Embed Size (px)
Citation preview

Ecological Entomology (2007), 32, 82–91
© 2007 The Authors82 Journal compilation © 2007 The Royal Entomological Society
Introduction
The current concept of habitat fragmentation emerged from the theory of island biogeography ( MacArthur & Wilson, 1967 ). The difficulty of extending this theory from island archipelagos to terrestrial systems of habitat patches was primarily based on the role of the surrounding matrix and edges ( Andrén, 1994; Davies et al. , 2001; Laurance et al. , 2002 ). This is because there are few cases where species habitat can be defined within the original sense of the island biogeography theory; that is, with
delimited boundaries and internally homogeneous patches sur-rounded by a single, unsuitable habitat. Specific plant – insect interactions, such as that of oak gall wasps, may constitute one of theses cases. Oak gall wasps are obligated to oviposit inside the tissues of their vegetal hosts, and the larvae develop in galls induced on specific plant tissues ( Stone & Schönrogge, 2003 ). Galls represent the extended phenotypes of gall-inducer (galler) genes ( Dawkins, 1982; Stone & Schönrogge, 2003 ), which con-stitutes a paradigmatic example of specific plant – insect interac-tions. Thus, galls represent discrete microhabitats that support relatively closed communities of specialist inhabitants ( Askew, 1961, 1971; Torossian, 1971; Ros-Farré & Pujade-Villar, 1998; Pujade-Villar, 2004 ); area and isolation of habitats can be de-fined by the distribution of the host-plant populations. Therefore,
Correspondence: Guillem Chust, AZTI, Tecnalia/Marine Research Division, Herrera kaia portualdea z/g, 20110 Pasaia (Gipuzkoa), Spain. E-mail: [email protected]
Gall wasps and their parasitoids in cork oak fragmented forests
G U I L L E M C H U S T 1 , L U C A S G A R B I N 2 and J U L I P U J A D E - V I L L A R 2 1 Laboratoire Evolution et Diversité Biologique, CNRS/UPS, Bâtiment 4R3, 118 route de Narbonne, 31062 Toulouse, France and
2 Universitat de Barcelona, Facultat de Biologia, Departament de Biologia Animal, Avda Diagonal 645, 08028 Barcelona, Spain
Abstract . 1. This paper explores the potential effects of host-plant fragmentation on cork oak gall wasp populations (Cynipidae, Hymenoptera) and on their predators, lethal inquilines, and parasitoids. To address this objective, galls were collected across a gradient of cork oak ( Quercus suber ) forest fragmentation in the East Pyrenees (Albera, Spain), and they were incubated to obtain the parasitism rates.
2. Two hypotheses were tested: (1) Host-plant fragmentation may induce a decline in gall wasp populations because of area and isolation effects on local extinction and dispersal; as a consequence of that, parasitoids may decline even more strongly in fragmented habitats than their prey. (2) Host-plant fragmentation may cause a decline in gall wasp parasitoid populations that, in turn, can lead to an ecological release in their prey populations.
3. Among the eight cork oak gall wasps sampled in the study area of Albera, the gall abundances of three species ( Callirhytis glandium , Callirhytis rufescens , and Andricus hispanicus ) were significantly related to forest fragmentation. The overall abundance of gall wasps was affected by a radius of ≈ 890 m surrounding landscape, presenting constant abundances with forest loss until forest cover is reduced at ≈ 40%; below that value the abundance increased rapidly. Three inquilines and 23 parasitoids species were recorded after gall incubation. In 25 cases, species of inquilines and parasitoids were newly recorded for the corresponding host in the Iberian peninsula.
4. Although the overall parasitism rate was high (1.1), it was uncorrelated with fragmentation and with overall cynipid abundance. These results indicate that host-plant fragmentation was correlated with higher abundance of gall wasps, whereas the parasitism rate could not explain this hyper-abundance in small forest fragments.
Key words . Cork oak , fragmentation , gall wasp , parasitism rate , parasitoid , Pyrenees , Quercus suber .

Gall wasps in cork oak fragmented forests 83
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 82–91
gall-inducing insects may be an appropriate biological model to study the effects of habitat fragmentation on populations.
In a single-species system, populations are expected to de-cline with decreasing habitat patch size and patch isolation ( Hanski, 1994 ). However, gall-inducing insects suffer frequent attacks from numerous parasitoids and inquilines ( Stone et al. , 2002 ). In this tritrophic system, parasitoids may greatly influ-ence gall-maker populations. In prey – predator systems, preda-tors and large-bodied species are predicted to decline or disappear from isolated habitat fragments, due to relatively greater energy and area requirements and smaller population sizes ( Holt, 1996 ). This hypothesis has been partially supported in experimental tropical forests ( Laurance et al. , 2002 ). Further, the loss of higher trophic levels can lead to ecological release in prey populations ( Kareiva, 1987 ). On the other hand, the responses of parasite – host interaction to habitat loss documented in the literature seem contradictory. For instance, parasitoid populations were reduced more than their phytophagous hosts on isolated plant patches of red clover ( Kruess, 1996 ) and Vicia sepium ( Kruess & Tscharntke, 2000 ). These findings agree with Holt’s (1996) predictions that food chain length should increase with area and decrease with isolation. In contrast, Doak (2000) and Cronin (2003) found, in different tritrophic systems, that the smallest or most isolated patches of host plant had the highest overall proportion of prey hosts parasitised. The cause of this distribution of parasitism is not well understood ( Cronin, 2003 ).
This paper explores the potential effects of the host-plant fragmentation on cork oak gall wasp populations (Cynipidae, Hymenoptera) and their predators (lethal inquilines and parasi-toids). The community of gall wasps inducing galls on the cork oak forests ( Q. suber ) in the East Pyrenees was studies, as well as the parasitoids and inquilines of these species. Galls were collected at 34 sites across a gradient of cork oak forest frag-mentation: from small patches embedded in an agricultural land to large forests. Two hypotheses were contrasted: (1) host-plant fragmentation may induce a decline in gall wasp populations because of area and isolation effects on local extinction and dis-persal ( Hanski, 1994 ); as a consequence of this, parasitoids may decline more strongly in fragmented habitats than their prey ( Holt, 1996 ). (2) Host-plant fragmentation may cause a decline
in gall wasp parasitoid populations that, in turn, can lead to an ecological release in their prey populations ( Kareiva, 1987 ).
