Embed Size (px)
Citation preview

1995;55:1798-1806. Cancer Res Kazue Matsumoto, Randal K. Wada, Joyce M. Yamashiro, et al. Neuroblastoma CellsAffects Survival, Differentiation, and Invasiveness of Human
TrkBExpression of Brain-derived Neurotrophic Factor and p145
Updated version
http://cancerres.aacrjournals.org/content/55/8/1798
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

[CANCER RESEARCH 55, 1798-1806, April 15, 1995]
Expression of Brain-derived Neurotrophic Factor and p!45TrkB Affects Survival,
Differentiation, and Invasiveness of Human Neuroblastoma Cells1
Kazue Matsumoto, Randal K. Wada, Joyce M. Yamashiro, David R. Kaplan, and Carol J. Thiele2
Cell and Molecular Biology Section, Pediatrie Branch, National Cancer Institute, Bethesda, Maryland 20892-1928 [K. M., C. J. T.¡:Department of Pediatrics, UCLA School ofMedicine, and the Gwynne Hazen Cherry Memorial Laboratories, Los Angeles, California 90024 [R. W., J. Y.]; and Eukaryotic Signal Transduclion Group, ABL-Basic Research
Program, National Cancer Institute, Frederick, Maryland 21702 [D. R. K.]
ABSTRACT
A large number of poor prognosis neuroblastoma (NB) tumors consti-tutively express brain-derived neurotrophic factor (BDNF) and variably
express the gene for its tyrosine kinase (Trk) receptor TrkB. Good prognosis NB tumors typically express high levels of TrkA mRNA, whichencodes the signal transducing receptor for nerve growth factor, pl40TrkA.
These neurotrophins are necessary for neural cell survival and differentiation. This study evaluates the effects of activation of the BDNF-TrkB
signal transduction pathway on the growth, survival, morphology, andinvasive capacity of NB cells. We find that the addition of BDNF to SY5Ycells induced to express pl45TrkB by retinole acid treatment does not
significantly affect cell proliferation yet will support cell survival. Activation of the BDNF-TrkB signal transduction pathway stimulates disaggre-
gation of cells and extension of neuritic processes which can be blocked bya BDNF-neutralizing antibody. Treatment of cells with K252a, an inhib
itor of Trk, reverses the cellular disaggregation. An evaluation of theeffects of BDNF and nerve growth factor on the ability of NB cells topenetrate basement membrane proteins indicated that BDNF stimulated a2-fold increase while nerve growth factor inhibited RA-SY5Y cell invasion. Thus, activation of the p 145 ' 'k" signal transduction pathway stim
ulates NB cell survival, disaggregation, and invasion; all characteristics ofmetastatic cells. Furthermore, these studies indicate that activation ofdifferent Trk signal transduction pathways in NB cells results in distinctdifferences in tumor cell biology and these may be relevant to the clinicalcourse of the patients.
INTRODUCTION
Human NB3 tumors are derived from pluripotent embryonal neural
crest cells. The neural crest is a transient embryonal structure thatarises during the closure of the neural tube, at which time cells migrateventrally and laterally to give rise to the sympathetic and sensoryganglia of the peripheral nervous system, the chromaffin cells of theadrenal gland, melanocytes, and a variety of other tissues. Furthermore, neural crest cells have high proliferative and migratory potentialcharacteristics also found in aggressive tumors (1). Several clinicalobservations suggest that NB tumors may arise from genetic alterations affecting processes controlling differentiation: (a) NB in situoccurs at a much higher rate than the frequency of NB in the population (2). (b) NB tumors exhibit a high degree of cellular heterogeneity reminiscent of different stages of neural cell development ranging from immature blasts to ganglion cells (3). (c) Cases ofspontaneous as well as therapy-induced maturation of tumors to
benign ganglioneuromas have been documented (4, 5).The survival, maturation, and function of neural cells is influenced
Received 10/18/94; accepted 2/15/95.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 This research was sponsored in part by the National Cancer Institute, NIH, Department of Health and Human Services, under Contract N01-CO-74101 with ABL (D. R. K.),
an NIH First Award (R. K. W.), and a grant from the Concern II Foundation (R. K. W.).2 To whom requests for reprints should be addressed, at Cell and Molecular Biology
Section, Pediatrie Branch, National Cancer Institute, Building 10/13N240, Bethesda, MD20892-1928.
3 The abbreviations used are: NBf neurobiastoma; NGF, nerve growth factor; BDNF,
brain-derived neurotrophic factor; RA, retinoic acid; NT, neurotrophin; Trk, tyrosinekinase. ' - -
by NTs encountered in the extracellular milieu during development.NGF is the prototypic NT in a family that includes BDNF, NT-3, andNT-4/5 (6-8). These polypeptides are structurally and functionally
related and are expressed in tissues, both in discreet yet often overlapping patterns during development (6-8). The effects of NTs are
mediated by the Trk family of tyrosine kinase receptors (TrkA, TrkB,and TrkC) that encode both full-length (pl30-145 kDa proteins) as
well as truncated variants that lack the intracellular Trk domain(9-12). Signals transduced by NGF are mediated primarily by TrkA,BDNF, NT-3, NT-4/5 by TrkB, and NT-3 predominantly by TrkC
(13).Analysis of NT receptors in NB tumors has shown that good
prognosis NB tumors express high levels of TrkA (14-17), while
many poor prognosis NB tumors and cell lines express BDNF (18,19)and tumors (18), but not typically cell lines, express either full-length
or truncated TrkB (18, 19). High levels of BDNF have been detectedin NB tumors and cell lines containing amplified N-myc genes, a
marker of poor prognosis in patients with NB tumors (20). NB celllines variably express TrkA and pVS1"â„¢(21, 22) and in many of these
cell lines signal transduction via endogenous TrkA is impaired (23).However, NGF signal transduction may be intact since NGF treatmentof a primary culture from an NB tumor stimulated cell survival (14).Furthermore, NGF treatment of NB cells expressing high levels ofTrkA by transfection cause early response gene activation, neuriteextension, and cell growth arrest in a NB cell line (23). Typically NBcell lines have been derived from poor prognosis NB tumors. Whilethese tumors express TrkB and BDNF, few NB cell lines expressreadily detectable levels of TrkB mRNA (18). Recently, we determined that many NB cell lines constitutively express BDNF, whiletheir ability to express high levels of TrkB is regulated by RA (18).The NB cell line, SY5Y (24), does not express high levels of BDNF;however, RA induces TrkB and neurite extension occurs with theaddition of exogenous BDNF (18).
The effects of BDNF-TrkB signal transduction on NB cell growth,
survival, and changes in cell morphology have been evaluated. Sincea number of poor prognosis NB tumors constitutively express BDNFand TrkB mRNA (19), we have used the SY5Y NB cell line becauseit does not constitutively express BDNF, thus enabling the biologicaleffects of activation of the TrkB signal transduction pathway byBDNF to be evaluated. In this study, we have found that BDNFincreases NB cell survival, induces neurite extension, cellulardisaggregation, and stimulates the invasiveness of NB cells.
MATERIALS AND METHODS
Cell Culture. The NB cell line SH-SY5Y (SY5Y) (24) was cultured in
RPMI 1640 (Media Tech, Herndon, VA) supplemented with 10% PCS(Biofluid, Rockville, MD), glutamine (2 HIM), penicillin (50 units/ml), andstreptomycin (50 M-g/ni') at 37°Cwith 5% CO2.
