Embed Size (px)
Citation preview

Author QueriesJournal: Proceedings of the Royal Society B
Manuscript: RSPB20142486
As the publishing schedule is strict, please note that this might be the only stage at which you are able to thoroughlyreview your paper.
Please pay special attention to author names, affiliations and contact details, and figures, tables and their captions.
If you or your co-authors have an ORCID ID please supply this with your corrections. More information about ORCIDcan be found at http://orcid.org/.
No changes can be made after publication.
SQ1 Your paper has exceeded the free page extent and will attract page charges.

rspb.royalsocietypublishing.org
ResearchCite this article: Xu X et al. 2015 Extantprimitively segmented spiders have recentlydiversified from an ancient lineage.Proc. R. Soc. B 20142486.http://dx.doi.org/10.1098/rspb.2014.2486
Received: 13 October 2014Accepted: 14 April 2015
Subject Areas:evolution, ecology, taxonomy and systematics
Keywords:living fossils, genetic diversity, plesiomorphies,vicariance, dispersal, ancestral areas
Authors for correspondence:Matjaz Kuntnere-mail: [email protected] Lie-mail: [email protected]
Electronic supplementary material is availableat http://dx.doi.org/10.1098/rspb.2014.2486 orvia http://rspb.royalsocietypublishing.org.
Extant primitively segmented spidershave recently diversified from anancient lineageXin Xu1, Fengxiang Liu1, Ren-Chung Cheng3, Jian Chen1, Xiang Xu5,Zhisheng Zhang6, Hirotsugu Ono7, Dinh Sac Pham8, Y. Norma-Rashid9,Miquel A. Arnedo10, Matjaz Kuntner1,3,4 and Daiqin Li2
1Centre for Behavioural Ecology and Evolution (CBEE), College of Life Sciences, Hubei University, Wuhan,People’s Republic of China2Department of Biological Sciences, National University of Singapore, Singapore, Singapore3Evolutionary Zoology Laboratory, Biological Institute ZRC SAZU, Ljubljana, Slovenia4Department of Entomology, National Museum of Natural History, Smithsonian Institution, Washington, DC, USA5College of Life Sciences, Hunan Normal University, Changsha, Hunan, People’s Republic of China6Key Laboratory of Eco-environments in Three Gorges Reservoir Region (Ministry of Education),School of Life Science, Southwest University, Chongqing, People’s Republic of China7Department of Zoology, National Museum of Nature and Science, 4-1-1 Amakubo, Tsukuba-shi,Ibaraki-ken 305-0005, Japan8Institute of Ecology and Biological Resources (IEBR), Vietnamese Academy of Science and Technology (VAST),18 Hoang Quoc Viet, Cau Giay, Hanoi, Vietnam9Institute of Biological Sciences, Faculty of Science, University of Malaya, Kuala Lumpur 50603, Malaysia10Institut de Recerca de la Biodiversitat, Departament de Biologia Animal, Universitat de Barcelona,Avinguda Diagonal 643, Barcelona 08028, Spain
Living fossils are lineages that retained plesiomorphic traits through long timeperiods. It is expected that such lineages have both originated and diversifiedlong ago. Such expectations have recently been challenged in some textbookexamples of living fossils, notably in extant cycads and coelacanths. Using aphylogenetic approach, we tested the patterns of the origin and diversificationof liphistiid spiders, a clade of spiders considered to be living fossils due totheir retention of arachnid plesiomorphies and their exclusive groupingin Mesothelae, an ancient clade sister to all modern spiders. Facilitated by orig-inal sampling throughout their Asian range, we here provide the phylogeneticframework necessary for reconstructing liphistiid biogeographic history. Allphylogenetic analyses support the monophyly of Liphistiidae and of eightgenera. As the fossil evidence supports a Carboniferous Euramerican originof Mesothelae, our dating analyses postulate a long eastward over-land disper-sal towards the Asian origin of Liphistiidae during the Paleogene (39–58 Ma).Contrary to expectations, diversification within extant liphistiid genera isrelatively recent, in the Neogene and Late Paleogene (4–39 Ma). While noover-water dispersal events are needed to explain their evolutionary history,the history of liphistiid spiders has potential to play prominently in vicariantbiogeographic studies.
1. IntroductionLiving fossils are those lineages that were once more diverse than today, or thathave survived over long evolutionary time scales with little morphologicalchange [1]. Textbook examples include plants such as the Gingko tree and thecycads, and animals such as coelacanth fishes and horseshoe crabs. Theseliving fossils are hypothesized to exhibit ancient roots and early diversificationpatterns [2,3]. However, such assertions are rarely tested within a time-calibratedphylogenetic framework. The latest dating analyses of extant cycads and coela-canths have discovered that although these lineages are ancient, their extantspecies are a surprisingly recent radiation [1,4]. Within arachnids, liphistiid spi-ders are widely recognized as ‘living fossils’ [5] as they are the only surviving
& 2015 The Author(s) Published by the Royal Society. All rights reserved.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
ARTICLE IN PRESS
RSPB20142486—23/4/15—20:27–Copy Edited by: Vinithalakshmi J.R.

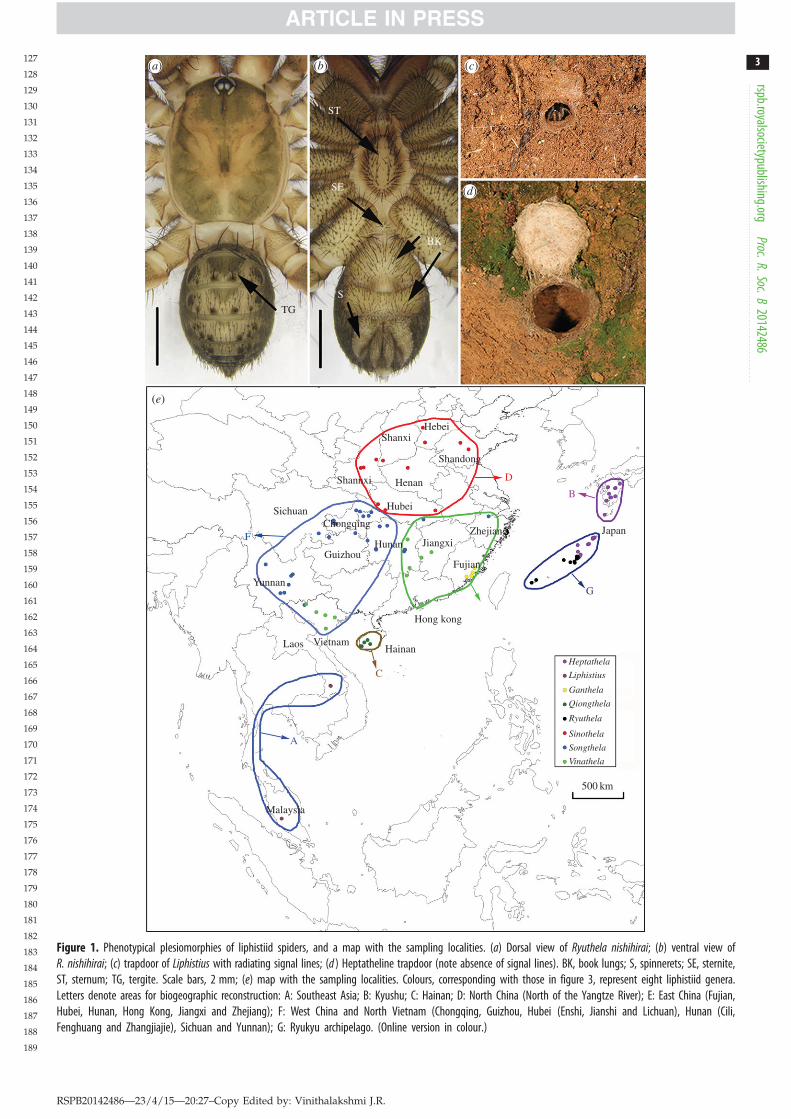
group of the suborder Mesothelae that have retained spiderplesiomorphies such as abdominal tergites and spinneretslocated in the middle of abdominal venter (figure 1a,b) [6–9].Because of this plesiomorphic morphology, the family andgenera of living liphistiid spiders native only to parts ofSoutheast and East Asia are assumed to be ancient and exhi-bit early diversification patterns. This assumption, however,remains untested.
