Embed Size (px)
Citation preview

Aquaculture, 66 (1987) 299-313 Elsevier Science Publishers B.V., Amsterdam - Printed in The Netherlands
299
Growth and Composition of the Swimming Muscle of Adult Atlantic Salmon (Salmo salar L.) During Long-Term Sustained Swimming
G.K. TOTLAND”, H. KRYVI’, K.A. JBDESTBL’, E.N. CHRISTIANSEN3, A. TANGERAS“ and E. SLINDE4
‘Zoological Laboratory, University of Bergen, Allegt. 41,5007 Bergen (Norway) ‘Department for Marine Zoology and Chemistry, Institute for Biology, University of Oslo, P.O. Box 1064,0316 Oslo 3 (Norway) 3Znstitute for Nutrition Research, University of Oslo, Oslo (Norway) 'Norbio A/S, P.O. Box 1137,500l Bergen (Norway) ‘To whom correspondence should be addressed.
(Accepted 21 May 1987)
ABSTRACT
Totland, G.K., Kryvi, H., Jsdestel, K.A., Christiansen, E.N., Tangerh, A. and Slinde, E., 1987. Growth and composition of the swimming muscle of adult Atlantic salmon (Salmo salar L.) during long-term sustained swimming. Aquaculture, 66: 299-313.
Atlantic salmon were raised in a swimming raceway on a commercial scale for 8 months, and the effects on growth and composition of the muscle were analysed. The weight gain was nearly 40% higher in fish reared in the raceway compared to the reference fish raised in ordinary cages. Measurements of fibre size distribution in red and white muscle mass revealed that the main difference in growth occurred in the white portion of the muscle. The raceway fish had a larger proportion of hypertrophied white fibres, but the inner red fibres were smaller. The reference fish had a lower content of stored glycogen and triglycerides in the inner red fibres than in the corre- sponding fibres in the exercised fish. Quality assessment according to industry-approved specifi- cations indicated 9.2% more superior-quality fish in the raceway group than in the reference
group.
INTRODUCTION
The health, quality and growth of fish are factors of prime importance in the fish farming industry. The environment of the fish influences these factors, and therefore great efforts are made to find optimal rearing conditions. It is of central interest in this respect to find the optimum conditions for muscle growth and feed conversion efficiency. Moderate exercise training of fish is known to enhance both these factors (Besner, 1980; Kuiper, 1982)) as well as survival
00448486/81/$03.50 0 1987 Elsevier Science Publishers B.V.

300
rate (Burrows, 1969). Trained fish also increase their critical swimming speed (Fairlinger and Beamish, 1978)) the ability to resist fatigue (Brett et al., 1958; Hammond and Hickman, 1966) and to withstand stress (Woodward and Smith, 1985). Fish seem to achieve this by adaptive changes both in their metabolism and the muscle. Previous studies on the effects of exercise training have been performed in relatively small swimming tanks or respirometers with small specimens which have been forced to swim at different water velocities for relatively short periods of time. The relevance of such studies to the conditions found in fish farms is largely unknown.
The present investigation was carried out on a commercial scale at a fish farm with Atlantic salmon (Salmo s&r L. ) (5000 fish, average initial weight 2 kg). The main purposes were to study the effect of long-term sustained swim- ming (8 months) on survival rate, muscle growth and composition, and also the fish quality. Fish muscle is suited for such studies since the fibre types form distinct populations. Biochemical characterizations can be performed on groups of pure muscle fibre types, free from effects of neighbouring fibres. The arrangement of muscle fibre types in the swimming muscle of the Atlantic salmon is similar to that of most teleost fish (see reviews by Bone, 1978; John- ston, 1981). The muscle can be divided into two separate parts: a lateral red portion which supports sustained activity, and a medial white part which makes up the bulk of the muscle and which is recruited for sustained swimming at higher speed, as well as for shorter bursts of activity.
The quality of fish muscle is determined by both the qualitative and quan- titative composition of muscle fibres and connective tissue. Knowledge of the relative contribution of the different components to the quality is still scanty. In the present experiment the group of fish which performed long-term train- ing was of the same stock of fish as that which experienced ordinary farming conditions. This may give clues to a better understanding of the relation between muscle characteristics and assessment of quality.
MATERIALS AND METHODS
The practical setup was designed by Noraqua A/S, Oslo, and conducted at Kvalos Fish Farm near M&y, Norway. The fish, Atlantic salmon (Salmo sub- L.) , were 2 years of age at the start of the experiment. During the experimental period the fish were fed daily TESS EDEL Dryfood (Skretting A/S, Stavan- ger) . The food was offered to satiation which implies that the food was with- drawn as soon as a marked reduction in the feeding activity was observed, thus allowing the daily food intake to be monitored.