NTs and Antibodies. NFG was obtained from Upstate Biotechnology, Inc.(Lake Placid, NY) or Genentech, Inc. (South San Francisco, CA). BDNF andNT-3 were kindly provided by R. Scott and J. Vaught (Cephalon, Inc., West
Chester, PA) or by J. Winslow, K. Nikolics, and Gene Burton (Genentech,Inc.). NT-4/5 was a gift from Arnon Rosenthal (Genentech, Inc.). The anti-
phosphotyrosine mAb 4G10 was provided by D. Morrison (National Cancer
1798
on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TrkB IN NEUROBLASTOMA
Fig. 1. Effect of RA on cell proliferation andexpression of TrkB mRNA and protein. A, effect ofdifferent concentrations of RA on SY5Y cell proliferation. SY5Y cells (5 X IO3) were plated in96-well plates and treated with solvent control or 1
pM, 1 nM, l P.M,or 5 ftM RA for varying times after24 h of preplating. B, effect of different concentrations of RA on TrkB mRNA induction. Total RNA(25 (ig) was isolated from SY5Y cells treated withcontrol solvent (Lane 0) or with the indicated concentrations of RA for 5 days. Northern blot wasanalyzed for expression of human TrkB. Glutaral-dehyde-3-phosphate dehydrogenase is shown to
control for sample loading. C, effect of differentconcentrations of RA on TrkB protein levels andautophosphorylation of TrkB. Lysate from cellstreated with the indicated concentrations of RA for7 days and prior to cell lysis treated with 100 ng/mlBDNF or control media for 5 min. Western blotwas analyzed using an anti-PTy antibody to determine autophosphorylation of pl45TrkB. Lines, mo
lecular weight standards of 200 kDa (upper) and100 kDa (lower). Densitometric analysis of appropriately exposed autoradiogram using a MicrotechImage Scanner and Image Analysis software (NIH)was used to estimate relative changes in gene orprotein expression.
Ù.O
B.
RA
TrkB
GAPDH •
C.
RA
u PTyr
—28S
Institute-Frederick Cancer Research and Development Center). Anti-TrkBantibody was generated as described (25-27). The anti-TrkB antibody does notrecognize TrkA (25-27). The BDNF-neutralizing antibody was kindly pro
vided by J. Carnahan (AMGEN, Thousand Oaks, CA) and generated asdescribed (28). The NGF-neutralizing antibody was purchased from Bohringer
Mannheim (Indianapolis, IN).Morphological Studies. SY5Y cells were plated in 24-well dishes at a
density of 2.5 X IO4 cells/well in duplicate. After 24 h, cells were treated with
various concentrations of NTs in combination with l UMall trans-RA (RA;
Sigma Chemical Company, St. Louis, MO) or solvent control for 6 days. Forantibody studies, a 1:100 dilution of anti-BDNF, anti-NGF, or preimmune
serum was added in the absence or presence of l UMRA or control solvent for6 days. For experiments using K252a (29, 30), SY5Y cells were plated at adensity of 5 X 103/well and after 24 h were treated with 5 /J.MRA and 10 ng/ml
BDNF. After 5 days, cells were washed and recultured in media containing 5/AMRA and 10 ng/ml BDNF in the presence of indicated concentrations ofK252a for 24 h. The photographs were taken with a Nikon Diaphot invertedmicroscope using Hoffman modulation lens. Cell counts were performed usinga hemocytometer and viability was assessed by trypan blue exclusion.
[3H]dThd Incorporation. SY5Y cells (5 X 103/well) were plated in quin
tuplicate in 96-well plates. After 24 h, cells were treated with l /J.MRA and 10
ng/ml BDNF. After 5 days, cultures were washed and medium was changed toserum free supplemented with 10 ng/ml BDNF or solvent control alone.Twenty h prior to harvest, cells were labeled with 1 /iCi/well [3H]dThd (ICN,Costa Mesa, CA). For RA dose-response experiments, 5 X IO3 cells/well were
plated in 96-well plates, then treated with 1 pM RA, 1 nvt RA, 1 /AMRA, 5 /XM
RA, or solvent control for varying times. For neuotrophin experiments, cellswere treated with 0, 10, and 100 ng/ml each of BDNF, NGF, or NT-3 for 6
days. For RA and BDNF concentration experiments, SY5Y cells were platedas described above and treated with a combination of 0, 10, or 100 ng/mlBDNF with 1 nM RA, 1 /AMRA, or solvent control. After 6 days, cells wereincubated with [3H]dThd for 20 h and harvested using an Inotech harvester
(Inotech, Lansing, MI), and counted in a Beckman LS1801 scintillationcounter (Beckman Instruments Inc., Fullton, CA).
RNA Analysis. RNA isolation and hybridization were performed as described previously (31, 32). Total RNA was extracted from subconflucnt150-cm2 plates of indicated cell lines. Total RNA (25 /ig) from each sample
was electrophoresed on 1% agarose-2.2 Mformaldehyde gels and transferred to
Nytran membranes (Schleicher and Schuell, Keene, NH) by capillary transfer.Blots were hybridized with 32P-labeled DNA. DNA inserts were isolated fromplasmids containing rat TrkB (33, 34), human TrkB,4 human TrkA (35), BDNF
(36), and glyceraldehyde-3-phosphate dehydrogenase. Blots were hybridized,
washed, and exposed to X-ray films as described previously (18). Rehybrid-
ization of membrane was performed after treating Nytran membranes for 1 hin 50% formamide and IX SSC at 75°C.
Protein Analysis. Cells (2 X 10") were plated in 150-cm2 dishes for 24 h
and treated with solvent control, 1 pM RA, 1 nM RA, 1 /¿MRA, or 5 /J.MRAfor 6 days. Cells were fed every 2 days with fresh RA media and were treatedwith 100 ng/ml BDNF for 5 min with rocking at 37°C.To detect TrkB protein,
cells were lysed, and the lysates were immunoprecipitated as described previously (18, 37, 38). The lysates were immunoprecipitated with anti-pan Trk203 (39). Immunoprecipitations (performed for 2-4 h at 4°C)were collected
with protein A-Sepharose and washed three times with NP40 lysis buffer and
once with water. The immunoprecipitates were boiled in sample buffer (2%SDS, 100 mM DTT, 10% glycerol, and 0.25% bromophenol blue) for 5 minand electrophoresed on 7.5% SDS-polyacrylamide gels (PAGE) before transferto nitrocellulose. Protein blots were probed overnight at 4°Cwith antiphos-
photyrosine antibody 4G10, and TrkB antibody was diluted in Tris-buffered
saline (pH 8.0) and 0.2% Tween 20. Blots were analyzed using an ECLchemiluminescence system (Amersham Corp., Arlington Heights, IL). Proteinblots were then stripped of antibody in 62.5 mM Tris (pH 6), 2% SDS, and 100HIM2-mercapotethanol at 70°Cfor 1 h and reprobed.
Invasion Assay. SY5Y cells were pretreated with 1 nM RA for 3 days. Asingle-cell suspension containing 5 X 10" cells in 0.2 ml of serum-freeIscove's modified Dulbecco's medium was placed in the upper compartment
4 Y. Kitajima, K. Matsumoto. M. Morrill, and C. J. Thiele, manuscript in preparation.
1799
on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
TrkB IN NEUROBLASTOMA
of a modified Boyden chamber (Falcon, Oxnard, CA) that had been coatedwith 40 ng Matrigel, an artificial basement membrane preparation (Collaborative Research, Bedford, MA). The bottom chamber held 0.5 ml medium andboth chambers contained 1 nM RA and varying amounts (0, 5, 10, or 50 ng/ml)of either BDNF or NGF. The cells were incubated for 24 h at 37°C.Under
these conditions, cells are unable to penetrate to the lower surface of the filterwithout elaborating proteolytic enzymes. After incubation, noninvasive cellsand Matrigel on the upper surface of the filter were removed using a moistcotton swab. Adherent invasive cells on the under surface of the filter werefixed with 2% paraformaldehyde, stained in 0.1% crystal violet, and countedunder high-power magnification. Nonadherent invasive cells found in thebottom chamber were collected by centrifugation and quantified by hemocy-
tometer. The values reported for each point represent the mean and SE ofquadruplicate experiments as a percentage of the control mean and SE values.
