Embed Size (px)
Citation preview

Functional characterization of the Xcs and Xps type IIsecretion systems from the plant pathogenic bacteriumXanthomonas campestris pv vesicatoria
Robert Szczesny*, Matthias Jordan*, Claudia Schramm, Steve Schulz, Virginie Cogez, Ulla Bonas and
Daniela Buttner
Institut fur Biologie, Bereich Genetik, Martin-Luther-Universitat Halle-Wittenberg, D–06099 Halle (Saale), Germany
Author for correspondence:Daniela Buttner
Tel: +49 345 5526293Email: [email protected]
halle.de
Received: 10 March 2010
Accepted: 22 April 2010
New Phytologist (2010) 187: 983–1002doi: 10.1111/j.1469-8137.2010.03312.x
Key words: amylase, bacterial spot disease,cellulase, HrpG, pepper, protease, tomato,type II secretion.
Summary
• Type II secretion (T2S) systems of many plant-pathogenic bacteria often secrete
cell wall-degrading enzymes into the plant apoplast.
• Here, we show that the Xps-T2S system from the plant pathogen Xanthomonas
campestris pv vesicatoria (Xcv) promotes disease and contributes to the trans-
location of effector proteins that are delivered into the plant cell by the type III
secretion (T3S) system.
• The Xcs-T2S system instead lacks an obvious virulence function. However,
individual xcs genes can partially complement mutants in homologous xps genes,
indicating that they encode functional components of T2S systems. Enzyme
activity assays showed that the Xps system contributes to secretion of proteases
and xylanases. We identified the virulence-associated xylanase XynC as a substrate
of the Xps system. However, homologs of known T2S substrates from other
Xanthomonas spp. are not secreted by the T2S systems from Xcv. Thus, T2S
systems from Xanthomonas spp. appear to differ significantly in their substrate
specificities.
• Transcript analyses revealed that expression of xps genes in Xcv is activated by
HrpG and HrpX, key regulators of the T3S system. By contrast, expression of xynC
and extracellular protease and xylanase activities are repressed by HrpG and HrpX,
suggesting that components and substrates of the Xps system are differentially
regulated.
Introduction
Virulence of Gram-negative plant pathogenic bacteria ofthe genus Xanthomonas often depends on protein secretionsystems that play a role in, for example bacterial adhesion,degradation of the plant cell wall, acquisition of nutrientsand suppression of plant defense (Preston et al., 2005;Buttner & Bonas, 2010). Targeted and random mutagenesisapproaches revealed a virulence function of several proteinsecretion systems from Xanthomonas species (spp.) includ-ing type II, type III, type IV and type V secretion systemsand ⁄ or their cognate substrates (Bonas et al., 1991; Ray
et al., 2000; Yang et al., 2000; Qian et al., 2005; He et al.,2007; Wang et al., 2008a; Das et al., 2009).
In our laboratory, we study virulence factors fromXanthomonas campestris pv vesicatoria (Xcv, also designatedXanthomonas euvesicatoria or Xanthomonas axonopodis pvvesicatoria; Jones et al., 2004), the causal agent of bacterialspot disease in pepper and tomato. The type III secretion(T3S) system is essential for colonization of the plant andtranslocates bacterial effector proteins directly into the plantcell cytosol where they interfere with cellular functions suchas defense responses to the benefit of the pathogen (Ghosh,2004; Gurlebeck et al., 2006; Zhou & Chai, 2008).Translocation of effector proteins into plant cells dependson extracellular Hrp (hypersensitive response and patho-genicity) pili that are associated with the membrane-spanning*These authors contributed equally to the work.
NewPhytologist Research
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002 983www.newphytologist.com

T3S apparatus and serve as protein transport channelsacross the plant cell wall (Buttner & Bonas, 2002; Ghosh,2004). Components of the T3S system are encoded by thechromosomal hrp gene cluster, which is specifically acti-vated when the bacteria enter the plant or are cultivated incertain minimal media (Buttner & Bonas, 2002).Expression of the hrp genes depends on the response regulatorHrpG, which controls – in most cases via the transcriptionalactivator HrpX – the expression of a genome-wide regulon,including effector genes and genes that encode predictedextracellular degradative enzymes (Wengelnik & Bonas,1996; Wengelnik et al., 1996b; Noel et al., 2001).
The contribution of the type II secretion (T2S) system tovirulence of Xcv has not yet been studied. Generally, T2Ssystems secrete toxins and degradative enzymes that aretransported across the inner bacterial membrane by the Secor TAT (twin-arginine translocation) system and often contri-bute to virulence of plant pathogenic bacteria (Sandkvist,2001; Cianciotto, 2005; Jha et al., 2005). Protein transportacross the outer bacterial membrane is mediated by the T2Sapparatus that consists of 12–15 components, most ofwhich are associated with the inner membrane or form aperiplasmic pseudopilus (Sandkvist, 2001). The continuousassembly and disassembly of the pseudopilus presumablypushes T2S substrates through the outer membrane secretin(Sandkvist, 2001; Cianciotto, 2005). Type II-dependentprotein transport is probably energized by a cytoplasmicATPase associated with the secretion apparatus at the innermembrane (Chen et al., 2005; Shiue et al., 2006, 2007).
Comparative genome sequence analyses of Xanthomonasspp. revealed that Xcv, X. campestris pv campestris (Xcc) andXanthomonas axonopodis pv citri (Xac) encode componentsof two different T2S systems, designated Xps and Xcs. Bycontrast, Xanthomonas oryzae pv oryzae (Xoo) and X. oryzaepv oryzicola (Xoc) contain only the Xps system (Lu et al.,2008). A virulence function has been reported for the Xpssystems from Xcc and Xoo (Dow et al., 1987; Ray et al.,2000; Sun et al., 2005). It is assumed that T2S substratesdegrade components of the plant cell wall, which is a physicalbarrier against invading microbes. In good agreement withtheir presumed role in cell wall degradation, type II-secretedcellulases, polygalacturonases and xylanases from Xcc and Xoowere shown to contribute to bacterial virulence (Rajeshwariet al., 2005; Hu et al., 2007; Jha et al., 2007; Wang et al.,2008a,b). Interestingly, experimental evidence suggests thatplant cell wall-degrading enzymes not only promote bacterialinvasion of the plant tissue but also activate defense responsesthat counteract microbial invasion (Ryan & Farmer, 1991;Liu et al., 2005; Chisholm et al., 2006; Jones & Dangl,2006; Bittel & Robatzek, 2007; Jha et al., 2007). Duringnatural infections, however, basal plant defense is suppressedby bacterial effector proteins that are translocated into theplant cell by the T3S system, suggesting a functional inter-play between T2S and T3S systems (Keshavarzi et al., 2004;
Metz et al., 2005; Jha et al., 2007; Zhou & Chai, 2008;Bartetzko et al., 2009). Notably, several T2S componentsand substrates from Xanthomonas spp. were shown to be co-regulated with T3S genes (Furutani et al., 2004; Wang et al.,2008b; Yamazaki et al., 2008).
In this study, we show that the Xps system from Xcvcontributes to bacterial virulence and type III effector proteintranslocation. No obvious role in virulence was observed forthe Xcs system, however, xcs genes can partly compensate lossof homologous xps genes. The virulence function of the Xpssystem presumably results from the secretion of extracellularproteases and xylanases. We identified the virulence-associatedxylanase XynC as a substrate of the Xps system and provideexperimental evidence that components and substrates of theXps system are differentially regulated.
Materials and Methods
Bacterial strains and growth conditions
Bacterial strains and plasmids used in this study are listed inTable 1. Escherichia coli cells were grown at 37�C in lyso-geny broth (LB) or Super medium (Qiagen), Xcv strainswere cultivated at 30�C in nutrient–yeast–glycerol (NYG)medium (Daniels et al., 1984) or in minimal medium A(Ausubel et al., 1996), which was supplemented withsucrose (10 mM) and casamino acids (0.3%). Plasmidswere introduced into E. coli by electroporation and into Xcvby conjugation, using pRK2013 as helper plasmid in tri-parental matings (Figurski & Helinski, 1979). Antibioticswere added to the media at the following final concen-trations: ampicillin, 100 lg ml)1; gentamycin 15 lg ml)1;kanamycin, 25 lg ml)1; rifampicin, 100 lg ml)1; spectino-mycin, 100 lg ml)1; tetracycline, 10 lg ml)1.
Plant material and plant inoculations
The near-isogenic pepper lines ‘Early Cal Wonder’ (ECW),ECW-10R and ECW-30R (Minsavage et al., 1990) weregrown at 25�C with 60–70% relative humidity and 16 hlight. Xcv strains were hand-inoculated with a needlelesssyringe into the intercellular spaces of pepper leaves atconcentrations of 2 · 108 CFU ml)1 in 1 mM MgCl2,unless stated otherwise. Disease symptoms and the hyper-sensitive response (HR) were scored over a period of2–10 d post inoculation (dpi). For better visualization ofthe HR, leaves were bleached in 70% ethanol. In plantagrowth curves were performed as described (Bonas et al.,1991). All experiments were repeated at least two times.
Enzyme activity assays
For the analysis of extracellular protease and xylanase activity,Xcv strains were adjusted to a density of 109 CFU ml)1 and
984 Research
NewPhytologist
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

Table 1 Bacterial strains and plasmids used in this study
Relevant characteristics Reference or source
Xcv
85-10 Pepper-race 2; wild type; Rifr Canteros (1990)85-10DhrcN hrcN deletion mutant of strain 85-10 Lorenz & Buttner (2009)85-10DhrcC hrcC deletion mutant of strain 85-10 Weber et al. (2005)85* 85-10 derivative containing the hrpG* mutation Wengelnik et al. (1999)85*DhrpX hrpX deletion derivative of strain 85* Noel et al. (2001)85*DhrpF hrpF deletion mutant of strain 85* Buttner et al. (2002)85-10Dxcs Deletion mutant of strain 85-10 lacking the complete xcs gene cluster This study85-10DxcsE xcsE deletion mutant of strain 85-10;
deleted in codons 17–482 (out of 496) of xcsE
This study
85-10DxcsD xcsD deletion mutant of strain 85-10;deleted in codons 166–376 (out of 690) from xcsD
This study
85-10DxpsE xpsE deletion mutant of strain 85-10;lacks codons 66-547 (out of 576) of xpsE
This study
85-10DxpsD xpsD deletion mutant of strain 85-10; lacks codons 196–384(out of 760) from xpsD and contains an additional frameshift
This study
85-10D4312 XCV4312 deletion mutant of strain 85-10;lacks codons 14-593 (out of 605) of XCV4312
This study
85-10DxynC xynC deletion mutant of strain 85-10; lacks codons 77-298(out of 407) of xynC and contains an additional frameshift
This study
85-10DxpsEDxcsE xpsE ⁄ xcsE double deletion mutant of strain 85-10 This study85-10DxpsEDxcsED4312
xpsE ⁄ xcsE ⁄ XCV4312 triple deletion mutant of strain 85-10 This study
85-10DxcsED4312 xcsE ⁄ XCV4312 double deletion mutant of strain 85-10 This study85-10DxpsED4312 xpsE ⁄ XCV4312 double deletion mutant of strain 85-10 This study85-10DxpsEDhrcN xpsE ⁄ hrcN double deletion mutant of strain 85-10 This study85-10DxpsDDxcsD xpsD ⁄ xcsD double deletion mutant of strain 85-10 This study85-10DxpsDDxcsE xpsD ⁄ xcsE double deletion mutant of strain 85-10 This study85-10DxpsDDxcsDDhrcC
xpsD ⁄ xcsD ⁄ hrcC triple deletion mutant of strain 85-10 This study
E. coli
DH5a F) recA hsdR17(rk),mk
+) F80dlacZ DM15 Bethesda ResearchLaboratories,Bethesda, MD, USA
DH5a kpir F) recA hsdR17(rk),mk
+) F80dlacZ DM15 [kpir] Menard et al. (1993)Plasmids
pBlueskript(II) KS Phagemid, pUC derivative; Apr StratagenepBBR1MCS-5 Broad-host-range vector; lac promoter; Gmr Kovach et al. (1995)pBBRxpsD pBBR1MCS-5 derivative encoding XpsD This studypBRM Golden Gate-compatible derivative of pBBR1MCS-5 with lac promoter This studypBRM-P Golden Gate-compatible derivative of pBBR1MCS-5 without lac promoter This studypBRM0031 pBRM derivative encoding XCV0031-c-Myc This studypBRM0959 pBRM derivative encoding XCV0959-c-Myc This studypBRM0960 pBRM derivative encoding XCV0960-c-Myc This studypBRM1823 pBRM derivative encoding XCV1823-c-Myc This studypBRM2049 pBRM derivative encoding XCV2049-c-Myc This studypBRM2571 pBRM derivative encoding XCV2571-c-Myc This studypBRM2918 pBRM derivative encoding XCV2918-c-Myc This studypBRM3013 pBRM derivative encoding XCV3013-c-Myc This studypBRM4074 pBRM derivative encoding XCV4074-c-Myc This studypBRMXc0705 pBRM derivative encoding Xc0705-c-Myc This studypBRMXc1849 pBRM derivative encoding Xc1849-c-Myc This studypBRMhrcN pBRM derivative encoding HrcN-c-Myc This studypDGW4M Derivative of pDSK602 containing attR1-CmR-ccdB-attR2
upstream of 4 · c-Myc epitope-encoding sequenceLorenz et al. (2008)
pDGW3669 pDGW4M derivative encoding XCV3669-c-Myc This studypDGW0722 pDGW4M derivative encoding XCV0722-c-Myc This studypDGW0965 pDGW4M derivative encoding XynC-c-Myc This studypDGW2569 pDGW4M derivative encoding XCV2569-c-Myc This studypDSK602 Broad-host-range vector; contains triple lacUV5 promoter; Smr Murillo et al. (1994)
NewPhytologist Research 985
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

