Embed Size (px)
Citation preview

EXTENSIVE TRAINING IS INSUFFICIENT TO PRODUCE THE WORK-ETHIC EFFECT IN PIGEONS
MARCO VASCONCELOS AND PETER J. URCUIOLI
PURDUE UNIVERSITY
Zentall and Singer (2007a) hypothesized that our failure to replicate the work-ethic effect in pigeons(Vasconcelos, Urcuioli, & Lionello-DeNolf, 2007) was due to insufficient overtraining followingacquisition of the high- and low-effort discriminations. We tested this hypothesis using the original work-ethic procedure (Experiment 1) and one similar to that used with starlings (Experiment 2) by providingat least 60 overtraining sessions. Despite this extensive overtraining, neither experiment revealed asignificant preference for stimuli obtained after high effort. Together with other findings, these datasupport our contention that pigeons do not reliably show a work-ethic effect.
Key words: work ethic, effort, within-trial contrast, overtraining, key peck, pigeons
_______________________________________________________________________________
In the original demonstration of the work-ethic effect (Clement, Feltus, Kaiser, & Zentall,2000), pigeons learned two simultaneous (S+vs. S2) discriminations, one preceded by afixed ratio (FR) 1 requirement to a center-keystimulus (low-effort trials) and the otherpreceded by an FR 20 requirement to thesame stimulus (high-effort trials). After acquir-ing both discriminations to high levels ofaccuracy and 20 subsequent overtrainingsessions, pigeons received periodic nondiffer-entially reinforced probe trials on which theyhad to choose between the high- and low-effortS+ stimuli and, on separate trials, the high-and low-effort S2 stimuli. Clement et al. founda significant preference both for the high-effort S+ and the high-effort S2, the work-ethiceffect (see also Kacelnik & Marsh, 2002;Friedrich & Zentall, 2004).
This result has been followed by a series ofother findings from the Zentall lab showing thatpigeons also prefer stimuli preceded by longover short delays (DiGian, Friedrich, & Zentall,2004), by the absence versus presence of food(Friedrich, Clement, & Zentall, 2005), and by arelatively aversive reinforcement contingency(Singer, Berry, & Zentall, 2007). Zentall andSinger (2007b; see also Clement et al., 2000)propose that this pattern of results reflects
greater conditioned reinforcing value of thepreferred stimuli arising from a larger contrastbetween the aversive states preceding them andthe reinforcing effect that follows them.
Be that as it may, the original work-ethiceffect has proved to be elusive. For example,we (Vasconcelos, Urcuioli, & Lionello-DeNolf,2007) were unable to replicate the effect in sixseparate experiments, and a similar failure-to-find was recently reported by Arantes andGrace (2008). Michael Colombo (personalcommunication, July 2, 2008) has also report-ed to us that his lab has been unable toreplicate the effect with pigeons. Indeed,Zentall and Singer (2007a) mention an un-published study by Klein and Zentall (2002)that was unable to reproduce the original(Clement et al., 2000) findings.
Zentall (2008) and Zentall and Singer(2007a) suggest that failures to replicate thework-ethic effect are probably due to insuffi-cient overtraining and cite data from relatedstudies in support of this (e.g., Friedrich &Zentall, 2004; Singer et al., 2007). Even thoughthe original demonstration employed 20 over-training sessions (the same number used byVasconcelos et al., 2007), they argued that 2 or3 times that number is probably required toreliably obtain preferences for high- over low-effort stimuli. According to them, the neces-sary within-trial contrast between prior eventsand the eventually preferred stimuli developsrather slowly.
In order to determine if this might indeedexplain most of the discrepancies betweenpigeon studies of the work-ethic effect, thepresent paper tested the extensive-overtrain-ing hypothesis in two different paradigms.
Preparation of this article was made possible by NationalInstitute of Mental Health Grant MH 66195 to Peter J.Urcuioli.
Correspondence concerning this article should beaddressed to Marco Vasconcelos or Peter J. Urcuioli,Department of Psychological Sciences, Purdue University,703 Third Street, West Lafayette, IN 47907-2081 (e-mail:[email protected] and [email protected]).
doi: 10.1901/jeab.2009.91-143
JOURNAL OF THE EXPERIMENTAL ANALYSIS OF BEHAVIOR 2009, 91, 143–152 NUMBER 1 (JANUARY)
143

Experiment 1 was conducted using the proce-dure described by Clement et al. (2000);Experiment 2 used an adaptation of Kacelnikand Marsh’s (2002) procedure with Europeanstarlings. In both experiments, pigeons receivedat least 60 overtraining sessions before theirpreference tests. Neither experiment, however,returned data indicating that extensive over-training yields a work-ethic effect like thatoriginally reported by Clement et al. (2000).
