Embed Size (px)
Citation preview

JOBNAME: JIP 68#1 96 PAGE: 1 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
The Structure and Behavior of Gypsy Moth (Lymantriadispar) Hemocytes
TARIQ M. BUTT* AND KATHLEEN S. SHIELDS†
*Entomology and Nematology Department, IACR-Rothamsted, Harpenden, Herts, AL5 2JQ, United Kingdom; and †USDA Forest Service,Northeastern Forest Experiment Station, Northeastern Center for Forest Health Research, 51 Mill Pond Road, Hamden, Connecticut 06514
Received July 6, 1995; accepted January 30, 1996
Light and electronmicroscope observations of gypsymoth larval hemocytes show seven distinct andreadily identifiable types of circulating cells: prohe-mocytes, granulocytes, coagulocytes, adipohemocytes,plasmatocytes, oenocytoids, and spherulocytes. In ad-dition to these seven, a large, motile, highly refractivecell was observed with the light microscope, but littlewas elucidated about its ontogeny or function. Plas-matocytes, granulocytes, and coagulocytes consti-tuted the largest portion of the hemocyte population.Plasmatocytes were distinguished by their largespindle shape and large dumbbell-shaped nuclei.These cells were motile and rapidly spread out on aglass microscope slide. Granulocytes characteristi-cally contained secretory granules with a microtubu-lar–filamentous substructure. They also containedvacuoles, microtubules, and substantial rough endo-plasmic reticulum. Both plasmatocytes and granulo-cytes were phagocytic. The fragile coagulocytes weredistinguished by the dilated perinuclear space, ab-sence of granules, and by ejection of the nucleus fromthe cell. Spherulocytes contained refractive, mem-brane-bound inclusions with a crystal-like substruc-ture and a central nucleus. Oenocytoids were typicallylarge, with smooth, hyaline cytoplasm and a tiny ec-centric nucleus. These particular cells increased innumber just before and during ecdysis. Adipohemo-cytes contained numerous lipid globules of differentsizes; these cells were particularly conspicuous in lateinstar larvae and pupae. Only the granulocytes andcoagulocytes appeared to be important in coagulation.During this process the granular–floccular contents ofthe granulocytes were extruded by exocytosis whilesome cells lysed. Coagulocytes were probably the firstto lyse. The coagulum was extremely sticky andtrapped other hemocytes which subsequently lysed.The debris from these cells was a component of thecoagulum which eventually became melanized. © 1996
Academic Press, Inc.
KEY WORDS: Hemocytes; ultrastructure; behavior; co-agulation; gypsy moth.
INTRODUCTION
Insect hemocytes are involved in wound repair, co-agulation, melanization, and immobilization of invad-ing organisms by encapsulation and/or phagocytosis(Ratcliffe and Rowley, 1979; Ratcliffe, 1993; Pech et al.,1995). In a recent light microscope study, gypsy mothplasmatocytes and coagulocytes (4 cystocytes) playedan active role in the cellular immune response to natu-ral protoplasts of the entomophthoraceous fungus En-tomophaga spp. (Butt and Humber, 1989). Plasmocytesand coagulocytes were reported to constitute over 90%of the hemocyte population (Butt and Humber, 1989)but accurate identification was precluded by the lim-ited resolution of the light microscope.It is clear from a review of the literature that there is
considerable confusion regarding the classification ofinsect hemocytes (e.g., Ravindranath, 1978; Gupta,1979, 1986). This problem is particularly true forgranulocytes (4 granular cells), and undifferentiatedcoagulocytes which are often confused with each other,and occasionally with other hemocyte types (Gregoire,1970; Gregoire and Goffinet, 1979; Gupta, 1979, 1986;Brehelin and Zachary, 1983, 1986; Russo et al., 1993).Because these cells are important components of theinsect immune system it is imperative that they shouldbe correctly identified and their functions understood.The aim of this study was to identify the different he-mocytes of gypsy moth larvae on the basis of their finestructure. Detailed observations were also made of thebehavior of hemocytes during coagulation to determinewhich cells are involved in this important event.
MATERIALS AND METHODS
Insects
Gypsy moth larvae were obtained from the U.S. For-est Service Insect Rearing Facility, (Hamden, CT). Alllarvae were of the New Jersey Standard Strain, rearedon a high wheat germ diet (Bell et al., 1981), at 27°C,60% RH, and 16:8 L:D photoperiod.
JOURNAL OF INVERTEBRATE PATHOLOGY 68, 1–14 (1996)ARTICLE NO. 0052
1
0022-2011/96 $18.00Copyright © 1996 by Academic Press, Inc.
All rights of reproduction in any form reserved.

JOBNAME: JIP 68#1 96 PAGE: 2 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
Light and Fluorescence Microscopy
Hemolymph samples were collected from 10 (5 male,5 female) second, third, fourth, and fifth instar larvaeapproximately 48hr postmolt, using the methods ofButt and Humber (1989). Samples were taken from anadditional 10 larvae at the “head slippage” stage ofeach instar (indicating a molt is imminent), and bloodwas also collected from 10 prepupae and ten 1- to2-day-old pupae by piercing the side of the insect andcollecting hemolymph on a microscope slide. Two he-molymph samples were collected from each insect. Onesample was observed under phase contrast, differentialinterference, and/or Hoffman modulation contrast,while acetoorcein was added to the other to stain nu-clei. A minimum of 100 cells from each sample wascharacterized, and the percentage of each hemocytetype was recorded. Additional samples were stained forsugars and lipids using 0.1% (w/v) Uvitex-BOPT (Ciba-Geigy)1 and 3 mg/ml Nile red (Kodak), respectively, andexamined by epifluorescence microscopy using the ap-propriate FITC and rhodamine filter combinations(Butt et al., 1989).
Scanning Electron Microscopy
Hemolymph was collected on a clean glass coverslipand the hemocytes were allowed to settle for 5 min. Thecells were fixed by immersing the coverslip in chilled3% (v/v) glutaraldehyde in 0.1 M sodium cacodylate, 2mM Ca2+ buffer, pH 7.2 for 15 min. After washing inbuffer for 5 min, they were postfixed in 1% (w/v) buff-ered osmium tetroxide, washed as before, then dehy-drated through a graded ethanol series, and criticalpoint dried in a Sorvall critical point drying system.Cells were coated with gold and examined in an ETECAutoscan scanning electron microscope at 10 kV.
Transmission Electron Microscopy
Method A. Hemolymph from four groups of 10 ran-domly selected fourth/fifth instar larvae was collected
in Eppendorf vials kept at 4°C. Samples were fixed in3% buffered (0.1 M sodium cacodylate, pH 7.2) and un-buffered glutaraldehyde, 4 and 15 min after pooling theblood. The cells were harvested in an Eppendorf 5415,washed five times with buffer, and the pellet was re-suspended in 2% (w/v) SeaPrep Agarose (FMC Corp).After setting at 4°C, the agar was cut into 3-mm cubes,fixed in osmium tetroxide, and stained en bloc withuranyl acetate according to Butt and Beckett (1984).Thin sections were stained with lead citrate and uranylacetate and examined in a Zeiss EM 10 or Philips EM300 transmission electron microscope at 60 kV.
Method B. Hemolymph collected from three repli-cates of 10 larvae (5 male + 5 female) at mid and latethird, fourth, and fifth instar was pooled and processedas above except for the following: operations were con-ducted at 4°C, the agarose step was omitted, ethanoland propylene oxide were used for dehydration, andmaterial was embedded in Epon 812 or Poly/Bed 812.
