Embed Size (px)
Citation preview

ESCUELA SUPERIOR Y TÉCNICA DE INGENIERÍA AGRARIA
INGENIERÍA DE BIOSISTEMAS
Enhancing Psyttalia concolor (Szépligeti)
(Hymenoptera: Braconidae) in olive
groves to increase biological control of
the olive fruit fly
_____________________________________
Presented by:
Lara Alina de Almeida Pinheiro
Directors:
Sónia Alexandra Paiva dos Santos
María del Pilar Medina Vélez
José Alberto Cardoso Pereira
_________________________________
León, 2019

ESCUELA SUPERIOR Y TÉCNICA DE INGENIERÍA AGRARIA
INGENIERÍA DE BIOSISTEMAS
Incrementar la presencia de Psyttalia
concolor (Szépligeti) (Hymenoptera:
Braconidae) en los olivares para mejorar
el control biológico de la mosca del olivo
_____________________________________
PhD candidate:
Lara Alina de Almeida Pinheiro
Supervisors:
Sónia Alexandra Paiva dos Santos
María del Pilar Medina Vélez
José Alberto Cardoso Pereira
_________________________________
León, 2019

I dedicate my PhD thesis to the people who
supported me daily and believed in my work.

The work developed within this PhD thesis is an integral part of the
project, “Olive crop protection in sustainable production under global
climatic changes: linking ecological infrastructures to ecosystem
functions” (EXCL/AGR-PRO/0591/2012), financed by FEDER funds
through the Competitiveness Factors Operational Program - COMPETE
and by National Funds through the Portuguese Foundation of Science and
Technology (FCT). L. Pinheiro thanks FCT, POPH-QREN and FSE for
SFRH/BD/103998/2014 grant.

The studies presented in this thesis were carried out in the
Agrobiotechnology group of the Higher Agricultural School of the
Polytechnic Institute of Bragança (Portugal) in collaboration with
Mountain Research Center and the laboratory of Crop Protection Unit,
ETSIA - Technical University of Madrid (Spain).

vi
AGRADECIMIENTOS
Agradezco mis alegrías. Agradezco profundamente a cada una de las personas que me
hizo feliz. Agradezco por haber conocido gente increíble a lo largo de esta caminata hacia
la obtención del título de Doctor, pero no solo las grandes ayudas importan. También los
pequeños gestos, las palabras, los abrazos, los consejos, las desilusiones y lo que se ofrece
sin nunca concretarse, fueron decisivos para mantenerme firme en esta etapa desafiante y
muchas veces solitaria que es escribir una tesis doctoral. He tenido días dulces y días
amargos, días de sol y días de tormenta. Hablé mucho, oí poco. Oí mucho, hablé poco.
Escribí mucho, escribí poco. Aprendí, subí, bajé, tropecé, caí, volví a subir, persistí, resistí
y no desistí… y así llegué al final de la tesis. Ahora solo me queda agradecer a todos lo
que formaron parte de esta aventura.
A la profesora Sónia Santos, mi orientadora de la tesis y mi madre en el mundo de la
investigación, agradezco por la oportunidad de hacer investigación a lo largo de estos 9
años. Agradezco por la disponibilidad dispensada en todas las situaciones, por la
transferencia de conocimientos y por sus sugerencias, que fueron muy valiosas para la
realización de este y de los otros proyectos que hemos tenido. Muchas gracias por todo.
A mi co-orientadora Pilar Medina, agradezco el modo en el que me apoyó en esta
investigación: una orientación científica estimulante, de confianza y de crecimiento. La
disponibilidad que siempre ha manifestado y la empatía con la que ha recibido mis ideas
fue uno de los estímulos que me ha permitido vencer las inseguridades de este proceso.
Muchas gracias.
Al profesor José Alberto, agradezco la transmisión de sus sabios conocimientos en
olivicultura, así como la búsqueda de financiación (por proyectos) que me ha permitido
disponer del material necesario para la investigación.
Agradezco a mis compañeros del laboratorio de Agrobiotecnología (CIMO, ESA-IPB,
Braganza, Portugal), a los que están en estos momentos y a los que ya pasaron por allá, y
a todo el personal del laboratorio de Protección de Cultivos (ETSIAAB-UPM, Madrid,
España). De éstos, debo destacar algunos por el cariño que me han dedicado y a quienes
siempre recordaré con buenos sentimientos:
Rosalina, Fátima y Cristina, fueron las amigas que estuvieron a mi lado durante esta
fase, por la compañía, la fuerza y el apoyo en ciertos momentos difíciles. Muchas Gracias.

vii
A Andrea (llena de garra, de energía positiva y de voluntad de cambiar el mundo),
Agus (pasión, garra, entrega y determinación son una presencia en su vida), Mar (¡tan
querida, tan dulce y tan talentosa!) y Celeste (llena de positividad y alegría, un verdadero
espíritu libre), quiero agradecer por todo el cariño, amistad y apoyo, así como darles un
aplauso de pie por su forma inspiradora de estar en la vida.
A mi gran amiga Soraia, que a lo largo de más de 20 años, ha sido siempre una amiga
muy leal, que estuvo siempre a mi lado cuando lo necesitaba, una persona en la que
realmente puedo confiar. Por eso, solo puedo agradecerle su sincera amistad y retribuirle
todo el cariño que me tiene. Sabes que eres una persona muy especial para mí, eres mi
hermana del corazón.
A mi Diogo (a quien amo con pasión) ¡Quiero agradecerte por todo! Gracias por
resolver mis dudas de inglés. Gracias por los momentos en que lloré y tú cariñosamente
me hiciste sonreír. Por los momentos en que perdí la paciencia, y tú viniste con palabras
amenas y dulces para calmarme. Por los momentos en que mi corazón estaba en pedazos,
y tú viniste lleno de amor. Por los momentos de alegría. Por los momentos en que, con
mucha esperanza, pensamos juntos en nuestro futuro.
A mi hermano y mis padres (¡las personas más lindas del universo!). A mi hermano,
le agradezco por todos los momentos y por todas las alegrías que vivimos juntos, incluso
por las peleas de la infancia [¡risas!]. Gracias por la presencia, por la palabra, por la
sonrisa, por el apoyo o por el simple recuerdo, que me dieron coraje y determinación para
trazar el camino hacia mis sueños. Te amo hermanito.
A mis Padres, gracias por enseñarme a caminar y así poder seguir mis propios pasos
hacia mis sueños. Gracias por la educación como ser humano, que hizo de mí la persona
que hoy soy. Gracias por estar siempre a mi lado. Gracias por haberos entregado en cuerpo
y alma a mí y a mi hermano, renunciando muchas veces a vuestros sueños, para que
muchas veces pudiéramos realizar los nuestros. Estoy y estaré eternamente agradecida
por todo lo que me habéis dedicado. Estoy muy orgullosa de ser vuestra hija y siento
mucha admiración por los padres que tengo. Gracias por todo. ¡Os quiero mucho!

viii
TABLE OF CONTENTS
Agradecimientos.....…………………………………………………………….. vi
List of Figures (in English)…………………………………………………….. xvi
List of Figures (in Spanish)…………………………………………...………… xviii
List of Tables (in English)………………………………………………………. xx
List of Tables (in Spanish)……………………………………………………… xxi
Abstract (in English)……………………………………………………………. xxii
Abstract (in Spanish)……...……………………………………………………. xxiv
Extended Abstract (in Spanish) ………………………………………………... x
xxvi
Photographs of the species studied in this thesis………………………………. li
Chapter 1: General Introduction………………………………………………. 52
1. Introduction……………………………………………………………… 53
1.1.Dietary requirements………………………………………………. 54
1.1.1. Carbohydrates………………………………………………… 54
1.1.2. Amino acid …………………………………………………… 55
2. Impacts of climate change on crop-pest-parasitoid interaction:
implications on biological control………………………...........................
55
3. References……………………………………………………………….. 58
Chapter 2: Objectives…………………………………………………………... 61
Chapter 3: Literature review…………………………………………………... 64
1. Abstract………………………………………………………………….. 65
2. Introduction……………………………………………………………… 65
3. Distribution of Psyttalia concolor……………………………………….. 67
4. Biology and life-history of Psyttalia concolor…………………………… 68
4.1.The life cycle of Psyttalia concolor…….…………………………… 68
4.2.Longevity of Psyttalia concolor adults………...……………………. 69

ix
4.3.Courtship and mating behavior………...……………………………. 70
4.4.Host-stage preference………...……………………………………... 70
4.5.Visual, olfactory and mechanical stimuli.……………………..…….. 72
4.6.Life-time fecundity…………………………….…………………..... 73
5. Ecology and habitat relationship…………………………………………. 74
5.1. Psyttalia concolor as a biological control agent of Bactrocera olea… 74
5.2. Thermal compatibility between Psyttalia concolor and Bactrocera
oleae………………………………………………………………….
75
5.3. Effect of pesticides on Psyttalia concolor…………………………… 76
6. Conclusion……………………………………………………………….. 77
7. References……………………………………………………………….. 77
Chapter 4: Sugars and honeydew increase longevity and parasitism capacity
of Psyttalia concolor (Hymenoptera: Braconidae)……………………………..
85
1. Abstract…………………………………………………………………... 86
2. Introduction……………………………………………………………… 86
3. Material and methods…………………………………………………….. 88
3.1. Insect rearing……………………………………………………….. 88
3.2. Food sources………………………………………………………... 89
3.3. Longevity and parasitism capacity of Psyttalia concolor fed on
different sugars………………………………………………………
89
3.3.1. Longevity……………………………………………………... 89
3.3.2. Parasitism capacity…………………………………….……… 90
3.4. Longevity of Psyttalia concolor fed on honeydew…………………. 91
3.5. Data analysis………………………………………………………... 91
3.5.1. Longevity……………………………………………………... 91
3.5.2. Parasitism capacity……………………………………………. 92
4. Results………………………………………………………………….... 92
4.1. Longevity and parasitism capacity of Psyttalia concolor fed on
different sugars………………………………………………………
92
4.1.1. Longevity……………………………………………………... 92
4.1.2. Parasitism capacity……………………………………………. 93
4.2. Longevity of Psyttalia concolor fed on honeydew…………………. 95

x
5. Discussion………………………………………………………………... 97
6. References……………………………………………………………….. 100
Chapter 5: Amino acids diet influence survival and reproductive capacity of
Psyttalia concolor (Hymenoptera: Braconidae), a parasitoid of tephritid
flies?.......................................................................................................................
104
1. Abstract…………………………………………………………….......... 105
2. Introduction……………………………………………………………… 105
3. Material and methods……………………………………………………. 107
3.1. Insect rearing…………………………………………………….…. 107
3.2. Food sources………………………………………………………... 108
3.3. Longevity…………………………………………………………... 108
3.4. Reproduction……………………………………………………….. 109
3.5. Data analysis………………………………………………………... 110
4. Results…………………………………………………………………… 110
4.1. Survival…………………………………………………………….. 110
4.2. Reproduction……………………………………………………….. 115
5. Discussion………………………………………………………….......... 116
6. References………………………………………………………….......... 119
7. Supplemental material…………………………………………………… 123
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor
(Szépligeti): a review…………………………………………………………….
124
1. Abstract………………………………………………………………….. 125
2. Introduction……………………………………………………………… 126
2.1. Effects of pesticides on natural enemies: methods for testing
compatibility..………………………………………………………..
127
2.2. Impact of pesticides on Bactrocera oleae - Psyttalia concolor
interaction……………………………………………………………
129
3. Impact of pesticides on parasitoid Psyttalia concolor: a review…………. 132
4. Laboratory case study: effects of different pesticides on mortality and
reproduction of Psyttalia concolor females……………………………….
137
4.1. Material and methods……………………………………………….. 137