RESULTS
Effect of NTs on NB Cell Growth and Survival. Since a numberof tumors derived from patients with an unfavorable prognosis express TrkB and BDNF, we evaluated the effects of BDNF on NBtumor ceil proliferation and survival. The SY5Y NB cell line consti-
tutively expresses TrkA yet does not express readily detectable TrkBand BDNF by Northern blot and Western blot analyses (18). Using theSY5Y NB cell model system, it is possible to study the effects of theaddition of exogenous BDNF on NB cells that lack pl45TrkB or can beinduced by RA to express pl45TrkB. Although RA is known to inhibit
NB cell growth (32, 41), an analysis of the effects of differentconcentrations of RA on SY5Y cell proliferation indicated that 1 nMRA had no significant effect on [3H]dThd uptake in SY5Y NB cells
while 1 juMRA resulted in a 50% reduction at 6 days. Concentrationsas high as 5 /AMRA are required to completely arrest SY5Y cellgrowth (Fig. 1A). Our previous experiments showed an increase inTrkB protein in NB cells treated with 1 nM RA (9-fold), and Trkprotein increased at 5 JXMRA (28-fold; Ref. 18). Northern blot
analysis indicated that concentrations of RA as low as 1 nM inducedboth the 8.4- and 9.5-kilobase TrkB mRNA species (8.5-fold) withincreasing levels of TrkB mRNA (28-fold) detected at 5 JAMRA(Fig. Iß).In addition, a 5-min BDNF treatment of 1 nM RA-treatedSY5Y cells caused a dramatic increase in p!45TrkB autophosphorylation
(94-fold), a level similar to that detected in 1 /AM(106-fold) or 5 /AMRA(102-fold)-treated cells (Fig. 1C). Autophosphorylation of pl45TrkB is an
early step in the BDNF-TrkB signal transduction pathway.
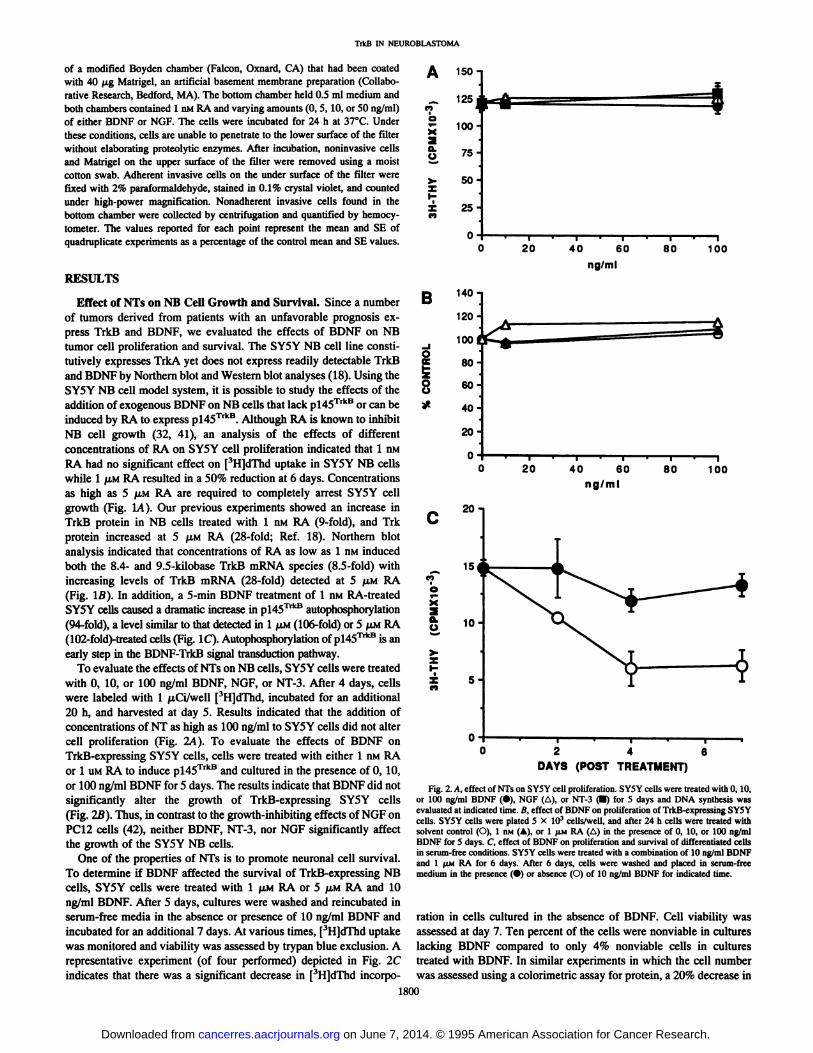
To evaluate the effects of NTs on NB cells, SY5Y cells were treatedwith 0, 10, or 100 ng/ml BDNF, NGF, or NT-3. After 4 days, cellswere labeled with 1 /xCi/well [3H]dThd, incubated for an additional
20 h, and harvested at day 5. Results indicated that the addition ofconcentrations of NT as high as 100 ng/ml to SY5Y cells did not altercell proliferation (Fig. 2A). To evaluate the effects of BDNF onTrkB-expressing SY5Y cells, cells were treated with either 1 nM RAor l UMRA to induce p!45TrkB and cultured in the presence of 0, 10,
or 100 ng/ml BDNF for 5 days. The results indicate that BDNF did notsignificantly alter the growth of TrkB-expressing SY5Y cells(Fig. 2B). Thus, in contrast to the growth-inhibiting effects of NGF onPC 12 cells (42), neither BDNF, NT-3, nor NGF significantly affect
the growth of the SY5Y NB cells.One of the properties of NTs is to promote neuronal cell survival.
To determine if BDNF affected the survival of TrkB-expressing NB
cells, SY5Y cells were treated with 1 JUMRA or 5 /AMRA and 10ng/ml BDNF. After 5 days, cultures were washed and reincubated inserum-free media in the absence or presence of 10 ng/ml BDNF andincubated for an additional 7 days. At various times, [3H]dThd uptake
was monitored and viability was assessed by trypan blue exclusion. Arepresentative experiment (of four performed) depicted in Fig. 2Cindicates that there was a significant decrease in [3H]dThd incorpo-
150
« 125noE 100-
a.u 75 -
50 -
25 -
B
ooc
8
140i
120 -
100
SO
40-
20 -
neX
O.u
20 40 60
ng/ml80 100
20 40 60
ng/ml80 100
246
DAYS (POST TREATMENT)
Fig. 2. A, effect of NTs on SY5Y cell proliferation. SY5Y cells were treated with 0,10,or 100 ng/ml BDNF (•),NGF (A), or NT-3 (•)for 5 days and DNA synthesis wasevaluated at indicated time. B, effect of BDNF on proliferation of TrkB-expressing SY5Ycells. SY5Y cells were plated 5 X IO3 cells/well, and after 24 h cells were treated with
solvent control (O), 1 nM (A), or l JÕMRA (A) in the presence of 0, 10, or 100 ng/mlBDNF for 5 days. C, effect of BDNF on proliferation and survival of differentiated cellsin serum-free conditions. SY5Y cells were treated with a combination of 10 ng/ml BDNFand 1 fAMRA for 6 days. After 6 days, cells were washed and placed in serum-freemedium in the presence (•)or absence (O) of 10 ng/ml BDNF for indicated time.
