Embed Size (px)
Citation preview

RESEARCH ARTICLE
Groucho Corepressor Proteins Regulate OticVesicle OutgrowthBaubak Bajoghli,† Narges Aghaallaei,† and Thomas Czerny*
The Groucho/Tle family of corepressor proteins is known to regulate multiple developmental pathways.Applying the dominant-negative effect of the short member Aes, we demonstrate here a critical role of thisgene family also for ear development. Misexpression of Aes in medaka embryos resulted in reduced size orloss of otic vesicles, whereas overexpression of the full-length Groucho protein Tle4 gave the oppositephenotype. These results are in close agreement with phenotypes observed for eye formation, suggesting asimilar role for Groucho/Tle proteins in the developmental pathways of both sensory organs. Furthermore,by using the heat-inducible HSE promoter, we observed reversible branching of the embryonic axis uponAes misexpression, indicating a transient duplication of the organizer. Groucho proteins, therefore, arecritical for organizer maintenance. Developmental Dynamics 233:760–771, 2005. © 2005 Wiley-Liss, Inc.
Key words: Medaka; Groucho; Aes; heat-inducible misexpression; otic vesicle; eye development; lens placode; axisduplication
Received 3 October 2004; Revised 21 January 2005; Accepted 22 January 2005
INTRODUCTION
Groucho or Transducin-like Enhancerof Split (Tle) proteins serve as non-DNA binding corepressors for specificsubsets of DNA binding transcriptionfactors. Upon interaction, they canswitch the activating potential of itsbinding partner to a repressing function(Valentine et al., 1998; Eberhard et al.,2000; Cai et al., 2003). Members of theGroucho/Tle family have been identifiedin vertebrates such as humans (Stifaniet al., 1992), mice (Miyasaka et al.,1993), Xenopus (Choudhury et al.,1997), zebrafish (Wulbeck and Campos-Ortega, 1997), and medaka (Lopez-Rioset al., 2003).
Groucho/Tle members are widely ex-pressed both during development andin the adult, in contrast to the more
limited expression pattern of their DNAbinding partners (Hartley et al., 1988;Stifani et al., 1992; Miyasaka et al.,1993; Schmidt and Sladek, 1993;Choudhury et al., 1997; Jimenez et al.,1997; Pflugrad et al., 1997; Sharief etal., 1997). The expression of Groucho/Tle members in diverse sensory organsindicates a role of this gene family dur-ing development of these organs. Forexample, it has been shown that Tle1and Tle4 are expressed in the develop-ing eye of medaka fish. In addition, Tle3has been found in the lens placodes ofmice (Leon and Lobe, 1997) andmedaka (Lopez-Rios et al., 2003). In Xe-nopus, Grg4 and Grg5 are expressed inotic vesicles (Molenaar et al., 2000).
An interesting member of Groucho/Tle family is Amino Enhancer of Split
(Aes) or Grg5, which exhibits strongsimilarity to the amino-terminal do-mains of other family members butlacks the C-terminal WD-40 repeats(Miyasaka et al., 1993). Aes/Grg5genes have been identified from vari-ous vertebrates (Schmidt and Sladek,1993; Choudhury et al., 1997; Mo-lenaar et al., 2000; Lopez-Rios et al.,2003). During early embryonic devel-opment, it is widely expressed, switch-ing to a tissue-specific pattern inadults (Choudhury et al., 1997; Mo-lenaar et al., 2000). The amino-termi-nal part of Aes/Grg5 harbors the Q-domain, a multimerization motif,facilitating interactions with otherGroucho family members. Lack ofbinding motifs for histone deacety-lases (HDAC; Brantjes et al., 2001),
Institute of Animal Breeding and Genetics, University of Veterinary Medicine, Vienna, AustriaGrant sponsor: Austrian Science Fund (FWF); Grant number: P15185.†B. Bajoghli and N. Aghaallaei contributed equally to this work.*Correspondence to: Thomas Czerny, Institute of Animal Breeding and Genetics, University of Veterinary Medicine, Veteri-narplatz 1, A-1210 Vienna, Austria. E-mail: [email protected]
DOI 10.1002/dvdy.20398Published online 28 April 2005 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 233:760–771, 2005
© 2005 Wiley-Liss, Inc.

nevertheless, interferes with bindingof this major repressive component toGroucho multiprotein complexes. Thederepression effect of Aes/Grg5 is fur-ther enhanced by the absence of theWD-40 repeats, responsible for mostprotein–protein interactions (Chen andCourey, 2000). This dominant-negativefunction has successfully been appliedto various experimental systems (Rooseet al., 1998; Ren et al., 1999).
Otic induction in vertebrates startsat mid-gastrula stage, initiated by fac-tors from hindbrain and subjacent me-soderm (reviewed in Whitfield et al.,2002; Riley and Phillips, 2003). Thisprocess occurs in a series of steps thatare likely to be regulated indepen-dently (Groves and Bronner-Fraser,2000; Liu et al., 2003). The best can-
didates for otic-inducing factors aremembers of the fibroblast growth fac-tor (Fgf) family of peptide ligands. Inparticular, Fgf8 and Fgf3 are suffi-cient to ectopically induce otic vesicles(Lombardo et al., 1998; Vendrell et al.,2000; Bajoghli et al., 2004; Phillips etal., 2004). In zebrafish acerebellar mu-tants, which contain a disrupted Fgf8gene, the otic vesicles show a reducedsize (Whitfield et al., 1996; Reifers etal., 1998). The same phenotype hasbeen observed in morpholino knock-down experiments for Fgf3 (Phillips etal., 2001; Leger and Brand, 2002; Ma-roon et al., 2002). Another candidatefor an otic-inducing factor is Wnt8. Inchick, an ortolog of the Wnt8 gene(Wnt8c) is expressed in the hindbrainbetween the two prospective otic anla-
gen and human Wnt8 can induce avariety of otic markers in chick em-bryos (Ladher et al., 2000). In ze-brafish, the indirect function of Wnt8in this process has been revealed; dis-ruption of the gene leads to a delay inpreotic expression of Fgf8 and Fgf3,thus, indirectly causing otic vesicles ofreduced size (Phillips et al., 2004). TheSox gene family also plays an impor-tant role in the development of thevertebrate ear (de Martino et al.,2000; Groves and Bronner-Fraser,2000; Koster et al., 2000; Chiang etal., 2001; Dutton et al., 2001; Saint-Germain et al., 2004). For example,overexpression of Sox3 in medakaleads to the formation of ectopic oticvesicle-like structures (Koster et al.,2000), whereas inactivation of Sox10
TABLE 1. Phenotypic Effects of Aes Misexpression in Medaka Embryos
DNA concentration 10 ng/�l 30 ng/�l 50 ng/�l
Number of induced embryosa 134 180 284Dead embryos 20 26 97Surviving embryos 114 154 187GFP positive after
inductionb105 (92%) 150 (97%) 170 (91%)
Malformations (%)c
Otic vesicle defects 9 (8.5%) 34 (23%) 54 (32%)Eye defects 8 (7.6%) 29 (19%) 57 (33%)Mid-hindbrain defects 7 (6.6%) 15 (10%) 45 (26%)Heart position 4 (4%) 23 (15%) 36 (21%)
aEmbryos were injected at the one-cell stage and GFP negative embryos were selected for heat treatment (39°C/2h) at the earlygastrula stage.
bPercentages were calculated from the surviving embryos.cPercentages were calculated from the GFP positive embryos.
TABLE 2. Eye and Otic Vesicle Phenotypes Observed after Aes Misexpression
Stage of activation one-cellaMid-Blastulab
Early-Gastrulac,d
Mid-Gastrulac
Late-Gastrulac 2-Somitec
Totally induced embryos 145 150 180 137 67 84Dead embryos 84 43 26 26 3 15Surviving embryos 61 107 154 111 64 69GFP positive after
induction- - 150 91 60 67
Malformations (%)e
Reduced otic vesicle 13% 15% 21% 18% 17% 19%Loss of otic vesicle 23% 6% 2% 2% 0% 0%Reduced eye size 18% 11% 14% 15% 13% 16%Loss of eyes 21% 5% 5% 5% 0% 0%
a,bThe concentrations for mRNA (a200 ng/�l) and DNA (bCMV:Aes; 200 ng/�l) were adjusted to result in similar percentages ofreduzed size phenotypes.
cThe GFP:HSE:Aes construct was injected at 30ng/�l. GFP negative embryos were selected for heat treatment (2h/39°C).dThese data correspond to those presented in Table 1 and are included for comparison.ePercentages were calculated from surviving embryos with GFP expression after induction.