Methods
Study area
The study area, at the southern foot of Albera massif (eastern Pyrenees, Spain), is 15 km in length from south to north, and 20 km from west to east ( Fig. 1). The vegetation is mostly evergreen due to the Mediterranean climate, with dry summers, mild winters, and maximum rainfall in early autumn. The study area was defined by the regional distribution of cork oak ( Quercus suber ) forests, which mainly grow in siliceous soils resulting from the alteration of granites. In the lowland plains (50 – 200 m a.s.l.), small patches of these forests are embedded in a matrix of agricultural land and pastures. Large forests, in the north-west of the study area, are generally maintained in hilly areas at higher altitudes (150 – 450 m); hence, they are more rainy than lowland plains. Cork oaks generally form open forests with a diverse shrub community and the scattered presence of other Quercus species ( Q. ilex , Q. coccifera , Q. pubescens ), fires and human extraction of cork favour the forest openness in this area.
Biology and sampling of gall wasps
Gall wasps (Cynipidae, Hymenoptera) induce a wide variety of gall types on twigs, roots, buds, leaves, inflorescences, and fruits. They exhibit complex life cycles, with cyclical sexual and parthenogenetic (agamic) generations, and each generation forms galls with unique morphology on specific host-plant modules ( Stone & Schönrogge, 2003 ). Galls of cork oak gall wasps were collected at different locations of the study area during late spring (from 24 May to 2 June 2004). In late spring, cork oaks develop new leaves and inflorescences, therefore con-stituting an optimal period to collect a wide variety of gall types ( Nieves-Aldrey, 1987 ). Gall types corresponding to eight gall
Fig. 1. Study area of Albera (East Pyrenees, Spain). Land covers classifi ed with SPOT image: dark grey, forest; white, herbaceous and agricultural land and other land covers. Squares indicate sites of gall wasp sampling.

84 Guillem Chust, Lucas Garbin & Juli Pujade-Villar
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 82–91
wasps species have been collected as follows. For galls induced in cork oak acorns: Callirhytis glandium (agamic form) and Callirhytis rufescens (agamic form), the number of acorns (ripe and fell) that at least one gall was induced inside the fruit was counted. For galls induced in leaves, twigs, buds, and inflorescences – Andricus burgundus (sexual form), Andricus grossulariae (= mayri ; sexual form), Andricus hispanicus (= niger ; sexual form), Neuroterus saliens (sexual form), and Plagiotrochus amenti (= suberi ; sexual form) – the number of galls was counted directly in the field by standing under the trees and collecting galls with a maximum reach of 2.5 m. Finally, Synophrus politus galls were also collected. This species is a very peculiar Cynipid because recent observations show that S. politus is an inquiline that attacks, at a very early stage, the sexual generation galls induced by the cynipid A. burgundus , according to Pujade-Villar et al. (2003) . The larva of S. politus controls the differentiation of the whole gall; the original tissues and structure of the host gall are rapidly reabsorbed and masked, and from this it results in the formation of a new gall very different to the inducer cynipid gall. The galls of A. burgundus are coalescent in buds; only one or two induced galls are attacked by S. politus , but all coalescent galls are destroyed and eliminated in a very short time with S. politus present. Synophrus politus was analysed as a galler (inducing galls in leaves, buds, and inflorescences). The gall inducers studied have complex cyclically parthenogenetic – heterogonic life cycles, and all of them, except N. saliens , are heteroecic (see the Appendix ). These heteroecic species need the presence of Q. suber and the Quercus oaks to close the cycle while the heterogonics close the cycle in the same species of host. Quercus pubescens stands are scattered over all area defined by Q. suber forests, in both large forests and agricultural areas.
Because the frequency of gallers in some study sites is related to the age of their hosts ( Price et al. , 1987; Cuevas-Reyes et al. , 2004 ), the sampling was restricted to trees that had flowered. Stands whose cork was not recently extracted were also selected to avoid potential sources of variation on gall abundance. Old galls were also taken into account. Galls collected in leaves, twigs, buds, and inflorescences were incubated for 1 year in the laboratory to obtain parasitoids and inquilines, and these data were used to estimate the rate of attack on galls.
Galls were collected at 34 sites across a gradient of cork oak forest fragmentation, from small patches, ≈ 0.2 ha, embedded in an agricultural land to continuous, large forests ( Fig. 1 ). For each site, a 40 × 40 m area of low density of trees (10 – 20 flowered stands) was defined. The selection of low-density forests avoids differences in gall densities because of illumi-nation degree of leaves (G. Chust, pers . obs.). Galls were sam-pled for 100 min at each site in 32 sites, 60 min for one site and 180 min for another site. The time unit of 100 min was defined according to a preliminary study of species-accumulation curves ( Fig. 2), in which galls were sampled in 18 adjacent tree stands for 10 min per tree. To estimate population densi-ties of C. glandium and C. rufescens , between 56 and 222 acorns were collected at each site. Five of the sites presented very low acorn density; less than 100 acorns per site were sampled in these sites.
Gall abundance was estimated for each cynipid species at each site as the number of counted galls divided by their corre-sponding unit of sampling effort (minutes or acorns), and stand-ardised by 100 units (i.e. the number of galls per 100 min or per 100 acorns). An overall measure of gall abundance of all cynipid species was obtained from the addition of the relative abundance of all species, where species relative abundance is the number of galls of a species at a site divided by the total number of galls of that species in all sites. Because cork oak gall wasps die after inquiline’s attack, overall parasitism rate was estimated as the rate of attack to galls by parasitoids and inquilines. Parasitism rate at each site was then estimated by the number of parasitoids and inquilines emerging after incubation divided by the number of gall wasps that emerged.
Fragmentation of cork oak forests
Optical satellite imagery was used to map cork oak forests of the area and to describe their fragmentation. An image was selected from the SPOT HRVIR satellite sensor (dated 30 June 1998) that covered the Albera massif. The image was georefer-enced by Spot Image to project the image in UTM map projec-tion. This geometric correction uses ground control points and a digital elevation model (DEM) to correct residual parallax errors due to relief. SPOT HRVIR has a spatial resolution of 20 × 20 m and records the image with four spectral bands (green, red, near-infrared and mid-infrared wavelengths).