Studies on the Coagulation of Gypsy Moth Blood
Hemolymph samples taken from third and fourth in-star larvae were allowed to coagulate on a glass micro-scope slide or in an Eppendorf vial for periods rangingfrom 5 sec to 45 min prior to being examined by light/fluorescence microscopy or processed for transmissionelectron microscopy, as described above.
RESULTS
General Observation
Seven types of hemocyte were identified in thin sec-tions: oenocytoids, prohemocytes, granulocytes, spher-ulocytes, coagulocytes, plasmatocytes, and adipohemo-cytes (Figs. 1–17). However, an additional hemocytewas observed with the light microscope which was usu-ally larger than plasmatocytes and much more refrac-tive (Fig. 1). Most differentiated or mature hemocytescould be readily identified by light microscopy but itwas difficult to distinguish between prohemocytes andimmature or undifferentiated plasmatocytes, and be-tween undifferentiated coagulocytes and other cells.
1 Mention of firms and/or proprietary product names does not con-stitute endorsement by the United States Department of Agricul-ture.
TABLE 1The Proportion (%) of Different Hemocyte Types at Different Developmental Stages of Gypsy Moth Larvae and Pupae
(n 4 10, ±SD)
Stage Plasmatocytesa Granulocytes Coagulocytes Sphereulocytes Oenocytoids Adipohemocytes
III 46.4 (11.3) 34.9 (11.6) 13.6 (4.3) 2.3 (1.7) 2.7 (0.8) <1.0III–IV 39.3 (8.8) 25.9 (10.4) 16.2 (6.5) 10.9 (5.3) 7.6 (4.2) <1.0IV 38.0 (6.0) 39.8 (6.4) 14.6 (4.4) 4.3 (2.2) 3.3 (0.9) <1.0IV–V 33.6 (9.7) 36.2 (9.8) 13.5 (4.1) 4.0 (2.4) 10.5 (3.3) 2.1 (1.8)V 37.6 (7.9) 41.3 (6.8) 12.2 (4.6) 1.4 (1.5) 3.5 (0.9) 4.0 (1.3)Prepupae 31.3 (12.1) 31.6 (11.8) 2.3 (1.3) 1.5 (1.4) 21.7 (4.8) 11.5 (4.3)Pupae 52.1 (10.1) 5.6 (4.2) 1.0 (1.1) <1.0 2.4 (1.6) 38.8 (9.7)
a Includes prohemocytes.
BUTT AND SHIELDS2
JOBNAME: JIP 68#1 96 PAGE: 3 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
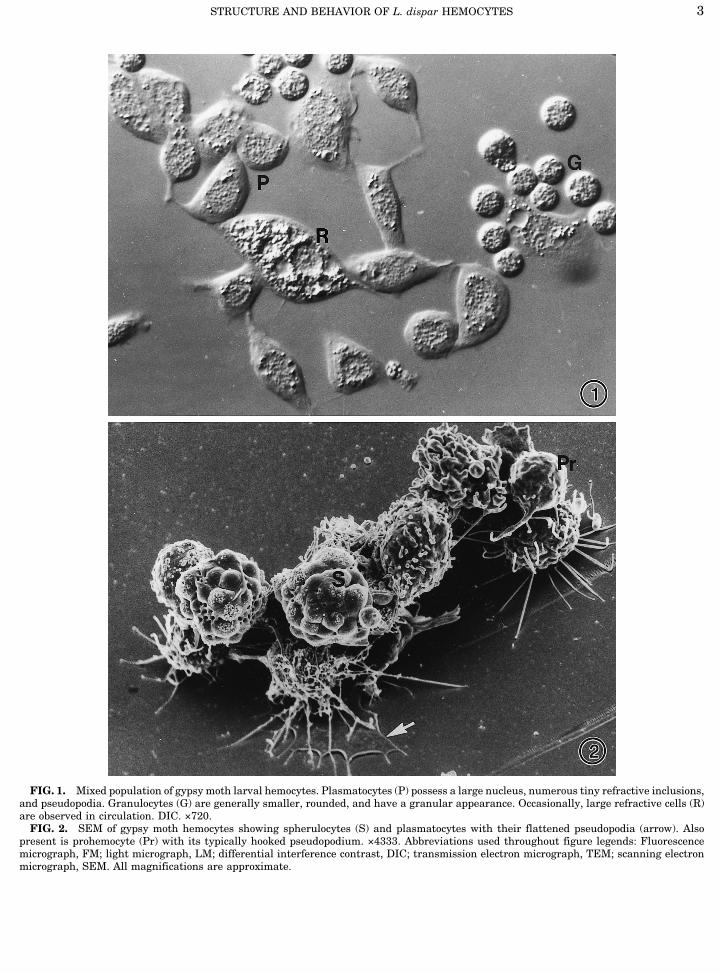
FIG. 1. Mixed population of gypsy moth larval hemocytes. Plasmatocytes (P) possess a large nucleus, numerous tiny refractive inclusions,and pseudopodia. Granulocytes (G) are generally smaller, rounded, and have a granular appearance. Occasionally, large refractive cells (R)are observed in circulation. DIC. ×720.FIG. 2. SEM of gypsy moth hemocytes showing spherulocytes (S) and plasmatocytes with their flattened pseudopodia (arrow). Also
present is prohemocyte (Pr) with its typically hooked pseudopodium. ×4333. Abbreviations used throughout figure legends: Fluorescencemicrograph, FM; light micrograph, LM; differential interference contrast, DIC; transmission electron micrograph, TEM; scanning electronmicrograph, SEM. All magnifications are approximate.
STRUCTURE AND BEHAVIOR OF L. dispar HEMOCYTES 3
JOBNAME: JIP 68#1 96 PAGE: 4 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
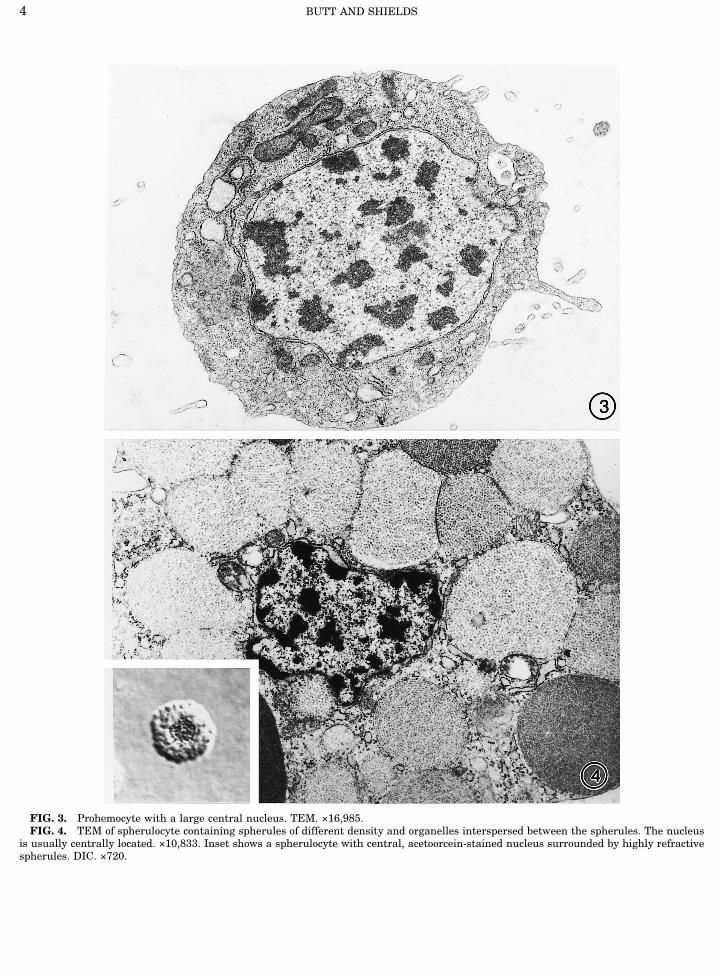
FIG. 3. Prohemocyte with a large central nucleus. TEM. ×16,985.FIG. 4. TEM of spherulocyte containing spherules of different density and organelles interspersed between the spherules. The nucleus
is usually centrally located. ×10,833. Inset shows a spherulocyte with central, acetoorcein-stained nucleus surrounded by highly refractivespherules. DIC. ×720.