ration in cells cultured in the absence of BDNF. Cell viability wasassessed at day 7. Ten percent of the cells were nonviable in cultureslacking BDNF compared to only 4% nonviable cells in culturestreated with BDNF. In similar experiments in which the cell numberwas assessed using a colorimetrie assay for protein, a 20% decrease in
1800
on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
TrkB IN NEUROBLASTOMA
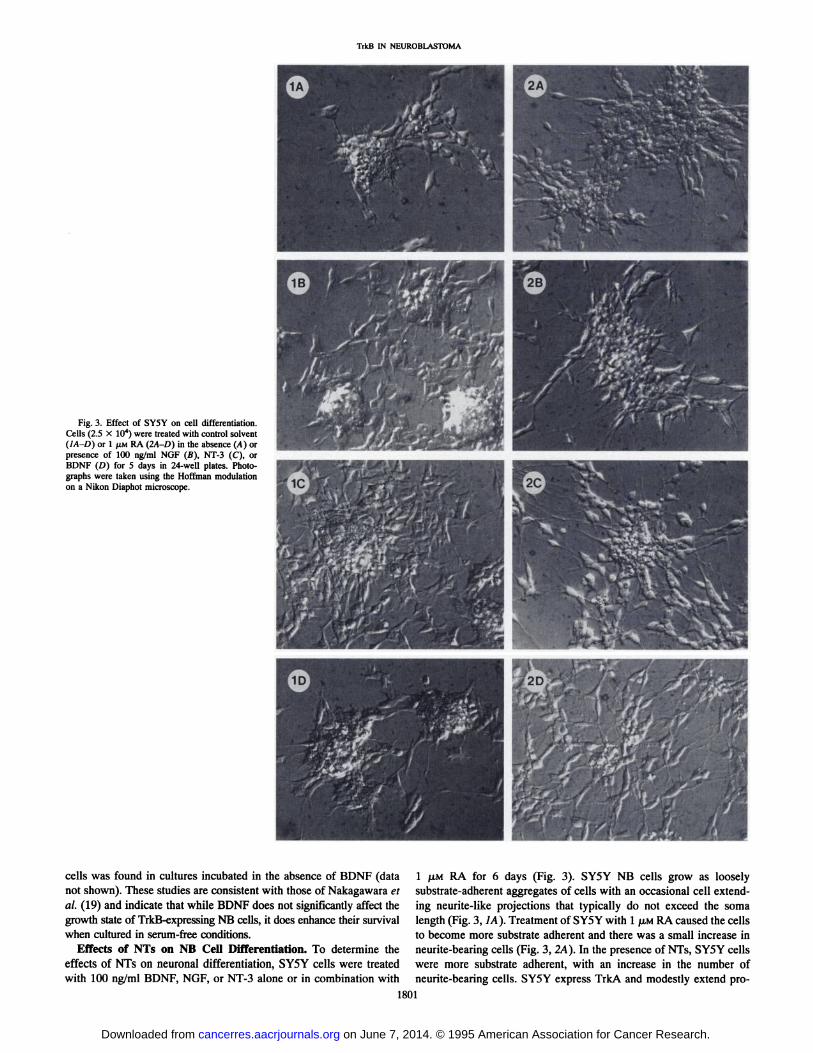
Fig. 3. Effect of SY5Y on cell differentiation.Cells (2.5 X IO4) were treated with control solvent(IA-D) or 1 /¿MRA (2A-D) in the absence (A) orpresence of 100 ng/ml NGF (B), NT-3 (C), orBDNF (D) for 5 days in 24-well plates. Photographs were taken using the Hoffman modulationon a Nikon Diaphot microscope.
cells was found in cultures incubated in the absence of BDNF (datanot shown). These studies are consistent with those of Nakagawara etal. (19) and indicate that while BDNF does not significantly affect thegrowth state of TrkB-expressing NB cells, it does enhance their survivalwhen cultured in serum-free conditions.
Effects of NTs on NB Cell Differentiation. To determine theeffects of NTs on neuronal differentiation, SY5Y cells were treatedwith 100 ng/ml BDNF, NGF, or NT-3 alone or in combination with
1 P.M RA for 6 days (Fig. 3). SY5Y NB cells grow as looselysubstrate-adherent aggregates of cells with an occasional cell extending neurite-like projections that typically do not exceed the soma
length (Fig. 3, IA). Treatment of SY5Y with l JU.MRA caused the cellsto become more substrate adherent and there was a small increase inneurite-bearing cells (Fig. 3, 2A). In the presence of NTs, SY5Y cells
were more substrate adherent, with an increase in the number ofneurite-bearing cells. SY5Y express TrkA and modestly extend pro-
1801
on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TrkB IN NEUROBLASTOMA
Fig. 4. BDNF-induced differentiation of RA-
treated SY5Y cells is dose dependent. Cells weretreated with media (A), 1.0 ng/ml (fi). 3 ng/ml (C), or10 ng/ml (D) BDNF in culture containing either 95%ethanol (1A-D) or l UM RA (2A-D) for 6 days.
Photographs were taken using a Nikon microscopewith Hoffman modulation.
1802
on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
TrkB IN NEUROBLASTOMA
BDNF(ng/ml)
PTyr
0 1 3 10—200K
—110K
TrkB—110K
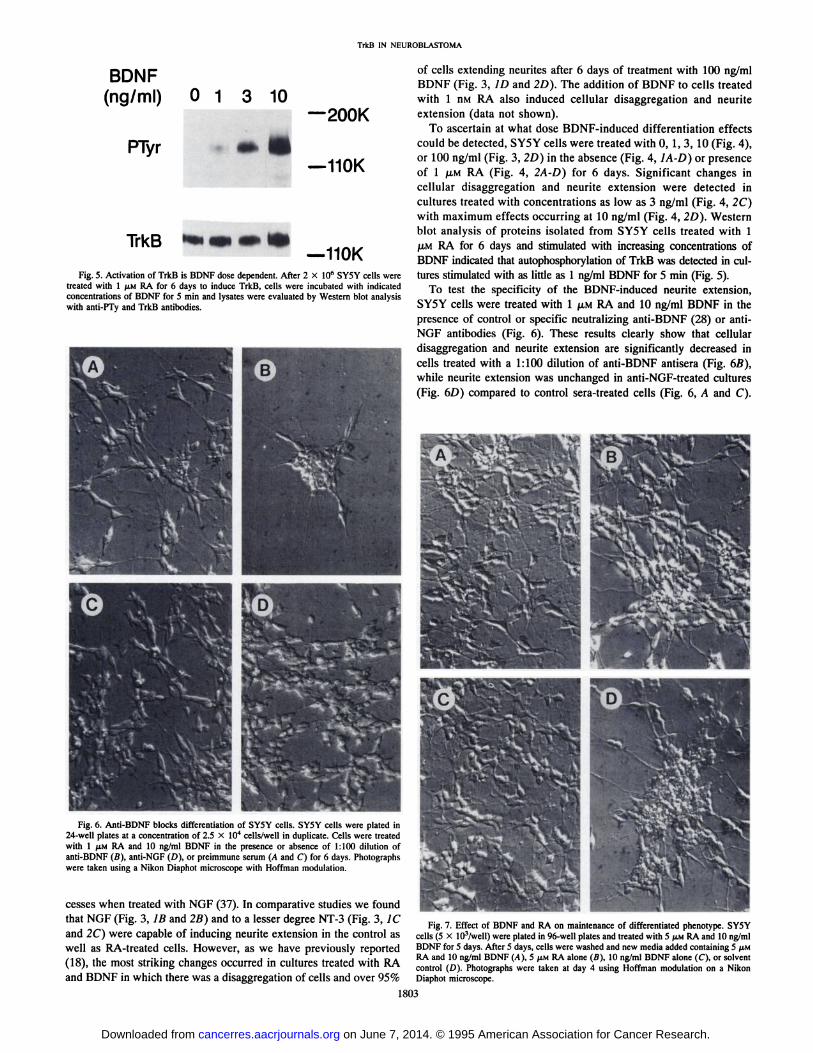
Fig. 5. Activation of TrkB is BDNF dose dependent. After 2 X 10" SY5Y cells were
treated with 1 JIM RA for 6 days to induce TrkB, cells were incubated with indicatedconcentrations of BDNF for 5 min and lysates were evaluated by Western blot analysiswith anti-PTy and TrkB antibodies.
