Embed Size (px)
Citation preview

www.e l sev i e r. com/ l oca te / i n t imp
International Immunopharmacology (2008) 8, 1291–1297
PRELIMINARY REPORT
Genistein down-modulates pro-inflammatory cytokinesand reverses clinical signs of experimental autoimmuneencephalomyelitisMarcio L. De Paula a, David H. Rodrigues b, Henrique C. Teixeira a,Michele M. Barsante a, Maria A. Souza a, Ana P. Ferreira a,⁎
a Department of Parasitology, Microbiology and Immunology, Biological Sciences Institute, Federal University of Juiz de Fora,Juiz de Fora, Minas Gerais, Brazilb Department of Biochemistry and Immunology, Biological Sciences Institute, Federal University of Minas Gerais,Belo Horizonte, Minas Gerais, Brazil
Received 29 February 2008; received in revised form 10 April 2008; accepted 5 May 2008
⁎ Corresponding author. Laboratory oParasitology, Microbiology and Immunotute, Federal University of Juiz de Fora,Brazil. Tel.: +55 32 2102 3214; fax: +55
E-mail address: [email protected]
1567-5769/$ - see front matter © 200doi:10.1016/j.intimp.2008.05.002
Abstract
Multiple sclerosis (MS) is the most common non-traumatic, disabling neurological humaninflammatory demyelinating disease of the central nervous system (CNS). Experimentalautoimmune encephalomyelitis (EAE) models MS and is characterized as a CD4+ T-helper type1 (Th1) cell-mediated autoimmune disease. It is characterized by an influx of activatedleukocytes into the CNS. Genistein, occurring abundantly in soy products, has apoptotic,antioxidant, and anti-inflammatory properties. In the present report, we investigated the use ofgenistein for the treatment of the murine model of MS. After induction of EAE with myelinoligodendrocyte glycoprotein 35–55 peptide (MOG35–55), we observed that genistein treatmentameliorated significantly the clinical symptoms, modulating pro- and anti-inflammatorycytokines. Moreover, we analyzed the leukocyte rolling and adherence in the CNS by performingintravital microscopy. Genistein treatment resulted in decreased rolling and adhering ofleukocytes as compared to the untreated group. Our data suggest that genistein might be apotential therapy for MS.© 2008 Elsevier B.V. All rights reserved.
KEYWORDSMultiple sclerosis;Experimental autoimmuneencephalomyelitis;Genistein;Cytokines;Intravital microscopy
f Immunology, Department oflogy, Biological Sciences Insti-36036-900, Juiz de Fora, M.G.,2102 3214..br (A.P. Ferreira).
8 Elsevier B.V. All rights reserved.
1. Introduction
MS is a chronic inflammatory disorder of the CNS with un-known etiology affecting approximately 2.5 million peopleworldwide [1,2]. A complex predisposing genetic trait and aninciting environmental insult such as infections agents ap-pear to be important in triggering the disease, comprising as

Table 1 Clinical score assessment
Part of the body Clinical signs Score a
Tail No clinical signs 0Loss of muscle tone in tail 1Paralysis 2
Hind-limb No clinical signs 0Weakness of one animal paw 1Weakness of both animal paws 2Paralysis of one animal paw 3Paralysis of both animal paws 4
Front-limb No clinical signs 0Weakness of any animal paw 1Paralysis of any animal paw 2
Bladder Continence 0Incontinence 1
a Numerical score arbitrarily established.
1292 M.L. De Paula et al.
hallmarks inflammation, demyelination and axonal damage[1,3]. Cytokines such as TNF-α, IFN-γ, IL-17 and IL-12, and tosome extent chemokines play a pivotal role in the establish-ment and maintenance of autoimmune disorders, acting inhighly complex networks, and often exert overlapping and inpart redundant functions by different cell types. Also, pro-inflammatory cytokines are thought to play a role in thepathogenesis of MS [4,5]. Due to its similarity to MS, EAE hasbeen used as an animal model for proof of concept studies ofMS therapy.
Both clinical and experimental evidence suggests that sexhormones in males and females are significant factorsresponsible for the sexual dimorphism in the immune response.The best-studied regulatory hormone, estrogen, is protectiveagainst the induction of EAE, demonstrating several immu-nosuppressive mechanisms. Phytoestrogens are a group ofbiologically active plant substances with a chemical structuresimilar to estrogen, exerting estrogenic and anti-estrogeniceffects [6]. Previous data suggest that the consumption ofphytoestrogens leads to protective effects againstmenopausalsymptoms and a variety of disorders, including diabetes andcancer [7,8]. Isoflavones make up the most common form ofphytoestrogens and are found in a variety of plants, especiallyin soy [9]. Genistein is the major bioactive isoflavone, dem-onstrating a variety of properties suchas induction of apoptosisin cancer cells and antioxidant effects [10,11]. However, littlehas beenelucidatedwith regard to its potential in autoimmunediseases caused by Tcell activation.