Groucho REGULATES OTIC VESICLE OUTGROWTH 761

Fig. 1.
Fig. 2.
Fig. 3.
762 BAJOGHLI ET AL.

in colourless mutants causes defectsin patterning of the otic vesicle andsmall ears (Whitfield et al., 1996).Pax8 a member of Pax2/5/8 family isone of the earliest markers of preoticdevelopment, first detectable at lategastrulation in zebrafish (Pfeffer etal., 1998) and Xenopus (Heller andBrandli, 1999). Loss of both Fgf8 andFgf3 function prevents Pax8 expres-sion in the ear primordium (Phillips etal., 2001; Leger and Brand, 2002). Al-though inactivation of Pax8 in micehas no effect on ear development(Mansouri et al., 1998), knock-down ofPax8 in zebrafish delays developmentof the otic placode (Hans et al., 2004).
Nothing is known about the role ofGroucho/Tle proteins during ear for-mation. In this report, therefore, weinvestigated the function of this genefamily in early otic development. For
this purpose, we applied the dominant-negative effect of Aes/Grg5 on otherfamily members. Using the recently de-veloped heat-inducible HSE-system formisexpression of Aes (Bajoghli et al.,2004), we observed hypoplasia of oticvesicles and eyes. These results werecomplemented by Tle4 overexpressionexperiments giving opposite pheno-types. In addition, we report that ec-topic Aes is able to induce partial axisduplications in medaka embryos.
RESULTS
Blocking GrouchoCorepressor Function inOtic Vesicles byMisexpression of Aes
The equilibrium between activatingand repressive functions is of major
importance for developmental deci-sions and Groucho proteins are keyfactors in such processes. We testedthe effects of these corepressors onotic vesicle formation. As a tool, weused Aes, a truncated Groucho pro-tein, known for its antagonistic func-tions on other members of the verte-brate Groucho family (Roose et al.,1998; Ren et al., 1999; Lopez-Rios etal., 2003). For misexpression, wechose the heat-inducible HSE system(Bajoghli et al., 2004), allowing ectopicactivation of the gene-of-interest in astage-dependent manner. The em-bryos were first induced during earlygastrulation, and the effects of Aesmisexpression were observed 48 hourslater. Between 91 and 97% of the sur-viving embryos developed green fluo-rescent protein (GFP) fluorescence(Table 1). GFP is used as a marker
TABLE 3. Effects of Aes and Tle4 Misexpression on Endogenous Eya1, Six1, and Sox3
Aes misexpression Tle4 misexpression
Sox3 Six1 Eya1 Eya1
Ectopic Repressiona Ectopic Repressiona Ectopic Repressionb Ectopicc Repression
0/31 10/31 0/31 7/31 0/26 17/26 18/43 0/43
aEmbryos with Sox3 or Six1 repression in the otic vesicle region.bEmbryos with Eya1 repression in the somites.cEmbryos with ectopic Eya1 in rhombencephalic ectoderm.
Fig. 1. Misexpression of Aes leads to size reduction or loss of otic vesicles. The heat-inducible Aes construct was coinjected with meganucleaseenzyme into one- or two-cell stage embryos. All embryos were heat treated at early gastrula (stage 13) for 2 hr at 39°C and analyzed for phenotypes2 days after induction. A: An embryo with a typical Aes-induced phenotype displays reduced size of the eye (arrowhead) and the otic vesicle (arrow).The midbrain is dramatically reduced, and the heart tube position is shifted by 90° to the affected side. B,B�: Embryos with smaller otic vesicles (B)exhibited green fluorescent protein (GFP) activity only at the affected side (B�; the dotted line demarcates the normal otic vesicle that is GFP-negative).C: An otic vesicle that started splitting. C�: GFP activity at the indentation indicates the involvement of Aes in this process. D,D�: An embryo with lossof an otic vesicle (D) in agreement with high levels of GFP activity (D�). The heart tube (h) of this embryo also pointed to the side of high misexpressionlevels. In all embryos, anterior is to the top (dorsal views). h, heart; mb, midbrain; ov, otic vesicle. Scale bar in A � 100 �m for A, 40 �m for B,B�, 35�m for C,C�, 60 �m for D,D�.
Fig. 2. Expression of otic vesicle marker genes after Aes activation. A–K: Dorsal (A–F) and lateral (G–K) views of embryos at stage 25 (19 somites)injected with HSE:Aes and induced at early gastrula stage (B,C,E,F,H,I,K). Anterior is to the top (A–F) or to the left (G–K). A,D: Wild-type expressionof Pax2 (A) and Eya1 (D) is shown. B,E: The expression of Pax2 (B) and Eya1 (E) was unaffected in embryos with reduced otic vesicle size. The bracketsindicate the different size of the otic vesicle territory in the affected and the normal half of the embryo. C,F: Loss of otic vesicles was associated withlack of Pax2 (C, arrowhead) and Eya1 (F, arrowhead) expression in this region. G: Sox3 is strongly expressed in the rhombencephalic ectoderm ofwild-type embryos. H: Misexpression of Aes leads to repression of Sox3 in this region (arrowhead). I: Same embryo as in H, but a different optical planeand an increase of background light makes visible the reduced size of the otic vesicle (marked by a red dotted line). J,K: Overexpression of Aes (K)represses the expression of Six1 in the otic vesicle region. MHB, midbrain–hindbrain boundary. Scale bars � 50 �m in A; 40 �m in B–F,J,K; 20 �min G–I.
Fig. 3. Misexpression of Tle4 leads to enlargement of otic vesicles. A–F: Dorsal views of embryos at stage 25 (A,B; 19 somites), stage 24 (C,E,F; 16somites) and stage 22 (D; 9 somites) with Tle4 induced at early gastrula stage; anterior is to the top (A–E) or to the left (F). A,B: Misexpression of Tle4resulted in embryos with enlarged otic vesicles (A), which correlated with the green fluorescent protein activity in the affected region (B; the dotted linedemarcates the normal otic vesicle). C,D: Expression of Eya1 (C) and Pax2 (D) was not affected by the enlargement of the otic vesicle. E: Ectopic oticvesicle-like structures were observed after Eya1 marker gene analysis (arrow; inset, lateral view, anterior to the left, dorsal to the top). F: The lensplacode was enlarged (arrow; the dotted line demarcates the eye field), or in rare cases, the eye became lost (arrowhead). Scale bar in A � 50 �mfor A,B, 35 �m for C, 25 �m for D; 40 �m for E, 65 �m for inset in E, 100 �m for F.
Groucho REGULATES OTIC VESICLE OUTGROWTH 763
gene in the HSE construct, indicatingthe efficiency of misexpression in theembryos.
As hypothesized, we found dramaticeffects on otic development for Aesmisexpression. The total number ofotic vesicle defects depended on theAes expression level. Increasing theDNA concentration from 10 to 50ng/�l concomitantly raised the ap-pearance of phenotypes from 9 to 32%(Table 1). The most prominent malfor-mation was a size reduction of the oticvesicles; typically, the affected organswere half the size of those of wild-typeembryos. In rare cases, they becamesplit or were lost completely. Aes,therefore, negatively affects otic vesi-cle outgrowth.
Consistent with the idea that in-terference with Groucho proteinfunction affects multiple develop-mental pathways, we observed addi-tional malformations throughout theembryo, most obvious in the eyes,Fig. 4.
Fig. 5.