Fig. 6. Anti-BDNF blocks differentiation of SY5Y cells. SY5Y cells were plated in24-welf plates at a concentration of 2.5 X IO4 cells/well in duplicate. Cells were treated
with 1 fiM RA and 10 ng/ml BDNF in the presence or absence of 1:100 dilution ofanti-BDNF (B), anti-NGF (D), or preimmune serum (A and C) for 6 days. Photographs
were taken using a Nikon Diaphot microscope with Hoffman modulation.
cesses when treated with NGF (37). In comparative studies we foundthat NGF (Fig. 3, IB and 2B) and to a lesser degree NT-3 (Fig. 3, 1C
and 2C) were capable of inducing neurite extension in the control aswell as RA-treated cells. However, as we have previously reported
(18), the most striking changes occurred in cultures treated with RAand BDNF in which there was a disaggregation of cells and over 95%
of cells extending neurites after 6 days of treatment with 100 ng/mlBDNF (Fig. 3, ID and 2D). The addition of BDNF to cells treatedwith 1 nM RA also induced cellular disaggregation and neuriteextension (data not shown).
To ascertain at what dose BDNF-induced differentiation effects
could be detected, SY5Y cells were treated with 0, 1, 3, 10 (Fig. 4),or 100 ng/ml (Fig. 3, 2D) in the absence (Fig. 4, ¡A-D)or presenceof l /UM RA (Fig. 4, 2A-D) for 6 days. Significant changes in
cellular disaggregation and neurite extension were detected incultures treated with concentrations as low as 3 ng/ml (Fig. 4, 2C)with maximum effects occurring at 10 ng/ml (Fig. 4, 2D). Westernblot analysis of proteins isolated from SY5Y cells treated with 1¡UMRA for 6 days and stimulated with increasing concentrations ofBDNF indicated that autophosphorylation of TrkB was detected in cultures stimulated with as little as 1 ng/ml BDNF for 5 min (Fig. 5).
To test the specificity of the BDNF-induced neurite extension,
SY5Y cells were treated with l JU.MRA and 10 ng/ml BDNF in thepresence of control or specific neutralizing anti-BDNF (28) or anti-
NGF antibodies (Fig. 6). These results clearly show that cellulardisaggregation and neurite extension are significantly decreased incells treated with a 1:100 dilution of anti-BDNF antisera (Fig. 6B),while neurite extension was unchanged in anti-NGF-treated cultures(Fig. 6D) compared to control sera-treated cells (Fig. 6, A and C).
Fig. 7. Effect of BDNF and RA on maintenance of differentiated phenotype. SY5Ycells (5 X lO'/well) were plated in 96-well plates and treated with 5 ;IM RA and 10 ng/ml
BDNF for 5 days. After 5 days, cells were washed and new media added containing 5 u,MRA and 10 ng/ml BDNF (A), 5 JXMRA alone (B), 10 ng/ml BDNF alone (C), or solventcontrol (D). Photographs were taken at day 4 using Hoffman modulation on a NikonDiaphot microscope.
1803
on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
TrkB IN NEUROBLASTOMA
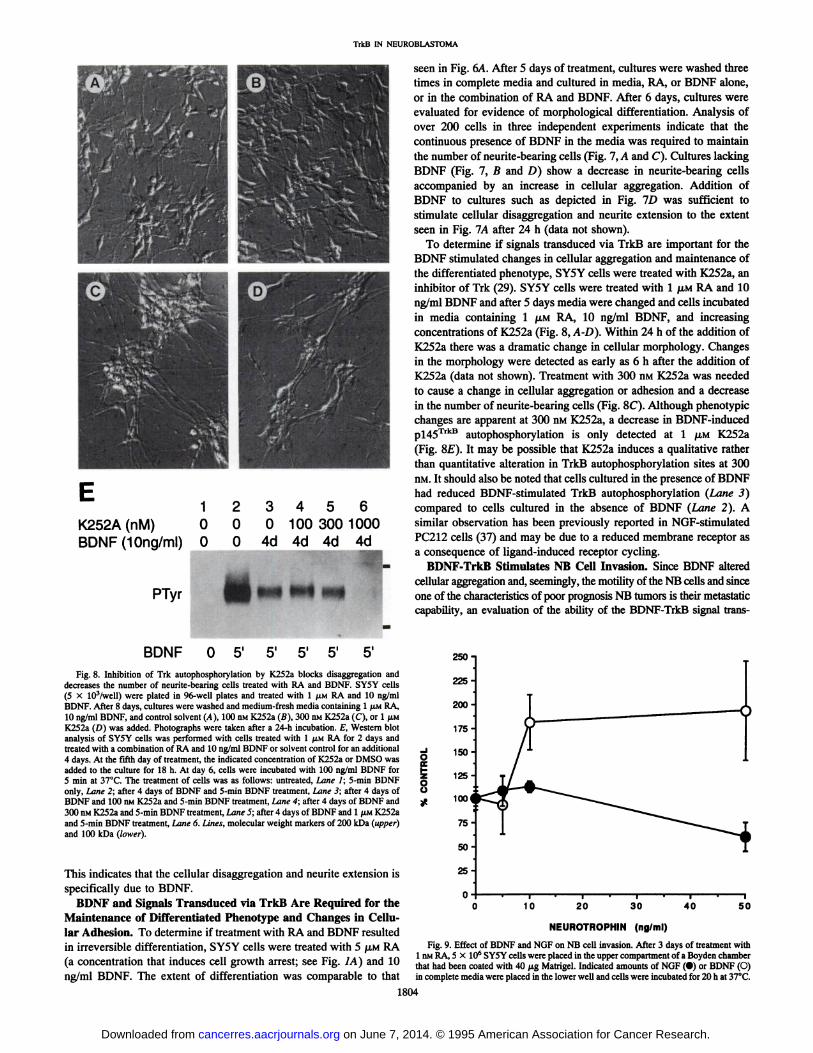
1K252A (nM) 0BDNF(10ng/ml) 0
PTyr
234560 0 100 300 10000 4d 4d 4d 4d
¿f,
^^^^ l^^^fc _mm ^9 mm
seen in Fig. 6A. After 5 days of treatment, cultures were washed threetimes in complete media and cultured in media, RA, or BDNF alone,or in the combination of RA and BDNF. After 6 days, cultures wereevaluated for evidence of morphological differentiation. Analysis ofover 200 cells in three independent experiments indicate that thecontinuous presence of BDNF in the media was required to maintainthe number of neurite-bearing cells (Fig. 7, A and C). Cultures lackingBDNF (Fig. 7, B and D) show a decrease in neurite-bearing cells
accompanied by an increase in cellular aggregation. Addition ofBDNF to cultures such as depicted in Fig. ID was sufficient tostimulate cellular disaggregation and neurite extension to the extentseen in Fig. 1A after 24 h (data not shown).
To determine if signals transduced via TrkB are important for theBDNF stimulated changes in cellular aggregation and maintenance ofthe differentiated phenotype, SY5Y cells were treated with K252a, aninhibitor of Trk (29). SY5Y cells were treated with l JU.MRA and 10ng/ml BDNF and after 5 days media were changed and cells incubatedin media containing 1 /XM RA, 10 ng/ml BDNF, and increasingconcentrations of K252a (Fig. 8, A-D). Within 24 h of the addition of
K252a there was a dramatic change in cellular morphology. Changesin the morphology were detected as early as 6 h after the addition ofK252a (data not shown). Treatment with 300 nM K252a was neededto cause a change in cellular aggregation or adhesion and a decreasein the number of neurite-bearing cells (Fig. 8C). Although phenotypicchanges are apparent at 300 nM K252a, a decrease in BDNF-inducedp!45TrkB autophosphorylation is only detected at l JU.MK252a
(Fig. 8E). It may be possible that K252a induces a qualitative ratherthan quantitative alteration in TrkB autophosphorylation sites at 300nM. It should also be noted that cells cultured in the presence of BDNFhad reduced BDNF-stimulated TrkB autophosphorylation (Lane 3)
compared to cells cultured in the absence of BDNF (Lane 2). Asimilar observation has been previously reported in NGF-stimulated
PC212 cells (37) and may be due to a reduced membrane receptor asa consequence of ligand-induced receptor cycling.