IFN-β has been approved for treatment of relapsing–remitting-MS and is currently the agent that is most broadlyused as an immunomodulatory and suppressive treatment. Itreduces exacerbation by only 30% and has a modest impacton disease progression. Indeed, it is a clear step forward inMS therapy, but the frequency of subcutaneous injections ofIFN-β, the flu-like symptoms that occur at the beginning oftherapy, the modest activity required of patients, and thetreatment failures are all reasons to search for better agents[5]. In this report, we sought to determine the clinical andbiological effects of isoflavone genistein on the myelin MOG-induced EAE model.
2. Materials and methods
2.1. Animals
Female C57Bl/6 mice 8–12 weeks old were obtained from the AnimalCare Facilities of the Federal University of Juiz de Fora (UFJF) andhoused in microisolator cages in the animal facility at the Laboratoryof Immunology. All procedures were in accordance with theprinciples of the Brazilian Code for the Use of Laboratory Animals.This project was approved by the Ethics Committee on the use oflaboratory animals from UFJF.
2.2. EAE induction
Groups of 3–6 animals were immunized or not subcutaneously (s.c.)at both sides of the tail base with 100 μg of MOG 35–55 peptide(Sigma Chemical Co., Saint Louis, USA) emulsified vol/vol incomplete Freund's adjuvant (CFA) (Sigma) supplemented with400 μg of attenuated Mycobacterium tuberculosis H37 RA (Difco,Detroit, USA). Pertussis toxin, 300 ng/animal (Sigma), was injectedintraperitoneally (i.p.) on the day of immunization and again 48 hlater. Non-immunized animals were used as control group. Animals
were monitored daily and neurological impairment was quantifiedon an arbitrary clinical scale.
2.3. Clinical assessment
Mice were weighed and observed daily for clinical signs of EAE up to21 days post-immunization (dpi). The clinical status was assessedscoring certain parts of the mice body individually according toTable 1. In case of death, mice were scored 15. The final clinicalscore was obtained adding all individual scores assessed.
2.4. Treatment with genistein
Micewere immunized and then divided into two groups. One receivedno treatment while the other was given 200 mg/kg body weight ofgenistein (Indofine Chemical Co., Hillsborough, USA) in dimethylsulfoxide (DMSO) (Sigma) 4% s.c. on daily basis [12]. The treatmentwas introduced 14 dpi for 7 days. In addition to the non-immunizedcontrol group that also received no genistein treatment, we usedanother non-immunized group in which DMSO 4% was applied only.
2.5. Cytokine production in the CNS
Brain tissue extracts were acquired from control and experimentalmice that were sacrificed by ketamine and xylazine overdose i.p. at21 dpi. Brains were removed after intravital microscopy, andhemispheres were stored on ice. Thereafter, the hemispheres werehomogenized in extraction solution (100 mg of tissue per 1 ml),containing: 0.4 MNaCl, 0.05% tween 20 (Merck&Co., Inc.,WhitehouseStation, USA), 0.5% bovine serum albumin (BSA), 0.1 M phenylmethyl-sulphonyl fluoride (PMSF), 0.1 M benzethonium chloride, 10 mMethylenediaminetetracetic acid (EDTA) and 20 kIU/ml aprotinin(Sigma), using Ultra-Turrax (IKA Works, Wilmington, USA). Brainhomogenate was spun at 10,000× g for 10min at 4 °C and supernatantswere collected and stored −70 °C. The concentration of IFN-γ, TNF-α,IL-12p40 and IL-10 in the supernatants of brain extraction, at 1:10dilution in 1% BSA in phosphate buffered saline (PBS), was assayed in anELISA set-up using commercially available antibodies and theconcentrations according to the procedures supplied by the manu-facturer (BD Biosciences Pharmingen, San Diego, USA).
2.6. Splenocyte culture
Suspensions of splenocytes were prepared on day 21. Cells werecultured at a density of 2×105/well in RPMI 1640 medium sup-plemented with 5% heat-inactivated fetal bovine serum, 2 mM
1293Genistein down-modulates pro-inflammatory cytokines and reverses autoimmune encephalomyelitis
L-glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin(RPMI 5%, Sigma), in a humidified incubator, at 37 °C and at 5% CO2
atmosphere. Cultures were stimulated with MOG35–55 peptide atthe final concentration of 10 μg/ml, concanavalin A (Sigma) at10 μg/ml, or genistein at 10 μg/ml. Supernatants were collectedafter 48 h and a quantitative ELISA was performed for IFN-γ, IL-10and TNF-α according to the manufacturer's recommendation (BDBiosciences Pharmingen).
