Embed Size (px)
Citation preview

Marine Pollution Bulletin xxx (2012) xxx–xxx
Contents lists available at SciVerse ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Functional composition of Chaetodon butterflyfishes at a peripheraland extreme coral reef location, the Persian Gulf
Morgan S. Pratchett a,⇑, Andrew S. Hoey a,b, David A. Feary c, Andrew G. Bauman a, John A. Burt d,Bernhard M. Riegl e
a ARC Centre of Excellence for Coral Reef Studies, James Cook University, Townsville Q4811, Australiab Red Sea Research Center, King Abdullah University of Science and Technology, Thuwal, Saudi Arabiac School of the Environment, University of Technology, Sydney, 123 Broadway, NSW 2007, Australiad Biology, New York University, Abu Dhabi, PO Box 129188, Abu Dhabi, United Arab Emiratese National Coral Reef Institute, Nova Southeastern University Oceanographic Center, 8000 N. Ocean Drive, Dania, FL 33004, United States
a r t i c l e i n f o
Keywords:ButterflyfishesCoral reefsDisturbanceDietary specialisationHabitat preferencesScleractinian corals
0025-326X/$ - see front matter � 2012 Published byhttp://dx.doi.org/10.1016/j.marpolbul.2012.10.014
⇑ Corresponding author. Tel.: +61 7 47815747; fax:E-mail address: [email protected] (M.S
Please cite this article in press as: Pratchett, M.Sthe Persian Gulf. Mar. Pollut. Bull. (2012), http:
a b s t r a c t
The functional composition of reef fish assemblages is highly conserved across large biogeographic areas,but it is unknown whether assembly rules hold at biogeographical and environmental extremes for coralreefs. This study examined the functional composition of butterflyfishes in the Persian Gulf, MusandamPeninsula, and Gulf of Oman. Only five species of butterflyfishes were recorded during this study, andmostly just in the Gulf of Oman. Unlike most locations in the Indo–Pacific where butterflyfish assem-blages are dominated by obligate corallivores, the only obligate corallivore recorded, Chaetodon melapte-rus, was rare or absent at all locations. The most common and widespread species was Chaetodonnigropunctatus, which is shown to be a facultative corallivore. The diversity of butterflyfishes in the Per-sian Gulf is likely to have been constrained by its’ biogeographical history and isolation, but functionalcomposition appears to be further affected by limited abundance of prey corals and harsh environmentalconditions.
� 2012 Published by Elsevier Ltd.
1. Introduction
Coral reefs are among the most threatened aquatic ecosystems(Hoegh-Guldberg and Bruno, 2010). It is estimated that 30% of theworld’s coral reefs are already lost, and up to 70% of reefs face asimilar fate within the next 50 years (Wilkinson, 2004), rivallingthe total global loss of tropical rainforests due to deforestation(Laurance, 1999). Major causes of coral reef degradation includeoverfishing (Bellwood et al., 2012), declining water quality (e.g.,Fabricius et al., 2005) and direct damage to corals, caused by coralmining and destructive fishing (e.g., Baird et al., 2005). Moreover,these enduring threats are now being compounded by global cli-mate change (Hughes et al., 2003; Hoegh-Guldberg et al., 2007).Coral reefs are particularly vulnerable to global climate change(Walther et al., 2002), due to the extreme thermal sensitivities ofthe key habitat forming organisms, scleractinian corals (Hoegh-Guldberg et al., 2007), which bleach and die during prolongedexposure to anomalous temperatures. Habitat degradation andcoral loss further impact the abundance and community composi-tion of coral reef associated organisms, especially highly specia-lised species with strong reliance on specific coral species
Elsevier Ltd.
+61 7 47816722.. Pratchett).
., et al. Functional composition//dx.doi.org/10.1016/j.marpolbu
(Pratchett et al., 2008a; Stella et al., 2011). It is likely therefore, sus-tained and ongoing climate change will greatly disrupt the func-tional composition of reef fish assemblages, which otherwiseappear to be structured according to highly conserved assemblyrules (Bellwood and Hughes, 2001; Bellwood et al., 2002).
Butterflyfishes (family Chaetodontidae) are among the mostspecialised of coral reef fishes, with many species having an obli-gate dependence on scleractinian corals for food (Reese, 1981;Tricas, 1989) and/or recruitment (Harmelin-Vivien, 1989; Pratchettet al., 2008b). Of the 78 species of butterflyfishes that live on coralreefs, 78% (61 species) feed at least in part on live corals (Cole et al.,2008). The remaining species feed on a variety of benthic and/orpelagic prey items, such as annelid worms, small crustaceans, mol-luscs, and macroalgae (Naglerkerken et al., 2009). Corallivorousfishes can be broadly divided into obligate or facultative coralli-vores (Reese, 1977, 1981; Harmelin-Vivien and Bouchon-Navaro,1983; Cox, 1994), which has important ramifications for theirdependence on coral resources (Harmelin-Vivien and Bouchon-Navaro, 1983) and susceptibility to disturbance (Reese, 1981;Williams, 1986). Corallivores that have diets containing more than80% coral exhibit disproportionate declines in abundance followingcoral loss, indicating that these species are obligately dependant oncoral for their survival (Pratchett et al., 2008a). Facultative coralli-vores, meanwhile, often feed opportunistically on live coral tissue
of Chaetodon butterflyfishes at a peripheral and extreme coral reef location,l.2012.10.014

2 M.S. Pratchett et al. / Marine Pollution Bulletin xxx (2012) xxx–xxx
but can withstand temporary declines in abundance of corals (e.g.,Williams, 1986), presumably by increasing consumption of non-coral prey (Pratchett et al., 2006).
Corallivorous fishes are an important component of reef fishassemblages. Despite initial assumptions that few fishes are capa-ble of feeding on corals (e.g., Connell, 1973), recent research hasshown that there are a high diversity of reef fishes that consumelive coral (Cole et al., 2008; Rotjan and Lewis, 2008). Moreover,these fishes consume significant quantities of coral (Hoey andBellwood, 2008; Cole et al., 2012), and provide an importanttrophic link provided between scleractinian corals and higher con-sumers (Glynn, 2004). Thus far, coral feeding has been documentedin 128 species and 11 families of coral reef fishes (Cole et al., 2008).However, butterflyfishes account for more than half of all knowncorallivores, and corallivory is considered synonymous with thisfamily (Cole et al., 2008). Not only do most butterflyfishes eat coral,butterflyfish assemblages in coral reef habitats across the Indo-Pa-cific tend to be dominated by obligate corallivores. On Australia’sGreat Barrier Reef, obligate corallivores accounted for 67.8% of but-terflyfishes recorded across 47 reefs surveyed for 18 years, and thefour most abundant species (Chaetodon rainfordi, Chaetodon lunul-atus, Chaetodon trifascialis, and Chaetodon aureofasciatus) were allobligate corallivores (Emslie et al., 2010). In contrast, no obligatecoral feeding fish are found in the Caribbean and butterflyfishassemblages are dominated by the facultative corallivore, Chaeto-don capistratus (Birkeland and Neudecker, 1981). To test whetherthis lack of obligate corallivores is attributable to low diversity ofbutterflyfishes in the Caribbean, or limitations in food availability,it is necessary to consider other peripheral and species depauper-ate locations. Few Chaetodon butterflyfishes are purported to occurin the Persian Gulf (Table 1), though several species (Chaetodon dia-leucos, Chaetodon collare, Chaetodon lunula, and Chaetodon leucople-ura) have recorded distributions extending to adjacent areas justoutside the Persian Gulf (Allen et al., 1998).