Experimental setup
The fish (mean length and weight: 56.3 cm and 2038 g, respectively) were divided into two groups of similar size, and both were kept in sea water at

301
seasonal temperatures (highest temperature, in October, 12 ‘C and lowest, in February, 2OC). One group was kept in a 300-m3 swimming raceway (length, 20 m; depth, 4.5 m; width, 4 m) , equipped with three streamsetters, each with 30-cm diameter propellers, fixed in one end. The water had a mean speed of 28.0 5 11.8 cm/s throughout the period. The mean calculated swimming speed dropped from 0.45 ? 0.23 L/s (body length/s + SD) at the start to 0.40 + 0.16 L/s at the end of the experiment. The maximum swimming speed was calcu- lated as 0.80 L/s, based on the water speed. The reference group was kept in standard cages with the same volume and stocking density as in the raceway, ca. eight fish/m3. At the start of the experiment this was equivalent to ca. 17 kg fish/m3. The water in the reference cage had a maximum speed of 5 cm/s. The direction of flow also varied, according to the tidal cycle.
Growth
The weight gain was recorded at intervals during the experimental period. A minimum of 50 fish from each group were weighed and their lengths meas- ured. At the end of the experiment, 500 fish were recorded. The weight of the visceral organs was also measured (10 fish from both groups on five occasions ) . The condition factor (CF) was calculated from the equation
CF=lOO~Weight (g)
(Length in cm) 3
The conversion rate was calculated as
Conversion rate = Food consumed/fish (kg)
Weight gain/fish (kg)
( n= 2257 and 2204 for raceway and reference groups, respectively.)
Morphometrical and histochemical analysis
Muscle blocks (approximately 0.5 cm3) were removed immediately after killing the animals by a quick blow to the head. Samples were taken from two locations: (1) close to the lateral line at the level of the anterior end of the dorsal fin (anterior), and (2) close to the lateral line at the level of the adipose fin (posterior). The samples were rapidly frozen by immersion in propane cooled to its melting point in liquid nitrogen, and transported in liquid nitrogen to the laboratory. Serial sections, 10pm thick, were cut on a cryostat and stained for activity of NADH-tetrazolium reductase (diaphorases-oxidative enzymes), glycogen (PAS), fat (Sudan B ) and connective tissue (collagen).
The volume fractions of red and white muscle tibres were measured on trans- verse sections of the whole tail taken from the area immediately in front of the adipose fin. From the individual muscle fibres cross-sectional area, circumfer-

302
Outer red Inner red
Outer whlte Inner white
Fig. 1. Section across the body of the fish to show the location of different muscle layers in the red and white portions of the muscle.
ence and form factor (F-factor’) were measured in four different layers of the muscle: outer red, inner red, outer white and inner white areas ( Fig. 1) . A total of 150 muscle frbres from each area was measured with a Leitz ASM 68K semi- automatic image analyser.
Determinations of glycogen, triglycerides, protein, dry matter and carotenoid pigments
Samples of muscle from the four different layers were analysed for glycogen and triglycerides. The samples were weighed, frozen in liquid nitrogen and pulverized. Triglycerides were analysed using a commercial enzymatic kit (including lipase) from Nycomed A/S, Oslo (Nycotest Triglycerides). Gly- cogen was extracted from pulverized muscle (100-200 mg protein) by 0.6 N HClO, as described by Keppler and Decker (1974). An aliquot of the extract was hydrolysed with amyloglucosidase for 160 min at 40°C. The hydrolysate was analysed for glucose content using a commercial enzymatic kit from Boeh- ringer Mannheim, F.R.G. (sucrose/glucose cat. no. 139 041). Correction for efficiency of glycogen hydrolysis was made using glycogen from oyster ( Sigma cat. no. G 8751) treated in the same way as the samples. Total nitrogen was measured as Kjeldahl N. Dry matter in the white muscle mass was determined after drying in an oven at 72°C for 2 days. The samples were taken from the front of the dorsal fin to above the lateral line, free from skin and bones. The carotenoids were extracted from the deep part of the white muscle with acetone and separated and quantified by HPLC using a Supelco LC-CN column and a mobile phase of n-hexane/2-propanol/N-ethyldiiso-propylamine (90:9:1)
(4nXarea) ‘Determined by the formula F= Circumference2

(Schiedt et al., 1981). In the standard solutions the concentrations of caroten- oids were determined spectrophotometrically using A :Tm of 2000 and 1600 for canthaxanthin and astaxanthin, respectively (Britton, 1985; Johnson et al., 1977).
Quality judgement and sensory analysis
At the completion of the experiment the fish were classified as superior, ordinary or quality-production fish according to standard Norwegian industry- approved specifications (Fiskeridirektoren, 1984). The sensory analysis was performed by a trained taste panel consisting of 12 persons at the Norwegian Food Research Institute. Ten fish from each group were evaluated by each judge on a nine-point scale for the following sensory properties: coarseness, firmness, fatness, juiciness, characteristic taste, colour and off-flavour.