2.7. Intravital microscopy
Intravital microscopy of the mouse cerebral microvasculature wasperformed as previously described [13]. Briefly, the mice wereanesthetized i.p. with a mixture containing 150 mg/kg ketamine and10 mg/kg xylazine and the tail vein was cannulated for administra-
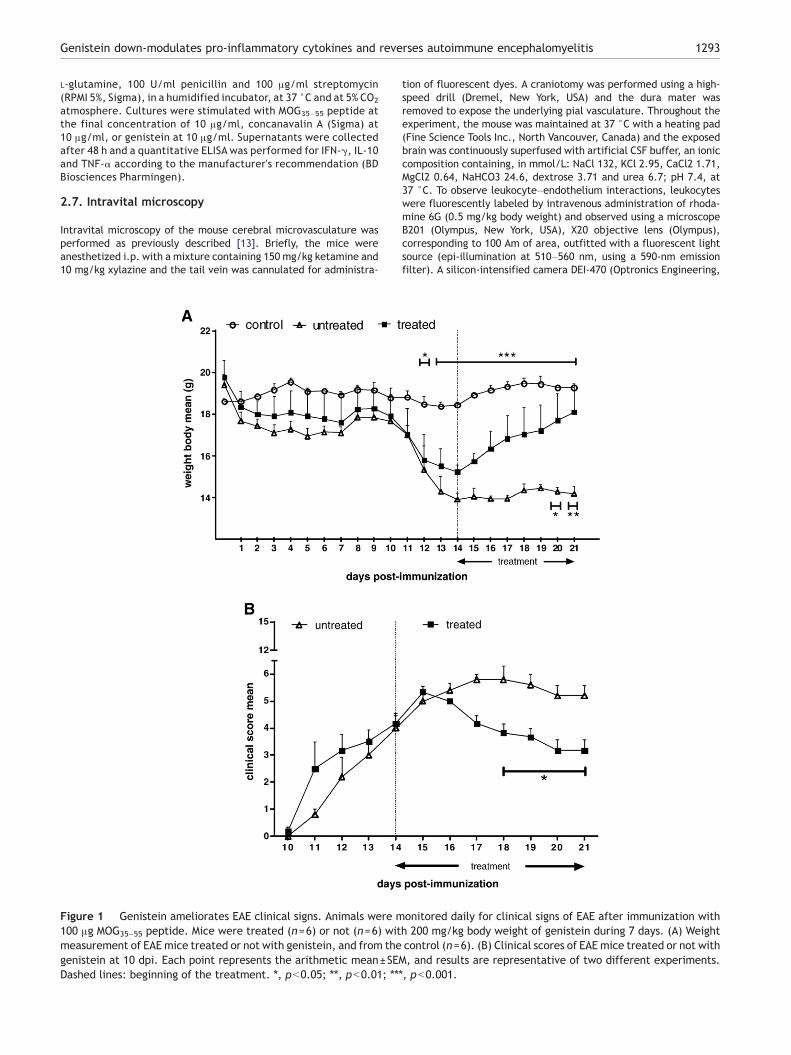
Figure 1 Genistein ameliorates EAE clinical signs. Animals were m100 μg MOG35–55 peptide. Mice were treated (n=6) or not (n=6) witmeasurement of EAE mice treated or not with genistein, and from thegenistein at 10 dpi. Each point represents the arithmetic mean±SEMDashed lines: beginning of the treatment. ⁎, pb0.05; ⁎⁎, pb0.01; ⁎⁎⁎
tion of fluorescent dyes. A craniotomy was performed using a high-speed drill (Dremel, New York, USA) and the dura mater wasremoved to expose the underlying pial vasculature. Throughout theexperiment, the mouse was maintained at 37 °C with a heating pad(Fine Science Tools Inc., North Vancouver, Canada) and the exposedbrain was continuously superfused with artificial CSF buffer, an ioniccomposition containing, in mmol/L: NaCl 132, KCl 2.95, CaCl2 1.71,MgCl2 0.64, NaHCO3 24.6, dextrose 3.71 and urea 6.7; pH 7.4, at37 °C. To observe leukocyte–endothelium interactions, leukocyteswere fluorescently labeled by intravenous administration of rhoda-mine 6G (0.5 mg/kg body weight) and observed using a microscopeB201 (Olympus, New York, USA), X20 objective lens (Olympus),corresponding to 100 Am of area, outfitted with a fluorescent lightsource (epi-illumination at 510–560 nm, using a 590-nm emissionfilter). A silicon-intensified camera DEI-470 (Optronics Engineering,
onitored daily for clinical signs of EAE after immunization withh 200 mg/kg body weight of genistein during 7 days. (A) Weightcontrol (n=6). (B) Clinical scores of EAE mice treated or not with, and results are representative of two different experiments., pb0.001.
1294 M.L. De Paula et al.
Goleta, USA) mounted on the microscope projected the image onto amonitor (Olympus). Rolling leukocytes were defined as white cellsmoving at a velocity less than that of erythrocyte cells. Leukocyteswere considered adherent to the venular endothelium if theyremained stationary for 30 s or longer.