EXPERIMENT 1
Pigeons were trained on two simultaneousdiscriminations, one preceded by a low-effortresponse requirement (FR 1) to one initial-link stimulus and the other preceded by ahigh-effort response requirement (FR 30) to adifferent initial-link stimulus. Signaling thedifferent work requirements should enhancethe probability of obtaining the work-ethiceffect (cf. Zentall & Singer, 2007b). Afterpigeons acquired each discrimination to highlevels of accuracy, they received 60 overtrain-ing sessions and were then tested for theirpreferences for high- over low- effort stimuli.
METHOD
Subjects
Six experimentally naı̈ve White Carneaupigeons obtained from the Palmetto PigeonPlant (Sumter, SC) were used. All were retiredbreeders and had continuous access to PurinaProGrains upon arrival in the lab in order todetermine their free-feeding body weights.Each pigeon’s weight was then graduallyreduced to and maintained at 80% of itsfree-feeding value by restricted feeding. Dur-ing experimental participation, food was pro-vided in the home cage only when food intakein a session was insufficient to maintain the80% body weight and on days on which theexperiment was not run.
Each pigeon was housed individually instainless-steel, wire-mesh cages in a tempera-ture- and humidity-controlled colony room ona 14:10 day-night cycle. The light portion ofthe cycle began at 07:00 hours. Grit and waterwere always available in the home cage.
Apparatus
A standard operant chamber consisting of aBRS/LVE (Laurel, MD) Model PIP-016 panel
inside a BRS/LVE Model SEC-002 enclosurewas used in this experiment. On the panelwere three horizontally aligned 25-mm-diame-ter response keys spaced 57 mm apart, centerto center, and positioned 75 mm from the topof the panel. Inline projectors (BRS/LVEModel IC-901-IDD) located behind the leftand right side keys were equipped with filtersfor displaying red, green, yellow, and bluehues. The projector behind the center keycontained a film for displaying a small whitedot and three white vertical or horizontal lineson black backgrounds (BRS/LVE Pattern No.692). A rear-mounted food hopper containingPurina ProGrains was accessible through a 58-mm-square opening located 130 mm below thecenter key. When the food hopper was raised,a small miniature bulb (ESB-28) in the metalhousing surrounding the hopper was lit. Apartially covered GE #1829 bulb located 76mm above the center key was used forchamber illumination; the opening in its metalcover directed light toward the ceiling. Venti-lation and masking noise was provided by aconstantly running blower fan attached to thechamber. All experimental events were con-trolled and recorded by an IBM-compatiblecomputer connected to the chamber throughan interface built in-house.
Procedure
The general procedure and proceduraldetails were virtually identical to that forGroup Line in Vasconcelos et al. (2007,Experiment 5), except that an FR 30 ratherthan an FR 40 requirement was used on high-effort trials. A condensed summary is providedbelow.
All pigeons were initially trained to eatreliably from the food hopper, after whichtheir key peck response to a small white dot onthe center key was shaped by the method ofsuccessive approximations. Over the next 12preliminary training sessions, pigeons ob-tained food by pecking one center-key linestimulus on a continuous reinforcement (FR1) schedule and the other line stimulus on aFR schedule whose parameter was raised from2-30 across successive sessions. The line–schedule pairings were counterbalanced acrossbirds, and the final multiple FR 1 FR 30schedule was in effect for two sessions. Each60-trial session contained an equal number of
144 MARCO VASCONCELOS and PETER J. URCUIOLI

randomized presentations of the vertical andhorizontal lines.
Next, pigeons were trained on two concur-rent simultaneous discriminations, one pre-ceded by a single peck (FR 1) to either thevertical or horizontal lines on the center key(low-effort trials) and the other by 30 pecks(FR 30) to the alternative center-key lines(high-effort trials). Completion of the require-ment on each trial turned off the line stimulusand immediately produced two hues (red andyellow, or blue and green) on the left and rightside keys, with the positions of the hues ineach set counterbalanced across trials. One setwas presented on FR 1 trials (low-effort S+ andlow-effort S2); the other set was presented onFR 30 trials (high-effort S+ and high-effortS2). A single peck to either hue within eachset turned off the other and initiated a 6-speriod that ended in food reinforcement if theS+ had been pecked or in nonreinforcement ifthe S2 had been pecked. Reinforcer durationwas constant within a session but varied from 2s through 6 s across sessions, as needed, inorder to maintain pigeons’ body weights at80% of their free-feeding values. The nonre-inforcement (timeout) period following an S2choice was always equal to the reinforcerduration.
The two simultaneous discriminations werepresented equally often in random order ineach 96-trial training session, with the con-straint that none of the four possible trial types(FR 1 vs. FR 30 x two left-right positions of thehue discriminative stimuli) occurred morethan three times in a row. The houselightwas only on during the 10-s intertrial intervalthat separated successive trials; it was turnedoff at the start of each trial and remained offuntil the end of the reinforcement or timeoutperiod. Each pigeon was trained to a criterionof at least 90% correct on both discriminationswithin a single session. Sixty overtrainingsessions then followed.