RESULTS
Behavioural observations
During the first 2 weeks of the experiment the fish in the raceway were located in the rear end of the tunnel, later the fish were more evenly distributed throughout the raceway tunnel. Occasional bursts of swimming were most fre- quent during feeding. The fish often alternated between swimming and gliding, and could also be observed drifting along with the water current. In the coldest periods the fish stayed mostly in the deeper parts of the tunnel. Throughout the experimental period they were generally absent from the surface waters. The fish in the reference group generally swam anticlockwise. This pattern was markedly broken up by a number of fishes swimming in all directions. The fish kept in the raceway regime had a larger appetite than the fish which expe- rienced ordinary fish-farm conditions, and this resulted in a 47% larger amount of food being offered to the raceway fish than the reference fish.
Growth and mortality
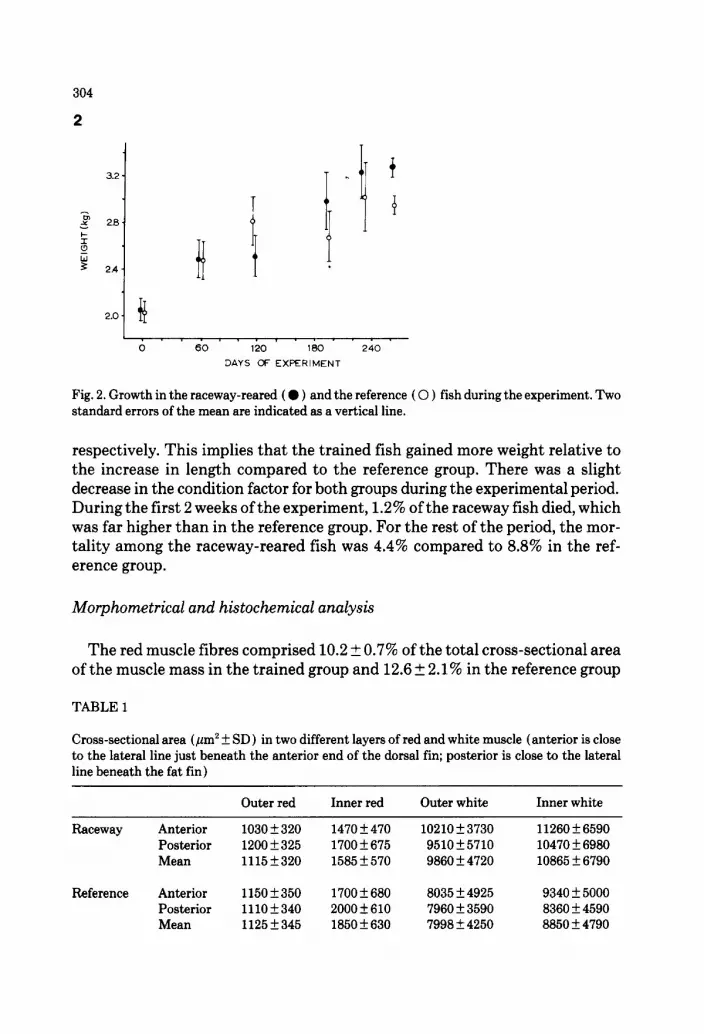
At the end of the experiment the trained fish had an average weight gain of 1.240 kg/fish (Fig. 2)) compared to 0.900 kg in the reference group. This rep- resents a 38% difference in growth (PC 0.05 ) . The main difference in growth took place in the swimming muscle; the growth of the internal organs was similar in the two groups. Since the trained fish had a better growth, the pro- portion of fish weighing 2-5 kg was higher in this group.
The conversion rate measured at the end of the experiment was 1.80 for the raceway fish and 1.73 for the reference group. At the end of the experiment the condition factor for the trained and the reference group was 0.97 and 0.93,
304
2
0 60 120 160 240
DAYS Cf EXPERIMENT
Fig. 2. Growth in the raceway-reared ( 0 ) and the reference ( 0 ) fish during the experiment. Two standard errors of the mean are indicated as a vertical line.
respectively. This implies that the trained fish gained more weight relative to the increase in length compared to the reference group. There was a slight decrease in the condition factor for both groups during the experimental period. During the first 2 weeks of the experiment, 1.2% of the raceway fish died, which was far higher than in the reference group. For the rest of the period, the mor- tality among the raceway-reared fish was 4.4% compared to 8.8% in the ref- erence group.
Morphometrical and histochemical analysis
The red muscle fibres comprised 10.2 2 0.7% of the total cross-sectional area of the muscle mass in the trained group and 12.6 + 2.1% in the reference group
TABLE I
Cross-sectional area (pm* f SD) in two different layers of red and white muscle (anterior is close to the lateral line just beneath the anterior end of the dorsal fin; posterior is close to the lateral line beneath the fat fin)
Outer red Inner red Outer white Inner white
Raceway Anterior 1030+320 1470f470 10210f3730 11260f6590 Posterior 1200f325 1700f675 9510f5710 10470f6980 Mean 1115f320 1585f570 9860 + 4720 10865f6790
Reference Anterior 1150+350 1700+680 8035 f 4925 9340 f 5000 Posterior lllOf340 2000+610 7960 f 3590 8360 f 4590 Mean 1125f345 1850f630 7998 + 4250 8850 f 4790

305
RED MUSCLE FIBRES
CROSS-SECTIONAL AREA(x102jm2)
Fig. 3. Frequency distribution of the cross-sectional areas of red muscle fibres of exercised and reference fish. Both inner and outer fibres are included. Each class is 100 pm’.