The purpose of this study was to quantify patterns of prey useby Chaetodon butterflyfishes, and assess their abundance fromthe Persian Gulf, along the Musandam Peninsula and to the Gulfof Oman, testing for changes in the functional composition of but-terflyfish assemblages across gradients in local environmental con-ditions. Coral reefs in the Persian Gulf are subject to extremeenvironmental conditions (Sheppard et al., 2010; Riegl et al.,2011), including the highest annual variability in seawater temper-atures (>20 �C range between extreme maximum and minimumtemperatures recorded at several locations in the gulf; Sheppardet al., 2010), very high salinity (>39 psu; Sheppard et al., 2010), sig-nificant seasonal fluctuations in insolation, and high levels of tur-bidity and sedimentation (Riegl, 1999;). Moreover, coral reefs inthis region have experienced frequent coral bleaching events since1996 (Burt et al., 2011a; Riegl et al., 2011). Given the need to betterunderstand coral reef resilience in the face of global climate change(McClanahan et al., 2007) the Persian Gulf may be an ideal locationto evaluate the role of extreme environmental conditions and
Table 1Distribution and diet of five sympatric Chaetodon species from the Persian Gulf and Sea o
Species Distribution
Chaetodon (Corallochaetodon)melapterus Guichenot, 1863
Persian Gulf and Gulf of Oman south to the Red Secoast of Yemen (Righton et al., 1996)
Chaetodon (Rabdophorus)nigropunctatus Sauvage, 1880
Persian Gulf and Gulf of Oman south to tropical EasAfrica (Allen et al., 1998)
Chaetodon (Rabdophorus)vagabundus Linnaeus, 1978
Widely distributed in the Indo–Pacific, and recordein the Persian Gulf (Shokri et al., 2005)
Chaetodon (Rabdophorus)gardineri Norman, 1939
North Western Indian Ocean: Gulf of Oman to theGulf of Aden (Allen et al., 1998)
Chaetodon (Rabdophorus) collareBloch, 1787
Indo-west Pacific, including Persian Gulf and northJapan (Allen et al., 1998)
Please cite this article in press as: Pratchett, M.S., et al. Functional compositionthe Persian Gulf. Mar. Pollut. Bull. (2012), http://dx.doi.org/10.1016/j.marpolbu
recurrent bleaching events in structuring coral reef assemblages.Moreover, this is the first quantitative study of feeding behaviourfor several species of butterflyfishes that are restricted to PersianGulf and/or northwestern Indian Ocean, including Chaetodon mel-apterus, and Chaetodon nigropunctatus.
2. Materials and methods
2.1. Field sampling
To test for spatial variation in abundance and composition ofcoral reef butterflyfishes, surveys were conducted at five locations(Fig. 1), including the southern Persian Gulf (Abu Dhabi), theMusandam Peninsula (Musandam, Fujairah) and the Gulf of Oman(Muscat, Ras Al Hadd). Each location was initially surveyed inNovember 2008, and then re-surveyed in January, 2012. Withineach location, abundance of butterflyfishes were quantified at mul-tiple sites (2–4 sites per location) using eight replicate 30 � 5 mbelt transects, following Feary et al. (2010). All sites were similarin terms of depth (c. 5–6 m), distance to shore (c. 100 m) and gen-eral habitat (Feary et al., 2010). Spatial and temporal variation incover and taxonomic composition of scleractinian corals was re-corded simultaneously. Benthic communities were sampled using0.25 m2 quadrats photographed at 3 m intervals on the same tran-sects sampled for fish (11 quadrats per transect). Photographs wereanalysed using 50 random point-intercepts in CPCe software(Kohler and Gill, 2006), with all corals identified to genus. Alltransects were run from haphazardly selected starting points andrun parallel to depth contours.
Sustained and ongoing monitoring is critical for understandingspatial and temporal changes in the ichthyofauna associated withcoral reef habitats throughout the Arabian region. However, therehas been a long-history of sampling of fishes and corals (albeit itvery patchy in time and space) in the southern Persian Gulf. Thisstudy brings together survey data from the coastline of Dubai,comparing the abundance and composition of butterflyfishes in1995 to extensive surveys conducted from 2008 to 2012. Impor-tantly, the period between 1995 and 2006 saw substantial changesin coastal environments off Dubai, affected by severe mass-bleachingof scleractinian corals in 1996 (Riegl, 1999) and extensive shorelinemodification associated with the construction of Palm Jumeirah(2002–2006) and other coastal developments (Sale et al., 2011).
2.2. Feeding observations
The range of prey types eaten by each species of butterflyfish, aswell as the proportional use of different prey types, was ascer-tained from field observations, following Pratchett (2005). In situfeeding observations were conducted opportunistically by MSPand ASH during surveys conducted in January 2012, such thatsampling intensity varied in approximate accordance with
f Oman.
Diet
a Reported to feed exclusively on coral polyps (Lieske and Myers, 1996)
t Assumed to be an obligate corallivore in Iran (Shokri et al., 2005), but otherspecies in this sub-genus are mostly facultative corallivores
d Benthic omnivore (Pratchett, 2005), which is known to feed on coral polyps insome locations (Cole et al., 2008)Reported to feed on coral polyps (Michael 2004), but no quantitativeassessment of dietary compositionReported to feed on coral polyps (Allen et al., 1998), but no quantitativeassessment of dietary composition
of Chaetodon butterflyfishes at a peripheral and extreme coral reef location,l.2012.10.014

Fig. 1. Map of study locations in the Persian Gulf, along the Arabian Peninsula and in the Gulf of Oman.
M.S. Pratchett et al. / Marine Pollution Bulletin xxx (2012) xxx–xxx 3
differential abundance of each butterflyfish species (Table 1).During each feeding observation, an individual butterflyfish wasfollowed for 3 min, recording the total number of bites taken fromeach type of hard-coral, soft-coral, or any other non-coral benthos.Corals were mostly identified to genus, though we pooled Favitesand Favia, due to concerns about the identification of somecolonies. No attempt was made to identify the specific source ofprey for butterflyfishes feeding on consolidated reef pavement,coral rubble and sand, but it was assumed that they were targetingsmall motile invertebrates, such as polychaetes and crustaceans,following Naglerkerken et al., (2009). Most butterflyfishes contin-ued to feed despite the presence of divers, but observations wereaborted on rare occasions where fish fled from the diver or soughtshelter within the reef matrix. In situ feeding observations provideecological data on the microhabitat from which the food items areselected, rather than actual items being ingested (Naglerkerkenet al., 2009). This may overestimate the reliance on live corals fornon-coral feeders that consume motile invertebrates from thesurface of live corals. However, in situ feed observations (cf. gutcontent analysis or stable istopes) are the only way to discernthe specific types of corals consumed by obligate coral feedingbutterflyfishes (Naglerkerken et al., 2009).
2.3. Statistical analyses
Feeding selectivity of coral-feeding butterflyfishes was assessedby comparing the proportional use of coral consumed (estimatedby the number of bites on each genus) to their relative availabilitywithin each site. Insufficient observations were conducted to testfor spatial variation in feeding preferences (e.g., among locations),but selectivity by each individual was assessed based on the spe-cific availability of corals in their local area, estimated fromphoto-transects. Dietary selectivity was analysed using log-likelihoodstatistics, following Pratchett (2007). Resource selection functionswere then used to see which coral genera were used more or lessfrequently than expected. Use was deemed disproportionate(either positively or negatively) to availability if the 95% CI didnot encompass 1 (Manly et al., 1993). To account for variation in
Please cite this article in press as: Pratchett, M.S., et al. Functional compositionthe Persian Gulf. Mar. Pollut. Bull. (2012), http://dx.doi.org/10.1016/j.marpolbu
sampling intensity among species, feeding rates, as well asdifferences in proportional coral consumption (Table 1), dietaryspecialisation was calculated based on the mean number ofdifferent corals (genera) consumed per 100 bites, followingPratchett (2007).