764 BAJOGHLI ET AL.

the midbrain–hindbrain region, andthe heart. The size of the eye wasreduced (Fig. 1A, arrowhead), in ex-treme cases leading to complete ab-sence of the eye. This strong effect oneye development has been describedpreviously for Aes mRNA injectionexperiments (Lopez-Rios et al., 2003)and, therefore, served as a referencefor Aes activity. In the midbrain–hindbrain region, we saw a dramaticreduction of midbrain size (Fig. 1A);in some cases, a quite variable phe-notype was observed, which turnedout to be caused by a partial axisduplication (see section Induction ofAxis Duplications by Aes in the Re-sults section). Furthermore, in Aes-injected embryos, the position of theheart tube appeared altered, point-ing straight away from the bodyaxis, either to the right side (Fig. 1A)or the left side of the embryo (Fig.1D). All described phenotypesshowed a strict dependence on Aesdose and appeared at similar ratios(Table 1).
We often observed a coincidence ofthe malformations on the same side ofthe embryos, either right or left. Typ-ical embryos developed, on the af-fected side, small eyes and small oticvesicles, a largely reduced midbrain,
and a heart tube pointing to the sameside (Fig. 1A). Such embryos appearedat a frequency of 15% for 50 ng/�l. Acomparison of the affected side withthe wild-type reference side of the em-bryo was of particular importance forqualifying size differences of the eyesand the otic vesicles. We thus couldunequivocally conclude a size reduc-tion for both sensory organs as a re-sult of Aes misexpression. Further ev-idence came from analysis of GFPexpression patterns. GFP is expressedin a bidirectional manner from theHSE promoter, thus marking Aes-misexpressing cells (Bajoghli et al.,2004). Tracing GFP fluorescence indi-cated a strict overlap of the describedphenotypes with high misexpressionlevels. Examples for hypoplasia orcomplete loss of otic vesicles areshown in Figure 1B and 1D, respec-tively. Splitting of the otic vesicle wasobserved rarely but also was corre-lated with GFP expression in an areawhere the splitting was initiated (Fig.1C). Later, a complete separation intotwo vesicles took place (data notshown). The heart tube strictlypointed to the side of high GFP ex-pression (Fig. 1D�). Thus, interferencewith Groucho function generates a re-producible phenotype in the embryo.
Time Dependence of the Aes-Induced Phenotypes
Deviations between the results ob-tained for the HSE-inducible system,compared with mRNA injections(Lopez-Rios et al., 2003; and J. Lopez-Rıos, personal communication) sug-gested a time dependence of the ef-fects of ectopic Aes activity in theembryos. As a major advantage, in-ducible systems can be activated dur-ing different time points, thus cover-ing different windows of competencefor developmental decisions. We,therefore, performed a time-course ex-periment by systematically varyinginduction from early gastrula to thetwo-somite stage (Table 2). To qualifyearlier stages, we included mRNA(translated immediately) and a DNAexpression construct (transcribed dur-ing mid-blastula). A reduction of oticvesicle size was observed for all timewindows of Aes activation at compara-ble rates, whereas complete loss was aphenotype preferentially seen forearly induction (23% for mRNA injec-tion). Almost identical results, bothfor size reduction and loss were ob-served for eye vesicles.
Misexpression of Aes Resultsin Repression of Six1 andSox3 in Otic Vesicles
To learn more about the molecularmechanisms, we investigated the ef-fect of ectopic Aes on otic placodemarker genes. We analyzed the ex-pression of Pax2, Eyes Absent 1(Eya1), Six1, and Sox3. In all verte-brates, Pax2 is expressed in preoticcells during early somitogenesis (Pfef-fer et al., 1998; Heller and Brandli,1999) until organogenesis (Whitfieldet al., 2002). Eya1 and Six1 are ex-pressed early on in the preplacodaldomain and later show distinct ex-pression patterns in the otic placode(Sahly et al., 1999; Koster et al., 2000;Bessarab et al., 2004). In medaka,Sox3 expression is found in all sensoryplacodes (Koster et al., 2000). Thesefour marker genes, therefore, can beused to analyze formation of the oticplacode.
Aes-injected embryos were first an-alyzed for Pax2 expression by whole-mount in situ hybridization (Fig.
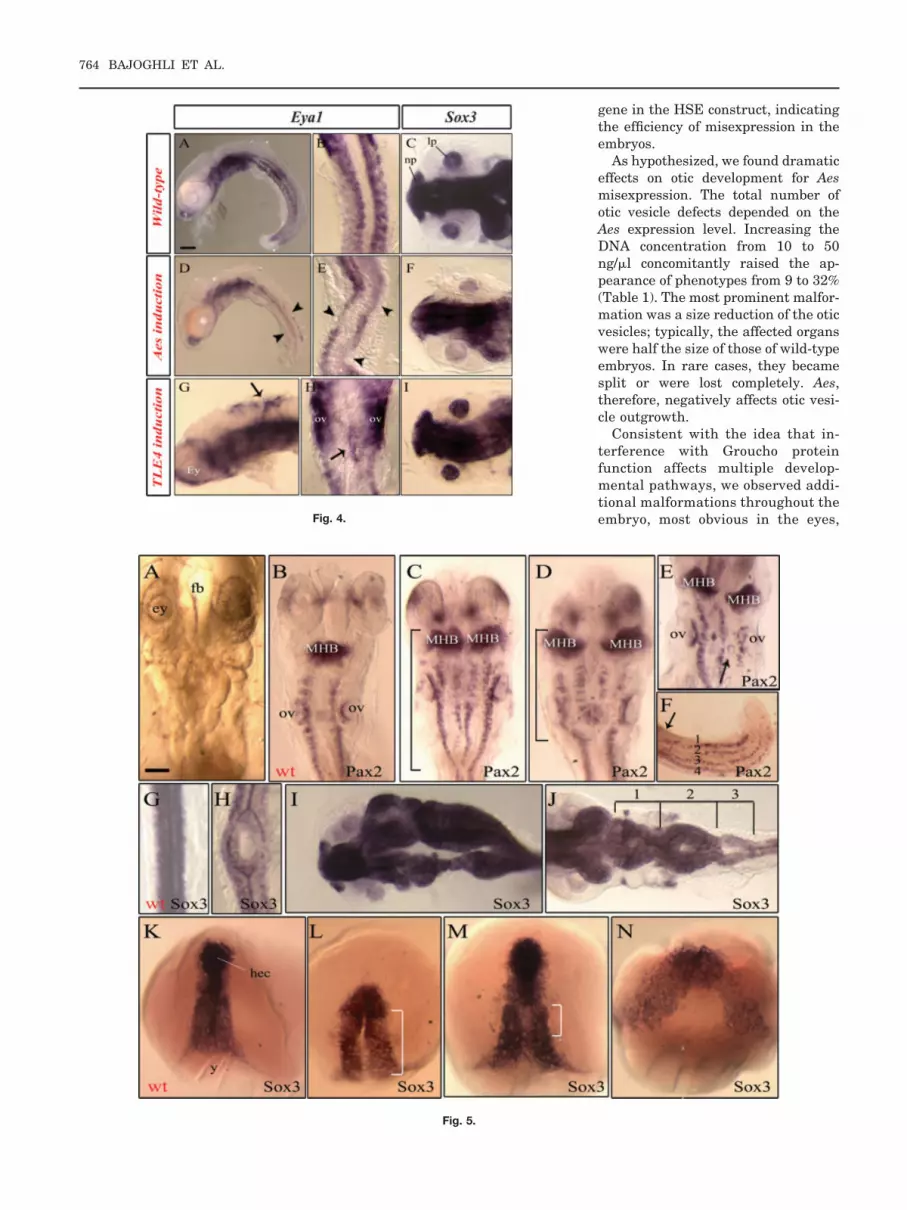
Fig. 4. Opposing effects of ectopic Tle/Aes on Eya1 and Sox3 expression. A–I: Lateral views (A,D,G)or dorsal views (B,C,E,F,H,I) of embryos at stage 27 (A,D; 24 somites), stage 24 (B,C,E,F,G,H,I; 16somites). In B,E, and H, anterior is to the top. Embryos were injected either with the Aes- or withthe Tle4-inducible construct. All injected embryos were heat induced at the same time during earlygastrula stage (for 39°C/2 hr). D,E,GH: Ectopic Aes leads to reduction of Eya1 in somites (D,E;arrowheads), but overexpression of Tle4 induces ectopic Eya1 in ectoderm (G,H; arrows). F,I: Theexpression of Sox3 in the lens placode was reduced in Aes-misexpressed embryos (F, arrowhead)in contrast to Tle4-injected embryos (I). ey, eye; lp, lens placode; nl, nasal placode; ov, otic vesicle.Scale bar in A � 25 �m for A,D, 50 �m for C,F,H,I, 70 �m for B,E, 20 �m for G.