4
I
16
0 T 12
t,
3 8
iii
: 4 LL
0
10 20 30 40
CROSS-SECTIONAL AREA(x10$n2)
Fig. 4. Frequency distribution of the cross-sectional areas of white muscle fibres of exercised and reference fish. Both inner and outer fibres are included. Each class is 1666 pm’.
(measured at the anterior level of the adipose fin). The cross-sectional area of the red muscle fibres sampled from the inner and outer layers was in general smaller in the anterior than in the posterior part of the fish (Table 1) , as opposed to the white fibres which were larger in the anterior than in the pos- terior part. The mean cross-sectional area of the outer red fibres was similar in the two groups (Table 1, Fig. 3 ) . The inner red fibres in the trained group had about a 15% smaller cross-sectional area than the corresponding fibres in the reference fish (Table 1, Fig. 3 ) .
306

307
The mean cross-sectional area of the white fibres in the trained fish was approximately 17% larger than the corresponding fibres in the reference group (Fig. 4). This was true for both the outer and inner parts of the white muscle and was mainly due to a population of extremely large fibres (shown in Fig. 7, compare with Fig. 8; see also Figs. 3 and 4).
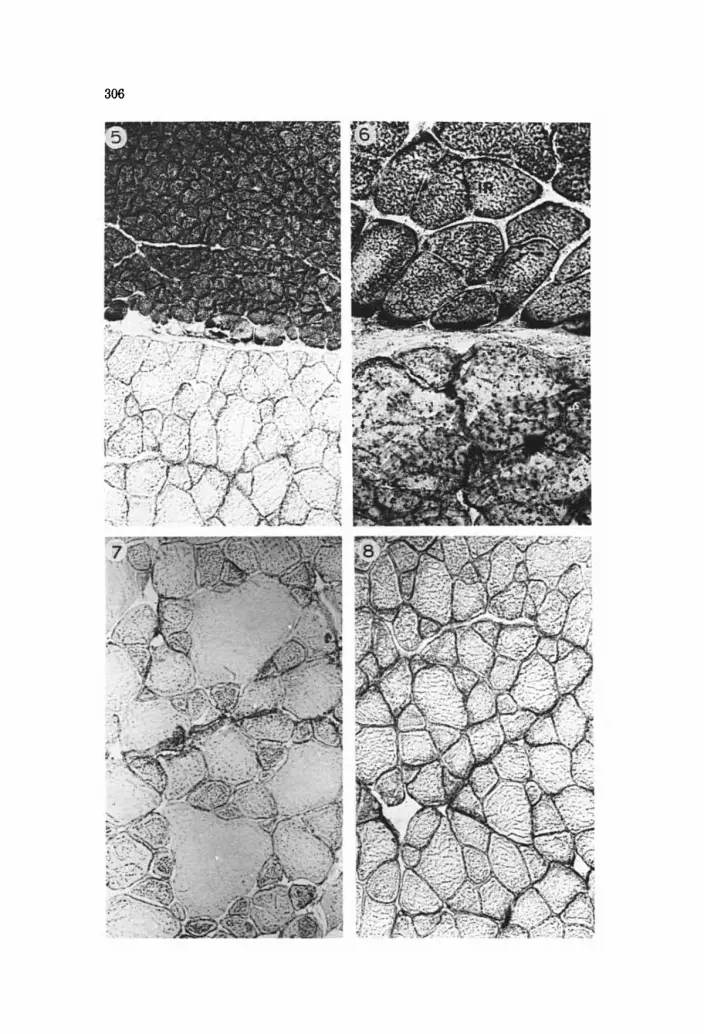
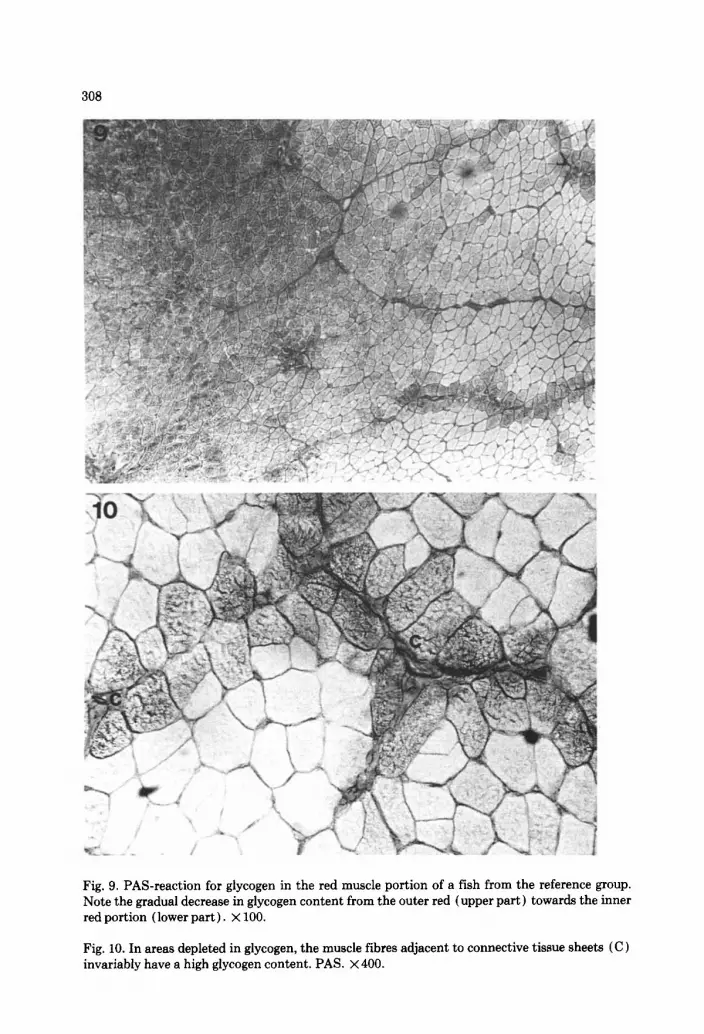
The red muscle fibres were round and homogeneous in cross-sectional area (F= 0.96). There was an abundance of mitochondria (Fig. 5)) and blood cap- illaries were numerous between the fibres. The innermost layers of the red fibres contained conspicuous subsarcolemmal mitochondrial accumulations (Fig. 6). A zonation was present in the distribution of glycogen, with a marked reduction in the PAS-positive material in the inner third of the red layer (Fig. 9). This was most evident in the reference fish. In the areas depleted in gly- cogen, the fibres adjacent to connective tissue sheets invariably had a high glycogen content (Fig. 10). A similar zonation was evident in fat content, with less fat in the inner red fibres. No difference was observed in the two groups in this respect. Fat was present both within and between the fibres.