Land cover data were generated through a supervised classi-fication of the SPOT image into three classes: (1) forest, (2) herbaceous and agricultural land (i.e. arable fields), and (3) other land covers (water surfaces, irrigated land, coniferous plantations). The land cover forest is largely dominated by cork oak forest surrounding sampling sites, whereas it includes holm
Replicates (trees)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Num
ber
of s
peci
es
0
1
2
3
4
5
Fig. 2. Species cumulative curve for galls sampled in leaves, twigs, buds, and infl orescences in 18 adjacent tree stands during 10 min for each tree. The cumulative curve indicated that beyond 6 – 10 trees, all cynipid species are present in the site. This indicates that beyond 60 – 100 min of sampling a set of different trees, all gall wasps species were present in the site. Andricus grossulariae was not present in this site.

Gall wasps in cork oak fragmented forests 85
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 82–91
oak ( Q. ilex ) and deciduous forests in the boundaries of the study area. Land cover was classified with a maximum likeli-hood procedure using the four spectral bands. The errors of omission and commission ( Stehman, 1997 ) were respectively, forest: 3.5% and 5.3%; agricultural and herbaceous land: 16.3% and 11.4%; other land covers: 0.1% and 0%.
To characterise habitat fragmentation, habitat loss was meas-ured as the fraction of forest cover in a landscape area. Given the variability of fragmentation descriptors resulting from the scale of observation in heterogeneous landscapes ( Turner et al. , 1989 ; see also Fig. 3), a multiscale approach ( Pearman, 2002; Chust et al. , 2003 ) was applied to characterise the fragmentation of cork oak forests at different spatial scales. The proportion of forest surrounding each sampling site was calculated at different scales (from 100 × 100 m to 2 × 2 km window size) ( Fig. 3 ).
Modelling gall wasp response to fragmentation
Two analyses were performed to test which of the proposed hypotheses is supported by cork oak gall wasps: (1) the response of gall abundance, at species level and for overall cynipid spe-cies, to the proportion of forest cover at different spatial scales; and (2) the response of parasitism rate to the proportion of forest at different scales and to overall cynipid species. To address these objectives, generalised additive models (GAMs) ( Hastie & Tibshirani, 1990 ) were used, which enables the fit of non-linear models. The non-linear relationships can also be approached by generalised linear models (GLMs) by introducing appropriate polynomial terms of the predictors to improve the fit of a linear
model; however, this task can be tedious and imprecise ( Guisan et al. , 2002 ). GAMs generalise GLMs ( Hastie & Tibshirani, 1990 ) and automatically identify appropriate transformations. In linear regression, the dependent variable Y is modelled by a set of predictors variables X i as
Y Xi i
i
p
= +∑=
a b e+1
(1)
where error � ~ N (0.1). In GAMs, the linear function is replaced by a non-linear function to give
Y f Xi i
i
p
(= +∑=
a e+ )1
(2)
where f i are unspecified smooth functions, and � can follow, apart from the Gaussian, other distributions such as binomial, Poisson and gamma. Smooth terms represented by penalised regression splines with smoothing parameters selected by gen-eralised cross-validation were used. One of the risks of applying these models with few data is to overfit the model by using many parameters; in general, bias decreases and variance (un-certainty) increases as the number of parameters in a model in-creases ( Burnham & Anderson, 2002 ). Given the relatively low number of cases for the present data (i.e. 34), overfitting was prevented by restricting the number of smooth terms to four. The model is fitted by maximum-likelihood estimation tech-nique through deviance reduction, where the explained devi-ance, 1 – (residual_deviance)/(null_deviance), is the equivalent to R 2 in least squares models ( Guisan & Zimmermann, 2000 ). The construction of the model for our data was carried out with mgcv package implemented in the R language ( Wood, 2000 ).
The response of gall abundance and parasitism rate to the proportion of forest cover was thus analysed with GAM at dif-ferent spatial scales. The scale at which the GAM accounts for the most biological (gall abundance or parasitism rate) deviance is called the optimal scale , which represents the extent of land-scape influence on local galler or parasitism incidences.
Modelling climate effects on gall wasps
Because small patches of forests in Albera are mainly located in lowland plains (whereas large forests are located at higher al-titudes, and hence more rainy), habitat fragmentation can covari-ate with xeric (dry) conditions. Xeric conditions are known to be associated with galling occurrence ( Price et al. , 1998 ). An expla-nation for this association is the proposal that endophyte fungi may cause significant gall insect mortality and that occurrence of such endophytes is reduced in xeric habitats ( Fernandes & Price, 1992 ). Humidity is also an important factor in caterpil-lar development in Callirhytis ( Nieves-Aldrey, 2001; Pujade-Villar, 2002 ). To discriminate between the effects of fragmentation and climatic conditions on gall abundance, a GAM was established between overall cynipid abundance and climatic conditions (i.e. mean annual precipitation and potential solar radiation) and habitat fragmentation (at the optimal scale identified at the previous section). Mean annual precipitation (mm year −1 ) and potential solar radiation (kJ m −2 day −1 ) were acquired from the digital climatic atlas of Catalonia at 180 m grid spacing ( Pons, 1996; Ninyerola et al. , 2000 ), available
Fig. 3. Effect of spatial scale to characterise forest fragmentation. Land cover in dark tone represents forest, and light grey represents herbaceous and agricultural land. The central circle represents the biological site where gall abundance were sampled. The fraction of forest cover is calculated over the squares with discontinuous border (two square window sizes are shown: 220 × 220 m and 1.1 × 1.1 km). The smallest scale presents a 100% of forest cover, whereas the largest scale contains less than 50%.

86 Guillem Chust, Lucas Garbin & Juli Pujade-Villar
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 82–91
online at http://magno.uab.es/atles-climatic/catala/cartografia.htm . Solar radiation was computed by taking into account the sun path through the day, the sun – earth distance, the atmospheric attenuation, incident angles on each point, and cast-shadow effects on each point ( Pons, 1996 ).
