Embed Size (px)
Citation preview

Toxicon, Vol. 33, No. 5, pp. 615-626. 1995 Cop&h1 8 1995 Else&r science Ltd
Printed in Onat Britain. All rigJm reserved 00414101195 59.50 + 0.00
FRACTIONATION OF BOTHROPS PIRAJAI SNAKE VENOM: ISOLATION AND CHARACTERIZATION OF PIRATOXIN-I,
A NEW MYOTOXIC PROTEIN
L. C. MANCUSO,’ M. M. CORREA,’ C. A. VIEIRA,’ 0. A. B. CUNHA,’ J.-J. LACHAT,2 H. S. SELISTRE DE ARAUJ0,3*4 C. L. OWNBY and J. R. GIGLIO’* ‘Dcpartamento de Bioquimica, and *Departamento de Morfologia, Faculdade de Medicina de RiberIo Preto, Universidade de Sfto Paulo, i4049-900 RibairZo Preto, SP, Brazil; 3Departamento de C&&as FisioEgicas, Universidade Federal de S&o Carlos, 13565905 Slo Carlos, SP, Brazil; and ‘Department of Physiological
Sciences, Oklahoma State University, Stillwater, OK 74978, U.S.A.
(Received 30 September 1994; accepted 5 December 1994)
L. C. Mancuso, M. M. Correa, C. A. Vieira, 0. A. B. Cunha, J.-J. Lachat, H. S. Selistre de Araujo, C. L. Ownby and J. R. Giglio. Fractionation of Bothrops pirujai snake venom: isolation and characterization of piratoxin-I, a new myotoxic protein. Toxicon 33, 615-626, 1995.-Whole desiccated venom of Bothrops pirajai was fractionated on a gel filtration (Sephadex G-75) column. Phospholipase AZ, arginine esterase and clotting activity profiles of the six fractions (SI to SVI) obtained were determined. Fraction SIV from the gel filtration column was subjected to chromatography on SP-Sephadex C-25. It was resolved into five subfractions (SIV-SP, to SIV-SP,). Fractions SIV-SP,, SIV-SP, and SIV-SP, showed phospholipase A2 activity but, among these fractions, only SIV-SP, was homogeneous. Induction of myonecrosis by SIV-SP3, SIV-SP, and SIV-SP, was demonstrated by their ability to release serum creatine kinase, and for SIV-SP,, to induce histological alterations in the injected mouse muscle. Chemical characterization by determination of mol. wts, isoelectric focusing and direct manual sequencing of the N-terminal region was performed for SIV-SP,, SIV-SP, and SIV-SP,. When compared with bothropstoxin-I, the myotoxin SIV-SP, showed the same total number of amino acid residues (12 1) and constant molar ratio for all but three amino acids. We have named this toxin piratoxin-I (PrTX-I).
INTRODUCTION
Snakes of the genus Bothrops represent a subject of great scientific, medical and social interest in Brazil, since they are responsible for 85-90% of the total ophidian accidents which occur in the country (Rosenfeld, 1971; Ferreira et al., 1992; Ribeiro et al., 1993).
Bothrops venoms contain several active toxins, of which an important group are the phospholipases A2 (E.C.3.1.1.4). The phospholipases A2 have, in addition to their digestive function, a range of biological activities which may include neurotoxicity; myotoxicity; initiation and/or inhibition of platelet aggregation; hemolytic, anticoagulant, convulsant,
* Author to whom correspondence should be addressed.
615

616 L. C. MANCUSO et al.
hypotensive, cardiotoxic and edema-inducing effects (Kini and Evans, 1989). Other components include coagulant, hemorrhagic, edema-inducing and bradykinin potentiating factors (Selistre et al., 1990a; Cintra et al., 1990).
Myotoxicity is defined as a specific venom action on skeletal muscle, affecting only muscle fibers and leaving other tissue structures such as connective tissue, nerves and vessels essentially unharmed (Mebs and Ownby, 1990). Several myotoxins have been isolated from bothropic venoms including those from Bothrops asper (Gutitrrez et al., 1984a, b; Lomonte and Gutierrez, 1989; Kaiser et al., 1990), B. nummzfer (Gutierrez et al., 1986). B. juraracussu (Homsi-Brandeburgo et al., 1988), B. insularis (Selistre et al., 1990~) and B. moojeni (Lomonte et al., 1990). Some of these myotoxins show anticoagulant activity when assayed on recalcified plasma. One of them, bothropstoxin-I, has recently been sequenced in this laboratory (Cintra et al., 1993).
In this paper we report the isolation and partial characterization of piratoxin-I (PrTX-I), a myotoxin from Bofhrops pirujui, a snake confined to the south of Bahia State, north-eastern Brazil (Hoge and Roman0 Hoge, 1978/1979). PrTX-I is compared with bothropstoxin-I (BthTX-I) from Bothrops jararacussu, a snake found in the central southern part of the country.