2.8. Statistical analysis
Results presented here represent at least two independent experi-ments and are presented as the mean±SEM. For clinical score andweight measurement analysis, two-way ANOVA was performed whilethe others were assessed by one-way ANOVA. All analyses werefollowed by the Bonferroni Multiple Comparison test (GraphPadPrism 5.00), and the differences were considered significant atpb0.05.
3. Results
3.1. Treatment with genistein ameliorates the EAE clinicalcourse
The initial clinical expression of the axonal damage in an EAEmodel is represented bywell-defined signs such asweight loss, tailparalysis and hind-limb weakness [14]. In this work, we investi-gated whether genistein would be effective in the treatment ofEAE induced in C57Bl/6mice. These animals were immunizedwithMOG35–55 peptide in adjuvant, and a group was treated daily withgenistein 3 days after the beginning of the clinical signs (14 dpi),when the disease was already established. EAE severity wasrecorded daily using a weight measurement and a clinical scorescale. The immunized mice presented disease signs of EAE such as
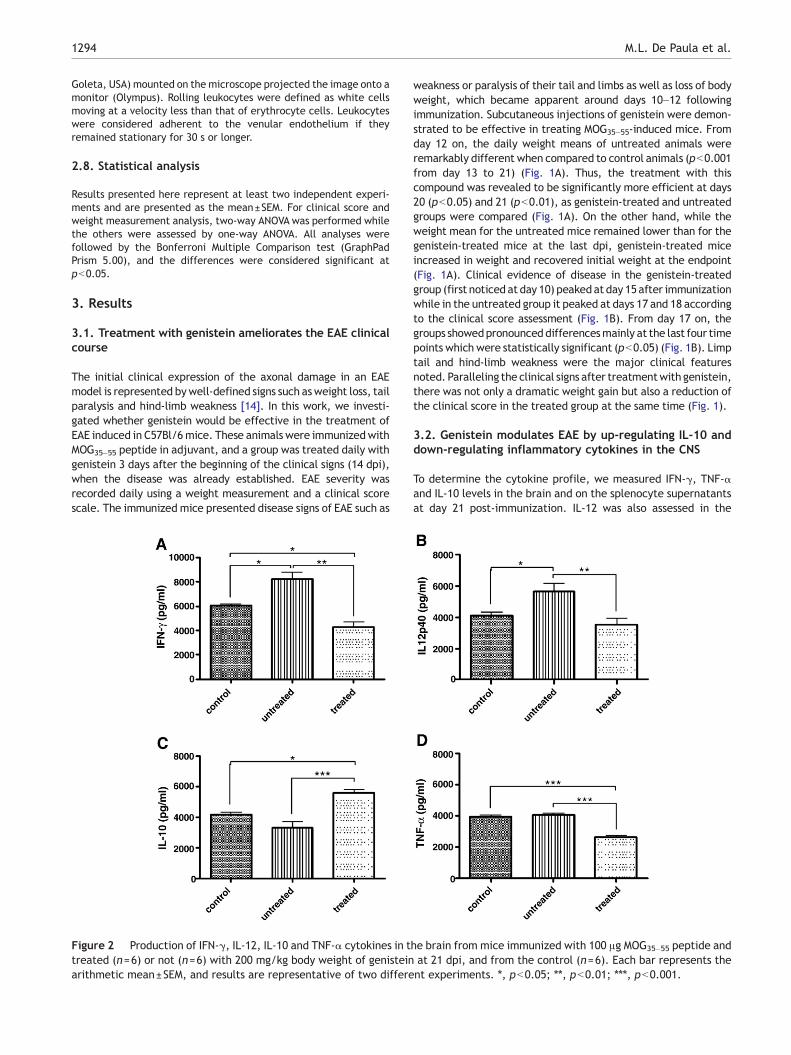
Figure 2 Production of IFN-γ, IL-12, IL-10 and TNF-α cytokines in ttreated (n=6) or not (n=6) with 200 mg/kg body weight of genisteinarithmetic mean±SEM, and results are representative of two differe
weakness or paralysis of their tail and limbs as well as loss of bodyweight, which became apparent around days 10–12 followingimmunization. Subcutaneous injections of genistein were demon-strated to be effective in treating MOG35–55-induced mice. Fromday 12 on, the daily weight means of untreated animals wereremarkably different when compared to control animals (pb0.001from day 13 to 21) (Fig. 1A). Thus, the treatment with thiscompound was revealed to be significantly more efficient at days20 (pb0.05) and 21 (pb0.01), as genistein-treated and untreatedgroups were compared (Fig. 1A). On the other hand, while theweight mean for the untreated mice remained lower than for thegenistein-treated mice at the last dpi, genistein-treated miceincreased in weight and recovered initial weight at the endpoint(Fig. 1A). Clinical evidence of disease in the genistein-treatedgroup (first noticed at day 10) peaked at day 15after immunizationwhile in the untreated group it peaked at days 17 and 18 accordingto the clinical score assessment (Fig. 1B). From day 17 on, thegroups showedpronounced differencesmainly at the last four timepoints which were statistically significant (pb0.05) (Fig. 1B). Limptail and hind-limb weakness were the major clinical featuresnoted. Paralleling the clinical signs after treatmentwith genistein,there was not only a dramatic weight gain but also a reduction ofthe clinical score in the treated group at the same time (Fig. 1).
