Embed Size (px)
Citation preview

Evidence for Context-sensitivity of Grasp Representations in Human Parietal and Premotor 1 Cortices 2
3 4
Mattia Marangon 1,2, Stéphane Jacobs 1,3, Scott H. Frey 1,4 5 1 Psychology Department, University of Oregon, Eugene, USA; 2 presently at University of Padua, 6 Department of General Psychology, Padua, Italy; 3 presently at INSERM U1028; CNRS 5292; 7 University Lyon I, Lyon Neuroscience Research Center, ImpAct team, Bron, France; 4 Lewis 8 Center for Neuroimaging, University of Oregon, Eugene, USA; 9 10 Correspondence to Scott H. Frey ([email protected]): 11 Department of Psychology, 1227 University of Oregon, Eugene, OR 97403, USA 12 13 Abstract: 248 words 14 Running Head: Context-specificity of grasp representations 15 16 17 18 19 20
21
Articles in PresS. J Neurophysiol (March 2, 2011). doi:10.1152/jn.00796.2010
Copyright © 2011 by the American Physiological Society.

Context-specificity of grasp representations 2Abstract 22 Grasp-related responses in neurons of the macaque rostral inferior parietal lobule (PF/PFG and the 23 anterior intraparietal area (AIP)) are modulated by task context. Event-related fMRI was used to 24 ask whether this is true in putative homologues of the human cortex, the human rostral inferior 25 parietal lobule (rIPL) and the anterior intraparietal sulcus (aIPS). Fifteen healthy, right-handed, 26 adults were required to select prospectively the most comfortable way to grasp a horizontally-27 oriented handle using the cued hand (left or right). In the NO ROTATION condition, the task was 28 simply to grasp the handle, while in the ROTATION condition the goal was to plan to grasp and 29 rotate it into a vertical orientation with the cued end (medial or lateral) pointing downward. In both 30 conditions, participants remained still and indicated their grip preferences by pressing foot pedals. 31 As in overt grasping, participants’ grip preferences were significantly influenced by anticipation of 32 the demands associated with handle rotation. Activity within the aIPS and rIPL increased 33 bilaterally in both the ROTATION and NO ROTATION conditions. Importantly, these responses 34 were significantly greater in the ROTATION vs. NO ROTATION condition. Similar context 35 effects were detected in the pre-supplementary motor area (pre-SMA), caudal intraparietal sulcus 36 (cIPS)/superior parietal lobule (SPL), bilateral dorsal (dPMC) and left ventral (vPMC) premotor 37 cortices. Grasp representations within the rIPL and aIPS are sensitive to predicted task demands and 38 play a role in context-sensitive grip selection. Moreover, the findings provide additional evidence 39 that areas involved in the sensorimotor control of grasp also contribute to feedforward planning. 40 41 Key words: inferior parietal lobule; grasping, action selection; fMRI; feedforward planning 42 43
44

Context-specificity of grasp representations 3Introduction 45 The choice of how to grasp an object (e.g., over- vs. under-hand) depends on sensory information 46 concerning the state of the body (e.g., current posture, range of motion) and the target object (e.g., 47 location, orientation), as well as anticipation of forthcoming task demands (e.g., intended object 48 rotation) (Johnson-Frey 2004). For instance, when grasping a handle with the intention to rotate it, 49 participants may select a less-comfortable initial grip in order to end in a more comfortable posture 50 (Rosenbaum and Jorgensen 1992). While much is currently understood about the neural 51 mechanisms involved in online sensorimotor control, comparatively little is known about brain 52 mechanisms that enable anticipated future task demands to influence the selection of movements, 53 including object-oriented grasping. 54 Context-sensitive representations of grasping. Neurophysiological studies in monkeys have 55 identified several areas in parietal and premotor cortices that are involved in grasp-related 56 sensorimotor transformations (Gardner et al. 2007; Rizzolatti and Luppino 2001). These include 57 the rostral inferior parietal lobule (rIPL) within the lateral bank of the intraparietal sulcus (area 58 AIP), area V6A (Fattori et al. 2009), ventral premotor (area F5) and the forelimb sector of dorsal 59 premotor (area F2) cortices (Raos et al. 2004). Neurons along the lateral convexity of the rIPL 60 (areas PF/PFG), that show increased activity during reach-to-grasp, are also sensitive to the larger 61 action context in which these movements are embedded; i.e., they are influenced by prediction of 62 forthcoming task demands. More precisely, a majority of neurons studied responded differentially 63 depending on whether the reach-to-grasp movements were to be followed by bringing the object to 64 the mouth, or placing it in a container (Fogassi et al. 2005; Fogassi and Luppino 2005). Likewise, 65 responses within AIP show selective premovement responses depending on the type of grasp that 66 the animals are instructed to perform during a forthcoming movement (Baumann et al. 2009). It 67 remains unknown whether responses in other regions (e.g., vPMC, dPMC) implicated in 68 sensorimotor control of grasp display such context sensitivity. 69

Context-specificity of grasp representations 4Similar to monkeys, human neuroimaging studies of visually-guided grasping demonstrate 70 consistent activation in rIPL extending into the anterior part of the intraparietal sulcus (aIPS) 71 (Binkofski et al. 1998; Culham et al. 2003; Frey et al. 2005). Likewise, transcranial magnetic 72 stimulation (TMS) over this region can disrupt sensorimotor control of grasp (Rice et al. 2007; Rice 73 et al. 2006; Tunik et al. 2005). Increased activity in premotor cortex has also been reported during 74 some studies of grasp execution (Binkofski et al. 1999; Ehrsson et al. 2000; Ehrsson et al. 2001; 75 Grafton et al. 1996). 76 In contrast to our understanding of the sensorimotor control of grasp, comparatively little is known 77 about the neural mechanisms involved in selecting actions on the basis of anticipated task demands. 78 Parietal and premotor regions show increased activity when participants prospectively decide 79 whether it would be more comfortable to grasp a handle, appearing in various orientations, in an 80 over- or under-hand grip despite the complete absence of overt hand movements (Johnson et al. 81 2002). More recently, volunteers were trained to grasp objects in many orientations with the hands 82 or a novel mechanical tool. Subsequent fMRI testing revealed increased activity within and along 83 the IPS (including aIPS and caudal IPS (cIPS)), left vPMC and dPMC during prospective grip 84 selection (Jacobs et al. 2010). Behavioral data provided strong evidence that these decisions were 85 based on accurate, effector-specific, internal representations. Interestingly, increases in neural 86 activity were the same regardless of the side (left or right) or effector (hand or tool) on which grip 87 selection decisions were based. These findings suggest that at least some of the parietal and 88 premotor regions involved in the on-line control of grasping also participate in forming effector-89 specific representations of grasping actions that are sensitive to anticipated task demands. Whether, 90 like cells in monkey rIPL and AIP, responses in one or more of these areas might be influenced by 91 anticipation of the demands of a movement subsequent to grasping the object is unknown. 92 Presently, we investigated this issue through use of a task in which participants were required to 93 choose whether an under- or over-hand power grip would be most natural to grasp a visually-94 presented handle while undergoing whole-brain, blood oxygen-level dependent (BOLD) functional 95

Context-specificity of grasp representations 5magnetic resonance imaging (fMRI). No overt movements were performed in this prospective 96 planning task, and grip preferences were instead expressed through button presses using the feet. 97 Critically, we varied the intended goal of the movement. In the NO ROTATION condition 98 participants were required to simply select what would be the most natural grip for engaging the 99 handle, while in the ROTATION condition the task involved choosing the grip that they would 100 prefer in order to rotate the handle to a cued orientation once in hand. On the basis of past research, 101 we expected that in the NO ROTATION condition participants would prefer the less awkward 102 overhand grip with either hand (Johnson 2000). However, in the ROTATION condition they would 103 select the grip that would allow them to complete the handle rotation in a comfortable posture 104 within the middle of their range of forearm supination-pronation. This end-state comfort effect is 105 well-established in overt motor performance (Rosenbaum and Jorgensen 1992), and is evidence of 106 context-sensitivity in action selection. 107 In accordance with neurophysiological evidence from monkeys indicating context sensitivity, we 108 predicted that the lateral convexity of the rIPL (Fogassi et al. 2005) and aIPS (Baumann et al. 2009) 109 would show increased activity when participants performed the ROTATION vs. NO ROTATION 110 condition of the prospective grip selection (PGS) task during BOLD fMRI. As reviewed earlier, 111 human left vPMC and bilateral dPMC and cIPS are also implicated in the prospective selection of 112 grasp (Jacobs et al. 2010). To the extent that representations in these areas are sensitive to 113 anticipated task demands that extend beyond prehension of the target handle, they should also show 114 greater activity during the ROTATION vs. NO ROTATION condition. 115 To our knowledge, the end-state comfort effect has not been investigated in a prospective planning 116 task. We therefore undertook a preliminary behavioral experiment in order to establish the effects 117 of anticipated handle rotation on overt grip selection (OGS) using a design similar to that of the 118 main neuroimaging study (Experiment 2). These results would then serve as a standard against 119 which the accuracy of prospectively selected grip preferences expressed during fMRI testing could 120 be compared. 121