The family Liphistiidae consists of 89 extant species-leveltaxa currently grouped in three genera, and displays an inter-esting geographical distribution confined to Southeast andEast Asia [10]. The group is a species-poor, but seeminglyancient lineage of relatively large, dispersal-limited, unu-sually long-lived (5–18 years), ground-dwelling spiders thatbuild trapdoor burrows (figure 1c,d) [8,9,11]. Since their dis-covery [12], classical authors [13] divide Liphistiidae intotwo subfamilies, Liphistiinae with a single genus, Liphistiusand 50 species, and Heptathelinae with 39 species withintwo genera, Heptathela and Ryuthela [10]. Liphistiines are geo-graphically separated from heptathelines, Liphistius occurringin Southeast Asia, and Heptathela and Ryuthela confined toEast Asia (figure 1e).
As the sister clade to all other known spiders, Mesothelaeshould play prominently in any attempts at reconstructing thearachnid tree of life and testing alternative biogeographichypotheses. Surprisingly, however, the research on Mesothelaeand Liphistiidae has thus far been devoid of comprehensivephylogenies. Although liphistiid exemplars are routinely usedin family level or higher spider phylogenetic research, theseworks either use solely morphological data [7–9,14–17], orwhere molecular data are used, employ a single or at themost two representatives of the family [18–21] except for arecent study reporting the molecular phylogeny for the genusRyuthela [22]. To date, no molecular phylogeny has beenavailable for the family and for other genera.
Representing an ancient lineage with a geographicallyrestricted distribution and very likely limited colonizationability, the biogeography of liphistiids is of great interest. Itis, therefore, surprising how little we know about the originof Mesothelae (the basal-most branch of all spiders) andLiphistiidae (the only Mesothelae family with extant species),and how geological and biological processes in the Earth’shistory might have affected them. The extant Mesothelaeare only distributed in East and Southeast Asia, from whereno Paleozoic arachnids are known, and the only fossil speciesis the Late Carboniferous Palaeothele montceauensis [23],known from the part of Euramerica that corresponds totoday’s France [23,24]. At that time in the Earth’s history(295 Ma), those regions where Mesothelae are found todaywere on landmasses isolated from Pangea [25–32]. Recentauthors [13,33] hypothesized that Mesothelae originated inEuramerica prior to its integration with Pangea, and thatduring Late Triassic the ancestor of Liphistiidae colonizedEast and Southeast Asia from Euramerica over one of severalpossible landmasses. The occurrence of Mesothelae on theislands of southern Japan (Kyushu and Okinawa Islands)was explained both through vicariant origin in the Tertiarywhen the Japanese island arc separated from mainland Asia[33], or alternatively, as a consequence of dispersal eventsover-land bridges from east China during the Pleistocene[9]. Considering the acute absence of a liphistiid phylogeny,none of the above biogeographic hypotheses have beenempirically tested.
Here we use molecular data from our original samplingto test the monophyly of Liphistiidae and its genera. Wethen devise a time-calibrated phylogeny to elucidate theorigin of the stem group Mesothelae and its major clades.We use these data to examine the biogeographic history ofthese spiders, and, given their sedentary lifestyle and longevolutionary history, propose that the lineage is an idealmodel for reconstructing past events. Finally, we test theassumption that present-day liphistiids exhibit not only earlyorigin but also diversification patterns, as would be expectedfrom their labelling as living fossils.
2. Material and methodsThe greater detailed methods are described in the electronic sup-plementary material.
(a) Material and phylogenetic inferenceOur study primarily focused on heptathelines (the liphistiids ofEast Asia), and we added two species of liphistiines from Laosand Malaysia (figure 1e). Our ingroup sampling thus had atotal of 124 liphistiid specimens from 105 localities (figure 1e).We sampled mygalomorph outgroups of the families Atypidaeand Ctenizidae and augmented original data with sequencesfrom GenBank (electronic supplementary material, table S1).We performed parsimony [34], maximum-likelihood (ML) [35]and Bayesian (BI) [36] phylogenetic analyses using nucleotidedata from five genes (CO1, 16S, 28S, H3, ITS2) (electronic sup-plementary material, table S2) and selecting the best fit DNAsubstitution model for each of the four partitions [37].
(b) Calibrations and divergence datingTo estimate divergence times within Liphistiidae, we devised amatrix whose taxon sample maximized fossil calibration points.Analyses conducted using only fossil-based time constraintsyielded extremely low estimates for mtDNA substitution rates,more than twice lower than any value reported in the literature[38–41]. Therefore, we conducted additional analyses withnormal distribution on the mtDNA rate prior based on infor-mation available in the literature. We linked the mitochondrialgenes (CO1 and 16S) in a single clock, assigning the substitutionrate parameter a normal prior with a mean 0.017 and standarddeviation 0.0045, as recently estimated for other spider lineages[42]. For the nuclear genes (28S, H3 and ITS2) and to speed-upanalyses, we restricted the parameter space by setting diffuse,uniform priors to the starting value of 0.00115, with the mini-mum and maximum bounds set to 0.0001 and 0.0115. Thesevalues are based on the universal substitution rate proposedfor arthropod mtDNA [43], and were selected under the assump-tion that the nuclear genes are about 1 order of magnitudeslower than mitochondrial (starting value) and generally nonuclear protein coding gene will show higher rates than themitochondrial genes (upper bound) [44].
Based on the jModeltest results, we used the substitutionmodels GTR ! I ! G for CO1, 16S, 28S and ITS2 and TN93 !I ! G for H3, and base frequencies were set to estimate exceptfor H3 and ITS2 which were set to equal. A starting tree includ-ing the time constraints was generated with starttree (http://bodegaphylo.wikispot.org/starttree_program) and incorporatedto the analysis by modifying the xml file. All analyses wererun assuming a lognormal relaxed clock for each gene partitionand a Yule tree prior. We specified an exponential prior for theyule.birthRate with a mean of 0.05, estimated using the pyuletool (available at https://github.com/joaks1/pyule). For eachanalysis, three independent chains were run for 50 million
rspb.royalsocietypublishing.orgProc.R.Soc.B
20142486
264
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
ARTICLE IN PRESS
RSPB20142486—23/4/15—20:27–Copy Edited by: Vinithalakshmi J.R.
Hebei
ST
(e)
SE
TG
BK
S
Shanxi
Shandong
HenanShannxi
Sichuan
FChongqing
Hunan Jiangxi
Fujian
Zhejiang Japan
B
D
C
A
G
HeptathelaLiphistius
Ganthela
Qiongthela
Ryuthela
Sinothela
Songthela
Vinathela
Guizhou
Hong kong
Yunnan
Laos Vietnam
Malaysia
Hainan
500 km
Hubei
(a) (b) (c)
(d)
Figure 1. Phenotypical plesiomorphies of liphistiid spiders, and a map with the sampling localities. (a) Dorsal view of Ryuthela nishihirai; (b) ventral view ofR. nishihirai; (c) trapdoor of Liphistius with radiating signal lines; (d ) Heptatheline trapdoor (note absence of signal lines). BK, book lungs; S, spinnerets; SE, sternite,ST, sternum; TG, tergite. Scale bars, 2 mm; (e) map with the sampling localities. Colours, corresponding with those in figure 3, represent eight liphistiid genera.Letters denote areas for biogeographic reconstruction: A: Southeast Asia; B: Kyushu; C: Hainan; D: North China (North of the Yangtze River); E: East China (Fujian,Hubei, Hunan, Hong Kong, Jiangxi and Zhejiang); F: West China and North Vietnam (Chongqing, Guizhou, Hubei (Enshi, Jianshi and Lichuan), Hunan (Cili,Fenghuang and Zhangjiajie), Sichuan and Yunnan); G: Ryukyu archipelago. (Online version in colour.)
rspb.royalsocietypublishing.orgProc.R.Soc.B
20142486
3127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
ARTICLE IN PRESS
RSPB20142486—23/4/15—20:27–Copy Edited by: Vinithalakshmi J.R.

generations. Chain convergence and correct mixing of eachMCMC chain was assessed with TRACER v. 1.6 [45]. The 10%of the first generation of each chain was removed as burninand the remaining values were combined into a single file withthe help of LogCombiner [46] and consensus trees were obtainedwith TreeAnnotator [46].
(c) Biogeographical analysesTo infer biogeographical events, we defined seven discretegeographical areas (figure 1e) and reconstructed ancestral distri-butions on a simplified matrix using three alternative methods inRASP v. 3.0 [47,48]: (i) statistical dispersal-vicariance analysis(S-DIVA; [47]), with a maximum of three areas per node; (ii) BIbinary MCMC analysis (BBM; [47,49]), with 5000 cycles using10 chains, sampling every 100 cycles, allowing for a maximumof three areas, giving a ‘custom’ root distribution, using theJC ! G substitution model; and (iii) dispersal-extinction-cladogenesis analysis (DEC; [50]), setting the maximum areasto three [48], with an unconstrained model allowing for equalrates of dispersal (1.0) between areas at any time.