BDNF-TrkB Stimulates NB Cell Invasion. Since BDNF alteredcellular aggregation and, seemingly, the motility of the NB cells and sinceone of the characteristics of poor prognosis NB tumors is their metastaticcapability, an evaluation of the ability of the BDNF-TrkB signal trans-
BDNF O 51 5'
Fig. 8. Inhibition of Trk autophosphorylation by K252a blocks disaggregation anddecreases the number of neurite-bearing cells treated with RA and BDNF. SY5Y cells(5 X 103/well) were plated in 96-well plates and treated with l JJ.MRA and 10 ng/mlBDNF. After 8 days, cultures were washed and medium-fresh media containing 1 JIMRA,
10 ng/ml BDNF, and control solvent (A), 100 nM K252a (B), 300 nM K252a (C), or l /IMK252a (D) was added. Photographs were taken after a 24-h incubation. £,Western blotanalysis of SY5Y cells was performed with cells treated with 1 /¿MRA for 2 days andtreated with a combination of RA and 10 ng/ml BDNF or solvent control for an additional4 days. At the fifth day of treatment, the indicated concentration of K252a or DMSO wasadded to the culture for 18 h. At day 6, cells were incubated with 100 ng/ml BDNF for5 min at 37°C.The treatment of cells was as follows: untreated. Lane I; 5-min BDNF
only. Lane 2; after 4 days of BDNF and 5-min BDNF treatment, Lane 3; after 4 days ofBDNF and 100 nM K252a and 5-min BDNF treatment, Lane 4; after 4 days of BDNF and300 nM K252a and 5-min BDNF treatment, Lane 5; after 4 days of BDNF and 1 fiu K252aand 5-min BDNF treatment. Lane 6. Lines, molecular weight markers of 200 kDa (upper)
and 100 kDa (lower).
This indicates that the cellular disaggregation and neurite extension isspecifically due to BDNF.
BDNF and Signals Transduced via TrkB Are Required for theMaintenance of Differentiated Phenotype and Changes in Cellular Adhesion. To determine if treatment with RA and BDNF resultedin irreversible differentiation, SY5Y cells were treated with 5 JJ.MRA(a concentration that induces cell growth arrest; see Fig. IA) and 10ng/ml BDNF. The extent of differentiation was comparable to that
o8s?250-225-200-175-150-125-
100i75
150-
25-
10 20 30 40 50
NEUROTROPHIN (ng/ml)
Fig. 9. Effect of BDNF and NGF on NB cell invasion. After 3 days of treatment with1 nM RA, 5 X IO6 SY5Y cells were placed in the upper compartment of a Boyden chamber
that had been coated with 40 /ng Matrigel. Indicated amounts of NGF (•)or BDNF (O)in complete media were placed in the lower well and cells were incubated for 20 h at 37°C.
1804
on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TrkB IN NEUROBLASTOMA
auction pathway to affect NB cell invasion was performed. SY5Y NBcells treated with 1 nM RA for 3 days were placed in the upper compartment of a Boyden chamber that had been coated with 40 /¿gMatrigel, anartificial basement membrane preparation. Under these conditions cellsare unable to penetrate to the lower surface of the filter without elaborating proteolytic enzymes. Complete media, with the addition of indicated amounts of BDNF or NGF, were placed in the lower chambers.After an overnight incubation, the filter was processed as described in"Materials and Methods" and invading cells were quantified. The results
represent the average total number of invasive cells from four independent assays and clearly show that concentrations of BDNF as low as 10ng/ml stimulate a 2-fold increase in the number of invasive NB cellscompared to untreated TrkB-expressing NB cells (Fig. 9). The effect ofNGF on the metastatic capability of 1 nMRA-treated SY5Y cells was also
evaluated. In contrast to the effects of BDNF on SY5Y cell invasion, theaddition of NGF at 50 ng/ml caused a decrease in invasiveness (Fig. 9).These results indicate that the activation of the TrkB signal transductionpathway by BDNF stimulates NB cell invasion, while the NGF-TrkA
signal transduction pathway may inhibit NB cell invasion.
DISCUSSION
The observations that TrkA is expressed by good prognosis NB tumors(14-17) has stimulated interest in determining the role and biological
consequences of the expression of the Trk family of receptors and theirligands in NB tumors. Recently we found that a number of NB cell linesconstitutively expressed BDNF but failed to express pl45trkB unless
stimulated with RA (18). In a study of poor prognosis NB tumors and celllines, it was found that poor prognosis NB tumors constitutively expressBDNF and variably express pl45TrkB or p95TrkB(19).
We have utilized RA to induce TrkB in the SY5Y cell line that doesnot constitutively produce readily detectable levels of BDNF or TrkB.Clearly, RA stimulates a number of changes in NB cells which mayaffect BDNF-TrkB signal transduction; however, the ability to addexogenous BDNF to SY5Y cell-expressing TrkB enables biological
and biochemical studies of the consequences of the activation of theTrkB signal transduction pathway by BDNF to be evaluated. Ourstudy did not reveal any evidence that BDNF (Fig. 4B) or the otherTrkB ligands, NT-3 and NT-4/5 (data not shown), significantly alteredcell proliferation in NB cells. The data indicated that under serum-free conditions, the survival of TrkB-expressing SY5Y cells was
maintained by treatment with BDNF (Fig. 5) and cell numbersdecreased in cultures lacking BDNF. This supports a previousfinding that TrkB mediated a survival response under conditions oflow serum in one NB cell line that was found to constitutivelyexpress pl45TrkB and BDNF (19). The survival capability of NTs
and their receptors is well documented in normal neural cellsand constitutive TrkB expression during tumorigenesis could impart a survival advantage to an immortal or tumorigenic cell.Furthermore, the biological effects of BDNF-TrkB signal trans
duction in NB cells are distinct from those noted in two othermodel systems in which TrkB-mediated signal transduction was
analyzed. In PC12 cells transfected with TrkB, treatment withBDNF arrests cell growth and induces differentiation (43), while inNIH-3T3 cells transfected with TrkB, transformed foci form (44).
These findings support the concept that the intracellular environment contributes to the potential diversity of biological responsesthat may occur upon activation of a signal transduction pathway. Furthermore, they underscore the necessity to study the biological effects ofactivation of endogenous receptors in their natural milieu.