Variation in abundance of Chaetodon butterflyfishes was ana-lysed using ANOVA, testing for differences between years (2008and 2012) and among locations (Abu Dhabi, Musandam, Fujairah,Muscat, and Ras Al Hadd). Variation among sites within locationswas not analysed due to inconsistencies in the number of sitessampled, and type III sums of squares were used to account forthe imbalanced sampling design. All count data was log10 trans-formed prior to analyses. Spatial and temporal variation in compo-sition of butterflyfish assemblages was then analysed usingMANOVA, testing for variation in relative abundance of butterfly-fish species among locations and between years. Correlation wasalso used to test whether variation in abundance of individual but-terflyfishes was related to local coral cover. It was expected a priori,that abundance of obligate corallivores would be strongly corre-lated with variation in live coral cover, but less so for facultativeand non-corallivores (Pratchett and Berumen, 2008). As such, sep-arate analyses were conducted for each species of butterflyfish.However, only those butterflyfishes that were sufficiently abun-dant and or widespread (C. nigropunctatus and C. melapterus) couldbe considered.
3. Results
3.1. Dietary composition
Feeding observations were completed for 96 individual butter-flyfishes from five species (Table 2). The majority of observations(60/96) were conducted for C. nigropunctatus, which was commonat all locations, whereas few feeding observations were conductedfor three rare species (C. collare, Chaetodon gardineri, and especiallyChaetodon vagabundus). Only one (out of five) species of butterfly-fish, C. melapterus, was recorded to feed exclusively on scleractin-ian corals (Table 2), and is thus considered to be an obligate
of Chaetodon butterflyfishes at a peripheral and extreme coral reef location,l.2012.10.014

Table 2Bite rates and coral use for 5 sympatric Chaetodon species from Arabian peninsula and Gulf of Oman. Bite rates are the mean number of bites taken during each three-minuteobservation. Coral use is presented for both the overall percentage of bites taken from scleractinian corals (% coral use), as well as the percentage of bites (of those taken fromcorals) taken from each of the four major coral taxa (Faviidae, Porites, Montipora and Acropora). Selectivity and significance of selectivity (p) is calculated only for corallivorousbutterflyfishes, because no attempt was made to discern specific prey items for fishes when feeding on non-coral substrates.
Species n Bite rate % Coral use % Faviidae % Porites % Montipora % Acropora Selectivity p
Obligate hard-coral feedersChaetodon melapterus 13 16.07 100.0 24.8 39.7 8.1 13.4 37.12 <0.01
Facultative corallivoresChaetodon nigropunctatus 60 13.38 27.7 36.2 41.7 0 3.2 50.91 0.09Chaetodon vagabundus 2 21.00 39.6 17.3 0 0 0 3.68 0.15
Non-corallivoresChaetodon gardineri 9 13.33 0.0 NA NAChaetodon collare 12 0.00 0.0 NA NA
-2
0
2
4
6
8
10
12
-0.5
0
0.5
1
1.5
2
2.5
3
3.5
4
Sele
ctio
n fu
nctio
ns
-4
-2
0
2
4
6
8
10
12
s s l l
(a) Chaetodon melapterus
(b) Chaetodon nigropunctatus
(c) Chaetodon vagabundus
4 M.S. Pratchett et al. / Marine Pollution Bulletin xxx (2012) xxx–xxx
corallivore. A further two species, C. nigropunctatus and C. vagabun-dus, were facultative corallivores, taking 27.7% and 39.6% of biteson live corals, respectively. The remaining species (C. gardineriand C. collare) did not take any bites from live corals. In the caseof C. gardineri, 95% of bites (114/120) across all individuals were ta-ken from consolidated carbonate pavement, with the remainingbites taken from either sand or rubble substrata. C. collare, mean-while, was never seen to take any bites on benthic substrata.Rather, all individuals of C. collare were observed in mid-water,3–5 m above the substratum. These fishes were making rapid,but small movements with their mouths, but it is unclear whetherthese fishes were actually feeding on planktonic prey. Bite rates forall benthic feeding butterflyfishes were fairly consistent, rangingfrom 13.33 bites per 3-min, for C. gardineri, up to 21.00 bites per3-min for C. vagabundus (Table 2).
Coral-feeding butterflyfishes (C. melapterus, C. nigropunctatus,and C. vagabundus) consumed a wide range of different corals(including at least nine different coral genera), mostly in accor-dance with their availability (Fig. 2). Only C. melapterus exhibitedsignificant feeding selectivity (Table 2), consuming Favia and Fa-vites, as well as Acropora disproportionately more than expectedbased on availability of these coral genera (Fig. 1). The predomi-nant genus eaten by C. melapterus was actually Porites (accountingfor 39.7% of bites), but these were eaten in approximate accordancewith their availability. Facultative corallivores, C. nigropunctatusand C. vagabundus, consumed a range of coral genera in approxi-mate accordance with their availability, but neither species con-sumed Montipora and several other genera (Fig. 2). Thepredominant corals consumed by C. nigropunctatus were Poritesand Pocillopora, but only Favia and Favites were consumed dispro-portionately more than expected from their availability (Fig. 2).The facultative corallivores (C. nigropunctatus and C. vagabundus)supplemented consumption of scleractinian corals with bites takenfrom other sessile organisms (e.g., tunicates and soft corals), butmost of their bites (up to 71.0%) were taken from consolidated car-bonate reef substrata devoid of any macro-invertebrates.
Fav
ia a
nd F
avit
e
Por
ite
Mon
tipo
ra
Acr
opor
a
Poc
illo
pora
Oth
er h
ard
cora
Non
-cor
a
Fig. 2. Dietary selection by coral-feeding butterflyfishes from the southern PersianGulf and Gulf of Oman. Selection functions significantly greater than 1 indicatedthat corals were used more than expected from their availability (i.e., selected),while selection functions less than 1 indicated that corals were used less thanexpected (i.e., avoided).
3.2. Spatial and temporal patterns of abundance
A total of 1180 butterflyfishes were counted across all 192 tran-sects surveyed in 2008 and 2012, giving a mean of 6.14 (±0.56 SE)butterflyfishes per 150 m2 (per transect). However, abundance ofbutterflyfishes varied significantly among locations and betweenyears (Table 3). Variation in abundance among locations was con-sistent between years, but significant declines in abundance wererecorded at Abu Dhabi, Musandam and Fujairah between 2008and 2012 (Fig. 3). At Abu Dhabi, mean abundance of butterflyfishesdeclined 75.6% from 2.56 (±1.19 SE) butterflyfishes per 150 m2 in2008, down to 0.65 (±0.28 SE) butterflyfishes per 150 m2 in 2012.Even at Fujairah, which had highest densities of butterflyfishes in
Please cite this article in press as: Pratchett, M.S., et al. Functional compositionthe Persian Gulf. Mar. Pollut. Bull. (2012), http://dx.doi.org/10.1016/j.marpolbu
2008 (14.83 ± 2.60 SE butterflyfishes per 150 m2), the abundanceof butterflyfishes declined by 67.9%. Declines in abundance wereapparent, but not significant at Muscat and Ras Al Hadd (Fig. 2).