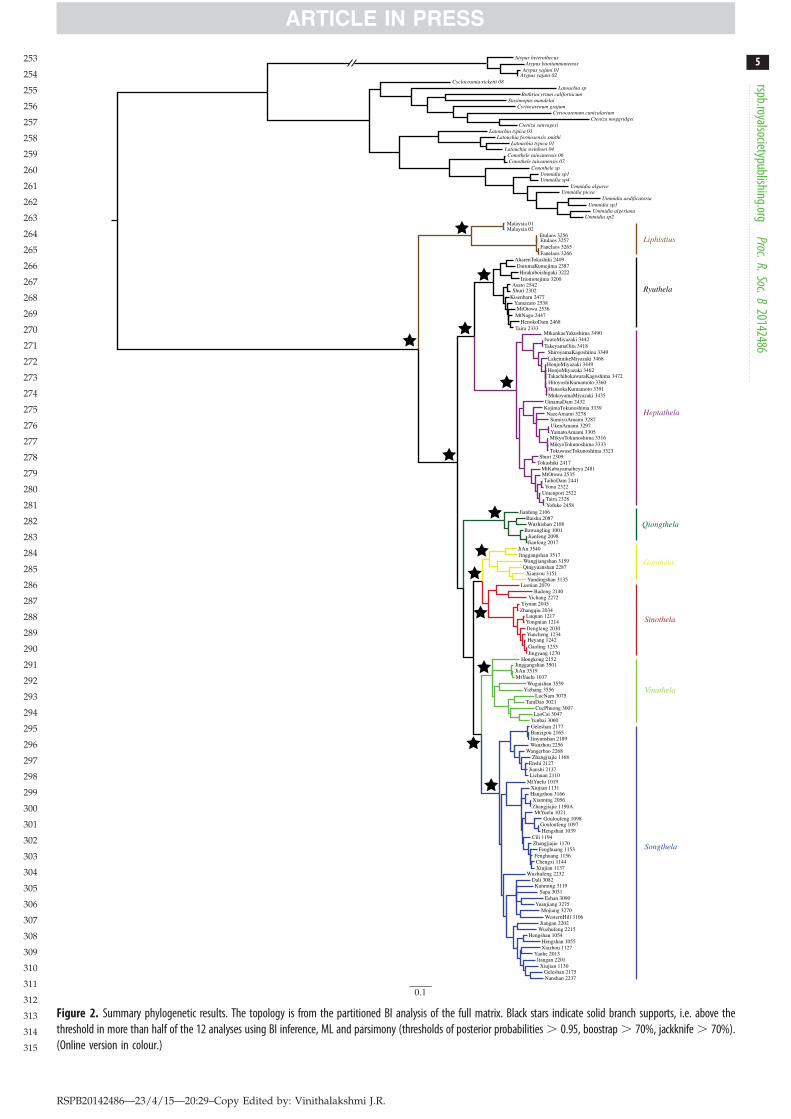
3. Results(a) Phylogenetic reconstructionAll analyses, BI, parsimony and ML under the four partitionschemes consistently recover Liphistiidae monophyly withhigh support values (posterior probability, hereafter PP, MLbootstrap support, BS and parsimony jackknife, JK, all reaching100%), and all analyses converged on similar within-familytopologies (figure 2). The monophyly of Liphistius, in our taxo-nomic sample represented by exemplars from Malaysia andLaos, is well supported (PP ! 1, JK ! 100 and BS ! 100) andthis clade was always sister to heptathelines, a clade unitingall remaining liphistiids (figure 2). The samples from Japanfit into two reciprocally monophyletic and highly supportedclades (figure 2), one including all species of Ryuthela fromRyukyu Island and southern part of Okinawa, and the otherclade consisting of some but not all Heptathela (those fromRyukyu Island and Kyushu; figure 2). The fact that all otherHeptathela species, those from the Asian mainland, fall intoother clades makes Heptathela paraphyletic. In an accompany-ing paper [51], we revise the genus level taxonomy based onthe phylogenetic evidence (this paper) and morphologicaland natural history diagnoses. Here, we refer to those taxo-nomic decisions by treating Liphistius Schiodte, 1849 andRyuthela Haupt, 1983 as valid genera, and Heptathela sensulato as six genera (nomenclature formalized in [51]): Ganthela,Heptathela sensu stricto, Qiongthela, Sinothela Haupt, 2003,Songthela Ono, 2000, Vinathela Ono, 2000.
(b) Divergence dating and biogeographicalreconstructions
Results of the analyses with and without the informedprior in the mtDNA substitution rate resulted in markedlydifferent timeframes of diversification for liphistiids (electronicsupplementary material, table S3 and figure S1; figure 3a).Informing the substitution rate resulted in a compression of theliphistiid clade towards the present, yielding dates about one-third younger than the fossil, unconstrained analysis (electronicsupplementary material, table S3). The estimated rate of substi-tution (ucld.mean) of the mtDNA under the unconstrained
fossil analysis yielded very low values, about twice lower thanthose reported in the literature [38–41] (electronic supplementarymaterial, table S4). In addition, some of the parameter valuesreported for the unconstrained fossil analysis (coefficient of vari-ation of the 28S and ITS2) were higher than those recommendedin the literature (greater than 1) (electronic supplementarymaterial, table S4). On the other hand, increasing the timerange of the maximum bound had little to no effect on the results(electronic supplementary material, tables S3 and S4, columnrates and rates (20%)).
The time-calibrated phylogenetic analyses based on fossil(exponential prior with 95% of the posterior density fallswithin the range 10% older than the minimum bound) andon substitution rates estimated the basal-most spider diver-gence between the Mesothelae (treated as a stem group)and the Opisthothelae (represented by Mygalomorphaeand Araneomorphae) at 297.6 Ma (295–306 Ma) (figure 3a).Given the estimated age of the extant mygalomorphs and ara-neomorphs at 245.3 Ma (240–259 Ma) and 280.8 (262–297 Ma),respectively, and a long, 250 million year Mesothelae branch,the Paleogene origin of the family Liphistiidae estimated at48 Ma (39–58 Ma) is relatively recent. Heptathelinae origin isestimated at 32.9 Ma (28–39 Ma). The origins of all liphistiidgenera fall into the Late Paleogene and Neogene (4–36 Ma).Within Heptathelinae, the clade uniting eastern islandOligocene taxa Heptathela (9.5; 7–12 Ma), Qiongthela (6.4;4–10 Ma) and Ryuthela (7.5; 5–10 Ma) originated at 30.6 Ma(25–36 Ma). The clade of a comparable Oligocene age (27.9;23–33 Ma) unites the mainland genera Ganthela (15.4;11–20 Ma), Sinothela (17.7; 14–22 Ma), Songthela (12.4;10–15 Ma) and Vinathela (16.2; 13–20 Ma). The extant diversityof the Heptathelinae genera can be traced to the Miocene(approx. 4–22 Ma), while Liphistius may have diversifiedslightly earlier (17.3; 11–24 Ma).
Ancestral area reconstructions indicate a number of likelydispersal and vicariant events (electronic supplementarymaterial, figure S2 and table S5). None of postulated dispersalevents in the liphistiid biogeographic history, however, needto invoke any over-water dispersal. The ancestral area for liphis-tiids is as likely to be Southeast as East Asia, which since thatgeological time (approx. 48 Ma) continued to be a single landmass. The heptatheline ancestors occupied the areas correspon-ding to today’s Hainan, East China or Ryukyu archipelago, atthe time (approx. 33 Ma) all part of continental Asia. Likewise,the ancestors of today’s island taxa (Heptathela s.s., Ryuthela,Qiongthela) occupied either Hainan or Ryukyu archipelagobefore they were islands (approx. 31 Ma). These three generadiversified vicariantly, before or when their ancestral areasbecame continental islands, i.e. when Kyushu separated fromthe Asian continent with the opening of the Japanese seaapproximately 15 Ma [53–55], and when Ryukyu archipelagowas gradually isolated from Asia approximately 10–5.3 Ma[56–61]. On the mainland, the separation of the lineagesSinothela and Ganthela (approx. 20; 16–25 Ma) correspondswell to the timing of the Yangtze River origin approximately23 Ma [55,62]. This suggests that their vicariant origin on eachside of the river is still mirrored in today’s distributions.Similarly, Vinathela and Songthela have never in their 23 Ma(19–28 Ma) history left the areas E and F (figure 1e) over Yangtzeto the north or over the ocean to the south and east, and neitherhas Ganthela in approximately 15 Ma (15–20 Ma). Similarly, theapproximately6 Ma (3–10 Ma) evolution of Qiongthela invokesno over-water dispersals from Hainan.
rspb.royalsocietypublishing.orgProc.R.Soc.B
20142486
4190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
ARTICLE IN PRESS
RSPB20142486—23/4/15—20:29–Copy Edited by: Vinithalakshmi J.R.