Context-specificity of grasp representations 6Experiment 1: Overt Grip Selection Task 122 Participants were asked to grasp a handle using either an over- or under-hand power grip under two 123 instructional conditions. The hand used varied from trial-to-trial. In the NO ROTATION control 124 condition, participants simply used the cued hand to grasp a horizontally oriented handle. In the 125 ROTATION condition, they were instructed to grasp the handle and rotate it 900, placing the cued 126 end of the handle downward. We expected that subjects would show a strong preference for the 127 overhand grip in the NO ROTATION condition. In the ROTATION condition, we expected that 128 participants would show an increased preference for underhand grips when the medial end (pointing 129 rightward on left hand trials and leftward on right hand trials) of the handle was cued. This 130 increased preference for the underhand grip would be considered evidence for context sensitivity of 131 grip selection. 132 Methods 133 In accordance with the Declaration of Helsinki, 13 healthy participants (8 females) (M = 26, SD = 134 3.8 years) gave informed consent to undertake the study. All participants were consistently right-135 handed according to the Edinburgh Handedness Inventory (Oldfield, 1971) and had normal or 136 corrected-to-normal vision. None had a history of neurological or psychiatric illness. 137 Apparatus. Both Experiments 1 and 2 were controlled with a laptop computer running Presentation 138 software (Neurobehavioral Systems Inc., http://nbs.neuro-bs.com/). A wooden rack was positioned 139 within reach of the participant, and the distance was adjusted to accommodate each participant’s 140 arm length. The rack was centered along the participant’s mid-saggital line. A 2” diameter wooden 141 handle (10” length) was suspended horizontally on the rack, with the square end pointing toward 142 the left or right depending on the trial. A wooden 2” x 2” x .5” square was attached to one end of 143 the handle, and a 2” diameter x .5” thick circle was centered on the opposite end. Lying horizontal 144 on the tabletop, directly in front of the rack, was a 20” flat LCD computer monitor fitted with a 145 clear Lexan cover. 146

Context-specificity of grasp representations 7Procedure. Every participant performed 144 trials divided into 3 blocks. Each block consisted of 4 147 repetitions of the 12 different trial types presented in pseudo-random order: 2 hand cues (left, right) 148 x 2 dowel orientations (square end pointing toward the left or right) x 3 target shapes (square, circle 149 or triangle). Figure 1A illustrates the graphical stimuli used in Experiment 2. These are similar to 150 actual handles and targets used here in Experiment 1, with the exception that the current handles 151 were unpainted. 152 Participants sat on a chair with their forearms resting on the tabletop and their left and right index 153 fingers depressing left and right response buttons, respectively. They wore a pair of liquid crystal 154 goggles (Translucent Technologies, Toronto). At the beginning of a run, the participant viewed a 155 left- or right-ward arrow that served to indicate which hand would be used on the subsequent trial. 156 The goggles then became opaque, preventing vision of the display. The experimenter placed the 157 handle on the rack with the square pointing to the left or right, according to the trial list and 158 depressed a key on the keyboard. The goggles became transparent after a variable 3-5s interval and 159 a target (square, circle or triangle) appeared on the monitor below the rack. If the target object was 160 a square or circle, then the participant was to release the response button and use the cued hand to 161 grasp the handle in a power grip, remove it from the rack, and rotate it 900 in order to place the 162 square or circular end (depending on the target’s shape) on the Lexan surface (ROTATION 163 condition). They were not required to align the end of the handle with the target. If the target was a 164 triangle, then they were instructed to use the cued hand to grasp the handle in a power grip and 165 place it lengthwise on the surface (NO ROTATION condition). Participants were told to use the 166 most natural grip (over- or under-hand) in both conditions, and to initiate their movements as soon 167 as possible after the goggles became transparent. Response times were measured from the onset of 168 vision until the response button was released. After each trial was completed, participants returned 169 their hand to the response button. This triggered the appearance of the 500ms hand cue for the next 170 trial, after which the goggles again became opaque and the process was repeated. The entirety of 171 each run was videotaped and grip preferences were coded off-line. 172

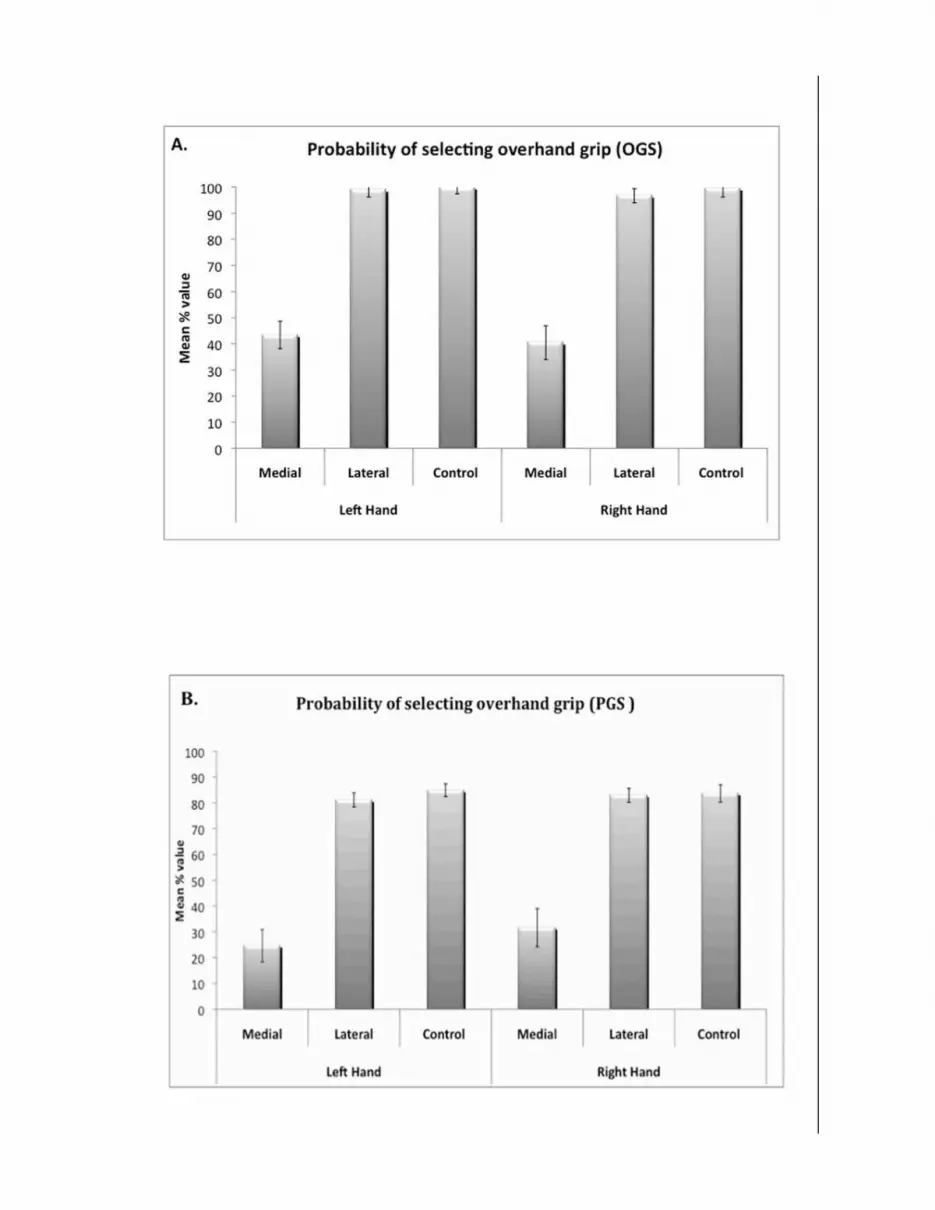
Context-specificity of grasp representations 8Data Analysis. As illustrated in Figure 1C, in order to enable direct comparisons of grip 173 preferences between hands, handle orientation on ROTATION trials was recoded. Medial trials 174 were those in which the cued end of the handle (square or circular depending on the target shape) 175 was pointing toward the participant’s midline. On lateral trials, the cued end was pointing away 176 from the midline. The mean probability of selecting the overhand grip for each condition was then 177 calculated separately for every participant and submitted to 2 (hand: left, right) x 3 (cue type: 178 lateral, medial, control) repeated measures ANOVA. A significance level of p < .05 was used for 179 all statistical tests. 180
------------------------------- 181 Insert Figure 1 about here 182 ------------------------------- 183
Trials with response times (RT) ± 2 SDs from the mean were defined as outliers and removed. 184 Excluded trials constituted 4% of the total number. 185 186 Results & Discussion 187 As expected, differences in grip preferences between hands were non-significant (F < 1.0). There 188 was a main effect of the target (medial, lateral, control) (F(1,11) = 28.08, MSe = 773.314, p < 189 0.001). As illustrated in Figure 2A, participants showed a strong preference for the overhand grip 190 when the lateral end of the handle matched the target, M =97.8%, SD = 10.3. This response pattern 191 did not differ significantly from that of the control condition (M = 99.31%, SD = 9.64), t(11) < 1.0. 192 As expected, subjects were less likely to select the overhand grip in the ROTATION condition 193 when the medial end of the handle was cued (M = 41.91%, SD = 24.21) as compared with either the 194 lateral end (t(11) = 5.614, p < 0.001) or control condition, t(11) = 5.611, p < 0.001 (Figure 2A). 195 Response times did not differ significantly between hands or target types, p < .20. 196