xi
4.1.1. Psyttalia concolor and Ceratitis capitata rearing conditions… 137
4.1.2. Pesticides…………………………………………………….. 137
4.1.3. Laboratory bioassays……………………………………….... 139
4.1.4. Contact bioassays…………………………………………….. 139
4.1.5. Ingestion bioassay……………………………………………. 141
4.1.6. Data analysis……………………………………………......... 141
4.2.Results………………………………………………………………. 141
4.2.1. Mortality of adult females……………………………………. 141
4.2.2. Sublethal effects on reproduction: attacked hosts and progeny
size…………………………………………………………..
143
4.3.Discussion…………………………………………………………... 143
4.3.1. Insecticide exposure………………………………………….. 143
4.3.2. Fungicide exposure…………………………………………... 150
4.3.3. Herbicide exposure…………………………………………... 151
4.3.4. Microbial and botanical exposure…………………………..... 152
5. Emergent problem: a risk for Psyttalia concolor survival………………... 154
6. Overview………………………………………………………………… 155
7. References……………………………………………………………….. 156
8. Supplemental material………………………………………………........ 167
Chapter 7: General conclusions………………………………………………... 169
1. General conclusions…………………………………………………….... 170
2. References……………………………………………………………….. 171

xii
TABLA DE CONTENIDOS
Agradecimientos.....…………………………………………………………….. vi
Relación de figuras (en ingles)………………………………………………….. xvi
Relación de figuras (en castellano)………………………...……………………
xviii
Relación de tablas (en ingles)…………………………………………………… xx
Relación de tablas (en castellano)……………………………………………… xxi
Resumen en ingles………………………………………………………………. xxii
Resumen en castellano………………………………………………………….. xxiv
Resumen extendido de la tesis………………………………………………….. xxvi
Fotografías de las especies estudiadas en esta tesis……………………………. li
Capítulo 1: Introducción General…………………………………………..….. 52
1. Introducción……………………………………………………………… 53
1.1.Requerimientos nutricionales……………………………………….. 54
1.1.1. Carbohidratos……………………………………………….… 54
1.1.2. Aminoácidos ……………………………………………….… 55
2. Impacto del cambio climatic en la interación planta-plaga-huésped:
implicaciones en el control biológico…………………………………….
55
3. Referencias………………………………………………………………. 58
Capítulo 2: Objetivos…………………………………………………………... 61
Capítulo 3: Revisión bibliográfica……………………………………………... 64
1. Resumen…………………………………………………………………. 65
2. Introducción……………………………………………………………... 65
3. Distribución de Psyttalia concolor………………………………………. 67
4. Biología e historia de vida de Psyttalia concolor………………………… 68
4.1.Ciclo de Psyttalia concolor…….……………………………………. 68
4.2.Longevidad de adultos de Psyttalia concolor………...……………… 69

xiii
4.3.Cortejo y apareamiento……..………...……………………………… 70
4.4.Estadío del huésped preferido………...……………………………… 70
4.5.Estimulos visuales, olfativos y táctiles.……………………..……….. 72
4.6.Fecundidad…………………………….………………….................. 73
5. Relaciones ecológicas y de hábitat………………………………………. 74
5.1.Psyttalia concolor como agente de control biologico de Bactrocera
oleae…………………………………………………………………. 74
5.2.Compatibilidad térmica entre Psyttalia concolor y Bactrocera oleae..
75
5.3.Efecto de los plaguicidas en Psyttalia concolor……………………… 76
6. Conclusiones…………………………………………………………….. 77
7. Bibliografía……………………………………………………………… 77
Capítulo 4: Los azúcares y la melaza incrementan la longevidad y la
capacidad parasítica de Psyttalia concolor (Hymenoptera:
Braconidae)……………………………………………………………………...
85
1. Resumen…………………………………………………………………. 86
2. Introducción……………………………………………………………… 86
3. Materiales y métodos…………………………………………………….. 88
3.1.Cría de insectos…………………………………………………….. 88
3.2.Fuentes alimenticias……………………………………………….... 89
3.3.Longevidad y capacidad parasítica de Psyttalia concolor alimentada
con diferentes azúcares………………………………………………
89
3.3.1. Longevidad…………………………………………………… 89
3.3.2. Capacidad parasítica…………………………………….……. 90
3.4.Longevidad de Psyttalia concolor alimentada con melazas…………. 91
3.5.Análisis de datos…………………………………………………….. 91
3.5.1. Longevidad…………………………………………………… 91
3.5.2. Capacidad parasítica………………………………………….. 92
4. Resultados………………………………………………………………... 92
4.1.Longevidad y capacidad parasitica de Psyttalia concolor alimentada
con diferentes azúcares………………………………………………
92
4.1.1. Longevidad…………………………………………………… 92
4.1.2. Capacidad parasítica………………………………………….. 93

xiv
4.2.Longevidad de Psyttalia concolor alimentada con melazas…………. 95
5. Discusión……………………………………………………………….... 97
6. Bibliografía………………………………………………………………. 100
Capítulo 5: ¿La dieta de aminoácidos influye en la supervivencia y
capacidad reproductora de Psyttalia concolor (Hymenoptera: Braconidae),
un parasitoide de tefrítidos?.................................................................................
104
1. Resumen……………………………………………………………......... 105
2. Introducción……………………………………………………………... 105
3. Materiales y métodos…………………………………………………….. 107
3.1.Cría de insectos……………………………………………………… 107
3.2.Fuentes alimenticias……………………………………………….... 108
3.3.Longevidad…………………………………………………………. 108
3.4.Reproducción……………………………………………………….. 109
3.5.Análisis de datos…………………………………………………….. 110
4. Resultados……………………………………………………………….. 110
4.1.Supervivencia……………………………………………………….. 110
4.2.Reproducción……………………………………………………….. 115
5. Discusión…………………………………………………………............ 116
6. Bibliografía…………………………………………………………........ 119
7. Material suplementario…………………………………………………... 123
Capítulo 6: Efectos secundarios de los plaguicidas en el parasitoide Psyttalia
concolor (Szépligeti): una revisión…………………………………...................
124
1. Resumen…………………………………………………………………. 125
2. Introducción……………………………………………………………… 126
2.1.Efectos de los plaguicidas en los enemigos naturales: métodos para
ensayar la compatibilidad.……………………………………………
127
2.2.Impacto de los plaguicidas en la interacción entre Bactrocera oleae -
Psyttalia concolor …………………………………………………...
129
3. Impacto de los plaguicidas en Psyttalia concolor: una revisión………...... 132
4. Estudio del caso en laboratorio: efectos de los diferentes plaguicidas en
la mortalidad y reproducción de las hembras de Psyttalia concolor……..
137

xv
4.1.Materiales y métodos……………………………………………….. 137
4.1.1. Condiciones de cría de Psyttalia concolor y Ceratitis capitata. 137
4.1.2. Plaguicidas………………………………………………….... 137
4.1.3. Condiciones de laboratorio…………………………………... 139
4.1.4. Ensayos por contacto………………………………………… 139
4.1.5. Ensayos por ingestión………………………………………... 141
4.1.6. Análisis de datos……………………………………………... 141
4.2.Resultados…………………………………………………………… 141
4.2.1. Mortalidad de hembras……………………………………….. 141
4.2.2. Efectos subletales en la reproducción: huéspedes atacados y
tamaño de la progenie………………………………………..
143
4.3.Discusión…………………………………………………………..... 143
4.3.1. Exposición a los insecticidas…………………………………. 143
4.3.2. Exposición a los fungicidas…………………………………... 150
4.3.3. Exposición a los herbicidas…………………………………... 151
4.3.4. Exposición a los biopesticidas…………………………........... 152
5. Plagas emergentes: un riesgo para la supervivencia de Psyttalia concolor.. 154
6. Resumen…………………………………………………………………. 155
7. Bibliografía………………………………………………………………. 156
8. Material suplementario………………………………………………....... 167
Capítulo 7: Conclusiones generales……………………………………………. 169
1. Conclusiones generales…………………………………………………... 170
2. Bibliografía………………………………………………………………. 171

xvi
LIST OF FIGURES
Figure 3.1. Geographical distribution of Psyttalia concolor: In black are represented
its native countries and in gray are represented the countries where it was
introduced……………………………………………………………………………
68
Figure 3.2. Psyttalia concolor laying one egg on an olive fruit fly larvae.…………... 68
Figure 3.3. Interaction between Psyttalia concolor female and Bactrocera oleae….. 74
Figure 4.1. A: Psyttalia concolor adult; B: Plexiglass cage used in the longevity
tests; C: Detail of feeding glass vials…………………………………………............
89
Figure 4.2. A: Plexiglass cage used in the parasitation tests; B: 3rd instar larvae of
C. capitata were offered to P. concolor; C: Detail of parasitism……………………..
90
Figure 4.3. Survival curves obtained by Kaplan–Meier estimates for Psyttalia
concolor females (A) and males (B) fed on single sugars…………………………….
92
Figure 4.4. Longevity (mean ± SE) (days) for Psyttalia concolor females (A) and
males (B) fed on different solutions of single sugars…………………………………
94
Figure 4.5. Survival curves obtained by Kaplan–Meier estimates for Psyttalia
concolor females (A) and males (B) fed on different honeydews…………………….
95
Figure 4.6. Longevity (mean ± SE) (days) for Psyttalia concolor females (A) and
males (B) fed on different honeydews………………………………………………..
96
Figure 5.1. Detail of plexiglass cage used in the longevity tests: A: Psyttalia
concolor adults; B: feeding glass vials……………………………………………….
108
Figure 5.2. A: Plexiglass cage used in the parasitation tests; B: 3rd instar larvae of
C. capitata offered to P. concolor……………………………………………………
109
Figure 5.3. Survival curves, obtained by Kaplan–Meier estimates, of adult females
(A) and males (B) of Psyttalia concolor fed on different food sources (W - water; D
- Glucose; Phe - Phenylalanine; Pro - Proline; Trp - Tryptophan; Mix – mixture of
D + Phe + Pro + Trp).………………………………………………………………...
111
Figure 5.4. Schoenfeld’s residuals plotted against the different levels of the
categorical variable “food resource” of the Cox PHMs fitted for adults of Psyttalia
concolor (Cox PHM1 for females: A, B, C, D; Cox PHM3 for males: E, F, G, H, I)…
114
Figure 5.5. Schoenfeld’s residuals plotted against the different levels of the
categorical variable “food resource” of the Cox PHM2 fitted for adults of Psyttalia
concolor (A, B, C, D) with the additional explanatory categorical variable “age at
death”………………………………………………………………………………..
115