Five preference-test sessions followed over-training. Each session included 36 test trialsrandomly intermixed among 60 baseline train-ing trials. Half of the test trials involved achoice between the two S+ stimuli fromtraining (low-effort S+ vs. high-effort S+) andthe other half involved a choice between thetwo S2 stimuli (low-effort S2 vs. high-effortS2). Twelve test trials (6 with the two S+stimuli and 6 with the two S2 stimuli) were
preceded by completion of the FR 1 require-ment to its corresponding center-key linestimulus, another 12 were preceded by com-pletion of the FR 30 requirement to itscorresponding line stimulus, and the remain-ing 12 involved the presentation of the two S+or two S2 stimuli without any center-keyresponse requirement (i.e., at the end of theITI). Choices on the test trials were nondiffer-entially reinforced 50% of the time. Type Ierror rate for all statistical tests reported hereand in the following experiment was set at .05.
RESULTS AND DISCUSSION
Pigeons learned the two simultaneous dis-criminations in an average of 3.2 sessions. Onepigeon was dropped from the experimentafter 30 overtraining sessions because itrepeatedly stopped for 3 hr or more at theonset of a high-effort (FR 30) trial. Duringovertraining, pigeons continued to performaccurately on both discriminations, averaging99.4% and 99.3% correct on low- and high-effort trials, respectively, over the last five ofthese sessions, F(1, 4) 5 0.63. Likewise, theyperformed accurately on both hue discrimina-tions during testing, averaging 96.0% and96.7% correct on low- and high-effort trials,respectively, on the first test session and 91.2%and 96.1% correct, respectively, over all fivetest sessions, F(1, 4) 5 1.00 and 3.45,respectively, ns.
Figure 1 shows each pigeon’s preference forthe high-effort S+ and for the high-effort S2averaged across the five test sessions. Thebottom right panel plots the correspondingpreferences averaged across all pigeons. Pref-erences are broken down by initial-link re-quirement: no previous stimulus (nps), FR 1,and FR 30. The only indication of a consistentpreference across pigeons was for the high-effort (FR 30) S+ on test trials initiated by a FR30 to the initial-link stimulus, t(4) 5 2.91, p ,.05. There was no corresponding evidence of aconsistent preference for that S+ on trialsinitiated by a FR 1 or trials that occurred withno previous (initial-link) stimulus, t (4) 5 0.19and 20.39, respectively. On S2 trials, 4 of the5 pigeons preferred the FR 30 S2 when testtrials were initiated by an FR 1 to the initial-link stimulus; the remaining pigeon (S3)showed a strong preference in the oppositedirection (i.e., for the FR 1 S2) on these trials.Overall, there was no evidence of a significant
EXTENSIVE TRAINING AND THE WORK-ETHIC EFFECT 145
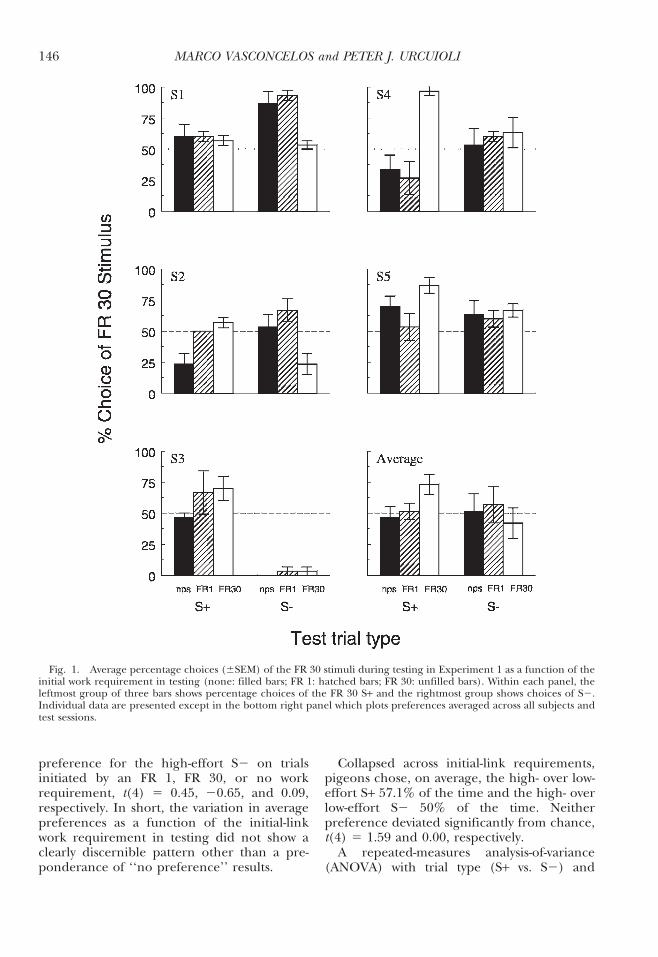
preference for the high-effort S2 on trialsinitiated by an FR 1, FR 30, or no workrequirement, t(4) 5 0.45, 20.65, and 0.09,respectively. In short, the variation in averagepreferences as a function of the initial-linkwork requirement in testing did not show aclearly discernible pattern other than a pre-ponderance of ‘‘no preference’’ results.