Context-specificity of grasp representations 9Adopting an overhand grip in the lateral cue condition and an underhand grip in the medial cue 197 condition allowed participants to complete handle rotations in a comfortable posture in the middle 198 of their range of hand pronation-supination. The fact that the current grip preferences exhibited this 199 pattern is further evidence that grip selection involves anticipation of forthcoming task demands 200 (Rosenbaum and Jorgensen 1992). Because participants continued to prefer an overhand grip in the 201 lateral cue condition, this result cannot be attributed to merely choosing overhand in the NO 202 ROTATION condition and underhand in the ROTATION condition. 203
------------------------------- 204 Insert Figure 2 about here 205 ------------------------------- 206
Experiment 2: fMRI Prospective Grip Selection Task 207 As expected on the basis of previous findings (Rosenbaum and Jorgensen 1992; Rosenbaum et al. 208 1992), participants in Experiment 1 adapted their grip preferences to the anticipated demands of a 209 forthcoming handle rotation. Experiment 2 asked whether this end-state comfort effect would also 210 be exhibited when participants were asked to decide how they would grasp the handle in both 211 ROTATION and NO ROTATION conditions while remaining still; i.e., in a purely prospective grip 212 selection (PGS) task that lacks overt movement and associated sensory feedback (Johnson 2000). If 213 so, then participants should again show an increased preference for underhand grips in the 214 ROTATION condition when the medial, but not the lateral, end of the handle is cued. To the extent 215 that this is the case, we reasoned that fMRI data acquired during performance of this PGS task 216 could be used to identify regions showing context sensitivity during the planning of reach-to-grasp 217 movements. 218 Based on earlier findings regarding prospective planning of grasp (Jacobs et al. 2010; Johnson et al. 219 2002), we expected that PGS in both the NO ROTATION and ROTATION conditions would be 220 associated with increased activity within and along the IPS (including aIPS and cIPS), dPMC, and 221

Context-specificity of grasp representations 10left vPMC. Further, results showing context-sensitivity in monkey rIPL (Fogassi et al. 2005; 222 Fogassi and Luppino 2005) and AIP (Baumann et al. 2009) suggest that putatively similar areas in 223 the human (rIPL & aIPS) might, in particular, show greater increases in activity during 224 ROTATION vs. NO ROTATION conditions. The reason for this expectation is that grip selection 225 in the ROTATION condition involves anticipating the additional demands that would be associated 226 with rotating the handle once in hand. Our use of whole brain imaging, also provided the 227 opportunity to assess whether other regions implicated in sensorimotor control and prospective 228 planning of grasp (cIPS, dPMC and left vPMC) might also show similar context sensitivity. If so, 229 then these findings could serve as an impetus for future single unit investigations in monkeys. 230 Methods 231 Fifteen (9 females) healthy participants (M = 24, SD = 4.3 years) took part in the study after giving 232 written informed consent according with the Declaration of Helsinki. None had participated in 233 Experiment 1, or had a history of neurological or psychiatric illness. All participants were 234 consistently right-handed according to the Edinburgh Handedness Inventory (Oldfield, 1971) and 235 had normal or corrected-to-normal vision. 236 Stimuli consisted of two graphically rendered handles, each with a square end and a round end 237 (Figure 1A). For one handle, the half with the square end was colored pink, and the other tan. The 238 second handle had these colors reversed. Each handle appeared in two orientations, one with the 239 square end on the left and the other with the square end on the right. The handles were realistically 240 shaded to give the appearance of three-dimensionality. As detailed below, on a given trial one of 241 these four handle permutations (2 handles x 2 orientations) was paired with one of three different 242 target objects, a circle, square or triangle. 243 Every trial consisted of six consecutive phases optimized for event-related fMRI testing (Figure 244 1B): 1) Hand cue--500ms left- or right-ward pointing arrow was presented centrally to inform the 245 participant on which hand (left or right) they should base their upcoming grip selection decision. 2) 246

Context-specificity of grasp representations 11Oversampling interval--variable delay (500, 1000,1500 or 2000ms) period time-locked to the 247 acquisition of a volume of functional MRI data (Miezin et al. 2000); 3) Grip Selection cue: 2500ms 248 handle stimulus presented centrally above one of 3 different targets (square, circle or triangle). 4) 249 Variable delay interval: (500, 1000, 1500ms). 5) Response cue: 2000ms duration colored squares 250 indicating which foot pedal (left or right) represented the pink or tan option on that particular trial. 251 Subjects were instructed to press the pedal whose color corresponded to where the thumb side of 252 their hand would be on the handle if grasped. Trial-by-trial variation in the identity of the pedals 253 forced participants to wait for this response cue before indicating their choice. This enabled 254 separation of activity associated with grip selection (our focus) from response button selection and 255 execution. 6) Inter-trial interval: (1500, 2000, 2500, 3000, 3500 or 4000ms) adjusted to bring each 256 trial to an equal 10s duration. The ordering of events within trial was necessarily fixed, due to the 257 sequential relationship between hand preparation, grip selection and response execution. However 258 oversampling and jitter interval durations occurred equally often for each trial type; i.e., 2 hands 259 (left, right) x 2 conditions (ROTATION, NO ROTATION). For a randomly selected 50% of the 260 trials within each run the left foot pedal was used to indicate the “pink” response, and for the 261 remainder this pedal indicated the “tan” response. 262 A centrally presented, white cross remained visible throughout the entirety of each functional run 263 and subjects were instructed to maintain fixation. Each participant performed 6 functional runs. 264 Each run consisted of 32, 10s long experimental trials (16 ROTATION trials--2 hand cues x 4 265 stimulus handles x 2 targets; 16 NO ROTATION trials—2 x 2 hand cues x 4 stimulus handles), 266 plus an additional 8 null trials of equal duration during which no stimuli were presented. The 267 doubling of the NO ROTATION trials was necessary to achieve equity with the total number of 268 ROTATION trials, making the identity of the forthcoming trial unpredictable. 269 Within each run these 40 trials (16 ROTATION, 16 NO ROTATION and 8 null) were presented in 270 pseudo-random order, subject to the constraint that no more than 3 trials of any single type could 271 occur sequentially. 272

Context-specificity of grasp representations 12The PGS task was similar to Experiment 1 except that no overt hand movements were involved. 273 Instead, in both the NO ROTATION and ROTATION conditions, participants were instructed to 274 identify whether they would prefer to place the thumb side of their cued hand on the pink or tan 275 side of the handle if grasping it in a power grip. This enabled us to code whether they preferred an 276 over- or under-hand grip for each trial. In the ROTATION condition, the shape of the target 277 matched one end of the dowel (square or circular target). The task was to select the grip that would 278 be preferred if intending to rotate the handle downward with the indicated hand, placing the cued 279 end on the target. Again, they were not asked to consider aligning the cued end and target shape. 280 On NO-ROTATION trials (triangular target) the task was to select the grip that would be preferred 281 if intending simply to grasp the handle with the cued hand and put it down lengthwise. Target 282 objects appeared centrally below each stimulus dowel (Figure 1A). Again, half of the total number 283 of trials consisted of the NO ROTATION control condition, while the remainder was equally split 284 between lateral and medial ROTATION trials. 285 Before the start of the scanning session, participants practiced the PGS task both outside the 286 scanner and inside an MRI simulator scanner (80 trials in each session). During both mock and 287 actual fMRI testing, participants were lying supine in the scanner with their arms extended at their 288 sides with the palms oriented vertically. They were unable to see their hands. Stimuli were 289 projected onto a screen at the back of the scanner and viewed in an angled mirror attached to the 290 head-coil. 291 Behavioral Data Analyses. As described for Experiment 1, handle orientation for the ROTATION 292 trials were recoded into medial and lateral in order to enable direct comparisons between hands 293 (Figure 1C). In addition, pink and tan responses on all trials were recoded into over- or under-294 hand. The mean probability of selecting the overhand grip for each condition was then calculated 295 separately for every participant and submitted to 2 (hand: left, right) x 3 (cue type: lateral, medial, 296 control) repeated measures ANOVA. Response times were not analyzed because they included not 297 only the time to select a grip, but also the variable delay and response cue intervals. 298

Context-specificity of grasp representations 13MRI data acquisition 299 Functional images were acquired on a Siemens Allegra 3T MRI system (Siemens, Erlangen, 300 Germany) equipped with echo planar imaging (EPI) capabilities, using a standard birdcage coil for 301 radio frequency transmission and signal reception. Blood oxygenation level-dependent sensitive 302 functional images were acquired using a single shot gradient EPI-sequence (TE/TR = 30/2000ms; 303 flip angle = 80°, 33 axial slices, voxel size = 4.0 x 4.0 x 4.0 mm; FOV = 200 mm, thickness = 304 3.0mm). A double gradient echo sequence was used to acquire a field map that was used to correct 305 EPI distortions. Two high-resolution anatomical images were acquired using a 3-D MP-RAGE 306 pulse sequence (TE/TR = 4.38/2500 ms; flip angle = 8.0°, 176 contiguous axial slices, thickness = 307 1.0 mm, voxel size = 1.0 x 1.0mm; FOV = 256 mm). Siemen’s Auto Align Scout and True FISP 308 sequences were executed for each participant at the beginning of each functional data collection run 309 in order to ensure that slices were prescribed in exactly the same positions across runs. DICOM 310 image files were converted to Nifti format using MRIconvert software 311 (http://lcni.uoregon.edu/~jolinda/MRIconvert/) 312 fMRI processing and analysis 313 fMRI data were preprocessed using the FSL toolbox (http://www.fmrib.ox.ac.uk/fsl) Version 4.0.2 314 (Smith et al. 2004). Pre-statistical processing was applied as follows: EPI dewarping using 315 PRELUDE + FUGUE (Jenkinson 2003; 2004), motion correction using MCFLIRT (Jenkinson et al. 316 2002), volumes were smoothed with a Gaussian kernel of 5mm (FWHM) in order to reduce the 317 noise, intensity normalized and filtered with a non linear high pass temporal filter (70.0 s). Skull 318 and surrounding tissue were removed using BET (Smith 2002). Registration to high-resolution 319 structural and/or standard space images was carried out using FLIRT (Jenkinson et al. 2002; 320 Jenkinson and Smith 2001). 321 Each fMRI run for a given subject was modeled separately at the first level. For each run, we 322 created a model with 4 explanatory variables (EVs) and their temporal derivatives to encode the 323