BUTT AND SHIELDS4
JOBNAME: JIP 68#1 96 PAGE: 5 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
The numbers of different types of hemocyte in larvaeand pupae is summarized in Table 1.
Prohemocytes
These cells were typically round or oval of variablesize (10–14 × 8–12 mm), containing dense cytoplasm
and a large central nucleus (Fig. 3). The nucleus tocytoplasm ratio was high, which helped differentiatethese cells from the plasmatocytes, granulocytes, andcoagulocytes. The plasma membrane was generallysmooth, with only a few protuberances. Prohemocytesnormally produced a single, hyaline, retractable pseu-dopodium which was bent at the apex (Figs. 2 and 3).
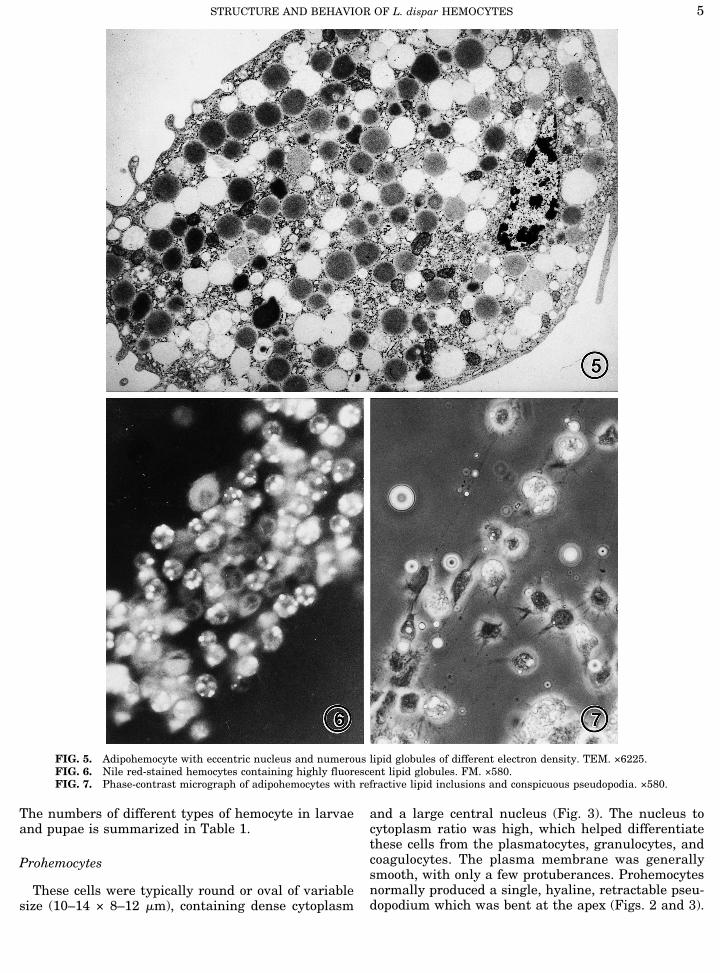
FIG. 5. Adipohemocyte with eccentric nucleus and numerous lipid globules of different electron density. TEM. ×6225.FIG. 6. Nile red-stained hemocytes containing highly fluorescent lipid globules. FM. ×580.FIG. 7. Phase-contrast micrograph of adipohemocytes with refractive lipid inclusions and conspicuous pseudopodia. ×580.
STRUCTURE AND BEHAVIOR OF L. dispar HEMOCYTES 5
JOBNAME: JIP 68#1 96 PAGE: 6 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
The cytoplasm hadmany free ribosomes, fewmitochon-dria, and sparse endoplasmic reticulum (ER). In thelight microscope, some cells with the dimensions andappearance of prohemocytes quickly differentiated intoreactive coagulocytes and recognizable granulocytes,
suggesting that the prohemocyte was the progenitor ofthese cell types. Prohemocytes constituted less than1.0% of the cells observed with light microscopy, andonly one obvious prohemocyte was found in thin sec-tions (Fig. 3) in spite of extensive examination of nu-
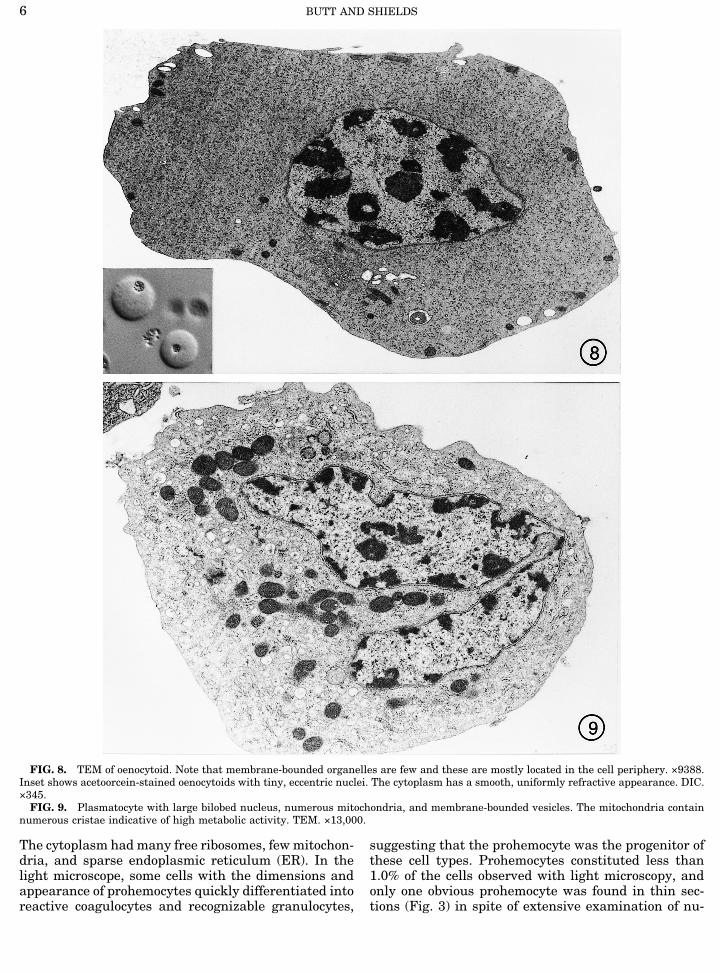
FIG. 8. TEM of oenocytoid. Note that membrane-bounded organelles are few and these are mostly located in the cell periphery. ×9388.Inset shows acetoorcein-stained oenocytoids with tiny, eccentric nuclei. The cytoplasm has a smooth, uniformly refractive appearance. DIC.×345.FIG. 9. Plasmatocyte with large bilobed nucleus, numerous mitochondria, and membrane-bounded vesicles. The mitochondria contain
numerous cristae indicative of high metabolic activity. TEM. ×13,000.
BUTT AND SHIELDS6
JOBNAME: JIP 68#1 96 PAGE: 7 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
merous samples, suggesting that the transformation ofthese cells must be rapid.