of Chaetodon butterflyfishes at a peripheral and extreme coral reef location,l.2012.10.014
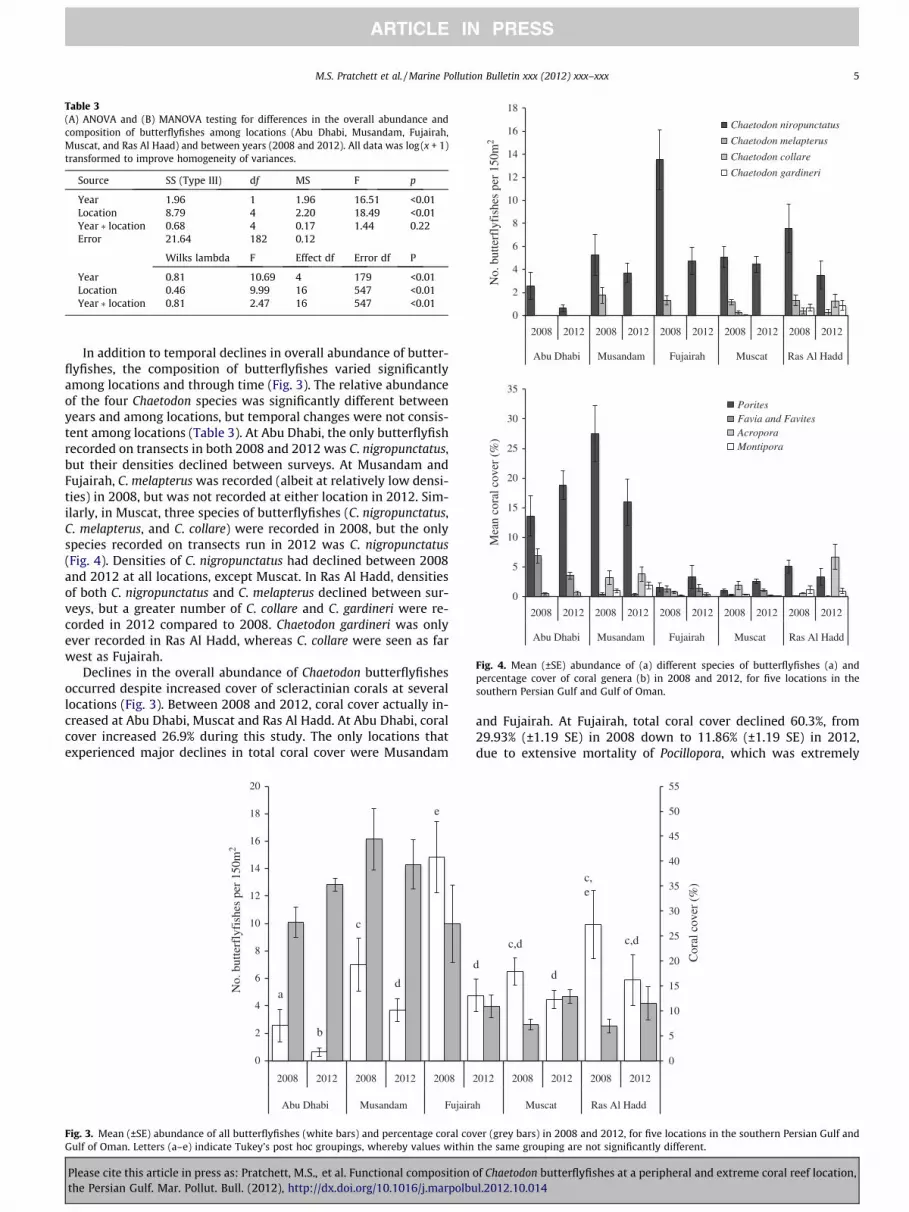
Table 3(A) ANOVA and (B) MANOVA testing for differences in the overall abundance andcomposition of butterflyfishes among locations (Abu Dhabi, Musandam, Fujairah,Muscat, and Ras Al Haad) and between years (2008 and 2012). All data was log(x + 1)transformed to improve homogeneity of variances.
Source SS (Type III) df MS F p
Year 1.96 1 1.96 16.51 <0.01Location 8.79 4 2.20 18.49 <0.01Year � location 0.68 4 0.17 1.44 0.22Error 21.64 182 0.12
Wilks lambda F Effect df Error df P
Year 0.81 10.69 4 179 <0.01Location 0.46 9.99 16 547 <0.01Year � location 0.81 2.47 16 547 <0.01
0
2
4
6
8
10
12
14
16
18
2008 2012 2008 2012 2008 2012 2008 2012 2008 2012
Abu Dhabi Musandam Fujairah Muscat
No.
but
terf
lyfi
shes
per
150
m2
Chaetodon niropunctatus
Chaetodon melapterus
Chaetodon collare
Chaetodon gardineri
0
5
10
15
20
25
30
35
2008 2012 2008 2012 2008 2012 2008 2012 2008 2012
Abu Dhabi Musandam Fujairah Muscat Ras Al Hadd
Mea
n co
ral c
over
(%
)
PoritesFavia and FavitesAcroporaMontipora
Ras Al Hadd
Fig. 4. Mean (±SE) abundance of (a) different species of butterflyfishes (a) andpercentage cover of coral genera (b) in 2008 and 2012, for five locations in thesouthern Persian Gulf and Gulf of Oman.
M.S. Pratchett et al. / Marine Pollution Bulletin xxx (2012) xxx–xxx 5
In addition to temporal declines in overall abundance of butter-flyfishes, the composition of butterflyfishes varied significantlyamong locations and through time (Fig. 3). The relative abundanceof the four Chaetodon species was significantly different betweenyears and among locations, but temporal changes were not consis-tent among locations (Table 3). At Abu Dhabi, the only butterflyfishrecorded on transects in both 2008 and 2012 was C. nigropunctatus,but their densities declined between surveys. At Musandam andFujairah, C. melapterus was recorded (albeit at relatively low densi-ties) in 2008, but was not recorded at either location in 2012. Sim-ilarly, in Muscat, three species of butterflyfishes (C. nigropunctatus,C. melapterus, and C. collare) were recorded in 2008, but the onlyspecies recorded on transects run in 2012 was C. nigropunctatus(Fig. 4). Densities of C. nigropunctatus had declined between 2008and 2012 at all locations, except Muscat. In Ras Al Hadd, densitiesof both C. nigropunctatus and C. melapterus declined between sur-veys, but a greater number of C. collare and C. gardineri were re-corded in 2012 compared to 2008. Chaetodon gardineri was onlyever recorded in Ras Al Hadd, whereas C. collare were seen as farwest as Fujairah.
Declines in the overall abundance of Chaetodon butterflyfishesoccurred despite increased cover of scleractinian corals at severallocations (Fig. 3). Between 2008 and 2012, coral cover actually in-creased at Abu Dhabi, Muscat and Ras Al Hadd. At Abu Dhabi, coralcover increased 26.9% during this study. The only locations thatexperienced major declines in total coral cover were Musandam
0
2
4
6
8
10
12
14
16
18
20
2008 2012 2008 2012 2008
Abu Dhabi Musandam Fujaira
No.
but
terf
lyfi
shes
per
150
m2
a
b
c
d
e
Fig. 3. Mean (±SE) abundance of all butterflyfishes (white bars) and percentage coral covGulf of Oman. Letters (a–e) indicate Tukey’s post hoc groupings, whereby values within
Please cite this article in press as: Pratchett, M.S., et al. Functional compositionthe Persian Gulf. Mar. Pollut. Bull. (2012), http://dx.doi.org/10.1016/j.marpolbu
and Fujairah. At Fujairah, total coral cover declined 60.3%, from29.93% (±1.19 SE) in 2008 down to 11.86% (±1.19 SE) in 2012,due to extensive mortality of Pocillopora, which was extremely
0
5
10
15
20
25
30
35
40
45
50
55
2012 2008 2012 2008 2012
h Muscat Ras Al Hadd
Cor
al c
over
(%
)
dd
c,d c,d
c,e
er (grey bars) in 2008 and 2012, for five locations in the southern Persian Gulf andthe same grouping are not significantly different.
of Chaetodon butterflyfishes at a peripheral and extreme coral reef location,l.2012.10.014

6 M.S. Pratchett et al. / Marine Pollution Bulletin xxx (2012) xxx–xxx
abundant in 2008. Importantly, cover of branching corals (e.g.,Acropora) was very low and actually declined at nearly all loca-tions, except Ras Al Hadd (Fig. 4). In locations where coral coveractually increased over time (e.g., Abu Dhabi), it was mostly dueto increases in abundance of Porites. Overall, there were no signif-icant correlations between abundance of butterflyfishes versuspercentage cover of scleractinian corals. The strongest correlationwas for C. nigropunctatus (R = �0.13), but the correlation wasnegative.
3.3. Long-term trends for Dubai
In 1995, before the first recorded mass-bleaching event and be-fore extensive coastal development in Dubai, mean densities of C.nigropunctatus were 2.35 (±0.81 SE) butterflyfishes per 150 m2 incoral-dominated habitats where Palm Jebel Ali was subsequentlyconstructed. Following extensive coastal modification (includingconstruction of both Palm Jebel Ali and Palm Jumeirah), mean den-sities of C. nigropunctatus on the only remaining hard subetrate (onbreakwaters with some coral growth), ranged from 0.19 (±0.81 SE)to 1.72 (±1.25 SE) butterflyfishes per 150 m2 in Dubai, dependingupon season and year. The only other butterflyfish recorded off Du-bai during surveys conducted in 1995 was C. melannotus, whichwas recorded on a single transect in 1995. This species was alsosighted in 2007, but was not recorded on transects, nor was it re-corded during surveys conducted in 2012.
