arise and evolve under
a variety of different geographic circumstances that do not always
neatly fit into sympatric versus allopatric categories.
ACKNOWLEGMENTSThe authors would like to thank S. Berlocher, A. Birke, O. Brunel,H. Dambroski, K. Filchak, D. Garcio, L. Guillne, and Z. Wang for assis-tance and moral support, as well as two anonymous reviewers for construc-tive criticisms and insightful comments for improving the manuscript. Inaddition, we are grateful for M. Pale’s help in collecting flies from Mexico.This research was supported by grants to JLF from the National ScienceFoundation, the U.S. Department of Agriculture and the State of Indiana21 Century Fund and to MA and JR from the Mexican Campana Na-cional Contra Moscas de la Fruta and CONACyT convenio 1100/596/04c-837-04.
LITERATURE CITEDBerlocher, S. H., B. A. McPheron, J. L. Feder, and G. L. Bush. 1993. A
revised phylogeny of the Rhagoletis pomonella (Diptera: Tephritidae)sibling species group. Ann. Entomol. Soc. Am. 86:716–727.
Berlocher, S. H. 2000. Radiation and divergence in the Rhagoletispomonella species group: inferences from allozymes. Evolution 54:543–557.
Bush, G. L. 1966. The taxonomy, cytology, and evolution of the genus Rhago-letis in North America (Diptera: Tephritidae). Bull. Mus. Comp. Zool.134:431–562.
———. 1969. Mating behavior, host specificity, and the ecological signif-icance of sibling species in frugivorous flies of the genus Rhagoletis(Diptera: Tephritidae). Am. Nat. 103:669–672.
Charlesworth, B. 1974. Inversion polymorphism in a two-locus genetic system.Genet. Res. 23:259–280.
Coyne, J. A., and H. A. Orr. 2004. Speciation. Sinauer Associates, Sunderland,MA, Pp. 545.
Feder, J. L., C. A. Chilcote, and G. L. Bush. 1988. Genetic differentia-tion between sympatric host races of the apple maggot fly Rhagoletispomonella. Nature 336:61–64.
Feder, J. L., and G. L. Bush. 1989. Gene frequency clines for host races ofRhagoletis pomonella in the midwestern United States. Heredity 63:245–266.
———. 1990. The geographic pattern of genetic differentiation betweenhost associated populations of Rhagoletis pomonella (Diptera: Tephri-tidae) in the eastern United States and Canada. Evolution 44:570–594.
Feder, J. L., T. A. Hunt, and G. L. Bush. 1993. The effects of climate, hostplant phenology and host fidelity on the genetics of apple and hawthorninfesting races of Rhagoletis pomonella. Entomol. Exp. Appl. 69:117–135.
Feder, J. L., J. B. Roethele, B. Wlazlo, and S. H. Berlocher. 1997a. Selectivemaintenance of allozyme differences between sympatric host races ofthe apple maggot fly. Proc. Natl. Acad. Sci. 94:11417–11421.
Feder, J. L., U. Stolz, K. M. Lewis, W. Perry, J. B. Roethele, and A.Rogers. 1997b. The effects of winter length on the genetics of apple andhawthorn races of Rhagoletis pomonella (Diptera: Tephritidae). Evolu-tion 51:1862–1876.
Feder, J. L., S. J. Berlocher, J. B. Roethele, H. Dambroski, J. J. Smith,W. L. Perry, V. Gavrilovic, K. E. Filchak, J. Rull, and M. Aluja.2003a. Allopatric genetic origins for sympatric host-plant shifts andrace formation in Rhagoletis. Proc. Natl. Acad. Sci. USA 100:10314–10319.
Feder, J. L., J. B. Roethele, K. E. Filchak, J. Niedbalksi, and J. Romero-Severson. 2003b. Evidence for inversion polymorphism related to sym-patric host race formation in the apple maggot fly, Rhagoletis pomonella.Genetics 163:939–953.
Feder, J. L., X. Xie, J. Rull, S. Velez, A. Forbes, H. Dambroski, K. Filchak, andM. Aluja. 2005. Mayr, Dobzhansky, Bush and the complexities of sym-patric speciation in Rhagoletis. Proc. Natl. Acad. Sci. USA 102:6573–6580.
Felsenstein, J. 1989. PHYLIP—phylogeny inference package. Cladistics5:164–166.
Filchak, K. E., J. B. Roethele, and J. L. Feder. 2000. Natural selection andsympatric divergence in the apple maggot, Rhagoletis pomonella. Nature407:739–742.
Hudson, R. R., and N. L. Kaplan. 1985. Statistical properties of the numberof recombination events in the history of a sample of DNA sequences.Genetics 111:147–164.
Kirkpatrick, M., and N. Barton. 2006. Chromosomal inversions, local adapta-tion, and speciation. Genetics 173:419–434.
1104 EVOLUTION MAY 2007

MEXICAN RHAGOLETIS
Kumar, S., K. Tamura, and M. Nei. 2004. MEGA3: integrated software formolecular evolutionary genetics analysis and sequence alignment. Brief.Bioinformatics 5:150–163.