Spherulocytes
These round–ovoid cells varied in size (9–13 × 11–19mm) and were readily distinguished by the numerous
refractive inclusions and comparatively small (3–4 mmdiameter) centrally located nucleus (Fig. 4, inset). Thespherules were responsible for the punctate appear-ance of the cells. Few mitotic cells were observed sug-gesting a slow turnover of these cells. Spherules variedin shape and size but were usually spherical, triangu-
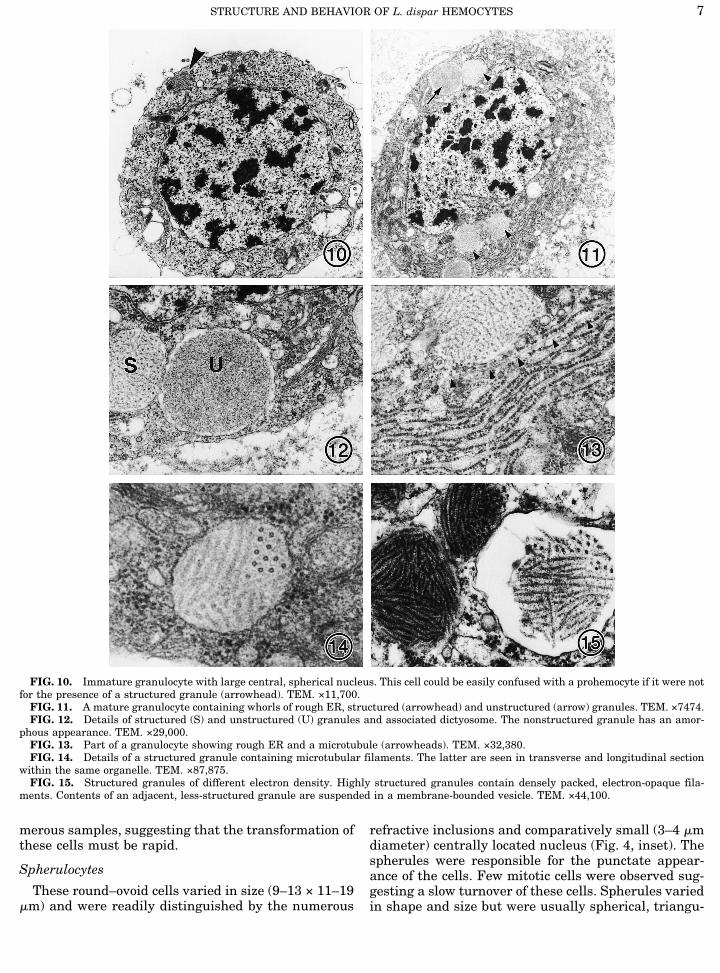
FIG. 10. Immature granulocyte with large central, spherical nucleus. This cell could be easily confused with a prohemocyte if it were notfor the presence of a structured granule (arrowhead). TEM. ×11,700.FIG. 11. A mature granulocyte containing whorls of rough ER, structured (arrowhead) and unstructured (arrow) granules. TEM. ×7474.FIG. 12. Details of structured (S) and unstructured (U) granules and associated dictyosome. The nonstructured granule has an amor-
phous appearance. TEM. ×29,000.FIG. 13. Part of a granulocyte showing rough ER and a microtubule (arrowheads). TEM. ×32,380.FIG. 14. Details of a structured granule containing microtubular filaments. The latter are seen in transverse and longitudinal section
within the same organelle. TEM. ×87,875.FIG. 15. Structured granules of different electron density. Highly structured granules contain densely packed, electron-opaque fila-
ments. Contents of an adjacent, less-structured granule are suspended in a membrane-bounded vesicle. TEM. ×44,100.
STRUCTURE AND BEHAVIOR OF L. dispar HEMOCYTES 7
JOBNAME: JIP 68#1 96 PAGE: 8 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
lar, oblong, or ovoid (ca. 1.6–3.2 mm diameter). Thesemembrane-bounded organelles had an internal sub-structure, a crystal-like lattice in concentric layers(Fig. 4). Some spherules were more electron-opaquethan others; this was due to the close packing of thelattice (Fig. 4). Golgi bodies, mitochondria, and roughER were interspersed among the spherules (Fig. 4).Motile spherulocytes produced short pseudopodiawhile sessile cells produced several, extremely fine fili-
podia of various lengths. Approximately 4.5% of larvalhemocytes were spherulocytes, but their numbers de-creased in prepupae (1.5%), and they comprised lessthan 1% of pupal hemocytes (Table 1).
Adipohemocytes
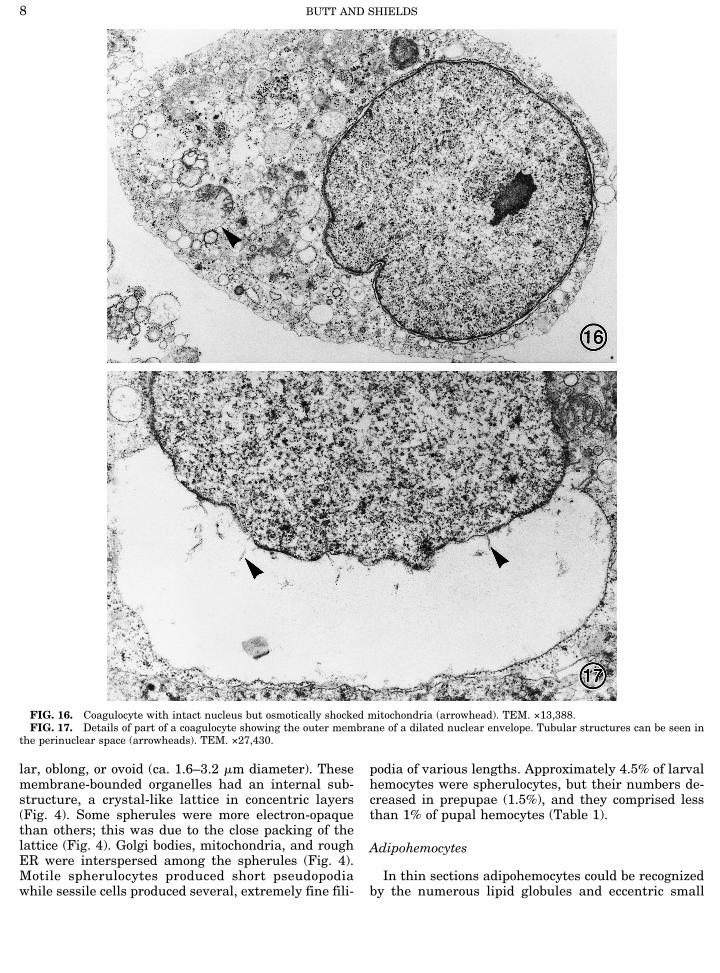
In thin sections adipohemocytes could be recognizedby the numerous lipid globules and eccentric small
FIG. 16. Coagulocyte with intact nucleus but osmotically shocked mitochondria (arrowhead). TEM. ×13,388.FIG. 17. Details of part of a coagulocyte showing the outer membrane of a dilated nuclear envelope. Tubular structures can be seen in
the perinuclear space (arrowheads). TEM. ×27,430.
BUTT AND SHIELDS8

JOBNAME: JIP 68#1 96 PAGE: 9 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
nucleus (Fig. 5). Interspersed among the globules wereGolgi bodies, short smooth cisternae, microtubules(MTs), and mitochondria. Fluorescence microscopy us-ing the lipid stain Nile red confirmed the presence oflipid globules in these cells (Fig. 6). Lipid staining inother hemocyte types was uniform and faint; they didnot contain prominent globules. Adipohemocytes fromsecond to fourth instar larvae contained one to threelipid globules per cell, while those from fifth instar lar-vae and especially from prepupae and pupae containednumerous large globules (not illustrated). Adipohemo-cytes were motile and produced pseudopodia and fili-podia (Fig. 7). Adipohemocytes comprised less than 1%of hemocytes in third and fourth instar larvae, buttheir numbers increased in the fifth instar (4.0%), inprepupae (11.5%), and especially in pupae (38.8%).