MATERIALS AND METHODS
Gel @ration, ion-exchange chromatography and chemical characterization Desiccated whole venom of B. pirajai (500 mg, from Instituto Butantan, SHo Paulo, Brazil) was extracted with
5 ml of 0.1 M ammonium bicarbonate buffer, pH 8.0, cleared by centrifugation for 5 min at 480 x g and subjected to gel filtration on a Sephadex G-75 (Pharmacia) column (3.7 x 115 cm) previously equilibrated and then eluted with the same buffer at 4°C. Fractions of 5 ml were collected at a flow rate of 30 ml/hr using an Ultrorac model 2130 fraction collector (LKB).
Fraction SIV (50 mg) of the gel filtration step was taken up into 3 ml of 0.05 M, pH 6.0 ammonium acetate buffer, cleared by centrifugation as before and applied to the top of a 2.2 x 63 cm column of SP-Sephadex C-25 (Pharmacia) column previously equilibrated and initially eluted with the same buffer at 4°C. Elution proceeded with a continuous convex concentration gradient up to 0.8 M buffer at pH 6.0. At this point, the upper 1 .O M buffer was changed to 2.0 M at pH 6.0. Fractions of 3 ml were collected at 20 ml/hr. Absorbances were determined at 280nm and conductivities were measured with a RC-16C Beckman conductimeter and converted to concentration values by means of a standard curve drawn with known ammonium acetate solutions. After centrifugation, an aliquot of the clear supernatant was used for the determination of protein concentration by the microbiuret method of Itzhaki and Gill (1964).
Details of material sources and subseauent steus including polvacrvlamide gel electronhoresis (PAGE), SDS-PAGE, isoelectric focusing, amino acid composition and d&t manual sequencing of the-l-terminal region have been described previously (Homsi-Brandeburgo et al., 1988). The amino acid composition was determined in an automatic analyzer equipped with a single 0.6 x 22 cm column of Beckman W-3 resin, operation range from 2 to 20 nmoles/residue.
Enzymatic assays Phospholipase A, and esterase activities were determined potentiometrically, using egg yolk and Na-p-tosyl-t-
arginine methyl ester (TAME, Sigma Chemical Co.), respectively, as substrates. Clotting activity was assayed on 0.1% (w/v) bovine fibrinogen in Tris buffer containing 1 mM CaCl, (Selistre and Giglio, 1987; Selistre et al., 199Oa). Proteolytic activity was determined on bovine casein (Merck) at I = 280 nm after 60 min of incubation at 37°C with 25 ng of each sample (Homsi-Brandeburgo et al., 1988).
Anticoagulant.activity was assayed by measuring th; recalcification time of fresh, platelet-poor, rat plasma. The plasma (0.5 ml) was incubated with saline or isolated venom fractions (0.1 ml), at 37°C for 3 min and the time for clotting after addition of 0.1 ml of 0.25 M CaCl, was measured (Selistre et al., 199Ob; Alvarado and Gutierrez, 1988).
Determination of ability of the toxins to induce myonecrosis of skeletal muscle in vivo The liberation of creatine kinase from damaged muscle cells was followed by use of the Granutest 2.5 Plus
(Merck) kit (Oliver, 1955) to measure the enzyme activity in mouse plasma. Three groups of animals (3540 g) were injected in the right gastrocnemius muscle, respectively, with 50 pg (_ 1.5 pg/g) of fractions SIV-SP, (n = 8)

Bothrops pirajai Snake Venom 617
SIV-SP, (n = 6) and SIV-SP, (n = 6) in 25 ~1 of pH 7.4 phosphate-buffered saline (PBS). After 3 hr, blood was collected from the abdominal cava and the serum separated by centrifugation at 480 x g for 3 min at room temperature. Control animals (n = 9) received PBS only.
For the morphological analysis of the alterations induced by SIV-SP, (* 1.5 pg/g), a small portion of the central region of the muscle was excised, placed in a 2.5% (v/v) glutaraldehyde fixing bath in 0.1 M, pH 7.4 phosphate buffer, cut into smaller fragments and then kept in this solution for 2 hr at 4°C. After three washes with the buffer, the fragments were subjected to secondary fixation in 1% (m/v) osmium tetroxide in the same buffer for 2 hr at 4°C. The material was subsequently dehydrated in graded ethanol and processed for embedding in Araldite resin 6005 kit (Ladd Res. Ind. Inc., Burlington, Vermont, U.S.A.). Sections 0.5 pm thick were cut using a DuPont-Sorvall ultramicrotome, stained with 1% (m/v) toluidine blue and examined under a light microscope. Light micrographs were made using a Carl Zeiss axiophot.
In addition experiments, adult female white mice, weighing 25-30 g, were injected i.m. into the dorsolateral aspect of the right thigh with 100 ~1 of SIV-SP, (3 pg/g) in physiologic saline (0.85% NaCl). Control animals received saline only.
Muscle samples were taken from the ventromedial aspect of the injected thigh at 5 hr after injection. The tissue was fixed and processed for embedding in plastic as previously described (Ownby et al., 1990). Initial fixation was for 2 hr at room temperature in 2% (v/v) cacodylate buffered glutaraldehyde (pH 7.4), followed by secondary fixation with 1% osmium tetroxide. Tissue was dehydrated in graded acetone and then embedded in Poly/bed 812 resin (Poly-Sciences, Warrington, PA, U.S.A.). Thick sections (1 pm) were cut from the plastic blocks with an MT-5000 ultramicrotome using glass knives and stained with toluidine blue.