incubated on NYG agar plates containing 1% skimmedmilk (AppliChem) and 0.1% remazol brilliant blue (RBB)xylan (Sigma-Aldrich), respectively (Vroemen et al., 1995).For detection of cellulase and amylase activities, we usedplates containing 1% carboxymethyl cellulose (CMC;Sigma-Aldrich) and 1% starch (AppliChem), respectively.The CMC plates were stained with 0.2% Congo red(Sigma-Aldrich) solution and destained with 0.5 M NaCl(Gough et al., 1988). Starch plates were stained withLugol’s solution (0.4% potassium iodide : 0.2% iodinesolution) and destained with 98% ethanol–acetone (1 : 1)(Hu et al., 1995). Bacteria were inoculated into holes thatwere punched out of the agar. Plates were incubated at 30�Cfor 1–2 d and bacteria were removed before documentation.All experiments were repeated at least two times.
Generation of deletion constructs
To delete xcsD, xpsD, xcsE, xpsE, XCV4312 and xynC,respectively, flanking regions were amplified by PCR usinggenomic DNA from Xcv strain 85-10 as template, and theamplicons were introduced into the suicide vector pOK1.Deletion constructs were introduced into the genome ofXcv by crossing-over events as described in Huguet et al.
(1998). Details on cloning strategies are available uponrequest. Primer sequences are listed in the Supporting Infor-mation, Table S1.
Generation of the Golden Gate-compatible expressionvectors pBRM and pBRM-P
The Golden Gate system allows efficient one step-cloningof PCR amplicons into a destination vector in a restriction–ligation reaction (Engler et al., 2008). The system is basedon type IIs restriction enzymes (e.g. BsaI) that cut DNAoutside of the enzyme’s recognition sites. The Golden Gate-compatible expression vectors pBRM and pBRM-P used inthis study are derived from pBBR1MCS-5 (see Table 1).To remove the internal BsaI site of pBBR1MCS-5, vectorDNA was amplified by PCR with primers pBBR1-for ⁄pBBR2-rev and pBBR3-for ⁄ pBBR4-rev and both fragmentswere religated in a one-step restriction–ligation reactionusing BsaI. The resulting vector pBBR1mod1 lacks theinternal BsaI site and contains a single EcoRI and a HindIIIsite that replace the polylinker. To generate pBRM, weamplified the lacZ alpha gene from pUC19 and the triple c-Myc epitope-encoding sequence from pC3003 with primerslacZ-for ⁄ lacZ-rev and myc-for ⁄ myc-rev, respectively, and
Table 1 (Continued)
Relevant characteristics Reference or source
pDSK604 Derivative of pDSK602 with modified polylinker Escolar et al. (2001)pDStrephrcN pDSK604 derivative encoding Strep-HrcN Lorenz & Buttner (2009)pDSMxpsE pDSK602 derivative encoding XpsE-c-Myc This studypDxpsE pDSK604 derivative encoding XpsE This studypDxopF1356 pDSK602 derivative encoding XopF11-200-AvrBs3D2 Buttner et al. (2006)pENTR ⁄ D TOPO Gateway system donor vector; Kmr InvitrogenpK18mobsac Suicide vector; oriVEc oriT sacB; Kmr Schafer et al. (1994)pK18Dxcs pK18mobsac derivative containing
flanking regions of the xcs gene clusterThis study
pK18DxpsE pK18mobsac derivative containing flanking regions of xpsE This studypLAFR6 RK2 replicon, Mob+ Tra); multicloning
site flanked by transcription terminators; TcrBonas et al. (1989)
pL6avrBs3356 pLAFR6 derivative encodingAvrBs31-200–AvrBs3D2 under control of the native promoter
Noel et al. (2003)
pL6xopC356 pLAFR6 derivative encoding XopC1-200–AvrBs3D2under control of the native promoter
Noel et al. (2003)
pL6xopJ356 pLAFR6 derivative encoding XopJ1-155–AvrBs3D2under control of the native promoter
Noel et al. (2003)
pOK1 Suicide vector; sacB sacQ mobRK2 oriR6K; Smr Huguet et al. (1998)pOKDxcsD pOK1 derivative containing flanking regions of xcsD This studypOKDxpsD pOK1 derivative containing flanking regions of xpsD This studypOKDxcsE pOK1 derivative containing flanking regions of xcsE This studypOKD0965 pOK1 derivative containing flanking regions of xynC This studypOKD4312 pOK1 derivative containing flanking regions of XCV4312 This studypOKDhrcC pOK1 derivative containing flanking regions of hrcC Weber et al. (2005)pRK2013 ColE1 replicon, TraRK+ Mob+; Kmr Figurski & Helinski (1979)pUC119 ColE1 replicon; Apr Vieira & Messing (1987)
Ap, ampicillin; Gm, gentamycin; Km, kanamycin; Rif, rifampicin; Sp, spectinomycin; Tc, tetracycline; r, resistant.
986 Research
NewPhytologist
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

cloned the corresponding fragments (digested with BpiI)into the EcoRI ⁄ HindIII sites of pBBR1mod, giving pBRM(Fig. S1). To generate pBRM-P, we amplified pBRM withprimers pBBR1-for and pBBR5-rev and ligated the PCRproduct in a one-step restriction–ligation reaction usingBsaI and ligase. The resulting vector pBBR1mod2 lacks thelac promoter and contains a single EcoRI and a HindIII sitethat replace the polylinker. We amplified the lac promoter,the lacZ alpha gene and the c-Myc epitope-encodingsequence from pBRM with primers lacZprom-for andmyc-Esp-rev. The corresponding PCR fragment andpBRM were digested with Esp3I and EcoRI ⁄ HindIII, respec-tively, and ligated, thus generating pBRM-P (see Fig. S1).
Generation of expression constructs
For the generation of expression constructs encodingcomponents or putative substrates of T2S systems, corre-sponding gene fragments were amplified by PCR from Xcvstrain 85-10, inserted into pENTR ⁄ D-TOPO and recom-bined into pDGW4M using GATEWAY technology(Invitrogen). Alternatively, PCR amplicons were clonedinto pBRM or pBRM-P in a one step restriction–ligationreaction as described in Engler et al. (2008). For fragmentswith an internal BsaI site, the reaction mix was incubatedwith ligase after heat inactivation for additional 20 min at37�C (Engler et al., 2008). All constructs are listed inTable 1.
Protein analysis and secretion experiments
For the analysis of bacterial protein accumulation in planta,bacteria were inoculated at a density of 109 CFU ml)1 intoleaves of susceptible ECW pepper plants. Proteins wereextracted 6 h post inoculation (hpi) from five leaf discs thatwere ground in 1 mM MgCl2. For in vitro T2S assays,bacteria were grown overnight in NYG medium, resus-pended in fresh NYG medium at a cell density of1.5 · 108 CFU ml)1 and incubated at 30�C for 2 h.Culture supernatants were separated from bacterial cells byfiltration and secreted proteins were precipitated by trichlo-roacetic acid as described in Rossier et al. (1999). Equalamounts of total bacterial cell extracts and culture super-natants were analysed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) andimmunoblotting. We used polyclonal antibodies specific forHrpF, AvrBs3, HrcJ and GroEL (Knoop et al., 1991;Rossier et al., 2000; Buttner et al., 2002; Stressgen), andmonoclonal anti-c-Myc (Roche) and anti-Strep antibodies(IBA GmbH). Horseradish peroxidase-labeled anti-rabbitand anti-mouse antibodies (Amersham Pharmacia Biotech)were used as secondary antibodies, and antibody reactionswere visualized by enhanced chemiluminescence (AmershamPharmacia Biotech).
RNA analysis
For transcript analyses via quantitative reverse-transcriptionPCR (qRT-PCR), bacteria were grown overnight in NYGmedium, adjusted to an optical density (OD, 600 nm) of0.2 and incubated for 4 h at 30�C. RNA was extractedusing a Trizol-based protocol and cDNA was synthesizedusing 4.5 lg RNA in a RevertAid H Minus First StrandcDNA-Synthesis Kit (Fermentas). xcsE, xpsE, xpsF and xopCtranscripts were amplified using primers xcsE_RT-for ⁄xcsE_RT-rev, xpsE_RT-for ⁄ xpsE_RT-rev, xpsF_RT-for ⁄xpsF_RT-rev and xopC_RT-for ⁄ xopC_RT-rev, respectively.The qRT-PCR was performed on an iCycler (Bio-Rad)using a SYBR Green-based PCR reaction mixture(ABsolute QPCR SYBR Green Fluorescein Mix; ABgeneLimited) and c. 9 ng of template cDNA. Details on PCRprofiles are available upon request. For comparative analysisof transcript levels of different genes, the efficiency of eachPCR reaction was determined using a dilution series oftemplate plotted into a standard curve. To ensure amplificationspecificity, amplicons were subjected to melting curveanalysis and analysed on 1.5% agarose gels. Transcript levelswere determined as technical triplicates and transcript levelsof constitutively expressed 16S rRNA (primers: 16S_RT-for ⁄ 16S_RT-rev) were used to account for differences incDNA amounts as described in the ABI user bulletin 2(Applied Biosystems).
Results
xcs and xps genes from Xcv strain 85-10 are expressedin vitro
Genome sequence analyses of Xcv strain 85-10 revealed thepresence of chromosomal xcs and xps gene clusters containing12 (xcsC to xcsN) and 11 (xpsD to xpsN) genes, respectively(Fig. 1a; Thieme et al., 2005). To investigate expression ofxcs and xps genes, we performed RT-PCR analysis withcDNA derived from Xcv strain 85-10 grown in complexNYG medium. Using primers specific for the predictedATPase-encoding genes xcsE and xpsE, cDNAs correspond-ing to fragments of both genes were amplified, suggestingthat xcsE and xpsE are expressed (Fig. 1b). This wasconfirmed by qRT-PCR analysis, which revealed that xpsEtranscript levels were c. 76-fold higher than xcsE levels (datanot shown).
The predicted secretin gene xpsD contributes tobacterial virulence
To investigate a possible contribution of the Xcs and Xpssystems to virulence of Xcv strain 85-10, we deleted xcsDand xpsD, respectively, which encode the predicted outermembrane secretins of the two T2S systems. As expected,
NewPhytologist Research 987
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com
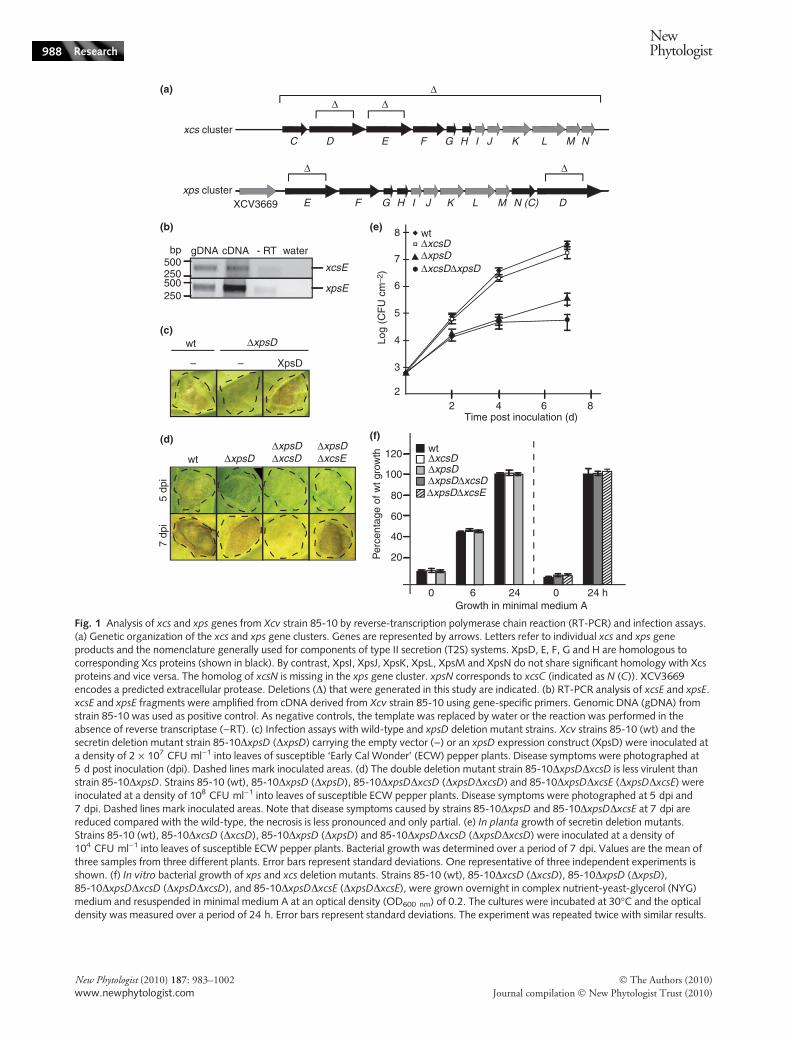
wt ΔxpsDΔxpsDΔxcsD
ΔxpsDΔxcsE
wt ΔxpsD
– XpsD–
22 4 6 8
wtΔxcsDΔxpsD
Time post inoculation (d)
Log
(CF
U c
m–2
)
8
7
6
5
4
3
ΔxcsDΔxpsD
C D E F G H I J K L M N
Δ ΔΔ
E DF G H I J K L M N (C)XCV3669
Δ Δ
xcs cluster
xps cluster
(a)
(b) (e)
(f)
(c)
(d)
5 dp
i7
dpi
gDNA cDNA - RT water
xcsE
xpsE
bp500250500250
0 6 24 0 24 h
20
40
60
80
100
120
Per
cent
age
of w
t gro
wth
wtΔxcsD
Growth in minimal medium A
ΔxpsDΔxpsDΔxcsDΔxpsDΔxcsE
Fig. 1 Analysis of xcs and xps genes from Xcv strain 85-10 by reverse-transcription polymerase chain reaction (RT-PCR) and infection assays.(a) Genetic organization of the xcs and xps gene clusters. Genes are represented by arrows. Letters refer to individual xcs and xps geneproducts and the nomenclature generally used for components of type II secretion (T2S) systems. XpsD, E, F, G and H are homologous tocorresponding Xcs proteins (shown in black). By contrast, XpsI, XpsJ, XpsK, XpsL, XpsM and XpsN do not share significant homology with Xcsproteins and vice versa. The homolog of xcsN is missing in the xps gene cluster. xpsN corresponds to xcsC (indicated as N (C)). XCV3669encodes a predicted extracellular protease. Deletions (D) that were generated in this study are indicated. (b) RT-PCR analysis of xcsE and xpsE.xcsE and xpsE fragments were amplified from cDNA derived from Xcv strain 85-10 using gene-specific primers. Genomic DNA (gDNA) fromstrain 85-10 was used as positive control. As negative controls, the template was replaced by water or the reaction was performed in theabsence of reverse transcriptase ()RT). (c) Infection assays with wild-type and xpsD deletion mutant strains. Xcv strains 85-10 (wt) and thesecretin deletion mutant strain 85-10DxpsD (DxpsD) carrying the empty vector ()) or an xpsD expression construct (XpsD) were inoculated ata density of 2 · 107 CFU ml)1 into leaves of susceptible ‘Early Cal Wonder’ (ECW) pepper plants. Disease symptoms were photographed at5 d post inoculation (dpi). Dashed lines mark inoculated areas. (d) The double deletion mutant strain 85-10DxpsDDxcsD is less virulent thanstrain 85-10DxpsD. Strains 85-10 (wt), 85-10DxpsD (DxpsD), 85-10DxpsDDxcsD (DxpsDDxcsD) and 85-10DxpsDDxcsE (DxpsDDxcsE) wereinoculated at a density of 108 CFU ml)1 into leaves of susceptible ECW pepper plants. Disease symptoms were photographed at 5 dpi and7 dpi. Dashed lines mark inoculated areas. Note that disease symptoms caused by strains 85-10DxpsD and 85-10DxpsDDxcsE at 7 dpi arereduced compared with the wild-type, the necrosis is less pronounced and only partial. (e) In planta growth of secretin deletion mutants.Strains 85-10 (wt), 85-10DxcsD (DxcsD), 85-10DxpsD (DxpsD) and 85-10DxpsDDxcsD (DxpsDDxcsD) were inoculated at a density of104 CFU ml)1 into leaves of susceptible ECW pepper plants. Bacterial growth was determined over a period of 7 dpi. Values are the mean ofthree samples from three different plants. Error bars represent standard deviations. One representative of three independent experiments isshown. (f) In vitro bacterial growth of xps and xcs deletion mutants. Strains 85-10 (wt), 85-10DxcsD (DxcsD), 85-10DxpsD (DxpsD),85-10DxpsDDxcsD (DxpsDDxcsD), and 85-10DxpsDDxcsE (DxpsDDxcsE), were grown overnight in complex nutrient-yeast-glycerol (NYG)medium and resuspended in minimal medium A at an optical density (OD600 nm) of 0.2. The cultures were incubated at 30�C and the opticaldensity was measured over a period of 24 h. Error bars represent standard deviations. The experiment was repeated twice with similar results.
988 Research
NewPhytologist
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