Malaysia 01Malaysia 02
Etulaos 3256Etulaos 3257Fanelaos 3265Fanelaos 3266
AharenTokashiki 2409DarumaKumejima 2387 Hirakuboishigaki 3222 Iriomotejima 3200
Asato 2542Shuri 2302
MtOtowa 2536MtNago 2447 HenokoDam 2468Taira 2333
Yamazato 2538Kisenbaru 2477
MtkankaeYakushima 3490IwatoMiyazaki 3442TakeyamaOita 3418
ShiroyamaKagoshima 3349LakemiikeMiyazaki 3468HonjoMiyazaki 3449
GinamaDam 2432KojimaTokunoshima 3339
HonjoMiyazaki 3462TakachihokawaraKagoshima 3472HitoyoshiKumamoto 3360HanaokaKumamoto 3391MukoyamaMiyazaki 3435
NazeAmami 3278SumiyoAmami 3287UkenAmami 3297YamatoAmami 3305MikyoTokunoshima 3316MikyoTokunoshima 3333TokuwaseTokunoshima 3323
Shuri 2309Tokashiki 2417
MtKubayamaiheya 2481MtOtowa 2535TaihoDam 2441Yona 2322
Untenport 2522Taira 2328Yofuke 2458
Jianfeng 2106Baisha 2087Wuzhishan 2108
Bawangling 1001Jianfeng 2098Jianfeng 2017
JiAn 3540Jinggangshan 3517
Wangjiangshan 3159Qingyuanshan 2287
Xianyou 3151Yundingshan 3135
Luotian 2079Badong 2140
Yichang 2272Yiyuan 2045Zhangqiu 2034
Luquan 1217Yongnian 1214Dengfeng 2030Yuncheng 1234Heyang 1242Gaoling 1255Jingyang 1270
Hongkong 2152Jinggangshan 3501JiAn 3519MtYuelu 1037
Wugaishan 3559Yizhang 3556
LucNam 3075TamDao 3021
CucPhuong 3007LaoCai 3047
Yenbai 3060Geleshan 2177Banzigou 2165Jinyunshan 2189Wanzhou 2256
Wangerbao 2268Zhangjiajie 1168
Enshi 2127Jianshi 2137Lichuan 2110
MtYuelu 1019Xiujian 1131Hangzhou 3166Xianning 2056Zhangjiajie 1190AMtYuelu 1021
Gouloufeng 1098Gouloufeng 1097Hengshan 1039
Cili 1194Zhangjiajie 1170
Fenghuang 1153Fenghuang 1156Chengxi 1144Xiujian 1137
Wuzhufeng 2232Dali 3082
Kunming 3119Sapa 3031
Eshan 3090Yuanjiang 3275
Mojiang 3270WesternHill 3106
Jiangan 2202Wuzhufeng 2215
Hengshan 1054Hengshan 1055Xiazhou 1127
Yanhe 2013Jiangan 2201
Xiujian 1130Geleshan 2175Nanshan 2237
Atypus heterothecusAtypus baotianmanensis
Atypus yajuni 01Atypus yajuni 02
Cyclocosmia ricketti 08Latouchia sp
Bothriocyrtum californicumStasimopus mandelai
Cyrtocarenum grajumCyrtocarenum cunicularium
Cteniza moggridgeiCteniza sauvagesi
Latouchia typica 03Latouchia formosensis smithi
Latouchia typica 01Latouchia swinhoei 04
Conothele taiwanensis 06Conothele taiwanensis 07
Conothele spUmmidia sp1Ummidia sp4
Ummidia algarveUmmidia picea
Ummidia aedificatoriaUmmidia sp3
Ummidia algerianaUmmidia sp2
Liphistius
Ryuthela
Heptathela
Qiongthela
Ganthela
Vinathela
Songthela
Sinothela
0.1
Figure 2. Summary phylogenetic results. The topology is from the partitioned BI analysis of the full matrix. Black stars indicate solid branch supports, i.e. above thethreshold in more than half of the 12 analyses using BI inference, ML and parsimony (thresholds of posterior probabilities . 0.95, boostrap . 70%, jackknife . 70%).(Online version in colour.)
rspb.royalsocietypublishing.orgProc.R.Soc.B
20142486
5253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
ARTICLE IN PRESS
RSPB20142486—23/4/15—20:29–Copy Edited by: Vinithalakshmi J.R.

4. DiscussionBristowe [5] referred to Liphistiidae as a ‘family of livingfossil spiders’ [5] because they ‘retained many charactersfrom their ancestors and resemble fossil spiders of theCarboniferous period’. Our extensive fieldwork over their pre-sent range (figure 1e) discovered numerous new localitiesand species and, albeit emphasizing heptathelines, secured ataxon sample needed for the first comprehensive species-level phylogeny of Liphistiidae. Our phylogenetic (figures 2and 3a) and biogeographic (electronic supplementarymaterial, figure S2) analyses allowed to rigorously test themonophyly of the family, its two subfamilies, its genera andspecies groups and the timing of the major divergencesand the ancestral ranges. Interesting strong evolutionary andbiogeographic patterns emerge. While extant liphistiidsindeed possess arachnid plesiomorphies (figure 1a,b) notshared with any other spiders, and the Mesothelae lineagetraces back as far as the Carboniferous, the origin of Liphistii-dae and its diversification are surprisingly much more recentthan once thought, repeating the patterns recently detectedalso in other textbook living fossils [1,4]. We trace the originof the family in Southeast or East Asia to the Late Paleogeneand Eocene (39–58 Ma), and the origins of all eight generato the Late Oligocene and Neogene (figure 3a; electronic sup-plementary material, figure S1 and table S3). However,liphistiids do belong to a very long evolutionary branch thatconnects them with the only mesothelean fossil known fromEurasia (figure 3a), and consequently, some of their
phenotypic traits resemble the hypothetically ancestral spi-ders. In the current absence of any additional fossil data, thislong phylogenetic branch suggests that mesothelean evolutionmust account for a very slow eastward move with possiblynumerous extinction events. Below, we discuss three possiblescenarios and suggest that eventual finding of new mesothe-lean fossils may support one over the other, depending onits location. Finally, we align liphistiid natural history, in par-ticular their sedentary lifestyle (figure 1c,d), to the emergingphylogenetic and biogeographic patterns to reveal that theyapparently lack over-water dispersal ability, and this makesthem ideal biogeographic models.
(a) Phylogeny and dating analysisWe provided the first phylogenetic hypothesis for the familyLiphistiidae, with a particular focus on the genera and speciesgroups within Heptathelinae. We used exemplars obtainedthrough our several years of sampling throughout theirrange, obtained data on five mitochondrial and nuclear mar-kers, and analysed them along with numerous mygalomorphoutgroups using three phylogenetic approaches (BI inference,ML and parsimony) under four partition schemes. The results(figures 2 and 3a) have converged on the following generalpatterns: (i) the monophyly of the family Liphistiidae is wellsupported; (ii) the genus Liphistius is the sister group toHeptathelinae, the latter containing all other liphistiids;(iii) within Heptathelinae, Ryuthela is monophyletic but thegenus ‘Heptathela’ s.l. as currently known is paraphyletic.
350 300 250 200 150 100 50 0
I II III IV V
VIII
IXVII
VI
Paleozoic Mesozoic Cenozoic
Carboniferous Permian Triassic Jurassic CretaceousPaleogene NeoP Eoc O Mi P P
H
Liphistius
Qiongthela
Ryuthela
Heptathela
Ganthela
Sinothela
Vinathela
Songthela
Mygalomorphs
Araneomorphs ‘Silk road’
‘stepping-on middle East’
Euramerica
Sibumasu
HebeiShanxi
Shannxi
Chongqing
Guizhou
Yunnan
Vietnam
Sichuan
North China
South China
Sibumasu Indochina
WB
Henan
Shandong
Hubei
Hunan LiangxiZhejiang
Fujian
Hong Kong
Hainan
Heptathela
Liphistius
Ganthela gen. nov.
Qiongthela gen. nov.
