Embed Size (px)
Citation preview

Functional Characterization of Human Myosin-18A and ItsInteraction with F-actin and GOLPH3*
Received for publication, June 28, 2013, and in revised form, August 14, 2013 Published, JBC Papers in Press, August 29, 2013, DOI 10.1074/jbc.M113.497180
Manuel H. Taft‡1, Elmar Behrmann§2, Lena-Christin Munske-Weidemann‡, Claudia Thiel‡, Stefan Raunser§,and Dietmar J. Manstein‡
From the ‡Institute for Biophysical Chemistry, Hannover Medical School, OE 4350, Carl-Neuberg-Strasse 1, 30625 Hannover,Germany and the §Department of Physical Biochemistry, Max-Planck-Institute of Molecular Physiology,44227 Dortmund, Germany
Background: Class-18A myosins share a unique N-terminal extension comprising a PDZ module and a KE-rich region.Results:Humanmyosin-18A binds F-actin via its motor domain in a nucleotide-dependentmanner and via the KE-rich region,modulated by direct interaction between the PDZ module and GOLPH3.Conclusion:Myosin-18A binds F-actin and recruits interaction partners to the cytoskeleton.Significance: This work establishes a molecular basis for myosin-18A mediated membrane-cytoskeleton interplay.
Molecular motors of the myosin superfamily share a genericmotor domain region. They commonly bind actin in an ATP-sensitive manner, exhibit actin-activated ATPase activity, andgenerate force andmovement in this interaction. Class-18myo-sins form heavy chain dimers and contain protein interactiondomains located at their uniqueN-terminal extension. Here, wecharacterized human myosin-18A molecular function in theinteraction with nucleotides, F-actin, and its putative bindingpartner, the Golgi-associated phosphoprotein GOLPH3. Weshow thatmyosin-18A comprises two actin binding sites. One islocated in the KE-rich region at the start of the N-terminalextension and appears to mediate ATP-independent binding toF-actin. The second actin-binding site resides in the genericmotor domain and is regulated by nucleotide binding in theabsence of intrinsic ATP hydrolysis competence. This coremotor domaindisplays its highest actin affinity in theADPstate.Electron micrographs of myosin-18A motor domain-decoratedF-actin filaments show a periodic binding pattern independentof the nucleotide state.We show that the PDZmodulemediatesdirect binding of myosin-18A to GOLPH3, and this interactionin turnmodulates the actin bindingproperties of theN-terminalextension. Thus, myosin-18A can act as an actin cross-linkerwith multiple regulatory modulators that targets interactingproteins or complexes to the actin-based cytoskeleton.
Myosins constitute a large superfamily of molecular motorsthat use the chemical energy provided by ATP hydrolysis tocyclically interact with filamentous F-actin and generate forceand movement (1). All myosins share a generic motor domain
that harbors the binding sites for ATP and F-actin. Based onsequence alignments of the motor domain, myosins can begrouped into 35 classes (2). In humans, 40 genes are found thatencode for myosins from 13 of these classes. The moleculardetails of themechanochemical transduction of energy bymyo-sins fromdifferent classes have been unraveledwith great accu-racy. Nevertheless, most myosins have not been characterizedin depth, in particular members of the myosin family, such asclass-18 myosins, which show distinct structural features set-ting them apart. Class-18 myosins are found in various species,from vertebrates to arthropods. They contain protein interac-tion domains that are located at their N terminus outside themotor domain (3). Like the founding member of this myosinclass,MysPDZ (now termedmousemyosin-18A), humanmyo-sin-18A comprises a region rich in lysine and glutamate resi-dues (KE) and a PDZ3module in its N-terminal extension. Thisdomain is followed by a genericmotor domain with an adjacentneck domain that can bind essential and regulatory light chains(4). The tail domain contains long stretches of coiled-coils thatsupport heavy chain dimerization (5). The molecular mass ofthe protein varies between 180 kDa for the shortest isoform,which lacks the N-terminal extension (myosin-18A�), and 233kDa for the longest isoform, termed myosin-18A�. A recentstudy identified the gene encoding human myosin-18A to bealternatively spliced in non-small cell lung cancer, leading toin-frame variations in the protein sequence (6). Furthermore,the gene was identified as a partner in the three-way chromo-somal translocation of stem cell leukemia-lymphoma syn-drome (7) and forms the fusion gene MYO18A-PDGFRB ineosinophilia-associated atypical myeloproliferative neoplasms(8). A three-way translocation of the highly promiscuous onco-geneMLL, a histonemethyltransferase, and the reciprocal part-ner geneMYO18Awas described in acutemyeloid leukemia (9).These studies suggest that functional myosin-18A protein is
* This work was supported by Deutsche Forschungsgemeinschaft Grants MA1081/19 –1 (to D. J. M.) and RA 1781/1–1 (to S. R.), Fonds der ChemischenIndustrie Grant 684052 (to E. B.), and the Max Planck Society (to S. R. andE. B.).
1 To whom correspondence should be addressed: Institut für Biophysikalis-che Chemie, Medizinische Hochschule Hannover, Carl-Neuberg-Str. 1,30625 Hannover, Germany. Tel.: 49-511-5328657; Fax: 49-511-5322909;E-mail: [email protected].
2 Present address: Institute of Medical Physics and Biophysics, Charité, Univer-sitätsmedizin Berlin, 10117 Berlin, Germany.
3 The abbreviations used are: PDZ, PSD-95/Discs-Large/ZO-1; TRITC, tetra-methylrhodamine isothiocyanate; M18A-MD, myosin-18A motor domain;mant, 2�/3�-O-(N-methyl-anthraniloyl); SH2 and SH3, Src homology 2 and3, respectively; TCEP, tris(2-carboxyethyl)phosphine; CM-loop, cardiomy-opathy loop.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 288, NO. 42, pp. 30029 –30041, October 18, 2013© 2013 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30029
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

required for the normal regulation of the cell cycle and thesuppression of key processes involved in cancer progression.Up to now, information on the biochemical properties of
class-18 myosins is scarce and in parts controversial. The func-tion of the unique N-terminal extension is only poorly under-stood (10, 11). In a recent study,Drosophilamyosin-18 has beenfound to be an actin-binding protein that does not bind nucle-otide, has no ATPase activity, and cannot actively translocateover actin filaments (12). A current publication of the samegroup on functional features of mouse myosin-18A reportsactin and nucleotide binding properties but no significantATPase activity and suggests that this myosin is not a tradi-tional motor (4). Moreover, the amino acid sequence of activesite elements of the myosin-18 motor domain exhibits changesin highly conserved regions, which can preventmyosin-18 fromproductively interacting with ATP in the same way as othermyosins.Furthermore, it has been shown that the N-terminal exten-
sion of humanmyosin-18A has an ATP-insensitive actin-bind-ing site outside the PDZ module (5). It was suggested that themotor domain of human myosin-18A does not bind to actin,because YFP-tagged motor domain constructs obtained fromcell lysates do not cosediment with actin. This observation is incontrast to the studies on Drosophila myosin-18 and mousemyosin-18A that attribute actin binding properties to themotor domain.Recently, it has been shown that myosin-18A� is a novel
binding partner of the PAK2��PIX�GIT1 complex (13). Thissuggests that myosin-18A may play an important role in regu-lating epithelial cell migration. The Rac/Cdc42-binding kinaseMRCK (myotonic dystrophy kinase-related Cdc42-bindingkinase) has been shown to associate with the myosin-18A�PDZ domain via a linker protein, LRAP35a (14). The resultingphosphorylation of the non-muscle RLC2A (myosin-2A regu-latory light chain) suggests an association ofmyosin-18A�withRLC2A. This hypothesis is supported by the fact that in vitromouse myosin-18A binds essential and regulatory light chainsvia its neck region (4). The tripartite MRCK�LRAP35a�myosin-18A� complex localizes to lamellar actomyosin bundles, wherenon-muscle myosin-2A drives the retrograde flow (14, 15).Therefore, myosin-18A can play a role the regulation and orga-nization of the actin cytoskeleton within lamellipodia.Additionally, Dippold et al. (16) have shown that GOLPH3
binds tomyosin-18A and connects theGolgi apparatus to F-ac-tin to provide a tensile force required for efficient tubule andvesicle formation. However, this function would presumablyimplicate active motor properties for myosin-18A.Here,we show that humanmyosin-18Acontains twodistinct
actin binding sites per heavy chain (four per dimer), one ofwhich is regulated by nucleotide binding and is capable of tar-geting interacting proteins to the actin cytoskeleton, where itcan function as an efficient and adjustable actin cross-linker.The PDZmodule is shown tomediate direct binding ofmyosin-18A to GOLPH3, and this interaction modulates the actinbinding properties of the unique N-terminal extension ofmyosin-18A.