Collapsed across initial-link requirements,pigeons chose, on average, the high- over low-effort S+ 57.1% of the time and the high- overlow-effort S2 50% of the time. Neitherpreference deviated significantly from chance,t(4) 5 1.59 and 0.00, respectively.
A repeated-measures analysis-of-variance(ANOVA) with trial type (S+ vs. S2) and
Fig. 1. Average percentage choices (6SEM) of the FR 30 stimuli during testing in Experiment 1 as a function of theinitial work requirement in testing (none: filled bars; FR 1: hatched bars; FR 30: unfilled bars). Within each panel, theleftmost group of three bars shows percentage choices of the FR 30 S+ and the rightmost group shows choices of S2.Individual data are presented except in the bottom right panel which plots preferences averaged across all subjects andtest sessions.
146 MARCO VASCONCELOS and PETER J. URCUIOLI

initial-link work requirement as factors wasalso run to evaluate trial-specific preferences.This ANOVA yielded no significant effects oftrial type, F(1, 4) 5 0.26, or initial-link workrequirement, F(2, 8) 5 0.50, but a significantinteraction between the two, F(2, 8) 5 10.57.This interaction likely reflects the significantpreference for the high-effort S+ stimulus butnot for the high-effort S2 stimulus on trialsinitiated by an FR 30 initial-link requirement.
Visual inspection of Figure 1 indicates thatthe preference results from Pigeon S3 mayhave been atypical and, thus, masked anoverall effect on the S2 preference trials.However, even with this bird removed fromthe analyses, there was no significant overallpreference for the high-effort S2, t(3) 5 1.92.Excluding this bird also did not change theresults of the repeated-measures ANOVA: nosignificant main effect of trial type, F(1, 3) 51.24, or initial-link work requirement, F(2, 6)5 1.36, but a significant interaction betweenthe two, F(2, 6) 5 15.05. Again, this reflects thespecificity of the high-effort S+ preference ontest trials initiated by a FR 30 initial-linkrequirement.
ANOVA on the first-session test results (notshown) revealed no significant effect of trialtype or initial-link requirement, F(1, 4) 5 0.31and F(2, 8) 5 3.17, respectively, or of theirinteraction, F(2, 8) 5 0.51. Pigeons preferredthe high-effort S+ and the high-effort S262.2% and 55.6% of the time, respectively,on the first session. Neither preference dif-fered significantly from chance, t(4) 5 1.54and 0.42, respectively. The results of theseanalyses were unchanged when the data fromPigeon S3 were excluded.
These results, once again, underscore theelusiveness of the work-ethic effect and dem-onstrate that extensive overtraining is insuffi-cient to produce the effect even using asignaled-effort procedure to optimize thechances of observing preference for a high-effort stimulus.
EXPERIMENT 2
Given our repeated failures to find a work-ethic effect in pigeons using the proceduredescribed by Clement et al. (2000), we decidedto explore the issue further using a differenteffort manipulation. In particular, we adaptedKacelnik and Marsh’s (2002) effort-based
procedure with starlings. In their experiment,starlings flew back and forth between twoperches located 1 m apart either 4 times or 16times. Food reinforcement was obtained bypecking one hue after 4 flights and by peckinganother hue after 16 flights. Periodically,starlings received reinforced probe trials inwhich they chose between the two S+ stimuliencountered during training. Kacelnik andMarsh found a statistically significant prefer-ence for the S+ preceded by 16 flights over theS+ preceded by 4 flights.
In our version of this procedure, we used a‘‘long box’’ containing two intelligence panels930 mm apart. Pigeons had to walk from onepanel to the other either 4 times (low-efforttrials) or 16 times (high-effort trials) in orderto obtain a discriminative stimulus that, whenpecked, produced food. Different S+ stimulifollowed the 4 and 16 runs. Pigeons were thenprobed for their S+ preferences after differentamounts of overtraining. As in Experiment 1and as in Kacelnik and Marsh’s (2002)procedure, different stimuli signaled the dif-ferent, forthcoming work requirements.
METHOD
Subjects
Four White Carneau pigeons obtained fromthe same supplier in Experiment 1 and housedand maintained as previously described servedin this experiment. All had limited experienceunrelated to the present contingencies.