Results
Descriptive analysis
A total of 3876 acorns were sampled, and 4 .6% of them were found to have galls induced by the two species of Callirhytis . The two species of Callirhytis did not present significant differ-ences in attack rate between sites with fewer acorns (less than 100) and areas with more acorns (anova, P = 0.507 for C. glandium , P = 0.333 for C. rufescens ). Average values standardised per 100 acorns were: 0.99 galls of C. glandium in sites with few acorns and 2.63 galls in sites with more acorns; 0.21 galls of C. rufescens in sites with few acorns and 2.11 galls in sites with more acorns. A total of 9733 galls induced in leaves, twigs, buds, and inflorescences were counted. Among them, N. saliens (sexual generation) and P. amenti (agamic generation) were the most abundant species, and A. grossulariae (sexual generation) the less frequent species. Galls of cynipid species studied occurred in more than a half of the sites, except A. grossulariae galls, which occurred only at seven sites (20.6%) ( Table 1). For this reason, A. grossulariae was excluded from the analysis at species level. The majority of recorded galls were either old (particularly of P. amenti and S. politus ) or recently eclosed; thus, only 2159 fresh galls were collected for incubation. Three inquilines and 23 parasitoids species were recorded after gall incubation ( Table 2). In 25 cases, species of inquilines and para-sitoids were newly recorded for the corresponding host in the Iberian peninsula. Neuroterus saliens had been attacked by a large number of these parasitoids (17 species), while A. hispani-cus only by two species. However, these differences depend on the number of parasitoid individuals that emerged after incuba-tion. The number of gall wasp and parasitoid individuals that emerged from incubations was high on galls of N. saliens (549
individuals of N. saliens and 499 parasitoid individuals) and low in other species ( Table 1 ); no emerged gall wasps were obtained from S. politus because these galls were very immature. For these reasons, only those sites where total emerged individuals were greater than 15 were considered for analysis of parasitism rate; that is in 23 sites (preserving the fragmentation gradient). The attack rate per species considering all sites was greatest in A. hispanicus (11.8 parasitoids per gall wasp emerged), followed by P. amenti and A. burgundus (more than two). Andricus grossulariae and N. saliens presented attack rates close to 1. The overall attack rate indicates that, on average, either one gall wasp or one parasitoid emerged from each gall.
Response of gall wasps to fragmentation and climatic conditions
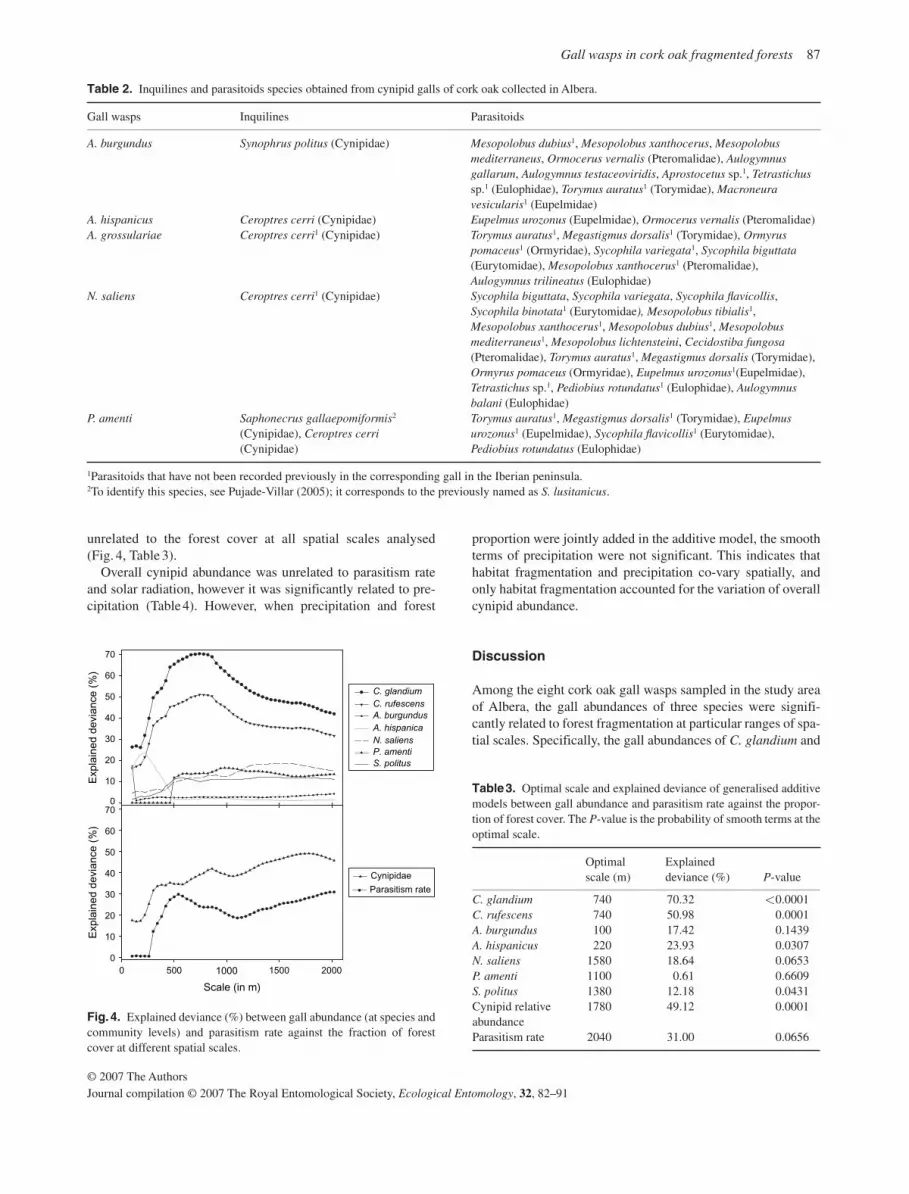
An additive model was fitted for each species and each spatial scale to relate the gall abundance and the fraction of forest cover. Among the seven gall wasps analysed, the gall abundance of C. glandium , C. rufescens , and A. hispanicus were signifi-cantly related to forest fragmentation at particular ranges of spatial scales, while the other four species were unrelated at all analysed scales ( Fig. 4). For these three species, the optimal scale could be identified and was the same for C. glandium and C. rufescens (740 × 740 m), but differed for A. hispanicus (220 × 220 m) ( Table 3). At these scales, the gall abundance of C. glandium and C. rufescens decreased with the proportion of forest cover, while A. hispanicus showed minimum abundances at intermediate values of forest proportion ( Fig. 5a – c).