xvii
Figure S5.1. Scheme of the risk of dying for each tested treatment: A: Females <
55 days; B: Females > 55 days and C: Males………………………………………...
123
Figure 6.1. Sequential scheme of standard method of IOBC for testing effects of
pesticides on natural enemies. * No additional tests needed; *2 Optional routes.
Figure adapted for Dohmen, 1998……………………………………………………
128
Figure 6.2. Biological relationship between olive fruit fly and beneficial insects and
pesticides application………………………………………………………………...
131
Figure 6.3. Contact bioassay: A: Psyttalia concolor female; B: Experimental arena
used to evaluate the mortality with (1) distilled water, (2) diet and (3) treated olive
branch; C: Olive tree (cv. Picual) leaves treated with a pesticide using hand spray;
D and E: Olive branch totally covered with the pesticide to dry……………………..
140
Figure 6.4. A: Parasitisation cages used to evaluate the sublethal effects of
pesticides on Psyttalia concolor reproduction. (1) Female wasp individually to
parasited, (2) 20 larvae (3rd instar) of C. capitata offered to P.
concolor………………………………………………………………………...........
140

xviii
RELACIÓN DE FIGURAS
Figura 3.1. Distribución geográfica de Psyttalia concolor: En negro están
representados los países en los cuales es nativa y en gris se representan los países
donde ha sido introducida……………………………………………………………
68
Figura 3.2. Psyttalia concolor poniendo un huevo en una larva de mosca del olivo… 68
Figura 3.3. Interacción entre hembra de Psyttalia concolor y Bactrocera oleae……. 74
Figura 4.1. A: Adulto de Psyttalia concolor; B: Caja de Plexiglás usada en los
ensayos de longevidad; C: Detalle de los bebederos…………………………………
89
Figura 4.2. A: Caja de Plexiglás utilizada en los ensayos de parasitación; B: El
tercer estadío larvario de C. capitata se ofreció a P. concolor; C: Detalle del
parasitismo…………………………………………………………………………...
90
Figura 4.3. Curvas de supervivencia de Kaplan–Meier para hembras de Psyttalia
concolor (A) y machos (B) alimentados en azúcares individuales...............................
92
Figura 4.4. Longevidad (media ± EE) (días) de hembras de Psyttalia concolor (A)
y machos (B) alimentados en diferentes diluciones de azúcares individuales..............
94
Figura 4.5. Curvas de supervivencia de Kaplan–Meier para hembras de Psyttalia
concolor (A) y machos (B) alimentados en diferentes azúcares……………………...
95
Figura 4.6. Longevidad (media ± EE) (días) de hembras de Psyttalia concolor (A)
y machos (B) alimentados en diferentes melazas…………………………………….
96
Figura 5.1. Detalle de la caja de Plexiglás utilizada en los ensayos de longevidad:
A: Adultos de Psyttalia concolor; B: bebederos……………………………………..
108
Figura 5.2. A: Caja de Plexiglás utilizada en los ensayos de longevidad; B: Tercer
estadío larvario de C. capitata ofrecido a P. concolor………………………………..
109
Figura 5.3. Curvas de supervivência de Kaplan–Meier de hembras (A) y machos
(B) of Psyttalia concolor alimentadas con diferentes recursos: (W - agua; D -
glucosa; Phe - Fenilalanina; Pro - Prolina; Trp - Triptófano; Mix – mezcla de D +
Phe + Pro + Trp). ……………………………………………….................................
111
Figura 5.4. Esquema de los resíduos Schoenfeld en función de los diferentes niveles
de la variable categórica “Fuente de alimento” del Cox PHMs para adultos de
Psyttalia concolor (Cox PHM1 para hembras: A, B, C, D; Cox PHM3 para machos:
E, F, G, H, I). ………………………………………………………………………...
114
Figura 5.5. Esquema de los resíduos Schoenfeld’s en función de los diferentes
niveles de la variable categórica “Fuente de alimento” del Cox PHM2 para adultos
de Psyttalia concolor (A, B, C, D) con la variable categórica adicional “edad de
muerte”…………………………………………………………………………........
115

xix
Figura S5.1. Esquema del riesgo de muerte para cada tratamiento ensayado: A:
Hembras < 55 días; B: Hembras > 55 días y C: Machos……………………………...
123
Figura 6.1. Esquema secuencial de los métodos estandarizados de la OILB para
ensayar efectos de plaguicidas en los enemigos naturales * No se necesitan ensayos
adicionales; *2 Rutas opcionales. Figura adaptada de Dohmen, 1998………………...
128
Figura 6.2. Relación biológica entre la mosca del olivo e insectos beneficiosos y la
aplicación de productos fitosanitarios………………………………………………..
131
Figura 6.3. Bioensayo de contacto: A: Psyttalia concolor hembra; B: Ensayo
experimental para evaluar la mortalidad con (1) agua destilada, (2) dieta y (3) ramas
de olivo tratadas; C: Hojas de olivo (cv. Picual) tratadas con un pulverizador
manual; D y E: Ramas de olivo completamente cubiertas con el producto
fitosanitario hasta su secado………………………………………………………….
140
Figura 6.4. A: Cajas de parasitación utilizadas para evaluar los efectos subletales de
los plaguicidas en la reproducción de Psyttalia concolor. (1) Hembra de P. concolor
dispuesta de forma individual para la parasitación de, (2) 20 larvas (tercer estadío
larvario) de C. capitata ………………………………………………………………
140

xx
LIST OF TABLES
Table 4.1. Percentage of attacked hosts and progeny size in the different treatments
with single sugars and solid diet…………………………………………...................
63
Table 5.1. Output para supuesto de riesgo proporcional (PHA) teste……………... 81
Table 5.2. Percentages of parasitoid emergence, attacked hosts and progeny size in
the different experiments with amino acids (mean ± Standard Error of the mean-
SE)…………………………………………………………………………………...
85
Table 6.1. Toxicity data of insecticides tested against Psyttalia concolor adult
females………………………………………………………………………………
102
Table 6.2. Toxicity data of fungicides and herbicides tested against Psyttalia
concolor adult females in different experiments……………………………………..
104
Table 6.3. Toxicity data of biopesticides tested against Psyttalia concolor adult
females………………………………………………………………………………
105
Table 6.4. Pesticides tested: type (herbicide, fungicide or insecticide), active
ingredient, trade name, mode of action and chemical group and code (HRAC, FRAC
and IRAC classification), concentration tested, dosis per application and maximum
application rates (MAPAMA, 2018) and main target in olive crops………………….
107
Table 6.5. Exposure to pesticides residues on olive tree leaves (Extended Laboratory
Test): Lethal effects measured as cumulative mortality of P. concolor females (%)
at 2, 4, 24, 48 and 72 hours after application (HAT) of different pesticides and IOBC
categories…………………………………………………………………………….
111
Table 6.6. Exposure via ingestion (Laboratory Test): Lethal effects measured as
cumulative mortality of P. concolor females (%) at 2, 4, 24, 48 and 72 hours after
application (HAT) of different pesticides and IOBC
categories…………………………………………………………………………….
113
Table 6.7. Percentages of attacked hosts, progeny size and sex/ratio of Psyttalia
concolor after 72 h exposure (contact or ingestion) to pesticides (mean values ±
standard error)………………………………………………………………………..
114
Table S6.1. Exposure of P. concolor to pesticides residues on olive tree leaves. Only
significant pairwise comparisons of treatments are presented………………………..
136
Table S6.2. Exposure of P. concolor to pesticides via ingestion. Only significant
pairwise comparisons of treatments are presented…………………………………..
137

xxi
RELACIÓN DE TABLAS
Tabla 4.1. Porcentaje de huéspedes no emergidos y ratio de parasitismo en los
diferentes tratamientos con azúcares individuales y dieta sólida……………………
63
Tabla 5.1. Evaluación del supuesto de riesgos proporcionales (PHA) teste ……...... 81
Tabla 5.2. Porcentaje de emergencia del parasitoide, huéspedes atacados y progenie
en los diferentes experimentos con aminoácidos (media ± error estándar de la
media)………………………………………………………………………………..
85
Tabla 6.1. Toxicidad de insecticidas en hembras de Psyttalia concolor…………….. 102
Tabla 6.2. Toxicidad de fungicidas y herbicidas en hembras de Psyttalia concolor…. 104
Tabla 6.3. Toxicidad de bioplaguicidas en hembras de Psyttalia concolor ………. 105
Tabla 6.4. Plaguicidas ensayados: tipo (herbicida, fungicida o insecticida),
ingrediente activo, nombre comercial, modo de acción y grupo químico y código
(clasificación HRAC, FRAC e IRAC), concentración de producto fitosanitario
ensayada (en 100 ml de agua), dosis por aplicación, máximo ratio de aplicación
(MAPAMA, 2018) y plaga/s que combate/n………………………………………...
107
Tabla 6.5. Exposición a los resíduos de plaguicidas en hojas de olivo (Ensayo de
laboratorio extendido): Mortalidad acumulada en hembras de P. concolor (%) a las
2, 4, 24, 48 y 72 horas tras la aplicación (HAT) de diferentes plaguicidas. Categorías
de la OILB…………………………………………………………………………...
111
Tabla 6.6. Exposición via ingestión (Ensayo de laboratorio): Mortalidad acumulada
en hembras de P. concolor (%) a las 2, 4, 24, 48 y 72 horas tras la aplicación (HAT)
de diferentes plaguicidas. Categorías de la OILB……………………………………
113
Tabla 6.7. Porcentajes de huéspedes atacados, progenie y ratio sexual de Psyttalia
concolor tras 72 horas de exposición (contacto o ingestión) a diferentes plaguicidas
(media ± error estándar)……………………………………………………………...
114
Tabla S6.1. Exposición de P. concolor a resíduos de plaguicidas en hojas de olivo.
Solo se presentan comparaciones por pares………………………………………….
136
Tabla S6.2. Exposición de P. concolor a plaguicidas vía ingestión. Solo se
presentan comparaciones por pares………………………………………………….
137

xxii
ABSTRACT
The olive fruit fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae), is the main pest of
the olive tree worldwide. In Trás-os-Montes, the levels of pest attack have increased
significantly in the last years. Among the complex of natural enemies, the braconid wasp
Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) is an important
endoparasitoid of the pest. However, pesticides application for pest management in olive
orchards might change parasitoids efficacy in biological control of B. oleae. The aim of
this work will be to study (a) the effect of non-crop plants on the fitness of P. concolor
and (b) evaluate the susceptibility of parasitoid to some pesticides used in Mediterranean
olive groves.
Sugars and amino acids are an important food source required by this parasitoid to
increase survival, fecundity and fitness. Moreover, is expected that sugars and amino
acids increase the potential of parasitoid as a biological control agent of B. oleae. On this
basis, we test the effect of (1) seven sugars: glucose and fructose, sucrose, trehalose and
melibiose, melezitose and sorbitol, (2) two types of honeydews excreted by hemipteran
pests present in olive groves, Saissetia oleae Olivier (Coccidae) and Euphyllura olivina
(Costa) (Psyllidae) and (3) three amino acids: phenylalanine, proline and tryptophan
(singly and mixed (Mix) )- on the longevity and reproduction of P. concolor. Survival
was checked every 24 hours and the reproductive parameters were assessed by calculating
the percentage attacked hosts and progeny size. Our results showed a positive relationship
between some carbohydrate (single sugars and honeydew) and amino acids and parasitoid
survival. For females, the sugars sorbitol and melibiose increased the longevity and the
amino acids phenylalanine and proline showed a significant reduction of death hazard.
While for males, glucose and fructose were the best food resources (hexose-nectars) and
the consumption of the Mix and phenylalanine reduce the risk of dying. Interestingly, P.
concolor benefited more from feeding on honeydew than on sugars, representing this food
item an important source of sugar in olive groves. Moreover, in general, female
parasitoids lived longer than males.
Concerning the reproduction, we also found that sugar solution increased the
reproductive performance more than amino acids. Sucrose are the sugar that benefit more
the reproduction while melezitose decreased significantly, comparatively to the other
sugars offered.