Aside from mediating neuronal survival, NTs stimulate morphological and biochemical differentiation of neural cells. In NB cells, theeffects of NTs alone are modest; however, treatment of cells with NTs
in combination with RA results in dramatic changes. BDNF stimulated cellular disaggregation and neurite extension to a degree notapparent in cultures treated with the other NTs. Cellular disaggregation and neurite extension occurred in cells treated with 1 nMand 1 /MMRA, indicating that the expression of a differentiated phenotype canoccur in proliferating cells. Since most NB cell lines and tumorsconstitutively produce BDNF, we focused our attention on the analysis of the effects of this NT. We found that concentrations as low as1 ng/ml stimulated autophosphorylation of pl45TrkB, while 3 ng/ml
consistently induced cellular disaggregation and neurite extension.Experiments utilizing the anti-BDNF blocking antibody and the wash
out experiments indicated the effects on cell morphology were specific and reversible. Cellular disaggregation and the proportion ofneurite-bearing cells was altered when the BDNF-TrkB signal transduction pathway was inhibited with Trk-specific tyrosine kinase in
hibitor K252a.The metastatic process has been shown to be a multistep process
marked by changes in cellular adhesion, attachment to basementmembrane, elaboration of proteolytic enzymes, locomotion, and additional changes in cell adhesion to form a metastatic nodule (45, 46).The changes in morphology noted in BDNF-treated TrkB-expressingSY5Y cells, the ability of a-BDNF antibodies to block this effect, andthe ability of the Trk inhibitor K252a to reverse BDNF-TrkB-inducedmorphological changes support the proposal that the BDNF-TrkB
signal transduction pathway stimulates the changes in cellular aggregation that are a prerequisite for metastasis. The finding that BDNFstimulates a 2-fold increase in NB cell invasion while another NT,
NGF, actually inhibits cell invasion supports the hypothesis that theBDNF-TrkB signal transduction pathway contributes to the metastaticbehavior of poor prognosis NB cells. Although the low-affinity NTreceptor p75NTR has been shown to stimulate melanoma cell invasion
(47), it is unlikely that this receptor mediates these effects in NB sinceNGF, which also binds p75NTR, inhibits cell invasion. We used 1 nM
RA to induce TrkB because this concentration does not affect theproliferation potential of SY5Y cells although it remains to be determined whether growth inhibition affects the invasion of NB cells. Thebiochemical mechanisms mediating the BDNF-TrkB-induced changes
in cellular adhesion and invasion are currently under investigation.The expression of BDNF and TrkB by NB tumor cells may be due
to their neural crest cell origin. The processes involved in dispersionof migratory neural crest cells and the subsequent aggregation of thesecells into tissues such as sympathetic ganglia or the adrenal medullaare analogous to those occurring in the metastatic process. A study byKalcheim and Gendreau (48), showed that BDNF and laminin, but notNGF, were necessary for the formation of sympathetic ganglia by migrating neural crest cells, indicating that the BDNF-TrkB signal trans
duction pathway may play a role in normal neural crest migration.Our findings that the activation of the BDNF-TrkB signal trans
duction pathway in NB cells stimulates cell survival and invasivenessis in sharp contrast to the biological effects stimulated by NGF. In thisstudy we show cellular invasion is stimulated by BDNF-induced
signal transduction, but inhibited by NGF. Studies by Nakagawara etal. (19) and confirmed in this report indicate that BDNF does notinhibit NB cell growth, yet supports NB cell survival. These resultsshow a contrasting role for the BDNF compared to another member ofthe NT family NGF, which induces cell growth arrest in NB cellsexpressing high levels of transfected TrkA (23) and survival ofprimary cultures of NB tumor cells (14). Thus, in NB cells in vitro, theexpression of TrkA or TrkB results in distinct biological responseswhen their respective signal transduction pathways are activated.
One of the hallmarks of poor prognosis NB tumors is that tumorsare metastatic and survive despite intensive chemotherapeutic regimens. Our observation that in NB cell lines activation of the BDNF-
1805
on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

TrkB IN NEUROBLASTOMA
TrkB signal transduction pathway increases Å“il survival and invasive-
ness suggests that this signal transduction pathway plays a role in theseprocesses and affects the biological behavior of these tumors which maycontribute to the poor outcome in these patients. Furthermore, the differential Trk gene expression noted in NB tumors supports the hypothesisthat good and poor prognosis NB tumors correspond to distinct lineagesof neural crest cells or cells at a different stage of development. Thisraises the possibility that the differences in the clinical course of patientswith these tumors may reflect differences in the basic biology of theneural crest cells at the time at which the genetic alterations leading totumorigenesis occurred. These findings suggest that the biological behavior of these tumors is affected by their expression of BDNF-TrkB.
Strategies aimed at interfering with this signal transduction pathway maybe useful in the treatment of poor prognosis NB tumors.
ACKNOWLEDGMENTS
We thank J. Carnahan for making available the BDNF-neutralizing antibody
before it was published. Additionally, we are grateful to J. Winslow, K.Nikolics, G. Burton, R. Scott, and J. Vaught for NT-3 and BDNF; A. Rosenthalfor NT-4/5; D. Morrison for anti-PTyr; D. Martin-Zanca for the human Trk
cDNA; D. Middlemas and T. Hunter for probes and cDNA to TrkB; L.Reichardt for the BDNF cDNA; and M. Glickman for the K252a. We alsothank J. Hughes for manuscript preparation and editorial assistance.
REFERENCES
1. LeDouarin, N. The Neural Crest. London: Cambridge University Press, 1981.2. Beckwith, J. B., and Perrin, E. V. In situ neuroblastomas: a contribution to the natural
history of neural crest tumors. Am. J. Pathol., 43: 1089-1104, 1963.3. Triche, T. J., Askin, F. B., and Rissane, J. M. Neuroblastoma, Ewing's sarcoma, and
the differential diagnosis of small-, round-, blue-cell tumors. In: M. Finegold (éd.),Pathology of Neoplasia ¡nChildren and Adolescents, pp. 145-195. Philadelphia:
W. B. Saunders, 1986.4. Fox, F., Davidson, J., and Thomas, L. Maturation of sympathicoblastoma and gan-
glioneuroma. Cancer (Phila.), 12: 108-116, 1959.5. Evans, A. E., Chatten, J., D'Angio, G. J., Gershon, G. M., Robinson, J., and
Schnaufer, L. A review of 17 IV-S neuroblastoma patients at the Children's Hospital
of Philadelphia. Cancer (Phila.), 45: 833-839, 1980.6. Levi-Montalcini, R. The nerve growth factor: thirty-five years later. Science
(Washington DC), 237: 1154-1164, 1987.7. Barde, Y-A. The nerve growth factor family. Prog. Growth Factor Res., 2: 237-248,
1990.8. Hefti, F., Dentón, T. L., Knusel, B., and Lapchak, P. A. Neurotrophic factors: what
are they and what are they doing? In: S. E. Loughtin and J. P. Fallón (eds.),Localization and Function of Neurotrophic Factors, pp. 25-49. New York: Academic
Press, Inc., 1992.9. Chao, M. V. Neurotrophin receptors: a window into neuronal differentiation. Neuron,
9: 583-593, 1992.
10. Barbacid, M., Lambelle, F., Pulido, D., and Klein, R. The trk family of tyrosineprotein kinase receptors. Biochim. Biophys. Acta, 7072: 115-127, 1991.
11. Middlemas, D. S., Lindberg, R. A., and Hunter, T. trkB, a neural receptor protein-tyrosine kinase: evidence for a full-length and two truncated receptors. Mol. CellBiol., //: 143-153, 1991.
12. Klein, R., Conway, D., Parada, L. F., and Barbacid, M. The trkB tyrosine proteinkinase gene codes for a second neurogenic receptor that lacks the catalytic kinasedomain. Cell, 61: 647-656, 1990.
13. Dechant, G., Rodriquez-Tebar, A., and Barde, Y. A. Neurotrophin receptors. Prog.Neurobiol., 42: 347-352, 1994.
14. Nakagawara, A., Arima-Nakagawara, M., Scavarda, N. J., Azar, C. G., Cantor, A. B., andBrodeur, G. M. Association between high levels of expression of the TRK gene andfavorable outcome in human neuroblastoma. N. Engl. J. Med., 328: 847-854, 1993.
15. Kogner, P., Barbany, G., Dominici, C., Castello, M. A., Raschella, G., and Persson,H. Coexpression of messenger RNA for TRK protooncogene and low affinity nervegrowth factor receptor in neuroblastoma with favorable prognosis. Cancer Res., 53:2044-2050, 1993.
16. Suzuki, T., Bogenmann, E., Shimada, H., Stram, D., and Seeger, R. C. Lack ofhigh-affinity nerve growth factor receptors in aggressive neuroblastomas. J. Nati.Cancer Inst., 85: 377-384, 1993.