4. Discussion
4.1. Dietary composition and specialisation
Corallivorous butterflyfishes can be broadly divided into obli-gate versus facultative corallivores, which differ greatly in theirdependence on coral resources and therefore, their susceptibilityto disturbance (Cole et al., 2008; Pratchett et al., 2008a). Obligatecorallivores (operationally defined based on >80% hard corals intheir diet; Pratchett et al., 2008a) tend to be much more prevalentthan facultative corallivores, but also exhibit disproportionate de-clines in abundance following coral loss (e.g., Pratchett et al.,2006). In this study, only C. melapterus fed exclusively on hard cor-als, taking bites from the surface of several different corals includ-ing Acropora, Pocillopora, Porites, as well as Favia and Favites. Theseobservations are consistent with those of Lieske and Myers (1996),who reported that C. melapterus feeds on live corals. Moreover, allbutterflyfishes within the subgenus Corallochaetodon, and espe-cially sister species to C. melapterus (e.g., Chaetodon trifasciatusand C. lunulatus), tend to be obligate corallivores that consume awide range of different corals (e.g., Bellwood et al., 2009).
As for C. melapterus, the dietary composition of C. nigropunctatushas never been rigorously quantified. Lieske and Myers (1996) re-ported that C. nigropunctatus feeds at least partially on coral pol-yps. Moreover, Shokri et al. (2005) revealed a strong positiverelationship between abundance of C. nigropunctatus and live coralcover on Iranian coral reefs and implicitly assumed that this spe-cies feeds primarily, if not exclusively, on live coral. Extensive feed-ing observations in the southern Persian Gulf and Gulf of Oman,show that C. nigropunctatus is a facultative corallivore and onlytakes a relatively small proportion (<20%) of its’ bites from live cor-als. Accordingly, most species with the subgenus Rabdophorus(including C. vagabundus and C. gardineri) are facultative or non-coral feeders, which feed mainly on small motile invertebrates(Pratchett, 2005). Some species of the subgenus Rabdophorus(e.g., C. lunula) are also known to feed on plankton (Harmelin-Vivien,1989), which might explain lack of benthic feeding recorded forC. collare. The extent to which these species actually consumescleractinian corals versus motile invertebrates may vary
Please cite this article in press as: Pratchett, M.S., et al. Functional compositionthe Persian Gulf. Mar. Pollut. Bull. (2012), http://dx.doi.org/10.1016/j.marpolbu
regionally, according to changes in the availability of corals versusother non-coral prey. It is rare however, that corals constitute morethan 50% of the diet for these species (but see Graham, 2007).
Most studies that have examined the specific diets of corallivor-ous butterflyfishes have revealed striking patterns of prey prefer-ence (Reese, 1981; Irons, 1989; Cox, 1994; Pratchett, 2005;Graham, 2007), whereby corallivores tend to eat only a small pro-portion of available corals, and predominantly Acropora or Pocillo-pora corals. Despite these inherent preferences, butterflyfishes mayincrease the range of coral prey consumed when preferred prey arenot available, either in certain habitats (Berumen et al., 2005) orfollowing habitat degradation (Pratchett et al., 2004). Selectivityrecorded in the southern Persian Gulf was much lower than hasbeen reported in other locations (e.g., Pratchett, 2007), which ispartly due to the limited resolution used to distinguish coral prey(i.e., genera versus species). Even so, there was limited evidence ofstrong feeding preferences, whereby most butterflyfishes feed pre-dominantly on the most common corals, Porites. High levels of eco-logical versatility are probably necessary for species persistencegiven limited availability of Acropora and Pocillopora, especiallywithin the southern Persian Gulf.
4.2. Functional composition of butterflyfish assemblages
The southern Persian Gulf is unique among locations that sup-port coral reef environments (Coles, 2003; Riegl, 2003; Sheppardet al., 2010), experiencing temperatures above and below thenormal limits for coral growth. Even so, coral-dominated habitatsand associated fish communities do occur within the Persian Gulf(Riegl, 1999; Burt et al., 2008, 2011b; Feary et al., 2010). However,there were remarkably few species of butterflyfishes recorded atsites surveyed within the Persian Gulf, and the functional compo-sition of butterflyfishes is very different to that recorded on coralreefs throughout much of the Indo–Pacific. At most locations,including the Red Sea (Bouchon-Navaro, 1980), Chagos (Pratchettet al., 2012), Australia’s Great Barrier Reef (Emslie et al., 2010)and French Polynesia (Berumen and Pratchett, 2006) obligate cor-al-feeding species dominate butterflyfish assemblages. In thesouthern Persian Gulf, however, the dominant butterflyfish wasC. nigropunctatus, while the only obligate coral feeding butterfly-fish, C. melapterus, was not recorded at Abu Dhabi (southern Per-sian Gulf). C. melapterus, as well as other butterflyfishes (C.vagabundus, C. collare) were recorded at Musandam (not alwayson transects), and is also reported from Iranian coral reefs, withinthe Persian Gulf (Shokri et al., 2005). This suggests that there areonly a few select species that can actually tolerate extreme envi-ronmental conditions (temperature and sedimentation) in thesouthern Persian Gulf (Sheppard et al., 1992; Coles, 2003). Alterna-tively, sustained and ongoing degradation of coral reef habitats(e.g., Riegl, 1999) may have led to recent extirpation and/or shiftsin the relative abundance of coral reef butterflyfishes (Shokriet al., 2005).
Since 1996, there have been significant changes to coral reefhabitats within the Persian Gulf (Riegl, 1999, 2003), largely charac-terised by declines in abundance of Acropora, and concomitant in-creases in abundance of faviids and Porites (Burt et al., 2008,2011a). Acropora corals are an important food source for corallivor-ous fishes (Pratchett, 2005; Cole et al., 2008), as well as providingcritical recruitment habitat for many butterflyfishes (Pratchettet al., 2008b). As such, local depletion of Acropora, even withoutsignificant declines in total coral cover, might account for lowdiversity and abundance of butterflyfishes recorded in the south-ern Persian Gulf (Berumen and Pratchett, 2006). Surveys of butter-flyfishes conducted before the 1996 mass-bleaching and extensivecoastal developments, revealed high abundance of C. nigropuncta-tus off Dubai. Densities of C. nigropunctatus were much lower along
of Chaetodon butterflyfishes at a peripheral and extreme coral reef location,l.2012.10.014

M.S. Pratchett et al. / Marine Pollution Bulletin xxx (2012) xxx–xxx 7
extensive breakwaters of Palm Jumeirah in 2007–2008, but this ispartly explained by the young age of these structures. Burt et al.(2009) showed that mature breakwaters (>25 years) have >50%coral cover and C. nigropunctatus is more abundant on these break-waters, than they are on natural reefs. Importantly, there is no evi-dence that coral-feeding butterflyfishes (e.g., C. melapterus) werepreviously abundant off Dubai (e.g., prior to the mass-bleachingand extensive modification of coastal habitats). The only other but-terflyfish recorded off Dubai was C. melannotus, and this was onlyrecorded on one transect in 1995. We cannot discount the possibil-ity that extinction filtering over longer timeframes caused selectiveremoval of highly specialised, obligate coral feeding butterflyfishes,which dominate butterflyfish assemblages in many other parts ofthe Indo–Pacific. More likely, however, is that the limited develop-ment of coral reefs ecosystems within the southern Persian Gulf(Sheppard et al., 2010) has limited the capacity for obligatecoral-feeding butterflyfishes to evolve, colonise or persist in thisregion (Burt et al., 2011b). Similarly, there are no obligatecoral-feeding butterflyfishes in the Caribbean, or far eastern Pacific,possibly due to the historical lack of key coral species (Cowmanand Bellwood, 2011).