xxiii
Insecticides are generally more toxic to parasitoid, above all organophosphates and
pyrethroids, while fungicides, herbicides and biopesticides have, in general, less effects
on mortality and reproductive parameters of P. concolor.
Keywords: Amino acids, honeydew, longevity, parasitoid, pesticides, sugars

xxiv
RESUMEN
La mosca del olivo, Bactrocera oleae (Rossi) (Diptera: Tephritidae), es la principal
plaga del olivo a nivel mundial. En Trás-os-Montes (Portugal), los niveles de ataque se
han incrementado de forma significativa en los últimos años. Entre el complejo de
enemigos naturales de la mosca del olivo, destaca el bracónido endoparasitoide Psyttalia
concolor (Szépligeti) (Hymenoptera: Braconidae). Sin embargo, la aplicación de
plaguicidas para el control de la plaga podría alterar la eficacia del potencial control
biológico que el parasitoide pudiera ejercer sobre la mosca del olivo. El objetivo de este
trabajo será estudiar: (a) los efectos de los recursos alimenticios que ofrecen las plantas
no cultivadas que aparecen en el olivar en el desarrollo y comportamiento de P. concolor
y (b) evaluar la susceptibilidad del parasitoide a determinados plaguicidas utilizados en
olivares mediterráneos.
Los azúcares y los aminoácidos son una fuente de alimento imprescindible para
incrementar la supervivencia y la capacidad reproductiva de P. concolor. Además, se
espera que los azúcares y los aminoácidos también incrementen del potencial de P.
concolor como agente de control biológico de B. oleae. En este contexto, analizamos los
efectos de (1) siete azúcares: glucosa y fructosa, sucrosa, trehalosa y melibiosa,
melecitosa y sorbitol, (2) dos tipos de melazas excretadas por plagas de hemípteros
presentes en los olivares como son Saissetia oleae Olivier (Coccidae) y Euphyllura
olivina (Costa) (Psyllidae) y (3) tres aminoácidos: fenilalanina, prolina y triptófano
(aislados y mezclados (Mix))- en la longevidad y reproducción de P. concolor. Se evaluó
la supervivencia cada 24 horas y los parámetros reproductivos se determinaron
calculando el porcentaje de huéspedes atacados y de parasitoides emergidos. Nuestros
resultados mostraron una relación positiva entre algunos carbohidratos (azúcares y
melazas) y aminoácidos y supervivencia del parasitoide. En el caso de las hembras, los
azúcares sorbitol y melibiosa incrementaron la longevidad y los aminoácidos fenilalanina
y prolina mostraron una reducción significativa del riesgo de muerte. Por el contrario, en
los machos, la glucosa y la fructosa fueron los mejores recursos alimenticios y el consumo
de la mezcla (Mix) y la fenilalanina redujo el riesgo de muerte. Curiosamente, las melazas
son mejor alimento que los azúcares para el parasitoide, siendo estas una fuente de
azúcares muy relevante en los olivares. Además, en general, las hembras sobrevivieron
más tiempo que los machos. Con respecto a la reproducción del parasitoide, encontramos

xxv
que las soluciones azucaradas influían más positivamente que los aminoácidos, siendo la
sucrosa la más efectiva y la que menos la melecitosa, en relación al resto de azúcares.
Cuando se revisó el efecto de los distintos fitosanitarios utilizados en olivar en la
supervivencia del parasitoide, se comprobó que los insecticidas son los más tóxicos,
especialmente organofosforados y piretroides, mientras que los fungicidas, herbicidas y
biopesticidas tienen menor efecto en la supervivencia y en los parámetros reproductivos
de P. concolor.
Palabras clave: Aminoácidos, azúcares, longevidad, melazas, parasitoide,
plaguicidas.

xxvi
RESUMEN EXTENDIDO DE LA TESIS
El cultivo del olivo es un uso muy importante de la tierra en la región mediterránea,
con una muy destacada importancia medioambiental, social y económica (Loumou and
Giourga, 2003). Sin embargo, es atacado por diferentes plagas que reducen su
rendimiento y, como consecuencia, la calidad del aceite de oliva y los ingresos de los
agricultores. Entre otras plagas, la mosca del olivo, Bactrocera oleae (Rossi) (Diptera:
Tephritidae), es la de mayor importancia en el mundo donde se cultiva el olivar (Daane
and Johnson, 2010). Esta plaga se controla principalmente con plaguicidas sintéticos,
cuyos efectos nocivos en el medioambiente y en la salud humana, así como en la
biodiversidad y en el uso potencial del control biológico son sobradamente conocidos.
Además, las aplicaciones químicas pueden dar lugar al repunte de la población plaga
por la reducción de los enemigos naturales o el desarrollo de la resistencia a insecticidas
(Soares, Pereira and Bastos, 2004; Hawkes et al., 2005). En los últimos años, B. oleae
tiene mayor importancia en la parte norte de Portugal, un lugar donde tradicionalmente el
ataque de la mosca ha sido bajo. Este incremento en el nivel de ataque está generando
preocupación entre los agricultores. En olivares sostenibles, el control de la mosca se
debería conseguir maximizando la acción de sus enemigos naturales. En este contexto,
innovar en el conocimiento de aspectos derivados de las interacciones plaga-parasitoide
es necesario y deberían ser explorados. Entre el complejo de enemigos naturales, el
bracónido Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) está considerado
como un endoparasitoide de importancia para el control de la mosca del olivo así como
de otros tefrítidos de importancia económica (Benelli and Canale, 2013). Además, en
laboratorio, P. concolor se cría fácilmente en un huésped alternativo, la mosca de la fruta
Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Se estudiaron, las interacciones
planta-parasitoide para favorecer el establecimiento de P. concolor en el olivar, así como
su capacidad para completar su ciclo de vida. Los parasitoides en fase adulta necesitan
recursos florales (polen y néctar) y también melazas para satisfacer sus necesidades
energéticas y completar la maduración de su aparato reproductor (Winkler et al., 2006)
Por tanto, la conservación o el aumento de poblaciones de P. concolor está estrechamente
ligado al mantenimiento de plantas no cultivadas (por ejemplo, márgenes de vegetación
o arbustos) dentro o alrededor del olivar. Esto implica el estudio de los efectos de plantas
no cultivadas como fuente de energía suplementaria (azúcares, aminoácidos y melazas)
en la supervivencia, reproducción y comportamiento de P. concolor, así como la

xxvii
comprensión de cómo contribuyen al mantenimiento del parasitoide en los olivares. La
investigación de estas interacciones finalmente suministrarán información valiosa para la
implementación de técnicas de control sostenibles.
Por tanto, el objetivo de este trabajo será el estudio de los diferentes componentes de
la interacción P. concolor-mosca del olivo, así como el efecto de las plantas no cultivadas
en el desarrollo y comportamiento P. concolor, con el fin de promover el control
biológico en los olivares. Para cumplir con este propósito, el programa de trabajo se
organice en tres partes con diferentes objetivos parciales:
Revisión bibliográfica: “BIOECOLOGÍA DE PSYTTALIA CONCOLOR (SZÉPLIGETI)
(HYMENOPTERA: BRACONIDAE) Y ESTRATEGIAS DE MANEJO PARA INCREMENTAR
EL CONTROL BIOLÓGICO DE LA MOSCA DEL OLIVO” (Capítulo 3).
Este capítulo disecciona los principales factores que contribuyen a la abundancia de P.
concolor en el olivar. Se sabe que los diferentes recursos alimenticios tienen un impacto
en la longevidad del parasitoide, pero su influencia en la interación plaga/parasitoide es
todavía desconocida, así como su efecto en el control biológico y, consecuentemente, en
las decisiones a tomar para un manejo sostenible.
El establecimiento de P. concolor en un nuevo hábitat depende de una combinación
de factores abióticos y bióticos. Algunos factores tales como el tamaño del fruto, peso,
color, dureza del epicarpo del fruto, estado fenológico del fruto del olivar y factores
químicos pueden influir en la atracción de la hembra P. concolor por las diferentes
variedades cultivadas del olivar. Está claro que la estructura del paisaje juega un papel
importante en la supervivencia del parasitoide, la nutrición y su actividad. Por tanto, es
fundamental estudiar los hábitos alimenticios y los requerimientos nutricionales y sus
consecuencias en la longevidad, desarrollo del insecto y reproducción. La abundancia de
huéspedes alternativas es igualmente importante, especialmente en ausencia de fruto en
el olivar. Teniendo esto en cuenta, es esencial encontrar plantas con tefrítidos alternativos
y conservar o incrementar la abundancia de estas plantas. Además, se debe profundizar
en el conocimiento de los mecanismos de interacción huésped - P. concolor, así como en
los del huésped en relación a estímulos asociados de comportamiento de las hembras (por
ejemplo, búsqueda con las antenas, con el ovipositor y vibrotaxis). Muchas cuestiones
sobre la interacción entre P. concolor y B. oleae necesitan todavía ser clarificadas, entre

xxviii
ellas, la identificación de los compuestos volátiles que están envueltos en esas
interacciones y que pueden influir en la identificación del huésped y en el comportamiento
ovipositor P. concolor.
Estos estudios contribuirán a mejorar las prácticas de manejo en el olivar, así como el
control biológico de la mosca del olivo mediante la atracción y conservación de P.
concolor en el cultivo. Además, se necesita profundizar en los estudios de longevidad de
P. concolor con el objetivo de obtener mayores ratios de parasitismo y, en consecuencia,
menores densidades de B. oleae. Otros enemigos naturales de la mosca del olivar también
podrían ser utilizados junto con P. concolor para optimizar el control.
Con el objetivo de estudiar el efecto de plantas no cultivadas espontáneas como
fuente de recursos suplementarios en la supervivencia, reproducción y
comportamiento de P. concolor y comprender cómo atraen y contribuyen a la
conservación del parasitoide en los olivares (Capítulos 4 y 5);
Se diseñaron experimentos en laboratorio para estudiar la alimentación de especímenes
adultos de P. concolor en:
(1) Azúcares: (Capítulo 4);
Se estima que el potencial de P. concolor como agente de control biológico de B. oleae
se incrementará cuando se suministren diferentes azúcares. Los azúcares son una fuente
importante de alimento que los adultos de este parasitoide necesitan para incrementar su
supervivencia, fecundidad y sanidad en general. El objetivo de este estudio fue estudiar
los efectos de siete azúcares (glucosa, fructosa, sucrosa, trehalosa, melibiosa, melecitosa
y sorbitol) en la longevidad y capacidad parasítica P. concolor. Además, evaluamos los
efectos de dos tipos de melaza excretada por plagas de hemípteros presentes en el olivar
Saissetia oleae Olivier (Coccidae) and Euphyllura olivina (Costa) (Psyllidae) en la
longevidad del parasitoide.
MATERIALES Y METODOS
Cría de insectos