The index of overall cynipid abundance was significantly related to forest fragmentation, where the best scale was identi-fied at 1.8 × 1.8 km (explained deviance: 49%, P = 0.0001) ( Fig. 4, Table 3 ). At this spatial scale, the relative cynipid abun-dance remained constant with forest loss until forest cover is reduced to ≈ 40% ( Fig. 5 ); below this, the relative abundance increased rapidly. Therefore, below a critical threshold of for-est cover, cynipids are more abundant in small or scattered patches of cork oak forests than in continuous forests. The rate of attack to cynipid galls, by parasitoids and inquilines, was
Table 1. Abundance of cork oak cynipid galls and parasitism rate sampled in 34 sites in Albera.
Mean abundance (per site)
Total abundance (all sites)
Occurrence at sites (%)
Emerged gall wasps
Emerged parasitoids and inquilines
Parasitism rate (all sites)
C. glandium 1 2.39 81.2 61.8 – – – C. rufescens 1 1.83 62.4 58.8 – – – A. burgundus 2 4.48 152.3 88.2 35 88 2.514 A. grossulariae 2 1.56 53.0 20.6 32 34 1.063 A. hispanicus 2 14.19 482.3 97.1 5 59 11.800 N. saliens 2 29.99 1019.7 88.2 549 499 0.909 P. amenti 2 27.01 918.4 100.0 7 20 2.857 S. politus 2 3.96 134.6 55.9 0 0 – Total 71.33 628 700 1.115
1 Abundance per 100 acorns. 2 Abundance per 100 min of sampling.
Gall wasps in cork oak fragmented forests 87
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 82–91
unrelated to the forest cover at all spatial scales analysed ( Fig. 4, Table 3 ).
Overall cynipid abundance was unrelated to parasitism rate and solar radiation, however it was significantly related to pre-cipitation ( Table 4). However, when precipitation and forest
proportion were jointly added in the additive model, the smooth terms of precipitation were not significant. This indicates that habitat fragmentation and precipitation co-vary spatially, and only habitat fragmentation accounted for the variation of overall cynipid abundance.
Discussion
Among the eight cork oak gall wasps sampled in the study area of Albera, the gall abundances of three species were signifi-cantly related to forest fragmentation at particular ranges of spa-tial scales. Specifically, the gall abundances of C. glandium and
Scale (in m)
0 500 1000 1500 2000
Exp
lain
ed d
evia
nce
(%)
Exp
lain
ed d
evia
nce
(%)
Cynipidae
Parasitism rate
0
10
20
30
40
50
60
70
0
10
20
30
40
50
60
70
C. glandium
C. rufescens
A. burgundus
A. hispanica
N. saliens
P. amenti
S. politus
Fig. 4. Explained deviance (%) between gall abundance (at species and community levels) and parasitism rate against the fraction of forest cover at different spatial scales.
Table 3. Optimal scale and explained deviance of generalised additive models between gall abundance and parasitism rate against the propor-tion of forest cover. The P -value is the probability of smooth terms at the optimal scale.
Optimal scale (m)
Explained deviance (%) P -value
C. glandium 740 70.32 <0.0001 C. rufescens 740 50.98 0.0001 A. burgundus 100 17.42 0.1439 A. hispanicus 220 23.93 0.0307 N. saliens 1580 18.64 0.0653 P. amenti 1100 0.61 0.6609 S. politus 1380 12.18 0.0431 Cynipid relative abundance
1780 49.12 0.0001
Parasitism rate 2040 31.00 0.0656
Table 2. Inquilines and parasitoids species obtained from cynipid galls of cork oak collected in Albera.
Gall wasps Inquilines Parasitoids
A. burgundus Synophrus politus (Cynipidae) Mesopolobus dubius 1 , Mesopolobus xanthocerus , Mesopolobus mediterraneus , Ormocerus vernalis (Pteromalidae), Aulogymnus gallarum , Aulogymnus testaceoviridis , Aprostocetus sp. 1 , Tetrastichus sp. 1 (Eulophidae), Torymus auratus 1 (Torymidae), Macroneura vesicularis 1 (Eupelmidae)
A. hispanicus Ceroptres cerri (Cynipidae) Eupelmus urozonus (Eupelmidae), Ormocerus vernalis (Pteromalidae) A. grossulariae Ceroptres cerri 1 (Cynipidae) Torymus auratus 1 , Megastigmus dorsalis 1 (Torymidae), Ormyrus
pomaceus 1 (Ormyridae), Sycophila variegata 1 , Sycophila biguttata (Eurytomidae), Mesopolobus xanthocerus 1 (Pteromalidae), Aulogymnus trilineatus (Eulophidae)
N. saliens Ceroptres cerri 1 (Cynipidae) Sycophila biguttata , Sycophila variegata , Sycophila fl avicollis , Sycophila binotata 1 (Eurytomidae ), Mesopolobus tibialis 1 , Mesopolobus xanthocerus 1 , Mesopolobus dubius 1 , Mesopolobus mediterraneus 1 , Mesopolobus lichtensteini , Cecidostiba fungosa (Pteromalidae), Torymus auratus 1 , Megastigmus dorsalis (Torymidae), Ormyrus pomaceus (Ormyridae), Eupelmus urozonus 1 (Eupelmidae), Tetrastichus sp. 1 , Pediobius rotundatus 1 (Eulophidae), Aulogymnus balani (Eulophidae)
P. amenti Saphonecrus gallaepomiformis 2 (Cynipidae), Ceroptres cerri (Cynipidae)
Torymus auratus 1 , Megastigmus dorsalis 1 (Torymidae), Eupelmus urozonus 1 (Eupelmidae), Sycophila fl avicollis 1 (Eurytomidae), Pediobius rotundatus (Eulophidae)
1 Parasitoids that have not been recorded previously in the corresponding gall in the Iberian peninsula. 2 To identify this species, see Pujade-Villar (2005) ; it corresponds to the previously named as S. lusitanicus .