EXPERIMENTAL PROCEDURES
Reagents—Standard chemicals, TRITC-phalloidin, and anti-FLAG antibody were purchased from Sigma-Aldrich; restric-tion enzymes, polymerases and DNA-modifying enzymes werepurchased fromMBI-Fermentas and Roche Applied Science.Plasmid Construction—The full-length cDNA clone
IRATp790A0771D (imaGenes GmbH, Berlin; GenBankTM
entry BC039612.1) of human myosin-18A was used as a tem-plate for the PCR amplification of DNA fragments encoding forthe constructs used in this study. Sequences encoding for theN-terminal subdomainsKE(aminoacids1–219;upstreamprimer,GCGGATCCATGGCTAATGCTCCCTCCTGC; downstreamprimer, GCTCGAGCCGGAGGGTAGGTGGGGGCAG), PDZ(amino acids 220–311; upstream primer, GCGGATCCGAGCT-GGAGCTGCAACGACGG; downstream primer, GCTCGAGT-GGAATGGGCTGCACCTTGAG), and KEPDZ (amino acids1–398; upstreamprimer as for KE; downstreamprimer, GCTCG-AGCTTCTCAACGTCATCCTCATCC)were amplified by PCR,digested with BamHI and XhoI, and subcloned into the plasmidpET23a(�) (Novagen) for the expression of C-terminal His-tagged proteins.The sequence coding for humanGOLPH3was amplified using
the full-length cDNA clone IRAUp969C1265D (imaGenesGmbH, Berlin; GenBankTM entry BC012123.1) as a templatefor PCR and subcloned into the plasmids pET23a(�) (upstreamprimer, GCGGATCCATGACCTCGCTGACCCAGCGC-AGC; downstream primer, GCTCGAGCTTGGTGAACGCC-GCCACCACC) and pGEX-6P2 (upstream primer as forpET23a(�); downstream primer, GCTCGAGTTACTTGGT-GAACGCCGCCACCACC), respectively.For the generation of human myosin-18A motor domain
constructs, the DNA sequences encoding for amino acids399–1185 (M18A-MD; upstream primer, GCGGATCCATG-GCTAATGCTCCCTCCTGC; downstream primer, GAAGC-TTTTATTTATCATCATCATCTTTATAATCTGTACATA-ATGCATCCCGCTGCTCCTCTAGACGTGC) or 220–1185(PDZ-M18A-MD; upstream primer, GCGGATCCGAGCTG-GAGCTGCAACGACGGCCC; downstream primer as forM18A-MD) were amplified by PCR, digested with BamHI andHindIII, and subcloned into the plasmid pFastBacDual (Invit-rogen). The downstream primers introduce C-terminal FLAGtags (DYKDDDDK) to facilitate purification. All plasmids wereconfirmed by sequencing.Protein Production and Purification—Plasmids for the pro-
duction of the N-terminal subdomains of Homo sapiens myo-sin-18A and theH. sapiensGOLPH3 protein were transformedinto Escherichia coli Rosetta pLys-S cells (Merck). Cells weregrown at 30 °C in LB medium (10 g of peptone, 10 g of yeastextract, and 5 g of NaCl per liter, pH 7.0), induced with 1 mM
isopropyl 1-thio-�-D-galactopyranoside atA600 � 0.6, grown at21 or 30 °C for 3–16 h, and harvested by centrifugation (4 °C,4000 � g).For the purification of His-tagged proteins, the cells were
resuspended in buffer (50mMHEPES, pH 8.0, 300mMNaCl, 10mM imidazole, Complete inhibitor mixture (Roche), 1 mg/mllysozyme, 1000 units of benzonase, 12mMMgCl2, 2 mMATP, 3mM benzamidine, 1 mM TCEP, 0.02% NaN3). After sonication
Biochemical Characterization of Human Myosin-18A
30030 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

and the addition of 1%Triton X-100, cell lysis was performed at4 °C for 30 min. Following centrifugation at 20,000 � g at 4 °C(Avanti J-30I, Beckmann Coulter), the supernatant was appliedto a nickel-nitrilotriacetic acid column (Qiagen, Hilden, Ger-many). The resin was washed with buffer (50 mM HEPES, pH8.0, 300 mM NaCl, 20 mM imidazole, 3 mM benzamidine, 1 mM
TCEP, 0.02% NaN3), and the bound protein was eluted with alinear gradient of this buffer with 500 mM imidazole. The pro-tein-containing fractions were dialyzed against storage buffer(50mMHEPES, pH8.0, 300mMNaCl, 3mMbenzamidine, 1mM
TCEP, 0.02% NaN3), concentrated, and applied to a gel filtra-tion column (HiLoad 16/600 Superdex 200 pg; GEHealthcare).The eluted pure protein was supplemented with 3% sucrose,flash-frozen in liquid nitrogen, and stored at �80 °C.The GST fusion construct of GOLPH3 was expressed in
E. coli Rosetta pLys-S cells. Cells were grown at 30 °C in LBmedium, inducedwith 1mM isopropyl 1-thio-�-D-galactopyra-noside at A600 � 0.6, grown at 30 °C for 3 h, and harvested bycentrifugation. Cell lysis was performed at 4 °C for 30 min inbuffer (50mMTris-HCl, pH 7.5, 300mMNaCl, 12mMMgCl2, 1mM benzamidine, Complete inhibitor mixture, 1 mg/mllysozyme, 1000 units of benzonase, 2 mM ATP, 1 mM PMSF, 1mM TCEP) and lysed with Triton X-100 (1% final concentra-tion). After centrifugation at 20,000� g at 4 °C, the supernatantwas applied to a glutathione-Sepharose column and washedwith buffer A (50 mM Tris-HCl, pH 7.5, 300 mM NaCl, 2 mM
MgCl2, 1 mM benzamidine, 1 mM TCEP). A linear gradient ofbuffer A and buffer B (buffer A containing 10 mM reduced glu-tathione) eluted the protein-containing fractions. GST-fusedPreScission protease was added to the protein solution thatsubsequently was dialyzed against storage buffer (50 mM Tris,pH 7.5, 200 mM NaCl, 1 mM EDTA, 1 mM benzamidine, 1 mM
TCEP). Cleaved GST and PreScission protease were removedby passing the protein solution over a glutathione-Sepharosecolumn, and the pure GOLPH3 protein was concentrated,snap-frozen in liquid nitrogen, and stored at �80 °C.The myosin-18A motor domain constructs (M18A-MD and
PDZ-M18A-MD) were overproduced in the baculovirus/Sf9system. The corresponding transfer vector was transformed inDH10Bac E. coli cells to generate recombinant bacmid, whichwas isolated and transfected in Sf9 insect cells usingCellfectin II(Invitrogen). Recombinant baculovirus was produced asdescribed by the manufacturer. Sf9 cells were infected withrecombinant baculovirus, collected 66 h postinfection, andstored at �80 °C. For purification, cells were lysed in buffer (50mMHEPES (pH 7.3), 300 mM KCl, 3 mMMgCl2, 0.1 mM EGTA,2 mM ATP, 5 mM �-mercaptoethanol, 5 mM benzamidine,Complete inhibitor mixture (Roche Applied Science)) by soni-cation and incubation at 4 °C for 30 min. The lysate was ultra-centrifuged (138,000 � g, 1 h), and the supernatant was appliedto �FLAG-M2 affinity gel (Sigma-Aldrich) and rotated for 2 hat 4 °C to ensure binding. The resin was transferred to a columnand washed with ATP buffer (50 mM HEPES (pH 7.5), 300 mM
KCl, 0.5 mM ATP, 0.1 mM EGTA, 3 mMMgCl2, 3 mM benzami-dine, 0.2% Triton X-100), wash buffer 1 (50 mM HEPES (pH7.5), 300 mM KCl, 0.1 mM EGTA, 3 mM MgCl2, 3 mM benzami-dine), and wash buffer 2 (50 mM HEPES (pH 7.5), 600 mM KCl,0.1 mM EGTA, 3 mM MgCl2, 3 mM benzamidine). The protein
was eluted with wash buffer 1 containing 0.1mg/ml FLAG pep-tide anddialyzed against storage buffer (50mMHEPES (pH7.5),300 mM KCl, 0.5 mM EDTA, 0.2 mM EGTA, 1 mMMgCl2, 1 mM
benzamidine, 1 mM DTT, 3% trehalose). The pure protein wasconcentrated, flash-frozen in liquid nitrogen, and stored at�80 °C.Rabbit skeletalmuscle actinwas purified as described byLeh-
rer and Kerwar (17). For selected experiments, F-actin was alsostabilized by the addition of equimolar concentrations of phal-loidin, because a perturbation of the interaction has beendescribed for some unconventional myosins (18). In all actininteraction assays, no significant differences were observedwhen using phalloidin-stabilized instead of non-treated actinfilaments.Kinetic Measurements—ATPase activities were measured at
25 °C with the NADH-coupled assay as described previously(19). Transient kinetic experiments were performed at 20 °Cwith either a Hi-tech Scientific SF-61 DX single mixingstopped-flow system (TgK Scientific Ltd., Bradford on Avon,UK) or an Applied Photophysics PiStar 180 instrument in assaybuffer (20mMMOPS (pH 7.0), 100mMKCl, 5mMMgCl2, 1mM
DTT) using procedures and kinetic models described previ-ously (Scheme 1) (20, 21). Error bars represent S.E. Mant-nu-cleotides (Jena Bioscience) were excited at 365 nmand detectedafter passing through a KV 389-nm cut-off filter.F-Actin Cosedimentation Assays—The interaction of puri-
fied myosin-18A subdomains with F-actin was assayed bycosedimentation. 0.2–2 �M protein was incubated on ice for 15min in assay buffer (20 mMMOPS (pH 7.0), 100 mM KCl, 5 mM
MgCl2, 1 mM TCEP) with varying concentrations of F-actin inthe presence or absence of nucleotides. The absence ofMg2�ATP or Mg2�ADP was guaranteed by the addition of 5mM EDTA and 1 unit/ml apyrase. The samples were subjectedto ultracentrifugation for 20 min at 170,000 � g in a BeckmanTLA-120.1 rotor (4 °C unless otherwise specified). The pelletswere resuspended in assay buffer to the same volume as thesupernatants, and both fractions were resolved by SDS-PAGE.Protein content in the supernatants and pellets was quantifiedby densitometric analysis of Coomassie Blue-stained gels usingthe program ImageJ (National Institutes of Health, Bethesda,MD). To determine actin affinities, the fraction of bound pro-tein in the pellet relative to the total protein contentwas plottedas a function of actin concentration, and a hyperbolic functionwas fitted to the data. In cases where the utilized myosin con-struct concentration exceeds the affinity, a standard quadraticequation was used to analyze the actin binding data as
SCHEME 1. Interaction scheme for actin and nucleotide binding of myo-sin. A, actin; M, myosin; N, nucleotide (T, ATP; D, ADP). For the equilibriumbinding constants, a notation is used that distinguishes between constants inthe absence and presence of actin by using italic type (kx, Kx) and boldfacetype (kx, Kx), respectively.
Biochemical Characterization of Human Myosin-18A
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30031
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

described previously (22–25). Averaged data of replicate bind-ing experiments using three different preparations are given inthe figures. Error bars represent S.E. All data analysis was per-formed with Origin 7.0 (Originlab).Differential Static Light Scattering—Protein stability was
assayed by following protein aggregation associated with heatdenaturation (26). To do so, protein solutions were diluted to aconcentration of 0.2mg/ml in a buffer containing 20mMMOPS(pH 7.0), 500 mM KCl, 5 mM MgCl2, 1 mM TCEP and heated insteps of 0.6 °C. Static light scatteringwasmeasured at each tem-perature step, and the resulting transition curve was fitted bya sigmoidal function. The transition midpoint gives a meas-ure for the aggregation temperature induced by proteindenaturation.Microscale Thermophoresis—Protein interaction studies
usingmicroscale thermophoresis were performed according toDuhr, Braun, and co-workers (27, 28). Protein was labeledaccording to NanoTemper using the MonolithTM NT.115 pro-tein labeling kit RED-NHS (amine-reactive). Experiments wereperformed using standard capillaries in the NanoTemperMonolithTM NT instrument for red dye fluorescence in buffer(25 mMHEPES (pH 7.3), 25 mM KCl, 5 mMMgCl2, 1 mMDTT).For high salt conditions, 25 mM KCl was replaced by 300 mM
NaCl. Error bars represent S.E.Negative Stain Electron Microscopy—Electron micrographs
of negatively stained complexes of F-actin decorated with themotor domain of myosin-18A were obtained as follows:H. sapiensmyosin-18A-MDwas incubated with F-actin in EM-buffer (20 mM HEPES (pH 7.0), 10 mM KCl, 5 mM MgCl2;optional: 5 mM ADP or ATP), adsorbed to freshly glow-dis-charged carbon-coated grids at a concentration of 0.1 mg/ml(1.1 �M M18A-MD and equimolar F-actin), and negativelystained with 0.75% uranyl formate. The specimen was visual-ized with a Jeol JEM-1400 transmission electron microscope(Jeol, Tokyo, Japan) at 120 kV.Bioinformatics—The alignment of the motor domain
sequence of Dictyostelium discoideum myosin-2 (amino acids1–765) with human (amino acids 1–1185) and mouse (aminoacids 1–1180) myosin-18A and Drosophila melanogastermyo-sin-18 (amino acids 1–1319) was performed with the ClustalWalgorithm as implemented in MegAlign 5.07 in the DNAStar
software package. The homology model of the human myosin-18A motor domain (residues 399–1185 of myosin-18A�) wasprepared using the I-TASSER protein structure and functionprediction server (29, 30) with the D. discoideum myosin-2motor domain as a template (31) (Protein Data Bank code1G8X). Figures were prepared with PyMOL (version 1.5.0.4Schrödinger, LLC).