3.2. Genistein modulates EAE by up-regulating IL-10 anddown-regulating inflammatory cytokines in the CNS
To determine the cytokine profile, we measured IFN-γ, TNF-αand IL-10 levels in the brain and on the splenocyte supernatantsat day 21 post-immunization. IL-12 was also assessed in the
he brain from mice immunized with 100 μg MOG35–55 peptide andat 21 dpi, and from the control (n=6). Each bar represents thent experiments. ⁎, pb0.05; ⁎⁎, pb0.01; ⁎⁎⁎, pb0.001.

Figure 3 Production of IL-10, IFN-γ and TNF-α cytokines byConA-driven splenocytes from mice immunized with 100 μgMOG35–55 peptide and treated (n=6) or not (n=6) with 200 mg/kg body weight of genistein at 21 dpi, and from the control mice(n=6). Each bar represents the arithmetic mean±SEM, and resultsare representative of duplicate cultures from two experiments. ⁎,pb0.05; ⁎⁎, pb0.01; ⁎⁎⁎, pb0.001.
Figure 4 Visualization of leukocyte–endothelium interactionfrom mice immunized with 100 μg MOG35–55 peptide and treated(n=4) or not (n=4) with 200 mg/kg body weight of genistein at21 dpi, and from the control (n=3). Intravital microscopy wasused to assess the rolling (A) and firm adhesion (B) of leukocytes/min on brain microvasculature. Each bar represents thearithmetic mean±SEM, and results are representative of twoindependent experiments. ⁎, pb0.05; ⁎⁎, pb0.01; ⁎⁎⁎, pb0.001.
1295Genistein down-modulates pro-inflammatory cytokines and reverses autoimmune encephalomyelitis
brain. Higher levels of IFN-γ and IL-12 were observed in theuntreated group than in the control group (pb0.05) (Fig. 2).Importantly, an impressive suppression not only of IFN-γ but alsoof IL-12 cytokine in the brain of the mice treated with genisteinin comparison to untreated mice (pb0.01) (Fig. 2). Likewise,even in relation to control mice, reduced IFN-γ levels werefound in the treated mice (pb0.05) (Fig. 2A). Conversely,elevated IL-10 levels were noted in the brain of the genistein-treated mice when compared to untreated (pb0.001) andcontrol groups (pb0.05) (Fig. 2C). The TNF-α assessment inthe brain revealed much lower levels in the genistein-treatedmice (pb0.001) compared to the untreated group (Fig. 2D).Surprisingly, no significant levels of TNF-α were observed in thebrain of the untreated mice in relation to control mice (Fig. 2D).Splenocytes were cultured and stimulated with specific and non-
specific antigens to assess cytokine levels on supernatants. Notonly IFN-γ but also IL-10 production diminished in cultures fromgenistein-treated mice in response to ConA which stimulatespolyclonal T cell response, as shown in Fig. 3A (pb0.01) and 3B(pb0.05). Also, the TNF-α production was reduced in thegenistein-treated group as compared to the untreated group,although with no statistical significance (Fig. 3C). In response toMOG35–55 peptide and genistein, no differences in regard tocytokine profile were detected in vitro (data not shown).
3.3. Leukocyte rolling and adherence are remarkablyaltered by genistein treatment
The leukocyte–endothelium interactions in the pial microcircula-tion of EAE mice treated or not with genistein were analyzed usingintravital microscopy at day 21 post-immunization. Firstly, asses-sing the number of rolling leukocytes, we found significantincreased events in the untreated group in comparison to thecontrol group (pb0.05) (Fig. 4A). In the genistein-treated mice,reduced rolling leukocytes were observed following EAE inductionas compared to untreated mice (pb0.05) (Fig. 4A). Secondly, wenoted an increasednumber of adhering leukocytes in the untreatedanimals in relation to the control animals (pb0.001) (Fig. 4B). Also,in the animals treatedwith genistein, there was a marked increaseof these events as compared to untreatedmice (pb0.001) (Fig. 4B).