Apparatus
A locally made operant chamber with astandard 3-key intelligence panel (BRS/LVEModel PIP-016) on each end was used. Thepigeon’s compartment (368 mm high 3 930mm wide 3 343 mm deep) was equipped withan aluminum grid floor. Three clear plasticpecking keys, 25 mm in diameter and spaced83 mm center-to-center, were about 250 mmabove the floor on each panel. Each key had aBRS/LVE Model IC-901-IDD in-line projectormounted behind it for displaying visual stim-uli. The center-key projectors could display awhite vertical line and an open white triangleboth on a black background (BRS/LVEPattern No. 696). The side-key projectorscould display red and green homogeneouscolored fields. Partially covered GE #1829bulbs located 57 mm above the center key on
EXTENSIVE TRAINING AND THE WORK-ETHIC EFFECT 147

each panel served as houselights. PurinaProGrains could be delivered in one of twomagazines (one on each panel) which, whenraised, were accompanied by lighting a smallminiature bulb (ESB-28) in a metal housingthat covered them. A 58-mm-square openingcentered approximately 130 mm below thecenter key on each panel allowed access to theback-mounted magazine. Ventilation andmasking noise were provided by two continu-ously running blower fans attached to thechamber. An IBM-compatible 386 computerlocated in an adjacent room controlled andrecorded all experimental events via a locallymade interface.
Procedure
Preliminary training. Pigeons were initiallytrained to obtain food by pecking at the redand green stimuli appearing on either side keyof the right panel, with side randomly inter-mixed within a single session. Next, pigeonswere trained to peck the vertical line and thetriangle on the center key of both panels. Inthese 60-trial preliminary training sessions, asingle peck to the stimulus on each trialturned it off and produced 3 s access to foodon the same panel on which the stimulusappeared. The houselight was on duringstimulus presentation and off during foodreinforcement and the ensuing 10-s ITI. Onlythe houselight located on the panel on whichthe stimuli appeared was operative duringthese sessions.
Next, birds were taught to alternate (walk)between the two panels. All trials began andterminated on the right panel. At the begin-ning of each trial, the right houselight and theright-panel center key were lit. A single peck tothe center-key stimulus turned it and the righthouselight off and lit the houselight and thecenter key on the opposite (left) panel. Again,a single peck at the left center-key stimulusturned it and the left houselight off and lit thehouselight and the center key of the rightpanel. This cycle was repeated until therequired number of runs between the twopanels was met. Completion of the require-ment turned off the houselight and produced2 to 6 s access to food, adjusted individually foreach pigeon in each session so as to maintainits 80% body weight. There were 60 trials persession separated by a dark 10-s ITI. On half ofthese trials, the center-key stimulus was the
vertical line; on the remaining half, it was theopen triangle. The number of runs required tocomplete a trial was gradually raised from 2 to8 across four sessions.
Baseline training I. During baseline train-ing, pigeons had to complete either 4 runs(low-effort trials) or 16 runs (high-effort trials)between the two panels. Each trial began withthe onset of the right houselight and theillumination of either a vertical line or an opentriangle on the right-panel center key. A singlepeck terminated this stimulus and the righthouselight and lit the left houselight and theleft-panel center key with the same stimulus.This cycle repeated up to the last run. On thefinal run, pecking the left-panel center-keystimulus turned it and the left houselight offand illuminated the right houselight and oneof the right-panel side keys (counterbalancedacross trials) with either red or green, depend-ing on the running requirement. A single peckto red (or green) turned it and the righthouselight off and produced 2 to 6 s access tofood, adjusted individually for each pigeon ineach session as before. A 10-s ITI withhouselights off then commenced.
For half of the pigeons, the red hueappeared on high-effort trials and the greenhue appeared on low-effort trials; for theremaining half, these contingencies werereversed. Likewise, for half of the birds, thevertical line appeared on the center keys onhigh-effort trials and the triangle appeared onthe center keys on low-effort trials, and viceversa for the remaining half. The two trialtypes occurred in a random order with theconstraints that each occurred 30 times per 60-trial session and that none occurred morethan three times in a row.
Preference testing I. After 25, 50, and 60baseline training sessions, pigeons received atest session in which they were probed for theirpreference for the high- versus low-effort S+.Each session included 12 probe-test trialsrandomly intermixed among 48 baseline train-ing trials, with the constraints that each baselinetrial type occurred at least once before the firsttest trial and that no two test trials occurred in arow. Four test trials were preceded by 4 runs,another four by 16 runs and, on the remainingfour, the two S+ stimuli simply appeared on theside keys at the end of the ITI (i.e., no runrequirement). On these trials, a single peck toeither S+ immediately turned off both stimuli
148 MARCO VASCONCELOS and PETER J. URCUIOLI

and the right houselight and produced foodwith a .5 probability independently of thepigeon’s choice. All other details were the sameas in baseline training.