RESULTS
Protein Expression and Purification—To study biochemicalproperties and protein interaction features of the differenthuman myosin-18A domains, the respective regions wereexpressed separately as discrete proteins (Fig. 1A). Expressionof the human myosin-18A motor domain (M18A-MD) in thebaculovirus/Sf9 system was 0.5–1 mg/liter of Sf9 cells at a den-sity of 1.6 � 106 cells/ml. The purity of M18A-MD followinganti-FLAG affinity chromatography was �95% based on Coo-massie-stained SDS-polyacrylamide gels (cf. Fig. 4A). To ensureproper folding of the motor domain construct, temperature-dependent unfolding of the protein was analyzed by differentialstatic light scattering (data not shown). The transition processis highly cooperative, and the transition temperature (TM �47.6 � 0.9 °C) is consistent with values previously reported formyosin (32, 33). The motor domain construct including thepreceding PDZ module, PDZ-M18A-MD, was expressed inconsiderably lower quantities (�0.1 mg/liter of Sf9 cells). TheN-terminal subdomain constructs of myosin-18A, KEPDZ, KE,and PDZ and the human GOLPH3 protein were expressed inE. coli at 2–10 mg/liter of bacteria culture. The purity of therespective proteins after nickel-nitrilotriacetic acid affinity andsize exclusion chromatography was 90–95%, as estimated fromCoomassie-stained SDS-polyacrylamide gels.Steady-state ATPase Activity of the Myosin-18A Motor
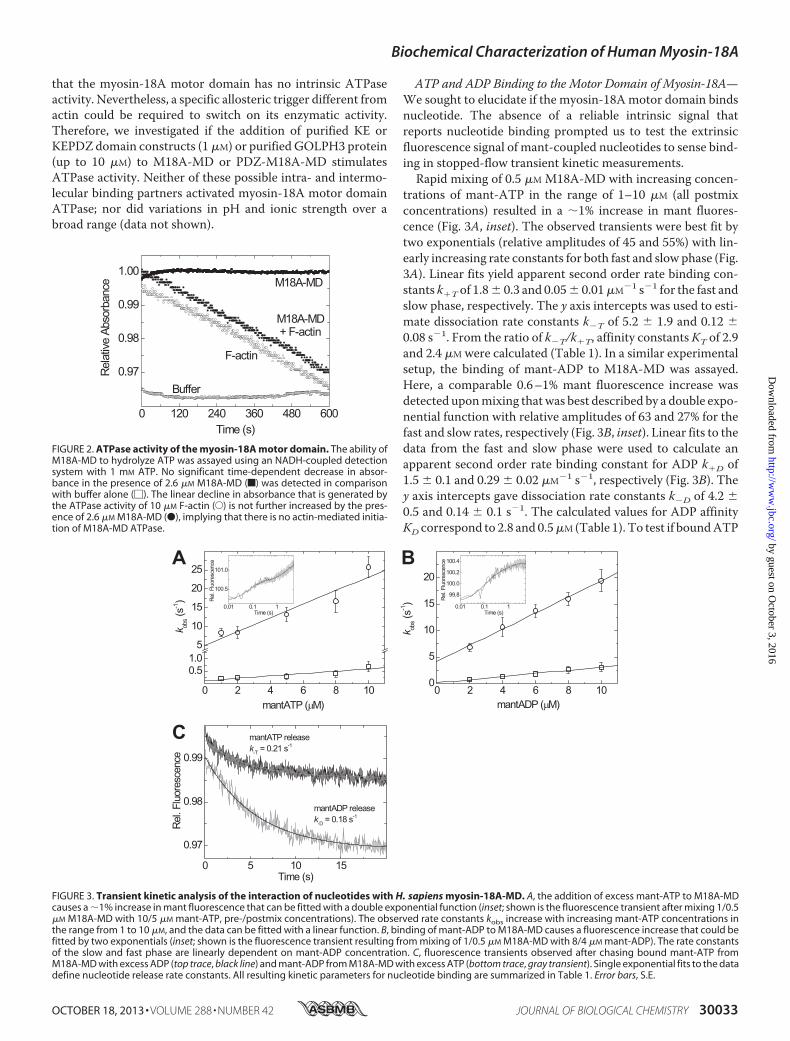
Domain—To explore the ability of M18A-MD to hydrolyzeATP, we performed NADH-coupled ATPase assays in theabsence and presence of F-actin (Fig. 2). We did not detect anysignificant basal ATPase activity for the motor domain con-struct, M18A-MD, or the motor domain including the preced-ing PDZ domain, PDZ-M18A-MD. Furthermore, ATPaseactivity could not be detected in the presence of up to 60 �M
F-actin for both motor domain constructs. These data imply
FLAGMD
HisPDZKE
M18A-MD (residue 399-1185, 800 aa, 88.5 kDa)
KEPDZ (residue 1-398, 406 aa, 45.4 kDa)
FLAGMDPDZPDZ-M18A-MD (residue 220-1185, 979 aa, 108.6 kDa)
HisKEKE (residue 1-219, 227 aa, 25.3 kDa)
HisPDZPDZ (residue 220-311, 105 aa, 11.8 kDa)
Hs myosin-18Aα (2054 aa, 233.1 kD)
FIGURE 1. Constructs used in this study. The schematic depicts the modular structure of the myosin-18A constructs investigated in this study. The abbrevi-ated construct names, number of amino acids, and calculated molecular masses are listed. The motor domain constructs PDZ-M18A-MD and M18A-MD areexpressed in the baculovirus/Sf9 system and purified via a C-terminal FLAG tag, whereas the N-terminal extension constructs are produced in E. coli andpurified via C-terminal His tags. Hs, H. sapiens.
Biochemical Characterization of Human Myosin-18A
30032 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
that the myosin-18A motor domain has no intrinsic ATPaseactivity. Nevertheless, a specific allosteric trigger different fromactin could be required to switch on its enzymatic activity.Therefore, we investigated if the addition of purified KE orKEPDZ domain constructs (1�M) or purified GOLPH3 protein(up to 10 �M) to M18A-MD or PDZ-M18A-MD stimulatesATPase activity. Neither of these possible intra- and intermo-lecular binding partners activated myosin-18A motor domainATPase; nor did variations in pH and ionic strength over abroad range (data not shown).
ATP and ADP Binding to the Motor Domain of Myosin-18A—We sought to elucidate if the myosin-18Amotor domain bindsnucleotide. The absence of a reliable intrinsic signal thatreports nucleotide binding prompted us to test the extrinsicfluorescence signal of mant-coupled nucleotides to sense bind-ing in stopped-flow transient kinetic measurements.Rapid mixing of 0.5 �M M18A-MD with increasing concen-
trations of mant-ATP in the range of 1–10 �M (all postmixconcentrations) resulted in a 1% increase in mant fluores-cence (Fig. 3A, inset). The observed transients were best fit bytwo exponentials (relative amplitudes of 45 and 55%) with lin-early increasing rate constants for both fast and slow phase (Fig.3A). Linear fits yield apparent second order rate binding con-stants k�T of 1.8� 0.3 and 0.05� 0.01�M�1 s�1 for the fast andslow phase, respectively. The y axis intercepts was used to esti-mate dissociation rate constants k�T of 5.2 � 1.9 and 0.12 �0.08 s�1. From the ratio of k�T/k�T, affinity constantsKT of 2.9and 2.4 �M were calculated (Table 1). In a similar experimentalsetup, the binding of mant-ADP to M18A-MD was assayed.Here, a comparable 0.6–1% mant fluorescence increase wasdetected uponmixing thatwas best described by a double expo-nential function with relative amplitudes of 63 and 27% for thefast and slow rates, respectively (Fig. 3B, inset). Linear fits to thedata from the fast and slow phase were used to calculate anapparent second order rate binding constant for ADP k�D of1.5 � 0.1 and 0.29 � 0.02 �M�1 s�1, respectively (Fig. 3B). They axis intercepts gave dissociation rate constants k�D of 4.2 �0.5 and 0.14 � 0.1 s�1. The calculated values for ADP affinityKD correspond to 2.8 and 0.5�M (Table 1). To test if boundATP
0 120 240 360 480 600
0.97
0.98
0.99
1.00
Buffer
F-actin
M18A-MD + F-actin
Time (s)
Rela
tive
Abso
rban
ce M18A-MD
FIGURE 2. ATPase activity of the myosin-18A motor domain. The ability ofM18A-MD to hydrolyze ATP was assayed using an NADH-coupled detectionsystem with 1 mM ATP. No significant time-dependent decrease in absor-bance in the presence of 2.6 �M M18A-MD (f) was detected in comparisonwith buffer alone (�). The linear decline in absorbance that is generated bythe ATPase activity of 10 �M F-actin (E) is not further increased by the pres-ence of 2.6 �M M18A-MD (●), implying that there is no actin-mediated initia-tion of M18A-MD ATPase.
0 2 4 6 8 10
0.51.0
5
10
15
20
25
0.01 0.1 1
100.5
101.0
Time (s)
Rel.
Fluo
resc
ence
mantATP (µM)
k obs
(s-1)
0 2 4 6 8 100
5
10
15
20
0.01 0.1 1
99.8
100.0
100.2
100.4
Rel.
Fluo
resc
ence
Time (s)
mantADP (µM)
k obs
(s-1)
A B
C
0 5 10 15
0.97
0.98
0.99
mantADP releasek-D = 0.18 s-1
Rel.
Fluo
resc
ence
Time (s)
mantATP releasek-T = 0.21 s-1
FIGURE 3. Transient kinetic analysis of the interaction of nucleotides with H. sapiens myosin-18A-MD. A, the addition of excess mant-ATP to M18A-MDcauses a 1% increase in mant fluorescence that can be fitted with a double exponential function (inset; shown is the fluorescence transient after mixing 1/0.5�M M18A-MD with 10/5 �M mant-ATP, pre-/postmix concentrations). The observed rate constants kobs increase with increasing mant-ATP concentrations inthe range from 1 to 10 �M, and the data can be fitted with a linear function. B, binding of mant-ADP to M18A-MD causes a fluorescence increase that could befitted by two exponentials (inset; shown is the fluorescence transient resulting from mixing of 1/0.5 �M M18A-MD with 8/4 �M mant-ADP). The rate constantsof the slow and fast phase are linearly dependent on mant-ADP concentration. C, fluorescence transients observed after chasing bound mant-ATP fromM18A-MD with excess ADP (top trace, black line) and mant-ADP from M18A-MD with excess ATP (bottom trace, gray transient). Single exponential fits to the datadefine nucleotide release rate constants. All resulting kinetic parameters for nucleotide binding are summarized in Table 1. Error bars, S.E.