1296 M.L. De Paula et al.
4. Discussion
MS is still considered a CD4+ Th1-mediated autoimmune dis-ease, which has socioeconomic importance second only totrauma in young adults [5]. Although a number of immuno-modulatory and immunosuppressive agents have beenapplied to MS treatment, better defined therapeutic strate-gies are required. Flavonoids and phytoestrogens have beentested in the EAE murine model, demonstrating that thesecompounds may have beneficial properties [15–17]. Due togenistein being the major bioactive isoflavone in soybeans,the biological properties of the phytoestrogens have beenattributed to it. Substantial data demonstrate that genisteinacts not only at the molecular level inhibiting the activity ofenzymes utilizing ATP such as tyrosine-specific proteinkinases but also at the cellular level inducing apoptosis andinhibiting cell proliferation, suppressing osteoclast and lym-phocyte functions, and exerting antioxidant effects [11,18].Herein, we investigated the effects of this compound on theEAE model by triggering the clinical disease in C57BL/6 micethrough injection of MOG. We noted that genistein hadsignificant beneficial effects on EAE decreasing severity notonly by down-regulating inflammatory cytokines but by af-fecting the trafficking of leukocytes into CNS as well.
Animal models of autoimmune diseases are providing avaluable means of analyzing the functional roles of cytokinesin the pathogenesis of autoimmunity. In our study, genisteintreatment significantly lowered pro-inflammatory cytokinelevels such as IFN-γ and IL-12 in the CNS. In addition, IFN-γproductionwas diminished on ConA-driven splenocytes. IFN-γactivates microglia to act as effector cells that damage CNScells via phagocytosis and the release of citotoxic factorsincluding glutamate, NO, superoxide, and pro-inflammatorycytokines [19]. Conversely, IFN-γ-knockout animals developnormal EAE compared to wild-type littermates [20]. DespiteIFN-γ possibly playing a relevant role in remission of MS by up-regulating pro-apoptotic proteins [21], the prevailing per-ception is that IFN-γ plays important roles in the initiationand development of MS. IL-12, a main stimulator of IFN-γ, haslong been known to drive Th1 polarization [22], and use ofanti-IL-12 can ameliorate EAE [23]. Yet in the absence of IL-12Rβ2, mice developed more severe clinical signs [24]. Atleast in part, IL-12 has been shown to act on the diseaseinitiation and in the CNS inflammatory events of MS [19].Next, we evaluated the local TNF-α levels in brain super-natants. TNF-α production is associated with Th1 responseand classically induces activation of a variety of cell types andexpression of adhesion molecules, cytokines, and chemo-kines in the CNS, which leads to CNS invasion by T cells andmonocytes. The expression of TNF-α in the CNS parallels thedisease course in EAE [4]. In TNF-α knockout mice, invasionof the CNS by immune cells is delayed, but later developsextensively. In humans, elevated serum TNF-α concentra-tions and PBMC secreting TNF-α have been reported in MSpatients [25]. According to kinetic studies in the EAE model,this cytokine peaks when clinical signs and T cell infiltrationare remarkable, and declines thereafter. Also, TNF-α can beproduced by different cell types at distinct times in thedisease process [26]. It may explain similar TNF-α amountsfound here in control and untreated groups at 21 dpi. BesidesIFN-γ and IL-12, genistein dramatically reduced TNF-α levelsafter 7 days of treatment in comparison to the untreated
group. Therefore, suppressed local TNF-α levels suggest aremarkable effect of genistein in MOG-induced EAE.
In order to rule out the possibility of genistein's vehicleinterferes with the results, we applied DMSO on non-im-munized animals. Similar results to control group were notedwhen only DMSO was used (data not shown). In the presentreport, we also analyzed IL-10 levels in the CNS and onculture splenocyte supernatants. Reduced IL-10 levels onConA-driven cultured splenocytes may be explained bydifferent responses in distinctive cells [27]. IL-10 has asprimary function to inhibit cytokine production by macro-phages such as TNF-α [28]. Genistein may diminish TNF-αlevels either directly, or indirectly by augmenting IL-10expression. Accumulated evidence has demonstrated that IL-10 is an important cytokine in the recovery from EAE. Incontrast, IL-10 was shown to play a dominant role in theregulation of EAE since IL-10−/− mice suffer a severe non-remitting disease [28]. Nevertheless, the consensus is thatIL-10 is the most potent regulator of EAE [29], and a cytokinecircuit involving IL-10 and IL-12 as counter-regulators hasbeen proposed to control disease progression [30]. In thiscontext, compounds such as genistein might become usefulin inflammatory disorders including MS.
A pathological hallmark of MS is the infiltration of immunecells across the endothelium of the blood brain barrier andtheir subsequent entry into the CNS [5]. Leukocyte extravasa-tion has been separated into discrete steps, which areassociated with interacting pairs of selectins and their ligands,integrins and cell-adhesion molecules, and particularly che-mokines and their receptors [31].To migrate into sites ofinflammation, leukocytes must first roll along the vesselbefore they firmly adhere and emigrate out of the micro-vasculature [32]. In the current work we performed intravitalmicroscopy to evaluate the leukocyte rolling and adhesion inthe brain microvasculature of EAE mice. Previous studies havedemonstrated the presence of chemokines in brain lesions andCSF of MS patients [33,34]. We noted a marked alteration notonly in leukocyte rolling but also in leukocyte adherence,suggesting that genistein acts indirectly on the interactionbetween cells and endothelium in the pialmicrovasculature bydown-regulating inflammatory cytokines as discussed above.Even though we did not evaluate the determinant moleculesinvolved in this process such as chemokines, integrins or cell-adhesion molecules, we cannot rule out that genistein mightalso act directly on them especially because phytoestrogenshave significantly higher affinities for estrogen receptors-β,which are located in high concentration in the brain [35].In addition, experimental evidence reported that genisteininhibits important enzymes responsible for up-regulatingadhesion proteins and integrins [36].