Machado, C. A., R. M. Kliman, J. A. Markert, and J. Hey. 2002. Infer-ring the history of speciation from multilocus sequence data: the caseof Drosophila pseudoobscura and its close relatives. Mol. Biol. Evol.19:472–488.
Mallet, J. 2005. Speciation in the 21st century. Review of “Speciation”, byJerry A. Coyne and H. Allen Orr. Heredity 95:105–109.
Marshall, C. J., and J. K. Liebherr. 2000. Cladistic biogeography of the Mex-ican transition zone. J. Biogeogr. 27:203–216.
McPheron, B. A., D. C. Smith, and S. H. Berlocher. 1988. Genetic dif-ferences between host races of Rhagoletis pomonella. Nature 336:64–66.
Navarro, A., and N. H. Barton. 2003. Chromosomal speciation and moleculardivergence-accelerated evolution in rearranged chromosomes. Science300:321–324.
Noor, M. A. F., K. L. Grams, L. A. Bertucci, and J. Reiland. 2001a. Chromo-somal inversions and the reproductive isolation of species. Proc. Natl.Acad. Sci. USA 98:12084–12088.
Noor, M. A. F., K. L. Grams, L. A. Bertucci, Y. Almendarez, J. Reiland, and K.R. Smith. 2001b. The genetics of reproductive isolation and the potentialfor gene exchange between Drosophila pseudoobscura and D. persimilisvia backcross hybrid males. Evolution 55:512–521.
Osada, N., and C.-I. Wu. 2005. Inferring the mode of speciation from genomicdata: a study of the great apes. Genetics 169:259–264.
Payseur, B. A., and M. W. Nachman. 2005. The genomics of speciation: inves-tigating the molecular correlates of X chromosome introgression acrossthe hybrid zone between Mus domesticus and Mus musculus. Biol. J.Linn. Soc. 84:523–534.
Posada, D., and K. A. Crandall. 1998. MODELTEST: testing the model ofDNA substitution. Bioinformatics 14:817–818.
Rieseberg, L. H. 2001. Chromosomal rearrangements and speciation. TrendsEcol. Evol. 16:351–358.
Rieseberg, L. H., J. Whitton, and K. Gardner. 1999. Hybrid zones and thegenetic architecture of a barrier to gene flow between two sunflowerspecies. Genetics 152:713–727.
Roethele, J. B., J. Romero-Severson, and J. L. Feder. 2001. Evidence forbroad-scale conservation of linkage map relationships between Rhago-letis pomonella and Drosophila melanogaster. Ann. Entomol. Soc. Am.94:936–947.
Rogers, S. M., and L. Bernatchez. 2005. Integrating QTL mapping and genomescans towards the characterization of candidate loci under parallel selec-tion in the lake whitefish (Coregonus clupeaformis). Mol. Ecol. 14:351–361.
Rull, J., M. Aluja, J. L. Feder, and S. H. Berlocher. 2006. The distributionand host range of hawthorn-infesting Rhagoletis pomonella (Diptera:Tephritidae) in Mexico. Ann. Entomol. Soc. Am. 99:662–672.
Saitou, N., and M. Nei. 1987. The neighbor-joining method: a new method forreconstructing phylogenetic trees. Mol. Biol. Evol. 4:406–425.
Schneider, S., D. Roessli, and L. Excoffier. 2000. Arlequin ver. 2.000: asoftware for population genetics data analysis. http://anthro.unige.ch/arlequin
Standley, P. C., and J. A. Steyermark. 1946. Flora of Guatemala. Fieldiana:botany. 24: IV. Chicago Natural History Museum, Chicago, IL.
Stump, A. D., M. C. Fitzpatrick, N. F. Lobo, S. Traore, N. F. Sagnon, C.Costantini, F. H. Collins, and N. J. Besansky. 2005. Centromere-proximaldifferentiation and speciation in Anopheles gambiae. Proc. Natl. Acad.Sci. 102:15930–15935.
Swofford, D. L. 2002. PAUP∗: phylogenetic analysis using parsimony (∗andother methods), ver. 4.0b10. Sinauer Associates, Sunderland, MA.
Turner, T. L., M. W. Hahn, and S. V. Nuzhdin. 2005. Genomic islands ofspeciation in Anopheles gambiae. PLoS Biol 3:1572–1578.
Wang, R. L., J. Wakeley, and J. Hey. 1997. Gene flow and natural selectionin the origin of Drosophila pseudoobscura and close relatives. Genetics147:1091–1106.
Associate Editor: M. Peterson
Supplementary MaterialThe following supplementary material is available for this article:
Figures S1–S16.
This material is available as part of the online article from:
http://www.blackwell-synergy.com/doi/abs/10.1111/j.1558-5646.2007.00091.x
(This link will take you to the article abstract).
Please note: Blackwell Publishing is not responsible for the content or functionality of any supplementary materials supplied by
the authors. Any queries (other than missing material) should be directed to the corresponding author for the article.
EVOLUTION MAY 2007 1105