White fibres were heterogeneous in cross-sectional area. They contained few mitochondria, little fat and were surrounded by few capillaries (Figs. 7 and 8). The most lateral white fibres contained more fat than the main muscle mass in both trained and untrained fish. Among the white fibres a distinct difference in the distribution of mitochondria was observed between the two test groups. In the trained fish, large fibres contained fewer mitochondria and the small fibres more mitochondria than in the reference group (Fig. 7). In the white muscle of the reference group, the mitochondria had a more even distribution (Fig. 8). No detectable changes were found in content and distribution of con- nective tissue components between the two groups.
Determinations of glycogen, triglycerides, protein, dry matter and carotenoid pigments
All muscle layers in both groups had a glycogen content between 5 and 10 mg/g muscle (Table 2). In all muscle types the trained fish contained more
Fig. 5. Transverse section to show the distribution of red and white muscle fibres. The red and white portions of the muscle are separated by a thin sheet of connective tissue. Section stained for NADH-tetrazolium reductase (NADH-TR) activity. Red portion - upper part; white portion - lower part X 100.
Fig. 6. Inner red and outer white fibres from the reference group. Note that the inner red fibres are larger and have more subsarcolemmal located mitochondria than the rest of the red fibres. NADH-TR. IR= inner red fibres. x 400.
Fig. 7. White muscle fibres from a raceway-reared fish. Note the extreme variation in cross-sec- tional area among the fibres and the difference in content of mitochondria. NADH-TR. x 100.
Fig. 8. White muscle fibres from a fish raised under ordinary fish-farm conditions. NADH-TR. x 100.
308
Fig. 9. PAS-reaction for glycogen in the red muscle portion of a fish from the reference group. Note the gradual decrease in glycogen content from the outer red (upper part) towards the inner red portion (lower part). x 100.
Fig. 10. In areas depleted in glycogen, the muscle fibres adjacent to connective tissue sheets (C) invariably have a high glycogen content. PAS. x400.