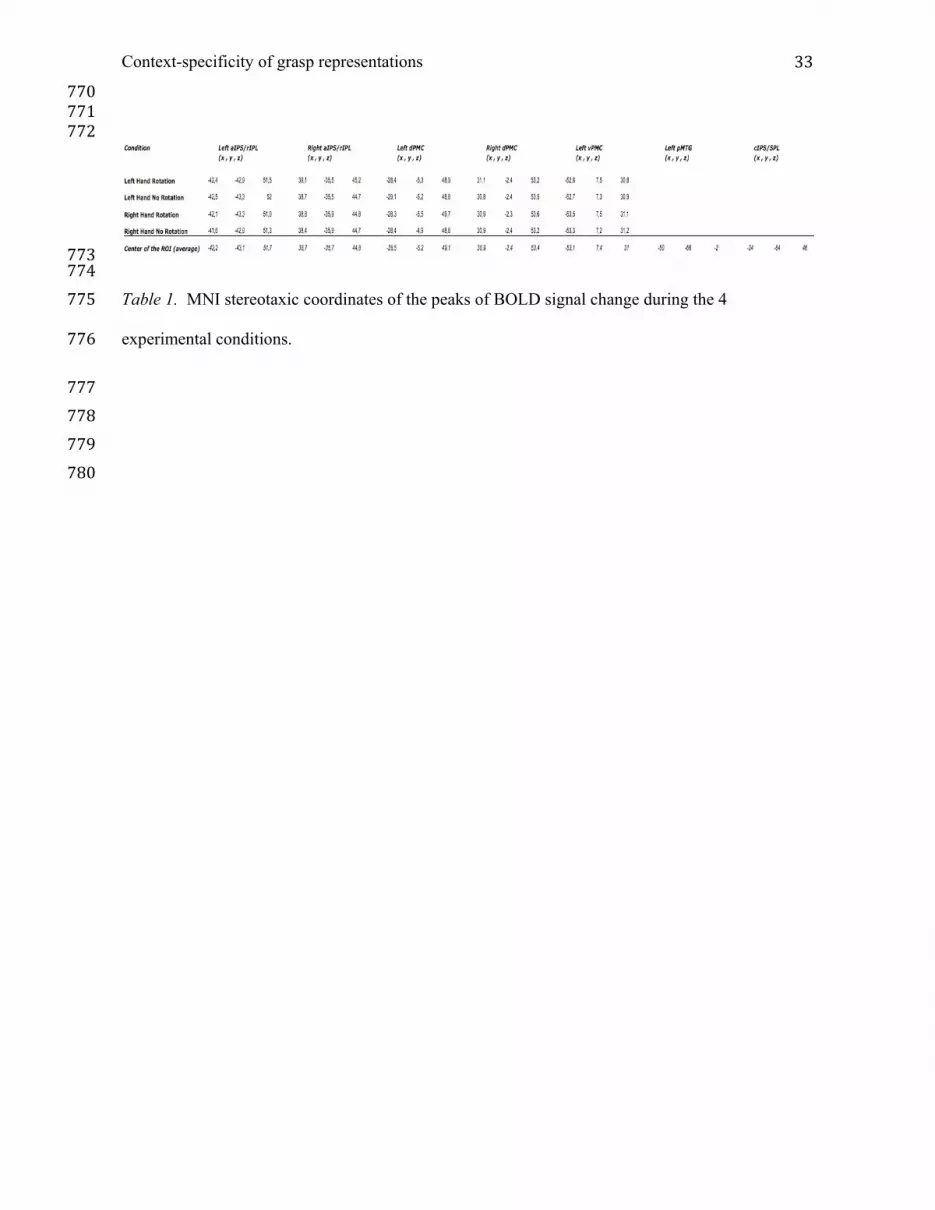
Context-specificity of grasp representations 14conditions under which grip selection decisions were performed in our 2 (hand: left, right) x 2 324 (goal: NO ROTATION, ROTATION) factorial design. These EVs were time-locked to the onset 325 of the handles and target objects (phase #3 above) and included the subsequent 3000ms (2500ms 326 stimulus duration plus the shortest delay interval of 500ms, Figure 1B). A fifth EV coded the 10s 327 null trials that were used as resting baseline. Orthogonal contrasts were used to test for differences 328 between each of the four experimental conditions and resting baseline. 329 Higher-level analyses were performed using FLAME Stage 1 (Beckmann et al. 2003; Woolrich et 330 al. 2004) to model and estimate random-effects components of mixed-effects variance. The 331 resulting first-level contrasts of parameter estimates (COPEs) served as inputs to second-level 332 (within subjects across runs) analyses. COPEs from the second-level analyses then served as inputs 333 to third-level (across participants) analyses. Z- (Gaussianised T/F) statistic images were 334 thresholded using Z > 2.3 and a (corrected) cluster significance threshold of P = 0.05 (Worsley 335 2001). 336 fMRI ANOVA. Next, a more targeted 2 (hand: left, right) x 2 (goal: NO ROTATION, ROTATION) 337 repeated-measures ANOVA was carried out only on those voxels from the initial whole brain 338 analysis that showed evidence of being modulated by the task at the group level; i.e., significant 339 increases in activity within at least one of the four experimental conditions relative to rest (Z > 2.3, 340 corrected cluster significance threshold of p = 0.05). 341 Anatomical localization was undertaken by manual comparison with an atlas (Duvernoy 1991) and 342 by overlaying activation maps on the population, landmark and surface-based atlas (PALs) of Van 343 Essen (Van Essen 2005) using CARET software (Van Essen et al. 2001). 344 Descriptive Region-of-Interest Analysis. Regions-of-interest (ROIs) were defined post-hoc within 345 bilateral rIPL/aIPS, dPMC and left vPMC. Spherical ROIs (5mm radius) were centered on the 346 mean x, y, z coordinates obtained by averaging across the four conditional peaks in each area 347 (Table 1). For every participant, mean percent signal change (PSC) relative to the resting baseline 348 was calculated separately within every ROI for each condition. Mean PSC was computed for each 349

Context-specificity of grasp representations 15individual and condition by taking the ratio of the COPEs (experimental condition > rest) and mean 350 voxel-intensity, and scaling by the peak height of a regressor formed by convolving an isolated 351 three second event with the double gamma hemodynamic response function (Mumford 2007). 352 Subject conditional means were then submitted to separate repeated measures ANOVAs for each 353 ROI with hand and condition as within-subjects factors. A Bonferroni-corrected threshold of p < 354 0.05 was used in significance tests. 355 Results & Discussion 356 Behavioral Data on Prospective Grip Selection. Similar to results of the overt grip selection task in 357 Experiment 1, no difference was detected between hands (F<1.0), and data was therefore pooled 358 across this variable (Figure 2b). As expected if prospectively selected grip preferences are context 359 sensitive, there was again a significant main effect of target (medial, lateral, control), F(1,14) = 360 43.687, MSe = 695.945, p < 0.001. As in Experiment 1, participants showed a strong preference 361 for overhand grips when the lateral end of the handle was cued (M = 82.3%, SD = 10.54) that did 362 not differ from that of the control condition (M = 84.3%, SD =11.32), t(14) = 1.409, p = .181. 363 Importantly, in the medial cue condition (M = 28.08%, SD = 26.60) they again displayed a 364 significantly lower preference for overhand grips compared to either the lateral cue condition [t(14) 365 = 6.65, p < .001] or the NO ROTATION control [t(14) = 6.85, p < .001]. This suggests that despite 366 the absence of overt movements and associated sensory feedback, prospectively chosen grips 367 involved accurately anticipating the demands associated with the intended handle rotations.. 368 Direct comparisons between the overt (Experiment 1) and prospective (Experiment 2) grip selection 369 tasks failed to detect a significant difference in grip preferences between tasks, F < 1.0 (cf. Figure 370 2, panel A & B). Likewise, the variable Task did not interact with the hand involved (F < 1.0). 371 Post-hoc, paired comparisons also failed to reveal any significant differences between specific 372 conditions of the PGS and OGS tasks, p > .20 in all cases. This suggests that despite 373 the absence of overt movements and associated sensory feedback, prospectively chosen grips 374 involved accurately anticipating the demands associated with the intended handle rotations. As 375