when bacteria were inoculated into leaves of susceptibleECW pepper plants, strain 85-10 induced water-soakedlesions (Fig. 1c). A similar phenotype was observed forstrains 85-10DxcsD and 85-10Dxcs, which lacks thecomplete xcs gene cluster, suggesting that xcs genes do notsignificantly contribute to virulence (data not shown;Fig. S2). By contrast, strain 85-10DxpsD led to reduced dis-ease symptoms (Fig. 1c). This effect was more pronouncedwhen bacteria were inoculated at low population densities(2 · 107 CFU ml)1; compare Fig. 1c,d). The xpsD mutantphenotype was complemented by ectopic expression ofxpsD, suggesting that it was specifically caused by the dele-tion of xpsD (Fig. 1c).
To investigate potential functional redundancies amongsecretin-encoding genes, we deleted both xpsD and xcsDfrom the genome of Xcv strain 85-10. XcsD shares 28%sequence identity with XpsD. Notably, the resulting doubledeletion mutant strain 85-10DxpsDDxcsD caused reduceddisease symptoms when compared with strain 85-10DxpsD(Fig. 1d). By contrast, the double deletion mutant strain85-10DxpsDDxcsE caused disease symptoms similar to thexpsD single deletion mutant (Fig. 1d). This suggests thatthe additional reduction in virulence of strain 85-10DxpsDDxcsD was specifically caused by the lack of xcsD.We therefore conclude that XcsD can at least partially com-pensate the loss of XpsD. In addition to disease symptoms,we analysed bacterial growth of wild-type and deletionmutant strains in susceptible ECW pepper plants. In plantagrowth of strain 85-10DxcsD 7 dpi was similar to that ofthe wild-type strain 85-10, whereas 85-10DxpsD numberswere c. 100-fold reduced compared with strain 85-10(Fig. 1e). Additional deletion of xcsD in strain 85-10DxpsDled to a further reduction of bacterial counts 7 dpi, which isin good agreement with the observed phenotypes (Fig. 1e).Reduced bacterial growth of single and double deletionmutants in planta was presumably not caused by a generalgrowth deficiency, because bacterial multiplication in mini-mal medium was not affected (Fig. 1f).
Characterization of Xcv mutants lacking predictedT2S-associated ATPases
To confirm that the virulence phenotypes of secretinmutants were caused by nonfunctional T2S systems and notby alterations in the outer membrane, we deleted xcsE, xpsEand XCV4312, respectively. XCV4312 encodes a predictedATPase with homology to XcsE and XpsE and is locatedoutside of the xcs and xps gene clusters of Xcv strain 85-10.Deletion of xpsE led to a significant reduction in diseasesymptoms and in planta bacterial growth (Fig. 2a,b).Reduced disease symptoms but not bacterial growth ofthe xpsE mutant was more pronounced than for strain85-10DxpsD, which was presumably a result of the func-tional redundancy of xpsD and xcsD (Fig. 2c,d; see above).
By contrast, xcsE and XCV4312 deletion mutants displayeda wild-type phenotype (Fig. 2a; data not shown).Furthermore, strain 85-10DxpsEDxcsE provoked diseasesymptoms comparable to strain 85-10DxpsE (Fig. 2a,c).Notably, however, deletion of xcsE in strain 85-10DxpsE ledto a further reduction of in planta bacterial growth, suggest-ing that in this respect XcsE (42% sequence identity withXpsE) can partly complement the xpsE deletion mutant(Fig. 2b). Reduced virulence and in planta growth of singleand double deletion mutants was presumably not caused bya general growth deficiency, because bacterial multiplicationin minimal medium was not affected (Fig. 2e). The xpsEmutant phenotype was complemented by ectopic expressionof xpsE or xpsE-c-myc, which encodes a C-terminally c-Mycepitope-tagged XpsE (Fig. 2f).
Type III-dependent effector protein translocation isreduced in the absence of xpsE
In addition to disease symptoms, we investigated whetherXps and Xcs systems from Xcv contribute to the inductionof plant defense responses that are triggered upon recogni-tion of type III effectors (also termed avirulence (Avr) pro-teins) in resistant plants carrying cognate resistance (R)genes. Avr protein-triggered plant defense is often associatedwith the HR, a rapid local plant cell death that restrictsbacterial multiplication (Dangl & Jones, 2001). For theanalysis of the HR induction, strains 85-10, 85-10Dxcs and85-10DxpsE were inoculated into leaves of resistant ECW-10Rpepper plants that carry the Bs1 resistance gene and inducethe HR upon recognition of the type III effector AvrBs1that is delivered by strain 85-10 (Ronald & Staskawicz,1988; Escolar et al., 2001). Deletion of the complete xcsgene cluster in strain 85-10 did not affect the HR (data notshown). By contrast, the HR induced by strain 85-10DxpsEwas significantly reduced when compared with the wild type(Fig. 3a). We then tested translocation of additional typeIII effector proteins including AvrBs3, XopC, XopJ andXopF1 using AvrBs3D2 as a reporter. AvrBs3D2 is a deriva-tive of the type III effector AvrBs3 lacking the secretion andtranslocation signal. However, AvrBs3D2 contains the effec-tor domain and is recognized in resistant ECW-30R pepperplants when fused to a functional T3S and translocationsignal (Szurek et al., 2002; Noel et al., 2003). As expected,strain 85-10 expressing AvrBs31-200-, XopC1-200-, XopJ1-155-and XopF11-200-AvrBs3D2 fusion proteins induced the HRin ECW-30R leaves (Szurek et al., 2002; Noel et al., 2003;Buttner et al., 2007; Fig. 3a). However, the HR induced bythe corresponding xpsE deletion mutant strains wasreduced, suggesting that XpsE is required for efficienteffector protein translocation (Fig. 3a).
To investigate whether reduced effector protein trans-location was caused by reduced T3S, we performed in vitroT3S assays with strains 85-10hrpG* (85*) and 85*DxpsE.
NewPhytologist Research 989
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

Both strains contain a constitutively active derivative of thekey regulator HrpG and therefore express T3S genes invitro under noninducing conditions (Wengelnik et al.,1999). For the analysis of in vitro T3S, strains 85* and
85*DxpsE were incubated in secretion medium and total cellextracts and culture supernatants were analysed by immuno-blotting. Fig. 3b shows that comparable amounts of theT3S translocon protein HrpF and the effector fusion proteins
– – XpsE XpsE-c-Myc
wt ΔxpsE
22 4 6 8
wtΔxcsEΔxpsE
Time post inoculation (d)
Log
(CF
U c
m–2
)
8
7
6
5
4
3
ΔxpsEΔxcsE
10
ΔhrcN
0 6 24 0 24 h
20
40
60
80
100
120
Per
cent
age
of w
t gro
wth
wtΔxcsE
Growth in minimal medium A
ΔxpsE
(a)
(c)
(d)
(e)
(f)
(b)
22 4 6 8
wtΔxpsD
Time post inoculation (d)
Log
(CF
U c
m–2
)
8
7
6
5
4
3
ΔxpsE
10
ΔhrcN
wt ΔxpsEΔxcsEΔxpsEΔxcsE
Δ4312ΔxcsEΔ4312
ΔxpsEΔ4312
ΔxpsEΔxcsEΔ4312
ΔxpsEΔxcsEΔxpsEΔxcsEΔ4312
wt ΔxpsDΔxpsEΔxpsEΔxcsE
ΔxpsDΔxcsD
Fig. 2 Analysis of deletion mutants deficient in the predicted Xps and Xcs system-associated ATPases. (a) The predicted T2S system-associatedATPase XpsE contributes to disease symptoms in pepper plants. Xcv strains 85-10 (wt), 85-10DxcsE (DxcsE), 85-10DxpsE (DxpsE),85-10DxpsEDxcsE (DxpsEDxcsE), 85-10D4312 (D4312), 85-10DxcsED4312 (DxcsED4312), 85-10DxpsED4312 (DxpsED4312) and85-10DxpsEDxcsED4312 (DxpsEDxcsED4312) were inoculated at population densities of 2 · 107 CFU ml)1 into leaves of susceptible ‘EarlyCal Wonder’ (ECW) pepper plants. Disease symptoms were photographed at 7 d post inoculation (dpi). Dashed lines delineate inoculatedareas. (b) Analysis of in planta bacterial growth of mutants deleted in predicted ATPases. Strains 85-10 (wt), 85-10DxcsE (DxcsE), 85-10DxpsE(DxpsE), 85-10DxpsEDxcsE (DxpsEDxcsE) and 85-10DhrcN (DhrcN, T3S mutant) were inoculated into leaves of ECW pepper plants. Bacterialmultiplication was determined over a period of 9 dpi. Values are the mean of three samples from three different plants. Error bars representstandard deviations. One representative of three independent experiments is shown. (c) Comparative infection assays with single and doubledeletion mutants. Strains 85-10 (wt), 85-10DxpsE (DxpsE), 85-10DxpsD (DxpsD), 85-10DxpsEDxcsE (DxpsEDxcsE) and 85-10DxpsDDxcsD
(DxpsDDxcsD) were inoculated into leaves of susceptible ECW pepper plants as described in (a). Disease symptoms were photographed 5 dpi.Dashed lines mark inoculated areas. (d) Analysis of in planta bacterial growth of xpsE and xpsD deletion mutants. Strains 85-10 (wt),85-10DxpsE (DxpsE), 85-10DxpsD (DxpsD) and 85-10DhrcN (DhrcN; T3S mutant) were inoculated into leaves of susceptible ECW pepperplants and bacterial multiplication was determined over a period of 8 dpi as described in (b). (e) In vitro bacterial growth of xps and xcs
deletion mutants. Strains 85-10 (wt), 85-10DxcsE (DxcsE), 85-10DxpsE (DxpsE), 85-10DxpsEDxcsE (DxpsEDxcsE), and 85-10DxpsEDxcsED4312 (DxpsEDxcsED4312), were grown overnight in complex NYG medium and resuspended in minimal medium A at anoptical density (OD600 nm) of 0.2. The cultures were incubated at 30�C and the optical density was measured over a period of 24 h. Error barsrepresent standard deviations. (f) Complementation of the xpsE mutant phenotype. Strains 85-10 (wt) and 85-10DxpsE (DxpsE) carrying theempty vector ()) or expression constructs encoding XpsE or XpsE-c-Myc as indicated were inoculated at a density of 2 · 107 CFU ml)1 intoleaves of susceptible ECW pepper plants. Disease symptoms were photographed 7 dpi. Dashed lines indicate the inoculated areas.
990 Research
NewPhytologist
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