309
- I
I 1 . 1 7 (
60 120 180 240
DAYS OF EXPERIMENT
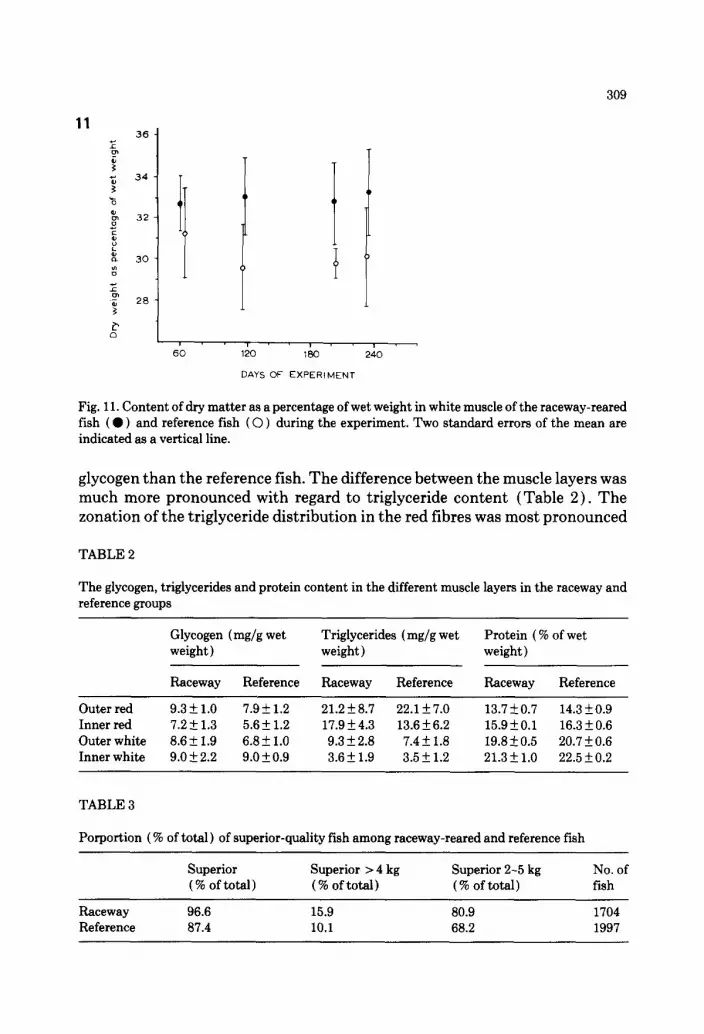
Fig. 11. Content of dry matter as a percentage of wet weight in white muscle of the raceway-reared fish ( 0 ) and reference fish ( 0 ) during the experiment. Two standard errors of the mean are indicated as a vertical line.
glycogen than the reference fish. The difference between the muscle layers was much more pronounced with regard to triglyceride content (Table 2). The zonation of the triglyceride distribution in the red fibres was most pronounced
TABLE 2
The glycogen, triglycerides and protein content in the different muscle layers in the raceway and reference groups
Outer red Inner red Outer white Inner white
Glycogen (mg/g wet Triglycerides (mg/g wet weight) weight )
Raceway Reference Raceway Reference
9.3+ 1.0 7.9 + 1.2 21.2 f 8.7 22.1* 7.0 7.2 + 1.3 5.6f 1.2 17.9 f 4.3 13.6? 6.2 8.6 + 1.9 6.8 f 1.0 9.3 f 2.8 7.4 + 1.8 9.0 + 2.2 9.0 f 0.9 3.6 + 1.9 3.5 * 1.2
Protein (% of wet weight)
Raceway Reference
13.71to.7 14.3 + 0.9 15.9fO.l 16.3 + 0.6 19.8 + 0.5 20.7 k 0.6 21.3 f 1.0 22.5 + 0.2
TABLE 3
Porportion (% of total) of superior-quality fish among raceway-reared and reference fish
Raceway Reference
Superior (% oftotal)
96.6 87.4
Superior > 4 kg (% oftotal)
15.9 10.1
Superior 2-5 kg (W of total)
80.9 68.2
No. of fish
1704 1997

310
in the reference group, which had a high fat concentration superficially and low in the inner red layer. The trained fish had a more even distribution of triglycerides in the different muscle layers. In all the different muscle layers the reference salmon had a slightly higher protein content than the trained fish (Table 2). The dry matter content was higher in the raceway fish than in the reference fish (Fig. 11) throughout the experiment.
No significant differences were found between the concentration of caroten- oids in the flesh of fishes reared in the raceway and the ordinary cage. The concentration of canthaxanthin in the fish reared in the raceway was 2.0 t 0.6 mg/kg wet weight compared to 1.7 ? 0.6 mg/kg in the control group (n= 10 for each group). The astaxanthin concentration in flesh from raceway-reared and control fishes was 4.7 & 1.9 and 4.3 Ifi 0.8 mg/kg wet weight, respectively (n= 10 for each group).
Quality judgement and sensory analysis
When the fish were slaughtered and classified, the proportion of fish in the superior-quality group was significantly higher among the trained fish (Table 3). The percentage of fish classified as lower quality was 3.4% in the trained group and 12.6% in the reference group (n x 2000 in each group). The test performed by the taste panel showed that off-flavour was somewhat lower and the characteristic taste was slightly higher in the trained fish. In both groups the flesh and the tail region was less coarse and less firm, but more juicy and had a higher characteristic taste than flesh from more anterior parts of the fish. The taste panel did not observe any difference in colour between the two groups.