Context-specificity of grasp representations 16with OGS, grips were chosen that would allow participants to complete the required actions 376 in a comfortable posture, i.e., they demonstrated the end-state comfort effect (Rosenbaum and 377 Jorgensen 1992; Rosenbaum et al. 1992). Results of fMRI testing provide insights into the neural 378 mechanisms involved in this prospective planning. 379 Neural Activity Associated with Prospective Grip Selection. In order to identify brain regions 380 whose activity was modulated by the PGS task, each of the 4 experimental conditions (2 (hand: 381 left, right) x 2 (goal: no rotation, rotation)) were separately contrasted against resting baseline. 382 Consistent with earlier results on prospective grip selection (Jacobs et al. 2010; Johnson et al. 2002) 383 very similar patterns of activity were found across all conditions (Figure 3). Areas showing 384 increased activity included: regions within and along the intraparietal sulcus (including the aIPS 385 and rIPL, and caudal IPS (cIPS)), dorsal premotor cortex (dPMC), superior parietal lobule (SPL), 386 pre-supplementary motor area (pre-SMA) and the lateral cerebellum. Likewise, activity in posterior 387 middle temporal gyrus (pMTG) and ventral premotor cortex (vPMC) only reached significance in 388 the left hemisphere (Jacobs et al. 2010). 389
------------------------------- 390 Insert Figure 3 about here 391 ------------------------------- 392
Next, a 2 (hand: left, right) x 2 (goal: no rotation, rotation) repeated measures ANOVA was 393 performed across all voxels that showed significantly increased activity relative to resting baseline 394 in at least one of the four experimental conditions. No areas showed a significant main effect of the 395 hand involved. However, numerous areas did manifest a main effect of goal, and planned contrasts 396 revealed that this resulted from greater increases in activity for the ROTATION vs. NO 397 ROTATION condition. As predicted on the basis of single unit recordings (Baumann et al. 2009; 398 Fogassi et al. 2005; Fogassi and Luppino 2005), this difference was detected in both aIPS and the 399 rIPL (Figure 4). These effects were bilateral, but more extensive in the left cerebral hemisphere. 400 In addition, a number of other regions involved in grasp representation also showed this goal effect, 401

Context-specificity of grasp representations 17including: left vPMC, dorsal premotor cortex (dPMC) and caudal IPS. This later increase extended 402 along the parieto-occipital sulcus (POS) and into the SPL, and might include putative V6A (Culham 403 and Valyear 2006). Similar increases were also observed in left pMTG and in pre-SMA. Possible 404 reasons for the involvement of these regions will be considered in the general discussion section 405 below. 406
------------------------------- 407 Insert Figure 4 about here 408 ------------------------------- 409
Descriptive ROI analysis. Regions of interest (ROIs) were defined post-hoc for those areas 410 previously implicated in sensorimotor control of grasp that showed an advantage for the 411 ROTATION vs. NO-ROTATION condition (bilateral aIPS/rIPL, dPMC, and left vPMC). With one 412 exception (right hand NO ROTATION in right aIPS/rIPL, p > .05) all experimental conditions 413 showed significantly greater activity than resting baseline in each ROI, p < .0001 in all cases (inset 414 panels, Figure 4). Right aIPS (t(14) = 2.97, p = 0.01) and right dPMC (t(14) = 6.53, p = 0.001) 415 both showed significantly greater activity when planning involved the contralateral (left) vs. the 416 ipsilateral (right) hand. (Figure 4). 417 General Discussion 418 As introduced earlier, the ways that we grasp objects are influenced by the movements that we 419 intend to perform once they are in hand, i.e., the ultimate goals of the action. While much is 420 currently known about the neural mechanisms involved in online sensorimotor control of grasping, 421 little is understood about the neural mechanisms responsible for the context-sensitivity of this 422 fundamental behavior. The current work yielded two main insights. We found that even in the 423 complete absence of movement and associated sensory feedback, prospectively selected grips 424 (Experiment 2) were highly consistent with those expressed during a similar task involving overt 425 movements (Experiment 1). Importantly, grip preferences in both circumstances showed clear 426

Context-specificity of grasp representations 18evidence of being influenced similarly by the anticipated demands of a forthcoming handle rotation. 427 In the ROTATION condition, they chose grips that would have allowed them to complete the 428 required action in a comfortable posture in the middle of their range of motion. To our knowledge, 429 this is the first demonstration of an end-state comfort effect in an action selection task that involves 430 only planning and no movement. The presence of this “end state comfort effect” in the PGS task is 431 consistent with the hypothesis that context sensitivity, as manifest in the selection of the 432 macroscopic aspects of grasp (e.g., under- vs. over-hand), can arise in a purely feed forward manner 433 (Frey 2010). Critically, these behavioral results allowed us to pursue the primary objective of this 434 work, to evaluate the presence of context-sensitive grasp representations in the rIPL. 435 As predicted based on evidence from single unit recordings in macaques (Baumann et al. 2009; 436 Fogassi et al. 2005), we detected greater increases in rIPL/aIPS during the ROTATION vs. NO 437 ROTATION condition of the PGS task. This suggests that these regions participate in the 438 formation of context-sensitive representations of grasp in the human brain. We now consider these 439 neurophysiological findings, as well as evidence for context-sensitivity of grasp representations in 440 other brain regions, in greater detail. 441 Context-sensitive grasp representations in the inferior parietal lobule. A considerable body of 442 evidence implicates the inferior parietal lobule of macaques (Gardner et al. 2007) and humans 443 (Castiello and Begliomini 2008) in constructing sensorimotor representations of grasp. Grasp-444 related activity of neurons in macaque AIP (Baumann et al. 2009) and rIPL (Fogassi et al. 2005) 445 further show sensitivity to the context in which these movements are performed. That is, activity 446 within these regions of the IPL is modulated according to the actions that the animal is instructed to 447 perform once the object is in hand. We find that human aIPS and rIPL both show increased activity 448 during prospective grip selection relative to resting baseline. Importantly, these increases were 449 significantly greater in the ROTATION vs. NO ROTATION condition (Figure 4). This suggests 450 that these areas of the human brain participate in context-sensitive representations of grasp. 451

Context-specificity of grasp representations 19An earlier investigation provided evidence that transcranial magnetic stimulation (TMS) delivered 452 soon after movement initiation over aIPS interferes with the ability to update motor programs on 453 the basis of sensory feedback (Tunik et al. 2005). Subsequent results suggest that these effects are 454 related to the execution, rather than the planning, phase of the grasping movement (Rice et al. 455 2006). However, there is also evidence that aIPS codes the goals, rather than movements, of 456 observed grasping actions (Hamilton and Grafton 2006), and a recent investigation of prospective 457 grip selection involving the hands or a recently mastered tool found that aIPS participates in grasp 458 planning even in the complete absence of movements (Jacobs et al. 2010). The present study 459 extends these observations by showing further increases in aIPS (and rIPL) activity when grip 460 selection decisions involve representing the demands of a subsequent object rotation. This finding 461 is consistent with the hypothesis that the aIPS (and we would add the rIPL) participates in 462 representing actions’ intended goals (Tunik et al. 2007). Further, we provide evidence that context 463 sensitivity is not peculiar to aIPS/rIPL, but is also found in a number of other regions known to be 464 involved in sensorimotor control of grasp. 465 Context-sensitive responses in other brain regions. As introduced earlier, in addition to rIPL and 466 AIP, single unit recordings reveal that neurons in PMv, PMd and V6a exhibit grasp-related 467 responses. Very recent findings also indicate context-sensitive response during movement 468 preparation in some grasp-related neurons located in the macaque rostral ventral premotor cortex 469 (area F5) (Bonini et al. 2010; Fluet et al. 2010). Together with an interconnected subdivision of the 470 rIPL (area PFG), F5 may play a key role in the organization of sequential, goal-directed actions 471 (Bonini et al. 2010). Similarly, in spite of the complete absence of movement, we find increased 472 activity in human left vPMC, located along the inferior portion of the precentral sulcus and 473 extending rostrally into both the inferior and middle frontal gyri. This is consistent with earlier 474 work on prospective grip selection with the hands or a recently learned tool (Jacobs et al. 2010), 475 and could be taken as evidence for a putatively homologous circuit for goal representation in the 476 human brain that is strongly left-lateralized (further discussion follows). Likewise, our whole-brain 477

Context-specificity of grasp representations 20coverage identified context-sensitive responses in bilateral dPMC, and cIPS/SPL during prospective 478 grip selection both here and in our previous work (Jacobs et al. 2010; Johnson et al. 2002)(Figure 479 4). This suggests that context-sensitivity during planning is a widespread property of parieto-480 premotor grasp representations. 481 Our grip selection tasks demand that participants choose the most comfortable hand orientation 482 (under- vs. over-hand) for grasping the stimulus object. Cells in V6A (Fattori et al. 2009) and the 483 distal forelimb sector of dorsal premotor cortex (PMd or F2) (Raos et al. 2004) code wrist (hand) 484 orientation during grasping. Increased activity in cIPS (extending into the POS and SPL), and 485 dPMC in all conditions of the present investigation might reflect similar coding within the human 486 brain (Figure 4). Importantly, the fact that these increases occurred in the absence of movements 487 suggests their involvement in feed-forward processing. As suggested below, along with aIPS/rIPL 488 and left vPMC, these areas may be involved in representing the state of the effectors during 489 internally simulated movements. Greater involvement during the ROTATION vs. NO ROTATION 490 condition would seem consistent with this hypothesis. 491 492 It is known that in macaques, pre-SMA (F6) is richly interconnected with prefrontal regions and 493 plays a role in premovement processes including selection and preparation (Luppino et al. 1991; 494 Luppino and Rizzolatti 2000) and may serve similar functions in humans (Picard and Strick 2001; 495 1996). The fact that pre-SMA activity shows evidence of context sensitivity here suggests that its 496 role must also include representing the different demands of the ROTATION and NO ROTATION 497 conditions. One possibility is that pre-SMA plays a role in the internal simulation of movements. 498 Increased pre-SMA activity is commonly found in a wide range of motor-cognitive tasks in human 499 neuroimaging studies that involve dynamically imagining or planning movements (Frey 2006; 500 Gerardin et al. 2000; Johnson et al. 2002). This region could be important for updating parieto-501 premotor representations of the effectors’ states over time. We will return to the issue of state 502 estimation shortly. 503