Fig. 5. A–N: Induction of Aes at early gastrula stage leads to partial axis duplication. A,C–F,L–N: Dorsalviews of embryos at stage 26 (A,C–F; 22 somites), stage 16 (L,M; late gastrulation), and stage 15 (N;mid-gastrulation) injected with the Aes-inducible construct at the one-cell stage. All embryos wereinduced at early gastrula (stage 13) at 39°C for 2 hr. Anterior (except F,I,J, left) is to the top. C,D,L,M:The brackets indicate the length of the duplication. A: An embryo with partial axis duplication in themidbrain–hindbrain region. C–F,H–N: This phenotype was analysed for Pax2 (C–F) and Sox3 (H–N)expression. C–E: Embryos with partial axis duplication display a duplicated midbrain–hindbrain bound-ary (MHB) and spinal cord neurons. The position and length of axis duplication differed betweenembryos (compare C and D). C–E,H,I: The axis duplication was observed in the anterior part (C–E,I) aswell as the posterior part of the embryos (F,H). F: Embryos with four rows of spinal cord neurons in thetail region (arrow indicating the beginning of the duplication). E: In some cases, otic vesicles wereobserved between the two neural tubes (arrow). J: We observed embryos with sequential branchingand fusion of the neural tube (the bracket and numbers mark the duplicated regions). L–N: Confirmingthe phenotype at later stages, analysis of embryos at late gastrula also revealed axis duplications.Strong green fluorescent protein–positive embryos were selected for whole-mount in situ hybridizationagainst Sox3. L–N: Variations in the extent of the duplicated region became visible, but head ectodermwas never affected. ey; eye; fb, forebrain; hec, head ectoderm; MHB, midbrain–hindbrain boundary; ov;otic vesicle; wt, wild-type; y, yolk. Scale bar in A � 100 �m for A,B,J, 80 �m for C–E, 60 �m for F–H,120 �m for I,K–N.
Groucho REGULATES OTIC VESICLE OUTGROWTH 765

2B,C). In general, the Pax2 expressionpattern appeared normal in otic vesi-cles with reduced size (Fig. 2B), al-though some early induced embryosdeveloped weak repression of thisgene (data not shown). On the con-trary, all endogenous Pax2 in this re-gion disappeared for embryos, whichdisplayed complete loss of an otic ves-icle (Fig. 2C, arrowhead). Eya1 ex-pression showed the same behaviorupon size reduction or loss of otic ves-icles (Fig. 2E,F). Of interest, in 65% ofthe embryos Eya1 expression in thesomites was reduced (17 of 26; Table3; Fig. 4D,E, arrowheads).
On the contrary, Six1 and Sox3 ex-pression was strongly repressed inotic vesicles of reduced size at a fre-quency of 23% (7 of 31) and 32% (10of 31), respectively (see Table 3). InFigure 2K,H, examples of this re-pression are shown. In some em-bryos, Sox3 expression in the lensplacodes also was reduced (4 of 31;Fig. 4F). Taken together, hypoplasiaof otic vesicles induced by Aes is ac-companied by normal expression forEya1, whereas Sox3 and Six1 ex-pression is reduced.
Overexpression of Tle4Leads to an Enlargement ofthe Otic Vesicle and EctopicActivation of Eya1
Our strategy was based on the inter-ference of Aes with Groucho proteinfunction. A clear prediction was,therefore, an opposite effect uponoverexpression of a full-length Grou-cho family member. We selected Tle4for this experiment, based on the ex-pression of this gene in Xenopus oticvesicles (Molenaar et al., 2000). Usingthe HSE system, we induced Tle4misexpression during early gastrula-tion. Upon low concentrations (10 ng/�l), 7% of the embryos (6 of 81) showedenlarged otic vesicles. Higher Tle4concentrations increased the percent-age of this phenotype to 13% (5 of 38)for 20 ng/�l and to 15% (17 of 111) for60 ng/�l. Regions of high Tle4 misex-pression correlated with this pheno-type, as concluded from GFP markergene expression (Fig. 3A,B). Most im-portantly, reduction of otic vesicle sizewas not detected in a single case,whereas complete loss of the eye ap-peared as a rare phenotype (Fig. 3F,
arrowhead). In addition, we observedin few embryos an enlargement of thelens (2 of 43; Fig. 3F, arrow), but at alow frequency compared with the al-terations of the otic vesicles. Whencoinjecting 20 ng/�l Tle4 together with20 ng/�l Aes (HSE-constructs inducedduring early gastrulation), no en-larged otic vesicles were detected anymore (n � 70) and, similarly, the re-duced otic vesicle phenotype of Aeswas largely reduced due to the pres-ence of the agonist (data not shown).These data further strengthen the op-posing effects of both proteins on oticvesicle outgrowth.
Tle1 overexpression experimentsresulted in enlargement of the eyefield, in addition Tle1 was shown toinduce patches of ectopic Six3 andPax6 expression, indicating the poten-tial of Groucho proteins to assist inthe formation of ectopic eye structures(Lopez-Rios et al., 2003). Using theHSE induction system, we could de-tect the formation of additional oticvesicles upon Tle4 overexpression. Foridentification, we used Eya1 as amarker (Fig. 3E, arrow). Contrary to asplitting of the vesicles, which we ob-served for Aes misexpression (Fig. 1C;adjacent vesicles), the position ofthese ectopic sensory organs seen forTle4 overexpression was quite distantfrom the endogenous vesicles (Fig. 3E,inset; ectopic vesicle at the dorsal sideof hindbrain).
As expected, Pax2 and Eya1 expres-sion within the enlarged otic vesicleswas normal (Fig. 3C,D). Contrary torepression of Eya1 induced by Aes,Tle4 overexpression led to ectopicEya1 in rhombencephalic ectoderm ata frequency of 42% (18 of 43; Fig.4G,H). Taken together, these datastrongly support a positive effect offull-length Groucho proteins for oticvesicle outgrowth in contrast to Aesand confirm the size determining ef-fect of this corepressor in developmentof the otic vesicle.
Induction of AxisDuplications by Aes
In several cases, Aes-injected embryosdeveloped a variable phenotype inneural tube-derived structures. In se-verely affected embryos, a duplicatedmidbrain–hindbrain region becamevisible (Fig. 5A). To get a clearer pic-
ture, we analyzed the expression ofthe midbrain–hindbrain boundarymarker Pax2. In situ hybridizationdata for this gene clearly indicated aduplication of the anterior neural tube(Fig. 5C–F). The two neural tubeswere completely separated, but other-wise showed a normal expression pat-tern for Pax2, a marker for differenti-ating neurons along the neural tube.Of interest, the duplication was par-tial, resulting in a fusion of the neuraltube posteriorly. We did not see fore-brain structures affected, but other-wise observed branching at differentpositions along the axis (Fig. 5F). Thelength of the duplicated region dif-fered between the embryos (Fig.5C,D). In extreme cases, where thetwo neural tubes were clearly sepa-rated from each other, we even ob-served otic vesicles at the expected po-sition between the two neural tubes(Fig. 5E, arrow).
Because misexpression with theHSE promoter generates a peak of Aesmisexpression, slowly declining withtime, we reasoned that position of thepartial duplication of the axis mightdepend on the time of induction. We,therefore, induced embryos at differ-ent time points during gastrulationand analyzed for axis duplication bywhole-mount in situ hybridizationagainst Pax2. We did not detect anytime dependence of this phenotype be-tween early- and mid-gastrula, butembryos induced during late gastruladid not display partial axis duplica-tions any more (data not shown).mRNA injections of Aes resulted inaxis duplications at a very low fre-quency (2 of 61; data not shown).
To back up the results for Pax2, weanalyzed the embryos with Sox3, amarker for neuroectoderm along theentire body axis (Koster et al., 2000).Labeling the neural tube with Sox3even better visualized the partial axisduplication (Fig. 5H–J). Applying thehigher resolution of this marker, weobserved embryos where splitting andfusion of the neural tube appearedseveral times along the axis (Fig. 5J).Of interest, we did not detect a singlecase of a complete axis duplication,suggesting that after the decline ofAes misexpression, axis induction con-tinues normally, resulting in a fusionof the duplicated structures.