Biochemical Characterization of Human Myosin-18A
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30033
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

can be exchanged by ADP and vice versa, we premixedM18A-MD with mant-labeled nucleotide and performed achasing experiment with an excess of the other nucleotide (Fig.3C). The observed transients can be described by single expo-nential functions in both cases and define dissociation rate con-stants k�T of 0.21 � 0.07 s�1 and k�D of 0.18 � 0.04 s�1.Althoughwewere not able to resolve a fast phase in the chasingexperiments, the obtained off-rates for the direct displacementare in good agreement with the values obtained from the yintercepts of the slow phase linear fits in Fig. 3, A and B. Thesedata demonstrate the ability of the myosin-18A motor domainto bind and interchange ATP and ADP nucleotides.Actin Binding Properties of the Myosin-18A Motor Domain—
Wemade use of F-actin cosedimentation assays to test the actinbinding ability of M18A-MD in different nucleotide states (Fig.4A). In the absence of actin, the isolatedmotor domain (0.5�M)was completely soluble under our assay conditions.When incu-bated with 4 �M F-actin at high ATP concentrations (5 mM) afraction of 35% of M18A-MD was bound to actin (as deter-mined by densitometry, described under “Experimental Proce-dures”). Under nucleotide-depleted rigor conditions, 44% oftotal M18A-MD remained unbound, whereas 56% was in theactin-bound state. In the ADP state, M18A-MD displayed thehighest actin affinity and was almost completely bound to actin(97%; Fig. 4A). Essentially the same results were obtained usingthe motor domain construct including the N-terminal PDZmodule, PDZ-M18A-MD.These data provide evidence that theprotein binds nucleotide and couples the conformational infor-mation to the actin binding site. Because ADP binding seems tostrengthen actin binding of M18A-MD, we analyzed its effecton F-actin cosedimentation in more detail. We found that theaddition of increasing ADP concentrations to the cosedimen-
tation assay (0.5 �Mmyosin, 4 �M F-actin) shifted the extent ofbound M18A-MD from 56% in the absence of nucleotide to95% at saturating ADP (Fig. 4B). Fitting of a hyperbola to thedata defines the ADP affinity of acto-M18A-MD as KAD �11.4� 3.8�M (see Scheme1 and legend for equilibriumbindingconstant notation). These data suggest a switching of myosin-18A motor domain conformation from a state that is compro-mised in actin binding to a fully actin binding-competent stateupon ADP binding. The F-actin cosedimentation assay results(Fig. 4A) imply a partitioning of myosin-18A between an actin-bound and dissociated state depending on bound nucleotide.To analyze the actin affinity of M18A-MD in different nucleo-tide states in more detail, we mixed 0.5 �M M18A-MD withincreasing concentrations of F-actin in the cosedimentationassay, and the fraction of bound myosin was plotted againstactin concentration (Fig. 4C). In all cases, a standard quadraticequation was fitted to the data to determine dissociation equi-librium constants (cf. Scheme 1); in the absence of nucleotide,M18A-MDhas an actin affinity ofKA � 86� 32 nM that is onlymarginally changed in the presence of ATP with KTA � 182 �155 nM. In the ADP state, the highest actin affinity could beobserved with KDA � 10 nM. To test the possibility of a slowequilibrium between an actin binding-competent state ofM18A-MD and a state that is unable to bind actin, we per-formed sequential actin rebinding experiments in the presenceof 1 mM ATP (Fig. 4D). In contrast to the mouse isoform, thehuman myosin-18A motor domain does not re-equilibrate togive the same distribution between incompetent and compe-tent actin binding states. In contrast, the M18A-MD fractionthat is incompetent to bind F-actin preserves this state evenafter 1 h of incubation at 25 °C. Essentially the same result wasobserved in the absence of nucleotide (data not shown). Never-
TABLE 1Summary of the rate and equilibrium constants of the interactions of the myosin-18A motor domain and N-terminal extension constructs withactin and nucleotidesExperimental conditions were as follows: 25 mM MOPS, 100 mM KCl, 5 mM MgCl2, 1 mM DTT, pH 7.0.
ConstantH. sapiensmyosin-18A
motor domain KEPDZ KEMus musculus
myosin-18A� S1aD. melanogastermyosin-18
motor domainb
Nucleotide binding to myosin(fast/slow phase)
Mant-ATP k�T (�M�1s�1) 1.8 � 0.3/0.05 � 0.01 0.12 � 0.01k�T (s�1)c 5.2 � 1.9/0.12 � 0.08 10.16 � 0.09k�T (s�1)d 0.21 � 0.07 NDe
KT (�M) 2.9/2.4 84.7Mant-ADP k�D (�M�1 s�1) 1.5 � 0.1/0.29 � 0.02 0.10 � 0.02
k-D (s�1)f 4.2 � 0.5/0.14 � 0.1 10.87 � 0.16k�D (s�1)g 0.18 � 0.04 10.3 � 0.3KD (�M) 2.8/0.5 108.7
Nucleotide binding to acto-myosinADP coupling KAD (�M) 11.4 � 3.8
KAD/KD 4.1/22.8Actin binding to myosinRigor KA (�M) 0.086 � 0.032 0.99 � 0.29 6.53 � 4.1 47.9 � 9.2 1.0 � 0.2
Fraction bound 0.61 � 0.06 1.0 � 0.09 1.0 � 0.27 0.248 � 0.078 0.83 � 0.05ADP KDA (�M) �0.01 ND ND ND ND
Fraction bound 0.97 � 0.08ATP KTA (�M) 0.182 � 0.155 1.02 � 0.24 3.92 � 6.75 54.0 � 29 1.0 � 0.1
Fraction bound 0.5 � 0.07 0.99 � 0.1 0.87 � 0.22 0.245 � 0.071 0.81 � 0.09a Guzik-Lendrum et al. (4).b Guzik-Lendrum et al. (12).c y intercept from the kobs versus mant-ATP plot.d ADP chasing experiment.e ND, not determined.f y intercept from the kobs versusmant-ADP plot.g ATP chasing experiment.
Biochemical Characterization of Human Myosin-18A
30034 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
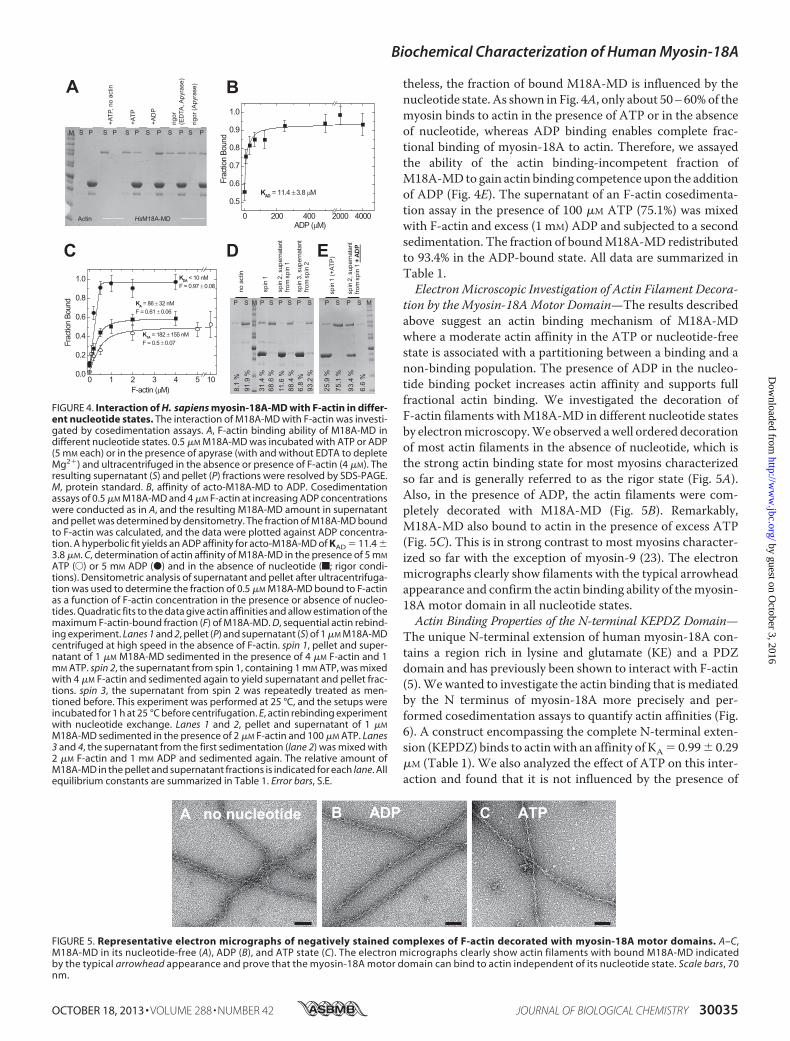
theless, the fraction of bound M18A-MD is influenced by thenucleotide state. As shown in Fig. 4A, only about 50–60%of themyosin binds to actin in the presence of ATP or in the absenceof nucleotide, whereas ADP binding enables complete frac-tional binding of myosin-18A to actin. Therefore, we assayedthe ability of the actin binding-incompetent fraction ofM18A-MDto gain actin binding competence upon the additionof ADP (Fig. 4E). The supernatant of an F-actin cosedimenta-tion assay in the presence of 100 �M ATP (75.1%) was mixedwith F-actin and excess (1 mM) ADP and subjected to a secondsedimentation. The fraction of boundM18A-MD redistributedto 93.4% in the ADP-bound state. All data are summarized inTable 1.ElectronMicroscopic Investigation of Actin Filament Decora-
tion by the Myosin-18AMotor Domain—The results describedabove suggest an actin binding mechanism of M18A-MDwhere a moderate actin affinity in the ATP or nucleotide-freestate is associated with a partitioning between a binding and anon-binding population. The presence of ADP in the nucleo-tide binding pocket increases actin affinity and supports fullfractional actin binding. We investigated the decoration ofF-actin filaments withM18A-MD in different nucleotide statesby electronmicroscopy.Weobserved awell ordered decorationof most actin filaments in the absence of nucleotide, which isthe strong actin binding state for most myosins characterizedso far and is generally referred to as the rigor state (Fig. 5A).Also, in the presence of ADP, the actin filaments were com-pletely decorated with M18A-MD (Fig. 5B). Remarkably,M18A-MD also bound to actin in the presence of excess ATP(Fig. 5C). This is in strong contrast to most myosins character-ized so far with the exception of myosin-9 (23). The electronmicrographs clearly show filaments with the typical arrowheadappearance and confirm the actin binding ability of themyosin-18A motor domain in all nucleotide states.Actin Binding Properties of the N-terminal KEPDZ Domain—