Context-specificity of grasp representations 21Despite a high degree of similarity between stimulus displays, an unexpected increase in left pMTG 504 was also found for the ROTATION vs. NO ROTATION comparison. This area has been shown to 505 increase activity during tasks that involve identification of familiar tools (Chao et al. 1999; Martin 506 et al. 1996) and shows a particular sensitivity to the motions of tools and utensils (Beauchamp et al. 507 2002). Yet, the present task involved neither familiar tools nor moving stimuli. Because both 508 ROTATION and NO ROTATION conditions involved identical objects, this difference also cannot 509 be attributed to the coding of objects’ structural properties, as has been reported in adjacent lateral 510 occipital complex (Kroliczak et al. 2008). Alternatively, left pMTG might be activated during the 511 internal simulation of grasping movements. As introduced below, prospective grip selection may 512 involve use of feed-forward processes to predict the sensory consequences associated with each 513 grip option. Perhaps increased activity in pMTG reflects the processing of predicted visual sensory 514 feedback that would accompany grasping movements. By this account, the greater response in the 515 ROTATION condition would reflect more complex movements than in the NO ROTATION 516 condition. 517 Areas involved in grip selection but not affected by context. Compared to rest, all experimental 518 conditions were associated with increased activity along the left MFG, and the ROTATION 519 condition for the left hand also involved increases in the right hemisphere (Figure 3). Increases in 520 left rostral MFG were also detected in all conditions of an earlier study of prospective grip selection 521 (Jacobs et al. 2010). In contrast to all other regions, however, MFG did not show evidence of 522 context sensitivity, i.e. responses did not differ significantly between the ROTATION vs. NO 523 ROTATION conditions. This suggests that MFG is involved in a processing component that is 524 common to all grip selection conditions, yet unaffected by the additional demands of planning for 525 handle manipulation. Based on previous findings (Schluter et al. 2001), a likely candidate is the 526 choice between the two grip options after the costs associated with each option have been 527 computed. 528

Context-specificity of grasp representations 22The lateral cerebellum also showed bilateral increases in activity for all conditions here, and in our 529 previous investigations of prospective grip selection (Jacobs et al. 2010; Johnson et al. 2002). 530 However, responses were unaffected by the goal manipulation (Figure 4). There is a considerable 531 body of evidence suggesting that the cerebellum may support internal models that predict the 532 sensory consequences of movements based on a copy the motor command (see reviews by 533 (Shadmehr and Krakauer 2008; Wolpert and Flanagan 2001). These predictions are viewed as 534 extending over relatively brief periods of time, slightly in advance of actual sensory feedback. 535 Involvement of the cerebellum here and in other in prospective planning tasks (Jacobs et al. 2010; 536 Johnson et al. 2002), raises the possibility of this structure being involved in forecasting 537 movements’ longer range sensory consequences for use in action selection (Frey 2010). The fact 538 that cerebellar activity does not increase further when planning involves handle rotation, however, 539 suggests that other areas may be involved in representing the sensory consequences of these 540 predictions (Figure 4). More precisely, it may be that the parietal and premotor regions that do 541 show context sensitivity are involved in maintaining and updating representations of how the state 542 of the limbs would be impacted by these predictions arising from internal models supported in the 543 cerebellum. 544 Left cerebral asymmetries in vPMC. Previous work revealed a strong left cerebral asymmetry in 545 vPMC during prospective grip selection regardless of the limb (left or right) or effector (hand or 546 novel tool) on which these judgments were based (Jacobs et al. 2010). We replicated this result, 547 and also showed that grasp-related responses within left vPMC are sensitive to the task context 548 (Figure 4). The significance of this cerebral asymmetry in prospective grasp planning is unclear. 549 Together, these findings point to the fact that the laterality within the sensorimotor system extends 550 to motor-cognitive planning functions (Willems et al. 2009). This is consistent with previous 551 findings on the specialized role for the left hemisphere in motor selection and attention processes in 552 healthy right-handed participants (Haaland and Harrington 1996; Rushworth et al. 1997; Rushworth 553 et al. 1998; Schluter et al. 2001) and in stroke patients (Laimgruber et al. 2005). 554

Context-specificity of grasp representations 23Hand-Independent Responses. It is important to stress that although participants’ behavioral 555 responses show strong evidence of taking into account the unique constraints of the two hands (i.e., 556 hand-dependence), our analyses focusing on areas involved in prospective grip selection failed to 557 detect any regions showing hand-dependent neural activity. The only indications of effector-558 specificity came from post-hoc ROI analyses showing an advantage for the contralateral hand in 559 right aIPS and right dPMC. The failure to detect evidence of effector-specificity in statistical 560 parametric maps is consistent with earlier results (Jacobs et al. 2010). It is tempting to interpret this 561 as evidence for limb-independent levels of motor representation. However, as stated earlier, our 562 behavioral data clearly show that prospective grip selection decisions were based on internal 563 representations that took into account the biomechanical constraints specific to each upper limb. 564 Taken together with the behavioral findings, these results thus demonstrate that common patterns of 565 neural activity across different effectors are a necessary, but not sufficient, source of evidence for 566 effector-independent levels of action representation. It is known that both effector-specific and 567 effector-nonspecific responses can be found in both parietal (Hyvarinen 1982) and premotor 568 (Rizzolatti et al. 1988) regions. This intermixing may lead to indistinguishable fMRI responses. 569 Limitations of the current work. One limitation of the current project is that during fMRI, 570 participants were required to plan, but not to execute, grasping actions. Whether activity would be 571 the same during the premovement planning phase of an actual grasping end-state task remains to be 572 shown. However, there is evidence indicating that in similar grip selection tasks, macroscopic 573 aspects of grasp (e.g., hand orientation) are chosen prior to movement onset (Stelmach et al. 1994). 574 The prospective task might involve processes, such as visual or kinesthetic/motor imagery, that may 575 not be engaged when planning forthcoming movements (Walsh and Rosenbaum 2009). While we 576 cannot rule out these possibilities, we did not detect any increases in visual areas of the brain that 577 were greater in the ROTATION vs. NO ROTATION condition, as would be expected if these 578 conditions differed in their demands on visualization. Engagement of the pre-SMA is consistent 579 with involvement of kinesthetic/motor imagery (Gerardin et al. 2000; Stephan et al. 1995). It is 580

Context-specificity of grasp representations 24also possible that the ROTATION condition was more demanding of spatial attention than the NO 581 ROTATION Control. Attentional demands might account for some of the increased activity in the 582 posterior parietal and even frontal cortex during the ROTATION condition (Shulman et al. 2002). 583 However, attentional processing alone cannot account for the effector- and goal-specific behaviors 584 exhibited in this study. Finally, it is tempting to interpret these results in terms of evidence for 585 functional homologies between monkey rIPS/AIP and human rIPL/aIPS. However, care must be 586 exercised when drawing such conclusions, especially in light of evidence for differences between 587 species in some IPS functions (Orban et al. 2004). 588 In conclusion, we present evidence that the human rIPL/aIPS is involved in forming context-589 sensitive grasp representations. Despite the complete absence of overt movements, this region 590 shows increased activity when selecting how best to grasp a handle for subsequent rotation. These 591 findings complement earlier observations of context-sensitive units in the macaque rIPL and AIP. 592 Further, we find evidence for context-sensitivity in a number of other regions that have previously 593 been implicated in sensorimotor control of grasping and/or action planning, including cIPS, dPMC, 594 left vPMC, and pre-SMA. Additional work is needed to determine the precise relationship between 595 these regions and context-sensitive behavior. 596 597
598

Context-specificity of grasp representations 25Acknowledgments 599 Experiment 1 fulfilled part of the requirements of an MA thesis submitted to the University of 600 Oregon by Evan Lohele-Conger. This work has been supported by grants to S.H.F. from the 601 National Institute of Neurological Disorders and Stroke (NIH/NINDS; # NS053962) and from the 602 Army Research Laboratory (ARO/ARL; # 49581-LS). S.J. held a grant from the Fyssen Foundation 603 during a portion of this work. We thank reviewers of earlier versions of this manuscript for their 604 constructive feedback. M.M. and S.H.F designed the experiment. M.M. and S.J. analyzed the data. 605 S.H.F. and M.M. wrote the manuscript. 606 607
608