Fish and coral assemblages in the Persian Gulf and Gulf of Omanare generally depauperate, which may be due to harsh local envi-ronmental conditions that limit the range of species that can livein these locations, or an inherently low species pool due to geo-graphical location and isolation (Riegl, 2003; Coles, 2003; Fearyet al., 2010; Burt et al., 2011b). Extreme temperatures recordedin the Persian Gulf are much higher and much lower than for othertropical enclosed seas, such as the Red Sea. In the southern PersianGulf the annual temperature range is typically >13 �C, compared to9–10 �C on the Mussandam Peninsula (Bauman et al., 2012). Incontrast, annual temperature fluctuations in sea surface tempera-ture for Eilat in the Red Sea are only 7–8 �C, ranging from 20 to21 �C in winter to 26–28 �C in summer (Labiosa et al., 2003). Thereare also well-established biogeographical barriers in the north-western Indian Ocean (e.g., Blum, 1989; Sheppard et al., 1992), sep-arating Arabian waters from the remainder of the Indian Ocean.This does not in itself explain the low diversity in the Persian Gulfand Gulf of Oman. The Red Sea and Gulf of Aden, for example, havea total of 11 species of Chaetodon butterflyfishes (Zekeria et al.,2005). However, the Persian Gulf is much shallower and youngercompared to the Red Sea and Gulf of Aden (Sheppard et al.,2010), which is likely to have constrained both evolution and col-onisation by reef-associated fishes. Our study revealed a system-atic decline in species richness of butterflyfishes from the Gulf ofOman, through the Strait of Hormuz, and down into the southernPersian Gulf. This is the expected pattern if reef fishes colonised,rather than evolved, within the Persian Gulf. However, the greatestdecline in species richness was between Muscat and Fujairah (inthe Gulf of Oman), whereas we might expect major differences iffish communities within versus outside of the Persian Gulf, dueto limited larval exchange through the narrow Straits of Hormuz.
Despite weak correlations between the abundance of butterflyf-ishes and live coral cover, regional differences in habitat structureare likely to influence butterflyfish assemblages across the PersianGulf and Gulf of Oman (Burt et al., 2011b). In the southern PersianGulf, high levels of sedimentation and limited areas of hard sub-strate constrain coral growth and reef formation (Sheppard et al.,2010). Even when corals do grow, there is limited vertical relief,thereby limiting the habitat availability and structure for coral reeffishes (Feary et al., 2010). This is reflected in the absence of special-ist, coral-feeding butterflyfishes. On the Musandam Peninsula andGulf of Oman, meanwhile, corals grow on abundant rock slopes, orconstruct extensive reef structures. Higher diversity of corals alsoprovides increased prey for obligate coral feeding butterflyfishes.Further, recent declines in the abundance of butterflyfishes are al-
Please cite this article in press as: Pratchett, M.S., et al. Functional compositionthe Persian Gulf. Mar. Pollut. Bull. (2012), http://dx.doi.org/10.1016/j.marpolbu
most certainly related to acute disturbances that have affectedreefs in the southern Persian Gulf and Sea of Oman (e.g., Baumanet al., 2010; Riegl et al., 2011). At Fujairah, for example, therewas comprehensive mortality of Acropora and Pocillopora in late2008, caused by a harmful algal bloom (HAB). Subsequent surveys,revealed a 71.5% decline in biomass of reef fishes, attributable toboth direct effects of the HAB on fishes as well as subsequent deg-radation of benthic habitats (Bauman et al., 2010). In the southernGulf, extremely high temperatures in 2010 resulted in extensivebleaching (Riegl et al., 2011). Most corals did recover, but subse-quent outbreaks of coral disease resulted in significant (up to90%) mortality among Acropora corals at some locations (Rieglet al., 2011) further reducing the abundance of Acropora in thesouthern Gulf.
There are many studies showing strong links between butterfly-fish abundance and coral cover (Pratchett and Berumen, 2008). Inparticular, abundance of butterflyfishes often declines in followingextensive coral depletion (Wilson et al., 2006; Pratchett et al.,2008a). Perhaps, even more importantly, significant changes inthe structure of coral assemblages can exert marked changes inthe structure of fish assemblages, even without any change in over-all coral cover (Berumen and Pratchett, 2006), due to the dispro-portionate influence of key coral taxa, such as Acropora.Sustained and ongoing changes in the structure of coral assem-blages within the Persian Gulf and Gulf of Oman, are likely there-fore, to lead to even more pronounced changes in the abundance,diversity and functional composition of reef fishes. Furthermore,these changes may be repeated at many other locations through-out the world, due to global climate change and increasing anthro-pogenic pressures on coral reefs.
4.3. Conclusions
This study revealed very low abundance and diversity of butter-flyfishes within the southern Persian Gulf and Gulf of Oman. Thelimited age and isolation of the Persian Gulf (Sheppard et al.,2010) is likely to have constrained both evolution and colonisationby reef-associated fishes. However, coral-feeding butterflyfisheswere rare from the Gulf of Oman to the Persian Gulf. The one spe-cies of butterflyfish that was prevalent across all locations (C.nigropunctatus) was found to be a facultative coral feeder, and onlyweakly reliant on live coral for food. This is not surprising given thelimited availability of Acropora and Pocillopora corals, which are themost preferred by most coral-feeding butterflyfishes (Pratchett,2005). Elsewhere in the Indo-Pacific, butterflyfish assemblagestend to be dominated by highly specialised, obligate coral feedingspecies (e.g., Emslie et al., 2010). The low diversity and uniquefunctional composition of butterflyfishes in the southern PersianGulf, likely reflects its peripheral location, but abundance and com-position of butterflyfishes has been further affected by recent coraldepletion. Extreme environmental conditions (most notably, ex-treme temperatures) within the Persian Gulf may also exceedphysiological tolerances of most fishes (Feary et al., 2010). If so,it will be interesting to assess physiological adaptations that en-able C. nigropunctatus to cope with these conditions, whereas mostother butterflyfishes cannot. Factors that influence the differentialresilience of fishes to both habitat degradation and increasing tem-peratures are critical for predicting likely changes in the functionalcomposition of reef fish assemblages due to ongoing global climatechange that will affect reefs throughout the world.
Acknowledgements
This research was funded by New York University Abu DhabiInstitute. M. Pratchett, A. Baird and A. Bauman were supportedby fellowships from the ARC Centre of Excellence for Coral Reef
of Chaetodon butterflyfishes at a peripheral and extreme coral reef location,l.2012.10.014

8 M.S. Pratchett et al. / Marine Pollution Bulletin xxx (2012) xxx–xxx
Studies, D. Feary was supported by a Chancellors Postdoctoral Fel-lowship within the University of Technology, Sydney, and A. Hoeyheld a fellowship from King Abdulla University of Science andTechnology (KAUST). Initial surveys were conducted under theauspices of the Nakheel-UNU-INWEH joint project ‘‘Strategic Man-agement of Marine Ecosystems in Nakheel Projects’’ as part ofbaseline data collection. Authors are grateful for constructive feed-back provided by A. Cole and anonymous reviewers.
References
Allen, G.R., Steene, R., Allen, M., 1998. A guide to Angelfishes and Butterflyfishes.Odyssey, Perth.
Baird, A.H., Campbell, S., Anggoro, A.W., Fadli, N., Herdiana, Y., Katawijaya, T.,Legawa, R., Mahyiddin, D., Mukminin, A., Pratchett, M.S., Rudi, E., Siregar, A.M.,Trilestari, S., 2005. Achenese reefs in the wake of the Tsunami. Curr. Biol. 15,1926–1930.
Bauman, A.G., Burt, J.A., Feary, D.A., Marquis, E., Usseglio, P., 2010. Tropical harmfulalgal blooms: an emerging threat to coral reef communities? Mar. Pollut. Bull.60, 2117–2122.
Bauman, A.G., Feary, D.A., Heron, S.F., Pratchett, M.S., Burt, J.A., 2012. Multipleenvironmental factors influence the spatial distribution and structure of reefcommunities in the northeastern Arabian Peninsula. Mar. Poll. Bull., in press.