RESULTS
Figure 1‘4 shows the gel filtration profile of B. pirujui snake venom on Sephadex G-75. Six fractions, named SI (a and b) to SVI were obtained. Phospholipase AZ, arginine esterase and clotting activity profiles are also shown.
Based on the enzymatic activity profiles and the elution volume of each fraction (Fig. lA), as well as PAGE migration of respective major components (Fig. 2A), fraction SIV was selected for further studies since its properties fitted well with the physicochemical and functional features of typical bothropic myotoxins such as BthTX-1 (Fig. 2B and C) and the myotoxins from B. insularis snake venom (Selistre et al., 199042). This fraction was resolved into five subfractions (Fig. lB), named SIV-SP, to SIV-SP,, by ion-exchange chromatography on SP-Sephadex C-25. Total protein and PLA, activity recovery data for the several fractions are shown in Table 1. A caseinolytic assay for SIV-SP,, SIV-SP, and SIV-SP, did not show any detectable activity.
The migration distance of SIV-SP, when compared with that of bothropstoxin (Fig. 2B) shows SIV-SP, to run slightly faster. SDS-PAGE in the presence of /I-mercaptoethanol indicates approximate mol. wts of 14,400, 14,800 and 15,500 for SIV-SPs, SIV-SP, and SIV-SP,, respectively. In non-reducing conditions, the mol. wt of SIV-SP, is close to 13,000 (Fig. 3).
Isoelectric focusing of SIV-SP, , SIV-SP, and SIV-SP, indicated an isolectric pH of 8.1, 8.2 and 8.2 for the three fractions, respectively. Figure 2C shows the isolectric focusing of BthTX- 1 and SIV-SP, .
The amino acid composition of SIV-SP, is shown in Table 2. Manual Edman-dansyl degradation of the six first amino acid residues of SIV-SP, to SIV-SP, revealed the following sequences:
SIV-SP, ,: Asp-Leu-Trp-Glx-Phe-Gly . . . . SIV-SP,.: Ser-Leu-Phe-Glx-Leu-Gly . . . . SIV-SPs : Ser-Leu-Phe-Glx-Leu-Gly . . . .
SIV-SP3 1 SIV-SP, and SIV-SPs show anticoagulant activity as indicated by dose-depen- dent recalcification times compared to that of the control.

618 L. C. MANCUSO ef al.
Table 3 shows the time (min) required for clotting of rat plasma at 37°C after incubation with the isolated toxins for 3 min and addition of 0.25 M CaCl, to a final 36mM concentration.
2.5
2.0
1.5
:
Es 1.0
a”
0.5
50 70 90 110 130 150 170 I90 210 230 250 270
TUBE NUMBER
- 0.6
% - 0.5
0 g 0.3
a 0.2
20 40 60 80 100 120 140 160 160 200 220 240 260 280
TUBE NUMBER
Fig. 1. Fractionation of Bothrops pirajai snake venom. (A) Gel filtration of 500 mg of desiccated venom on Sephadex G-75 (3.7 x 115 cm) at 4°C. The gel was equilibrated and eluted with 0.1 M, pH 8.0 ammonium bicarbonate buffer. Flow rate: 30 ml/hr; fraction volume: 5 ml/tube; c.t.: clotting time (se@. (B) Ion-exchange chromatography of 50 mg of fraction SIV (from 1A) on a 2.2 x 63 cm column of SP-Sephadex C-25 at 4°C. Elution started with 0.05 M, pH 6.0 ammonium acetate buffer, followed by a continuous concentration gradient of the
same buffer up to 1.4 M. Flow rate: 20 ml/hr; fraction volume: 3 ml/tube.

Bothrops pirajai Snake Venom 619
Fig. 2. PAGE and electrofocusing of isolated toxins. (A and B) PAGE in b-alanine-acetic acid buffer pH 4.5, for 3 hr at 20 mA, using a polyacrylamide gel concentration of 10% (m/v). A 0.2% (w/v) Coomassie Brilliant Blue G-250 in methanol-water (1: 1, v/v) was used for staining. (A lanes) 1, Whole venom; 2, SIa; 3, Sib; 4, SII; 5, SIII; 6, SIV; 7, SIV-SP,; 8, SIV-SP,; 9, SIV-SP,; 10, SIV-SP,; 11, SW-SP,. (B lanes) 1, SIV-SP,; 2, BthTX-I. (C) Isoelectric focusing on 5% (m/v) polyacrylamide gel. The gel contained 1% Buffalyte (Pharmacia), 0.17% (v/v) TEMED, 10% (m/v) sucrose and 0.075% (m/v) ammonium persulfate. The cathode was preliminary soaked in 1 M NaOH and ‘the anode in 1 M H,PO,. A prerun of 30 min at 100 V and 40 mA was carried out, followed by a two-step run at 750 V (40 to 2 mA) for
135min and 155OV (5 to OmA) for 75 min. Lane 1, BthTX-I; lane 2, SW-SP,.