Context-specificity of grasp representations 26Literature Citations 609 Baumann MA, Fluet MC, and Scherberger H. Context-specific grasp movement representation 610 in the macaque anterior intraparietal area. J Neurosci 29: 6436-6448, 2009. 611 Beauchamp MS, Lee KE, Haxby JV, and Martin A. Parallel visual motion processing streams 612 for manipulable objects and human movements. Neuron 34: 149-159, 2002. 613 Beckmann CF, Jenkinson M, and Smith SM. General multilevel linear modeling for group 614 analysis in FMRI. Neuroimage 20: 1052-1063, 2003. 615 Binkofski F, Buccino G, Posse S, Seitz RJ, Rizzolatti G, and Freund HJ. A fronto-parietal 616 circuit for object manipulation in man: evidence from an fMRI-study. European Journal of 617 Neuroscience 11: 3276– 3286, 1999. 618 Binkofski F, Dohle C, Posse S, Stephan KM, Hefter H, Seitz RJ, and Freund HJ. Human 619 anterior intraparietal area subserves prehension: a combined lesion and functional MRI activation 620 study. Neurology 50: 1253-1259., 1998. 621 Bonini L, Rozzi S, Serventi FU, Simone L, Ferrari PF, and Fogassi L. Ventral premotor and 622 inferior parietal cortices make distinct contribution to action organization and intention 623 understanding. Cereb Cortex 20: 1372-1385, 2010. 624 Castiello U, and Begliomini C. The cortical control of visually guided grasping. Neuroscientist 14: 625 157-170, 2008. 626 Chao LL, Haxby JV, and Martin A. Attribute-based neural substrates in temporal cortex for 627 perceiving and knowing about objects. Nat Neurosci 2: 913-919, 1999. 628 Culham JC, Danckert SL, DeSouza JF, Gati JS, Menon RS, and Goodale MA. Visually guided 629 grasping produces fMRI activation in dorsal but not ventral stream brain areas. Exp Brain Res 153: 630 180-189, 2003. 631 Culham JC, and Valyear KF. Human parietal cortex in action. Curr Opin Neurobiol 16: 205-212, 632 2006. 633

Context-specificity of grasp representations 27Duvernoy HM. The human brain: surface, blood supply, and three-dimensional sectional 634 anatomy. New York: Springer Wien, 1991. 635 Ehrsson HH, Fagergren A, Jonsson T, Westling G, Johansson RS, and Forssberg H. Cortical 636 activity in precision- versus power-grip tasks: an fMRI study. J Neurophysiol 83: 528-536, 2000. 637 Ehrsson HH, Fagergren E, and Forssberg H. Differential fronto-parietal activation depending on 638 force used in a precision grip task: an fMRI study. Journal of Neurophysiology 85: 2613-2623, 639 2001. 640 Fattori P, Breveglieri R, Marzocchi N, Filippini D, Bosco A, and Galletti C. Hand orientation 641 during reach-to-grasp movements modulates neuronal activity in the medial posterior parietal area 642 V6A. J Neurosci 29: 1928-1936, 2009. 643 Fluet MC, Baumann MA, and Scherberger H. Context-specific grasp movement representation 644 in macaque ventral premotor cortex. J Neurosci 30: 15175-15184, 2010. 645 Fogassi L, Ferrari PF, Gesierich B, Rozzi S, Chersi F, and Rizzolatti G. Parietal lobe: from 646 action organization to intention understanding. Science 308: 662-667, 2005. 647 Fogassi L, and Luppino G. Motor functions of the parietal lobe. Curr Opin Neurobiol 15: 626-648 631, 2005. 649 Frey SH. Forecasting the long-range consequences of manual and tool use actions: 650 neurophysiological, behavioral and computational considerations. (pp. 295-313). Motor control: 651 Theories, experiments and applications 652 2010. 653 Frey SH, Vinton D, Norlund R, and Grafton ST. Cortical topography of human anterior 654 intraparietal cortex active during visually guided grasping. Brain Res Cogn Brain Res 23: 397-405, 655 2005. 656 Frey SHG, V.E. Neural bases of learning of a multi-step task through observation. In: Cognitive 657 Neuroscience Society. San Francisco: 2006. 658

Context-specificity of grasp representations 28Gardner EP, Babu KS, Reitzen SD, Ghosh S, Brown AS, Chen J, Hall AL, Herzlinger MD, 659 Kohlenstein JB, and Ro JY. Neurophysiology of prehension. I. Posterior parietal cortex and 660 object-oriented hand behaviors. J Neurophysiol 97: 387-406, 2007. 661 Gerardin E, Sirigu A, Lehéricy S, Poline JB, Gaymard B, Marsault C, Agid Y, and Le Bihan 662 D. Partially overlapping neural networks for real and imagined hand movements. Cerebral cortex 663 (New York, NY : 1991) 10: 1093-1104, 2000. 664 Grafton ST, Fagg AH, Woods RP, and Arbib MA. Functional anatomy of pointing and grasping 665 in humans. Cereb Cortex 6: 226-237., 1996. 666 Haaland KY, and Harrington DL. Hemispheric asymmetry of movement. Curr Opin Neurobiol 667 6: 796-800., 1996. 668 Hamilton AF, and Grafton ST. Goal representation in human anterior intraparietal sulcus. J 669 Neurosci 26: 1133-1137, 2006. 670 Hyvarinen J. Posterior parietal lobe of the primate brain. Physiol Rev 62: 1060-1129, 1982. 671 Jacobs S, Danielmeier C, and Frey SH. Human anterior intraparietal and ventral premotor 672 cortices support representations of grasping with the hand or a novel tool. J Cogn Neurosci 22: 673 2594-2608, 2010. 674 Jenkinson M. Fast, automated, N-dimensional phase-unwrapping algorithm. Magn Reson Med 49: 675 193-197, 2003. 676 Jenkinson M. Improving the registration of B0-disorted EPI images using calculatedcost function 677 weights. In: Tenth Int Conf on Functional Maping of the Human Brain2004. 678 Jenkinson M, Bannister P, Brady M, and Smith S. Improved optimization for the robust and 679 accurate linear registration and motion correction of brain images. Neuroimage 17: 825-841, 2002. 680 Jenkinson M, and Smith S. A global optimisation method for robust affine registration of brain 681 images. Med Image Anal 5: 143-156, 2001. 682 Johnson SH. Thinking ahead: the case for motor imagery in prospective judgements of prehension. 683 Cognition 74: 33-70, 2000. 684

Context-specificity of grasp representations 29Johnson SH, Rotte M, Grafton ST, Hinrichs H, Gazzaniga MS, and Heinze HJ. Selective 685 activation of a parietofrontal circuit during implicitly imagined prehension. Neuroimage 17: 1693-686 1704, 2002. 687 Johnson-Frey SH, McCarty, M., & Keen, R. Reaching beyond spatial perception: effects of 688 intended future actions on visually guided prehension. Visual Cognition 11: 371-399, 2004. 689 Kroliczak G, McAdam TD, Quinlan DJ, and Culham JC. The human dorsal stream adapts to 690 real actions and 3D shape processing: a functional magnetic resonance imaging study. J 691 Neurophysiol 100: 2627-2639, 2008. 692 Laimgruber K, Goldenberg G, and Hermsdorfer J. Manual and hemispheric asymmetries in the 693 execution of actual and pantomimed prehension. Neuropsychologia 43: 682-692, 2005. 694 Luppino G, Matelli M, Camarda RM, Gallese V, and Rizzolatti G. Multiple representations of 695 body movements in mesial area 6 and the adjacent cingulate cortex: an intracortical 696 microstimulation study in the macaque monkey. J Comp Neurol 311: 463-482., 1991. 697 Luppino G, and Rizzolatti G. The Organization of the Frontal Motor Cortex. News Physiol Sci 15: 698 219-224., 2000. 699 Martin A, Wiggs CL, Ungerleider LG, and Haxby JV. Neural correlates of category-specific 700 knowledge. Nature 379: 649-652., 1996. 701 Miezin FM, Maccotta L, Ollinger JM, Petersen SE, and Buckner RL. Characterizing the 702 hemodynamic response: effects of presentation rate, sampling procedure, and the possibility of 703 ordering brain activity based on relative timing. Neuroimage 11: 735-759, 2000. 704 Mumford JA. A guide to calculating percent change with Featquery. Unpublihsed technical report. 705 http://mumford.bol.ucla.edu/perchange_guide.pdf. 706 Orban GA, Van Essen D, and Vanduffel W. Comparative mapping of higher visual areas in 707 monkeys and humans. Trends Cogn Sci 8: 315-324, 2004. 708 Picard N, and Strick PL. Imaging the premotor areas. Curr Opin Neurobiol 11: 663-672, 2001. 709