XopJ1-155–AvrBs3D2 and XopC1-200–AvrBs3D2, whichwere selected for in vitro T3S assays, were detected in theculture supernatants of both strains. These results suggestthat the Xps system is not essential for the activity of theT3S system per se.
Next, we investigated at which time-point after infectionthe reduction in effector protein translocation is detectable.For this, we blocked bacterial protein synthesis in infectedleaf tissue of ECW-10R pepper plants by treatment withspectinomycin. Infiltration of spectinomycin 0 and 4 hpiabolished the HR induction by strains 85-10 and 85-10DxpsE (Fig. 3c). However, strain 85-10 induced the HRwhen spectinomycin was infiltrated 6 hpi, whereas no HRinduction was observed for strain 85-10DxpsE (Fig. 3c).This suggests that the amounts of effector proteins trans-located within 6 hpi by the wild-type, but not by the xpsEdeletion mutant are sufficient to elicit the HR. ReducedHR induction 6 hpi was presumably not caused by differ-ences in bacterial counts of strains 85-10 and 85-10DxpsE,which were comparable 6 hpi (Fig. 3d).
We also analysed the accumulation of Hrp and effectorproteins 6 hpi by immunoblotting. Expression of hrp genesis specifically activated when the bacteria enter the apoplast,presumably by plant-derived molecules (Schulte & Bonas,1992; Wengelnik et al., 1996a,b). When proteins wereextracted 6 hpi from leaves of susceptible ECW pepperplants that were inoculated with Xcv wild-type and xpsEdeletion mutants, comparable levels of the predicted lipo-protein HrcJ and the effector fusion XopF1-200-AvrBs3D2were detected in both 85-10 and 85-10DxpsE (Fig. 3e,f). Aslight reduction in HrpF levels was observed for the xpsEdeletion mutant, however, previous experiments indicatedthat this is not sufficient to cause a reduction in HR induc-tion (C. Lorenz & D. Buttner, unpublished). We concludefrom our experiments that xpsE is required for efficienteffector protein translocation.
The xpsE mutant phenotype can be complementedin trans
It is conceivable that the reduced virulence of strain 85-10DxpsE results from a lack of extracellular bacterialproteins. We therefore wondered whether this deficiencycan be complemented in trans. For this, we coinoculatedstrain 85-10DxpsE with strains 85-10DhrcN and85-10DhrcNDxpsE, respectively, both deficient in T3S andtherefore nonpathogenic (Lorenz & Buttner, 2009; Fig. 4).Disease symptoms in ECW pepper leaves were partiallyrestored when strain 85-10DxpsE was coinoculated withstrain 85-10DhrcN (Fig. 4). This was not observed whenstrain 85-10DxpsE was coinoculated with strain85-10DhrcNDxpsE, which is deficient in the Xps system,suggesting that the restoration of the wild-type phenotypewas specifically caused by Xps substrates (Fig. 4). Similarly,
HR induction of strain 85-10DxpsE in resistant ECW-10Rpepper plants was enhanced upon coinoculation with strain85-10DhrcN but not with strain 85-10DhrcNDxpsE. Wetherefore conclude that the efficient translocation of effectorproteins by the xpsE deletion mutant can be restored in trans.
The Xps system contributes to extracellular proteaseand xylanase activities
Next, we investigated the contribution of the Xcs and Xpssystem to the secretion of degradative enzymes, includingproteases, xylanases, cellulases and amylases, by suitableplate assays (see the Materials and Methods section). Strain85-10 secretes extracellular proteases that degrade milk pro-teins, which is visible as a cleared halo around the bacteria(Noel et al., 2001; Fig. 5a). The analysis of xcs and xpsmutants revealed that halo formation on milk plates by xcsmutants was like wild type (Fig. 5a). By contrast, no halowas detected for strains 85-10DxpsE and 85-10DxpsD,suggesting that the Xps system is required for extracellularprotease activity (Fig. 5b).
To analyse the activity of extracellular xylanases, bacteriawere incubated on NYG agar plates containing RBB xylan.Degradation of xylan is visible as a clear zone around thebacteria. Deletion of xps, but not of xcs genes resulted insignificantly reduced extracellular xylanase activity 1 dpi(Fig. 5a,b). Notably, when bacteria were incubated for 2 don RBB xylan plates there was residual extracellular xylanaseactivity for xps deletion mutants (see Fig. 5b). Becausesimilar results were observed for the triple deletion mutants85-10DxpsEDxcsED4312 and 85-10DxpsDDxcsDDhrcC, theresidual xylanase activity of xps deletion mutants waspresumably not caused by cross-complementation byhomologous genes (Fig. 5c). Strain 85-10DxpsDDxcsDDhrcC lacks both secretins of the T2S systems and HrcC,which is the secretin of the T3S system and shares 25%identity with XpsD. Reduced activity of extracellularproteases and xylanases in xpsD and xpsE deletion mutantswas complemented by ectopic expression of xpsD and xpsE-c-myc, respectively (Fig. 5d,e).
Extracellular amylase and cellulase activities areindependent of the Xcs and Xps systems
In addition to extracellular protease and xylanase activities,we analysed the activities of extracellular cellulases andamylases on plates containing CMC and starch, respec-tively. To visualize substrate degradation, CMC plates werestained with Congo red and starch plates with Lugol’s solution.Unexpectedly, extracellular cellulase and amylase activitiesin xcs and xps deletion mutants were like wildtype (Fig. 6a,b). Similar results were obtained with thetriple deletion mutants 85-10DxpsEDxcsED4312 and85-10DxpsDDxcsDDhrcC (Fig. 6c,d), suggesting that the
NewPhytologist Research 991
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

Xcs and Xps systems are not required for secretion ofamylases and cellulases from Xcv. Notably, however, secre-tion of amylases and cellulases in Xcc and Xoo depends onthe T2S system (Hu et al., 1992; Ray et al., 2000; Furutaniet al., 2004; Sun et al., 2005).
Analysis of candidate T2S substrates
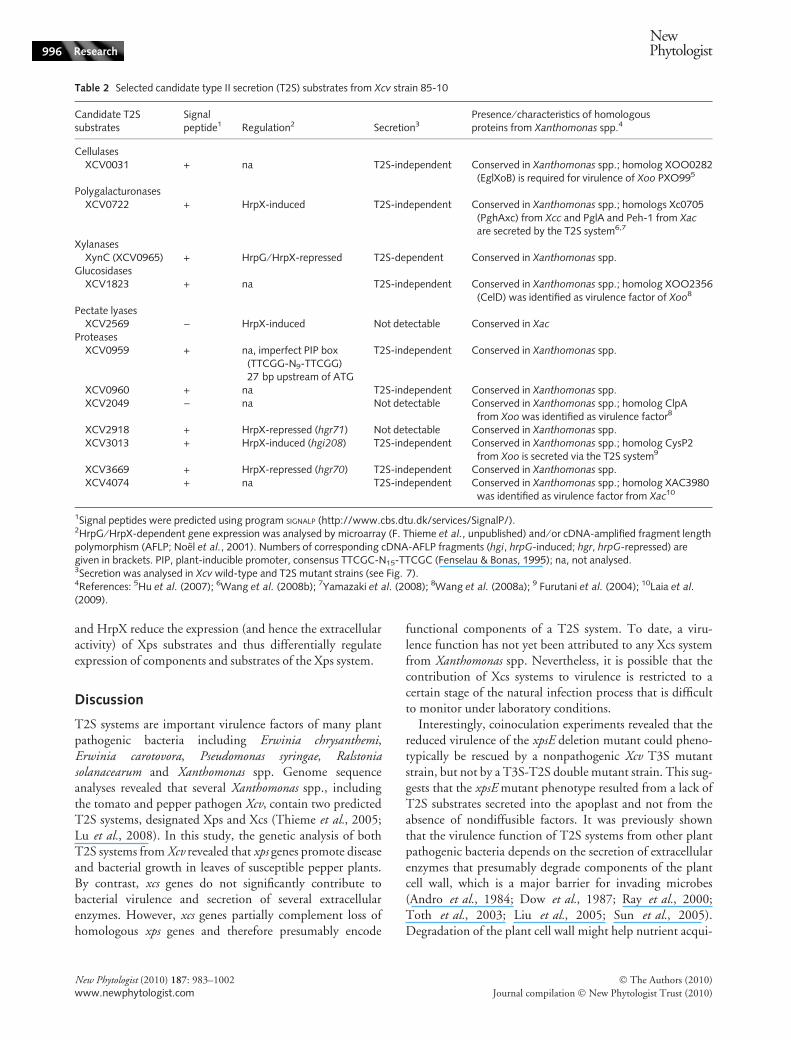
Next, we analysed secretion of candidate T2S substratesincluding a putative cellulase (XCV0031), polygalacturonase
(XCV0722), xylanase (XCV0965), glucosidase (XCV1823),pectate lyase (XCV2569) and several proteases (XCV0959,XCV0960, XCV2049, XCV2918, XCV3013, XCV3669,XCV4074; Table 2). Candidate proteins were selected basedon homologies to known T2S substrates and virulence fac-tors from Xanthomonas spp. and ⁄ or their coregulation withthe T3S system (Noel et al., 2001; F. Thieme et al., unpub-lished; see Table 2). All proteins were synthesized in Xcv asC-terminally c-Myc epitope-tagged derivatives. Wild-typeand T2S mutant strains carrying the respective expression
(a) (d)
(e)
(b)
(c)
AvrBs31-200-AvrBs3Δ2
XopC1-200-AvrBs3Δ2
XopF11-200-AvrBs3Δ2
XopJ1-155-AvrBs3Δ2
ECW-10R
wt ΔxpsE
ECW-30R
wt ΔxpsE
wt ΔxpsE
TE SN
HrpF
XopJ1-155-AvrBs3Δ2
XopC1-200-AvrBs3Δ2
95
95
kDa
95
wt
Δxps
E
ECW-10R
– spec 0 hpi
+ spec
1×10
8 cfu ml –1
4 hpi 6 hpi
wt ΔxpsE
Δxcs
HrpF
Unspecific signal
HrcJ
GroEL
95
kDa
55
26
17
–
6 hpi
Ponceau
2
3
4
5
6
7
24 hpi6 hpi
Log
(CF
U c
m–2
)
8
9
0 hpi
*
wt ΔxpsE
Δxcs
95
kDa –XopF11-200-AvrBs3Δ2
(f)
6 hpi
Ponceau
55
26
34
43
55
34
43
72
992 Research
NewPhytologist
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

constructs were incubated in NYG medium, and total cellextracts and culture supernatants were analysed by immuno-blotting using a c-Myc epitope-specific antibody.
Most proteins except the predicted pectate lyaseXCV2569-c-Myc and the predicted proteases XCV2918-c-Myc and XCV2049-c-Myc were detected in the culturesupernatant of the wild-type strain, indicating that theywere secreted (Fig. 7a,b). Unexpectedly, for the predictedproteases XCV0959 and XCV3669 we detected two pro-teins in the culture supernatants including smaller and ⁄ orlarger proteins than the proteins detected in total cellextracts. Smaller proteins are presumably cleavage productswhereas larger proteins might result from a specific proteinmodification in the culture supernatant. However, it cannotbe excluded that secreted XCV0959-c-Myc and XCV3669-
c-Myc are part of protein complexes that are not dissolvedby SDS-PAGE. Note that XCV0959-c-Myc was less stablein cell extracts of T2S mutant strains. Unexpectedly, c-Mycepitope-tagged derivatives of XCV0031, XCV0722,XCV0959, XCV0960, XCV1823, XCV3013, XCV3669and XCV4074 were detected in the culture supernatants ofT2S-deficient strains, suggesting that secretion of these pro-teins was independent of both T2S systems (Fig. 7a,c). Thiswas not caused by cell lysis as, for example, the cytoplasmicT3S system-associated ATPase HrcN that was ectopicallyexpressed as Strep or c-Myc epitope-tagged derivative wasonly detectable in total cell extracts (Fig. 7a).
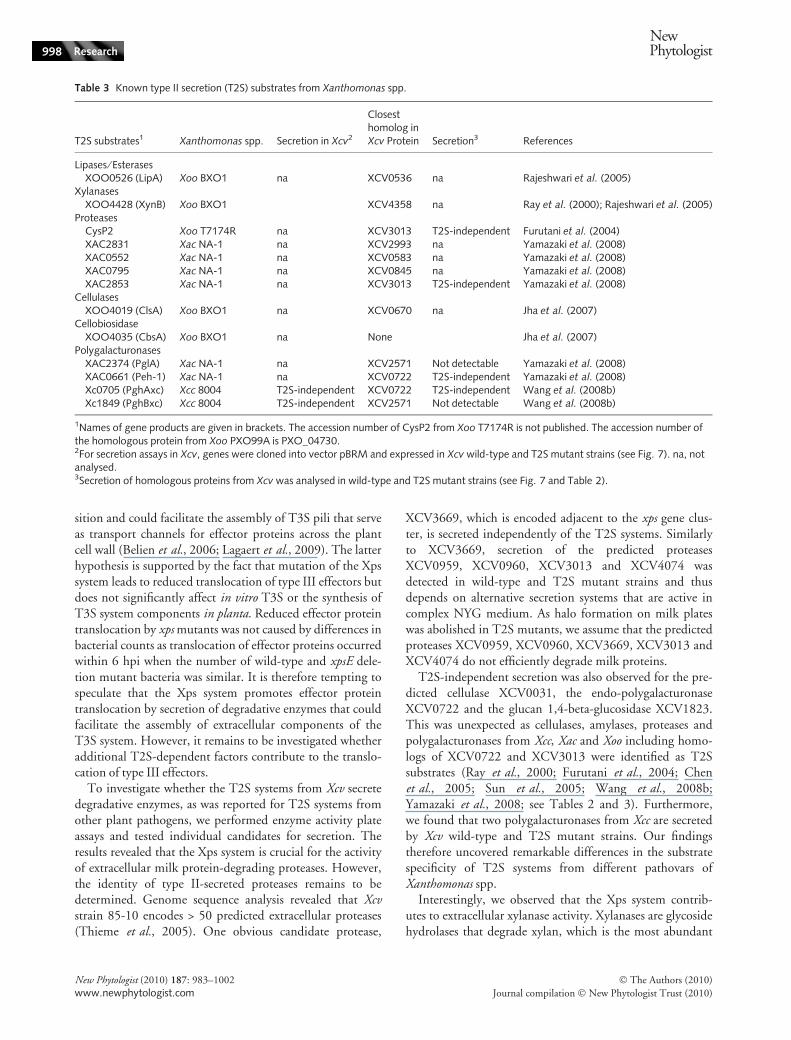
We also investigated secretion of two polygalacturonases(PghAxc and PghBxc) from Xcc that were previously identi-fied as T2S substrates (Wang et al., 2008b; Table 3). The
Fig. 3 The Xps system contributes to type III-dependent effector protein translocation. (a) XpsE contributes to the induction of the hypersensitiveresponse (HR). Strains 85-10 (wt) and 85-10DxpsE (DxpsE) were inoculated at a density of 2 · 107 CFU ml)1 into leaves of resistant ‘EarlyCal Wonder’ (ECW)-10R pepper plants. Similarly, both strains carrying AvrBs31-200-, XopC1-200-, XopJ1-155- and XopF11-200-AvrBs3D2 fusionproteins, respectively, were inoculated into leaves of ECW-30R pepper plants that trigger the HR upon recognition of AvrBs3. For the bettervisualization of the HR, leaves were bleached in ethanol 2 d post inoculation (dpi). Dashed lines indicate the inoculated areas. (b) In vitro T3Sassays with wild-type and xpsE deletion mutants. Strains 85* (wt) and 85*DxpsE (DxpsE) were incubated in T3S-inducing medium and totalcell extracts (TE) and culture supernatants (SN) were analyzed by immunoblotting, using antibodies specific for the T3S translocon proteinHrpF and the effector protein AvrBs3, respectively. XopJ1-155–AvrBs3D2 and XopC1-200–AvrBs3D2 were ectopically expressed in both strains.Upper signals correspond to HrpF, XopJ1-155-AvrBs3D2 and XopC1-200–AvrBs3D2, respectively, lower signals are degradation products. (c)Timing of effector protein translocation. Xcv strains 85-10 (wt) and 85-10DxpsE (DxpsE) were inoculated into leaves of resistant ECW-10Rpepper plants at bacterial densities of 1 · 108 CFU ml)1. Infected leaf areas were infiltrated with spectinomycin (10 mg ml)1) 0, 4 and 6 hpito block bacterial protein synthesis. Leaves were bleached in ethanol 2 dpi. Dashed lines indicate the inoculated areas. (d) In planta bacterialmultiplication of wild-type and xpsE deletion mutant strains. Xcv strains 85-10 (wt; dark tinted bars) and 85-10DxpsE (DxpsE; light tinted bars)were inoculated into leaves of susceptible ECW pepper plants at a density of 1 · 108 CFU ml)1 and bacterial growth was determined 0, 6 and24 hpi. The asterisk indicates a significant difference with P < 0.05 based on the results of an unpaired Student’s t test. (e) Analysis of bacterialprotein accumulation in wild-type and T2S mutant strains isolated from susceptible pepper plants. Xcv strains 85-10 (wt), 85-10Dxcs (Dxcs)and 85-10DxpsE (DxpsE) were inoculated into leaves of susceptible ECW pepper plants at a density of 109 CFU ml)1. Proteins were extractedfrom uninfected plant material ()) or infected plant material 6 hpi and analysed by Ponceau staining and immunoblotting, using HrpF-,HrcJ- and GroEL-specific antibodies. The signal in the fourth panel represents an unknown plant protein that is nonspecifically detected by theHrpF-specific antibody and shows that comparable protein amounts were loaded. (f) Analysis of effector protein accumulation in wild-typeand T2S mutant strains isolated from susceptible pepper plants. Xcv strains 85-10 (wt), 85-10Dxcs (Dxcs) and 85-10DxpsE (DxpsE) wereinoculated into leaves of susceptible ECW pepper plants at a density of 109 CFU ml)1. Proteins were extracted as described in (e) and analysedby Ponceau staining and immunoblotting, using an AvrBs3-specific antibody. XopF1-200–AvrBs3D2 was ectopically expressed from a respectiveexpression construct. The upper signal corresponds to XopF1-200–AvrBs3D2 and lower bands are degradation products.
wt ΔxpsE
+ ΔhrcN+ MgCl2+ ΔhrcN ΔxpsEΔhrcNMgCl2
ΔhrcNΔxpsE
ECW
ECW-10R
+ ΔhrcN+ MgCl2+ ΔhrcN ΔxpsE
Fig. 4 The xpsE mutant phenotype can be complemented in trans. Xcv strains 85-10 (wt) and 85-10DxpsE (DxpsE), respectively, were mixed1 : 1 with MgCl2, strain 85-10DhrcN (DhrcN) or strain 85-10DhrcNDxpsE (DhrcNDxpsE) as indicated and inoculated at a final concentration of2 · 107 CFU ml)1 into leaves of susceptible ‘Early Cal Wonder’ (ECW) and resistant ECW-10R pepper plants; MgCl2, strains 85-10DhrcN and85-10DhrcNDxpsE were included as negative controls. Disease symptoms were photographed at 5 d post inoculation (dpi). For the bettervisualization of the hypersensitive response (HR), leaves were bleached in ethanol 1 dpi. Dashed lines mark the inoculated areas.
NewPhytologist Research 993
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