Ryuthela
Sinothela
Songthela
Vinathela
Cambodia
Borneo
Sumatra500 km
QiangtangLhasa
Thailand
Malaysia
Laos
Myanmar
TaiwanRyukyu Islands
Kyushu
North China
2
4
5
6
7
8
3 11
9
10
12
13
14
15
1
297.6
48
32.9
17.3
30.6
9.5
23.4
7.5
6.4
27.9
12.4
16.2
15.4
17.7
20.2
22.9
291280.9
245.3
**
*
**
**
**
**
* **
**
**
**
* *
**
**
**
**PP > 95%* PP > 50%
(a) (b)
Figure 3. Divergence time estimates and geographical distribution of liphistiid genera. (a) Chronogram from BEAST with inferred node ages in million years and95% confidence intervals (bars), estimated using fossil calibrations and informed priors on the mtDNA substitution rates. The arrow indicates the fossil age ofPalaeothele montceauensis but does not imply direct ancestry. Black numbered circles represent the major nodes matching those in electronic supplementarymaterial, figure S2. Triangles with Roman numbers mark relevant geological events that may have affected the dispersal routes and diversification of mesothelidand liphistiid lineages. I, Pangea started to form; II, Pangea started to break-up and Sibumasu terrane contacted with Eurasia; III, Sibumasu terrane accreted to theIndochina, South China and North China plates; IV, North China plate connected to Eurasia; V, India collided with Eurasia; VI, the Yangtze river formed; VII, Japaneseislands separated from East Asia; VIII, Taiwan formed; IX, Hainan Island separated from the mainland. (b) General distribution maps of the eight genera delimited inthis study, and the demarcation of main continental plates and terranes that formed Southeast and East Asia (in part from [52]). Inset map shows the two mostplausible of the three mutually exclusive hypotheses explaining the eastward dispersal routes of liphistiid ancestors (simplified on today’s land masses, but see textfor details). WB, West Burma. (Online version in colour.)
rspb.royalsocietypublishing.orgProc.R.Soc.B
20142486
6316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
ARTICLE IN PRESS
RSPB20142486—23/4/15—20:29–Copy Edited by: Vinithalakshmi J.R.

Liphistiid monophyly has never been questioned, and theclade has been recovered in all morphological and molecularanalyses that focused on higher level phylogenetics [7–9,14,15,17–20,63]. However, these studies only included asmall subset of liphistiid diversity, typically one Liphistiusand/or one Heptathela [7,8,18–20]. Our study is the first totest and confirm liphistiid monophyly with a comprehensivetaxon sampling and using molecular data.
Two distinct clades can be identified within Liphistiidae:Liphistiinae and Heptathelinae. Liphistiinae contains onlyLiphistius whose monophyly has not been disputed [9,13].Our dating analysis estimates the crown group Liphistius tobe approximately 17 Ma, but its stem may be as old asapproximately 48 Ma. Its sister lineage, Heptathelinae thatcontains all other liphistiids from China, Japan and Vietnam,originated during the Eocene/Oligocene. Our results unequi-vocally confirm the monophyly of Ryuthela in agreementwith morphological [9,13] and preliminary molecular data-sets [22]. However, all phylogenetic evidence refutes themonophyly of ‘Heptathela’ s.l., contradicting most recenttaxonomic literature [13,64].
The monophyly of each of the mainland Chinese generaSongthela, Vinathela, Sinothela, and in part, Ganthela is wellsupported in the phylogeny, and also by the dating analysisand morphological diagnostics [51]. The exception is the subop-timal clade support for Ganthela in the parsimony analyses, andpoorly resolved relationships within Songthela in most analyses.The apparent conflict in the data may be due to inadequatetaxonomic sampling in south and southwest China. The positionof the genus Qiongthela from Hainan remains unresolved.With relatively low support, the phylogenetic analyses place itas sister to all mainland taxa (figure 2), but the dating analysisplaces it as sister to the doublet Ryuthela and Heptathela s. str.,the remaining two island genera (figure 3a; electronic sup-plementary material, table S5). More sampling and data areneeded to resolve this topological instability for the threegenera that may be crucial for the understanding of liphistiidisland biogeographic patterns.
(b) Historical biogeographyMesothelid spiders are sedentary, dispersal-limited spiderswhose evolutionary history dates back to the very origin ofspiders in the Late Devonian. They have therefore alreadyattracted considerable biogeographical interest [9,33,64–66],but our view is that they should play a more prominent rolein future reconstructions of biogeographical histories. As allprior hypotheses have been devoid of molecular data, thehere presented phylogenetic framework and the inferredchronogram (figures 2 and 3a) shed new light on the originand biogeography of the group. Interestingly, the recon-structed ancestral areas (electronic supplementary material,figure S2) and dispersal versus vicariant events (electronic sup-plementary material, table S5) do not require any explanationsof over-water dispersal in liphistiid biogeographic history.Apparently, effective water barriers include not only bodiesof ocean, but also major rivers, notably Yangtze.
The available fossil evidence supports the ‘Euramericanorigin hypothesis’ for Mesothelae (figure 3b inset; [13,33]).Our various dating analyses agree that the spider origin, andthus the basal-most split between Mesothelae and Opisthothelewas in the Late Carboniferous and Early Permian, approxi-mately 295–320 Ma (figure 3a; electronic supplementary
material figure S1 and table S3). During the Late Carboniferousand Permian, the Laurentia and the Baltica plates togetherformed the equatorially situated Euramerica [28,28,31,67].It was there that the only known Mesothelae fossil livedaround 295 Ma [23,24]. By contrast, the regions inhabitedtoday by extant liphistiids (Southeast and East Asia) corre-spond to isolated microplates of that time, and are notknown to contain any Palaeozoic arachnid fossils. Therefore,all evidence points to the origin of Mesothelae in Euramerica.
The long evolutionary history from the Mesothelae originin Euramerica to modern time is estimated at about 298 Ma.Prior to our dating analysis, the node Liphistiidae was tradition-ally viewed as ‘primitive’ and ancient, but in fact our datinganalysis places its origin at ‘only’ approximately 48 Ma, andtherefore, recovers an extraordinarily long (approx. 250 Ma)branch between the time of the mesothelean fossil (295 Ma)and the origin of Liphistiidae. Even the alternative analysisthat only used fossil data unconstrained for mtDNA substi-tution rates uncovered a relatively recent origin of Liphistiidaeat approximately 124 Ma, implying a minimum estimate ofthe mesothelean branch of 177 Ma. This means that liphistiidancestors must have arrived east over Laurasia (or Gondwana,see below) over a long geological time.
The available paleogeographical and palaeoclimatologicaldata [28,29,31,52,67–71] allow for three possibilities of such along journey over land from Laurasia to Asia (figure 3b; seeelectronic supplementary material): (i) the liphistiid ancestorsmay have taken a southern route along the northern marginsof Gondwana, to which Euramerica connected during theLate Carboniferous [33], then drifted north towards SoutheastEurasia on Sibumasu, i.e. eastern Burma, western Thailand,northwestern peninsular Malaysia and a part of Sumatra[68–70,72–74]. We name this scenario the ‘Out of Gondwana’hypothesis; (ii) they may have taken a route along thesouthern margin of Laurasia by stepping onto the Cimmeriancontinent strip [33]. This is the ‘Stepping-on Middle East’hypothesis (figure 3b, inset); and finally (iii) they may havetravelled through northern Laurasia and on to the NorthChina plate after it had connected to Laurasia during Mid-Jurassic. We name this the ‘Silk road’ hypothesis (figure 3b,inset). Wherever the over-land dispersal took the liphistiidancestors during this long evolutionary time, they (alongwith any branches they may have formed) must have repeat-edly gone extinct along the way. Finding new fossilMesothelae representatives from the Late Carboniferous toEocene is therefore expected, but depending on the routetaken, these fossils could be on either of the continentalmasses of Gondwana, Central and North Asia, Sibumasu orChina. See electronic supplementary material for discussionon plausibility of these hypotheses and their predictions.
Liphistiidae diverged in the Paleogene and diversifiedduring the Neogene (figure 3a; electronic supplementarymaterial figure S1 and table S3). The estimated diversificationtimes and ancestral area reconstructions (figure 3a; electronicsupplementary material figures S1 and S2, tables S3 and S5)suggest that the origins of Heptathela s. str., Qiongthela, andRyuthela, the three genera exclusively inhabiting the southwes-tern Japanese islands and Hainan Island, predate the break-upof the islands form the mainland estimated at 15 Ma for Japanand approximately 2–2.5 Ma for Hainan [53,55,56,75–77]. Ourresults thus support Schwendinger’s hypothesis [33], andrefute the explanation of liphistiids dispersing onto Japaneseislands over-land bridges during the Pleistocene [9]. Likewise,
rspb.royalsocietypublishing.orgProc.R.Soc.B
20142486
7379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
ARTICLE IN PRESS
RSPB20142486—23/4/15—20:29–Copy Edited by: Vinithalakshmi J.R.

the occurrence of Qiongthela on Hainan Island requires no adhoc over-water dispersal events.
We can now also explain the absence of liphistiids fromTaiwan. The island namely formed de novo about 9 Maduring the collision of the Philippine Sea plate with theEurasian plate [78]. In accordance with their sedentary terres-trial life history and an over 298 Ma long evolutionary historyof slow, over-land dispersal, the best explanation at hand isthat liphistiids have never set their tarsi on Taiwan, noteven during relatively short lived land bridges that formedbetween the southern Japanese islands and the Asianmainland during the Pleistocene glacial cycles [79–82].