xxix
Psyttalia concolor se crió sobre C. capitata en la Unidad de Protección de Cultivos,
ETSIAAB-UPM, siguiendo la metodología descrita por Jacas and Viñuela (1994). Ambos
insectos se desarrollaron en unas condiciones de 25 ± 2ºC de temperatura, 75±5% de
humedad and un fotoperíodo 16:8h luz:oscuridad. Los parasitoides no fueron alimentados
ni bebieron agua antes del comienzo de los experimentos. C. capitata se utilize como
modelo para estudiar el efecto en los diferentes recursos alimenticios en la capacidad
parasítica de P. concolor.
Alimento
Se evaluaron siete azúcares: dos monosacáridos (D(+)-Glucosa 1-hydratada
(RFE, USP, BP, Ph. Eur., DAB) PRS-CODEX, PanReac AppliChem y D(+)-Fructosa,
99%, ACROS Organics), tres disacáridos ((D+)-Sacarosa, + 99% ACROS Organics;
D(+)-Trehalosa dihydratada, 99%, ACROS Organics and Melibiosa, 98 % Sigma-
Aldrich)), un trisacárido (D(+)-Melezitosa monohidratada, 99+%, Acros Organics) y un
azúcar alcohol (Sorbitol BioUltra, ≥99.5% (HPLC), Sigma-Aldrich). Estos azúcares se
seleccionaron basándose en que son los componentes mayoritarios de los azúcares
procedentes de plantas (néctar floral and melaza - ver Wäckers, 2001). La concentración
utilizada en todas las soluciones azucaradas fue 1 M, que constituye la concentración más
alta de azúcares encontrados en néctar floral y melazas (Wäckers, 2001).
Para el tratamiento con melazas (experimento 2), se recogieron melazas de S. oleae y
E. olivina en plantones de olivar de 2 años de edad mantenidos en una cámara climática
en las siguientes condiciones: 24 ± 1ºC de temperatura, 70 ± 5% de humedad relativa y
un fotoperíodo de 16:8 h (L:D) en la Escuela de Agricultura del Instituto Politécnico de
Braganza. Un pedazo de Parafilm® (5 cm x 5 cm) se colocó bajo ramas infestadas con
cada uno de los productores de melaza durante 24 horas. Después, las gotas de melaza
excretadas sobre el Parafilm® se ofrecieron a los parasitoides
Longevidad y capacidad parasítica de adultos de Psyttalia concolor alimentados con
diferentes azúcares
Longevidad
En los experimentos de longevidad, cinco hembras y machos de P. concolor recién
emergidos (< 24 h) (Fig. 4.1. A) fueron colocados en una caja de plexiglás (12 cm de
diámetro x 5 cm de altura) con un agujero en la tapa (5.5 cm de diámetro) cubierto con

xxx
una malla, con el objetivo de favorecer la ventilación. Cada tratamiento constó de seis
replicas, cada una de ellas constituídas por una caja.
Cada solución azucarada se ofreció de forma individual en un recipiente de cristal (2
ml) tapado con Parafilm®, en el que se había introducido un trozo de bayeta para
favorecer la capilaridad. Los recipientes de vidrio se sujetaron a la base de la caja con un
pedazo de plastilina. En cada caja, se colocaron dos recipientes, el que contenía la dilución
azucarada y otro con agua destilada. Como control negativo se utilizó solamente agua
destilada y como control positivo una mezcla de levadura de cerveza y azúcar (1:4). Los
recipientes con azúcares y dieta sólida se rellenaron diariamente, cambiándose
completamente cada semana, mientras que la melecitosa se renovó diariamente debido a
su propensión a cristalizar. La supervivencia del parasitoide se evaluó diariamente.
Capacidad parasítica
Los parámetros de capacidad parasítica, porcentaje de huéspedes atacados (porcentaje
de pupas sin emergencia de mosca de la fruta) y el porcentaje de la descendencia
(porcentaje de parasitoides emergidos de las pupas parasitadas) se evaluaron usando cajas
similares a las de los experimentos de longevidad, con una variante: un agujero adicional
(5.5 cm de diámetro) en la base de la caja cubierto con una malla. Se situó una hembra
fecundada en la caja y se alimentó con un azúcar sencillo o dieta sólida (control).
Después de siete días, el tercer estadío larvario de C. capitata se ofreció a hembra de
P. concolor en una proporción de 20 larvas/hembra situándolas en lo alto de un bote de
vidrio cubierto con una pieza Parafilm® (4 cm de diámetro) durante una hora (según
Bengochea et al. (2014) y este procedimiento se repitió durante cinco días consecutivos.
Posteriormente, las larvas de C. capitata fueron transferidas a placas Petri y situadas en
una cámara visitable a 25 ± 2ºC, 75 ± 5% y 16: 8 h luz: oscuridad hasta la emergencia.
Se anotó el tiempo transcurrido desde el estado de pupa hasta la emergencia de los adultos
C. capitata. Solo se tuvieron en cuenta los datos obtenidos de los últimos cuatro días de
parasitación, puesto que estudios previos han demostrado que necesitan al menos un día
para aprender a parasitar y los datos del primer día son muy erráticos. Un total de 25
hembras por tratamiento fueron utilizadas en los experimentos.
Longevidad de Psyttalia concolor alimentada en melazas
Grupos de 10 machos o hembras recién emergidos (12-24 h edad) fueron colocados en
cajas de plexiglás (7.5 cm diámetro x 4.5 cm altura) con un agujero en la tapa cubierto

xxxi
por una malla para facilitar la ventilación. Cada caja contenía (1) melaza (S. oleae o E.
olivina) + agua destilada o (2) 10% miel multifloral de montaña (Bragança Mountain) +
agua destilada (control positivo) o (3) solamente agua destilada (control negativo). El
agua y la miel fueron suministrados en tubos para microcentrífuga de 1.5 ml con una tira
de bayeta y Parafilm®. La melaza se suministró como se ha descrito en el epígrafe 3.2.
La dieta ofrecida a los adultos se reemplazó a diario. Las cajas con los parasitoides se
mantuvieron en una cámara climática (25 ± 2ºC, 75 ± 5% y 16:8 h) como la descrita
anteriormente y la supervivencia de los adultos en cada caja se evaluó hasta que murió el
último adulto superviviente. Cada tratamiento se realize con 30 parasitoides de cada sexo.
Análisis de datos
Longevidad
La longevidad media y los errores estándar fueron calculados de forma separada para
cada uno de los sexos de P. concolor. Todos los análisis y gráficos se desarrollaron con
el programa R (R core team 2014). Para los análisis de supervivencia, la función survfit
[en el paquete survival ] fue utilizada para estimar las diferentes curvas de supervivencia
según el método de Kaplan–Meier. La curva de supervivencia Kaplan–Meier de los
tratamientos individuales fue comparada con log-rank test survdiff. El modelo no
paramétrico de riesgos proporcionales de Cox (function coxph [en el paquete survival ])
se utilizó para analizar el efecto de la fuente de alimentación en la supervivencia del
parasitoide. La hipótesis de riesgos proporcionales se evaluó por inspección visual usando
la función cox.zph del mismo paquete.
Capacidad parasítica
El efecto de los azúcares en el porcentaje de huéspedes atacados y de parasitoides
emergidos fue analizado utilizando la prueba de Kruskal-Wallis, mediante la función
Kruskal del paquete agricolae. El análisis de los datos se realizó con el programa R.
RESULTADOS Y DISCUSIÓN
Los azúcares y las melazas ensayadas en este trabajo presentaron variación con
respecto a su efecto en la longevidad de machos y hembras. Nuestros resultados
mostraron que: (1) hembras y machos de P. concolor tuvieron diferentes preferencia o
necesidades de azúcares, y (2) los azúcares tienen mayor efecto positivo en las hembras
que en los machos para un azúcar dado. De la misma forma, Williams III et al. (2015)

xxxii
encontraron que las hembras de P. lounsburyi (Silvestri) (Hymenoptera: Braconidae)
tenían mayor longevidad que los machos. Es posible que este efecto se deba a que las
hembras tienen mayores requerimientos nutricionales que los machos por su mayor gasto
energético buscando huésped y ovipositando (Zhang et al., 2014).
Las hembras de P. concolor vivieron más cuando se alimentaron de azúcares
típicamente presentes en los néctares de plantas de la zona mediterránea (y sólo
esporádicamente encontrados en melazas) (Petanidou, 2005), por ejemplo, sorbitol y
melibiosa. Aparte de su valor nutricional, Burke and Waddell (2011) demostraron que el
sorbitol es un azúcar que se mantiene mayor tiempo en la memoria, una característica
muy importante para reforzar la memoria del apetito, aunque no se considere un azúcar
dulce. Además, la acumulación de sorbitol en parasitoides está también asociada a la
tolerancia a bajas temperaturas (Bale, 2002), lo que favorece la presencia de los
parasitoides en agroecosistemas donde el invierno es más severo. Por tanto, de acuerdo
con estos hallazgos, las flores cuyo néctar contenga sorbitol y melibiosa deberían estar
presentes o ser preservadas en los olivares, tales como Asparagus acutifolius L.
(Asparagaceae) y Convolvulus arvensis L. (Convolvulaceae) (Petanidou, 2005; Nave et
al., 2017b)
En este estudio, P. concolor alimentada en sucrosa vivió menos tiempo comparado con
los otros azúcares ensayados y el control positivo. Por el contrario, otros estudios
evidenciaron el efecto positivo de la alimentación con sucrosa en otros bracónidos tales
como Cotesia glomeratae (L.) (Lee and Heimpel, 2008), Chelonus elaeaphilus Silvestri
(Nave et al., 2017a) and P. lounsburyi (Williams III et al., 2015). Esto demostró que la
influencia de los azúcares en la longevidad varía según las species, así como la capacidad
para digerirlos y sintetizarlos (see Wäckers, 1999; Wäckers et al., 2006; Tompkins,
Wratten and Wäckers, 2010).
En especies sinovigénicas, como P. concolor, el porcentaje de huéspedes atacados es
un factor importante para la eficiencia del control biológico. En nuestro estudio, las
hembras alimentadas con sucrosa incrementaron el porcentaje de huéspedes atacados y
de los parasitoides emergidos en relación a los otros azúcares ofrecidos, sugiriendo que
este azúcar podría rápidamente remplazar la energía gastada en la reproducción.
Curiosamente, en los tratamientos con azúcar melecitosa se redujo el porcentaje de
huéspedes atacados (de forma significativa) y de parasitoides emergidos. Este efecto
puede ser debido al hecho de que melecitosa es un azúcar que cristaliza rápidamente, lo
cual dificulta la ingestión por el parasitoide (Wäckers, 2000).