Baseline training II. Next, the baselinecontingencies were reversed. Both the center-key stimulus and the S+ previously presentedon high-effort trials were now assigned to thelow-effort trials, and vice versa for the stimuliinitially presented on low-effort trials. All otherdetails for these 70 sessions were the same as inBaseline Training I.
Preference testing II. Pigeons were againtested in a single session for their preferencebetween the two S+ stimuli after 20, 50, and 60baseline training sessions. After 70 sessions, 5test sessions were given. Details of thesesessions were identical to Preference Testing I.
RESULTS AND DISCUSSION
The average latencies to peck the initial,center-key stimulus on low- and high-efforttrials over the five training sessions prior to thefirst test session were 2.65 s and 21.13 s,respectively. This latency difference was signif-icant, F(1, 3) 5 26.86, as were the correspond-ing differences for the five training sessionspreceding the other two prereversal sessions,Fs(1, 3) 5 56.40 and 21.87. After the baselinecontingencies were reversed, the averagedifference in center-key-peck latencies priorto the first test session approached signifi-cance, F(1, 3) 5 8.66, p 5 .06, and wassignificant for the baseline sessions precedingthe remaining test sessions, smallest F(1, 3) 511.15. Overall, these analyses confirm thatpigeons could discriminate the forthcominglow- versus high-effort requirement uponseeing the center-key stimulus.
Figure 2 shows each pigeon’s preference forthe high-effort S+ during pre- and postreversaltests broken down according to the runrequirement preceding the appearance ofthe high- versus low-effort S+ stimuli. We plotthe data only from the tests following 25 and60 (or 70) pre- and postreversal trainingsessions1 because repeated-measures ANOVAs
revealed no significant effect of test session, allFs , 1.00 (see Vasconcelos & Urcuioli, 2008,Figure 1, for the average results from theremaining test sessions).
The initiating run requirement had nosignificant effects on the preferences observedin any test, all Fs(2, 6) , 1.00. Moreimportantly, there was no consistent between-subject pattern of preference in either the pre-or postreversal tests. Likewise, there was noconsistent pattern of preference within sub-jects in their pre- versus postreversal tests.Throughout a total of 11 test sessions, none ofthe preferences for the high-effort stimulusdeviated significantly from chance, all ts(3) $21.57 and # 1.00.
1 The data at 70 training sessions after reversal corre-spond to the first of the five test sessions that were given. Arepeated-measures ANOVA with test session and initiatingevent as factors confirmed that neither had a significantinfluence on pigeons’ choices, F(4, 12) 5 0.28 and F(2, 6)5 1.68, respectively.
Fig. 2. Percentage choices of the high-effort (16-runs)stimulus for each bird during testing in Experiment 2 as afunction of the initial work requirement. The left panelpresents preferences after 25 and 60 prereversal trainingsessions. The right panel presents the data after 25 and 70postreversal training sessions.
EXTENSIVE TRAINING AND THE WORK-ETHIC EFFECT 149

In short, extensive overtraining using anadaptation of Kacelnik and Marsh’s (2002)procedure for pigeons did not produce signif-icant preferences for a high-effort S+, contraryto Kacelnik and Marsh’s findings with star-lings. Given that we ran only 4 pigeons(compared to the 12 starlings used by Kacelnikand Marsh), a legitimate concern is that wehad insufficient statistical power to detect awork-ethic effect. However, if the variabilityobserved here is indicative of the variability wewould have seen with other pigeons, theresults would have been substantially the same.
Of course, the use of different species(starlings vs. pigeons) and different responses(fly vs. walk) makes comparisons betweenexperiments complicated. It is known, forinstance, that flying costs much more energythan walking for starlings (Bautista, Tinber-gen, & Kacelnik, 2001). The issue, however, ishow effortful (energetically costly) are differ-ent amounts of walking for pigeons and howthese map onto different degrees of aversive-ness (Zentall & Singer, 2007b). Our manipu-lation was not predicated on absolute costsbut, rather, on the relative aversiveness ofwalking 16 times between panels in thecontext of the alternative 4 walks. Shapiro,Siller, and Kacelnik (2008), for example, havefound that the latency to accept food sourcesthat differ in amount and delay to reward isaffected not only by the rate (amount/delay)associated with the present option, but also bythe rate associated with the alternative option.Briefly, latencies to accept a currently present-ed option increased with higher alternativereward rates. The fact that pigeons were muchslower to initiate a high-effort than a low-efforttrial clearly indicates that they were sensitive tothe different walking requirements. If thoselatency differences also reflect differentialaversiveness, then within-trial contrast shouldhave yielded preferences for the high-effort S+(e.g., Zentall & Singer, 2007b). But no suchpreferences were observed.