Taken together, our results shed more light on potentialalternative therapies that might be applied to MS. Limitingthe activation and migration of immune cells into the CNSappears to be an interesting approach to developing a noveltreatment for MS.
Acknowledgments
This work was supported by grants from CAPES, CNPqand FAPEMIG. Language assistance was provided by DanielStockdell.

1297Genistein down-modulates pro-inflammatory cytokines and reverses autoimmune encephalomyelitis
References
[1] Weiner HL, Selkoe DJ. Inflammation and therapeutic vaccina-tion in CNS diseases. Nature 2002;420:879–84.
[2] Steinman L, Zamvil S. Transcriptional analysis of targets inmultiple sclerosis. Nat Rev Immunol 2003;3:483–92.
[3] Dyment D, Ebers G, Sadovnick AD. Genetics of multiple sclerosis.Lancet Neurol 2004;3:104–10.
[4] Begolka WS, Miller SD. Cytokines as intrinsic and exogenousregulators of pathogenesis in experimental autoimmune ence-phalomyelitis. Res Immunol 1998;149:771–81.
[5] Sospedra M, Martin R. Immunology of multiple sclerosis. AnnuRev Immunol 2005;23:683–747.
[6] Kuiper GGJM, Lemmen JG, Carlsson B, Corton JC, Safe SH, vander Saag PT, et al. Interaction of estrogenic chemicals andphytoestrogens with estrogen receptor {beta}. Endocrinology1998;139:4252–63.
[7] Bhathena SJ, Velasquez MT. Beneficial role of dietary phytoestro-gens in obesity and diabetes. Am J Clin Nutr 2002;76: 1191–201.
[8] Lamartiniere CA, Cotroneo MS, Fritz WA, Wang J, Mentor-Marcel R, Elgavish A. Genistein chemoprevention: timing andmechanisms of action in murine mammary and prostate. J Nutr2002;132:552S–5S.
[9] Tham DM, Gardner CD, Haskell WL. Potential health benefitsof dietary phytoestrogens: a review of the clinical, epidemio-logical, and mechanistic evidence. J Clin Endocrinol Metab1998;83:2223–35.
[10] Wei H, Bowen R, Cai Q, Barnes S, Wang Y. Antioxidant andantipromotional effects of the soybean isoflavone genistein.Proc Soc Exp Biol Med 1995;208:124–30.
[11] Polkowski K, Mazurek AP. Biological properties of genistein. Areview of in vitro and in vivo data. Acta Pol Pharm 2000;57:135–55.
[12] Yellayi S, Naaz A, Szewczykowski MA, Sato T, Woods JA, ChangJ, et al. The phytoestrogen genistein induces thymic and im-mune changes: a human health concern? Proc Natl Acad Sci USA2002;99:7616–21.
[13] Carvalho-Tavares J, Hickey MJ, Hutchison J, Michaud J, SutcliffeIT, Kubes P. A role for platelets and endothelial selectins in tumornecrosis factor-{alpha}-induced leukocyte recruitment in thebrain microvasculature. Circ Res 2000;87: 1141–8.
[14] Leadbetter EA, Bourque CR, Devaux B, Olson CD, Sunshine GH,Hirani S, et al. Experimental autoimmune encephalomyelitisinduced with a combination of myelin basic protein and myelinoligodendrocyte glycoprotein is amelioratedbyadministration of asingle myelin basic protein peptide. J Immunol 1998;161:504–12.
[15] Hendriks JJA, Alblas J, van der Pol SMA, van Tol EAF, DijkstraCD, de Vries HE. Flavonoids influence monocytic GTPaseactivity and are protective in experimental allergic encepha-litis. J Exp Med 2004;200:1667–72.
[16] Muthian G, Bright JJ. Quercetin, a flavonoid phytoestrogen,ameliorates experimental allergic encephalomyelitis by block-ing IL-12 signaling through JAK-STAT pathway in T lymphocyte.J Clin Immunol 2004;24:542–52.
[17] Singh NP, Hegde V, Hofseth LJ, Nagarkatti M, Nagarkatti P.Resveratrol (trans-3, 5,4′-trihydroxystilbene) ameliorates experi-mental allergic encephalomyelitis primarily via induction ofapoptosis in T cells involving activation of aryl hydrocarbonreceptor and estrogen receptor. Mol Pharmacol 2007;72:1508–21.