17. Borrello, M. G., Bongarzone, 1., Pierotti, M. A., Luksch, R., Gasparini, M., Collini,P., Pilotti, S., Rizzetti, M. G., Mondellini, P., and DeBernardi, B. trk and retproto-oncogene expression in human neuroblastoma specimens: high frequency of trkexpression in non-advanced stages. Int. J. Cancer, 54: 540-545, 1993.
18. Kaplan, D. R., Matsumoto, K., Lucarelli, E., and Thiele, C. J. Induction of TrkB byretinole acid mediates biologic responsiveness to BDNF and differentiation of humanneuroblastoma cells. Neuron, //: 321-331, 1993.
19. Nakagawara, A., Azar, C. G., Scavarda, N. J., and Brodeur, G. M. Expression and
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
function of TRK-B and BDNF in human neuroblastomas. Mol. Cell Biol., 14:759-767, 1994.Brodeur, G. M., Seeger, R. C., and Schwab, M. Amplification of N-myc in untreated
human neuroblastomas correlates with advanced disease stage. Science (WashingtonDC), 224: 1121-1124, 1984.Baker, D. L., Reddy, U. R., Pleasure, D., Thorpe, C. L., Evans, A. E., Cohen, P. S.,and Ross, A. H. Analysis of nerve growth factor receptor expression in humanneuroblastoma and neuroepithelioma cell lines. Cancer Res., 49: 4142-4146, 1989.
Azar, C. G., Scavarda, N. J., Reynolds, C. P., and Brodeur, G. M. Multiple defects ofthe nerve growth factor receptor in human neuroblastomas. Cell Growth & Differ., 1:421-428, 1990.Matsushima, H., and Bogenmann, E. Expression of trkA cDNA in neuroblastomasmediates differentiation in vitro and in vivo. Mol. Cell Biol., 13: 7447-7456, 1993.
Biedler, J. L., Helson, L., and Spengler, B. A. Morphology on growth, tumorigenicityand cytogenetics of human neuroblastoma cells in continuous culture. Cancer Res.,33: 2643-2649, 1973.
Soppet, D., Escandron, E., Maragos, J., Middlemas, D. S., Reid, S. W., Blair, J.,Burton, L. E., Stanton, B. R., Kaplan, D. R., Hunter, T., Nikolics, K., and Parada,L. F. The neurotrophic factors brain-derived neurotrophic factor and neurotrophin-3are ligands for the trkB tyrosine kinase receptor. Cell, 65: 895-904.
Knüsel,B., Rabin, S. J., Hefti, F., and Kaplan, D. R. Regulated neurotrophin receptorresponsiveness during neuronal migration and early differentiation. J. Neurosci., 14:1542-1554, 1994.Allendoerfer, K. L., Gabelli, R. J., Escandón, E., Kaplan, D. R., Nikolics, K., andShatz, C. J. Regulation of neurotrophin receptors during the maturation of themammalian visual system. J. Neurosci., 14: 1795—1811,1994.Ghosh, A., Carnahan, J., and Greenberg, M. E. Requirement for BDNF in activity-dependent survival of cortical neurons. Science (Washington DC), 263: 1618-1623,
1994.Knüsel,B., and Hefti, F. K-252 compounds: Modulators of neurotrophin signaltransduction. J. Neurochem., 59: 1987-1996, 1992.Matsuda, Y., and Fukuda, J. Inhibition by K252a, a new inhibitor of protein kinases,of nerve growth factor-induced neunte growth of chick embryo dorsal root ganglioncells. Neurosci. Lett., 87: 11-17, 1988.Thiele, C. J., Reynolds, C. P., and Israel, M. A. Decreased expression of N-m>cprecedes retinoic acid-induced morphological differentiation of human neuroblastoma. Nature (Lond.), 313: 404-406, 1985.Thiele, C. J., Deutsch, L. A., and Israel, M. A. The expression of multiple protoon-
cogenes is differentially regulated during retinoic acid induced maturation of humanneuroblastoma cell lines. Oncogene, 3: 281-288, 1988.Middlemas, D. S., Lindberg, R. A., and Hunter, T. trkB, a neural receptor protein-tyrosine kinase: evidence for a full-length and two truncated receptors. Mol. CellBiol., 11: 143-153, 1991.
Klein, R., Conway, D., Parada, L. F., and Barbacid, M. The trkB tyrosine proteinkinase gene codes for a second neurogenic receptor that lacks the catalytic kinasedomain. Cell, 61: 647-656, 1990.Martin-Zanca, D., Oskam, R., Mitra, G., Copeland, T., and Barbacid, M. Molecularand biochemical characterization of the human trk proto-oncogene. Mol. Cell Biol.,9: 24-33, 1989.Jones, K. J., and Reichardt, L. F. Molecular cloning of a human gene that is a memberof the nerve growth factor family. Proc. Nati. Acad. Sci. USA, 87: 8060-8064, 1990.Kaplan, D. R., Martin-Zanca, D., and Parada L. F. Tyrosine phosphorylation andtyrosine kinase activity of the Irk proto-oncogene product induced by NGF. Nature(Lond.), 350: 158-160, 1991a.Kaplan, D. R., Hempstead, B. L., Martin-Zanca, D., Chao, M. V., and Parada, L. F.The trk proto-oncogene product: a signal transducing receptor for nerve growthfactor. Science (Washington DC), 252: 554-558, 1991b.Hempstead, B. L., Rabin, S. J., Kaplan, L., Reid, S, Parada, L. F., and Kaplan, D. R.Overexpression of the trk tyrosine kinase rapidly accelerates nerve growth factor-induced differentiation. Neuron, 9: 1-20, 1992.Sidell, N. Retinoic acid-induced growth inhibition and morphologic differentiation ofhuman neuroblastoma cells in vitro. !. Nati. Cancer Inst., 68: 589-593, 1982.
Greene, L. A., and Tischler, A. Establishment of a nonadrenergic clonal line of ratadrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Nati.Acad. Sci. USA, 73: 2424-2428, 1976.
Smith, D. S., King, C. S., Pearson, E., Gittinger, C. K., and Landreth, G. Selectiveinhibition of nerve growth factor-stimulated protein kinases by K-252a and 5'-5-
methyladenosine in PC12 cells. J. Neurochem., 53: 800-806, 1989.
Squinto, S. P., Stitt, T. N., Aldrich, T. H., Davis, S., Bianco, S. M., Radziejewski, C.,Glass, D. J., Masiakowski, P., Furth, M. E., Valenzuela, D. M., DiStefano, P. S., andYancopoulos, G. D. trkB encodes a functional receptor for brain-derived neurotrophicfactor and neurotrophin-3 but not nerve growth factor. Cell, 65: 885-893, 1991.
Glass, D. J., Nye, S. H., Hantzopoulos, P., Macchi, M. J., Squinto, S. P., Goldfarb, M.,and Yancopoulos, G. D. TrkB mediates BDNF/NT-3-dependent survival and proliferation in fibroblasts lacking the low affinity NGF receptor. Cell, 66: 405-413,1991.
Liotta, L. A., and Kohn, E. Cancer invasion and metastasis. J. Am. Med. Assoc., 263:1123, 1990.Hynes, R., and Lander, A. D. Contact and adhesive specificities in the association,migration, and targeting of cells and axons. Cell, 68: 303—322,1992.
Herrmann, J. L., Menter, D. G., Hamada, J., Marchetti, D., Nakajima, M., andNicolson, G. L. Mediation of NGF-stimulated extracellular matrix invasion by thehuman melanoma low-affinity p75 neurotrophin receptor: melanoma p75 functionsindependently of trkA. Mol. Cell Biol., 4: 1205-1216, 1993.Kalcheim, C., and Gendreau, M. Brain-derived neurotrophic factor stimulates survival and neuronal differentiation in cultured avian neural crest. Dev. Brain Res., 41:79-86, 1988.
1806
on June 7, 2014. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