To examine earlier stages of axis
766 BAJOGHLI ET AL.

induction, we analyzed Sox3 expres-sion during late gastrulation (Aes in-duced at early gastrula). The duplica-tion could clearly be detected at thisearly stage (Fig. 5L–N) and appearedin GFP-expressing embryos at a fre-quency of 36% (17 of 47). The extentand distance of the duplicated neuro-ectodermal structures differed dra-matically (compare Fig. 5L–N). There-fore, Aes transiently interferes withnormal axis induction.
DISCUSSION
The study of mutants is the keymethod for analysis of gene function.Unfortunately, several gene familiesexhibit a high degree of functionaloverlap. A typical example is theGroucho/Tle family. The amino acidsequences of these proteins are highlyconserved between different Grouchofamily members, in particular in theC-terminal WD-40 repeats, the majorprotein–protein interaction domain(Choudhury et al., 1997; Fisher andCaudy, 1998; Li, 2000). This findingexplains why diverse Groucho/Tlemembers often interact with the sametranscription factors (Choi et al.,1999; Ren et al., 1999; Eberhard et al.,2000). Therefore, the analysis of mu-tants or morpholino knockdown ex-periments will only cover subsets ofthe complete Groucho corepressorfunction. The Groucho-related proteinAes/Grg5 lacks the WD-40 repeats,thus blocking many protein–proteininteractions. Due to deviations in theamino acid sequence, this protein doesnot interact with HDAC1 (Brantjes etal., 2001), a major source of repressivefunction conferred by Groucho pro-teins (Chen et al., 1999; Brantjes etal., 2001). With its N-terminal Q-do-main, Aes/Grg5 nevertheless can mul-timerize with other Groucho proteins,thus explaining the dominant-nega-tive effect seen for this short memberof the Groucho gene family (Roose etal., 1998; Ren et al., 1999).
By overexpressing Aes in medakaembryos, we applied the suppressiveeffect on all Groucho family membersat the same time. This strategy re-sulted in a typical phenotype showingeyes, midbrain, and otic vesicles of re-duced size. A critical control for thisdominant-negative approach was theapplication of full-length Tle proteins.
Shifting the equilibrium toward therepressive function, overexpression ofTle4 indeed resulted in opposite phe-notypes.
Groucho/Tle ProteinsStrongly Affect Otic VesicleDevelopment
The role of Groucho proteins in oticvesicle development has not been in-vestigated to date. Expression analy-sis has shown that in Xenopus, Grg4,and Grg5 are active during early oticvesicle development (Molenaar et al.,2000). In zebrafish Gro1 (a Tle3 homo-logue) transcripts were also detectedin this region (Wulbeck and Campos-Ortega, 1997). In medaka, neitherTle1, Tle3, or Tle4 are expressed inearly otic vesicles (Lopez-Rios et al.,2003), but we have data that two newfull-length Groucho family membersare activated in this region (N.Aghaallaei, B. Bajoghli, I. Walter, andT. Czerny, submitted).
When interfering with Groucho pro-tein function, we observed two differ-ent phenotypes in ear development,namely absence or reduced size of theotic vesicle. Loss of this structure inprinciple could be the extreme versionof a reduced size phenotype, but dif-ferences in time dependence indicatean involvement of Groucho proteinfunction in two separate developmen-tal pathways. Misexpression of Aes bymRNA injection generated embryoswith loss of otic vesicles at a high fre-quency (23%). The same phenotypewas rarely observed when Aes was in-duced at mid-gastrula (2%) but wasnot detected for Aes activation at laterstages. On the contrary, size reductionof the otic vesicle was observed atroughly the same frequency (between13 and 21%) for all Aes activationtimes tested (up to the two-somitestage). In addition, unspecific pertur-bation effects of surrounding tissue,preventing proper induction, cannotbe excluded for loss of otic vesicles.Take together, an early involvementof Groucho protein function is sug-gested for preplacodal stages (initia-tion phase), whereas a continuouslyacting system determining otic vesiclesize is sensitive to Aes during allstages (maintenance phase).
To characterize the molecular basis ofthese phenotypes, we analyzed the ex-
pression of otic vesicle marker genes. Inembryos lacking an otic vesicle due toAes misexpression, transcripts for Pax2were lost in this region, whereas other-wise Pax2 expression was almost nor-mal in these embryos. Pax2 is expressedearly in otic placodes (three-somitestage) and has an important regulatorfunction during subsequent otic devel-opment (for review, see Riley and Phil-lips, 2003). Nevertheless, disruption ofmouse Pax2 does not alter otic induc-tion (Torres et al., 1996) and does notaffect Eya1 and Six1 expression (Xu etal., 1999; Zheng et al., 2003; Burton etal., 2004). Eya1 is expressed earlier(end of gastrulation) than Pax2 and af-fects the early inductive signallingevents involved in otic vesicle forma-tion, as demonstrated by Eya1-deficientmice (Xu et al., 1999). Although not af-fected in formed otic vesicles, Aes misex-pression resulted in a dramatic down-regulation of Eya1 in the somites.Furthermore, ectopic expression wasdetected in rhombencephalic ectodermupon Tel4 overexpression, thus clearlyindicating the sensitivity of this gene toGroucho activity, which similarly mightplay a role in early induction of oticvesicles. Even more striking was the re-duction of Six1 expression for vesicles ofreduced size. A highly conserved net-work of interaction has been describedfor Pax-Six-Eya-Dach genes acting dur-ing organogenesis (Bonini et al., 1997;Heanue et al., 1999; Li et al., 2003).Coexpression of these genes during oticdevelopment suggests functional inter-actions, strongly supported by gene in-activation experiments. Tle/Aes misex-pression, therefore, might criticallyaffect correct formation of this gene net-work during otic vesicle formation.
Furthermore, Sox3 expression inthis region appeared strongly re-duced. Sox3 has been implicated pre-viously in sensory organ development(Koster et al., 2000). Misexpression ofSox3 in medaka embryos leads to theformation of ectopic otic vesicle struc-tures, which express the markersPax2 and Eya1. Overexpression of adominant-negative form of Sox3,which encodes the DNA-bindingHMG-domain without transactivationdomain, leads to dysgenesis of endog-enous sensory organs (Koster et al.,2000). Thus, Sox3 is also a good can-didate to control both initiation andsize regulation of these organs.
Groucho REGULATES OTIC VESICLE OUTGROWTH 767

Groucho/Tle proteins exhibit theirfunction by binding to other transcrip-tion factors. The question, therefore,arises, which up-stream regulatorscould confer the Aes effect on Six1,Eya1, and Sox3. Among the genes ex-pressed during otic vesicle develop-ment, members of the Pax2/5/8 sub-family have been identified asinteraction partners for Groucho/Tleproteins (Eberhard et al., 2000; Cai etal., 2003). As discussed before, Pax2inactivation does not affect Eya1 orSix1 expression and Pax5 is expressedlater during inner ear development.On the contrary, Pax8 can be detectedin preotic cells during late gastrula-tion in all vertebrates, representingone of the earliest markers in this re-gion (Pfeffer et al., 1998; Heller andBrandli, 1999). Loss of Pax8 functionin the mouse does not exhibit an oticphenotype (Mansouri et al., 1998), butknockdown of Pax8 in zebrafish delaysotic placode development (Hans et al.,2004). This variability of phenotypescould be explained by the high degreeof functional overlap between mem-bers of the Pax2/5/8 subfamily. Pax8and Pax2 are sequentially expressed,and combined inactivation of bothgenes in zebrafish results in completeloss of otic vesicles (Hans et al., 2004).Interaction with Groucho proteins hasbeen described also for members of theSix-family (Kobayashi et al., 2001;Zhu et al., 2002; Lopez-Rios et al.,2003). Thus, Six3 could be identifiedto confer the effect of Aes/Tle misex-pression on eye development, result-ing in similar phenotypes comparedwith ear formation (see below). Here,autoregulation of Six3 could be maderesponsible for the effect on its ownexpression (Zhu et al., 2002) andmight similarly affect Six1. Therefore,currently Six1 is the best candidatefor a critical target of Groucho proteinfunction during otic vesicle formation.