88 Guillem Chust, Lucas Garbin & Juli Pujade-Villar
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 82–91
C. rufescens decreased with the proportion of forest cover, while A. hispanicus showed minimum abundances at intermediate val-ues of proportion forest cover. The overall index of cynipid abundance was significantly related to the forest fragmentation, where the best scale was identified at 1.8 × 1.8 km. This scale represents the extent of landscape influence on local galler abun-dances. The local abundance of gall wasps is thus affected by a radius of ≈ 890 m surrounding landscape, presenting constant abundances with forest loss until forest cover is reduced at ≈ 40%; below that value the abundance increased rapidly. Therefore, below a critical threshold of forest cover, cynipids are increasingly more abundant in small and isolated patches of cork oak forests than in continuous forests. These results do not agree with the first hypothesis based on Holt’s (1996) predic-tions, in which host-plant fragmentation would induce a decline in gall wasp populations and, particularly, on parasitoids. The results do not support the second hypothesis: host-plant frag-mentation causes a higher density of gall wasp abundance be-cause of a decline in gall wasp parasitoid populations, following Kareiva (1987) . The parasitism rate in Albera was uncorrelated with fragmentation and with overall cynipid abundance; hence,
the hyper-abundance of gall wasps in small forest fragments can not be explained. As noted, the parasitism rate could only be estimated at 23 sites. Therefore, a supplementary effort of sam-pling should be carried out in the future to obtain better estimates of parasitism and, hence, to establish the validation or rejection of that hypothesis.
Other potential reasons for the positive effects of host frag-mentation on cynipid abundance were explored because of the lack of correlation between gall wasp abundance and parasitism rate. First, as mentioned, most cork oak gall wasp species need the presence of deciduous oaks to close the cycle, and Q. pubes-cens stands are scattered over all the studied area. It is possible that this oak species is the limiting factor of gall wasp popula-tions in this area. If so, in agricultural-dominated landscapes, these galler populations would be concentrated in the few ref-uges that are the small patches of cork oaks; this would explain the high density of galler populations. A sampling scheme in future work, based on partitioning the fragmentation of both hosts, would permit this possibility to be tested. Second, some evidence supports the possibility that forest management or the distance to urban areas may influence the overall abundance of
Table 4. Explained deviance of generalised additive models between cynipid relative abundance against parasitism rate, xeric conditions (precipita-tion, solar radiation), and precipitation plus forest proportion (scale: 1780 × 1780 m).
Parasitism rate Precipitation Solar radiation Precipitation+Forest proportion
Explained deviance (%) 1.27 24.60 0.34 52.60 P -value 0.6004 0.0098 0.7439 0.1414 (Precipitation)
0.0011 (Forest proportion)
Fig. 5. Smooth functions (continuous line) of generalised additive models fi tting gall abun-dance ( ○ ) with respect to the proportion of forest cover at the corresponding optimal scales. Abun-dance values and smooth functions are centred on zero. Upper and lower discontinuous lines represent confi dence bands for smooth function. (a) Callirhytis glandium (scale: 740 × 740 m), (b) Callirhytis rufescens (scale: 740 × 740 m), (c) Andricus hispanicus (scale: 220 × 220 m), (d) overall cynipids (scale: 1780 × 1780 m).

Gall wasps in cork oak fragmented forests 89
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 82–91
gall wasps on Quercus robur ( Skrzypczynska, 2001 ). Cork oaks in forest patches surrounded by an agricultural land are sub-jected to different conditions than stands in larger forests. Although galls may represent discrete microhabitats, the quality of the matrix may influence the interactions with species other than parasitoids (e.g. cork oak endophytic community, Gange et al. , 2002 , abundance of galler predators). Galler predators, such as birds and rodents ( Schönrogge et al. , 1999; Pujade-Villar, 2004 ), are not reported on these cork oak gall wasps. Third, edge effects should also be taken into account when small patches are sam-pled because part of sampling includes the effects of edges. However, cork oak forms open forests in Albera and the sam-pling was designed in particularly low density forests; this should prevent edge effects because biophysical properties (at least for illumination) in the forest interior are similar to that at the edges (G. Chust, pers . obs.). Fourth, the fruit production of cork oaks can present variability among sites; thus, in areas with few acorns, the same number of gallers may lead to high number of galls per acorn. The analysis performed did not support this possibility; the abundance of the two species of Callirhytis per acorn did not present significant differences between sites with few acorns and areas with more acorns.
The rate of gall wasps against parasitoids and inquilines emerged was high ( ≈ 1:1); this reinforces the idea that parasi-toids and inquilines could influence gall wasp populations. Several studies reported top-down effects in regulating gall wasp populations ( Stone et al. , 2002 ), especially those caused by parasitoids (e.g. Moriya et al. , 1989 ). Cynipid inquilines are also known to have a significant role in local extinctions of pop-ulations such as Xanthoteras politum , which was affected by one Ceroptres and one Synergus species (only identified to ge-nus according to Washburn & Cornell, 1981 ). In the Palaearctic, inquilines are the main source of mortality in other gall wasp species, such as Cynips divisa and Andricus legitimus by Synergus pallipes (= nervosus ) and Synergus clandestinus re-spectively, according to Sitch et al. (1988) and Wiebes-Rijks (1980) . Thus, despite being an important part of cynipid gall communities, inquilines are probably the most neglected com-ponent within the system.
Xeric conditions were not associated with gall insect occur-rence. This is not to say that dry habitats do not influence cork oak gall wasp populations at larger, biogeographical scales, as it takes place in other galler species ( Price et al. , 1998; Gange et al. , 2002 ). The sampling scheme was not designed in the present study to detect xeric conditions, and therefore the varia-tion of such dry conditions among sites was too small. This per-mits establishment of the fact, however, that habitat fragmentation is the main factor influencing galler occurrence at the landscape scale.
Although the results presented here do not concern the process generating the community structure of such a tritrophic system, this work reports, for the first time, the impact of cork oak forest fragmentation on the cynipid gall wasp species. Further, since galls represent discrete microhabitats that support relatively closed communities of specialist inhabitants and their parasitoids, and the ease with which galls can be collected and incubated, gall insects appeared to be an appropriate biological model to study the effects of habitat fragmentation in tritrophic systems.