Oenocytoids
These relatively large cells (30–35 mm) rapidly be-came spherical on microscope slides. They typicallypossessed smooth, homogeneous cytoplasm, sparse or-ganelles, and a tiny eccentric nucleus (Fig. 8, inset).Only the nucleus could be identified with the light mi-croscope, but in thin sections tiny mitochondria and afew membrane-bound vesicles and cisternae that werelocated predominantly in the cell periphery were ob-served (Fig. 8). Clusters of polyribosomes were foundthroughout the cytoplasm. Approximately 3% of hemo-cytes from midinstar larvae were oenocytoids, butthere was a conspicuous increase in numbers of thiscell type during ecdysis. Oenocytoids represented 7.6%of the hemocyte population at the conclusion of instarIII, 10.5% at the conclusion of IV, and 21.7% in prepu-pae (Table 1).
Plasmatocytes
These motile, pleomorphic cells constituted 39% ofthe total hemocyte population in larvae and 52.1% inprepupae (Figs. 1 and 9). Plasmatocytes were observedto phagocytose cell debris and have been reported tophagocytose microorganisms in the hemolymph and toencapsulate foreign cells (Butt and Humber, 1989).Plasmatocytes varied in size, and these cells were prob-ably at various stages of maturation. In freshly isolatedhemolymph, they were mostly spindle-shaped (8–20 ×5–9 mm), but a few were oval or spherical (8–20 mmdiameter). The large central nucleus (5–9 × 3–6 mm)was often bilobed or fusiform and occasionally spheri-cal. Nuclei contained numerous patches of electron-opaque chromatin (Fig. 9). Mitotic nuclei were ob-served in all preparations (not illustrated).Plasmatocytes contained dense cytoplasm, several
Golgi bodies, numerous mitochondria, free ribosomes,membrane-bounded vesicles, and phagocytic vacuoles(Fig. 9). Cisternae of both smooth and rough ER weredispersed throughout the cytoplasm but were rarely
arranged in stacks. Plasmatocytes lacked any electron-opaque or 0structured0 granules but possessed numer-ous MTs which were present in the cell cortex inter-spersed among the cisternae of rough ER and in pseu-dopodia.The pseudopodia of plasmatocytes readily spread
over the substratum forming a broad, thin, clear mem-brane. From the ends of this flat membrane fine filipo-dial extensions were produced (Fig. 2). The pseudopo-dium not only firmly adhered the cell to the substra-tum but also played an active role in cytokinesis andphagocytosis.
Granulocytes
In the light microscope, granulocytes appeared asmotile, spherical cells (7–9 mm diameter), with aprominent central nucleus (3–6 mm diameter) whichcontained several patches of heterochromatin (Fig. 1).These cells contained few to numerous refractive inclu-sions. On glass slides they produced one to two broadhyaline pseudopodia or lamellipodia which encircledthe cell. Granulocytes contained few to numerous re-fractive inclusions. They were more stable than coagu-locytes and the perinuclear space did not dilate. Thinsections revealed that granulocytes were the only he-mocytes to possess granules (Figs. 10 and 11). The con-tents of mature granulocytes included: Golgi bodies,multivesicular bodies, dense cytoplasm, MTs, exten-sive rough ER with most cisternae arranged in stacks,numerous mitochondria, membrane-bounded vesicles,and smooth ER (Figs. 11–13).Two types of membrane-bounded granules were ob-
served. The first, which we refer to as structured gran-ules, contained electron-opaque filaments with a fuzzycoating (Figs. 12–15). The filaments varied in lengthbut in cross section resembled MTs with a diameter of17–20 nm. The microtubular filaments of the struc-tured granules were arranged in bundles (Fig. 14).Each granule contained one or more of these bundleswhich were stacked at different angles. The contents ofthe structured granule varied in opacity and were oftensuspended in the vesicle (Fig. 15). The second type ofgranule we refer to as the unstructured granule (Fig.12). These organelles contained a homogenous, elec-tron-opaque, granular-flocculent material which wasexpelled from the cell. The contents of structured gran-ules were never discharged. Very few granules wereobserved containing both the fuzzy, microtubular ele-ments as well as the flocculent material, suggestingthat one might convert into the other rapidly. Bothgranules varied in size but the unstructured granuleswere generally larger than the structured ones, prob-ably due to hydration and swelling. In a particular cell,the diameter of structured and unstructured granulesmeasured 350–840 and 1000 nm, respectively. Distri-bution of these granules varied; some cells possessed
STRUCTURE AND BEHAVIOR OF L. dispar HEMOCYTES 9

JOBNAME: JIP 68#1 96 PAGE: 10 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
only the structured and others only unstructured gran-ules, but most possessed both. Granulocytes comprised35.6% of larval hemocytes, but only 5.6% of pupal he-mocytes (Table 1).
Coagulocytes
Differentiated (i.e., reactive) coagulocytes consti-tuted 14.0% of the hemocytes observed from larvae and1.0% of hemocytes from pupae, as determined by lightmicroscopy. The total number of coagulocytes in circu-lation probably was greater, as nonreactive coagulo-cytes could not easily be distinguished from prohemo-cytes and undifferentiated plasmocytes. Although thevast majority of coagulocytes was round (8–19 mm di-ameter), some cells were oblong (14.5 ± 4 mm width, 17± 7.5 mm length) and a few exceeded 40 mm in length.Coagulocytes lacked granules and stacked rough ERbut contained numerous membrane-bounded vesicles(Fig. 16). Based on measurements from electron micro-graphs nuclei of undifferentiated cells were ca. 4 mmdiameter. As observed with the light microscope, nucleiof differentiated cells were round and measured 8.2 ±5.5 mm in diameter (i.e., area enclosed by the innermembrane of the nuclear envelope). In thin sectionsthey appeared to contain less electron-opaque chroma-tin than the other hemocytes (Figs. 16 and 17).During differentiation, these motile and extremely
fragile cells followed two pathways: either the nuclearenvelope dilated and the cells became permeable todyes or they lysed. The former was most common andentailed separation of the outer membrane of thenuclear envelope while the inner membrane remainedrelatively intact. Tubular material was occasionally ob-served in the dilated perinuclear space (Fig. 17).