Mechanistically, the effect of Grou-cho proteins on the formation of oticvesicles and eyes is quite unexpected.Down-regulation of the repressing ac-tivity by Aes misexpression leads toreduced size or complete loss, whereasenhanced repression results in hyper-plasia of these sense organs. In a lin-ear genetic pathway, the effect ofGroucho proteins, therefore, could betransmitted only indirectly by anotherfactor, which itself would suppress
sensory organ development. In princi-ple, Tcf/Lef transcription factorswould be good candidates for trans-mission of a Groucho effect. They areknown to interact with Groucho fam-ily members (Brantjes et al., 2001),making the canonical wnt-signallingpathway dependent on the Tle/Aesequilibrium. In zebrafish, wnt signal-ling has been proposed to play a rolein otic placode induction, by regulat-ing expression of Fgf3 and Fgf8 in thehindbrain. Furthermore, injection ofWnt antagonists (GSK3b; Saint-Ger-main et al., 2004) and Wnt8 morpho-lino oligos (Phillips et al., 2004) re-sulted in otic vesicles of reduced size.Therefore, Tcf/Lef proteins play apositive role in the proposed path-way, the phenotypic consequencesfor Tle/Aes misexpression neverthe-less would be contradictory to ourresults. Provided that Pax2/5/8 andSix genes also positively affect oticvesicle outgrowth, the same criterionwould argue against these gene fam-ilies. Interestingly Six genes are reg-ulated by a feedback loop (Zhu et al.,2002) and quite surprisingly Tle-corepressors binding to Six proteinsenhance gene expression. It will berevealing, therefore, to study in de-tail the mechanism of Groucho pro-tein function during sensory organdevelopment.
Parallel Pathways of OticVesicle and EyeDevelopment
Groucho/Tle family members havebeen identified previously to partici-pate in eye development (Kobayashiet al., 2001; Zhu et al., 2002; Lopez-Rios et al., 2003). Gain-of-functionanalysis of Tle1 resulted in ectopic ex-pression of the eye retina marker Rx2(Lopez-Rios et al., 2003). Similarly, weobserved an enlargement of the eyefield upon Tle4 overexpression in ourexperiments. Blocking Groucho/Tlefunction by overexpression of the dom-inant-negative Aes led to reduction orloss of the eye (Lopez-Rios et al.,2003). A detailed analysis of the timedependence in our experiments indi-cates that loss of the eye is a pheno-type for early Aes activation (mostprominent for mRNA injection),whereas a size reduction is seen at
similar frequencies up to the two-somite stage.
We found extensive similarities be-tween otic vesicle and eye phenotypesin our experiments. Typical embryoswith unilateral Aes misexpressionshowed size reduction of both eye andotic vesicles, similarly Tle4 overex-pression resulted in the opposite phe-notypes for both organs. Unexpectedwere the highly corresponding statis-tics for these malformations, when an-alyzed for different induction times.Similar ratios, both for complete lossas well as for size reduction were seenfor all time points tested. These datasuggest equal pathways for early de-velopment of both organs.
After initiation of eye development,a complicated network of genetic in-teractions starts. Six3 and Pax6 arekey players in this process (Hill et al.,1991; Ton et al., 1991; Quiring et al.,1994; Oliver et al., 1995; Loosli et al.,1999), and Six genes have been iden-tified to confer the Tle corepressor ef-fect (Kobayashi et al., 2001; Zhu et al.,2002; Lopez-Rios et al., 2003). Our ex-periments suggest that the basic ar-chitecture of the developmental path-ways for both sensory organs have tobe similar, strongly suggesting an im-portant function for Six family mem-bers in otic development. The exacttargets still have to be identified, butour results support the prominent roleof Groucho corepressor proteins dur-ing sensory organ formation.
Induction of Aes DuringGastrulation Leads toPartial Axis Duplication
The genetic network controlling theinduction of the anterior/posterioraxis in vertebrates is well understood(for reviews, see De Robertis et al.,2000; Schier, 2001). Gain-of-functionanalysis has identified multiple com-ponents of the Wnt-signalling path-way with the potential to initiate thisprocess (Moon and Kimelman, 1998).Downstream effectors of this pathwayare Tcf/Lef transcription factors,which interact with �-catenin in thenucleus (Behrens et al., 1996; Mo-lenaar et al., 1996; van de Wetering etal., 1997). In the absence of �-cateninactivation, Tcf/Lef transcription fac-tors are transformed into transcrip-tional repressors through interaction
768 BAJOGHLI ET AL.

with Groucho/Tle proteins (Roose etal., 1998; Brantjes et al., 2001). Thus,Grg4 inhibits axis formation, whereasAes/Grg5 enhances the axis-inducingpotential of misexpressed effectors ofwnt signalling (Roose et al., 1998).Dorsal injection of Aes/Grg5 mRNAalone into Xenopus embryos did notresult in significant phenotypes(Roose et al., 1998). We rarely ob-served a duplication of the endoge-nous axis after mRNA injection (3.2%)but at a frequency of 36% upon Aesactivation during early gastrulation.The duplications mainly affected themidbrain and the hindbrain but, insome cases, also more posterior re-gions of the spinal chord. We neversaw duplicated heads. In all cases, thebranched neural tubes fused again ata more posterior position, consistentwith a transient effect of Aes afterheat-shock induction.
Taken together, misexpression ofAes/Grg5 through application of aheat-inducible promoter has shownphenotypes in sensory organ develop-ment and organizer formation. Duringotic vesicle and eye formation, Grou-cho proteins seem to control both ini-tiation as well as size regulation. Thenext step will be to identify the tran-scription factors conferring the effectsof these corepressors during otic vesi-cle development.
EXPERIMENTALPROCEDURES
Fish Strains andMaintenance
Medaka embryos and adults of theCab inbred strain were used for allexperiments (Loosli et al., 2000).Adult fish were kept under a repro-duction regimen (14 hr light/10 hrdark) at 26°C. Embryos were collecteddaily immediately after spawning.Embryonic stages were determinedaccording to Iwamatsu (2004).
DNA, RNA Injection, andHeat-Shock Treatment
Medaka Aes (Lopez-Rios et al., 2003)and mouse Tle4 (Eberhard et al.,2000) cDNAs were cloned into theHSE expression construct (Bajoghli etal., 2004). DNA was coinjected withthe I-SceI meganuclease enzyme as
described (Thermes et al., 2002) intosingle blastomeres at the one- to two-cell stage. After injection, the embryoswere incubated at 28°C. Before heat-shock treatment, the embryos wereselected for background activity underthe fluorescence microscope. Heattreatment was performed for 2 hr at39°C as described previously (Bajoghliet al., 2004). After heat shock, the em-bryos were incubated at 28°C. In alltransient experiments, backgroundGFP expression was seen in 0–10% ofthe injected embryos, which wereeliminated from further analysis. As acontrol group, 100 embryos were in-jected with GFP:HSE:Aes at 30 ng/�l.Twenty-four hours after injection, fiveembryos exhibited background GFPexpression. The remaining embryos(n � 95) were incubated at 28°C untilhatching. All embryos were GFP-neg-ative and did not show any pheno-types (data not shown). For DNA in-jections (CMV:Aes), a pCS2 expressionconstruct containing the medaka AescDNA was used. Messenger RNA ofAes was in vitro transcribed using theSP6 Message Machine Kit (Ambion).The CMV:Aes DNA and the mRNAwere injected in 1� Yamamoto buffer.
Whole-Mount In SituHybridization
Embryos were fixed in 4% paraformal-dehyde/2PTW (2 � PBS at pH 7.5,0.1% Tween-0). The chorion of em-bryos beyond gastrula stages was re-moved. Whole-mount in situ hybrid-ization was performed at 65°C asdescribed (Quiring et al., 2004) usingDIG-labeled probes for Sox3, Eya1,Six1, and Pax2 (Koster et al., 2000).
ACKNOWLEDGMENTSWe thank Viktoriya Titova for cloningof DNA constructs, Jochen Wittbrodtand Felix Loosli for providing in situprobes (Sox3, Six1, Pax2, Eya1), Ja-viar Lopez-Rıos for medaka Aes cDNAand Dirk Eberhard for mouse Tle4cDNA. The work was supported by theAustrian Genome Project GEN-AU.