corresponding genes (Xc0705 and Xc1849) were amplifiedfrom Xcc strain 8004 and expressed as C-terminally c-Mycepitope-tagged derivatives in Xcv strains 85-10 and85-10DxpsEDxcsED4312. When analysed for secretion,PghAxc-c-Myc and PghBxc-c-Myc were detected in theculture supernatants of both Xcv strains, suggesting thatthey were secreted but independently of the T2S systems(Fig. 7d). We therefore conclude that T2S substrate recog-nition differs in Xcv and Xcc.
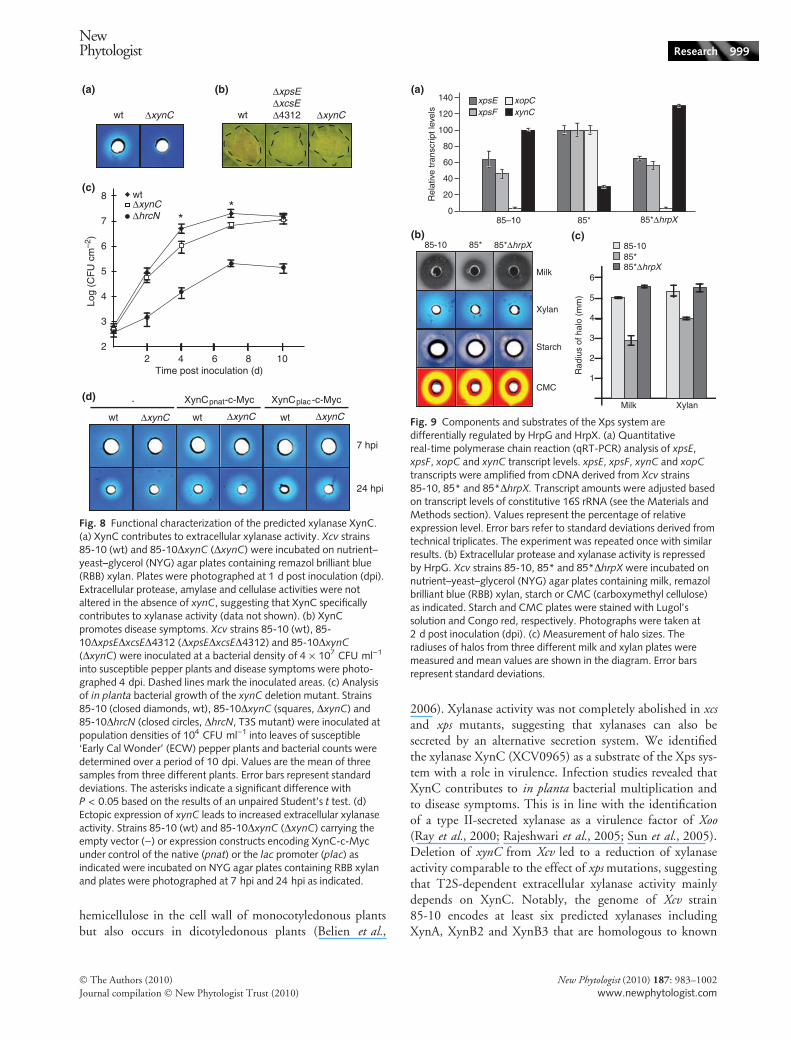
The xylanase XynC is secreted by the Xps system andcontributes to virulence
In contrast to most candidate substrates tested, secretionof the predicted xylanase XCV0965 (hereafter referred toas XynC) was severely reduced in strain 85-
10DxpsEDxcsED4312 when compared with the wild-typestrain (Fig. 7a). Secretion assays with single deletionmutants revealed that secretion of XynC-c-Myc was depen-dent on xps but not on xcs genes (Fig. 7e). To investigatethe contribution of the predicted xylanase XynC to extra-cellular xylanase activity and bacterial virulence, we deletedthe corresponding gene from the genome of Xcv strain85-10. When strains 85-10 and 85-10DxynC were grownon NYG agar plates containing RBB xylan, as describedearlier, extracellular xylanase activity was severely reduced instrain 85-10DxynC (Fig. 8a). We also analysed a potentialrole of XynC in virulence. When bacteria were inoculatedinto leaves of susceptible ECW pepper plants, strain85-10DxynC induced reduced disease symptoms comparedwith the wild-type strain 85-10 (Fig. 8b). Furthermore,deletion of xynC resulted in reduced bacterial counts in
wt Δxcs ΔxcsE ΔxcsD
Milk
Xylan
Xylan
Milk
wt ΔxpsD
- XpsD-
wt ΔxpsE
- XpsE-c-Myc-
Xylan
Milk
1 dpi2 dpi
wt
ΔxpsEΔxcsEΔ4312 wt
ΔxpsDΔxcsDΔhrcC
wt ΔxpsE ΔxpsD
Milk
Xylan1 dpi
2 dpi1 dpi
2 dpi
(a)
(c)
(d)
(b)
(e)
Fig. 5 Analysis of extracellular protease and xylanase activities in xcs and xps deletion mutants. (a) The Xcs system is dispensable for extra-cellular protease and xylanase activity. Xcv strains 85-10 (wt), 85-10Dxcs (Dxcs), 85-10DxcsE (DxcsE) and 85-10DxcsD (DxcsD) wereincubated on nutrient–yeast–glycerol (NYG) agar plates containing milk or remazol brilliant blue (RBB) xylan. Plates were photographed at 1 dpost inoculation (dpi). (b) The Xps system contributes to extracellular protease and xylanase activity. Xcv strains 85-10 (wt), 85-10DxpsE
(DxpsE) and 85-10DxpsD (DxpsD) were incubated on NYG agar plates containing milk or RBB xylan and plates were photographed at 1 dpiand 2 dpi as indicated. (c) Extracellular xylanase activity is not completely abolished in T2S mutants. Strains 85-10 (wt), 85-10DxpsEDxcsE
D4312 (DxpsEDxcsED4312) and 85-10DxpsDDxcsDDhrcC (DxpsDDxcsDDhrcC) were incubated on NYG agar plates containing RBB xylan.Plates were photographed at 1 and 2 dpi as indicated. (d) Complementation of extracellular protease and xylanase activities in the xpsE dele-tion mutant. Strains 85-10 (wt) and 85-10DxpsE (DxpsE) carrying the empty vector ()) or an xpsE-c-myc expression construct (XpsE-c-Myc)were incubated on NYG plates containing milk or RBB xylan and plates were photographed at 1 dpi. (e) Complementation of extracellularprotease and xylanase activities in the xpsD deletion mutant. Strains 85-10 (wt) and 85-10DxpsD (DxpsD) carrying the empty vector ()) or anxpsD expression construct (XpsD) were incubated on NYG plates containing milk or RBB xylan and plates were photographed at 1 dpi.
994 Research
NewPhytologist
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