Behavior of Hemocytes during Coagulation
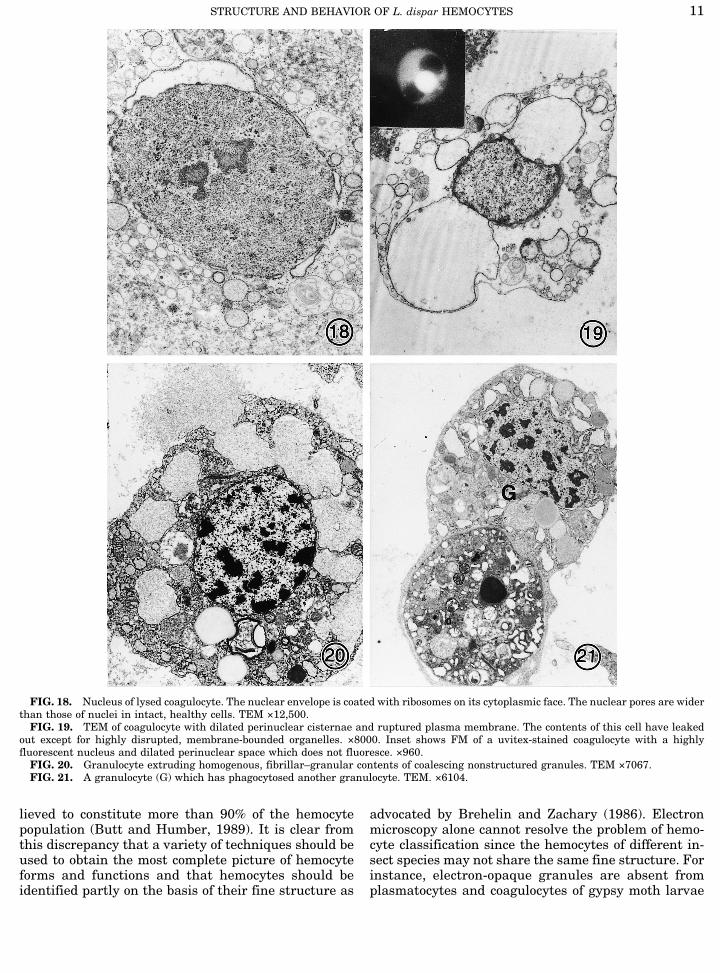
The first cells to undergo visible change during co-agulation were the coagulocytes and granulocytes. Cy-tological changes were observed in coagulocytes withinthe first 30 sec of bleeding, with maximum transfor-mation occurring within 15–30 min. Fluorescence mi-croscopy showed that coagulocytes became rapidly per-meabilized, imbibing water and dyes, and in bright-field they had a prominent outline. These featureshelped distinguish these cells from other hemocytes.In thin sections the cytoplasm of coagulocytes was
less dense than that of other hemocytes, and numerousribosomes coated the surface of the nuclear envelopeand of membrane-bounded vesicles (Fig. 18). The poresof nuclei from lysed cells were often wider than thosefrom intact cells (compare Figs. 16 and 18). However,in some ruptured coagulocytes nuclear pores were notobserved (Fig. 19). Permeabilized coagulocytes stainedwith the fluorochrome Uvitex, which binds to b-glu-cans (Fig. 19, inset). Staining of the nucleus was gen-erally more intense than that of the cytoplasm, while
the perinuclear space did not stain at all (Fig. 19 in-sert). Dilation of the perinuclear space resulted in theouter membrane becoming detached and expandingwithout any apparent changes being detected in theinner membrane (Figs. 17–19). Some nuclei were ex-truded from the cell without damaging the plasmamembrane, presumably through the uncontrolled ex-pansion of the perinuclear space. It was not uncommonfor some coagulocytes to lyse spontaneously, with orwithout dilation of the nuclear envelope. Cytoplasmfrom the lysed cells contributed toward the coagulum.Two patterns of behavior were observed in granulo-
cytes during coagulation. The first activity entaileddischarge of flocculent coagulum by exocytosis (Fig.20). The lobed appearance of the normally spherical,granular inclusions suggested fusion of these organ-elles during exocytosis (Fig. 20). The second patterninvolved cell lysis. The chromatin of lysing cells wasless electron-opaque than that of intact cells. In addi-tion, some granulocytes phagocytosed cell debris andother hemocytes, including other granulocytes (Fig.21). Phagocytic vacuoles contained organelles fromlysed hemocytes, including granules from granulo-cytes.Lysis of granulocytes and coagulocytes always initi-
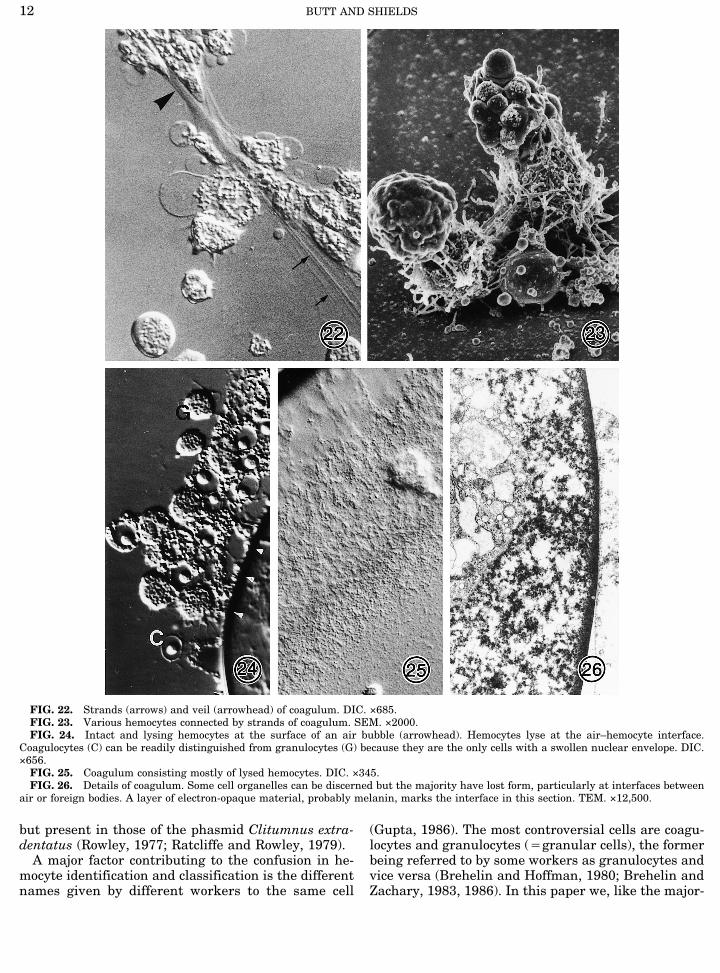
ated changes in other hemocytes including intact co-agulocytes and granulocytes. These changes includedextension of filipodia and strands of coagulum, en-hanced phagocytosis in plasmatocytes and granulo-cytes, and lysis of hemocytes that became entrapped inthe coagulum (Figs. 22 and 24–26). Hemocytes ap-peared to become sticky and readily adhered to eachother (Figs. 22–24). The filipodial and/or coagulumstrands often fused to form a veil emanating from orsurrounding hemocytes (Fig. 22).Cells at the air–hemolymph interface were the first
to lyse but a chain reaction occurred resulting in lysisof adjacent cells (Figs. 24 and 25). This pattern wasrepeated at the surface of debris which was not of he-mocoel origin. Cell lysis was normally limited to a nar-row zone, i.e., it was not extensive. Ultimately, the co-agulum became melanized (Figs. 25 and 26).
DISCUSSION
Hemocyte Classification
Gypsy moth larvae possess eight different types ofhemocyte, whereas in other Lepidoptera only five arefrequently reported; these include plasmatocytes,granulocytes, spherulocytes, oenocytoids, and prohe-mocytes (e.g., Akai and Sato, 1973; Beaulaton andMonpeyssin, 1976; Essawy et al., 1985; Raina, 1976).Approximately 90% of the gypsy moth larval hemo-
cyte population was composed of plasmatocytes, granu-locytes, and coagulocytes. In an earlier light micro-scope study plasmatocytes and coagulocytes were be-
BUTT AND SHIELDS10
JOBNAME: JIP 68#1 96 PAGE: 11 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
lieved to constitute more than 90% of the hemocytepopulation (Butt and Humber, 1989). It is clear fromthis discrepancy that a variety of techniques should beused to obtain the most complete picture of hemocyteforms and functions and that hemocytes should beidentified partly on the basis of their fine structure as
advocated by Brehelin and Zachary (1986). Electronmicroscopy alone cannot resolve the problem of hemo-cyte classification since the hemocytes of different in-sect species may not share the same fine structure. Forinstance, electron-opaque granules are absent fromplasmatocytes and coagulocytes of gypsy moth larvae
FIG. 18. Nucleus of lysed coagulocyte. The nuclear envelope is coated with ribosomes on its cytoplasmic face. The nuclear pores are widerthan those of nuclei in intact, healthy cells. TEM ×12,500.FIG. 19. TEM of coagulocyte with dilated perinuclear cisternae and ruptured plasma membrane. The contents of this cell have leaked
out except for highly disrupted, membrane-bounded organelles. ×8000. Inset shows FM of a uvitex-stained coagulocyte with a highlyfluorescent nucleus and dilated perinuclear space which does not fluoresce. ×960.FIG. 20. Granulocyte extruding homogenous, fibrillar–granular contents of coalescing nonstructured granules. TEM ×7067.FIG. 21. A granulocyte (G) which has phagocytosed another granulocyte. TEM. ×6104.