The unique N-terminal extension of human myosin-18A con-tains a region rich in lysine and glutamate (KE) and a PDZdomain and has previously been shown to interact with F-actin(5).Wewanted to investigate the actin binding that is mediatedby the N terminus of myosin-18A more precisely and per-formed cosedimentation assays to quantify actin affinities (Fig.6). A construct encompassing the complete N-terminal exten-sion (KEPDZ) binds to actinwith an affinity of KA� 0.99� 0.29�M (Table 1). We also analyzed the effect of ATP on this inter-action and found that it is not influenced by the presence of
0 200 400 2000 4000
0.5
0.6
0.7
0.8
0.9
1.0
KAD = 11.4 ±3.8 µM
ADP (µM)Fr
actio
n Bo
und
P S P S M
spin
1 (+
ATP
)
spin
2, s
uper
nata
ntfro
msp
in1
+ AD
P
25.9
%
75.1
%
93.4
%
6.6
%
P S M P S P S P S
noac
tin
spin
1
spin
2, s
uper
nata
ntfro
msp
in1
spin
3, s
uper
nata
ntfro
msp
in2
8.1
%91
.9 %
31.4
%68
.6 %
11.6
%88
.4 %
6.8
%93
.2 %
Actin HsM18A-MD
M S P S P S P S P S P S P
+ATP
, no
actin
+ATP
+AD
P
rigor
(ED
TA, A
pyra
se)
rigor
(Apy
rase
)A B
C D
0 1 2 3 4 5 100.0
0.2
0.4
0.6
0.8
1.0
KTA = 182 ± 155 nMF = 0.5 ± 0.07
KDA < 10 nMF = 0.97 ± 0.08
Frac
tion
Boun
d
F-actin (µM)
KA = 86 ± 32 nMF = 0.61 ± 0.06
E
FIGURE 4. Interaction of H. sapiens myosin-18A-MD with F-actin in differ-ent nucleotide states. The interaction of M18A-MD with F-actin was investi-gated by cosedimentation assays. A, F-actin binding ability of M18A-MD indifferent nucleotide states. 0.5 �M M18A-MD was incubated with ATP or ADP(5 mM each) or in the presence of apyrase (with and without EDTA to depleteMg2�) and ultracentrifuged in the absence or presence of F-actin (4 �M). Theresulting supernatant (S) and pellet (P) fractions were resolved by SDS-PAGE.M, protein standard. B, affinity of acto-M18A-MD to ADP. Cosedimentationassays of 0.5 �M M18A-MD and 4 �M F-actin at increasing ADP concentrationswere conducted as in A, and the resulting M18A-MD amount in supernatantand pellet was determined by densitometry. The fraction of M18A-MD boundto F-actin was calculated, and the data were plotted against ADP concentra-tion. A hyperbolic fit yields an ADP affinity for acto-M18A-MD of KAD � 11.4 �3.8 �M. C, determination of actin affinity of M18A-MD in the presence of 5 mM
ATP (E) or 5 mM ADP (●) and in the absence of nucleotide (f; rigor condi-tions). Densitometric analysis of supernatant and pellet after ultracentrifuga-tion was used to determine the fraction of 0.5 �M M18A-MD bound to F-actinas a function of F-actin concentration in the presence or absence of nucleo-tides. Quadratic fits to the data give actin affinities and allow estimation of themaximum F-actin-bound fraction (F) of M18A-MD. D, sequential actin rebind-ing experiment. Lanes 1 and 2, pellet (P) and supernatant (S) of 1 �M M18A-MDcentrifuged at high speed in the absence of F-actin. spin 1, pellet and super-natant of 1 �M M18A-MD sedimented in the presence of 4 �M F-actin and 1mM ATP. spin 2, the supernatant from spin 1, containing 1 mM ATP, was mixedwith 4 �M F-actin and sedimented again to yield supernatant and pellet frac-tions. spin 3, the supernatant from spin 2 was repeatedly treated as men-tioned before. This experiment was performed at 25 °C, and the setups wereincubated for 1 h at 25 °C before centrifugation. E, actin rebinding experimentwith nucleotide exchange. Lanes 1 and 2, pellet and supernatant of 1 �M
M18A-MD sedimented in the presence of 2 �M F-actin and 100 �M ATP. Lanes3 and 4, the supernatant from the first sedimentation (lane 2) was mixed with2 �M F-actin and 1 mM ADP and sedimented again. The relative amount ofM18A-MD in the pellet and supernatant fractions is indicated for each lane. Allequilibrium constants are summarized in Table 1. Error bars, S.E.
A no nucleotide B ADP C ATP
FIGURE 5. Representative electron micrographs of negatively stained complexes of F-actin decorated with myosin-18A motor domains. A–C,M18A-MD in its nucleotide-free (A), ADP (B), and ATP state (C). The electron micrographs clearly show actin filaments with bound M18A-MD indicatedby the typical arrowhead appearance and prove that the myosin-18A motor domain can bind to actin independent of its nucleotide state. Scale bars, 70nm.
Biochemical Characterization of Human Myosin-18A
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30035
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from
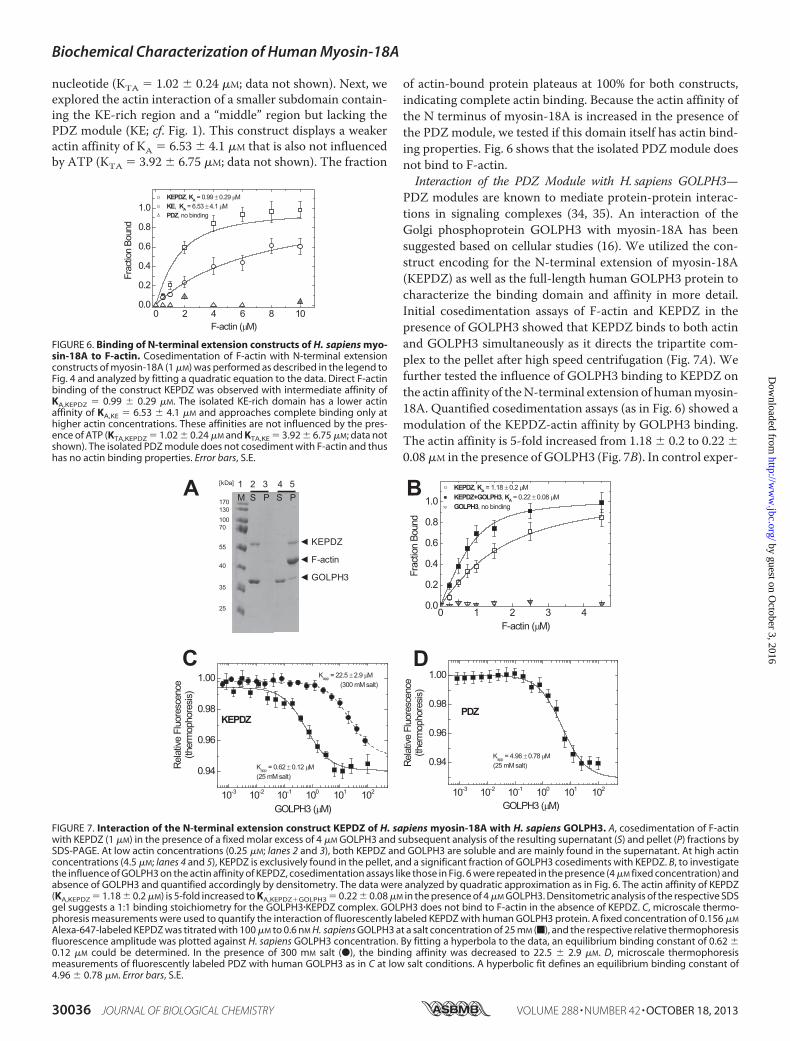
nucleotide (KTA � 1.02 � 0.24 �M; data not shown). Next, weexplored the actin interaction of a smaller subdomain contain-ing the KE-rich region and a “middle” region but lacking thePDZ module (KE; cf. Fig. 1). This construct displays a weakeractin affinity of KA � 6.53 � 4.1 �M that is also not influencedby ATP (KTA � 3.92 � 6.75 �M; data not shown). The fraction
of actin-bound protein plateaus at 100% for both constructs,indicating complete actin binding. Because the actin affinity ofthe N terminus of myosin-18A is increased in the presence ofthe PDZ module, we tested if this domain itself has actin bind-ing properties. Fig. 6 shows that the isolated PDZ module doesnot bind to F-actin.Interaction of the PDZ Module with H. sapiens GOLPH3—
PDZ modules are known to mediate protein-protein interac-tions in signaling complexes (34, 35). An interaction of theGolgi phosphoprotein GOLPH3 with myosin-18A has beensuggested based on cellular studies (16). We utilized the con-struct encoding for the N-terminal extension of myosin-18A(KEPDZ) as well as the full-length human GOLPH3 protein tocharacterize the binding domain and affinity in more detail.Initial cosedimentation assays of F-actin and KEPDZ in thepresence of GOLPH3 showed that KEPDZ binds to both actinand GOLPH3 simultaneously as it directs the tripartite com-plex to the pellet after high speed centrifugation (Fig. 7A). Wefurther tested the influence of GOLPH3 binding to KEPDZ onthe actin affinity of theN-terminal extension of humanmyosin-18A. Quantified cosedimentation assays (as in Fig. 6) showed amodulation of the KEPDZ-actin affinity by GOLPH3 binding.The actin affinity is 5-fold increased from 1.18 � 0.2 to 0.22 �0.08 �M in the presence of GOLPH3 (Fig. 7B). In control exper-
0 2 4 6 8 100.0
0.2
0.4
0.6
0.8
1.0KEPDZ, KA = 0.99 ± 0.29 µMKE, KA = 6.53 ± 4.1 µMPDZ, no binding
F-actin (µM)
Frac
tion
Boun
d
FIGURE 6. Binding of N-terminal extension constructs of H. sapiens myo-sin-18A to F-actin. Cosedimentation of F-actin with N-terminal extensionconstructs of myosin-18A (1 �M) was performed as described in the legend toFig. 4 and analyzed by fitting a quadratic equation to the data. Direct F-actinbinding of the construct KEPDZ was observed with intermediate affinity ofKA,KEPDZ � 0.99 � 0.29 �M. The isolated KE-rich domain has a lower actinaffinity of KA,KE � 6.53 � 4.1 �M and approaches complete binding only athigher actin concentrations. These affinities are not influenced by the pres-ence of ATP (KTA,KEPDZ � 1.02 � 0.24 �M and KTA,KE � 3.92 � 6.75 �M; data notshown). The isolated PDZ module does not cosediment with F-actin and thushas no actin binding properties. Error bars, S.E.