planta 4–7 dpi (Fig. 8c). We conclude from these data thatXynC is an active xylanase, which is secreted by the Xpssystem and contributes to bacterial virulence.
Notably, ectopic expression of xynC-c-myc under controlof the native or the lac promoter in the wild-type or thexynC deletion mutant led to increased extracellular xylanaseactivity in vitro, which was already detectable 5–7 hpi,whereas halo formation for the wild-type strain was onlyobserved 24 hpi (Fig. 8d). However, xynC-c-myc expressiondid not reproducibly restore the virulence of the xynCdeletion mutant (data not shown). We speculate that lack ofreproducible complementation resulted from the enhancedxylanase activity which might not be favorable for the bacterialinteraction with the host (note that xynC is down-regulatedby HrpG; see below).
xps genes and T2S substrates are differentiallyregulated by HrpG and HrpX
Given the potential interplay between T2S and T3Ssystems, we speculated whether xps genes and xynC areregulated by HrpG and HrpX, which are the key regulatorsof the T3S system (Wengelnik & Bonas, 1996; Wengelniket al., 1996b). For this, we performed qRT-PCR analysis ofstrains 85-10, 85* and 85*DhrpX (deleted in the transcrip-tional activator HrpX) grown in NYG medium using
primers specific for xpsE, xpsF and xynC, respectively. Theamounts of both xpsE and xpsF transcripts were increased instrain 85* when compared with strains 85-10 and85*DhrpX, suggesting that expression of xpsE and xpsF isenhanced by HrpG and HrpX (Fig. 9a). As control forHrpG ⁄ HrpX-dependent gene expression we analysed tran-script abundance of the type III effector gene xopC, which isinduced by HrpG and HrpX (Noel et al., 2003; Fig. 9a).In contrast to xpsE, xpsF and xopC, however, xynC transcriptamounts were significantly reduced in strain 85* comparedwith strains 85-10 and 85*DhrpX, suggesting that expres-sion of xynC is downregulated by HrpG and HrpX(Fig. 9a).
We also analysed potential effects of HrpG and HrpX onextracellular enzyme activities. For this, we performed plateassays as described above with strains 85-10, 85* and85*DhrpX. It was previously shown that extracellular prote-ase activity of Xcv is suppressed by HrpG and HrpX (Noelet al., 2001; Fig. 9b,c). Similarly to protease activity, extra-cellular xylanase activity was decreased in strain 85* whencompared with strains 85-10 and 85*DhrpX (Fig. 9b,c).This is in good agreement with the reduced xynC transcriptlevels in strain 85* (see above). In contrast to protease andxylanase activities, amylase and cellulase activities that areindependent of the T2S systems were slightly increased instrain 85* (Fig. 9b,c). Overall, our results suggest that HrpG
wt ΔxpsE ΔxpsD
CMC
Starch
wt ΔxcsE ΔxcsD
CMC
Starch
wt
ΔxpsEΔxcsEΔ4312 wt
ΔxpsDΔxcsDΔhrcC
Starch
CMC
CMC
Starch
(c)
(a)
(d)
(b)
Fig. 6 Extracellular cellulase and amylase activity is not affected in xcs and xps deletion mutants. (a) Cellulase and amylase activities of xcs
single deletion mutants. Xcv strains 85-10 (wt), 85-10DxcsE (DxcsE) and 85-10DxcsD (DxcsD) were incubated at an optical density of 1.0 at600 nm on nutrient–yeast–glycerol (NYG) agar plates containing CMC (carboxymethyl cellulose) or starch as indicated. Cellulose was stainedwith Congo red and starch was visualized by Lugol’s solution. Plates were photographed at 2 d post inoculation (dpi). (b) Cellulase andamylase activities of xps single deletion mutants. Xcv strains 85-10 (wt), 85-10DxpsE (DxpsE) and 85-10DxpsD (DxpsD) were incubated onNYG agar plates containing CMC or starch as described in (a). Plates were photographed at 2 dpi. (c) Cellulase and amylase activities of anATPase-deficient triple deletion mutant. Strains 85-10 (wt) and 85-10DxpsEDxcsED4312 (DxpsEDxcsED4312) were incubated on NYG agarplates containing CMC or starch as described in (a). Photographs were taken at 2 dpi. (d) Cellulase and amylase activities of a secretin-deficienttriple deletion mutant. Strains 85-10 (wt) and 85-10DxpsDDxcsDDhrcC (DxpsDDxcsDDhrcC) were incubated on NYG agar plates containingCMC or starch as described in (a). Photographs were taken at 2 dpi.
NewPhytologist Research 995
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com
and HrpX reduce the expression (and hence the extracellularactivity) of Xps substrates and thus differentially regulateexpression of components and substrates of the Xps system.
Discussion
T2S systems are important virulence factors of many plantpathogenic bacteria including Erwinia chrysanthemi,Erwinia carotovora, Pseudomonas syringae, Ralstoniasolanacearum and Xanthomonas spp. Genome sequenceanalyses revealed that several Xanthomonas spp., includingthe tomato and pepper pathogen Xcv, contain two predictedT2S systems, designated Xps and Xcs (Thieme et al., 2005;Lu et al., 2008). In this study, the genetic analysis of bothT2S systems from Xcv revealed that xps genes promote diseaseand bacterial growth in leaves of susceptible pepper plants.By contrast, xcs genes do not significantly contribute tobacterial virulence and secretion of several extracellularenzymes. However, xcs genes partially complement loss ofhomologous xps genes and therefore presumably encode
functional components of a T2S system. To date, a viru-lence function has not yet been attributed to any Xcs systemfrom Xanthomonas spp. Nevertheless, it is possible that thecontribution of Xcs systems to virulence is restricted to acertain stage of the natural infection process that is difficultto monitor under laboratory conditions.
Interestingly, coinoculation experiments revealed that thereduced virulence of the xpsE deletion mutant could pheno-typically be rescued by a nonpathogenic Xcv T3S mutantstrain, but not by a T3S-T2S double mutant strain. This sug-gests that the xpsE mutant phenotype resulted from a lack ofT2S substrates secreted into the apoplast and not from theabsence of nondiffusible factors. It was previously shownthat the virulence function of T2S systems from other plantpathogenic bacteria depends on the secretion of extracellularenzymes that presumably degrade components of the plantcell wall, which is a major barrier for invading microbes(Andro et al., 1984; Dow et al., 1987; Ray et al., 2000;Toth et al., 2003; Liu et al., 2005; Sun et al., 2005).Degradation of the plant cell wall might help nutrient acqui-
Table 2 Selected candidate type II secretion (T2S) substrates from Xcv strain 85-10
Candidate T2Ssubstrates
Signalpeptide1 Regulation2 Secretion3
Presence ⁄ characteristics of homologousproteins from Xanthomonas spp.4
CellulasesXCV0031 + na T2S-independent Conserved in Xanthomonas spp.; homolog XOO0282
(EglXoB) is required for virulence of Xoo PXO995
PolygalacturonasesXCV0722 + HrpX-induced T2S-independent Conserved in Xanthomonas spp.; homologs Xc0705
(PghAxc) from Xcc and PglA and Peh-1 from Xacare secreted by the T2S system6,7
XylanasesXynC (XCV0965) + HrpG ⁄ HrpX-repressed T2S-dependent Conserved in Xanthomonas spp.
GlucosidasesXCV1823 + na T2S-independent Conserved in Xanthomonas spp.; homolog XOO2356
(CelD) was identified as virulence factor of Xoo8
Pectate lyasesXCV2569 ) HrpX-induced Not detectable Conserved in Xac
ProteasesXCV0959 + na, imperfect PIP box
(TTCGG-N9-TTCGG)27 bp upstream of ATG
T2S-independent Conserved in Xanthomonas spp.
XCV0960 + na T2S-independent Conserved in Xanthomonas spp.XCV2049 ) na Not detectable Conserved in Xanthomonas spp.; homolog ClpA
from Xoo was identified as virulence factor8
XCV2918 + HrpX-repressed (hgr71) Not detectable Conserved in Xanthomonas spp.XCV3013 + HrpX-induced (hgi208) T2S-independent Conserved in Xanthomonas spp.; homolog CysP2
from Xoo is secreted via the T2S system9
XCV3669 + HrpX-repressed (hgr70) T2S-independent Conserved in Xanthomonas spp.XCV4074 + na T2S-independent Conserved in Xanthomonas spp.; homolog XAC3980
was identified as virulence factor from Xac10
1Signal peptides were predicted using program SIGNALP (http://www.cbs.dtu.dk/services/SignalP/).2HrpG ⁄ HrpX-dependent gene expression was analysed by microarray (F. Thieme et al., unpublished) and ⁄ or cDNA-amplified fragment lengthpolymorphism (AFLP; Noel et al., 2001). Numbers of corresponding cDNA-AFLP fragments (hgi, hrpG-induced; hgr, hrpG-repressed) aregiven in brackets. PIP, plant-inducible promoter, consensus TTCGC-N15-TTCGC (Fenselau & Bonas, 1995); na, not analysed.3Secretion was analysed in Xcv wild-type and T2S mutant strains (see Fig. 7).4References: 5Hu et al. (2007); 6Wang et al. (2008b); 7Yamazaki et al. (2008); 8Wang et al. (2008a); 9 Furutani et al. (2004); 10Laia et al.
(2009).
996 Research
NewPhytologist
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

wtΔxpsEΔxcsE wt
ΔxpsEΔxcsEkDa
TE SN
XCV2569-c-Myc 55
43
XCV2918-c-Myc 43
XCV3013-c-Myc
XynC-c-Myc
XCV0031-c-Myc
XCV1823-c-Myc72
43
55
43
Strep-HrcN43
XCV4074-c-Myc55
ΔxcsEΔxpsEΔ4312wt wtkDa
TE SN
ΔxcsEΔxpsEΔ4312
wt ΔxcsE ΔxcsD ΔxpsE
TE
SN
XynC-c-Myc
55kDa
55
72
95
XCV3013-c-Myc
XCV0031-c-Myc
XCV1823-c-Myc72
43
43
XCV4074-c-Myc55
XCV0722-c-Myc72
ΔxcsDΔxpsDwt wtkDa
TE SN
ΔxcsDΔxpsD
72
95
PghAxc-c-Myc
ΔxcsEΔxpsEΔ4312wt wtkDa
TE SN
ΔxcsEΔxpsEΔ4312
55
72 PghBxc-c-Myc
72
95
55
43
43
XCV3669-c-Myc
HrcN-c-Myc
XCV2918-c-Myc
XCV0722-c-Myc72
XCV2569-c-Myc55
XCV3669-c-Myc XCV3669-c-Myc
XCV2049-c-Myc 96
XCV0959-c-Myc
XCV0960-c-Myc55
XCV0959-c-Myc7255
95
72
55
95
GroEL55
Strep-HrcN
GroEL
55
55
(a)
(b)
(c)
(d)
(e)
Fig. 7 Secretion experiments with T2S candidate substrates. (a) Analysis of in vitro secretion of predicted T2S substrates by Xcv. Strains 85-10(wt) and 85-10DxpsEDxcsED4312 (DxpsEDxcsED4312) carrying expression constructs encoding XCV3669-c-Myc, HrcN-c-Myc, XCV2918-c-Myc, XCV0031-c-Myc, XCV0722-c-Myc, XCV0959-c-Myc, XCV0960-c-Myc, XCV1823-c-Myc, XCV2569-c-Myc, XCV3013-c-Myc,XCV4074-c-Myc, XynC-c-Myc and Strep-HrcN, respectively (see also Table 2), were incubated in NYG medium. Total cell extracts (TE) andculture supernatants (SN) were analysed by immunoblotting, using c-Myc or Strep epitope- and GroEL-specific antibodies. The upper signalsin strains 85-10 and 85-10DxpsEDxcsED4312 carrying XCV0959-c-Myc and XCV3669-c-Myc, respectively, correspond to the full-lengthXCV0959-c-Myc and XCV3669-c-Myc proteins, lower bands are degradation products. Comparable amounts of the general chaperone GroELin TE of strains 85-10 (wt) and 85-10DxpsEDxcsED4312 (DxpsEDxcsED4312) carrying XCV0959-c-Myc show that similar protein amountswere loaded. Note that GroEL is not exclusively cytoplasmic (Vanet & Labigne, 1998; C. Lorenz & D. Buttner, unpublished). Strep-HrcN,HrcN-c-Myc, XCV2918-c-Myc and XCV2569-c-Myc served as lysis controls in strains that contained c-Myc epitope-tagged T2S candidatesubstrates. One representative blot for Strep-HrcN, HrcN-c-Myc, XCV2918-c-Myc and XCV2569-c-Myc is shown. Experiments were repeatedat least four times with similar results. (b) The predicted pectate lyase XCV2569 and the putative proteases XCV2918 and XCV2049-c-Myc arenot detectable in the culture supernatant. Strains 85-10 (wt) and 85-10DxpsEDxcsE (DxpsEDxcsE) carrying XCV2569-c-myc, XCV2918-c-mycand XCV2049-c-myc expression constructs, respectively, were incubated in NYG medium and TE and SN were analysed by immunoblottingusing a c-Myc epitope-specific antibody. (c) In vitro secretion assays with wild-type strains and T2S mutants deleted in both predicted T2Ssecretin-encoding genes. Strains 85-10 (wt) and 85-10DxpsDDxcsD (DxpsDDxcsD) carrying expression constructs encoding XCV3669-c-Myc,XCV0031-c-Myc, XCV0722-c-Myc, XCV0959-c-Myc, XCV1823-c-Myc, XCV3013-c-Myc and XCV4074-c-Myc, respectively, were incubatedin NYG medium and TE and SN were analysed by immunoblotting using a c-Myc epitope-specific antibody. (d) Secretion of PghBxc (Xc1849)and PghAxc (Xc0705) in Xcv is independent of the T2S system. Strains 85-10 (wt) and 85-10DxpsEDxcsED4312 (DxpsEDxcsED4312) carryingexpression constructs encoding PghBxc-c-Myc and PghAxc-c-Myc, respectively, were incubated in NYG medium and TE and SN wereanalysed by immunoblotting using c-Myc epitope-, Strep epitope- and GroEL-specific antibodies. Strep-HrcN was expressed from an ectopicplasmid and served as lysis control. (e) Efficient secretion of XynC-c-Myc depends on XpsE. Strains 85-10 (wt), 85-10DxcsE (DxcsE),85-10DxcsD (DxcsD) and 85-10DxpsE (DxpsE) expressing xynC-c-myc from a corresponding expression construct were incubated in NYGmedium and TE and SN were analysed as described in (c).
NewPhytologist Research 997
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com
sition and could facilitate the assembly of T3S pili that serveas transport channels for effector proteins across the plantcell wall (Belien et al., 2006; Lagaert et al., 2009). The latterhypothesis is supported by the fact that mutation of the Xpssystem leads to reduced translocation of type III effectors butdoes not significantly affect in vitro T3S or the synthesis ofT3S system components in planta. Reduced effector proteintranslocation by xps mutants was not caused by differences inbacterial counts as translocation of effector proteins occurredwithin 6 hpi when the number of wild-type and xpsE dele-tion mutant bacteria was similar. It is therefore tempting tospeculate that the Xps system promotes effector proteintranslocation by secretion of degradative enzymes that couldfacilitate the assembly of extracellular components of theT3S system. However, it remains to be investigated whetheradditional T2S-dependent factors contribute to the translo-cation of type III effectors.
To investigate whether the T2S systems from Xcv secretedegradative enzymes, as was reported for T2S systems fromother plant pathogens, we performed enzyme activity plateassays and tested individual candidates for secretion. Theresults revealed that the Xps system is crucial for the activityof extracellular milk protein-degrading proteases. However,the identity of type II-secreted proteases remains to bedetermined. Genome sequence analysis revealed that Xcvstrain 85-10 encodes > 50 predicted extracellular proteases(Thieme et al., 2005). One obvious candidate protease,
XCV3669, which is encoded adjacent to the xps gene clus-ter, is secreted independently of the T2S systems. Similarlyto XCV3669, secretion of the predicted proteasesXCV0959, XCV0960, XCV3013 and XCV4074 wasdetected in wild-type and T2S mutant strains and thusdepends on alternative secretion systems that are active incomplex NYG medium. As halo formation on milk plateswas abolished in T2S mutants, we assume that the predictedproteases XCV0959, XCV0960, XCV3669, XCV3013 andXCV4074 do not efficiently degrade milk proteins.
T2S-independent secretion was also observed for the pre-dicted cellulase XCV0031, the endo-polygalacturonaseXCV0722 and the glucan 1,4-beta-glucosidase XCV1823.This was unexpected as cellulases, amylases, proteases andpolygalacturonases from Xcc, Xac and Xoo including homo-logs of XCV0722 and XCV3013 were identified as T2Ssubstrates (Ray et al., 2000; Furutani et al., 2004; Chenet al., 2005; Sun et al., 2005; Wang et al., 2008b;Yamazaki et al., 2008; see Tables 2 and 3). Furthermore,we found that two polygalacturonases from Xcc are secretedby Xcv wild-type and T2S mutant strains. Our findingstherefore uncovered remarkable differences in the substratespecificity of T2S systems from different pathovars ofXanthomonas spp.
Interestingly, we observed that the Xps system contrib-utes to extracellular xylanase activity. Xylanases are glycosidehydrolases that degrade xylan, which is the most abundant
Table 3 Known type II secretion (T2S) substrates from Xanthomonas spp.
T2S substrates1 Xanthomonas spp. Secretion in Xcv2
Closesthomolog inXcv Protein Secretion3 References
Lipases ⁄ EsterasesXOO0526 (LipA) Xoo BXO1 na XCV0536 na Rajeshwari et al. (2005)
XylanasesXOO4428 (XynB) Xoo BXO1 XCV4358 na Ray et al. (2000); Rajeshwari et al. (2005)
ProteasesCysP2 Xoo T7174R na XCV3013 T2S-independent Furutani et al. (2004)XAC2831 Xac NA-1 na XCV2993 na Yamazaki et al. (2008)XAC0552 Xac NA-1 na XCV0583 na Yamazaki et al. (2008)XAC0795 Xac NA-1 na XCV0845 na Yamazaki et al. (2008)XAC2853 Xac NA-1 na XCV3013 T2S-independent Yamazaki et al. (2008)
CellulasesXOO4019 (ClsA) Xoo BXO1 na XCV0670 na Jha et al. (2007)
CellobiosidaseXOO4035 (CbsA) Xoo BXO1 na None Jha et al. (2007)
PolygalacturonasesXAC2374 (PglA) Xac NA-1 na XCV2571 Not detectable Yamazaki et al. (2008)XAC0661 (Peh-1) Xac NA-1 na XCV0722 T2S-independent Yamazaki et al. (2008)Xc0705 (PghAxc) Xcc 8004 T2S-independent XCV0722 T2S-independent Wang et al. (2008b)Xc1849 (PghBxc) Xcc 8004 T2S-independent XCV2571 Not detectable Wang et al. (2008b)
1Names of gene products are given in brackets. The accession number of CysP2 from Xoo T7174R is not published. The accession number ofthe homologous protein from Xoo PXO99A is PXO_04730.2For secretion assays in Xcv, genes were cloned into vector pBRM and expressed in Xcv wild-type and T2S mutant strains (see Fig. 7). na, notanalysed.3Secretion of homologous proteins from Xcv was analysed in wild-type and T2S mutant strains (see Fig. 7 and Table 2).
998 Research
NewPhytologist
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com
hemicellulose in the cell wall of monocotyledonous plantsbut also occurs in dicotyledonous plants (Belien et al.,
2006). Xylanase activity was not completely abolished in xcsand xps mutants, suggesting that xylanases can also besecreted by an alternative secretion system. We identifiedthe xylanase XynC (XCV0965) as a substrate of the Xps sys-tem with a role in virulence. Infection studies revealed thatXynC contributes to in planta bacterial multiplication andto disease symptoms. This is in line with the identificationof a type II-secreted xylanase as a virulence factor of Xoo(Ray et al., 2000; Rajeshwari et al., 2005; Sun et al., 2005).Deletion of xynC from Xcv led to a reduction of xylanaseactivity comparable to the effect of xps mutations, suggestingthat T2S-dependent extracellular xylanase activity mainlydepends on XynC. Notably, the genome of Xcv strain85-10 encodes at least six predicted xylanases includingXynA, XynB2 and XynB3 that are homologous to known
wt ΔxynC
22 4 6 8
wtΔxynC
Time post inoculation (d)
Log
(CF
U c
m–2
)
8
7
6
5
4
3
ΔhrcN
10
**
wt ΔxynC
XynC -c-Mycpnat XynC -c-Mycplac
wt ΔxynC wt ΔxynC
-
7 hpi
24 hpi
wt ΔxynC
ΔxpsEΔxcsEΔ4312
(a)
(c)
(d)
(b)
Fig. 8 Functional characterization of the predicted xylanase XynC.(a) XynC contributes to extracellular xylanase activity. Xcv strains85-10 (wt) and 85-10DxynC (DxynC) were incubated on nutrient–yeast–glycerol (NYG) agar plates containing remazol brilliant blue(RBB) xylan. Plates were photographed at 1 d post inoculation (dpi).Extracellular protease, amylase and cellulase activities were notaltered in the absence of xynC, suggesting that XynC specificallycontributes to xylanase activity (data not shown). (b) XynCpromotes disease symptoms. Xcv strains 85-10 (wt), 85-10DxpsEDxcsED4312 (DxpsEDxcsED4312) and 85-10DxynC(DxynC) were inoculated at a bacterial density of 4 · 107 CFU ml)1
into susceptible pepper plants and disease symptoms were photo-graphed 4 dpi. Dashed lines mark the inoculated areas. (c) Analysisof in planta bacterial growth of the xynC deletion mutant. Strains85-10 (closed diamonds, wt), 85-10DxynC (squares, DxynC) and85-10DhrcN (closed circles, DhrcN, T3S mutant) were inoculated atpopulation densities of 104 CFU ml)1 into leaves of susceptible‘Early Cal Wonder’ (ECW) pepper plants and bacterial counts weredetermined over a period of 10 dpi. Values are the mean of threesamples from three different plants. Error bars represent standarddeviations. The asterisks indicate a significant difference withP < 0.05 based on the results of an unpaired Student’s t test. (d)Ectopic expression of xynC leads to increased extracellular xylanaseactivity. Strains 85-10 (wt) and 85-10DxynC (DxynC) carrying theempty vector ()) or expression constructs encoding XynC-c-Mycunder control of the native (pnat) or the lac promoter (plac) asindicated were incubated on NYG agar plates containing RBB xylanand plates were photographed at 7 hpi and 24 hpi as indicated.
0
20
40
60
80
100
85–10 85* 85*ΔhrpX
Rel
ativ
e tr
ansc
ript l
evel
s
120
140
85-10 85* 85*ΔhrpX
Milk
CMC
Xylan
Starch
xpsE xopC
1
2
3
4
5
Rad
ius
of h
alo
(mm
)
6
Milk Xylan
85-1085*85*ΔhrpX
xynCxpsF
(b) (c)
(a)
Fig. 9 Components and substrates of the Xps system aredifferentially regulated by HrpG and HrpX. (a) Quantitativereal-time polymerase chain reaction (qRT-PCR) analysis of xpsE,xpsF, xopC and xynC transcript levels. xpsE, xpsF, xynC and xopC
transcripts were amplified from cDNA derived from Xcv strains85-10, 85* and 85*DhrpX. Transcript amounts were adjusted basedon transcript levels of constitutive 16S rRNA (see the Materials andMethods section). Values represent the percentage of relativeexpression level. Error bars refer to standard deviations derived fromtechnical triplicates. The experiment was repeated once with similarresults. (b) Extracellular protease and xylanase activity is repressedby HrpG. Xcv strains 85-10, 85* and 85*DhrpX were incubated onnutrient–yeast–glycerol (NYG) agar plates containing milk, remazolbrilliant blue (RBB) xylan, starch or CMC (carboxymethyl cellulose)as indicated. Starch and CMC plates were stained with Lugol’ssolution and Congo red, respectively. Photographs were taken at2 d post inoculation (dpi). (c) Measurement of halo sizes. Theradiuses of halos from three different milk and xylan plates weremeasured and mean values are shown in the diagram. Error barsrepresent standard deviations.
NewPhytologist Research 999
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