620 L. C. MANCUSO et al.
Myonecrosis was assayed by injecting mice i.m. with purified subfractions (1.5 pg/g body weight) and determining plasma creatine kinase 3 hr later. The levels of enzyme activity induced by SIV-SP,, SW-SP, and SW-SP, were l&l2 times higher than the control (Table 4). In addition, myonecrosis induced by SIV-SP, (1.5 and 3 pg/g body weight) was followed by histopathological analysis of injected muscle sections (Fig. 4) which were appropriately fixed for light microscopic examination 3 and 5 hr after injection. Examin- ation of the sections of skeletal muscle clearly demonstrates that SIV-SP, causes myonecrosis. There was intercellular edema, and leukocytes were present in the connective tissue after 3 hr (Fig. 5). Some muscle cells had delta lesions (triangular-shaped pale areas beneath a ruptured plasma membrane), and there was no evidence of vascular lesions or hemorrhage. Muscle cells from control mice injected with 0.85% NaCl were normal in appearance.
DISCUSSION
The profile of clotting, esterase and PLA, activities (Fig. 1) of B. pirajai snake venom fits well with those of other bothropic venoms (Seegers and Ouyang, 1979; Iwanaga and Suzuki, 1979; Selistre and Giglio, 1987; Homsi-Brandeburgo et al., 1988; Kini and Evans, 1989: Valiente et al., 1992). Although SIV-SP,, SIV-SP, and SIV-SP, showed significant PLA, activity, SIV-SP, and SIV-SP, were not pure (Fig. 2A), so were not used in further studies. SIV-SP, , SIV-SP, and SIV-SP, , however, showed single PAGE bands and passed the subsequent purity tests: isoelectric focusing (figures for SIV-SP, and SIV-SP, not shown) and N-terminal manual sequence analysis, up to at least the sixth amino acid residue. Not surprisingly, SIV-SP, showed complete sequence homology with BthTX-I (Cintra et al., 1993) in this region. SIV-SP,, like BthTX-I (Homsi-Brandeburgo et al., 1988) is devoid of PLA, activity, at least on the conventional egg-yolk substrate.
Table 1. Total protein and PLA, activity recoveries during the fractionation of Bothrops pirajai snake venom
Protein* Enzymatic activity? Fraction (mg) Ret (%) (U/mg) Total U Ret (%)
Whole venom 293.1 100.0 26.2 7685.3 100.0 SIa 18.4 6.3 - Sib 28.3 9.7 - - SII 20.0 6.8 -
SIII 64.5 22.0 67.7 4364.7 56.8 SIV 121.9 41.6 20.3 2481.7 32.3 sv 20.9 7.1 - - SVI 12.8 4.4 - -
Total 286.8 97.9 6846.4 89.1
SIV 121.9 100.0 20.3 2481.7 100.0 SIV-SP, 7.6 6.2 12.8 96.8 3.9 SIV-SP, 3.1 2.5 10.8 33.2 1.3 SIV-SP, 23.0 18.8 72.2 1659.6 66.9 SIV-SP, 17.5 14.0 residual - - SIV-SP, 57.6 47.3 residual - Total 108.7 89.2 - 1789.6 72.1
* Microbiuret method (Itzhaki and Gill, 1964). t De Haas et a/. (1968).

Bothrops pirajai Snake Venom 621
5.0 - BSA
b
3 iz
3
g Q 4.0 -
s
3.0 I I I I I I I I
1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0
MIGRATION DISTANCE (cm 1
Fig. 3. Molecular weight of SIV-SP, by SDS-PAGE. Mol. wt markers are: bovine serum albumin, BSA (67,000); ovalbumin, OA (44,000); carbonic anhydrase, CA (30,000); citochrome c, Cit. c (12,400). The values found for SW-SP, were 15,500
and 13,000, in the presence and absence of 1% (v/v) /3-mercaptoethanol, respectively.
Based on this lack of PLAz activity, high similarity in the amino acid composition and high sequence homology when compared to BthTX-I (work in progress), SIV-SP, is expected to be also a Lysh9 PLA,-like myotoxin since Asp*, which is required for PLA, activity (Kini and Evans, 1987, 1989) was replaced by Lys,, in BthTX-I (Cintra et al., 1993). Despite the high sequence homology that we expect between SIV-SP, and BthTX-I, a subtle difference in the electrophoretic migration was repeatedly de- tected (Fig. 2B), showing that SIV-SP, is slightly more basic. This conclusion was confirmed by ion-exchange chromatography (SIV-SP; required a higher pH for elution from SP-Sephadex than BthTX-I) and by the amino acid composition (SIV-SP, has one Lys more and two Asx less, besides two additional Ser and one Ala less than BthTX-I). Based on the genetic code, we may speculate that probable point mutations matching with the above results are: Asn c+ Lys, Asn ++ Ser and Alao Ser. In the three cases, the predicted mutations correspond to a single base change in the re- spective mRNA triplets. The mutation Asn t* Lys is probably responsible for the slightly higher basicity of SIV-SP,. A similar point mutation (Ala,% + Pro,,) occurred when a mutant ribitol dehydrogenase evolved in a continuous culture of Klebsiella aerogenes on xylitol (Dothie et al., 1985). The mol. wt of SIV-SP,, approximately 13,000 by SDS-PAGE in the absence and 15,500 in the presence of /3-mercaptoethanol (Fig. 3), and 13,652 from the amino acid composition (Table 2), suggests a structural change in the reduced toxin large enough to produce a lower electrophoretic mi- gration. TOX 3W-c