xxxiii
In general, la porcentaje de parasitoides emergidos observado en P. concolor
alimentado en azúcares tuvo rango que osciló entre 50-60%, lo que indica un riesgo de
puesta limitada de huevos en el campo, similar al observado en otras especies (see
Heimpel and Rosenheim, 1998). Los nutrientes necesarios para la producción de huevos
son parcialmente obtenidos de proteínas y lípidos, mientras que los carbohidratos
incrementan la duración de la fecundidad en los parasitoides (Benelli et al., 2017). Es
importante considerar que los parasitoides necesitan otros nutrientes básicos tales como
vitaminas, sales minerales, esteroides y ácidos nucleicos para su normal desarrollo y
reproducción.
La glucosa y la fructosa, dos de los principales azúcares presentes en varios néctares
florales y melazas, son los responsables de la mayor longevidad de los machos. Estos
resultados indican que los machos de P. concolor parecen ser más generalistas que las
hembras, sugiriendo que los néctares donde las hexosas son dominantes beneficiarían más
a los machos que a las hembras. Estos azúcares se pueden encontrar en olivar en plantas
silvestres con flores del tipo Raphanus raphanistrum (Brassicaceae) and Malva neglecta
(Malvaceae) o en plantas con néctar ricos en hexosas, tales como Daucus carota
(Apiaceae), Conopodium majus (Apiaceae), Foeniculum vulgare (Apiaceae), Asparagus
acutifolius (Asparagaceae) and Anarrhinum bellidifolium (Scrophulariaceae) (Nave et al.,
2017a).
Por otro lado, los machos que se alimentaron exclusivamente con melibiosa tuvieron
menor longevidad (aproximadamente 10 días, mientras que aquellos alimentados con
glucosa o fructosa, tenían una longevidad media de más del doble) Este dato concuerda
con un estudio realizado en Cotesia glomerata (L.) (Hymenoptera: Braconidae) sobre
cuantificación de comida ingerida que sugería que, contrariamente a la melibiosa, la
glucosa y la fructosa fueron inmediatamente convertidas en energía, cumpliendo con los
requisitos nutricionales de los parasitoides (Hausmann, Wäckers and Dorn, 2005).
Hoy en día, la investigación en longevidad de los parasitoides alimentados en melazas
es contradictoria, ya que algunos estudios indican que las melazas no incrementan la
supervivencia de los parasitoides, mientras que otros muestran lo contrario, aunque
siempre como una fuente de alimentación menos rica que los néctares o las soluciones de
miel (reviewed by Wäckers and van Rijn, 2005). Sin embargo, hay una gran variación de
tipos de melaza con diferente contenido en carbohidratos que da lugar a diferentes
longevidades en los parasitoides (Tena et al., 2018). Además, una menor longevidad
puede ser atribuida a una elevada viscosidad y a la tendencia de algunas melazas a

xxxiv
cristalizar rápidamente (Faria, Wäckers and Turlings, 2008). En nuestro estudio, la
melaza no fue un alimento de inferior calidad desde el punto de vista del incremento de
la supervivencia de las hembras y machos de P. concolor, obteniéndose incluso valores
más altos que con los azúcares sencillos. La melaza de S. oleae y E. olivina fue un buen
recurso alimenticio para las hembras, obteniéndose un mayor valor de longevidad que
con miel diluída (control positivo). Sin embargo, para los machos, la melaza de E. olivina
dio lugar al valor más alto de longevidad, mientras que S. oleae and la miel mostraron
resultados similares. Por el contrario, Villa et al. (2017) encontraron que las melazas de
E. olivina y S. oleae redujeron el riesgo de muerte de las hembras de Elasmus flabellatus
(Fonscolombe) (Hymenoptera: Eulophidae) en comparación con otras flores evaluadas,
pero no con una solución de miel (control positivo)
Los resultados de Wang et al. (2011) mostraron que la melaza de S. oleae incrementó
la longevidad de los parasitoides P. humilis (Silvestri) (Hymenoptera: Braconidae) y
Scutellista caerulea (Fonsc.) (Hymenoptera: Pteromalidae) cuando fueron alimentados
con ella una única vez.
En el campo, cuando las flores producen poco o ningún nectar, las melazas de E. olivina
y S. oleae pueden ser la principal fuente de carbohidratos disponible (Wäckers, 2005).
Sin embargo, este recurso solo se encuentra disponible durante unos pocos meses al año
y, con frecuencia, a muy baja densidad (Williams III et al., 2015). Por otro lado, una de
las ventajas de esta fuente de alimento es que los productores de melaza pueden estar
asociados al cultivo, a las malas hierbas próximas al cultivo o incluso a la vegetación de
los (Wäckers, van Rijn and Heimpel, 2008). Por tanto, la presencia de melazas cuando
el néctar no es abundante extiende la posibilidad de supervivencia de (incluso con una
única comida) y potencialmente incrementa la eficiencia del control biológico de las
poblaciones de B. oleae.
Una manipulación táctica que se ha propuesto como alternativa al manejo de las
plantas con flores implica la aplicación de soluciones de carbohidratos y/o proteínas a los
cultivos para que actúen como suplemento alimenticio, también conocido como
pulverización de comida artificial o melazas artificiales (Wade et al. 2008a, b; Tena et al.
2015). Esta estrategia adquiere una importancia primordial en cultivos cuya vegetación
natural se elimina mediante prácticas tales como el arado y la aplicación de herbicidas,
dando lugar a una reducción severa de los recursos florales (Walters, 2011). Además, la
escasez de plantas durante el verano mediterráneo puede eventualmente limitar los

xxxv
recursos azucarados durante un periodo de tiempo (debido a la baja calidad y cantidad de
los recursos) (Segoli and Rosenheim, 2013; Tena et al., 2016; Kishinevsky et al., 2017).
En conclusión, merece la pena destacar que los experimentos individualizando al
parasitoide en cajas no es necesariamente representativo de sus necesidades básicas en el
campo, pero sí son una indicación de la importancia de ciertos recursos alimenticios en
la vida de los parasitoides (Wäckers, 2001). Bajo condiciones más realistas, las
necesidades nutricionales de P. concolor serán más altas debido a su comportamiento de
búsqueda de huéspedes y de la parasitación en sí misma. Nuestros hallazgos suponen que
una buena fuente de carbohidratos (azúcares o melazas) es importante para el
mantenimiento de P. concolor en el campo, especialmente en la longevidad y oviposición
de la hembra del depredador. Por tanto, es importante suministrar comida mediante el
manejo de las plantas con flores o un suplemento artificial de comida. Considerando los
beneficios del suplemento de azúcares para los parasitoides, es importante desarrollar un
sistema de suministro de estos azúcares en el campo, ya sea mediante contenedores, gotas
en tronco, pulverizado de hojas o metodologías que permitan la conservación y la
disponibilidad de los azúcares en campo por un período de tiempo mayor.
(2) Aminoácidos: (Capítulo 5);
Psyttalia concolor es uno de los enemigos naturales asociados a la mosca del olivo, B.
oleae, más importante. La composición de los recursos alimenticios, usualmente en forma
de polen, néctar y melazas, son directamente responsables de la sanidad de los adultos del
parasitoide. Los insectos, en general, difieren mucho en sus preferencias nutritivas,
generalmente cuando se trata de aminoácidos. El efecto de los aminoácidos que entran a
formar parte de los recursos alimenticios del parasitoide en los olivares, tanto en su
supervivencia como en su éxito reproductivo no está del todo investigado. El objetivo de
este estudio es conocer el efecto de tres aminoácidos (fenilalanina, prolina y triptófano)
en la sanidad de P. concolor. Mientras el triptófano y la fenilalanina son aminoácidos
esenciales para los insectos, la prolina es un aminoácido muy común, pero no esencial.
Todos ellos se encuentran en el néctar, el polen y las melazas (Blüthgen, Gottberger and
Fiedler, 2004; Human and Nicolson, 2006; Petanidou et al., 2006; Leroy et al., 2011).
Nuestra hipótesis es que una dieta suplementada con uno o una mezcla de aminoácidos
puede ser beneficiosa para el ciclo de vida de P. concolor ya que (1) incrementaría su
longevidad; y (2) mejoraría la eficacia parasítica en larvas de B. oleae.

xxxvi
MATERIALES Y METODOS
Cría de insectos
Psyttalia concolor se crió sobre C. capitata en la Unidad de Protección de
Cultivos, ETSIAAB-UPM, siguiendo la metodología descrita por Jacas and Viñuela
(1994). Ambos insectos se desarrollaron en unas condiciones de 25 ± 2ºC de temperatura,
75±5% de humedad and un fotoperíodo 16:8h luz:oscuridad.
Fuentes alimenticias
Tres aminoácidos, L-Triptófano (Trp) (Sigma-Aldrich; ≥ 98%), L-Fenilalanina (Phe)
(Sigma-Aldrich; ≥ 98%), y L-Prolina (Pro) (Sigma-Aldrich; ≥ 99%) – fueron ensayados
de forma individual y mezclados a una concentración de 0.1 mM. Todos los aminoácidos
(ensayados de forma individual o mezclados) se disolvieron en D(+)-Glucosa (Sigma-
Aldrich; ≥ 99.5%) a una concentración 1 M.
La concentración de aminoácidos fue seleccionada basándonos en los experimentos
llevados a cabo Blüthgen and Fiedler (2004). Agus destilada y D(+)-Glucosa 1M fueron
utilizados como control positive y negative, respectivamente.
Longevidad
Para los experimentos de longevidad, cinco hembras y cinco machos recién emergidos
(< 24 h) de P. concolor se colocaron en una caja de plexiglás (12 cm de diámetro x 5 cm
de altura) con un agujero en la tapa (5.5 cm de diámetro) cubierto por una malla para
ventilación. Cada tratamiento consistió en seis repeticiones de 1 caja cada una.
Las soluciones de aminoácidos + glucosa se ofrecieron en un recipiente de cristal (2
ml) con una tira de bayeta y tapada con Parafilm®. Los recipientes se sujetaron a la base
de la caja de plexiglás con un pedazo de Plastilina. Se añadió además otro recipiente igual
con agua destilada. Las soluciones se rellenaron todos los días y se cambiaron cada
semana. La mortalidad de los individuos de P. concolor se registró a diario y los insectos
muertos se retiraron de las cajas y se almacenaron a –20ºC.
Reproducción
Treinta parejas (< 24 h) fueron colocadas en cajas de plexiglás (una por pareja) de
manera similar a las utilizadas en los experimentos de longevidad, pero con un agujero
adicional en el fondo de la caja (5.5 cm de diámetro) cubierto con una malla. Después de

xxxvii
siete días, larvas de tercer estadío de C. capitata fueron ofrecidas a hembras de P.
concolor en proporción 20 larvas/hembra situándolas en la parte alta de un bote de cristal
cubierto con un trozo de Parafilm® (4 cm de diámetro). Las larvas fueron ofrecidas
durante una hora, de acuerdo con Bengochea et al. (2014) y este procedimiento se repitió
durante cinco días. Posteriormente, estas larvas se transfirieron a placas Petri y colocadas
en una cámara visitable a 25 ± 2ºC, 75 ± 5% y 16: 8 h luz: oscuridad hasta la emergencia.
Se registró el tiempo transcurrido desde el estado de pupa hasta emergencia. El porcentaje
de huéspedes atacados (porcentaje de pupas sin emergencia de mosca de la fruta) y el
porcentaje de la descendencia (porcentaje de parasitoides emergidos de las pupas
parasitadas) fue evaluado. Solo cuatro días de parasitación se utilizaron en el análisis de
datos debido a la necesidad de aprender a parasitar de la hembra. Un total de 30 hembras
fueron estudiadas de forma individual en cada tratamiento.
Análisis de datos
Las curvas de supervivencia se representaron para cada tratamiento con estimaciones
de Kaplan-Meier utilizando las funciones Surv y survfit del paquete de supervivencia
(Therneau, 2014) en R (R core team, 2014).
El modelo no paramétrico de riesgos proporcionales de Cox (Cox PHM), utilizando la
función coxph del mismo paquete, se ajustaron para hembras y machos con el fin de
analizar el efecto de diferentes recursos alimentarios en la supervivencia de los adultos.
El supuesto de riesgo proporcional (PHA) de la regresión de Cox se confirmó
probando la no correlación entre los residuos de Schoenfeld y el tiempo de supervivencia
utilizando la función cox.zph del mismo paquete y trazando los residuos de Schoenfeld
suavizados contra los diferentes niveles de la variable categórica (Therneau y Grambsch,
2000). Cuando no se logró la PHA, en primer lugar, se identificó el momento específico
en que se violó la PHA en las parcelas residuales suavizadas de Schoenfeld. Luego, se
creó una variable categórica adicional (edad al morir) con dos niveles (Y: parasitoides
que mueren más jóvenes, O: parasitoides que mueren más viejos) para obtener diferentes
proporciones de riesgo para los parasitoides que mueren antes y después del punto
específico en que se violó el PHA. La función cluster en las fórmulas del modelo se utilizó
para indicar el intervalo con el fin de tener en cuenta las observaciones no independientes.
Los parámetros reproductivos calculados fueron: (a) media del parasitoide emergente
[± Error estándar (SE)], (b) porcentaje del huésped atacado [media ± Error estándar (SE)]
y (c) el porcentaje del tamaño de la progenie [media ± Error estándar (SE)].