GENERAL DISCUSSION
In two different paradigms, we examinedthe possibility that pigeons would show pref-erences for stimuli following high effort overstimuli following low effort if they receivedextensive overtraining after discriminationacquisition (Experiment 1) or extensive train-
ing with singly presented S+ stimuli (Experi-ment 2). Experiment 1 was closely modeledafter Clement et al.’s (2000) procedure withtwo exceptions. First, work requirements weresignaled. According to Zentall and Singer(2007b), this should increase the chances ofobserving such preferences. Second, an FR 30instead of an FR 20 was used as the initial-linkrequirement on high-effort trials, increasingthe disparity between the high- and low-efforttrials. Experiment 2 used an effort-basedmanipulation that had previously proveneffective in producing preferences for a high-effort S+ in starlings (Kacelnik & Marsh, 2002).Our version of their procedure requiredpigeons to walk from one end of a long boxto the other end different numbers of times(again, signaled by different initial-link stimu-li) in order to obtain the terminal-link S+stimuli. The results of these two experimentswere exactly the same: Pigeons did not exhibitan overall preference for the high-effortdiscriminative stimuli from training.
There was some variation in preference as afunction of the initial-link requirement, butthis was mostly inconsistent across pigeons.Indeed, the only statistically significant varia-tion was found in Experiment 1 where pigeonspreferred the high- over low-effort S+ whenthose stimuli appeared after the high-effort(FR 30) requirement. This suggests thattraining may have resulted in a conditionaldiscrimination in which choice of a particularS+ was cued by the initial-link requirementthat preceded it. But even here, the data areinconsistent given that there were no corre-sponding preferences for the low- over high-effort S+ when they appeared after the low-effort (FR 1) or no requirement. Preferencevariations as a function of initial-link require-ments have been previously noted by us(Vasconcelos et al., 2007) and by Arantes andGrace (2008), but neither a consistent nor asingle pattern emerges. Coupled with the lackof such variation in Clement et al. (2000),there does not appear to be a reliable effecthere either, which should not be surprisinggiven that successful baseline performancesdid not require a conditional discrimination.
One might surmise that our findings of nooverall preference simply mean that what wecall ‘‘low effort’’ and ‘‘high effort’’ are, in fact,not functionally different for pigeons. Thisconcern can be answered in a number of ways.
150 MARCO VASCONCELOS and PETER J. URCUIOLI

First, if these schedules are not functionallydifferent along the dimension(s) importantfor within-trial contrast (Zentall & Singer,2007b), then this clearly supports our conten-tion (Vasconcelos et al., 2007) that the originalclaim of a work-ethic effect in pigeons (Clem-ent et al., 2000) was a Type I error. It wouldalso bring into question the reliability of other,similar findings (e.g. Friedrich & Zentall,2004). Second, number of pecks necessary tocomplete the high-effort (FR 30) requirementin Experiment 1 was 50% greater than thenumber of pecks necessary to complete thehigh-effort (FR 20) requirement in Clement etal. (2000) and our attempted replications ofthat study (Vasconcelos et al., 2007, Experi-ments 1 through 4). This increase shouldenhance the probability that the two initial-link schedule requirements would be func-tionally different. Third, the literature indi-cates that a large FR requirement has aversivequalities for pigeons, especially when alternat-ed within a session with a smaller FR require-ment (Flory, 1969; see also Azrin, 1961, andThompson, 1965). In view of this, it would berather surprising if there were no differentialaversiveness associated with a 30:1 differencein signaled initial-link work requirements.Moreover, we observed that the pigeons inExperiment 1 often paused for very lengthyperiods of time on FR 30 trials (indeed, onepigeon had to be dropped from the experi-ment because it routinely stopped respondingaltogether on these trials), indicating a cleardifference between this schedule and the FR 1schedule.
Questions about functional differences, orlack thereof, may be even more apropos toExperiment 2. Here, our manipulation waspredicated on the assumption that walkingback and forth between the ends of a long box4 versus 16 times for pigeons was akin to flyingback and forth 4 versus 16 times betweenperches for starlings (Kacelnik & Marsh,2002). The two types of responding may bevastly different in energetic consequences, aswe previously acknowledged, but we can restassured that the work requirement differencefor pigeons was large enough to have a majoreffect on their behavior: They took eight timeslonger, on average, to initiate walking on thehigh-effort trials than on the low-effort trials.Indeed, this measure allows us to avoid thecircularity of postulating differential aversive-
ness on the basis of the very preference resultsthat those differences are meant to explain. Ofcourse, we did not obtain the predictedpreference for a high-effort S+ (Zentall &Singer, 2007b) despite pigeons’ long-latencyreactions to the initial-link stimulus thatsignaled it.