Bellwood, D.R., Hughes, T.P., 2001. Regional-scale assembly rules and biodiversity ofcoral reefs. Science 292, 1532–1535.
Bellwood, D.R., Hoey, A.S., Hughes, T.P., 2012. Human activity selectively impactsthe ecosystem roles of parrotfishes on coral reefs. Proc. Roy. Soc. B. 279, 1621–1629.
Bellwood, D.R., Klanten, S., Pratchett, M.S., Konow, N., van Herwerden, L., 2009.Evolutionary history of the butterflyfishes (F. Chaetodontidae) and the rise ofcoral feeding fishes. J. Evolution. Biol. 23, 335–349.
Bellwood, D.R., Wainwright, P.C., Fulton, C.J., Hoey, A.S., 2002. Assembly rules andfunctional groups at global biogeographical scales. Funct. Ecol. 16, 557–562.
Berumen, M.L., Pratchett, M.S., 2006. Recovery without resilience. Persistentdisturbance and long-term shifts in the structure of fish and coralcommunities at Tiahura Reef, Moorea. Coral Reefs 25, 647–653.
Berumen, M.L., Pratchett, M.S., McCormick, M.I., 2005. Within reef variation in thediet and condition of two coral feeding butterflyfish (Pices: Chaetodontidae).Mar. Ecol. Prog. Ser. 287, 217–227.
Birkeland, C., Neudecker, S., 1981. Foraging behaviour of two CaribbeanChaetodontidae: Chaetodon cpistratus and C. aculeatus. Copeia 1981, 169–178.
Blum, S.D., 1989. Biogeography of the Chaetodonitidae: an analysis of allopatryamong closely related species. Environ. Biol. Fish. 25, 9–31.
Bouchon-Navaro, Y., 1980. Quantitative distribution of the Chaetodontidae on afringing reef of the Jordanian coast (Gulf of Aqaba – Red Sea). Tethys 9, 247–251.
Burt, J., Al-Harthi, S., Al-Cibahy, A., 2011a. Long-term impacts of bleaching events onthe world’s warmest reefs. Mar. Environ. Res. 72, 225–229.
Burt, J., Bartholomew, A., Usseglio, P., 2008. Are artificial reefs surrogates of naturalhabitats for corals and fish in Dubai, United Arab Emirates? Coral Reefs 28, 663–675.
Burt, J., Bartholomew, A., Usseglio, P., Bauman, A., Sale, P.F., 2009. Recovery of coralsa decade after a bleaching event in Dubai, United Arab Emirates. Mar. Biol. 154,27–36.
Burt, J., Feary, D., Bauman, A., Usseglio, P., Cavalcante, G., Sale, P., 2011b.Biogeographic patterns of reef fish community structure in the northeasternArabian Peninsula. ICES J. Mar. Sci. 68, 1875–1883.
Cole, A.J., Lawton, R.J., Wilson, S.K., Pratchett, M.S., 2012. Consumption of tabularacroporid corals by reef fishes: a comparison with plant–herbivore interactions.Funct. Ecol. 26, 307–316.
Cole, A.J., Pratchett, M.S., Jones, G.P., 2008. Diversity and functional importance ofcoral-feeding fishes on tropical coral reefs. Fish Fish. 9, 286–307.
Coles, S.L., 2003. Coral species diversity and environmental factors in the ArabianGulf and the Gulf of Oman: a comparison to the Indo–Pacific region. Atoll Res.Bull. 507 (2003), 1–19.
Connell, J.H., 1973. Population ecology of reef building corals. In: Jones, O.A.,Endean, R. (Eds.), Biology and Geology of Coral Reefs, II, Biology 1. AcademicPress, NY and, London, pp. 205–245.
Cowman, P.F., Bellwood, D.R., 2011. Coral reefs as drivers of cladogenesis:expanding coral reefs, cryptic extinction events, and the development ofbiodiversity hotspots. J. Evolution. Biol. 24, 2543–2562.
Cox, E.F., 1994. Resource use by corallivorous butterflyfishes (FamilyChaetodontidae) in Hawaii. Bull. Mar. Sci. 54 (2), 535–545.
Emslie, M.J., Pratchett, M.S., Cheal, A.J., Osborne, K., 2010. Great Barrier Reefbutterflyfish community structure: the role of shelf position and benthiccommunity type. Coral reefs 29, 705–715.
Fabricius, K., De’ath, G., McCook, L., Turak, E., Williams, D.Mc.B., 2005. Changes inalgal, coral and fish assemblages along water quality gradients on the inshoreGreat Barrier Reef. Mar. Pollut. Bull. 51, 384–398.
Feary, D.A., Burt, J.A., Bauman, A.G., Usseglio, P., Sales, P.F., Cavalcante, G.H., 2010.Fish communities on the world’s warmest reefs: what can they tell us about theeffects of climate change in the future? J. Fish Biol. 77, 1931–1947.
Please cite this article in press as: Pratchett, M.S., et al. Functional compositionthe Persian Gulf. Mar. Pollut. Bull. (2012), http://dx.doi.org/10.1016/j.marpolbu
Graham, N.A.J., 2007. Ecological versatility and the decline of coral feeding fishesfollowing climate driven coral mortality. Mar. Biol. 153, 127–199.
Glynn, P.W., 2004. High complexity food webs in low-diversity Eastern Pacific reef-coral communities. Ecosystems 7, 358–367.
Harmelin-Vivien, M., 1989. Implications of feeding specialization on therecruitment processes and community structure of butterflyfishes. Environ.Biol. Fish. 25, 101–110.
Harmelin-Vivien, M.L., Bouchon-Navaro, Y., 1983. Feeding diets and significance ofcoral feeding among chaetodontid fishes in Moorea (French Polynesia). CoralReefs 2, 119–127.
Hoey, A.S., Bellwood, D.R., 2008. Cross-shelf variation in the role of parrotfishes onthe Great Barrier Reef. Coral Reefs 27, 37–47.
Hoegh-Guldberg, O., Bruno, J.F., 2010. The impact of climate change on the world’smarine ecosystems. Science 328, 1523–1528.
Hoegh-Guldberg, O., Mumby, P.J., Hooten, A.J., Steneck, R.S., Greenfield, P., Gomez,E., Harvell, C.D., Sale, P.F., Edwards, A.J., Caldeira, K., Knowlton, N., Eakin, C.M.,Inglesias-Prieto, R., Muthiga, N., Bradbury, R.H., Dubi, A., Hatziolos, M.E., 2007.Coral reefs under rapid climate change and ocean acidification. Science 318,1737–1742.
Hughes, T.P., Baird, A.H., Bellwood, D.R., Card, M., Connolly, S.R., Folke, C., Grosberg,R., Hoegh-Guldberg, O., Jackson, J.B.C., Kleypas, J., Lough, J.M., Marshall, P.,Nystrom, M., Palumbi, S.R., Pandolfi, J.M., Rosen, B., Roughgarden, J., 2003.Climate change, human impacts and the resilience of coral reefs. Science 301,929–933.
Irons, D.K., 1989. Temporal and areal feeding behaviour of the butterflyfish,Chaetodon trifascialis, at Johnston Atoll. Environ. Biol. Fish. 25, 187–193.
Kohler, K.E., Gill, S.M., 2006. Coral point count with excel extensions (CPCe): a visualbasic program for the determination of coral and substrate coverage usingrandom point count methodology. Comput. Geosci. 32, 1259–1269.
Labiosa, R.G., Arrigo, K.R., Genin, A., Monismith, S.G., van Dijken, G., 2003. Theinterplay between upwelling and deep convective mixing in determining theseasonal phytoplankton dynamics in the Gulf of Aqaba: evidence from SeaWiFSand MODIS. Limnol. Oceanogr. 48, 2355–2368.
Laurance, W.F., 1999. Reflections on the tropical deforestation crisis. Biol. Conserv.91, 109–117.