REFERENCES
Bajoghli B, Aghaallaei N, Heimbucher T,Czerny T. 2004. An artificial promoterconstruct for heat-inducible misexpres-
sion during fish embryogenesis. Dev Biol271:416–430.
Behrens J, von Kries JP, Kuhl M, Bruhn L,Wedlich D, Grosschedl R, Birchmeier W.1996. Functional interaction of beta-catenin with the transcription factorLEF-1. Nature 382:638–642.
Bessarab DA, Chong SW, Korzh V. 2004.Expression of zebrafish six1 during sen-sory organ development and myogenesis.Dev Dyn 230:781–786.
Bonini NM, Bui QT, Gray-Board GL, War-rick JM. 1997. The Drosophila eyes ab-sent gene directs ectopic eye formation ina pathway conserved between flies andvertebrates. Development 124:4819–4826.
Brantjes H, Roose J, van De Wetering M,Clevers H. 2001. All Tcf HMG box tran-scription factors interact with Groucho-related co-repressors. Nucleic Acids Res29:1410–1419.
Burton Q, Cole LK, Mulheisen M, ChangW, Wu DK. 2004. The role of Pax2 inmouse inner ear development. Dev Biol272:161–175.
Cai Y, Brophy PD, Levitan I, Stifani S,Dressler GR. 2003. Groucho suppressesPax2 transactivation by inhibition ofJNK-mediated phosphorylation. Embo J22:5522–5529.
Chen G, Courey AJ. 2000. Groucho/TLEfamily proteins and transcriptional re-pression. Gene 249:1–16.
Chen G, Fernandez J, Mische S, CoureyAJ. 1999. A functional interaction be-tween the histone deacetylase Rpd3 andthe corepressor groucho in Drosophiladevelopment. Genes Dev 13:2218–2230.
Chiang EF, Pai CI, Wyatt M, Yan YL,Postlethwait J, Chung B. 2001. Two sox9genes on duplicated zebrafish chromo-somes: expression of similar transcrip-tion activators in distinct sites. Dev Biol231:149–163.
Choi CY, Kim YH, Kwon HJ, Kim Y. 1999.The homeodomain protein NK-3 recruitsGroucho and a histone deacetylase com-plex to repress transcription. J BiolChem 274:33194–33197.
Choudhury BK, Kim J, Kung HF, Li SS.1997. Cloning and developmental ex-pression of Xenopus cDNAs encoding theEnhancer of split groucho and relatedproteins. Gene 195:41–48.
de Martino S, Yan YL, Jowett T, Postleth-wait JH, Varga ZM, Ashworth A, AustinCA. 2000. Expression of sox11 gene du-plicates in zebrafish suggests the recip-rocal loss of ancestral gene expressionpatterns in development. Dev Dyn 217:279–292.
De Robertis EM, Larrain J, OelgeschlagerM, Wessely O. 2000. The establishmentof Spemann’s organizer and patterningof the vertebrate embryo. Nat Rev Genet1:171–181.
Dutton KA, Pauliny A, Lopes SS, ElworthyS, Carney TJ, Rauch J, Geisler R, Haff-ter P, Kelsh RN. 2001. Zebrafish colour-less encodes sox10 and specifies non-ec-tomesenchymal neural crest fates.Development 128:4113–4125.
Groucho REGULATES OTIC VESICLE OUTGROWTH 769

Eberhard D, Jimenez G, Heavey B, Bus-slinger M. 2000. Transcriptional repres-sion by Pax5 (BSAP) through interactionwith corepressors of the Groucho family.EMBO J 19:2292–2303.
Fisher AL, Caudy M. 1998. Groucho pro-teins: transcriptional corepressors forspecific subsets of DNA-binding tran-scription factors in vertebrates and in-vertebrates. Genes Dev 12:1931–1940.
Groves AK, Bronner-Fraser M. 2000. Com-petence, specification and commitmentin otic placode induction. Development127:3489–3499.
Hans S, Liu D, Westerfield M. 2004. Pax8and Pax2a function synergistically inotic specification, downstream of theFoxi1 and Dlx3b transcription factors.Development 131:5091–5102.
Hartley DA, Preiss A, Artavanis-TsakonasS. 1988. A deduced gene product fromthe Drosophila neurogenic locus, en-hancer of split, shows homology to mam-malian G-protein beta subunit. Cell 55:785–795.
Heanue TA, Reshef R, Davis RJ, MardonG, Oliver G, Tomarev S, Lassar AB,Tabin CJ. 1999. Synergistic regulation ofvertebrate muscle development byDach2, Eya2, and Six1, homologs ofgenes required for Drosophila eye forma-tion. Genes Dev 13:3231–3243.
Heller N, Brandli AW. 1999. Xenopus Pax-2/5/8 orthologues: novel insights into Paxgene evolution and identification ofPax-8 as the earliest marker for otic andpronephric cell lineages. Dev Genet 24:208–219.
Hill RE, Favor J, Hogan BL, Ton CC, Saun-ders GF, Hanson IM, Prosser J, JordanT, Hastie ND, van Heyningen V. 1991.Mouse small eye results from mutationsin a paired-like homeobox-containinggene. Nature 354:522–525.
Iwamatsu T. 2004. Stages of normal devel-opment in the medaka Oryzias latipes.Mech Dev 121:605–618.
Jimenez G, Paroush Z, Ish-Horowicz D.1997. Groucho acts as a corepressor for asubset of negative regulators, includingHairyandEngrailed.GenesDev11:3072–3082.
Kobayashi M, Nishikawa K, Suzuki T,Yamamoto M. 2001. The homeobox pro-tein Six3 interacts with the Grouchocorepressor and acts as a transcriptionalrepressor in eye and forebrain formation.Dev Biol 232:315–326.
Koster RW, Kuhnlein RP, Wittbrodt J.2000. Ectopic Sox3 activity elicits sen-soryplacodeformation.MechDev95:175–187.
Ladher RK, Anakwe KU, Gurney AL,Schoenwolf GC, Francis-West PH. 2000.Identification of synergistic signals initi-ating inner ear development. Science 290:1965–1967.
Leger S, Brand M. 2002. Fgf8 and Fgf3 arerequired for zebrafish ear placode induc-tion, maintenance and inner ear pattern-ing. Mech Dev 119:91–108.
Leon C, Lobe CG. 1997. Grg3, a murineGroucho-related gene, is expressed inthe developing nervous system and in
mesenchyme-induced epithelial struc-tures. Dev Dyn 208:11–24.
Li SS. 2000. Structure and function of theGroucho gene family and encoded tran-scriptional corepressor proteins from hu-man, mouse, rat, Xenopus, Drosophilaand nematode. Proc Natl Sci Counc Re-pub China B 24:47–55.
Li X, Oghi KA, Zhang J, Krones A, BushKT, Glass CK, Nigam SK, Aggarwal AK,Maas R, Rose DW, Rosenfeld MG. 2003.Eya protein phosphatase activity regu-lates Six1-Dach-Eya transcriptional ef-fects in mammalian organogenesis. Na-ture 426:247–254.
Liu D, Chu H, Maves L, Yan YL, MorcosPA, Postlethwait JH, Westerfield M.2003. Fgf3 and Fgf8 dependent and in-dependent transcription factors are re-quired for otic placode specification. De-velopment 130:2213–2224.
Lombardo A, Isaacs HV, Slack JM. 1998.Expression and functions of FGF-3 inXenopus development. Int J Dev Biol 42:1101–1107.
Loosli F, Winkler S, Wittbrodt J. 1999.Six3 overexpression initiates the forma-tion of ectopic retina. Genes Dev 13:649–654.
Loosli F, Koster RW, Carl M, Kuhnlein R,Henrich T, Mucke M, Krone A, WittbrodtJ. 2000. A genetic screen for mutationsaffecting embryonic development inmedaka fish (Oryzias latipes). Mech Dev97:133–139.
Lopez-Rios J, Tessmar K, Loosli F, Witt-brodt J, Bovolenta P. 2003. Six3 and Six6activity is modulated by members of thegroucho family. Development 130:185–195.