622 L. C. MANCUSO et al.
Table 2. Amino acid composition of SIV-SP, compared with that of BthTX-I
Amino acid SW-SP, * BthTX-It
Asx 12 14 Thr 5 5 Ser 6 4 Glx 7 7 Pro 6 6 Gly 10 10 Ala 5 6 Half-Cys 14 14 Val 45 4 Met 1 1 Ile 3 3 Leu 10 10 Tyr 8 8 Phe 2 2 His 2 2 LYS 20 19 Arg 5 5 Trp 1 1 Total 121-122 121 Minimum mol. wt 13,652 13,720
* Half-cystine residues were estimated as cysteic acid after oxidation by performic acid, and Trp after hydrolysis with 3 M 2-mercaptoethanesulfonic acid. Values for Ser and Thr were obtained by extrapolation to the intercept at zero time of hydrolysis.
7 Data obtained from the complete sequence of BthTX-I (Cintra et al., 1993).
Asx and Glx stands for Asp +Asn and Glu+Gln, re- spectively.
Myotoxic activity was assayed using two methods: determination of serum creatine kinase, which is released from damaged muscle fibers together with other enzymes and myoglobin, and direct histological examination of sections of the injected muscle. The latter is considered a superior method since other factors besides myonecrosis, such as severe hemorrhage, can induce an increase of serum creatine kinase activity. Furthermore, the sensitivity of the enzyme assay is such that moderate myonecrosis may not lead to significant changes in the level of creatine kinase activity (Mebs and Ownby, 1990). The type of myonecrosis that we observed corresponds closely with that described for other
Table 3. Anticoagulant activity of the isolated myotoxins
Recalcilication time (min) Dose (yg) SIV-SP, SIV-SP, SIV-SP, Control
0.01 16.4 - 7.5 0.04 >45 - 0.16 r45 - 7.8 0.62 245 15.8 11.3 - 1.25 - 16.5 - 2.5 - 16.5 - 2.5 >45 42.7 11.7 -
10.0 - >45 22.8 50.0 - - 34.8 -
100.0 29.5 >45
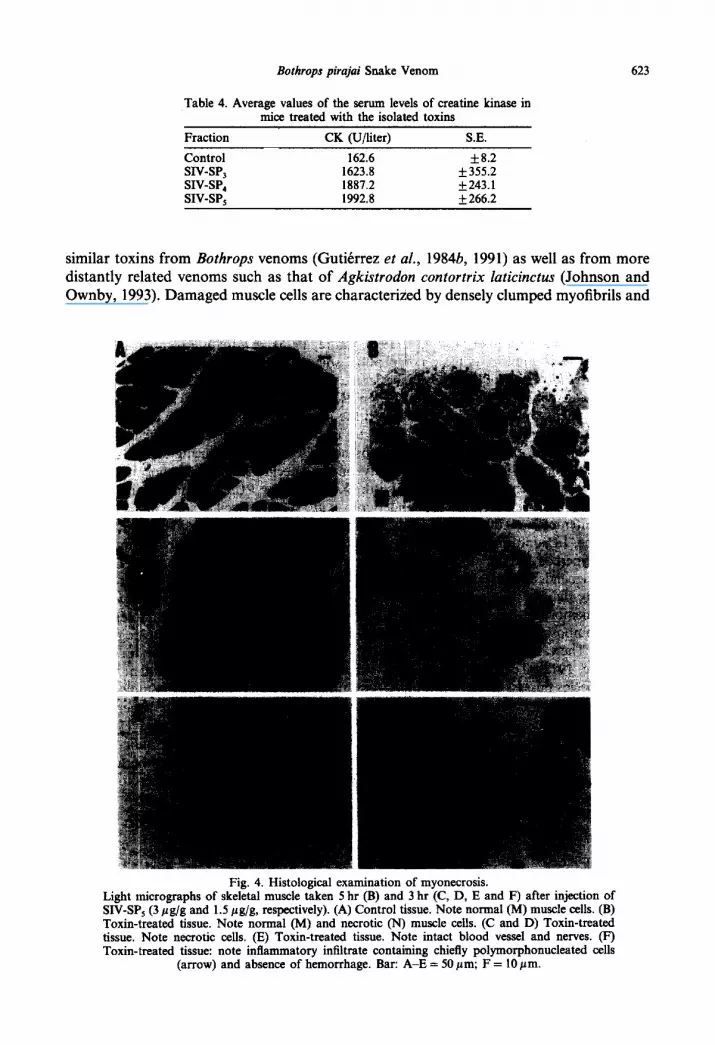
Bothrops pirajui Snake Venom 623
Table 4. Average values of the serum levels of creatine kinase in mice treated with the isolated toxins
Fraction CK (U/liter) S.E.