Lieske, E., Myers, R., 1996. Collins Pocket Guide. Coral Reef Fishes. Indo–Pacific &Caribbean Including the Red Sea. Harper Collins Publishers.
Manly, B.F.J., McDonald, L.L., Thomas, D.L., 1993. Resource Selection by Animals.Chapman and Hall, London.
McClanahan, T.R., Ateweberhan, M., Graham, N.A.J., Wilson, S.K., Sebastian, C.R.,Guillaume, M.M.M., Bruggeman, J.H., 2007. Western Indian Ocean coralcommunities, bleaching responses and susceptibility to extinction. Mar. Ecol.Prog. Ser. 337, 1–13.
Michael, S.W., 2004. Angelfishes and Butterflyfishes: Reef Fishes Series. T.F.H.Publications, Neptune City.
Naglerkerken, I., van der Velde, G., Wartenbergh, S.L.J., Nugues, M.M., Pratchett,M.S., 2009. Cryptic dietary components reduce dietary overlap amongsympatric butterflyfishes (Chaetdontidae). J. Fish Biol. 75, 1123–1143.
Pratchett, M.S., 2005. Dietary overlap among coral-feeding butterflyfishes(Chaetodontidae) at Lizard Island, northern Great Barrier Reef. Mar. Biol. 148,373–382.
Pratchett, M.S., 2007. Dietary selection by coral-feeding butterflyfishes(Chaetodontidae) on the Great Barrier Reef Australia. Raffles Bull. Zool. S14,155–160.
Pratchett, M.S., Wilson, S.K., Berumen, M.L., McCormick, M.I., 2004. Sub-lethaleffects of coral bleaching on an obligate coral feeding butterflyfish. Coral Reefs23, 352–356.
Pratchett, M.S., Berumen, M.L., 2008. Inter-specific variation in distributions anddiets of coral reef butterflyfishes (Teleostei: Chaetodontidae). J. Fish Biol. 73,1730–1747.
Pratchett, M.S., Marnane, M.J., Berumen, M.L., Eagle, J.E., Pratchett, D.J., 2008a.Habitat associations of juvenile versus adult butterflyfishes. Coral Reefs 27,541–551.
Pratchett, M.S., Munday, P.L., Wilson, S.K., Graham, N.A.J., Cinner, J.E., Bellwood, D.R.,Jones, G.P., Polunin, N.V.C., McClanahan, T.R., 2008b. Effects of climate-inducedcoral bleaching on coral-reef fishes: ecological and economic consequences.Oceanogr. Mar. Biol. Annu. Rev. 46, 251–296.
Pratchett, M.S., Wilson, S.K., Baird, A.H., 2006. Declines in the abundance ofChaetodon butterflyfishes (Chaetodontidae) following extensive coral depletion.J. Fish Biol. 69, 1269–1280.
Pratchett, M.S., Graham, N.A.J., Cole, A.J., (in press). Specialist corallivores dominatebutterflyfish assemblages in coral-dominated reef habitats. J. Fish Biol.
Reese, E.S., 1977. Coevolution of corals and coral feeding fishes of the familyChaetodonitidae. In: Proceedings of the 3rd International Coral Reef Symposium2, pp. 267–274.
Reese, E.S., 1981. Predation on corals by fishes of the family Chaetodontidae:implications for conservation and management of coral reef ecosystems. Bull.Mar. Sci. 31, 594–604.
Righton, D., Kemp, J., Ormond, R., 1996. Biogeography, community structure anddiversity of Red Sea and western Indian Ocean butterflyfishes. J. Mar. Biol.Assoc. UK 76, 223–228.
Riegl, B., 1999. Corals in a non-reef setting in the southern Arabian Gulf (Dubai,UAE): fauna and community structure in response to recurring mass mortality.Coral Reefs 18, 63–73.
Riegl, B., 2003. Climate change and coral reefs: different effects in two high-latitudeareas (Arabian Gulf, South Africa). Coral Reefs 22, 433–446.
of Chaetodon butterflyfishes at a peripheral and extreme coral reef location,l.2012.10.014

M.S. Pratchett et al. / Marine Pollution Bulletin xxx (2012) xxx–xxx 9
Riegl, B.M., Purkis, S.J., Al-Cibahy, A.S., Abdel-Moati, M.A., Hoegh-Guldberg, O., 2011.Present limits to heat-adaptability in corals and population-level responses toclimate extremes. PLoS One 6, e24802.
Rotjan, R.D., Lewis, S.M., 2008. Impact of coral predators on tropical reefs. Mar. Ecol.Prog. Ser. 367, 73–91.
Sale, P.F., Feary, D.A., Burt, J.A., Bauman, A.G., Cavalcante, G.H., Drouillard, K.G.,Kjerfve, B., Marquis, E., Trick, C.G., Usseglio, P., Van Lavieren, H., 2011. Thegrowing need for sustainable ecological management of marine communities ofthe Persian Gulf. Ambio 40, 4–17.
Sheppard, C.R.C., Al-Husiani, M., Al-Jamali, F., Al-Yamani, F., Baldwin, R., Bishop, J.,Benzoni, F., Dutrieux, E., Dulvy, N.K., Durvasula, S.R.V., Jones, D.A., Loughland, R.,Medio, R., Nithyanandan, M., Pilling, G.M., Polikarpov, I., Price, A.R.G., Purkis, S.,Riegl, B., Saburova, M., Samimi Namin, K., Taylor, O., Wilson, S., Zainal, K., 2010.The Gulf: a young sea in decline. Mar. Pollut. Bull. 60, 13–38.
Sheppard, C.R.C., Price, A.R.G., Roberts, C., 1992. Marine ecology of the Arabianregion: Patterns and processes in extreme tropical environments. AcademicPress, London.
Shokri, M.R., Fatemi, S.M.R., Crosby, M.P., 2005. The status of butterflyfishes(Chaetodontidae) in the northern Persian Gulf, I.R. Iran. Aquat. Conserv.: Mar.Freshwat. Ecosyst. 15, S91–S99.
Please cite this article in press as: Pratchett, M.S., et al. Functional compositionthe Persian Gulf. Mar. Pollut. Bull. (2012), http://dx.doi.org/10.1016/j.marpolbu
Stella, J.S., Pratchett, M.S., Hutchings, P.A., Jones, G.P., 2011. Diversity, importanceand vulnerability of coral-associated invertebrates. Oceanogr. Mar. Biol. Ann.Rev. 49, 43–104.
Tricas, T.C., 1989. Prey selection by coral-feeding butterflyfishes: strategies tomaximise the profit. Environ. Biol. Fish. 25, 175–185.
Walther, G.R., Post, E., Convey, P., Menze, A., Parmesan, C., Beebee, T.J.C., Fromentin,J.M., Hoegh-Guldberg, O., Bairlein, F., 2002. Ecological responses to recentclimate change. Nature 416, 389–395.
Wilkinson, C.R., 2004. Status of Coral Reefs of the World: 2004. Australian Instituteof Marine Science, Townsville.
Williams, D.Mc.B., 1986. Temporal variation in the structure of reef slope fishcommunities (central Great Barrier Reef): short-term effects of Acanthasterplanci infestation. Mar. Ecol. Prog. Ser. 28, 157–164.
Wilson, S.K., Graham, N.A.J., Pratchett, M.S., Jones, G.P., Polunin, N.V.C., 2006.Multiple disturbances and the global degradation of coral reefs: are reef fishesat risk or resilient? Glob. Change Biol. 12, 2220–2234.
Zekeria, Z.A., Afeworki, Y., Videler, J.J., 2005. The distribution patterns of Red SeaChaetodontidae assemblages. Aquat. Conserv.: Mar. Freshwat. Ecosyst. 15, S71–S76.
of Chaetodon butterflyfishes at a peripheral and extreme coral reef location,l.2012.10.014
























![STEP7_VOL1 [Persian]](https://img.dokumen.tips/doc/110x75/63211d438a1d893baa0ce9de/step7vol1-persian.jpg)