DISCUSSION
The aim of this study was to examine the effect of sustained exercise on the swimming muscle of adult Atlantic salmon. In ordinary fish-farm regimes the size of the cage and the stocking density impose constraints on the locomotory possibilities of fish. Particularly for the larger species like salmon, the loco- motory repertoire is obviously limited. Many of the behavioural patterns char- acteristic of the species in the wild may be physically inhibited in ordinary fish-farm regimes.
Due to the water current and the shape of the swimming tunnel, the fish reared in the raceway were stimulated to swim in a more natural pattern. The distribution of fish in one end of the raceway at the beginning of the experi- ment implied that the start of an exercise period was stressful to the fish. Dav- ison and Goldspink (1977) mention that up to 20% of the brown trout exercised at 1.5 and 3 L/s died during the first 2 days of their experiment, and that this is common in exercise experiments of fish. This was also experienced in the

311
present study where a higher number of fish in the raceway died compared with the reference group during the first 2 weeks of the experiment. This is in contrast to the result for the remaining experimental period where the mor- tality among the fish in the reference group was twice that of raceway reared fish. In conclusion, we recommend that the water current is gradually increased during the first weeks of similar setups.
The significantly higher appetite in the raceway fish is probably caused by the water current which forces the fish to swim more actively. The better water exchange, improved swimming activities and less confusion during feeding (Milinski, 1977) provided better physical conditions, may have reduced aggression and probably explain the better appetite. The fact that fewer eye bites and skin wounds were observed among the raceway fish than among the reference fish indicates less aggression. This is also reflected in the higher qual- ity of the raceway fish.
The food conversion rate was of the same magnitude in the two groups. The experimental setup did not, however, allow the loss of food to be recorded.
The increase in weight during the experimental period was rather low. This was probably mainly due to low temperatures during a large part of the exper- imental period.
Previous histological studies have shown that when fish are compelled to swim at a certain speed for long periods of time, the muscle fibres used for the propulsion become hypertrophied ( Greer-Walker, 1971; Greer-Walker and Pull, 1973; Greer-Walker and Emerson, 1978; Johnston and Moon, 1980). Our results indicate that the muscle fibres in the outer red portion of the muscle are used in a similar way in the two groups. The muscle fibres in the inner red portion of the reference group are larger than the corresponding fibres in the trained group, indicating that these fibres are more heavily involved in swimming activity. This is also supported by the reduced stores of glycogen and triglyc- erides in the same layers.
The white muscle fibres in the raceway fish have obviously been more actively used than the corresponding fibres in the fish from the reference group. The white fibres of the raceway fish had a 17% larger mean cross-sectional area, which was mainly due to the appearance of some new large-diameter fibres. This may explain most of the increased growth in the fish reared in the race- way. It also indicates that most of the growth is due to hypertrophy in the white portion of the muscle, and not to an increase in the number of muscle fibres. This has also been suggested by Weatherley et al. (1980), who found no increase in fibre numbers in rainbow trout larger than 50 cm. Previous studies have indicated that a water speed greater than l-l.5 L/s was necessary to induce growth in white muscle in trout and salmon (Davison and Goldspink, 1977; Besner, 1980; Nahhas et al., 1982; Kuiper, 1982). These studies, however, were performed with small specimens ( 6-15 cm), which have a different swimming efficiency (Webb, 1975) than the larger specimens used in our study. Based

312
on Webb’s results, it can be concluded that the white fibres will be recruited at lower relative swimming speeds in larger specimens. In addition, it was observed that the bursts in swimming were more frequently performed by the raceway fish than by the reference fish. We can therefore suggest that the bursts in swimming are responsible for the marked hypertrophy of white fibres and that the sustained swimming is responsible for the increase in concentration of mitochondria observed in the smaller white fibres. The increase in aerobic capacity is in agreement with the study of Davie et al. (1986) who found a similar effect in trout after sustained swimming. Based on our histological observations, then, it is reasonable to infer that the long-term exercise increased both aerobic and anaerobic capacity in the white portion of the muscle.
The content of dry matter did not change significantly in the swimming muscle during the experimental period. The increase in body weight must therefore be an increase in solid matter. The relative composition of protein, glycogen and triglycerides was influenced by exercise. The raceway-reared fish contain more glycogen and triglycerides and less protein than the fish reared in the ordinary cages. This seems to have little influence on the taste properties of the fish. In the fish from both groups the tail region obtained higher taste scores than the anterior part of the fish. The tail region contains a higher proportion of red muscle fibres than the anterior part. This indicates a corre- spondence between the biochemical and histochemical profiles of the muscle and its taste properties. We also suggest that the hypertrophy in the white muscle portion of the trained fish increased the firmness of the fish, and that it is partly responsible for the higher proportion of superior-quality fish.
The main advantages of the raceway rearing system are a higher weight gain, a lower mortality, and a better quality of fish. Increased expenses were mainly electricity needed to run the propellers (4 kW) and a higher food consumption. In the present experiment the net economic gain was significant.