5. ConclusionOur results combined with the fossil record indicate thatMesothelae originated in Euramerica in the Carboniferousand Permian boundary, and suggest that the period that fol-lowed was marked by a long eastward over-land dispersal ofliphistiid ancestors towards the origin of Liphistiidae inSoutheast or East Asia. The notable pattern seen is an up to250-Ma-long lag between the stem of Mesothelae and theorigin of extant Liphistiidae, combined with the occurrenceof likely extinction events during the evolutionary historyof the family. Since their origin in the Paleogene Asia is rela-tively recent, we argue that liphistiids are modern spidersthat happened to have retained certain characters consideredplesiomorphic within spiders. As such, their labelling asliving fossils is accurate from the phenotypical stance, how-ever, their diversification is much more recent than expected.
Organismal dispersal abilities profoundly affect theirdiversification patterns [83] and biogeographic work benefitsfrom identifying such lineages that due to their ancient originsand limited dispersal abilities retain their genetic imprint andbiogeographic signal [84]. The family Liphistiidae should joinother prominent litter or soil dwelling spiders [40,85–88] and
be considered an excellent model for biogeographic researchon a par with selected few phylogenetically old terrestrialarthropod lineages, such as harvestmen [89], scorpions [90],centipedes [91] and velvet worms [84]. Both the reconstructedbiogeographic history of Liphistiidae and their natural historynamely suggest that these spiders are very poor and slowdispersers that, rather than ever move over-water or overshort-lived land bridges, prefer to maintain their restrictedranges, or ride vicariantly on drifting continents.
Data accessibility. DNA sequences can be accessed via GenBank(KP229800-KP230400); phylogenetic matrices can be accessed viaDryad: http://dx.doi.org/10.5061/dryad.b8d6m; and all other data-sets supporting this study are made available as part of electronicsupplementary material.Acknowledgements. We thank numerous people for local assistance, helpand advice: Gary Ades, Paul Crow, Yorkie Wong and Zoie Wong inHong Kong, Xianjin Peng, Luyu Wang, Bo Wu, Chengqiong Wu,Tingbang Yang and Zizhong Yang in China, Chu Thi Thao andNeuyen Thi Dinh in Vietnam, Zoltan Korsos, Mamoru Toda and BoWu in Japan. We thank Ingi Agnarsson, Danwei Huang, Wei ShongHwang, Nina Vidergar and Matjaz Gregoric for kind help and/oradvice on molecular resources and data analyses, Peter Jager for pro-viding information on collection locality in Laos, Gonzalo Giribet forcommenting on an early draft, and Reta Mehta as well as two anon-ymous reviewers for their insightful comments. Finally, we thank thestaff of the Centre for Behavioural Ecology and Evolution (HubeiUniversity) and the Behavioural Ecology and Sociobiology Lab(National University of Singapore) for support, in particular, ZhanqiChen, Seok Ping Goh, Xiaoguo Jiao, Hongze Li, Jie Liu, Yu Peng,Xiaoyan Wang, Chen Xu, Long Yu and Zengtao Zhang.Funding statement. This work was supported in part by the NSFC (grantno. 31272324) and the Singapore Ministry of Education (AcRF Tier 1grant no. R-154-000-591-112) grants to D.L., and by the SlovenianResearch Agency grant nos. P1-10236 and MU-PROM/12-001 to M.K.Authors’ contributions. D.L., M.K. and X.X. designed this study; D.L.,M.K., F.X.L., H.O., D.S.P., Y.N-R., X.X., X.X. and Z.S.Z. conductedfieldwork; and X.X., R.-C.C., M.A.A., D.L. and M.K. performeddata analyses. All the authors contributed to the writing, revisingof the manuscript and gave final approval for publication.
References
1. Nagalingum NS, Marshall CR, Quental TB, Rai HS,Little DP, Mathews S. 2011 Recent synchronousradiation of a living fossil. Science 334, 796 – 799.(doi:10.1126/science.1209926)
2. Norstog K, Nicholls TJ. 1997 The biology of thecycads. Ithaca, NY: Comstock Pub. Associates.
3. Watson J, Cusack HA. 2005 Cycadales of theEnglish Wealden. Palaeontogr. Soc. Monogr. 622,1 – 189.
4. Inoue JG, Miya M, Venkatesh B, Nishida M. 2005The mitochondrial genome of Indonesian coelacanthLatimeria menadoensis (Sarcopterygii:Coelacanthiformes) and divergence time estimationbetween the two coelacanths. Gene 349, 227 – 235.(doi:10.1016/j.gene.2005.01.008)
5. Bristowe WS. 1975 A family of living fossil spiders.Endeavour 34, 115 – 117. (doi:10.1016/0160-9327(75)90130-1)
6. Pocock RI. 1892 XXXVIII. - Liphistius and its bearingupon the classification of spiders. Ann. Mag. NatHist. Ser. 6 10, 306 – 314. (doi:10.1080/00222939208677416)
7. Platnick NI, Gertsch WJ. 1976 The suborders ofspiders: a cladistic analysis (Arachnida, Araneae).Am. Mus. Nov. 2607, 1 – 15.
8. Coddington JA, Levi HW. 1991 Systematics andevolution of spiders (Araneae). Annu. Rev. Ecol. Syst.22, 565 – 592. (doi:10.1146/annurev.es.22.110191.003025)
9. Haupt J. 2003 The Mesothelaea—monograph of anexceptional group of spiders (Araneae: Mesothelae)(Morphology, behaviour, ecology, taxonomy,distribution and phylogeny). Zoologica 154, 1 – 102.
10. World Spider Catalog. 2015 World spider catalog. version16. Bern, Switzerland: Natural History Museum. Seehttp://wsc.nmbe.ch (accessed 13 April 2015).
11. Bristowe WS. 1976 A contribution to the knowledgeof liphistiid spiders. J. Zool. Lond. 178, 1 – 6.(doi:10.1111/j.1469-7998.1976.tb02260.x)
12. Schiodte JC. 1849 Om en afigende sloegt afspindlernes orden. Naturhistorisk Tidsskr. 2,617 – 624.
13. Schwendinger PJ, Ono H. 2011 On two Heptathelaspecies from southern Vietnam, with a discussion
of copulatory organs and systematics of theLiphistiidae (Araneae: Mesothelae). Rev. Suisse Zool.118, 599 – 637.
14. Haupt J. 1983 Vergleichende Morphologie derGenitalorgane und Phylogenie der liphistiomorphenWebspinnen (Araneae: Mesothelae). I. Revision derbisher bekannten Arten. Z. Zool. Syst. Evol. Forsch.21, 275 – 293. (doi:10.1111/j.1439-0469.1983.tb00296.x)
15. Haupt J. 1984 Comportement sexuel, morphologiegenitale et phylogenese des araigneesliphistiomorphes. Rev. Arachnol. 5, 161 – 168.
16. Platnick N, Goloboff PA. 1985 On the monophylyof the spider suborder Mesothelae (Arachnida:Araneae). New York Entomol. Soc. 93, 1265 – 1270.
17. Kraus O, Kraus M. 1993 Divergent transformation ofchelicerae and original arrangement of eyes inspiders (Arachnida, Araneae). Mem. Queensl. Mus.33, 579 – 584.
18. Agnarsson I, Coddington JA, Kuntner M. 2013Systematics—progress in the study of spiderdiversity and evolution. In Spider research in the
rspb.royalsocietypublishing.orgProc.R.Soc.B
20142486
8442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
ARTICLE IN PRESS
RSPB20142486—23/4/15—20:29–Copy Edited by: Vinithalakshmi J.R.

21st century (ed. D Penney), pp. 58 – 111.Manchester, UK: Siri Scientific Press.