Mansouri A, Chowdhury K, Gruss P. 1998.Follicular cells of the thyroid gland re-quire Pax8 gene function. Nat Genet 19:87–90.
Maroon H, Walshe J, Mahmood R, KieferP, Dickson C, Mason I. 2002. Fgf3 andFgf8 are required together for formationof the otic placode and vesicle. Develop-ment 129:2099–2108.
Miyasaka H, Choudhury BK, Hou EW, LiSS. 1993. Molecular cloning and expres-sion of mouse and human cDNA encod-ing AES and ESG proteins with strongsimilarity to Drosophila enhancer of splitgroucho protein. Eur J Biochem 216:343–352.
Molenaar M, van de Wetering M, Ooster-wegel M, Peterson-Maduro J, Godsave S,Korinek V, Roose J, Destree O, CleversH. 1996. XTcf-3 transcription factor me-diates beta-catenin-induced axis forma-tion in Xenopus embryos. Cell 86:391–399.
Molenaar M, Brian E, Roose J, Clevers H,Destree O. 2000. Differential expressionof the Groucho-related genes 4 and 5during early development of Xenopuslaevis. Mech Dev 91:311–315.
Moon RT, Kimelman D. 1998. From corti-cal rotation to organizer gene expression:toward a molecular explanation of axisspecification in Xenopus. Bioessays 20:536–545.
Oliver G, Mailhos A, Wehr R, CopelandNG, Jenkins NA, Gruss P. 1995. Six3, amurine homologue of the sine oculisgene, demarcates the most anterior bor-der of the developing neural plate and isexpressed during eye development. De-velopment 121:4045–4055.
Pfeffer PL, Gerster T, Lun K, Brand M,Busslinger M. 1998. Characterization ofthree novel members of the zebrafishPax2/5/8 family: dependency of Pax5 andPax8 expression on the Pax2.1 (noi) func-tion. Development 125:3063–3074.
Pflugrad A, Meir JY, Barnes TM, MillerDM, 3rd. 1997. The Groucho-like tran-scription factor UNC-37 functions withthe neural specificity gene unc-4 to gov-ern motor neuron identity in C. elegans.Development 124:1699–1709.
Phillips BT, Bolding K, Riley BB. 2001.Zebrafish fgf3 and fgf8 encode redundantfunctions required for otic placode induc-tion. Dev Biol 235:351–365.
Phillips BT, Storch EM, Lekven AC, RileyBB. 2004. A direct role for Fgf but notWnt in otic placode induction. Develop-ment 131:923–931.
Quiring R, Walldorf U, Kloter U, GehringWJ. 1994. Homology of the eyeless geneof Drosophila to the Small eye gene inmice and Aniridia in humans. Science265:785–789.
Quiring R, Wittbrodt B, Henrich T, Rami-alison M, Burgtorf C, Lehrach H, Witt-brodt J. 2004. Large-scale expressionscreening by automated whole-mount insitu hybridization. Mech Dev 121:971–976.
Reifers F, Bohli H, Walsh EC, Crossley PH,Stainier DY, Brand M. 1998. Fgf8 is mu-tated in zebrafish acerebellar (ace) mu-tants and is required for maintenance ofmidbrain-hindbrain boundary develop-ment and somitogenesis. Development 125:2381–2395.
Ren B, Chee KJ, Kim TH, Maniatis T.1999. PRDI-BF1/Blimp-1 repression ismediated by corepressors of the Grouchofamily of proteins. Genes Dev 13:125–137.
Riley BB, Phillips BT. 2003. Ringing in thenew ear: resolution of cell interactions inotic development. Dev Biol 261:289–312.
Roose J, Molenaar M, Peterson J, Huren-kamp J, Brantjes H, Moerer P, van deWetering M, Destree O, Clevers H. 1998.The Xenopus Wnt effector XTcf-3 inter-acts with Groucho-related transcrip-tional repressors. Nature 395:608–612.
Sahly I, Andermann P, Petit C. 1999. Thezebrafish eya1 gene and its expressionpattern during embryogenesis. DevGenes Evol 209:399–410.
Saint-Germain N, Lee YH, Zhang Y, Sar-gent TD, Saint-Jeannet JP. 2004. Speci-fication of the otic placode depends onSox9 function in Xenopus. Development131:1755–1763.
Schier AF. 2001. Axis formation and pat-terning in zebrafish. Curr Opin GenetDev 11:393–404.
Schmidt CJ, Sladek TE. 1993. A rat ho-molog of the Drosophila enhancer of split
770 BAJOGHLI ET AL.

(groucho) locus lacking WD-40 repeats.J Biol Chem 268:25681–25686.
Sharief FS, Tsoi SC, Li SS. 1997. cDNAcloning and genomic organization of en-hancer of split groucho gene from nema-tode Caenorhabditis elegans. BiochemMol Biol Int 43:327–337.
Stifani S, Blaumueller CM, Redhead NJ,Hill RE, Artavanis-Tsakonas S. 1992.Human homologs of a Drosophila En-hancer of split gene product define anovel family of nuclear proteins. NatGenet 2:119–127.
Thermes V, Grabher C, Ristoratore F,Bourrat F, Choulika A, Wittbrodt J, JolyJS. 2002. I-SceI meganuclease mediateshighly efficient transgenesis in fish.Mech Dev 118:91–98.
Ton CC, Hirvonen H, Miwa H, Weil MM,Monaghan P, Jordan T, van HeyningenV, Hastie ND, Meijers-Heijboer H,Drechsler M, et al. 1991. Positional clon-ing and characterization of a paired box-and homeobox-containing gene from theaniridia region. Cell 67:1059–1074.
Torres M, Gomez Pardo E, Gruss P. 1996.Pax2 contributes to inner ear patterningand optic nerve trajectory. Development122:3381–3391.
Valentine SA, Chen G, Shandala T, Fer-nandez J, Mische S, Saint R, Courey AJ.1998. Dorsal-mediated repression re-quires the formation of a multiproteinrepression complex at the ventral si-lencer. Mol Cell Biol 18:6584–6594.
van de Wetering M, Cavallo R, Dooijes D,van Beest M, van Es J, Loureiro J, YpmaA, Hursh D, Jones T, Bejsovec A, PeiferM, Mortin M, Clevers H. 1997. Armadillocoactivates transcription driven by theproduct of the Drosophila segment polar-ity gene dTCF. Cell 88:789–799.
Vendrell V, Carnicero E, Giraldez F,Alonso MT, Schimmang T. 2000. Induc-tion of inner ear fate by FGF3. Develop-ment 127:2011–2019.
Whitfield TT, Granato M, van Eeden FJ,Schach U, Brand M, Furutani-Seiki M,Haffter P, Hammerschmidt M, Heisen-berg CP, Jiang YJ, Kane DA, Kelsh RN,Mullins MC, Odenthal J, Nusslein-Vol-
hard C. 1996. Mutations affecting devel-opment of the zebrafish inner ear andlateral line. Development 123:241–254.
Whitfield TT, Riley BB, Chiang MY, Phil-lips B. 2002. Development of the ze-brafish inner ear. Dev Dyn 223:427–458.
Wulbeck C, Campos-Ortega JA. 1997. Twozebrafish homolugues of the Drosophilaneurogenic gene groucho and their pat-tern of transcription during early embry-ogenesis. Dev Genes Evol 207:156–166.
Xu PX, Adams J, Peters H, Brown MC,Heaney S, Maas R. 1999. Eya1-deficientmice lack ears and kidneys and showabnormal apoptosis of organ primordia.Nat Genet 23:113–117.
Zheng W, Huang L, Wei ZB, Silvius D,Tang B, Xu PX. 2003. The role of Six1 inmammalian auditory system develop-ment. Development 130:3989–4000.
Zhu CC, Dyer MA, Uchikawa M, Kondoh H,Lagutin OV, Oliver G. 2002. Six3-medi-ated auto repression and eye developmentrequires its interaction with members ofthe Groucho-related family of co-repres-sors. Development 129:2835–2849.
Groucho REGULATES OTIC VESICLE OUTGROWTH 771





