and predicted xylanases from other plant pathogenic bacte-ria (Thieme et al., 2005). It remains to be investigatedwhether deletion of additional xylanase-encoding genes inXcv leads to a further reduction of bacterial virulence. It wasreported previously that inactivation of single genes for T2Ssubstrates often has a minor effect on bacterial virulencepresumably owing to functional redundancies amongsecreted proteins (Rajeshwari et al., 2005; Jha et al., 2007).
Interestingly, extracellular xylanase activity of Xcv isrepressed by HrpG and HrpX, the key regulators of theT3S system. This was confirmed by qRT-PCR analysis ofxynC transcript levels. Similarly, extracellular protease activ-ity, which also depends on the Xps system, is repressed byHrpG and HrpX (Noel et al., 2001). We thereforeconclude that production of certain T2S substrates fromXcv, including proteases and xylanases, is negatively affectedby HrpG and HrpX. Our observations contrast with thefinding that synthesis of T2S substrates from Xoo and Xcc isactivated by HrpG and HrpX (Furutani et al., 2004; Wanget al., 2008b). Notably, however, HrpG and HrpX fromXcv do not only negatively affect extracellular protease andxylanase activity but also promote expression of xps genes.HrpG ⁄ HrpX-dependent activation of xps gene expressionwas recently also reported for Xac and suggests that T2Sand T3S systems are coregulated (Yamazaki et al., 2008).To our knowledge, our findings provide the first experi-mental evidence for a differential regulation of componentsand substrates of a T2S system. It remains to be investigatedwhether HrpG ⁄ HrpX-dependent suppression of extracellularenzyme activities promotes the bacterial interaction withthe host. It is tempting to speculate that the HrpG ⁄ HrpX-dependent reduction of extracellular protease activityduring T3S prevents degradation of extracellular compo-nents or substrates of the T3S system.
In conclusion, we show that the Xps system contributesto virulence and secretes the xylanase XynC, which isrequired for disease symptoms and in planta bacterialgrowth of Xcv. Intriguingly, our data strongly suggest thatthe virulence function of the Xps system from Xcv might beto facilitate type III-dependent effector protein trans-location. Another key finding is the remarkable differencein regulation and identity of T2S substrates in differentXanthomonas spp., which clearly highlights the importanceof comparative studies of protein secretion systems in differentXanthomonas pathovars.
Acknowledgements
We are grateful to S. Marillonnet for helpful suggestions onthe Golden Gate cloning strategy, to C. Lorenz for generat-ing pBRM and pBRMhrcN, to J. Hausner for constructionof pBRM-P, to C. Kretschmer for sequencing and B.Rosinsky for greenhouse work. Part of this work wassupported by grants from the Deutsche Forschungsge-
meinschaft, the Sonderforschungsbereich SFB 648 ‘MolekulareMechanismen der Informationsverarbeitung in Pflanzen’ toDB and UB, and the Federal Ministry of Education andResearch (BMBF GenoMik+) to UB.
References
Andro T, Chambost JP, Kotoujansky A, Cattaneo J, Bertheau Y, Barras
F, Van Gijsegem F, Coleno A. 1984. Mutants of Erwinia chrysanthemidefective in secretion of pectinase and cellulase. Journal of Bacteriology160: 1199–1203.
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA,
Struhl K. 1996. Current protocols in molecular biology. New York, NY,
USA: John Wiley & Sons, Inc.
Bartetzko V, Sonnewald S, Vogel F, Hartner K, Stadler R, Hammes UZ,
Bornke F. 2009. The Xanthomonas campestris pv vesicatoria type III
effector protein XopJ inhibits protein secretion: evidence for interference
with cell wall-associated defense responses. Molecular Plant–MicrobeInteractions 22: 655–664.
Belien T, Van Campenhout S, Robben J, Volckaert G. 2006. Microbial
endoxylanases: effective weapons to breach the plant cell-wall barrier or,
rather, triggers of plant defense systems? Molecular Plant–MicrobeInteractions 19: 1072–1081.
Bittel P, Robatzek S. 2007. Microbe-associated molecular patterns
(MAMPs) probe plant immunity. Current Opinion in Plant Biology 10:
335–341.
Bonas U, Schulte R, Fenselau S, Minsavage GV, Staskawicz BJ, Stall RE.
1991. Isolation of a gene-cluster from Xanthomonas campestris pv.
vesicatoria that determines pathogenicity and the hypersensitive
response on pepper and tomato. Molecular Plant–Microbe Interactions 4:
81–88.
Bonas U, Stall RE, Staskawicz B. 1989. Genetic and structural
characterization of the avirulence gene avrBs3 from Xanthomonascampestris pv. vesicatoria. Molecular and General Genetics 218: 127–136.
Buttner D, Bonas U. 2002. Getting across-bacterial type III effector
proteins on their way to the plant cell. EMBO Journal 21: 5313–5322.
Buttner D, Bonas U. 2010. Regulation and secretion of Xanthomonasvirulence factors. FEMS Microbiology Reviews 34: 107–133.
Buttner D, Lorenz C, Weber E, Bonas U. 2006. Targeting of two effector
protein classes to the type III secretion system by a HpaC- and HpaB-
dependent protein complex from Xanthomonas campestris pv. vesicatoria.
Molecular Microbiology 59: 513–527.
Buttner D, Nennstiel D, Klusener B, Bonas U. 2002. Functional analysis
of HrpF, a putative type III translocon protein from Xanthomonascampestris pv. vesicatoria. Journal of Bacteriology 184: 2389–2398.
Buttner D, Noel L, Stuttmann J, Bonas U. 2007. Characterization of the
non-conserved hpaB-hrpF region in the hrp pathogenicity island from
Xanthomonas campestris pv. vesicatoria. Molecular Plant–MicrobeInteractions 20: 1063–1074.
Canteros BI. 1990. Diversity of plasmids and plasmid-encoded phenotypictraits in Xanthomonas campestris pv. vesicatoria. PhD thesis, University of
Florida, Gainesville, FL, USA.
Chen Y, Shiue SJ, Huang CW, Chang JL, Chien YL, Hu NT, Chan NL.
2005. Structure and function of the XpsE N-terminal domain, an
essential component of the Xanthomonas campestris type II secretion
system. Journal of Biological Chemistry 280: 42356–42363.
Chisholm ST, Coaker G, Day B, Staskawicz B. 2006. Host–microbe
interactions: shaping the evolution of the plant immune response. Cell124: 803–814.
Cianciotto NP. 2005. Type II secretion: a protein secretion system for all
seasons. Trends in Microbiology 13: 581–588.
Dangl JL, Jones JDG. 2001. Plant pathogens and integrated defence
responses to infection. Nature 411: 826–833.
1000 Research
NewPhytologist
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