ACKNOWLEDGEMENTS
We thank Noraqua A/S (Oslo) for providing material and facilities for this investigation.
REFERENCES
Besner, M., 1980. Endurance training: an affordable rearing strategy to increase food conversion efficiency, stamina, growth and survival of coho salmon smelts (Oncorhynchus klsutch) . Ph.D. dissertation, University of Washington, Seattle, WA, 200 pp.
Bone, Q., 1978. Locomotor muscle. In: W.S. Hoar and D.J. Randall (Editors), Fish Physiology, Vol. VII. Academic Press, New York, NY, pp. 362-423.
Brett, J.R., Hollands, M. and Alderdice, D.E., 1958. The effect of temperature on the cruising speed of young sockeye and coho salmon. J. Fish. Res. Board Can., 15: 587-605.

313
Britton, G., 1985. General carotenoid methods. Meth. Enzymol., 111: 113-149. Burrows, R.E., 1969. The influence of fingerling quality on adult salmon survivals. Trans. Am.
Fish. Sot., 98: 777-784. Davie, P.S., Wells, R.M.G. and Tetens, V., 1986. Effects of sustained swimming on rainbow trout
muscle structure, blood oxygen transport, and lactate dehydrogenase isozymes: evidence for increased aerobic capacity of white muscle. J. Exp. Zool., 237: 159-171.
Davison, W. and Goldspink, G., 1977. The effect of prolonged exercise on the lateral musculature of the brown trout (Salmo trutta). J. Exp. Biol., 70: 1-12.
Fairlinger, S. and Beamish, F.W.H., 1978. Changes in blood chemistry and critical swimming speed of largemouth bass, Micropterus salmoides, with physical conditioning. Trans. Am. Fish. Sot., 107: 523-527.
Fiskeridirektaren, 1984. Melding fra fiskeridirekteren: Regler for kvalitetsgradering av oppdr- ettsfisk. Nor. Fiskeoppdrett, 9: 34-35.
Greer-Walker, M., 1971. Effect of starvation and exercise on the skeletal muscle fibres of the cod (Gadus morhua L.) and the coalfish (Gadus uirens L.) , respectively. J. Cons. Int. Explor. Mer., 33: 421-427.
Greer-Walker, M. and Emerson, L., 1978. Sustained swimming speeds and the myotomal muscle function in the trout (Salmo gairdneri) . J. Fish Biol., 13: 475-481.
Greer-Walker, M. and Pull, G.A., 1973. Skeletal muscle function and sustained swimming speeds in the coalfish (Gadus virens L.). Comp. Biochem. Physiol. A, 44: 495-502.
Hammond, B.R. and Hickman, C.P., Jr., 1966. The effect of physical conditioning on the metab- olism of lactate, phosphate, and glucose in rainbow trout (Salmo gairdneri) . J. Fish. Res. Board Can., 23: 65-83.
Johnson, E.A., Conklin, D.E. and Lewis, M.J., 1977. The yeast Phaffia rhodozyma as a dietary pigment source for salmonids and crustaceans. J. Fish. Res. Board Can., 34: 2417-2421.
Johnston, I.A., 1981. Structure and function of fish muscles. In: M.H. Day (Editor), Zool. Sot. Symp. Vertebrate Locmotion, Vol. 45. Academic Press, London, pp. 71-113.
Johnston, I.A. and Moon, T.W., 1980. Endurance training in fast and slow muscles. J. Comp. Biochem., 135: 147-156.
Keppler, D. and Decker, K., 1974. Glycogen, determination with amyloglucosidase. In: H.U. Berg- meyer (Editor), MethodsofEnzymatic AnaIysisVol. 3. Academic Press, London, pp. 1127-1131.
Kuiper, J., 1982. Salmon thrive on exercise. Fish Farmer, 5(4): 9-10. Milinski, M., 1977. Experiments on the selection by predators against spatial oddity of their prey.
Z. Tierpsychol., 43: 311-325. Nahhas, R., Jones, N.V. and Goldspink, G., 1982. Some aspects of sustained training of rainbow
trout (Salmo gairdneri). J. Fish Biol., 20: 351-353. Schiedt, K., Leuenberger, F. and Vecchi, M., 1981. Natural occurrence of enantiomeric and meso-
astaxanthin. 5. Ex. wild salmon (Salmo salar and Oncorhynchus). Helv. Chim. Acta, 64: 449-457.
Weatherley, A.H., Gill, H.S. and Rogers, S.C., 1980. The relationship between mosaic muscle fibres and size in rainbow trout (Salmo gairdneri). J. Fish Biol., 17: 603-610.
Webb, P.W., 1975. Hydrodynamics and energetics of fish propulsion. Bull. Fish. Res. Board Can., 190: 158 pp.
Woodward, J.J. and Smith, L.S., 1985. Exercise training and the stress response in rainbow trout (Salmo gairdneri) . J. Fish Biol., 26: 435-447.





