Control 162.6 *fl.2 SWSP, 1623.8 f 355.2 SIV-SP, 1887.2 k243.1 SIV-SP, 1992.8 f 266.2
similar toxins from Bothrops venoms (Gutitrrez et al., 19846, 1991) as well as from more distantly related venoms such as that of Agkistrodon contortrix luticinctus (Johnson and Ownby, 1993). Damaged muscle cells are characterized by densely clumped myofibrils and
Fig. 4. Histological examination of myonecrosis. Light micrographs of skeletal muscle taken 5 hr (B) and 3 hr (C, D, E and F) after injection of SIV-SP, (3 pg/g and 1.5 pg/g, respectively). (A) Control tissue. Note normal (M) muscle cells. (B) Toxin-treated tissue. Note normal (M) and necrotic (N) muscle cells. (C and D) Toxin-treated tissue. Note necrotic cells. (E) Toxin-treated tissue. Note intact blood vessel and nerves. (F) Toxin-treated tissue: note inflammatory infiltrate containing chiefly polymorphonucleated cells
(arrow) and absence of hemorrhage. Bar: A-E = 50 pm; F = 10 pm.

624 L. C. MANCUSO et al.
ruptured plasma membranes at early time periods (15 min to 6 hr) and by disorganized myofibrils and phagocytic cells at later time periods (12-24 hr).
Typical Brazilian bothropic venoms are usually hemorrhagic (Mandelbaum et al., 1976, 1982, 1984). With these venoms, myonecrosis appears only after massive intramuscular hemorrhage (Queiroz et al., 1985) and is probably secondary to disruption of local microcirculation (Ownby et al., 1978). Histological examination of the SIV-SP, injected skeletal muscle does not reveal any hemorrhage (Fig. 4E), suggesting that muscular lesion occurs through a direct alteration of the cell membrane. The myonecrosis is associated with a pronounced inflammatory reaction and invasion by chiefly polymorphonuclear leuko- cytes (Fig. 4F).
Myotoxicity is apparently independent of proteolytic activity since typical myotoxins, such as bothropstoxin-I itself (Homsi-Brandeburgo et al., 1988) are devoid of this activity. Furthermore, some small, polypeptide myotoxins (4000-5000 mol. wt) such as crotamine (Goncalves and Polson, 1974; Giglio, 1975; Laure, 1975), myotoxin a (Cameron and Tu, 1978) and the homologous toxic polypeptide isolated from the midget faded rattlesnake (Crotulus viridis concolor) venom (Bieber et al., 1987; Engle et al., 1983) are devoid of any enzymatic activity. Within this context, even PLA, activity is apparently not a prerequisite for myotoxicity, but we cannot yet discard the possibility that hydrolytic degradation of the membrane phospholipids may help to disorganize the structure of muscle cell membranes. Also, the several reports of absence of PLAz activity using egg-yolk lecitins as substrate do not exclude PLA, activity on the specific membrane phospholipids of muscle cells. Evidence was found that BthTX-II, another PLA,-like myotoxin from B. jarurucussu snake venom (Homsi-Brandeburgo et al., 1988), as well as some other Bothrops toxins, interact and disrupt the phospholipid bilayer of the skeletal muscle plasma membrane. In addition, an increase in the cytosolic calcium levels induced by myotoxins causes hypercontraction of myofilaments, mitochondrial alterations and activation of calcium-dependent proteases and phospholipases (Gutierrez et al., 1991). It is therefore very likely that the first event in the SIV-SP,-dependent myonecrosis is disruption of the plasma membrane that leads to an influx of calcium into the cytosol and mitochondria.
The composition of the venom of snakes of the same species varies with geographical location. The variability is shown in both the structure and function of the venom proteins (Willemse, 1978). These differences may, ultimately, promote speciation, so that within a genus, species possess toxins with high degree of chemical and functional homology. This is the case with crotamine, from Crotulus durissus terriJicus of South America and myotoxin a from Crotulus viridis viridis from North America (Cameron and Tu, 1977).
According to Kini and Evans (1987), PLAz enzymes exhibiting strong anticoagulant activity possess a specific anticoagulant site around residues 54-77 whereas the myotoxic site is around residues 89-97. The complete sequencing of SIV-SP, (myotoxic and anticoagulant) and SIV-SP, (myotoxic) and attempts to isolate active segments of the polypeptide chain containing these specific sites are in progress in our laboratory. SIV-SP, will be called piratoxin-I (PrTX-I).
Acknowledgements-The authors are indebted to Coordenaclo de Aperfeipoamento de Pessoal de Nivel Superior (CAPES), Conselho National de Desenvolvimento Cientiico e Tecnocologico (CNPq) and Fundacao de Apoio ao Ensimo, Pesquisa e AssistBncia do Hospital das Clinicas da Faculdade de Medicina de Ribeirgo Preto (FAEPA) for financial support. Thanks are also due to Centro Interdepartamental de Quimica de Proteinas (CIQP) and Dr E. C. Arantes for running the amino acid analyses and the electrofocusing, respectively, to Mrs M. I. C. Fonseca for typing and to Dr Richard J. Ward for revising the manuscript.