Daniels MJ, Barber CE, Turner PC, Sawczyc MK, Byrde RJW, Fielding
AH. 1984. Cloning of genes involved in pathogenicity of Xanthomonascampestris pv. campestris using the broad host range cosmid pLAFR1.
EMBO Journal 3: 3323–3328.
Das A, Rangaraj N, Sonti RV. 2009. Multiple adhesin-like functions of
Xanthomonas oryzae pv. oryzae are involved in promoting leaf
attachment, entry, and virulence on rice. Molecular Plant–MicrobeInteractions 22: 73–85.
Dow JM, Milligan DE, Jaison L, Barber CE, Daniels MJ. 1987. A gene
cluster in Xanthomonas campestris required for pathogenicity controls the
excretion of polygalacturonate lyase and other enzymes. Physiological andMolecular Plant Pathology 31: 261–271.
Engler C, Kandzia R, Marillonnet S. 2008. A one pot, one step,
precision cloning method with high throughput capability. PLoSONE 3: e3647.
Escolar L, Van den Ackerveken G, Pieplow S, Rossier O, Bonas U. 2001.
Type III secretion and in planta recognition of the Xanthomonasavirulence proteins AvrBs1 and AvrBsT. Molecular Plant Pathology 2:
287–296.
Fenselau S, Bonas U. 1995. Sequence and expression analysis of the hrpBpathogenicity operon of Xanthomonas campestris pv. vesicatoria which
encodes eight proteins with similarity to components of the Hrp, Ysc,
Spa, and Fli secretion systems. Molecular Plant–Microbe Interactions 8:
845–854.
Figurski D, Helinski DR. 1979. Replication of an origin-containing
derivative of plasmid RK2 dependent on a plasmid function provided intrans. Proceedings of the National Academy of Sciences, USA 76: 1648–
1652.
Furutani A, Tsuge S, Ohnishi K, Hikichi Y, Oku T, Tsuno K, Inoue Y,
Ochiai H, Kaku H, Kubo Y. 2004. Evidence for HrpXo-dependent
expression of type II secretory proteins in Xanthomonas oryzae pv. oryzae.
Journal of Bacteriology 186: 1374–1380.
Ghosh P. 2004. Process of protein transport by the type III secretion
system. Microbiology and Molecular Biology Reviews 68: 771–795.
Gough CL, Dow JM, Barber CE, Daniels MJ. 1988. Cloning of two
endoglucanase genes of Xanthomonas campestris pv. campestris: analysis of
the role of the endoglucanase in pathogenesis. Molecular Plant-MicrobeInteractions 1: 275–281.
Gurlebeck D, Thieme F, Bonas U. 2006. Type III effector proteins from
the plant pathogen Xanthomonas and their role in the interaction with
the host plant. Journal of Plant Physiology 163: 233–255.
He YQ, Zhang L, Jiang BL, Zhang ZC, Xu RQ, Tang DJ, Qin J, Jiang
W, Zhang X, Liao J et al. 2007. Comparative and functional genomics
reveals genetic diversity and determinants of host specificity among
reference strains and a large collection of Chinese isolates of the
phytopathogen Xanthomonas campestris pv. campestris. Genome Biology 8:
R218.
Hu NT, Hung MN, Chiou SJ, Tang F, Chiang DC, Huang HY, Wu CY.
1992. Cloning and characterization of a gene required for the secretion
of extracellular enzymes across the outer membrane by Xanthomonascampestris pv. campestris. Journal of Bacteriology 174: 2679–2687.
Hu NT, Hung MN, Liao CT, Lin MH. 1995. Subcellular location of
XpsD, a protein required for extracellular protein secretion by
Xanthomonas campestris pv. campestris. Microbiology 141: 1395–1406.
Hu J, Qian W, He C. 2007. The Xanthomonas oryzae pv. oryzae eglXoBendoglucanase gene is required for virulence to rice. FEMS MicrobiologyLetters 269: 273–279.
Huguet E, Hahn K, Wengelnik K, Bonas U. 1998. hpaA mutants of
Xanthomonas campestris pv. vesicatoria are affected in pathogenicity but
retain the ability to induce host-specific hypersensitive reaction.
Molecular Microbiology 29: 1379–1390.
Jha G, Rajeshwari R, Sonti R. 2005. Bacterial type two secretion system
secreted proteins: double-edged swords for plant pathogens. MolecularPlant-Microbe Interactions 18: 891–898.
Jha G, Rajeshwari R, Sonti RV. 2007. Functional interplay between two
Xanthomonas oryzae pv. oryzae secretion systems in modulating virulence
on rice. Molecular Plant-Microbe Interactions 20: 31–40.
Jones JD, Dangl JL. 2006. The plant immune system. Nature 444: 323–
329.
Jones JB, Lacy GH, Bouzar H, Stall RE, Schaad NW. 2004.
Reclassification of the xanthomonads associated with bacterial spot
disease of tomato and pepper. Systematic and Applied Microbiology 27:
755–762.
Keshavarzi M, Soylu S, Brown I, Bonas U, Nicole M, Rossiter J,
Mansfield J. 2004. Basal defenses induced in pepper by
lipopolysaccharides are suppressed by Xanthomonas campestris pv.
vesicatoria. Molecular Plant-Microbe Interactions 17: 805–815.
Knoop V, Staskawicz B, Bonas U. 1991. Expression of the avirulence gene
avrBs3 from Xanthomonas campestris pv. vesicatoria is not under the
control of hrp genes and is independent of plant factors. Journal ofBacteriology 173: 7142–7150.
Kovach ME, Elzer PH, Hill DS, Robertson MA, Farris MA, Roop RM II,
Peterson KM. 1995. Four new derivatives of the broad-host-range
cloning vector pBBR1MCS, carrying different antibiotic-resistance
cassettes. Gene 166: 175–176.
Lagaert S, Belien T, Volckaert G 2009. Plant cell walls: protecting the
barrier from degradation by microbial enzymes. Seminars in Cell andDevelopmental Biology 20: 1064–1073.
Laia ML, Moreira LM, Dezajacomo J, Brigati JB, Ferreira CB, Ferro MI,
Silva AC, Ferro JA, Oliveira JC. 2009. New genes of Xanthomonas citrisubsp. citri involved in pathogenesis and adaptation revealed by a
transposon-based mutant library. BMC Microbiology 9: 12.
Liu H, Zhang S, Schell MA, Denny TP. 2005. Pyramiding unmarked
deletions in Ralstonia solanacearum shows that secreted proteins in
addition to plant cell-wall degrading enzymes contribute to virulence.
Molecular Plant-Microbe Interactions 18: 1296–1305.
Lorenz C, Buttner D. 2009. Functional characterization of the type III
secretion ATPase HrcN from the plant pathogen Xanthomonascampestris pv. vesicatoria. Journal of Bacteriology 191: 1414–1428.
Lorenz C, Kirchner O, Egler M, Stuttmann J, Bonas U, Buttner D.
2008. HpaA from Xanthomonas is a regulator of type III secretion.
Molecular Microbiology 69: 344–360.
Lu H, Patil P, Van Sluys MA, White FF, Ryan RP, Dow JM,
Rabinowicz P, Salzberg SL, Leach JE, Sonti R et al. 2008.
Acquisition and evolution of plant pathogenesis-associated gene
clusters and candidate determinants of tissue-specificity in
Xanthomonas. PLoS ONE 3: e3828.
Menard R, Sansonetti PJ, Parsot C. 1993. Nonpolar mutagenesis of
the ipa genes defines IpaB, IpaC, and IpaD as effectors of Shigellaflexneri entry into epithelial cells. Journal of Bacteriology 175: 5899–
5906.
Metz M, Dahlbeck D, Morales CQ, Al Sady B, Clark ET, Staskawicz BJ.
2005. The conserved Xanthomonas campestris pv. vesicatoria effector
protein XopX is a virulence factor and suppresses host defense in
Nicotiana benthamiana. Plant Journal 41: 801–814.
Minsavage GV, Dahlbeck D, Whalen MC, Kearny B, Bonas U,
Staskawicz BJ, Stall RE. 1990. Gene-for-gene relationships specifying
disease resistance in Xanthomonas campestris pv. vesicatoria–pepper
interactions. Molecular Plant–Microbe Interactions 3: 41–47.
Murillo J, Shen H, Gerhold D, Sharma A, Cooksey DA, Keen NT. 1994.
Characterization of pPT23B, the plasmid involved in syringolide
production by Pseudomonas syringae pv. tomato PT23. Plasmid 31: 275–
287.
Noel L, Thieme F, Gabler J, Buttner D, Bonas U. 2003. XopC and XopJ,
two novel type III effector proteins from Xanthomonas campestris pv.
vesicatoria. Journal of Bacteriology 185: 7092–7102.
Noel L, Thieme F, Nennstiel D, Bonas U. 2001. cDNA-AFLP analysis
unravels a genome-wide hrpG-regulon in the plant pathogen
NewPhytologist Research 1001
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com

Xanthomonas campestris pv. vesicatoria. Molecular Microbiology 41:
1271–1281.
Preston GM, Studholme DJ, Caldelari I. 2005. Profiling the secretomes
of plant pathogenic Proteobacteria. FEMS Microbiology Reviews 29:
331–360.
Qian W, Jia Y, Ren SX, He YQ, Feng JX, Lu LF, Sun Q, Ying G, Tang
DJ, Tang H et al. 2005. Comparative and functional genomic analyses
of the pathogenicity of phytopathogen Xanthomonas campestris pv.
campestris. Genome Research 15: 757–767.
Rajeshwari R, Jha G, Sonti RV. 2005. Role of an in planta-expressed
xylanase of Xanthomonas oryzae pv. oryzae in promoting virulence on
rice. Molecular Plant–Microbe Interactions 18: 830–837.
Ray SK, Rajeshwari R, Sonti RV. 2000. Mutants of Xanthomonas oryzaepv. oryzae deficient in general secretory pathway are virulence deficient
and unable to secrete xylanase. Molecular Plant–Microbe Interactions 13:
394–401.
Ronald PC, Staskawicz BJ. 1988. The avirulence gene avrBs1 from
Xanthomonas campestris pv. vesicatoria encodes a 50-kDa protein.
Molecular Plant–Microbe Interactions 1: 191–198.
Rossier O, Van den Ackerveken G, Bonas U. 2000. HrpB2 and HrpF
from Xanthomonas are type III-secreted proteins and essential for
pathogenicity and recognition by the host plant. Molecular Microbiology38: 828–838.
Rossier O, Wengelnik K, Hahn K, Bonas U. 1999. The Xanthomonas Hrp
type III system secretes proteins from plant and mammalian pathogens.
Proceedings of the National Academy of Sciences, USA 96: 9368–9373.
Ryan CA, Farmer EE. 1991. Oligosaccharide signals in plants: a current
assessment. Annual Review of Plant Physiology and Plant MolecularBiology 42: 651–674.
Sandkvist M. 2001. Biology of type II secretion. Molecular Microbiology40: 271–283.
Schafer A, Tauch A, Jager W, Kalinowski J, Thierbach G, Puhler A.
1994. Small mobilizable multi-purpose cloning vectors derived from the
Escherichia coli plasmids pK18 and pK19: selection of defined deletions
in the chromosome of Corynebacterium glutamicum. Gene 145: 69–73.
Schulte R, Bonas U. 1992. Expression of the Xanthomonas campestris pv.
vesicatoria hrp gene cluster, which determines pathogenicity and
hypersensitivity on pepper and tomato, is plant inducible. Journal ofBacteriology 174: 815–823.
Shiue SJ, Chien IL, Chan NL, Leu WM, Hu NT. 2007. Mutation of a
key residue in the type II secretion system ATPase uncouples ATP
hydrolysis from protein translocation. Molecular Microbiology 65: 401–
412.
Shiue SJ, Kao KM, Leu WM, Chen LY, Chan NL, Hu NT. 2006. XpsE
oligomerization triggered by ATP binding, not hydrolysis, leads to its
association with XpsL. EMBO Journal 25: 1426–1435.
Sun QH, Hu J, Huang GX, Ge C, Fang RX, He CZ. 2005. Type-II secretion
pathway structural gene xpsE, xylanase- and cellulase secretion and
virulence in Xanthomonas oryzae pv. oryzae. Plant Pathology 54: 15–21.
Szurek B, Rossier O, Hause G, Bonas U. 2002. Type III-dependent
translocation of the Xanthomonas AvrBs3 protein into the plant cell.
Molecular Microbiology 46: 13–23.
Thieme F, Koebnik R, Bekel T, Berger C, Boch J, Buttner D, Caldana C,
Gaigalat L, Goesmann A, Kay S et al. 2005. Insights into genome
plasticity and pathogenicity of the plant pathogenic bacterium
Xanthomonas campestris pv. vesicatoria revealed by the complete genome
sequence. Journal of Bacteriology 187: 7254–7266.
Toth IK, Bell KS, Holeva M, Birch PR. 2003. Soft rot erwiniae: from
genes to genomes. Molecular Plant Pathology 4: 17–30.
Vanet A, Labigne A. 1998. Evidence for specific secretion rather than
autolysis in the release of some Helicobacter pylori proteins. Infection andImmunity 66: 1023–1027.
Vieira J, Messing J. 1987. Production of single-stranded plasmid DNA.
Methods of Enzymology 153: 3–11.
Vroemen S, Heldens J, Boyd C, Henrissat B, Keen NT. 1995. Cloning
and characterization of the bgxA gene from Erwinia chrysanthemi D1
which encodes a beta-glucosidase ⁄ xylosidase enzyme. Molecular andGeneral Genetics 246: 465–477.
Wang JC, So BH, Kim JH, Park YJ, Lee B-M, Kang HW. 2008a. Genome-
wide identification of pathogenicity genes in Xanthomonas oryzae pv.
oryzae by transposon mutagenesis. Plant Pathology 57: 1136–1145.
Wang L, Rong W, He C. 2008b. Two Xanthomonas extracellular
polygalacturonases, PghAxc and PghBxc, are regulated by type III
secretion regulators HrpX and HrpG and are required for virulence.
Molecular Plant–Microbe Interactions 21: 555–563.
Weber E, Ojanen-Reuhs T, Huguet E, Hause G, Romantschuk M,
Korhonen TK, Bonas U, Koebnik R. 2005. The type III-dependent
Hrp pilus is required for productive interaction of Xanthomonascampestris pv. vesicatoria with pepper host plants. Journal of Bacteriology187: 2458–2468.
Wengelnik K, Bonas U. 1996. HrpXv, an AraC-type regulator, activates
expression of five of the six loci in the hrp cluster of Xanthomonascampestris pv. vesicatoria. Journal of Bacteriology 178: 3462–
3469.
Wengelnik K, Marie C, Russel M, Bonas U. 1996a. Expression and
localization of HrpA1, a protein of Xanthomonas campestris pv.
vesicatoria essential for pathogenicity and induction of the
hypersensitive reaction. Journal of Bacteriology 178: 1061–1069.
Wengelnik K, Rossier O, Bonas U. 1999. Mutations in the regulatory
gene hrpG of Xanthomonas campestris pv. vesicatoria result in
constitutive expression of all hrp genes. Journal of Bacteriology 181:
6828–6831.
Wengelnik K, Van den Ackerveken G, Bonas U. 1996b. HrpG, a key hrpregulatory protein of Xanthomonas campestris pv. vesicatoria is
homologous to two-component response regulators. Molecular Plant–Microbe Interactions 9: 704–712.
Yamazaki A, Hirata H, Tsuyumu S. 2008. HrpG regulates type II
secretory proteins in Xanthomonas axonopodis pv. citri. Journal of GeneralPlant Pathology 74: 138–150.
Yang B, Zhu W, Johnson LB, White FF. 2000. The virulence factor
AvrXa7 of Xanthomonas oryzae pv. oryzae is a type III secretion
pathway-dependent nuclear-localized double-stranded DNA-binding
protein. Proceedings of the National Academy of Sciences, USA 97:
9807–9812.
Zhou JM, Chai J. 2008. Plant pathogenic bacterial type III effectors
subdue host responses. Current Opinion in Microbiology 11: 179–185.
Supporting Information
Additional supporting information may be found in theonline version of this article.
Fig. S1 Schematic representation of the Golden Gate-compatible expression vectors pBRM and pBRM-P.
Fig. S2 Deletion of the complete xcs gene cluster does notsignificantly affect bacterial virulence.
Table S1 Primers used in this study.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting informationsupplied by the authors. Any queries (other than missingmaterial) should be directed to the New Phytologist CentralOffice.
1002 Research
NewPhytologist
� The Authors (2010)
Journal compilation � New Phytologist Trust (2010)
New Phytologist (2010) 187: 983–1002
www.newphytologist.com





