Acknowledgements
This research was supported by the Working Community of Pyrenees (Communauté de Travail des Pyrénées, Generalitat de Catalunya – Conseil Régional Midi-Pyrénées).
References
Andrén , H . ( 1994 ) Effects of habitat fragmentation on bird and mam-mals in landscapes with different proportions of suitable habitat: a review . Oikos , 71 , 355 – 366 .
Askew , P . ( 1961 ) On the biology of the inhabitants of oak galls of Cynipidae (Hymenoptera) in Britain . Transactions of the Society for British Entomology , 14 , 237 – 268 .
Askew , P . ( 1971 ) Parasitic Insects . Heinemann Education Books , London . Burnham , K.P. & Anderson , D.R . ( 2002 ) Model Selection and Multi-
modal Inference: A Practical Information – Theoretic Approach , 2nd edn . Springer , New York .
Chust , G. , Pretus , J.L.I. , Ducrot , D. , Bedòs , A. & Deharveng , L . ( 2003 ) Response of soil fauna to landscape heterogeneity: determining opti-mal scales for biodiversity modeling . Conservation Biology , 17 , 1712 – 1723 .
Cronin , J.T . ( 2003 ) Patch structure, oviposition behavior and the distri-bution of parasitism risk . Ecological Monographs , 73 , 283 – 300 .
Cuevas-Reyes , P. , Quesada , M. , Hanson , P. , Dirzo , R. & Oyama , K . ( 2004 ) Diversity of gall-inducing insects in a Mexican tropical dry forest: the importance of plant species richness, life-forms, host plant age and plant density . Journal of Ecology , 92 , 707 – 716 .
Davies , K.F. , Melbourne , B.A. & Margules , C.R . ( 2001 ) Effects of within- and between-patch processes on community dynamics in a fragmentation experiment . Ecology , 82 , 1830 – 1846 .
Dawkins , R . ( 1982 ) The Extended Phenotype: the Gene as a Unit of Selection . Oxford University Press , Oxford .
Doak , P . ( 2000 ) The effects of plant dispersion and prey density on parasitism rates in a naturally patchy habitat . Oecologia , 122 , 556 – 567 .
Fernandes , G.W. & Price , P.W . ( 1992 ) The adaptive signifi cance of in-sect gall distribution: survivorship of species in xeric and mesic habitats . Oecologia , 90 , 14 – 20 .
Gange , A.C. , Croft , R. & Wu , W . ( 2002 ) Gall insect and endophytic fungal co-occurrence in a xeric and mesic site . Ecological Ento-mology , 27 , 362 – 365 .
Guisan , A. , Edwards , T.C. Jr & Hastie , T . ( 2002 ) Generalized linear and generalized additive models in studies of species distributions: set-ting the scene . Ecological Modelling , 157 , 89 – 100 .
Guisan , A. & Zimmermann , N.E . ( 2000 ) Predictive habitat distribution models in ecology . Ecological Modelling , 135 , 147 – 186 .
Hanski , I . ( 1994 ) Patch-occupancy dynamics in fragmented landscapes . Trends in Ecology and Evolution , 9 , 131 – 135 .
Hastie , T.J. & Tibshirani , R.J . ( 1990 ) Generalized Additive Models . Chapman & Hall , London .
Holt , R.D . ( 1996 ) Temporal and spatial aspects of food web structure and dynamics . Food Webs: Integration of Patterns and Dynamics ( ed . by E. A. Polis & K. O. Winemiller ), pp. 255–257. Chapman & Hall , London .
Kareiva , P . ( 1987 ) Habitat fragmentation and the stability of predator – prey interactions . Nature , 326 , 388 – 390 .
Kruess , A . ( 1996 ) Folgen der Lebensraum-Fragmentierung für Pfl anze-Herbivor-Parasitoid-Gesellschaften: Artendiversität und Interak-tionen . Haupt , Bern .
Kruess , A. & Tscharntke , T . ( 2000 ) Species richness and parasitism in a fragmented landscape: experiments and fi eld studies with insects on Vicia sepium . Oecologia , 122 , 129 – 137 .

90 Guillem Chust, Lucas Garbin & Juli Pujade-Villar
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 82–91
Laurance , W.F. , Lovejoy , T.E. , Vasconcelos , H.L. , Bruna , E.M. , Didham , R.K. , Stouffer , P.C . et al . ( 2002 ) Ecosystem decay of Ama-zonian forest fragments: a 22-year investigation . Conservation Biol-ogy , 16 , 605 – 618 .
MacArthur , R.H. & Wilson , E.O . ( 1967 ) The Theory of Island Biogeog-raphy . Princeton University Press , Princeton, NJ .
Moriya , S. , Inoue , K. , Otake , A. , Shiga , M. & Mabuchi , M . ( 1989 ) Decline of the chestnut gall-wasp population, Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) after the establishment of Torymus sinensis Kamijo (Hymenoptera: Torymidae) . Applied Entomology and Zoology , 24 , 231 – 33 .
Nieves-Aldrey , J.L . ( 1987 ) Estado actual de conocimiento de la sub-família Cynipidae (Hym., parasitica, Cynipidae) en la Península Ibérica (Hym., Cynipidae) . EOS , 63 , 179 – 195 .
Nieves-Aldrey , J.L . ( 2001 ). Hymenoptera, Cynipidae. Fauna Ibérica , Vol. 16 ( ed . by M. A. Ramos et al. ). Museo Nacional de Ciencias Naturales . CSIC , Madrid .
Ninyerola , M. , Pons , X. & Roure , J.M . ( 2000 ) A methodological approach of climatological modelling of air temperature and precipi-tation through GIS techniques . International Journal of Climatology , 20 , 1823 – 1841 .
Pearman , P.B . ( 2002 ) The scale of community structure: habitat varia-tion and avian guilds in tropical forest understory . Ecological Monographs , 72 , 19 – 39 .
Pons , X . ( 1996 ) Estimación de la Radiación Solar a partir de modelos digitales de elevaciones. Propuesta metodológica. VII . Coloquio de Geografía Cuantitativa, Sistemas de Información Geográfi ca Y Teledetección ( ed . by J. Juaristi and I. Moro ), pp. 87–97. Vitoria , Gasteiz .