Bothrops pirajai Snake Venom 625
REFERENCES
Alvarado, J. and Guti&rez, J. M. (1988) Anticoagulant effect of myotoxic phospholipase A, isolated from the venom of the snake Bothrops arper (Viper&e). Bee. Biol. Trop. 36 (2B), 563-565.
Bieber, A. L., McParland, R. H. and Becker, E. R. (1987) Amino acid sequence of myotoxins from Crotalus uiridis concolor. Toxicon 28, 667680.
Cameron, D. and Tu, A. T. (1977) Characterization of myotoxin a from the venom of prairie rattlesnake (CrotaIur viridLs viridis). Biochemistry 16, 2546-2553.
Cameron, D. and Tu, A. T. (1978) Chemical and functional homology of myotoxin a from prairie rattlesnake venom and crotamine from South American rattlesnake venom. Biochim. Biophys. Acta 532, 147-W.
Cintra, A. C. O., Vieira, C. A. and Giglio, J. R. (1999) Primary structure and biological activity of bradykinin potentiating peptides from Bothrops insularis snake venom. J. Protein Chem. 9, 221-227.
Ciitra, A. C. O., Marangoni, S., Oliveira, B. and Giglio, J. R. (1993) Bothropstoxin-I: amino acid sequence and function. .?. Protein Chem. 12, 57-64.
Dothie, J. M., Giglio, J. R., Moore, C. B., Taylor, S. S. and Hartley, B. S. (1985) Ribitol dehydrogenase of Klebsiella aerogenes: sequence and properties of wild-type and mutant strains. Biochem. J. 230, 569-578.
Engle, C. M.., Becker, R. R., Bailey, T. and Bieber, A. L. (1983) Characterization of two myotoxic proteins from venom of Crotalus viridis concolor. J. Toxic.-Toxin Rev. 2, 267-283.
Ferreira, M. L., Moura-da-Silva, A. M., Franca, F. 0. S., Cardoso, J. L. and Mota, I. (1992) Toxic activities of venoms from nine Bothrops species and their correlation with lethality and necrosis. Toxicon 38.1603-1608.
Giglio, J. R. (1975) Analytical studies on crotamine hydrochloride. Ar&yt. Biochem. 69, 207-221: Goncalves, J. M. and Polson, A. (1947) The electrouhoretic analvsis of snake venoms. Arch. Biochem. 13.
253-259. Gutibrrez, J. M., Ownby, C. L. and Odell, G. V. (1984u) Isolation of a myotoxin from Bothrops asper venom:
partial characterization and action on skeletal muscle. Toxicon 22, 115128. Gutierrez, J. M., Ownby, C. L. and Odell, G. V. (1984b) Pathogen&s of myonecrosis induced by crude venom
and a myotoxin of Bothrops asper. Exp. Mol. Path. 48, 367379. Gutierrez, J. M., Lomonte, B. and Cerdas, L. (1986) Isolation and partial characterization of a myotoxin from
the venom of the snake Bothrops nummifer. Toxicon 24, 885-894. Gutibrrez, J. M., Nunez, J., Diaz, C., Cintra, A. C. O., Homsi-Brandeburgo, M. I. and Giglio, J. R. (1991)
Skeletal muscle degeneration and regeneration after injection of bothropstoxin-II, a phospholipase A, isolated from the venom of the snake Bothrops jararacussu. Exp. Mol. Path. 55, 217-229.
Hoge, A. R. and Roman0 Hoge, S. A. R. W. L. (1978/79) Sinopse das serpentes peconhentas do Brasil. Mem. Inst. Butantan 42143, 373-496.
Homsi-Brandeburgo, M. I., Queiroz, L. S., Santo-Neto, H., Rodrigues-Simioni, L. and Giglio, J. R. (1988) Fractionation of Bothrops jararacussu snake venom: partial chemical characterization and biological activity of bothropstoxin. Toxicon 26, 615627.
Itzhaki, R. F. and Gill, D. M. (1964) A micro-biuret method for estimating proteins. Anafyt. Biochem. 9,401-410. Iwanaga, S. and Suzuki, T. (1979) Enzymes in snake venoms. In: Snake Venoms. Handbook of Experimental
Pharmacology, Vol. 52, p. 105 (Lee, C. Y., Ed.). New York: Springer. Johnson, E. K. and Ownby, C. L. (1993) Isolation of a myotoxin from the venom of Agkistrodon contortrix
laticinctus (broad-banded copperhead) and pathogenesis of myonecrosis induced by it in mice. Toxicon 31, 243-255.
Kaiser, I. I., Gutitrrez, J. M., Plummer, D., Aird, S. D. and Odell, G. V. (1990) The amino acid sequence of myotoxic phospholipase from the venom of Bothrops asper. Arch. Biochem. Biophys. 278, 319-325.