STRUCTURE AND BEHAVIOR OF L. dispar HEMOCYTES 11
JOBNAME: JIP 68#1 96 PAGE: 12 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
but present in those of the phasmid Clitumnus extra-dentatus (Rowley, 1977; Ratcliffe and Rowley, 1979).A major factor contributing to the confusion in he-
mocyte identification and classification is the differentnames given by different workers to the same cell
(Gupta, 1986). The most controversial cells are coagu-locytes and granulocytes (4granular cells), the formerbeing referred to by some workers as granulocytes andvice versa (Brehelin and Hoffman, 1980; Brehelin andZachary, 1983, 1986). In this paper we, like the major-
FIG. 22. Strands (arrows) and veil (arrowhead) of coagulum. DIC. ×685.FIG. 23. Various hemocytes connected by strands of coagulum. SEM. ×2000.FIG. 24. Intact and lysing hemocytes at the surface of an air bubble (arrowhead). Hemocytes lyse at the air–hemocyte interface.
Coagulocytes (C) can be readily distinguished from granulocytes (G) because they are the only cells with a swollen nuclear envelope. DIC.×656.FIG. 25. Coagulum consisting mostly of lysed hemocytes. DIC. ×345.FIG. 26. Details of coagulum. Some cell organelles can be discerned but the majority have lost form, particularly at interfaces between
air or foreign bodies. A layer of electron-opaque material, probably melanin, marks the interface in this section. TEM. ×12,500.
BUTT AND SHIELDS12

JOBNAME: JIP 68#1 96 PAGE: 13 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
ity of workers, refer to cells containing structuredgranules as granulocytes. Another factor adding to theconfusion is the interpretation of micrographs. For ex-ample, the nuclei of plasmatocytes are often bilobedand if the attenuated portion stains poorly or is onlyevident in serial sections then the plasmatocyte may beinterpreted by some workers as having two nuclei(Gupta, 1986).
Adipohemocytes, Spherulocytes, and Oenocytoids
Adipohemocytes, spherulocytes, and oenocytoids arethe least controversial of hemocyte types. Adipohemo-cytes were conspicuous in late instar larvae and pupaeof L. dispar. It is possible that they assist in the depo-sition of fat reserves needed during the pupal stagewhen the insect is not feeding. However, adipohemo-cytes were less obvious in late instar larvae ofHeliothisarmigera (Essawy et al., 1985), suggesting that this cellhas other functions or is less important for some in-sects. According to Raina (1976) adipohemocytes ofPectinophora gossypiella are derived from granulocytesbut we found no evidence for this in gypsy moth. How-ever, gypsy moth adipohemocytes might represent themature stage of the large, highly refractive cells (Fig.1) that we observed occasionally in light microscopepreparations of larval hemolymph.The spherules of gypsy moth spherulocytes exhibit
an internal substructure, a crystal-like lattice in con-centric layers, similar to that of other Lepidoptera in-cluding Spodoptera littoralis (Harpaz et al. 1969) andHeliothis virescens (Cook et al., 1985). In other speciessuch as Melolontha melolontha (Devauchelle, 1971)and C. extradentatus (Rowley, 1977) the spherules areelectron-opaque without any visible structure.Gypsy moth oenocytoids resemble those of other in-
sects such asGalleria mellonella (Ratcliffe and Rowley,1979) in being large, hyaline cells with a small eccen-tric nucleus. Their numbers increase during ecdysissuggesting they play a key role in this process. Theyare known to contain phenoloxidase (Iwama andAshida, 1986), so perhaps they assist in melanizationof the cuticle.
Coagulocytes versus Granulocytes
The coagulocyte and granulocyte are the mostlywidely distributed hemocyte types reported in inverte-brates, and the most controversial (Gregoire and Gof-finet, 1979; Brehelin and Zachary, 1986; Gupta, 1986).Granulocytes and nonreactive coagulocytes are noteasily distinguished from each other with the light mi-croscope or from prohemocytes or immature plasmato-cytes.The fine structure of granulocytes differs radically
from that of coagulocytes and other gypsy moth hemo-cytes. The granulocyte is the only cell which containsstructured granules and a well-developed endomem-
brane system (rough and smooth ER, Golgi apparatus).The structured granules in gypsy moth granulocytesresemble those in granulocytes of C. extradentatus(Rowley, 1977), Leucophaea maderae (Hagopian,1971), Antheraea pernyi (Beaulaton and Monpeyssin,1976), Locusta migratoria, G. mellonella, and manyother insect species (Brehelin et al., 1978; Brehelin andZachary, 1986; Gupta, 1979, 1986). The unstructuredelectron-opaque granules, the contents of which aredischarged into the hemolymph, probably representthe final stage in the development of the structuredgranule. The fact that unstructured granules arelarger and their contents more diffuse than those ofstructured granules suggests that the microtubularfilaments are dissolved following imbibition of water.Tightly packed bundles of filaments suspended in themembrane-bounded granule also suggest uptake of wa-ter. If the swelling of granules was an artifact of speci-men preparation, we would expect other membrane-bounded organelles in the same cell to be swollen, butthis was not so.The fine structure of undifferentiated gypsy moth
coagulocytes differs markedly from that of the granu-locytes although this is not apparent in the light mi-croscope. Coagulocytes, like those from other insectspecies are hyaline, with a poor to moderately devel-oped endomembrane system and a dilated nuclear en-velope. The latter characteristic distinguishes coagulo-cytes from all the other hemocyte types (Gregoire,1970; Gregoire and Goffinet, 1979). These cells are ex-tremely fragile and are the first to show visible cyto-logical changes upon removal from the insect.
Behavior of Coagulocytes and Granulocytesduring Coagulation
Coagulation in L. dispar appears to correspond topattern III as described by Gregoire (1970) and essen-tially involves hemocyte lysis starting with the coagu-locytes and granulocytes. Hemocyte lysate appears totrigger a number of events in the hemolymph: forma-tion of strands, cells becoming sticky, enhanced phago-cytosis, and/or further lysis of hemocytes.Strands in the hemolymph have been reported by
other workers (Rowley, 1977; Gregoire and Goffinet,1979). The sticky structures trapped cell componentsand other hemocytes. It is not clear whether they weregenerated from lysate or were filopodial extensionsfrom plasmatocytes or granulocytes. According toWago (1983, 1984) the filopodia signal other hemocytesto become phagocytic or partake in encapsulation. He-mocytes in contact with, or in the vicinity of thestrands lost their form and ultimately lysed. Hemocytelysate is known to “prime” hemocytes. Soderhall et al.(1984) have shown that hemocyte lysate enhances theencapsulation reaction of crayfish hemocytes. Butt andHumber (1989) reported that plasmatocytes become
STRUCTURE AND BEHAVIOR OF L. dispar HEMOCYTES 13

JOBNAME: JIP 68#1 96 PAGE: 14 SESS: 12 OUTPUT: Fri Jun 21 17:17:00 1996/xypage/worksmart/tsp000/71389k/8pu
modified and will envelope other plasmatocytes andcoagulum surrounding foreign cells.Coagulation of gypsy moth hemolymph as observed
on a glass microscope slide culminates in melanizationof the comparatively homogeneous coagulum. The co-agulum is similar in appearance whether it forms atthe surface of an air bubble, the edge of a coverslip, orthe surface of host-derived or foreign debris. If coagu-lation of hemolymph is allowed to proceed in an Ep-pendorf vial it also culminates in hemocyte lysis andmelanization, but the hemolymph is more gelatinous.According to Bohn (1986) clotting agents are releasedfrom granulocytes and/or coagulocytes which interactwith plasma proteins to induce gelation. He believesthat the hemocyte coagulogen (clotting protein) is con-tained in the granules of hemocytes and that thesereact with the plasma coagulogen (a lipophorin) to formthe clot. Lipophorin is a major component of gypsymoth hemolymph (unpublished). So it is highly feasiblethat the strands we described represented strands ofcoagulum rather than filopodial extensions.