10-3 10-2 10-1 100 101 102
0.94
0.96
0.98
1.00
Kapp = 4.96 ± 0.78 µM(25 mM salt)
PDZ
GOLPH3 (µM)
Rela
tive
Fluo
resc
ence
(ther
mop
hore
sis)
0 1 2 3 40.0
0.2
0.4
0.6
0.8
1.0KEPDZ, KA = 1.18 ± 0.2 µMKEPDZ+GOLPH3, KA = 0.22 ± 0.08 µMGOLPH3, no binding
F-actin (µM)
Frac
tion
Boun
d
A B
C
10-3 10-2 10-1 100 101 102
0.94
0.96
0.98
1.00
KEPDZ
Kapp = 22.5 ± 2.9 µM(300 mM salt)
Kapp = 0.62 ± 0.12 µM(25 mM salt)
GOLPH3 (µM)
Rela
tive
Fluo
resc
ence
(ther
mop
hore
sis)
M S P S P1 2 3 4 5
◄ KEPDZ
◄ F-actin
◄ GOLPH3
[kDa]
17013010070
55
40
35
25
D
FIGURE 7. Interaction of the N-terminal extension construct KEPDZ of H. sapiens myosin-18A with H. sapiens GOLPH3. A, cosedimentation of F-actinwith KEPDZ (1 �M) in the presence of a fixed molar excess of 4 �M GOLPH3 and subsequent analysis of the resulting supernatant (S) and pellet (P) fractions bySDS-PAGE. At low actin concentrations (0.25 �M; lanes 2 and 3), both KEPDZ and GOLPH3 are soluble and are mainly found in the supernatant. At high actinconcentrations (4.5 �M; lanes 4 and 5), KEPDZ is exclusively found in the pellet, and a significant fraction of GOLPH3 cosediments with KEPDZ. B, to investigatethe influence of GOLPH3 on the actin affinity of KEPDZ, cosedimentation assays like those in Fig. 6 were repeated in the presence (4 �M fixed concentration) andabsence of GOLPH3 and quantified accordingly by densitometry. The data were analyzed by quadratic approximation as in Fig. 6. The actin affinity of KEPDZ(KA,KEPDZ � 1.18 � 0.2 �M) is 5-fold increased to KA,KEPDZ�GOLPH3 � 0.22 � 0.08 �M in the presence of 4 �M GOLPH3. Densitometric analysis of the respective SDSgel suggests a 1:1 binding stoichiometry for the GOLPH3�KEPDZ complex. GOLPH3 does not bind to F-actin in the absence of KEPDZ. C, microscale thermo-phoresis measurements were used to quantify the interaction of fluorescently labeled KEPDZ with human GOLPH3 protein. A fixed concentration of 0.156 �M
Alexa-647-labeled KEPDZ was titrated with 100 �M to 0.6 nM H. sapiens GOLPH3 at a salt concentration of 25 mM (f), and the respective relative thermophoresisfluorescence amplitude was plotted against H. sapiens GOLPH3 concentration. By fitting a hyperbola to the data, an equilibrium binding constant of 0.62 �0.12 �M could be determined. In the presence of 300 mM salt (●), the binding affinity was decreased to 22.5 � 2.9 �M. D, microscale thermophoresismeasurements of fluorescently labeled PDZ with human GOLPH3 as in C at low salt conditions. A hyperbolic fit defines an equilibrium binding constant of4.96 � 0.78 �M. Error bars, S.E.
Biochemical Characterization of Human Myosin-18A
30036 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

iments, GOLPH3 was completely soluble under the assay con-ditions and did not bind to F-actin. To analyze and quantify thedirect interaction of KEPDZ and GOLPH3, we took advantageof microscale thermophoresis (27, 28). This newly developedmethod allows the determination of the binding affinitybetween a fluorescently labeled and a non-labeled protein. WetitratedAlexa-647-labeledKEPDZproteinwith increasing con-centrations of GOLPH3. The observed GOLPH3 concentra-tion-dependent decrease in the thermophoresis fluorescenceamplitude (Fig. 7C) was fittedwith a hyperbola to give an appar-ent equilibrium binding constant Kapp of 0.62 � 0.12 �M at lowsalt conditions. In the presence of high salt concentrations (300mMNaCl), the binding affinity was decreased to 22.5 � 2.9 �M.This result confirms a direct binding event between GOLPH3and the N-terminal extension of myosin-18A. To test if theisolated PDZ module is sufficient to mediate binding of myo-sin-18A to GOLPH3, we repeated the thermophoresis experi-ment with purified fluorescently labeled PDZ protein (Fig. 7D).The results show that the PDZ module binds to GOLPH3 butwith lower affinity (4.96 � 0.78 �M at low salt conditions) thanthe complete N terminus (KEPDZ construct).
DISCUSSION
In the present study, we aimed to elucidate the molecularproperties of three fundamental humanmyosin-18A functionaldomains: the KE-rich region and the PDZ module, whichtogether constitute the N-terminal extension, as well as thegeneric myosin motor domain. We established the solubleexpression and purification of different constructs, using twoexpression systems; a minimal construct encompassing thecore motor domain (M18A-MD) and a motor domainconstruct including the N-terminally located PDZ module(PDZ-M18A-MD)were successfully produced in the Sf9/insectcell system. The N-terminal extension construct (KEPDZ) aswell as the separate KE-rich region (KE) and the PDZ module(PDZ) were produced in E. coli and purified to homogeneity. Inaddition, we expressed the phosphoprotein GOLPH3 as a GSTfusion protein in E. coli and utilized protease cleavage to obtaina “tag-free” version for interaction studies. Using these con-structs, we studied the biochemical properties of the individualmyosin-18A domains to assemble an overall picture of the pro-tein’s molecular mechanism.We show that the core myosin-18A motor domain binds
mant-labeled ATP and ADP nucleotides but does not exhibitintrinsic basal or actin-activated ATPase activity. Nucleo-tide binding modulates the actin affinity of the motordomain and regulates its partitioning between an actin bind-ing-competent and -incompetent state. The ADP-boundstate has the highest actin affinity, and complete fractionalbinding of the myosin-18A motor domain to the actin fila-ment is only observed in this state. These results are sup-ported by the fact that in electron micrographs, we observecomplete decoration of F-actin filaments with the typicalarrowhead appearance of the myosin motor domain in allnucleotide states. Like generic myosin motors, the myosin-18A motor domain appears to dock with similar orientationand employing conserved surface contacts to the actin fila-ment. Accordingly, the myosin-18A motor domain has pre-
served the myosin-inherent ability to bind nucleotide and tocouple the conformational information from the nucleotidebinding pocket to the actin binding site.The core motor domain of myosin-18A, omitting the N-ter-
minal extension, comprises 786 amino acids. It is considerablyextended comparedwith human skeletal musclemyosin-2 (701amino acids without the N-terminal SH3-like subdomain) orD. discoideum myosin-2 (685 amino acids without SH3). Theunderlying insertions are distributed over the nucleotide bind-ing pocket elements (switch-2 region), the actin bindingregions (cardiomyopathy loop (CM-loop) and activation loop),and prominent surface loops (near the SH2 domain) of themotor domain. We prepared a structural model of the motordomain using the I-TASSER protein structure and functionprediction server (29, 30). The model shows the location ofthese inserts and their likely effect on conserved structural ele-ments in the myosin motor domain (Fig. 8, A and B; alignmentdepicted in Fig. 8C).The switch-2 region of myosin-18A displays an insertion of
six residues, and the conserved glutamic acid in switch-2 ischanged to glutamine (pink in Fig. 8,A andB). Previously, it hasbeen shown that this residue plays a key role in the mechanismof chemo-mechanical coupling (36) and that mutation of thisresidue abolishes the cellular functions of myosin-2 in D. dis-coideum cells (19). Accordingly, the elongation of the switch-2region in myosin-18A by a flexible linker (GGSARGA) and thedisturbed salt bridge between switch-1 and switch-2 are possi-ble structural determinants for the absent intrinsic ATPaseactivity of the motor domain.Furthermore, the region preceding the SH2 domain contains
a 29-residue surface loop extension (yellow in Fig. 8, A and B).This flexible loop is located near the entrance of the nucleotidebinding pocket and comprises multiple serine residues. Weobserved biphasic ATP andADPbinding kinetics thatmay pos-sibly be attributed to the existence of two structural states ofthis extended SH2 loopmodulating nucleotide interactions andtriggering slow and fast binding and release kinetics. This inter-pretation is supported by the fact that both slow and fast ratesfor ATP and ADP binding are dependent on nucleotide con-centration. The biphasic nucleotide binding behavior has notbeen observed for the mouse myosin-18A S1 construct (4).Also, the overall nucleotide affinities of human M18A-MD areabout 30 times higher than for the mouse isoform, and theADP-mediated switching to full binding ability has not beendescribed before. Because mouse and human isoforms of myo-sin-18A share complete amino acid sequence conservation inthe SH2 loop extension, this is not the only determinant for thenucleotide binding properties of myosin-18A.The CM-loop is located at the tip of the motor domain and
involved in the actomyosin binding interface. Remarkably, inmyosin-18A, the loop is considerably modified because itcontains 14 additional amino acids (blue in Fig. 8A). Thisenables higher flexibility of the loop and alters the actinbinding properties in comparison with generic myosinmotor domains. A more flexible and elongated CM-loopmight render the motor’s actin affinity more independentfrom cleft closure and thereby permit high actin affinity that
Biochemical Characterization of Human Myosin-18A
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30037
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

is less influenced by the nucleotide state of the active sitethan in other myosins.The activation loop, which is part of the actin-myosin inter-
face, is located within helix HR in the helix-loop-helix actinbinding motif (orange in Fig. 8A) (37, 38). In human myosin-18A, this loop is elongated by 13 extra amino acids that intro-duce additional positive charges (three arginine residuesinstead of one in D. discoideum myosin-2), which interfereswith regular activation by this loop (cf. Fig. 8C).