Kini, R. M. and Evans, H. J. (1987) Structur&function relationships of phospholipases: the anticoagulant region of phospholipases A*. J. biol. Chem. 262, 14402-14407.
Kini, R. M. and Evans, H. J. (1989) A model to explain the pharmacological effects of snake venom phospholipases A,. Toxicon 27, 613635.
Laure, C. J. (1975) Die primlrstruktur des crotamins. Hoppe-Seyler’s Z. Physiol. Chem. 316, 213-215. Lomonte, B. and Gutitrrez, J. M. (1989) A new muscle damaging toxin, myotoxin II, from the venom of the
snake Bothrops asper (terciopelo). Toxicon 27, 725-733. Lomonte, B., Gutierrez, J. M., Furtado, M. F., Otero, R., Rosso, J.-P., Vargas, O., Carmona, E. and Rovira,
M. E. (1990) Isolation of basic myotoxins from Bothrous mooieni and Bothrops atrox snake venoms. Toxicon _ _ 2% 1137-1146.
Mandelbaum, F. R., Reichl, A. P. and Assakura, M. T. (1976) Some physical and biochemical characteristics of HF2, one of the hemorrhagic factors in the venom of Bothrops jararaca. In: Animal, Plant and Microbial Toxins, Vol. 1, 111-121 (Oshaka, A. E., Hayashi, K. and Sawai, Y., Eds). New York: Plenum Press.
Mandelbaum, F. R., Reichl, A. P. and Assakura, M. T. (1982) Isolation and characterization of a oroteolvtic enzyme from the venom of the snake Bofhrops jararaca (jamraca). Toxicon 20, 955-972. _ _
Mandelbaum, F. R., Assakura, M. T. and Reichl. A. P. (1984) Characterization of the two hemorrhaaic factors isolated from the venom of Bothrops neuwedi (jararaca pintada). Toxicon 22, 193-206.
Mebs, D. and Ownby, C. L. (1990) Myotoxic components of snake venoms: their biochemical and biological activities. Pharm. Ther. 48, 223-236.

626 L. C. MANCUSO et al.
Oliver, I. T. (1955) A spectrophotometric method for the determination of creatine phosphokinase and myokinase. Biochem. J. 61, 116122.
Ownby, C. L., Bjamason, J. and Tu, A. T. (1978) Hemorrhagic toxins from rattlesnake (Crotalw arrox) venom. Pathogenesis of hemorrhage induced by three purified toxins. Am. J. Path. 93, 201-210.
Ownby, C. L., Nika, T., Imai, K. and Sugihara, H. (1990) Pathogenesis of hemorrhagic induced by bilitoxin, a hemorrhage toxin isolated from the venom of the common cantil (Agkistrodon bihneatus bilinearus). Toxicon 28, 837-846.
Queiroz, L. S., Santo Neto, H., Assakura, M. T., Reich, A. P. and Mandelbaum, F. R. (1985) Pathological changes in muscle caused by haemorrhagic and proteolytic factors from Bothrops jararaca snake venom. Toxicon 23, 341-345.
Ribeiro, L. A., Pires de Campos, V. A. F., Albuquerque, M. J. and Takoaka, N. Y. (1993) Acidente ofidico no estado de SBo Paulo. Rev. Ass. Med. Brasil 39, 47.
Rosenfeld, G. (1971) Symptomatology, pathology and treatment of snake bites in South America. In: Venomous Animals and their Venoms, Vol. II, pp. 345-384 (Biicherl, W., Buckley, E. and Deulofeu, V., Eds). New York: Academic Press.
Seegers, W. H. and Ouyang, C. (1979) Snake venom and blood coagulation. In: Snake Venoms. Handbook of Experimenlal Pharmacology, Vol. 52, p. 704 (Lee, C. Y., Ed.). New York: Springer.
Selistre, H. S. and Giglio, J. R. (1987) Isolation and characterization of a thrombin-like enzyme from the venom of the snake Bofhrops insularis (jararaca ilhoa). Toxicon 25, 1135-l 144.
Selistre, H. S., Queiroz, L. S., Cunha, 0. A. B., De Souza, G. E. P. and Giglio, J. R. (1990a) Isolation and characterization of hemorrhagic, myonecrotic and edema-inducing toxins from Bothrops inszdaris (jararaca ilhoa) snake venom. Toxicon Z?, 261-273.
Selistre, H. S., Giglio, J. R. and Homsi-Brandeburgo, M. I. (19906) Atividade anticoagulante de toxinas dos venenos de Bothrops insularis e Bothrops jararacussu. Mem. Inst. Butantan 52 (Suppl.), 73-74.
Valiente, C., Moreno, E., Sittenfeld, A., Lomonte, B. and Gutitrrez, J. M. (1992) An electrophoretic study on phospholipase A, isoenzymes in the venoms of Central American Crotalinae snakes. Toxicon 30, 815-823.
Williense, G. T. (1978) Individual variation in snake venom. Camp. Biochem. Physiol. 61B, 553-557.





