19. Bond JE, Garrison NL, Hamilton CA, Godwin RL,Hedin M, Agnarsson I. 2014 Phylogenomics resolvesa spider backbone phylogeny and rejects aprevailing paradigm for orb web evolution. Curr.Biol. 24, 1765 – 1771. (doi:10.1016/j.cub.2014.06.034)
20. Fernandez R, Hormiga G, Giribet G. 2014Phylogenomic analysis of spiders revealsnonmonophyly of orb weavers. Curr. Biol. 24,1772 – 1777. (doi:10.1016/j.cub.2014.06.035)
21. Sharma PP, Kaluziak ST, Perez-Porro AR, Gonzalez VL,Hormiga G, Wheeler WC, Giribet G. 2014Phylogenomic interrogation of Arachnida revealssystemic conflicts in phylogenetic signal. Mol. Biol.Evol. 31, 2963 – 2984. (doi:10.1093/molbev/msu235)
22. Tanikawa A. 2013 Phylogeny and genetic variationin the spiders of the genus Ryuthela (Araneae:Liphistiidae). Acta Arachnol. 62, 41 – 49. (doi:10.2476/asjaa.62.41)
23. Selden PA. 1996 Fossil mesothele spiders. Nature379, 498 – 499. (doi:10.1038/379498b0)
24. Selden PA. 1996 First fossil mesothele spider, fromthe Carboniferous of France. Rev. Suisse Zool. 2,585 – 596.
25. McLoughlin S. 2001 The breakup history ofGondwana and its impact on pre-Cenozoic floristicprovincialism. Aust. J. Bot. 49, 271 – 300. (doi:10.1071/BT00023)
26. Scotese CR. 2002 Plate tectonic maps andcontinental drift animations. PALEOMAP Project.See www.scotese.com/earth.htm (accessed11 October 2014).
27. Rogers JJW, Santosh M. 2003 Supercontinents inearth history. Gondwana Res. 6, 375. (doi:10.1016/S1342-937X(05)70993-X)
28. Metcalfe I. 2002 Permian tectonic framework andpalaeogeography of SE Asia. J. Asian Earth Sci. 20,551 – 566. (doi:10.1016/S1367-9120(02)00022-6)
29. Metcalfe I. 2011 Tectonic framework andPhanerozoic evolution of Sundaland. Gondwana Res.19, 3 – 21. (doi:10.1016/j.gr.2010.02.016)
30. Seton M et al. 2012 Global continental andocean basin reconstructions since 200 Ma. EarthSci. Rev. 113, 212 – 270. (doi:10.1016/j.earscirev.2012.03.002)
31. Metcalfe I. 2013 Gondwana dispersion and Asianaccretion: tectonic and palaeogeographic evolutionof eastern Tethys. J. Asian Earth Sci. 66, 1 – 33.(doi:10.1016/j.jseaes.2012.12.020)
32. Torsvik TH, Cocks LRM. 2013 Gondwana from topto base in space and time. Gondwana Res. 24,999 – 1030. (doi:10.1016/j.gr.2013.06.012)
33. Schwendinger PJ. 2009 Liphistius thaleri, a newmesothelid spider species from southern Thailand(Araneae, Liphistiidae). Contrib. Nat. Hist. 12,1253 – 1268.
34. Goloboff PA, Farris JS, Nixon KC. 2008 TNT, a freeprogram for phylogenetic analysis. Cladistics 24,774 – 786. (doi:10.1111/j.1096-0031.2008.00217.x)
35. Zwickl DJ. 2006 Genetic algorithm approaches forthe phylogenetic analysis of large biological
sequence datasets under the maximum likelihoodcriterion. PhD thesis, The University of Texasat Austin.
36. Ronquist F et al. 2012 MrBayes 3.2: efficientBayesian phylogenetic inference and model choiceacross a large model space. Syst. Biol. 61, 539 – 542.(doi:10.1093/sysbio/sys029)
37. Darriba D, Taboada GL, Doallo R, Posada D. 2012jModelTest 2: more models, new heuristics andparallel computing. Nat. Methods 9, 772. (doi:10.1038/nmeth.2109)
38. Bond J, Hedin M, Ramirez M, Opell B. 2001 Deepmolecular divergence in the absence ofmorphological and ecological change in theCalifornian coastal dune endemic trapdoor spiderAptostichus simus. Mol. Evol. 10, 899 – 910. (doi:10.1046/j.1365-294X.2001.01233.x)
39. Macıas-Hernandez NE, Oromı P, Arnedo MA. 2010Integrative taxonomy uncovers hidden speciesdiversity in woodlouse hunter spiders (Araneae,Dysderidae) endemic to the Macaronesianarchipelagos. Syst. Biod. 8, 531 – 553. (doi:10.1080/14772000.2010.535865)
40. Bidegaray-Batista L, Arnedo MA. 2011 Gonewith the plate: the opening of the WesternMediterranean basin drove the diversification ofground-dweller spiders. BMC Evol. Biol. 11, 317.(doi:10.1186/1471-2148-11-317)
41. Planas E, Ribera C. 2014 Uncovering overlookedisland diversity: colonization and diversification ofthe medically important spider genus Loxosceles(Arachnida: Sicariidae) on the Canary Islands.J. Biogeogr. 41, 1255 – 1266. (doi:10.1111/jbi.12321)
42. Bidegaray-Batista L, Ferrandez MA, Arnedo MA. 2014Winter is coming: Miocene and Quaternary climatic shiftsshaped the diversification of Western-MediterraneanHarpactocrates (Araneae, Dysderidae) spiders. Cladistics30, 428 – 446. (doi:10.1111/cla.12054)
43. Brower AVZ. 1994 Rapid morphological radiation andconvergence among races of the butterfly Heliconiuserato inferred from patterns of mitochondrial DNAevolution. Proc. Natl Acad. Sci. USA 91, 6491 – 6495.(doi:10.1073/pnas.91.14.6491)
44. Opatova V, Arnedo MA. 2014 From Gondwana toEurope: inferring the origins of MediterraneanMacrothele spiders (Araneae, Hexathelidae) and thelimits of the family Hexathelidae. Invert. Syst. 28,361 – 374.
45. Drummond AJ, Rambaut A. 2007 BEAST: Bayesianevolutionary analysis by sampling trees. BMC Evol.Biol. 7, 214. (doi:10.1186/1471-2148-7-214)
46. Drummond AJ, Suchard MA, Xie D, Rambaut A.2012 Bayesian phylogenetics with BEAUti and theBEAST 1.7. Mol. Biol. Evol. 29, 1969 – 1973. (doi:10.1093/molbev/mss075)
47. Yu Y, Harris AJ, He XJ. 2010 S-DIVA (StatisticalDispersal-Vicariance Analysis): a tool for inferringbiogeographic histories. Mol. Phylogenet. Evol. 56,848 – 850. (doi:10.1016/j.ympev.2010.04.011)
48. Yu Y, Harris AJ, He XJ. 2014 RASP (reconstructancestral state in phylogenies) 3.0. See http://mnh.scu.edu.cn/soft/blog/RASP.
49. Nylander JAA, Olsson U, Alstrom P, Sanmartın I.2008 Accounting for phylogenetic uncertainty inbiogeography: a Bayesian approach to dispersal –vicariance analysis of the thrushes (Aves: Turdus).Syst. Biol. 57, 257 – 268. (doi:10.1080/10635150802044003)
50. Ree RH, Smith SA. 2008 Maximum likelihoodinference of geographic range evolution by dispersallocal extinction and cladogenesis. Syst. Biol. 57,4 – 14. (doi:10.1080/10635150701883881)
51. Xu X, Liu FX, Chen J, Ono H, Li D, Kuntner M. 2015 Agenus level taxonomic revision of primitivelysegmented spiders (Mesothelae: Liphistiidae). Zookeys488, 121 – 151. (doi:10.3897/zookeys.488.8726)
52. Metcalfe I. 1998 Palaeozoic and Mesozoic geologicalevolution of the SE Asian region: multidisciplinaryconstraints and implications for biogeography. InBiogeography and geological evolution of SE Asia(eds R Hall, JD Holloway), pp. 25 – 41. Leiden,The Netherlands: Backhuys Publishers.
53. Otofuji Y, Matsuda T, Nohda S. 1985 Opening modeof the Japan Sea inferred from paleomagnetism ofthe Japan arc. Nature 317, 603 – 604. (doi:10.1038/317603a0)
54. Otofuji Y, Itaya T, Matsuda T. 1991 Rapid rotation ofSouthwest Japan: paleomagnetism and K-Ar ages ofMiocene volcanic rocks of southwest Japan.Geophys. J. Intern. 105, 397 – 405. (doi:10.1111/j.1365-246X.1991.tb06721.x)
55. Yin A. 2010 Cenozoic tectonic evolution of Asia: apreliminary synthesis. Tectonophysics 488,293 – 325. (doi:10.1016/j.tecto.2009.06.002)
56. Kimura M. 2000 Paleogeography of the RyukyuIslands. Tropics 10, 5 – 24. (doi:10.3759/tropics.10.5)
57. Kimura M. 2002 The formation age of the RyukyuArc and migration of biota to the arc. Naha,Okinawa: Okinawa Times Co.
58. Kimura M. 2003 Land connections between Eurasiancontinent and Japanese islands—related to humanmigration. Mig. Diff. 4, 14 – 33.