Context-specificity of grasp representations 30Picard N, and Strick PL. Motor areas of the medial wall: a review of their location and functional 710 activation. Cereb Cortex 6: 342-353, 1996. 711 Raos V, Umilta MA, Gallese V, and Fogassi L. Functional properties of grasping-related neurons 712 in the dorsal premotor area F2 of the macaque monkey. J Neurophysiol 92: 1990-2002, 2004. 713 Rice NJ, Tunik E, Cross ES, and Grafton ST. On-line grasp control is mediated by the 714 contralateral hemisphere. Brain Res 1175: 76-84, 2007. 715 Rice NJ, Tunik E, and Grafton ST. The anterior intraparietal sulcus mediates grasp execution, 716 independent of requirement to update: new insights from transcranial magnetic stimulation. J 717 Neurosci 26: 8176-8182, 2006. 718 Rizzolatti G, Camarda R, Fogassi L, Gentilucci M, Luppino G, and Matelli M. Functional 719 organization of inferior area 6 in the macaque monkey. II. Area F5 and the control of distal 720 movements. Exp Brain Res 71: 491-507, 1988. 721 Rizzolatti G, and Luppino G. The cortical motor system. Neuron 31: 889-901., 2001. 722 Rosenbaum DA, and Jorgensen MJ. Planning macroscopic aspects of manual control. Human 723 Movement Science 11: 61-69, 1992. 724 Rosenbaum DA, Vaughan J, Barnes HJ, and Jorgensen MJ. Time course of movement 725 planning: selection of handgrips for object manipulation. J Exp Psychol Learn Mem Cogn 18: 1058-726 1073, 1992. 727 Rushworth MF, Nixon PD, Renowden S, Wade DT, and Passingham RE. The left parietal 728 cortex and motor attention. Neuropsychologia 35: 1261-1273., 1997. 729 Rushworth MF, Nixon PD, Wade DT, Renowden S, and Passingham RE. The left hemisphere 730 and the selection of learned actions. Neuropsychologia 36: 11-24., 1998. 731 Schluter ND, Krams M, Rushworth MF, and Passingham RE. Cerebral dominance for action in 732 the human brain: the selection of actions. Neuropsychologia 39: 105-113, 2001. 733 Shadmehr R, and Krakauer J. A computational neuroanatomy for motor control. Experimental 734 Brain Research 185: 359-381, 2008. 735

Context-specificity of grasp representations 31Shulman GL, d'Avossa G, Tansy AP, and Corbetta M. Two attentional processes in the parietal 736 lobe. Cereb Cortex 12: 1124-1131, 2002. 737 Smith SM. Fast robust automated brain extraction. Hum Brain Mapp 17: 143-155, 2002. 738 Smith SM, Jenkinson M, Woolrich MW, Beckmann CF, Behrens TE, Johansen-Berg H, 739 Bannister PR, De Luca M, Drobnjak I, Flitney DE, Niazy RK, Saunders J, Vickers J, Zhang 740 Y, De Stefano N, Brady JM, and Matthews PM. Advances in functional and structural MR image 741 analysis and implementation as FSL. Neuroimage 23 Suppl 1: S208-219, 2004. 742 Stelmach GE, Castiello U, and Jeannerod M. Orienting the finger opposition space during 743 prehension movements. J Mot Behav 26: 178-186, 1994. 744 Stephan KM, Fink GR, Passingham RE, Silbersweig D, Ceballos-Baumann AO, Frith CD, 745 and Frackowiak RS. Functional anatomy of the mental representation of upper extremity 746 movements in healthy subjects. J Neurophysiol 73: 373-386., 1995. 747 Tunik E, Frey SH, and Grafton ST. Virtual lesions of the anterior intraparietal area disrupt goal-748 dependent on-line adjustments of grasp. Nat Neurosci 8: 505-511, 2005. 749 Tunik E, Rice NJ, Hamilton A, and Grafton ST. Beyond grasping: representation of action in 750 human anterior intraparietal sulcus. Neuroimage 36 Suppl 2: T77-86, 2007. 751 Van Essen DC. A Population-Average, Landmark- and Surface-based (PALS) atlas of human 752 cerebral cortex. Neuroimage 28: 635-662, 2005. 753 Van Essen DC, Drury HA, Dickson J, Harwell J, Hanlon D, and Anderson CH. An integrated 754 software suite for surface-based analyses of cerebral cortex. J Am Med Inform Assoc 8: 443-459, 755 2001. 756 Walsh M, and Rosenbaum D. Deciding how to act is not achieved by watching mental movies. 757 Journal of Experimental Psychology: Human Perception and Performance 35: 1481-1489, 2009. 758 Willems RM, Toni I, Hagoort P, and Casasanto D. Body-specific motor imagery of hand 759 actions: neural evidence from right- and left-handers. Front Hum Neurosci 3: 39, 2009. 760 Wolpert DM, and Flanagan JR. Motor prediction. Curr Biol 11: R729-732., 2001. 761

Context-specificity of grasp representations 32Woolrich MW, Behrens TEJ, Beckmann CF, Jenkinson M, and Smith SM. Multi-level linear 762 modelling for FMRI group analysis using Bayesian inference. NeuroImage 21: 1732-1747, 2004. 763 Worsley KJ. Statistical analysis of activation images. In: Functional MRI: An Introduction to 764 Methods, edited by Jezzard P, Matthews PM, and Smith SM. USA: Oxford University Press, 2001, 765 p. 251-270. 766 767 768 769
Context-specificity of grasp representations 33 770 771 772
773 774 Table 1. MNI stereotaxic coordinates of the peaks of BOLD signal change during the 4 775 experimental conditions. 776 777 778 779 780

Context-specificity of grasp representations 34Figure Legends 781 Figure 1. Schematic illustrations of the stimuli and paradigms used in Experiments 1 and 2. In 782 Experiment 1 (overt grip selection), actual handles were used, while in Experiment 2 (prospective 783 grip selection) stimuli were computer images with photo-realistic shading to provide the illusion of 784 three-dimensionality. (A) An example of grip selection stimuli from three different trials. 785 Assuming that the right hand was cued, moving from left to right in the figure are illustrations of a: 786 1) ROTATION trial with the lateral end of the handle cued, 2) NO ROTATION trial, and 3) 787 ROTATION trial with medial end of the handle cued. (B) Illustration of a possible event sequence 788 during a prospective grip selection trial in Experiment 2. Participants were instructed to decide 789 whether it would be more comfortable to place the thumb side of their cued hand (as indicated by 790 the arrow) on the pink or tan end of the handle if grasping it in a power grip. If the target was a 791 circle or square, they were to base their grip selection on consideration of the need to rotate the 792 handle to align the corresponding end of the handle (ROTATION condition). If, as illustrated here, 793 the target was a triangle, then they were instructed to decide on which end on the handle they would 794 place their thumb if simply grasping the handle without the intention to rotation (NO ROTATION 795 condition). (C) Biomechanical constraints governing pronation-supination of the left and right 796 hands are mirror symmetrical (i.e., 1800 out of phase). In order to directly compare grip selection 797 preferences across left and right hands, ROTATION trials were therefore organized according to 798 whether the end of the handle cued by the target shape was pointing toward (medial) or away from 799 (lateral) the participant’s midline. These are illustrated for the both overhand (upper half) and 800 underhand (lower half) possibilities. 801 Figure 2. Grip preference data from Experiment 1 (overt grip selection; panel A) and Experiment 802 2 (prospective grip selection; panel B). Direct comparisons of grip preferences between overt and 803 prospective tasks failed to detect any differences. As detailed in the text, participants in both 804 experiments preferred overhand grips in the NO ROTATION and when the lateral end of the hand 805

Context-specificity of grasp representations 35was cued in the ROTATION condition. By contrast, when the medial end of the handle was cued 806 in the ROTATION condition, participants showed a preference for the underhand grip in both tasks. 807 Figure 3. Areas showing significant increases in activity relative to resting baseline for the four 808 experimental conditions. Group average data (z ≥ 2.3, p < 0.05, cluster-based correction for 809 multiple comparisons) are overlaid onto the partially inflated PALs atlas in Caret 5.5 using the 810 multi-fiducial mapping procedure. Areas showing significant activation after multi-fiducial 811 mapping are represented in warm hues. Pale green areas were significantly activated in the group 812 FSL analysis, but did not survive the multi-fiducial correction for intra-subject anatomical 813 variability. Note the similarity of increases across all four conditions, and left cerebral asymmetries 814 in rostral posterior parietal and premotor regions. 815 Figure 4. Results of the 2 (Hand: left hand, right hand) x 2 (Goal: Rotation, No Rotation) 816 repeated measures ANOVA. This more targeted analysis included voxels that were activated 817 significantly relative to resting baseline in at least one of the four conditions (see Figure 3). Table 818 1 provides details on the coordinates used to center these 5mm radius spherical ROIs. No areas 819 showed a significant effect of Hand. However, premotor and parietal regions, including rIPL/aIPS 820 did show significantly greater increases when grip selection involved considering the demands 821 associated with handle ROTATION as compared to NO ROTATION. These effects were bilateral 822 within the rostral IPL and in dorsal premotor cortex, but were restricted to the left cerebral 823 hemisphere in the posterior middle temporal and ventral premotor cortices (see text for details). No 824 regions showed the opposite effect. The lower row illustrates the same data group results displayed 825 on neurologically-oriented, axial slices of the group-average, T1-weighted structural scan. Inset 826 panels illustrate results of descriptive ROI analyses (see Method). These post-hoc tests provide 827 some evidence for responses that are affected by the hand involved in planning. Right aIPS and 828 right dPMC both showed significantly greater activity when planning involved the contralateral 829 (left) vs. the ipsilateral (right) hand. With the exception of left dPMC, all ROIs showed a 830

Context-specificity of grasp representations 36significant interaction between hand and goal, reflecting greater increases in activity when choosing 831 a grip for the contralateral hand with the goal of handle rotation (see text for details). 832 833