The actin affinities of the humanmyosin-18Amotor domainin the absence and presence of nucleotide are 2 orders of mag-nitude stronger than for mouse myosin-18A S1 (Table 1).Accordingly, for the mouse isoform, no decoration of F-actinfilaments could be observed by electron microscopic investiga-tion. In contrast, M18A-MD shows the classic arrowhead dec-oration of F-actin filaments. The sequence comparison (Fig.8C) of the actin binding regions (CM-loop and activation loop)displays only subtle differences between the two myosin iso-
A B
C CM-loop__________CM-loop___________
384 VNPSVLEKALM--------EPRILAGRDLVAQHLNVEKS------SSSRDALVK Dd myosin-2 MD704 CSLEELSSAIFKHQHKGGTLQRSTSFRQGPEESGLGDGTGPKLSALECLEGMAA Hs myosin-18A MD704 CSLEELSSAIFKHQLKGGTLQRSTSFRQGPEESGLGEGT--KLSALECLEGMAS Mm myosin-18A MD861 VNLEDLSSAAF-----GLTQPNAPNGGLSPSKSPTSDTG--HEWAWECLEALVI Dm myosin-18 MD
Switch-2DIXGFE
450 IGVLDISGFEIFKVN------SFEQLCINYTNEKLQQFFNHHMFKLEQEE Dd myosin-2 MD785 MMIVDTPGFQNPEQGGSARGASFEELCHNYTQDRLQRLFHERTFVQELER Hs myosin-18A MD783 MMIVDTPGFQNPEWGGSARGASFEELCHNYAQDRLQRLFHERTFLQELER Mm myosin-18A MD935 IMLIDTPGFQNPASCGQQVGATLADLRHNYLQERLQMLFHHTTLVAPRDR Dm myosin-18 MD
Activation loop___ activation-loop____
513 TIDLIDGRQPP----------------GILALLDEQSVFPNATDNTLITKL Dd myosin-2 MD854 SVAAVDQASHQSLVRSLAR---TDEARGLLWLLEEEALVPGASEDTLLERL Hs myosin-18A MD852 SVAAVDQASH--LVRSLAH---ADEARGLLWLLEEEALVPGATEDALLDRL Mm myosin-18A MD1005 LISLIDKAPQNHVVRSSQRDLREHDRRGMLWLLDEEAIYPNSNDDTFLERL Dm myosin-18 MD
Region preceding the SH-2 helix___SH-2____
657 IPNNK--------------------QLPAK---------LEDKVVLDQLRGS Dd myosin-2 MD1048 LPVAEGWAGEPRSASSRRVSSSSELDLPSGDHCEAGLLQLDVPLLRTQLRGS Hs myosin-18A MD1044 LPVAEGWPGEPRSASSRRVSSSSELDLPPGDPCEAGLLQLDVSLLRAQLRGS Mm myosin-18A MD1197 LLQHN--AGKHTKYTANGSPSSAAGQVSSE------EEMVNVPLLRSQLRGS Dm myosin-18 MD
N-terminus
C-terminus
FIGURE 8. Structural model of the H. sapiens myosin-18A motor domain. The D. discoideum myosin-2 motor domain structure (Protein Data Bank code1G8X) was used as a template to generate a homology model of the human myosin-18A motor domain (residues 399 –1185 of the full-length sequence).A, the overall fold of the motor domain of human myosin-18A displays high similarity with generic myosin motor domains. Nevertheless, H. sapiensmyosin-18A contains four major insertions in the motor domain sequence, which are located near switch-2 (pink; 6 residues), at the CM-loop (blue; 14residues), at the activation loop (orange; 13 residues), and preceding the SH2 helix (yellow; 29 residues). ADP is shown in a stick representation with blackcarbon atoms; the orange sphere designates the location of the Mg2� ion. B, close-up view of the nucleotide binding pocket. The molecule was subjectedto a left-handed rotation of about 45º around a vertical axis through the Mg2� ion. Important features of the binding pocket are colored as follows: cyan,P-loop (GSSGSGKT); red, switch-1 (NGNATR); light blue, switch-2 (DTPGFQ). C, multiple-sequence alignment of the motor domain (MD) of D. discoideum(Dd) myosin-2 and the motor domains of human (Hs) myosin-18A, mouse (Mm) myosin-18A, and D. melanogaster (Dm) myosin-18. Important myosinmotor domain features are indicated and labeled. The gray shaded box marks Glu-459 (D. discoideum), which constitutes the salt bridge with Arg-238 inD. discoideum myosin-2.
Biochemical Characterization of Human Myosin-18A
30038 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

forms. Nevertheless, in their study, Guzik-Lendrum et al. (4)utilized an S1 construct comprising a short N-terminalsequence preceding the motor domain and a neck region withbound essential and regulatory light chains. A previous studyrevealed differences for chicken skeletal musclemyosin S1withwild-type versus truncated essential light chains in actin bind-ing and ATPase activity, suggesting a direct interaction of theessential light chain N terminus with actin that is regulated bythe SH3-like subdomain ofmyosin (39). Accordingly, one couldspeculate about a similar mechanism for myosin-18A, wherethe short N-terminal sequence of the mouse myosin-18A S1construct used in the study of Guzik-Lendrum et al. (4) and thebound light chains modulate the actin (and maybe also nucle-otide) affinity of myosin-18A.The biochemical characterization of modules that reside in
the large N-terminal extension of human myosin-18A� pro-vides information on the molecular function of this myosin inthe cellular context. Two previous studies proposed ATP-inde-pendent actin binding for the N-terminal subdomain of myo-sin-18A� (5, 11). In both studies, immunoprecipitation fromhuman cell line-derived cell lysates showed an interaction ofthe N-terminal extension of myosin-18A with actin filaments.In expanding these studies, we utilized purified bacteriallyexpressed N-terminal subdomain constructs to define bindingaffinities and saturation ratios for F-actin. We confirm thatthe actin binding site resides within the KE-rich subdomain(amino acids 1–219 of humanmyosin-18A�) with intermediateaffinity and full binding ability. Although the isolated PDZmodule does not interact with actin, it significantly strengthensthe actin affinity of the N-terminal extension. Remarkably, thedirect interaction of GOLPH3 with the PDZ module leads tochanges in the actin binding properties, and this interplay actsas a modulator for the KE motif-mediated cytoskeleton inter-action. Likewise, the affinity of the PDZmodule with GOLPH3is significantly enhanced by the presence of the KE-rich region(Fig. 7,C andD). This further confirms an interdependent con-nection between the neighboring functional domains KE andPDZ. The observed independence of the binding affinities fromATP confirms the assumption of Isogawa et al. (5) that thisinteraction is ATP-insensitive. The N-terminal extension ofmyosin-18A could therefore be used by the protein to cross-link F-actin filaments to higher order complexes in a GOLPH3binding-regulated fashion.We determined the ADP affinity of myosin-18A in the actin-
bound state (KAD) to be 12 �M, which is in the range ofreported physiological concentrations: 12 �M MgADP in brainand 6�MMgADP in restingmuscle or 43�MMgADP inmuscleafter heavy exercise (40); 40 to 140�M (relaxed/contracted) freeADP in the cytosol of smooth muscle cells (41, 42). Within thecytosol, local high ADP concentrations are expected in areas ofhigh ATP turnover, such as myosin filaments or near mem-branes were ion pumps are located. Transiently increased ADPlevelsmay facilitate strong actin binding of all four binding sitesof a dimeric myosin-18A molecule, resulting in stable cross-linking of F-actin filaments or an attenuation of myosin fila-ment contractility. However, in all possible nucleotide states, afraction of the motor domain is strongly bound to the actinfilaments. It is therefore likely that in cellular conditions, a
release factor is needed to detach myosin-18A from the actincytoskeleton to allow redistribution.When feeding all equilibrium binding constants of M18A-MD
for actin and ADP into Scheme 1, the product should equal 1.Using the values determined in this study, we obtain a product of0.029. This discrepancy suggests the presence of at least one addi-tional step in the interaction ofM18A-MDwith actin and/orADPthat is not resolved by our experiments.For human myosin-18A, a number of confirmed or putative
binding partners have been described (13, 14, 16, 43, 44). Theycan in principle act as modulators of myosin-18A function orstimulate enzymatic activity when bound to the protein. Fur-thermore, the presence of the complete N-terminal extensionmay be necessary for motor domain catalytic function as forexample in myosin-3A the presence of the N-terminal kinasedomain happens to modulate the kinetics of the motor domain(45, 46). Nevertheless, Guzik-Lendrum et al. (4) report nucleo-tide and actin binding but no significant ATPase activity for arecombinant mouse myosin-18A� motor construct, whichcontains the complete N-terminal domain.In an attempt to characterize the interaction of myosin-
18A with GOLPH3, we were able to prove the direct bindingof the phosphoprotein to the N-terminal extension of myo-sin-18A. Because the affinity is ionic strength-dependent, wesuppose that charge-charge interactions are responsible forhigh affinity binding. We could show a direct effect ofGOLPH3 on myosin-18A function because its binding to thePDZ module enhances the actin affinity of the N-terminalextension. Moreover, the GOLPH3�myosin-18A complexserves as a junction between the Golgi membrane and thecytoskeleton and may thus be highly regulated, presumablyby other associated proteins.The motor domain of myosin-18A appears to have evolved to
serve as an actin cross-linker, whose activity is modulated in anucleotide- and cargo-dependentmanner. Direct actin binding tothe motor domain involves generic actin binding motifs and con-servedalbeit attenuatedcommunicationpathwaysbetweennucle-otide and actin binding regions. Actin binding to the N-terminalextension is modulated by binding of GOLPH3 and potentiallyother cargo molecules to the PDZ module. Non-muscle myo-sin-2Aandmyosin-18Ahavebeen shown to share the sameessen-tial and regulatory light chains (4).Moreover, they colocalize nearthe cell periphery in lamellar actomyosin bundles (14). Therefore,it is tempting to speculate thatmyosin-18A is part of these bipolarfilaments actingasmediatorbetweenmembraneandcytoskeletoncomponents.Otherpotential roles ofmyosin-18A include a ratch-et-like function, where the positive strain resulting from the pro-ductive interaction of non-muscle myosin-2A with actin pushesthemolecule forward to the next actin binding site, where it snapsinto place. In the same manner, strain-induced conformationalchanges can affect nucleotide and actin affinity, enabling myosin-18A to work as an efficient strain sensor within the contractilemachinery.
Acknowledgments—We thank Michael Radke, Henning Grosskopf,Georg Adler-Gunzelmann, and Michal Stanczak for help anddiscussions.
Biochemical Characterization of Human Myosin-18A
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30039
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

REFERENCES1. Bloemink, M. J., and Geeves, M. A. (2011) Shaking the myosin family tree.