xxxviii
La función ANOVA se usó para analizar las diferencias entre los tratamientos con
aminoácidos, seguido de una comparación por pares con Tukey`s rango
estudentizado (HSD) a p = 0.05.
RESULTADOS Y DISCUSIÓN
Existen varios estudios que demuestran que los azúcares ejercen un efecto positive en
las especies de himenópteros (Wäckers, 2001; Winkler et al., 2005; Harvey et al., 2012).
Sin embargo, nutrientes tales como las proteínas, los aminoácidos y las vitaminas pueden
también ser importantes para la sanidad de los parasitoides (Harvey et al., 2012). Sin
embargo, el efecto de estas fuentes alimenticias sobre la supervivencia y el éxito
reproductivo de P. concolor no es muy conocido. Este estudio tiene como objetivo
determinar el efecto de tres aminoácidos (triptófano, fenilalanina y prolina) y una mezcla
de los tres en la longevidad y parámetros reproductivos del parasitoide P. concolor, con
el objetivo de analizar su utilidad como recursos alimenticios potenciales.
Nuestros resultados son el primer estudio de los efectos del consumo de aminoácidos
en la supervivencia y reproducción de P. concolor, y su conocimiento puede ayudar a
explicar la necesidad de ciertas fuentes de alimento (polen, néctar y/o melazas) por el
parasitoide y mejorar el control de la plaga. Los alimentos ricos en aminoácidos son
extremadamente importantes en programas de control biológico para asegurar una
generación de hembras reproductivas, la prolongación del período de oviposición, el
incremento de la puesta de huevos, el ratio de maduración y la proporción de la ratio
sexual (Coskun and Emre, 2015). Nuestros datos demostraron que: (1) los recursos
alimenticios son fundamentales para hembras y machos de P. concolor y (2) el riesgo de
muerte asociado al consumo de diferentes dietas depende del sexo.
No fue una sorpresa que los parasitoides alimentados en agua exclusivamente tuvieron
una supervivencia muy corta en relación a aquellos bien alimentados. La adición de
aminoácidos a una concentración de 0.1 mM a la dieta no incrementó la supervivencia de
los parasitoides significativamente. Además, mientras no se detectaron diferencias entre
los aminoácidos y la glucosa (control positivo) en los machos cuando se observaron las
curvas de supervivencia, el consumo de fenilalanina produjo una menor supervivencia en
comparación con los valores obtenidos con glucosa. Aunque estos resultados indican que
los azúcares son necesarios para la supervivencia P. concolor, la concentración de
aminoácidos utilizada es probablemente insuficiente para satisfacer las necesidades
nutricionales de los parasitoides. El hecho de que añadir aminoácidos a la dieta no

xxxix
incrementase la supervivencia de los parasitoides no implica que no sean un importante
factor para su desarrollo y reproducción. De hecho, si consideramos el ratio de
probabilidad de muerte (Cox PHM), el riesgo de muerte a cualquier tiempo se redujo
tanto en hembras como en machos con ciertos aminoácidos. Independientemente de su
edad, las hembras alimentadas con fenilalanina y Prolina mostraron una reducción
significativa del riesgo de muerte, mientras que para los machos el riesgo de muerte se
redujo con el consumo de la mezcla de aminoácidos, seguido por fenilalanina. El efecto
de fenilalanina y prolina en la reducción del riesgo de muerte puede ser el resultado de la
fuerte actividad fagoestimulante de ambos aminoácidos (Carter et al., 2006; Petanidou et
al., 2006). La fenilalanina es esencial para el crecimiento del insecto y su desarrollo
(Coskun and Emre, 2015). Además, la fenilalanina es uno de los aminoácidos más
comunes en el néctar de las flores de especies de plantas (Petanidou et al., 2006)
asegurando una nutrición óptima en el campo. Por otro lado, la prolina puede
potencialmente ser utilizada en el metabolism del vuelo y la diapausa (Li et al., 2014;
Micheu, Crailsheim and Leonhard, 2000). Según nuestros resultados, el triptófano no
redujo el riesgo de muerte de hembras y machos de P. concolor. Este aminoácido
constituye un importante componente del exoesqueleto de los insectos y es
particularmente importante para la esclerotización de la cutícula (ver Hidalgo et al.,2014).
El triptófano también juega un importante papel en los receptores químico-sensorales de
los insectos (Nicolson and Thornburg, 2007) y fue atractivo para, por ejemplo, las
hembras de crisopas (Chrysoperla carnea Stephens, Hagen et al., 1976). De hecho, la
pulverización en campo de L-triptófano en olivares incrementó la abundancia de C.
carnea, un enemigo natural de la mariposa del olivo, Prays oleae (Bernard) (ver McEwen,
Jervis and Kidd, 1994).
Además, nuestros resultados de reproducción mostraron que añadir aminoácidos
(fenilalanina, prolina, triptófano y la mezcla correspondiente) no mejoró los parámetros
reproductivos en comparación con el control (glucosa). Curiosamente, solo los
parasitoides alimentados con prolina redujeron el porcentaje de hembras emergidas y el
de huéspedes atacados comparados con la glucosa. Además, los valores de porcentaje de
la descendencia fueron mucho más variables y, dependiendo el experimente oscilaron
entre el 14 y el 78%. Comparado con la glucosa, no hubo por tanto diferencias
significativas, salvo cuando el parasitoide se alimentó de la mezcla de aminoácidos. El
efecto de la prolina en los parámetros reproductivos parece ser diferente no solo de la
concentración sino también del tipo de dieta (ratio de proteínas (aminoácidos):

xl
carbohidratos) y de la especie de parasitoide (Hajirajabi et al., 2016). Según Hajirajabi et
al. (2016), la prolina tuvo efectos positives en el ratio de descendencia (mayor producción
de hembras), período de oviposición, fecundidad, fertilidad y ratio reproductivo para el
parasitoide Trissolcus grandis Thompson (Hymenoptera: Scelionidae). La prolina
ingerida por P. concolor podría haber sido utilizada como energía para funciones
somáticas (mantenimiento y locomoción) como se describe en la literatura (e.g. Arrese
and Soulages, 2010). Así, la prolina podría ser utilizada como energía rápida para
desplazamientos cortos y la glucosa para vuelos más largos (Carter et al., 2006).
Hay también evidencias de que la fenilalanina es un posible estimulante para los
parasitoides (Barrett and Schmidt, 1991). Sin embargo, nuestros resultados demostraron
que el consumo de fenilalanina no incrementaba la reproducción de los parasitoides.
Según Grenier (2012), altas concentraciones de fenilalanina pueden ser tóxicas para los
parasitoides y, puesto que en el presente estudio no afectó negativamente a la
supervivencia, podemos concluir que probablemente fueran metabolitos secundarios los
que indujeran la toxicidad en los procesos reproductivos. Por otro lado, el triptófano no
complete todos los requerimientos nutricionales para una correcta reproducción como se
observó también en Chrysoperla carnea (McEwen, Jervis and Kidd, 1994).
En parasitoides sinovigénicos como P.concolor, se parte de un número más bajo de
huevos maduros (la carga de huevos inicial), pero pueden ir madurando huevos a lo largo
de toda su vida adulta (Ellers, Sevenster and Driessen, 2000). Por ello, las hembras
necesitan obtener proteínas con mayor demanda de aminoácidos asociado a la producción
de huevos. Sin embargo, algunos parasitoides producen huevos mientras se alimentan de
fuentes azucaradas (Rivero and Casas, 1999). Esto podría sugerir que para el caso de P.
concolor, la ingestión de azúcares podría ser un requisito imprescindible para la
reproducción. Es también posible que el éxito reproductivo de la hembra no dependa
exclusivamente de la calidad y disponibilidad de los aminoácidos. Por ejemplo, para el
ectoparasitoide sinovigénico Eupelmus vuilletti (Crawford) (Hymenoptera, Eupelmidae),
sola la proteína obtenida por host-feeding incrementa la producción de huevos y la
supervivencia (Giron, Pincebourde and Casas, 2004). Además, algunos parasitoides
utilizan lípidos y proteínas de reserve acumuladas durante las fases larvarias para la
producción de huevos y la maduración (Cicero, Sivinski and Aluja, 2012).
En resumen, el consumo de aminoácidos afecta a la supervivencia de P. concolor y
tiene importantes implicaciones en la interacción huésped-parasitoide. Futuras

xli
investigaciones son necesarias para confirmar la importancia de suplementar la dieta para
maximizar la longevidad y reproducción del parasitoide en el campo.
Efectos secundarios de los productos fitosanitarios en el parasitoide
Psyttalia concolor (Szépligeti): Revisión y caso de estudio en laboratorio
La aplicación de plaguicidas para el manejo de las plagas del olivar podría modificar
la eficacia del control biológico de B. oleae. Cuando los plaguicidas son pulverizados en
los olivares, P. concolor puede ser contaminado por aplicación directa en forma de gotas,
por contacto con superficies tratadas o por ingestión de recursos alimenticios
contaminados (por ejemplo, alimentarse en agua contaminada, polen, néctar o melazas
disponible en partes de la planta tratadas). La aplicación de un plaguicida no solo reduce
la plaga, sino también las poblaciones del parasitoide porque necesitan cierta población
de la plaga para sobrevivir y dependen de su capacidad de recuperarse de los efectos
tóxicos del producto fitosanitario. Identificar plaguicidas selectivos con bajos efectos en
P. concolor y conocer el momento adecuado de aplicación o liberación de los parasitoides
proporciona una oportunidad de uso del parasitoide bajo IPM. El impacto causado por los
plaguicidas es importante para determinar el desarrollo de futuras estrategias de manejo
que conserven y/o incremente los niveles de población de los parasitoides en los olivares.
Con esta premisa, este trabajo es una recopilación de toda la información existente sobre
plaguicidas ensayados en P. concolor, que se presenta como revisión en la Tabla 6.1.
(insecticidas), Tabla 6.2. (fungicidas y herbicidas) y Tabla 6.3. (biopesticidas). La
información sobre plaguicidas que están actualmente registrados en la Unión Europea
se revisan en Tablas 6.1. y 6.2.
ESTUDIO DEL CASO EN LABORATORIO: EFECTO DE DIFERENTES PLAGUICIDAS EN LA
MORTALIDAD Y LA REPRODUCCIÓN DE HEMBRAS DE PSYTTALIA CONCOLOR
MATERIALES Y MÉTODOS
Condiciones de cría de Psyttalia concolor y Ceratitis capitata
Los adultos de P. concolor proceden de la parasitación en una cría masiva de la mosca C.
capitata instalada en un laboratorio (Unidad de Protección de Cultivos, ETSIAAB-UPM,
Madrid, Spain). C. capitata se utilizó como huésped de sustitución de B. oleae. La cría
de ambos insectos se mantuvo en una cámara visotable con condiciones