Price , P.W. , Fernandes , G. , Lara , A.C. , Brawn , J. , Gerling , D. , Barrios , H . et al . ( 1998 ) Global patterns in local numbers of insect galling species . Journal of Biogeography , 25 , 581 – 591 .
Price , P.W. , Roininen , H. & Tahvanainen , J . ( 1987 ) Plant age and attack by the bud galler, Euura mucronata . Oecologia , 73 , 334 – 337 .
Pujade-Villar , J . ( 2002 ) Una presentación excelente para un volumen con demasiados errores: Nieves-Aldrey (2001), Hymenoptera, Cynipidae , Fauna Ibérica . Boletín de la Asociación Española de Entomología , 26 , 143 – 159 .
Pujade-Villar , J . ( 2004 ) Les gales dels roures: una font de recursos per a molts organismes . Habitats , 9 , 11 – 25 .
Pujade-Villar , J . ( 2005 ) About two valid species: Saphonecrus gallae-pomiformis (Boyer de Fonscolombe, 1832) n. comb. & Synergus facialis Hartig, 1840 (Hym., Cynipidae: Synergini) . Boletín de la Asociación Española de Entomología , 29 , 137 – 138 .
Pujade-Villar , J. , Melika , G. , Ros-Farré , P. , Ács , Z. & Csóka , G . ( 2003 ) Cynipid inquiline wasps of Hungary, with taxonomic notes on the Western Palaearctic fauna (Hymenoptera: Cynipidae, Cynipinae, Synergini) . Folia Entomologica Hungarica , 64 , 121 – 170 .
Ros-Farré , P. & Pujade-Villar , J . ( 1998 ) Relacions tròfi ques en galles de la forma agàmica d’ Andricus kollari (Hartig, 1843) . Butlletí de la Institució Catalana d’Història Natural , 66 , 95 – 108 .
Schönrogge , K. , Walker , P. & Crawley , M.J . ( 1999 ) Complex life-cycles in Andricus kollari (Hymenoptera, Cynipidae) and their impact on associated parasitoid and inquiline species . Oikos , 84 , 293 – 301 .
Sitch , T.A. , Grewcock , D.A. & Gilbert , F.S . ( 1988 ) Factors affecting components of fi tness in a gall-making wasp ( Cynips divisa Hartig) . Oecologia , 76 , 371 – 375 .
Skrzypczynska , M . ( 2001 ) Studies on insects causing galls on the leaves of pedunculate oak Quercus robur in southern Poland . Anzeiger für Schädlingskunde , 74 , 40 – 42 .
Stehman , S.V . ( 1997 ) Selecting and interpreting measures of thematic classifi cation accuracy . Remote Sensing of Environment , 62 , 77 – 89 .
Stone , G.N. & Schönrogge , K . ( 2003 ) The adaptive signifi cance of insect gall morphology . Trends in Ecology and Evolution , 18 , 512 – 522 .
Stone , G.N. , Schönrogge , K. , Atkinson , R.J. , Bellido , D. & Pujade- Villar , J . ( 2002 ) The population biology of oak gall wasps (Hymenoptera: Cynipidae) . Annual Review of Entomology , 47 , 633 – 668 .
Torossian , G . ( 1971 ) Faune secondaire des galles de Cynipidae; I: Étude systematique des fourmies et des principaux arthropodes récoltés dans les galles . Insectes Sociaus , 3 , 135 – 154 .
Turner , M.G. , O’Neill , R.V. & Gardner , R.H. & Milne , B.T . ( 1989 ) Effects of changing spatial scale on the analysis of landscape pattern . Landscape Ecology , 3 , 153 – 162 .
Walker , P . ( 2002 ) Two new records for cynipid oak galls (Cynipidae: Hymenoptera) in Britain . Cecidology , 17 , 64 – 67 .
Washburn , J.O. & Cornell , H.V . ( 1981 ) Parasitoids, patches, and phe-nology: their possible role in the local extinction of a cynipid gall wasp population . Ecology , 62 , 1597 – 1607 .
Wiebes-Rijks , A.A . ( 1980 ) The identity of the gall-wasp causing stunted acorns (Hymenoptera, Cynipidae) . Netherlands Journal of Zoology , 30 , 243 – 253 .
Wood , S.N . ( 2000 ) Modelling and smoothing parameter estimation with multiple quadratic penalties . Journal of the Royal Statistical Society, B , 62 , 413 – 428 .
Accepted 9 August 2006
Gall wasps in cork oak fragmented forests 91
© 2007 The AuthorsJournal compilation © 2007 The Royal Entomological Society, Ecological Entomology, 32, 82–91
Appendix Biology of cork oak gall wasps in the Iberian peninsula sampled in Albera. The entries in bold are the gall forms used in this study. In the heterogonic simple cycle, the alternation of host between the two generations of gallers is not obligate, while in heterogonic heteroecic host, alternation is obligate.
Gallwasps
Body size of sexual forms 1 (female) (mm)
Body size of asexual forms 1 (mm)
Gall of sexual form
Gall of asexual form Cycle
C. glandium 1.8 – 2.0 2.5 – 3.0 Twigs of deciduous oak Acorns of evergreen oak
Heterogonic, heteroecic
C. rufescens 2.0 – 2.5 2.6 Twigs of deciduous oak Acorns of evergreen oak
Heterogonic, heteroecic
A. burgundus 1.4 – 1.6 – Buds of cork oak Unknown, but probably in buds of deciduous oak
Probably heterogonic, heteroecic
A. grossulariae 2 2.0 – 2.4 2.8 – 4.0 Infl orescences of cork oak
Buds of deciduous oak Heterogonic, heteroecic
A. hispanicus 1.7 – 2.0 3.5 – 4.5 Buds of cork oak Buds of deciduous oak Heterogonic, heteroecic N. saliens 1.7 – 1.9 1.3 Unripe acorns of
cork oak Leaves of cork oak Heterogonic simple
P. amenti 3 1.6 – 2.1 2.8 – 3.0 Twigs of cork oak Leaves, infl orescences and twigs of cork oak
Probably heterogonic simple
1 Source: Nieves-Aldrey (2001) . 2 According to Walker (2002) . 3 According to Garbin et al. (unpublished) .





