Both sets of results are consistent withprevious findings (Arantes & Grace, 2008;Vasconcelos et al., 2007) and indicate thatcontrary to Zentall and Singer’s hypothesis(2007a), insufficient overtraining is not re-sponsible for failures to replicate the work-ethic effect in pigeons. Zentall (2008) hasrecently argued that previous experience withlean schedules of reinforcement may beanother factor contributing to the null find-ings that we and others have reported. Again,both experiments reported here speak againstthis argument given that our pigeons hadeither no prior experimental history (Experi-ment 1) or had experience with matching-to-sample (Experiment 2) which, at asymptote,involves nearly continuous reinforcement.
At least three different sets of researchershave now reported a failure to replicate thework-ethic effect in pigeons: ourselves (Vascon-celos et al., 2007; the present experiments),Arantes and Grace (2008) and Colombo(personal communication, July 2, 2008). Inclosing, we should point out that the naturalreaction of wondering why the effect is foundby some (e.g., Clement et al., 2000) but not byothers presupposes that there is an effect to befound. However, if the original work-ethicdemonstration was indeed a Type I error, thereis no effect to be found and no conflict existsother than that occurring by chance.
REFERENCES
Arantes, J., & Grace, R. G. (2008). Failure to obtain valueenhancement by within-trial contrast in simultaneousand successive discriminations. Learning & Behavior,36, 1–11.
Azrin, N. H. (1961). Time-out from positive reinforce-ment. Science, 133, 382–383.
Bautista, L. M., Tinbergen, J., & Kacelnik, A. (2001). Towalk or to fly? How birds choose among foragingmodes. Proceedings of the National Academy of Sciences,98, 1089–1094.
Clement, T. S., Feltus, J. R., Kaiser, D. H., & Zentall, T. R.(2000). ‘‘Work ethic’’ in pigeons: Reward value isdirectly related to the effort or time required toobtain the reward. Psychonomic Bulletin & Review, 7,100–106.
EXTENSIVE TRAINING AND THE WORK-ETHIC EFFECT 151

DiGian, K. A., Friedrich, A. M., & Zentall, T. R. (2004).Discriminative stimuli that follow a delay have addedvalue for pigeons. Psychonomic Bulletin & Review, 11,889–895.
Flory, R. K. (1969). Attack behavior in a multiple fixed-ratio schedule of reinforcement. Psychonomic Science,16, 156–157.
Friedrich, A. M., Clement, T. S., & Zentall, T. R. (2005).Discriminative stimuli that follow the absence ofreinforcement are preferred by pigeons over thosethat follow reinforcement. Learning & Behavior, 33,337–342.
Friedrich, A. M., & Zentall, T. R. (2004). Pigeons shift theirpreference toward locations of food that take moreeffort to obtain. Behavioural Processes, 67, 405–415.
Kacelnik, A., & Marsh, B. (2002). Cost can increasepreference in starlings. Animal Behaviour, 63, 245–250.
Klein, E. D., & Zentall, T. R. (2002). [Failure to replicatethe ‘‘work ethic’’ effect in pigeons.] Unpublished rawdata.
Shapiro, M. S., Siller, S., & Kacelnik, A. (2008). Simulta-neous and sequential choice as a function of rewarddelay and magnitude: Normative, descriptive andprocess-based models tested in the European starling(Sturnus vulgaris). Journal of Experimental Psychology:Animal Behavior Processes, 34, 75–93.
Singer, R. A., Berry, L. M., & Zentall, T. R. (2007).Preference for a stimulus that follows an aversiveevent: Contrast or delay reduction? Journal of theExperimental Analysis of Behavior, 87, 275–285.
Thompson, D. M. (1965). Time-out from fixed-ratioreinforcement: A systematic replication. PsychonomicScience, 2, 109–110.
Vasconcelos, M., Urcuioli, P. J., & Lionello-DeNolf, K. M.(2007). Failure to replicate the ‘‘work ethic’’ effect inpigeons. Journal of the Experimental Analysis of Behavior,87, 383–399.
Vasconcelos, M., & Urcuioli, P. J. (2008). Certainties andmysteries in the within-trial contrast literature: A replyto Zentall (2008). Learning & Behavior, 36, 23–25.
Zentall, T. R. (2008). Within-trial contrast: When you see itand when you don’t. Learning & Behavior, 36, 19–22.
Zentall, T. R., & Singer, R. A. (2007a). Within-trial contrast:When is a failure to replicate not a Type I error? Journalof the Experimental Analysis of Behavior, 87, 401–404.
Zentall, T. R., & Singer, R. A. (2007b). Within-trial contrast:Pigeons prefer conditioned reinforcers that follow arelatively more than a less aversive event. Journal of theExperimental Analysis of Behavior, 88, 131–149.
Received: December 6, 2007Final Acceptance: September 5, 2008
152 MARCO VASCONCELOS and PETER J. URCUIOLI





