ACKNOWLEDGMENT
IACR-Rothamsted receives grant-aided support from the BBSRC.
REFERENCES
Akai, H. and Sato, S. 1973. Ultrastructure of the larval hemocytes ofthe silkworm, Bombyx mori L. (Lepidoptera: Bombycidae). Int. J.Insect Morphol. Embryol. 2, 207–231.
Beaulaton, J. and Monpeyssin, M. 1976. Ultrastructure et cyto-chimie des hemocytes d’Antheraea pernyi, Guer. (Lepidoptera, At-tacidae) au cous du cinquieme age larvaire. J. Ultrastruct. Res. 55,143–156.
Bell, R. A. Owens, C. D. Shapiro, M. and Tardif, J. G. R. 1981. Massrearing and virus production. In “The Gypsy Moth: Research to-ward Integrated Pest Management” (C. C. Doane and M. L. Mc-Manus, Eds.), pp. 599–655. USDA, Washington, DC.
Bohn, H. 1986. Hemolymph clotting in insects. In “ Immunity inInvertebrates” (M. Brehelin, Ed.) pp. 188–207. Springer-Verlag,Berlin.
Brehelin, M. and Hoffmann, J. A. 1980. Phagocytosis of inert par-ticles in Locusta migratoria and Galleria mellonella: Study of ul-trastructure and clearance. J. Insect Physiol. 26, 103–111.
Brehelin, M. and Zachary, D. 1983. About insect plasmatocytes andgranular cells. Dev. Comp. Immunol. 7, 683–686.
Brehelin, M. and Zachary, D. 1986. Insect haemocytes: A new clas-sification to rule out the controversy. In “Immunity in Inverte-brates” (M. Brehelin, Ed.), pp. 36–48. Springer-Verlag, Berlin.
Brehelin, M. Zachary, D. and Hoffman, J. A. 1978. A comparativeultrastructural study of blood cells from nine insect orders. CellTissue Res. 195, 45–57.
Butt, T. M. and Beckett, A. 1984. Ultrastructure and Behavior of thespindle pole body of the aphidpathogenic fungus Erynia neoaphi-dis. Protoplasma 121, 61–71.
Butt, T. M. and Humber, R. A. 1989. Response of gypsy moth hemo-cytes to natural fungal protoplasts of three Entomophaga species(Zygomycotina: Entomophthorales). J. Invertebr. Pathol. 53, 121–123.
Butt, T. M. Hoch, H. C. Staples, R. C. and St. Leger, R. J. 1989. Use
of fluorochromes in the study of fungal cytology and differentia-tion. Exp. Mycol. 13, 303–320.
Cook, D. Stoltz, D. B. and Pauley, C. 1985. Purification and prelimi-nary characterization of insect spherulocytes. Insect Biochem. 15,419–426.
Devauchelle, G. 1971. Etude ultrastructurale des hemocytes du Co-leoptere Melolontha melolontha (L). J. Ultrastruct. Res. 34, 492–516.
Essawy, M. Maleville, A. and Brehelin, M. 1985. The hemocytes ofHeliothis armigera: Ultrastructure, functions, and evolution in thecourse of larval development. J. Morphol. 186, 255–264.
Gregoire, C. 1970. Haemolymph coagulation in arthropods. Symp.Zool. Soc. London 27, 45–74.
Gregoire, C. and Goffinet, G. 1979. Controversies about the coagu-locyte. In 0Insect Hemocytes0 (A. P. Gupta, Ed.), pp. 189–229.Cambridge Univ. Press, Cambridge.
Gupta, A. P. 1979. “Insect Hemocytes.” Cambridge Univ. Press,Cambridge.
Gupta, A. P. 1986. “Hemocytic and Humoral Immunity.” Wiley, NewYork.
Hagopian, M. 1971. Unique structures in the insect granular hemo-cytes. J. Ultrastruc. Res. 36, 646–658.
Harpaz, I. Kislev, N. and Zelcer, A. 1969. Electron-microscopic stud-ies on hemocytes of the Egyptian cottonworm, Spodoptera littora-lis (Boisduval) infected with a nuclear polyhedrosis virus, as com-pared to non-infected hemocytes. I. Noninfected hemocytes. J. In-vertebr. Pathol. 14, 175–185.
Iwama, R. and Ashida, M. 1986. Biosynthesis of prophenoloxidase inhemocytes of larval hemolymph of the silkworm, Bombyx mori.Insect Biochem. 16, 547–555.
Pech, L. L. Trudeau, D. and Strand, M. R. 1995. Effects of basementmembrane on the behavior of hemocytes from Pseudoplusia in-cludens (Lepidoptera: Noctuidae): Development of an in vitro en-capsulation assay. J. Insect Physiol. 41, 801–807.
Raina, A. K. 1976. Ultrastructure of the larval haemocytes of thepink bollworm Pectinophora gossypiella (Lepidoptera: Gelechi-idae). Int. J. Insect Morphol. Embryol. 5, 187–195.
Ratcliffe, N. A. 1993. Cellular defense responses of insects: Unre-solved problems. In “Parasites and Pathogens of Insects” (N. E.Beckage, S. N. Thompson, and B. A. Frederici, Eds.), Vol. 1, pp.267–304. Academic Press, San Diego.
Ratcliffe, N. A. and Rowley, A. F. 1979. A comparative synopsis ofthe structure and function of the blood cells of insects and otherinvertebrates. Dev. Comp. Immunol. 3, 189–243.
Ravindranath, M. H. 1978. The individuality of plasmtocytes andgranular hemocytes of arthropods: A review.Dev. Comp. Immunol.2, 581–594.
Rowley, A. F. 1977. The role of the haemocytes of Clitumnus extra-dentatus in haemolymph coagulation. Cell Tissue Res. 182, 513–524.
Russo, J. Allo, M-R. Nenon, J-P. and Brehelin, M. 1993. The hemo-cytes of the mealybugs Phenacoccus manihoti and Planococcuscitri (Insecta: Homoptera) and their role in capsule formation.Can. J. Zool. 72, 252–258.
Soderhall, K. Vey, A. and Ramstedt, M. 1984. Hemocyte lysate en-hancement of fungal spore encapsulation by crayfish hemocytes.Dev. Comp. Immunol. 8, 23–29.
Wago, H. 1983. Cellular recognition of foreign materials by Bombyxmori phagocytes: II. Role of hemolymph and phagocyte filopodia inthe cellular reactions. Dev. Comp. Immunol. 7, 199–208.
Wago, H. 1984. In vitro evidence for the requirement of filipodialelongation for the progress of phagoyctosis by phagocytic granularcells of the silkworm, Bombyx mori. Dev. Comp. Immunol. 8, 7–14.
BUTT AND SHIELDS14





