[18] Gossner G, Choi M, Tan L, Fogoros S, Griffith KA, Kuenker M.Genistein-induced apoptosis and autophagocytosis in ovariancancer cells. Gynecol Oncol 2007;105:23–30.
[19] Platten M, Steinman L. Multiple sclerosis: trapped in deadlyglue. Nat Med 2005;11:252–3.
[20] Willenborg DO, Fordham SA, Staykova MA, Ramshaw IA, CowdenWB. IFN- {gamma} is critical to the control of murine autoimmune
encephalomyelitis and regulates both in the periphery and inthe target tissue: a possible role for nitric oxide. J Immunol1999;163:5278–86.
[21] Takeuchi H, Wang J, Kawanokuchi J, Mitsuma N, Mizuno T,Suzumura A. Interferon-gamma induces microglial-activation-induced cell death: a hypothetical mechanism of relapse andremission in multiple sclerosis. Neurobiol Dis 2006;22:33–9.
[22] Xiao BG, Ma CG, Xu LY, Link H, Lu CZ. IL-12/IFN-gamma/NO axisplays critical role in development of Th1-mediated experi-mental autoimmune encephalomyelitis. Mol Immunol 2008;45:1191–6.
[23] Leonard JP, Waldburger KE, Goldman SJ. Prevention of exper-imental autoimmune encephalomyelitis by antibodies againstinterleukin 12. J Exp Med 1995;181:381–6.
[24] Zhang GX, Gran B, Yu S, Li J, Siglienti I, Chen X, et al. Inductionof experimental autoimmune encephalomyelitis in IL-12 recep-tor-{beta}2-deficient mice: IL-12 responsiveness is not requiredin the pathogenesis of inflammatory demyelination in the cen-tral nervous system. J Immunol 2003;170:2153–60.
[25] Ozenci V, Kouwenhoven M, Huang YM, Kivisakk P, Link H.Multiple sclerosis is associated with an imbalance betweentumour necrosis factor-alpha (TNF-alpha)- and IL-10-secretingblood cells that is corrected by interferon-beta (IFN-beta)treatment. Clin Exp Immunol 2000;120:147–53.
[26] Juedes AE, Hjelmström P, Bergman CM, Neild AL, Ruddle NH.Kinetics and cellular origin of cytokines in the central nervoussystem: insight into mechanisms of myelin oligodendrocyteglycoprotein-induced experimental autoimmune encephalo-myelitis. J Immunol 2000;164:419–26.
[27] McGeachy MJ, Anderton SM. Cytokines in the induction andresolution of experimental autoimmune encephalomyelitis.Cytokine 2005;32:81–4.
[28] Bettelli E, Prabhu Das M, Howard ED, Weiner HL, Sobel RA,Kuchroo VK. IL-10 is critical in the regulation of autoimmuneencephalomyelitis as demonstrated by studies of IL-10- and IL-4-deficient and transgenic mice. J Immunol 1998;161:3299–306.
[29] Bettelli E, Nicholson LB, Kuchroo VK. IL-10, a key effectorregulatory cytokine in experimental autoimmune encephalo-myelitis. J Autoimmun 2003;20:265–7.
[30] Segal BM, Dwyer BK, Shevach EM. An interleukin (IL)-10/IL-12immunoregulatory circuit controls susceptibility to autoim-mune disease. J Exp Med 1998;187:537–46.
[31] Ransohoff RM, Kivisakk P, Kidd G. Three or more routes forleukocyte migration into the central nervous system. Nat RevImmunol 2003;3:569–81.
[32] Kerfoot SM, Kubes P. Overlapping roles of P-selectin and {alpha}4 integrin to recruit leukocytes to the central nervous systemin experimental autoimmune encephalomyelitis. J Immunol2002;169:1000–6.
[33] Sorensen TL, Tani M, Jensen J, Pierce V, Lucchinetti C, FolcikVA, et al. Expression of specific chemokines and chemokinereceptors in the central nervous system of multiple sclerosispatients. J Clin Invest 1999;103:807–15.
[34] Glass WG, Hickey MJ, Hardison JL, Liu MT, Manning JE, Lane TE.Antibody targeting of the CC Chemokine ligand 5 results indiminished leukocyte infiltration into the central nervoussystem and reduced neurologic disease in a viral model ofmultiple sclerosis. J Immunol 2004;172:4018–25.
[35] Kuiper GGJM, Carlsson B, Grandien K, Enmark E, Haggblad J,Nilsson S, et al. Comparison of the ligand binding specificity andtranscript tissue distribution of estrogen receptors {alpha} and{beta}. Endocrinology 1997;138:863–70.
[36] Muller DM, Pender MP, Greer JM. Chemokines and chemokinereceptors: potential therapeutic targets in multiple sclerosis.Curr Drug Targets Inflamm Allergy 2004;3:279–90.





