59. Ota H. 1998 Geographic patterns of endemism andspeciation in amphibians and reptiles of the RyukyuArchipelago, Japan, with special reference to theirpaleogeographical implications. Res. Popul. Ecol. 40,189 – 204. (doi:10.1007/BF02763404)
60. Ota H. 2000 The current geographic faunal patternsof reptiles and amphibians of the RyukyuArchipelago and adjacent regions. Tropics 10,51 – 62. (doi:10.3759/tropics.10.51)
61. Otsuka H, Takahashi A. 2000 Pleistocene vertebratefaunas in the Ryukyu Islands: their migration andextinction. Tropics 10, 25 – 40. (doi:10.3759/tropics.10.25)
62. Zheng HB, Clift PD, Wang P, Tada R, Jia JT, He MY,Jourdan F. 2013 Pre-Miocene birth of the YantzeRiver. Proc. Natl Acad. Sci. USA 110, 7556 – 7561.(doi:10.1073/pnas.1216241110)
63. Bond JE, Hendrixson BE, Hamilton CA, Hedin M.2012 A reconsideration of the classification of thespider infraorder Mygalomorphae (Arachnida:Araneae) based on three nuclear genes andmorphology. PLoS ONE 7, e38753. (doi:10.1371/journal.pone.0038753)
rspb.royalsocietypublishing.orgProc.R.Soc.B
20142486
9505
506
507
508
509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
524
525
526
527
528
529
530
531
532
533
534
535
536
537
538
539
540
541
542
543
544
545
546
547
548
549
550
551
552
553
554
555
556
557
558
559
560
561
562
563
564
565
566
567
ARTICLE IN PRESS
RSPB20142486—23/4/15—20:29–Copy Edited by: Vinithalakshmi J.R.

64. Ono H. 2000 Zoogeographic and taxonomic noteson spiders of the subfamily Heptathelinae (Araneae,Mesothelae, Liphistiidae). Mem. Natl Sci. Mus. 33,145 – 151.
65. Paik K. 1953 A study on the geographicaldistribution of Heptathela kimurai Kishida. ActaArachnol. 13, 53 – 68. (doi:10.2476/asjaa.13.63)
66. Haupt J. 2003 Zoogeography in southern Japan asrevealed by ground-living arachnids. Rev. SuisseZool. 110, 133 – 139.
67. Cox CB, Moore PD. 2005 Biogeography: an ecologicaland evolutionary approach, 7th edn. Malden, MA:Blackwell Publishing.
68. Ali JR, Aitchison JC. 2005 Greater India. EarthSci. Rev. 72, 169 – 188. (doi:10.1016/j.earscirev.2005.07.005)
69. Ali JR, Aitchison JC. 2006 Positioning PaleogeneEurasia problem: solution for 60 – 50 Ma andbroader tectonic implications. Earth Planet. Sci. Lett.251, 148 – 155. (doi:10.1016/j.epsl.2006.09.003)
70. Ali JR, Aitchison JC. 2008 Gondwana to Asia: platetectonics, paleogeography and the biologicalconnectivity of the Indian sub-continent from theMiddle Jurassic through latest Eocene(166 – 35 Ma). Earth Sci. Rev. 88, 145 – 166. (doi:10.1016/j.earscirev.2008.01.007)
71. Torsvik TH, Cocks LRM. 2004 Earth geography from400 to 250 Ma: a palaeomagnetic, faunal and faciesreview. J. Geol. Soc. Lond. 161, 555 – 572. (doi:10.1144/0016-764903-098)
72. Golonka J, Krobicki M, Pajak J, Giang NV, ZuchiewiczW. 2006 Phanerozoic palaeogeography of SoutheastAsia. Geolines 20, 40 – 43.
73. Golonka J. 2007 Phanerozoic paleoenvironment andpaleolithofacies maps. Mesozoic. Geologia 33,211 – 264.
74. Clouse RM, Giribet G. 2010 When Thailand was anisland—the phylogeny and biogeography of mite
harvestmen (Opiliones, Cyphophthalmi, Stylocellidae)in Southeast Asia. J. Biogeogr. 37, 1114 – 1130.(doi:10.1111/j.1365-2699.2010.02274.x)
75. Zeng ZX, Zeng XZ. 1989 Physical geography ofHainan Island. Beijing, China: Science Press.
76. Zhao HT, Wang LR, Yuan JY. 2007 Origin and timeof Qiongzhou Strait. Marine Geol. Quat. Geol. 27,33 – 40.
77. Huang Y, Guo X, Ho SY, Shi H, Li J, Li J, Cai B, WangY. 2013 Diversification and demography of theoriental garden lizard (Calotes versicolor) on HainanIsland and the adjacent mainland. PLoS ONE 8,e64754. (doi:10.1371/journal.pone.0064754)
78. Sibuet JC, Hsu SK. 2004 How was Taiwan created?Tectonophysics 379, 159 – 181. (doi:10.1016/j.tecto.2003.10.022)
79. Wageman JM, Hilde TMC, Emery KO. 1970 Structuralframework of East China-Sea and Yellow-Sea. Am.Assoc. Pet. Geol. Bull. 54, 1611 – 1643.
80. Ye XQ. 1982 On the formation and developmentof the geology and geomorphology of Taiwan.J. Central China Teachers College 1982, 83 – 89.
81. Juan VCC. 1986 Thermal-tectonic evolution of theYellow sea and East-China-Sea—Implication fortransformation of continental to oceanic-crust andmarginal basin formation. Tectonophysics 125,231 – 244. (doi:10.1016/0040-1951(86)90016-8)
82. Teng LS. 1992 Geotectonic evolution of Tertiarycontinental margin basins of Taiwan. Petrol. Geol.Taiwan 27, 1 – 19.
83. Agnarsson I, Cheng RC, Kuntner M. 2014 A multi-clade test supports the intermediate dispersalmodel of biogeography. PLoS ONE 9, e86780.(doi:10.1371/journal.pone.0086780)
84. Murienne J, Daniels SR, Buckley TR, Mayer G, GiribetG. 2014 A living fossil tale of Pangaeanbiogeography. Proc. R. Soc. B 281, 20132648.(doi:10.1098/rspb.2013.2648)
85. Hendrixson B, Bond J. 2007 Molecular phylogenyand biogeography of an ancient Holarctic lineage ofmygalomorph spiders (Araneae: Antrodiaetidae:Antrodiaetus). Mol. Phylogenet. Evol. 42, 738 – 755.(doi:10.1016/j.ympev.2006.09.010)
86. Hedin M, Starrett J, Hayashi C. 2013 Crossing theuncrossable: novel trans-valley biogeographicpatterns revealed in the genetic history of low-dispersal mygalomorph spiders (Antrodiaetidae,Antrodiaetus) from California. Mol. Ecol. 22,508 – 526. (doi:10.1111/mec.12130)
87. Opatova V, Bond JE, Arnedo MA. 2013 Ancientorigins of the Mediterranean trap-door spiders ofthe family Ctenizidae (Araneae, Mygalomorphae).Mol. Phylogenet. Evol. 69, 1135 – 1145. (doi:10.1016/j.ympev.2013.08.002)
88. Wood HM, Matzke NJ, Gillespie RG, Griswold CE.2013 Treating fossils as terminal taxa in divergencetime estimation reveals ancient vicariance patternsin the palpimanoid spiders. Syst. Biol. 62,264 – 284. (doi:10.1093/sysbio/sys092)
89. Giribet G, Sharma PP, Benavides LR, Boyer SL,Clouse RM, de Bivort BL, Kawauchi GY, Murienne J,Schwendinger PJ. 2012 Evolutionary andbiogeographic history of the harvestman suborderCyphophthalmi (Arachnida, Opiliones)—an ancientand global group of arachnids. Biol. J. Linn. Soc.105, 92 – 130. (doi:10.1111/j.1095-8312.2011.01774.x)
90. Bryson Jr RW, Savary WE, Prendini L. 2013Biogeography of scorpions in the Pseudouroctonusminimus complex (Vaejovidae) from south-westernNorth America: Implications of ecologicalspecialization for pre-Quaternary diversification.J. Biogeogr. 40, 1850 – 1860.
91. Giribet G, Edgecombe GD. 2012 Reevaluating theArthropod tree of life. Annu. Rev. Entomol. 57,167 – 186. (doi:10.1146/annurev-ento-120710-100659)
rspb.royalsocietypublishing.orgProc.R.Soc.B
20142486
10568
569
570
571
572
573
574
575
576
577
578
579
580
581
582
583
584
585
586
587
588
589
590
591
592
593
594
595
596
597
598
599
600
601
602
603
604
605
606
607
608
609
610
611
612
613
614
615
616
617
618
619
620
621
622
623
624
625
626
627
628
629
630
ARTICLE IN PRESS
RSPB20142486—23/4/15—20:29–Copy Edited by: Vinithalakshmi J.R.





