Biochemical kinetics defines four types of myosin motor. Semin. Cell Dev.Biol. 22, 961–967
2. Odronitz, F., and Kollmar, M. (2007) Drawing the tree of eukaryotic lifebased on the analysis of 2,269 manually annotated myosins from 328 spe-cies. Genome Biol. 8, R196
3. Furusawa, T., Ikawa, S., Yanai, N., and Obinata, M. (2000) Isolation of anovel PDZ-containing myosin from hematopoietic supportive bone mar-row stromal cell lines. Biochem. Biophys. Res. Commun. 270, 67–75
4. Guzik-Lendrum, S., Heissler, S. M., Billington, N., Takagi, Y., Yang, Y.,Knight, P. J., Homsher, E., and Sellers, J. R. (2013) Mammalian myosin-18A, a highly divergent myosin. J. Biol. Chem. 288, 9532–9548
5. Isogawa, Y., Kon, T., Inoue, T., Ohkura, R., Yamakawa, H., Ohara, O., andSutoh, K. (2005) The N-terminal domain of MYO18A has an ATP-insen-sitive actin-binding site. Biochemistry 44, 6190–6196
6. Langer, W., Sohler, F., Leder, G., Beckmann, G., Seidel, H., Gröne, J.,Hummel,M., and Sommer, A. (2010) Exon array analysis using re-definedprobe sets results in reliable identification of alternatively spliced genes innon-small cell lung cancer. BMC Genomics 11, 676
7. Walz, C., Chase, A., Schoch, C., Weisser, A., Schlegel, F., Hochhaus, A.,Fuchs, R., Schmitt-Gräff, A., Hehlmann, R., Cross, N. C., and Reiter, A.(2005) The t(8;17)(p11;q23) in the 8p11 myeloproliferative syndromefuses MYO18A to FGFR1. Leukemia 19, 1005–1009
8. Walz, C., Haferlach, C., Hänel, A., Metzgeroth, G., Erben, P., Gosenca, D.,Hochhaus, A., Cross, N. C., and Reiter, A. (2009) Identification of aMYO18A-PDGFRB fusion gene in an eosinophilia-associated atypicalmyeloproliferative neoplasm with a t(5;17)(q33–34;q11.2). Genes Chro-mosomes Cancer 48, 179–183
9. Ussowicz, M., Jaskowiec, A., Meyer, C., Marschalek, R., Chybicka, A.,Szczepanski, T., and Haus, O. (2012) A three-way translocation of MLL,MLLT11, and the novel reciprocal partner gene MYO18A in a child withacute myeloid leukemia. Cancer Genet. 205, 261–265
10. Mori, K., Furusawa, T., Okubo, T., Inoue, T., Ikawa, S., Yanai, N., Mori,K. J., andObinata,M. (2003)Genome structure and differential expressionof two isoforms of a novel PDZ-containing myosin (MysPDZ) (Myo18A).J. Biochem. 133, 405–413
11. Mori, K., Matsuda, K., Furusawa, T., Kawata, M., Inoue, T., and Obi-nata, M. (2005) Subcellular localization and dynamics of MysPDZ(Myo18A) in live mammalian cells. Biochem. Biophys. Res. Commun.326, 491–498
12. Guzik-Lendrum, S., Nagy, A., Takagi, Y., Houdusse, A., and Sellers, J. R.(2011) Drosophila melanogaster myosin-18 represents a highly diver-gent motor with actin tethering properties. J. Biol. Chem. 286,21755–21766
13. Hsu, R.-M., Tsai, M.-H., Hsieh, Y.-J., Lyu, P.-C., and Yu, J.-S. (2010) Iden-tification of MYO18A as a novel interacting partner of the PAK2/�PIX/GIT1 complex and its potential function in modulating epithelial cell mi-gration.Mol. Biol. Cell 21, 287–301
14. Tan, I., Yong, J., Dong, J. M., Lim, L., and Leung, T. (2008) A tripartitecomplex containing MRCK modulates lamellar actomyosin retrogradeflow. Cell 135, 123–136
15. Heissler, S. M., and Manstein, D. J. (2013) Nonmuscle myosin-2. Mix andmatch. Cell. Mol. Life Sci. 70, 1–21
16. Dippold, H. C., Ng, M. M., Farber-Katz, S. E., Lee, S.-K., Kerr, M. L.,Peterman, M. C., Sim, R., Wiharto, P. A., Galbraith, K. A., Madha-varapu, S., Fuchs, G. J., Meerloo, T., Farquhar, M. G., Zhou, H., andField, S. J. (2009) GOLPH3 bridges phosphatidylinositol-4-phosphateand actomyosin to stretch and shape the Golgi to promote budding.Cell 139, 337–351
17. Lehrer, S. S., and Kerwar, G. (1972) Intrinsic fluorescence of actin. Bio-chemistry 11, 1211–1217
18. Diensthuber, R. P.,Müller,M., Heissler, S.M., Taft,M. H., Chizhov, I., andManstein, D. J. (2011) Phalloidin perturbs the interaction of human non-muscle myosin isoforms 2A and 2C1 with F-actin. FEBS Lett. 585,767–771
19. Furch, M., Fujita-Becker, S., Geeves, M. A., Holmes, K. C., and Manstein,
D. J. (1999) Role of the salt-bridge between switch-1 and switch-2 of Dic-tyosteliummyosin. J. Mol. Biol. 290, 797–809
20. Bagshaw, C. R. (1975) The kinetic mechanism of the manganous ion-de-pendent adenosine triphosphatase of myosin subfragment 1. FEBS Lett.58, 197–201
21. Taft, M. H., Hartmann, F. K., Rump, A., Keller, H., Chizhov, I., Manstein,D. J., and Tsiavaliaris, G. (2008) Dictyosteliummyosin-5b is a conditionalprocessive motor. J. Biol. Chem. 283, 26902–26910
22. Kurzawa, S. E., andGeeves,M. A. (1996) A novel stopped-flowmethod formeasuring the affinity of actin for myosin head fragments using micro-gram quantities of protein. J. Muscle Res. Cell. Motil. 17, 669–676
23. Nalavadi, V., Nyitrai, M., Bertolini, C., Adamek, N., Geeves, M. A., andBähler,M. (2005)Kineticmechanismofmyosin IXB and the contributionsof two class IX-specific regions. J. Biol. Chem. 280, 38957–38968
24. Struchholz, S., Elfrink, K., Pieper, U., Kalhammer, G., Honnert, U., Grüt-zner, A., Linke, W. A., Liao, W., and Bähler, M. (2009) Functional role ofthe extended loop 2 in the myosin 9b head for binding F-actin. J. Biol.Chem. 284, 3663–3671
25. Heissler, S. M., and Manstein, D. J. (2011) Comparative kinetic and func-tional characterization of the motor domains of human nonmuscle myo-sin-2C isoforms. J. Biol. Chem. 286, 21191–21202
26. Senisterra, G. A.,Markin, E., Yamazaki, K., Hui, R., Vedadi,M., andAwrey,D. E. (2006) Screening for ligands using a generic and high-throughputlight-scattering-based assay. J. Biomol. Screen. 11, 940–948
27. Duhr, S., and Braun, D. (2006) Why molecules move along a temperaturegradient. Proc. Natl. Acad. Sci. U.S.A. 103, 19678–19682
28. Wienken, C. J., Baaske, P., Rothbauer, U., Braun, D., and Duhr, S. (2010)Protein-binding assays in biological liquids using microscale thermopho-resis. Nat. Commun. 1, 100
29. Roy, A., Kucukural, A., and Zhang, Y. (2010) I-TASSER. A unified plat-form for automated protein structure and function prediction. Nat. Pro-toc. 5, 725–738
30. Wu, S., Skolnick, J., and Zhang, Y. (2007) Ab initio modeling of smallproteins by iterative TASSER simulations. BMC Biol. 5, 17
31. Kliche, W., Fujita-Becker, S., Kollmar, M., Manstein, D. J., and Kull, F. J.(2001) Structure of a genetically engineeredmolecularmotor.EMBO J.20,40–46
32. Zolkiewski, M., Redowicz, M. J., Korn, E. D., and Ginsburg, A. (1996)Thermal unfolding of Acanthamoeba myosin II and skeletal muscle myo-sin. Biophys. Chem. 59, 365–371
33. Ponomarev, M. A., Furch, M., Levitsky, D. I., and Manstein, D. J. (2000)Charge changes in loop 2 affect the thermal unfolding of themyosinmotordomain bound to F-actin. Biochemistry 39, 4527–4532
34. Ponting, C. P., Phillips, C., Davies, K. E., and Blake, D. J. (1997) PDZdomains. Targeting signalling molecules to sub-membranous sites.BioEssays 19, 469–479
35. Ranganathan, R., and Ross, E. M. (1997) PDZ domain proteins. Scaffoldsfor signaling complexes. Curr. Biol. 7, R770–R773
36. Onishi, H., Kojima, S., Katoh, K., Fujiwara, K., Martinez, H. M., and Mo-rales,M. F. (1998) Functional transitions inmyosin. Formation of a criticalsalt-bridge and transmission of effect to the sensitive tryptophan. Proc.Natl. Acad. Sci. U.S.A. 95, 6653–6658
37. Várkuti, B. H., Yang, Z., Kintses, B., Erdélyi, P., Bárdos-Nagy, I., Kovács,A. L., Hári, P., Kellermayer, M., Vellai, T., and Málnási-Csizmadia, A.(2012) A novel actin binding site of myosin required for effective musclecontraction. Nat. Struct. Mol. Biol. 19, 299–306
38. Behrmann, E., Müller, M., Penczek, P. A., Mannherz, H. G., Manstein,D. J., and Raunser, S. (2012) Structure of the rigor actin-tropomyosin-myosin complex. Cell 150, 327–338
39. Lowey, S., Saraswat, L. D., Liu, H., Volkmann, N., and Hanein, D. (2007)Evidence for an interaction between the SH3 domain and the N-terminalextension of the essential light chain in class II myosins. J. Mol. Biol. 371,902–913
40. Roth, K., and Weiner, M. W. (1991) Determination of cytosolic ADP andAMP concentrations and the free energy of ATP hydrolysis in humanmuscle and brain tissues with 31P NMR spectroscopy.Magn. Reson. Med.22, 505–511
41. Krisanda, J. M., and Paul, R. J. (1983) Phosphagen and metabolite content
Biochemical Characterization of Human Myosin-18A
30040 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 42 • OCTOBER 18, 2013
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

during contraction in porcine carotid artery. Am. J. Physiol. 244,C385–C390
42. Khromov, A., Somlyo, A. V., and Somlyo, A. P. (1998)MgADP promotes acatch-like state developed through force-calcium hysteresis in tonicsmooth muscle. Biophys. J. 75, 1926–1934
43. Yang, C.-H., Szeliga, J., Jordan, J., Faske, S., Sever-Chroneos, Z., Dorsett, B.,Christian, R. E., Settlage, R. E., Shabanowitz, J., Hunt,D. F.,Whitsett, J. A., andChroneos, Z. C. (2005) Identification of the surfactant proteinA receptor 210as the unconventional myosin 18A. J. Biol. Chem. 280, 34447–34457
44. Matsui, K., Parameswaran, N., Bagheri, N., Willard, B., and Gupta, N.(2011) Proteomics analysis of the ezrin interactome in B cells reveals anovel association with Myo18a�. J. Proteome Res. 10, 3983–3992
45. Dosé, A. C., Ananthanarayanan, S., Moore, J. E., Burnside, B., and Yengo,C.M. (2007) Kineticmechanismof humanmyosin IIIA. J. Biol. Chem. 282,216–231
46. Dosé, A. C., Ananthanarayanan, S., Moore, J. E., Corsa, A. C., Burnside, B.,andYengo, C.M. (2008) The kinase domain alters the kinetic properties ofthe myosin IIIA motor. Biochemistry 47, 2485–2496
Biochemical Characterization of Human Myosin-18A
OCTOBER 18, 2013 • VOLUME 288 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 30041
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Stefan Raunser and Dietmar J. MansteinManuel H. Taft, Elmar Behrmann, Lena-Christin Munske-Weidemann, Claudia Thiel,
F-actin and GOLPH3Functional Characterization of Human Myosin-18A and Its Interaction with
doi: 10.1074/jbc.M113.497180 originally published online August 29, 20132013, 288:30029-30041.J. Biol. Chem.
10.1074/jbc.M113.497180Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/288/42/30029.full.html#ref-list-1
This article cites 46 references, 14 of which can be accessed free at
by guest on October 3, 2016
http://ww
w.jbc.org/
Dow
nloaded from





