xlii
medioambientales controladas (25±2◦C, 75±10% humedad relativa y 16:8 L:D
fotoperíodo). Para profundizar más en los detalles de las crías, ver Jacas y Viñuela
(1994b) y Bengochea et al. (2014).
La elección de hembras de P. concolor para los experimentos se debió a su mayor
longevidad con respecto a los machos (Pinheiro et al., 2017), su mayor tolerancia a los
plaguicidas y a que las hembras son las responsables de la parasitación en campo (Jacas
y Viñuela, 1994a).
Plaguicidas
Se evaluaron ocho plaguicidas registrados para diferentes usos contra plagas
importantes del olivar en Portugal y/o en España (DGAV, 2018; MAPAMA, 2018): un
herbicida (gliphosato), tres insecticidas (beta-ciflutrin, deltametrin and zeta-cipermetrin)
and cuatro fungicidas (difenoconazol, dodina, tebuconazol and trifloxistrobin). Los datos
de los nombres comerciales, grupo químico y sitio de acción de acuerdo con la
clasificación de HRAC, FRAC e IRAC, la máxima concentración recomendada de
acuerdo con MAPAMA (2018) y la plaga/s que combate están resumidos en la Tabla 6.4.
El dimetoato, un insecticida sistémico, se utilizó como control positivo debido a su
eficacia controlando varias plagas del olivar y porque además es el insecticida más usado
contra B. oleae en áreas mediterráneas, como son los olivares de la península Ibérica
(Ruiz-Torres and Montiel-Bueno 2007; Pavlidi et al., 2018). El agua destilada sirvió
como control negativo.
Análisi de laboratorio
Los adultos de Psyttalia concolor fueron expuestos a los plaguicidas por dos vías:
contacto con hojas pulverizadas con el plaguicida o ingestión del mismo. Los parámetros
evaluados fueron: (1) mortalidad de la hembra del parasitoide P. concolor (%) y (2)
reproducción, como los huéspedes atacados (%) (percentaje de pupas sin emergencia de
la mosca) y descendencia (%) (porcentaje de parasitoides emergidos de los individuos
parasitados) (ver Bengochea et al., 2014 para más detalle). Los experimentos se realizaron
a las mismas condiciones controladas que se describen en el epígrafe anterior.
Ensayo por contacto

xliii
Tras la emergencia de adultos de P. concolor, diez hembras fecundadas (< 48-h-edad)
fueron colocadas en una caja de plexiglás (12 cm de diámetro x 5 cm de altura) que tenía
un orificio en la parte superior (5.5 cm de diámetro) cubierto con una malla para
ventilación. En cada caja, se suministró un recipiente de cristal (30 x 35 mm2) con agua
destilada, así como comida (levadura de cerveza y azúcar en las proporciones 1:4) en
tapones de plástico (24 x 6 mm2) ad libitum a hembras de P. concolor. En cada caja se
introdujo una rama de olivo (cv. Picual; 7 años de edad) con seis hojas cada una
procedente de olivos situados en los invernaderos de ETSIAAB-UPM que habían sido
tratada con la correspondiente solución de cada producto comercial o agua (control
negativo) utilizando pulverizadores manuales. Las ramitas habían de estar secas cuando
fueron ofrecidas al parasitoide (aproximadamente, 1 hora después de la aplicación). En
los ensayos de mortalidad se evaluaron cinco repeticiones por tratamiento. Los efectos de
los plaguicidas se estudiaron a las 2, 4, 24, 48 y 72 horas después del tratamiento (HAT).
Después de 72 horas, se eligieron 25 hembras en total (cinco hembras por repetición) en
aquellos cuyo ratio de supervivencia fue mayor del 50% para estudiar los efectos
subletales en la reproducción durante 5 días. Los datos de la primera parasitación se
rechazaron porque pueden ser muy erráticos debido a que las hembras necesitan al menos
un día para aprender a parasitar.
Durante cinco días consecutivos, y 1 hora por día, se ofrecieron larvas de tercer estadío
larvario de C. capitata a hembras de P. concolor en una proporción de 20 larvas por
hembra. Las larvas expuestas a la parasitación se transfirieron a una placa Petri para su
pupación y se evaluaron los porcentajes de huéspedes atacados y descendencia
aproximadamente un mes después de la parasitación. La caja de parasitación es similar
a la descrita en los test de mortalidad , pero con un agujero adicional (5.5 cm de diámetro)
en la parte inferior de la caja cubierta con una malla. Las larvas fueron ofrecidas a P.
concolor en un cuadrado de Parafilm® (10,2 cm2) localizado en la parte alta del bote de
plástico sujeto con dos gomas a la caja de parasitación con el fin de inmovilizar a las
larvas (ver Bengochea et al., 2014 para más detalles).
Ensayo de ingestión
En este experimento, cada unidad experimental estuvo formada por una caja que
contenía un recipiente de cristal (2 ml) con la correspondiente solución de cada plaguicida
y dieta sólida localizada en un tapón de plástico y suministrada ad libitum. No se
introdujeron ramas de olivo. Se utilizaron diez hembras de menos de 24 horas por

xliv
repetición y 5 repeticiones por tratamiento. La mortalidad y la reproducción se evaluaron
como se ha descrito en el ensayo previo.
Análisis de los datos
Los porcentajes de mortalidad no cumplieron las premisas del análisis ANOVA y se
aplicó un análisis no paramétrico de Kruskal-Wallis test, con una comparación dos a dos.
Los huéspedes atacados y la descendencia se compararon con un análisis de varianza
ANOVA y el test LSD. Los análisis estadísticos se realizaron con el paquete estadístico
IBM-SPSS, version 24.0 (SPSS Inc. IBM Company, 2016). La distribución normal de los
resíduos y la homogeneidad de varianzas se evaluó mediante los análisis de Kolmogorov–
Smirnov y Levene, respectivamente. Todos los análisis se realizaron con un nivel de
significación del 5 %. Los datos se muestran en las tablas 6.5., 6.6. y 6.7. (valores medios
± ES).
Posteriormente, los datos se interpretaron de acuerdo con las categorías de la OILB
(Sterk et al., 1999): 1, inócuo (< 30%); 2, ligéramente tóxico (30-79%); 3,
moderadamente tóxico (80-99%); y 4, tóxico (>99%) para los ensayos de laboratorio
(exposición vía ingestión) y como 1, inócuo (< 25%); 2, ligeramente tóxico (25-50%); 3,
moderadamente tóxico (51-75%); y 4 = tóxico (>75%) para ensayos de laboratorio
extendido (cuando las hembras se expusieron a residuos en hojas de olivo tratadas).
RESULTADOS Y DISCUSIÓN
Tras revisar la literatura disponible se puede concluir que:
1) En general, fungicidas, herbicidas y biopesticidas tienen menos efectos en la
mortalidad y en los parámetros reproductivos que los insecticidas, aunque algunos
fungicidas son muy tóxicos para P. concolor. Aunque hay una variabilidad significativa
en la respuesta a los insecticidas por parte del parasitoide, hay muchas evidencias de que
los insecticidas neurotóxicos son más dañinos que otros compuestos como los reguladores
del crecimiento de los insectos. De hecho, los compuestos más utilizados contra la mosca
del olivo, organofosforados y piretroides, son bastante a menudo muy tóxicos para las
hembras de P. concolor. Además, aunque no ha sido muy estudiado, se sabe que los
insecticidas neurotóxicos afectan al ratio sexual de la progenie y causa un incremento
significativo en la proporción de machos de la mayoría de las especies de bracónidos. Tal
y como describieron Biondi et al., (2013), puesto que los bracónidos tienen
partenogénesis arrenotóquica (huevos no fértiles producen solo machos) posiblemente el

xlv
mayor número de machos en la progenie se deba a la esterilidad de los mismos o a
alteraciones del comportamiento reproductor de las hembras de los parasitoides una vez
expuestos a insecticidas neurotóxicos.
2) A pesar de que el impacto de los fungicidas en P. concolor es menor que el de los
insecticidas, también deberían ser estudiados porque causan efectos relevantes cuando se
aplican de forma foliar. No hay datos, por otra parte, del uso de mezclas
fungicidas+insecticidas.
3) La mayoría de los estudios han sido llevados a cabo en laboratorio (bajo
“condiciones estandarizadas”), con diferentes vías de exposición (residual, ingestión…)
y con criterios de evaluación que incluían únicamente la mortalidad después de tres días
(efectos letales) y efectos en la reproducción (porcentaje de huéspedes atacados y
descendencia) (efectos subletales). Sin embargo, también pueden ocurrir efectos nocivos
en la longevidad, aprendizaje, comportamiento, neurofisiología, fisiología e inmunología
(Desneux, Decourtye and Delpuech, 2007; Talebi, Kavousi and Sabahi, 2008).
4) La mayoría de los estudios de laboratorio consideran el efecto de un único producto
fitosanitario, pero no los efectos sinérgicos de varios plaguicidas que pueden utilizarse al
mismo tiempo en campo.
5) Los efectos de la persistencia de los productos fitosanitarios no ha sido apenas
explorada, salvo raras excepciones.
6) Los resultados de los ensayos de laboratorio deberían ser completados con datos
de campo y apenas hay referencias bibliográficas en este sentido.
Estos estudios ecotoxicológicos asociados a los riesgos de los productos fitosanitarios
deberían ayudar a implementar cambios en los modelos de aplicación (por ejemplo, el
paso de compuestos de amplio espectro con dosis altas a compuestos selectivos utilizados
a dosis más bajas) (Amano and Haseeb, 2001). Además, estos estudios permitirán la
optimización del momento de liberación de los parasitoides tras las aplicaciones de
fitosanitarios (cuando se maximice la supervivencia de los mismos) y dará lugar a un
control sostenible, estableciendo como premisa previa el uso de los plaguicidas menos
tóxicos para P. concolor como un pre-requisito para el control de B. oleae.
REFERENCES

xlvi
Amano, H. and Haseeb, M. (2001). Recently-proposed methods and concepts of testing
the effects of pesticides on the beneficial mite and insect species: study limitations
and implications in IPM. Applied Entomology and Zoology, 36 (1): 1-11.
Arrese, E.L. and Soulages, J.L. (2010). Insect Fat Body: Energy, Metabolism, and
Regulation. Annual Review of Entomology, 55(1): 207-225.
Bale, J.S. (2002). Insects and low temperatures: from molecular biology to distributions
and abundance. Philosophical Transactions of the Royal Society B: Biological
Sciences, 357(1423): 849-862.
Barrett M. and Schmidt, J.M. (1991). A comparison between the amino acid composition
of an egg parasitoid wasp and some of its hosts. Entomologia Experimentalis et
Applicata, 59(1): 29-41.
Benelli, G. and Canale, A. (2013). Do tephritid-induced fruit volatiles attract males of the
fruit flies parasitoid Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae)?
Chemoecology, 23(3):191-199.
Benelli, G., Giunti, G., Tena, A., Desneux, N., Caselli, A. and Canale, A. (2017). The
impact of adult diet on parasitoid reproductive performance. Journal of Pest Science,
90: 807-823.
Bengochea, P., Budia, F., Viñuela, E. and Medina, P. (2014). Are kaolin and copper
treatments safe to the olive fruit fly parasitoid Psyttalia concolor? Journal of Pest
Science, 87: 351-359.
Biondi, A., Zappalà, L., Stark, J.D. and Desneux, N. (2013). Do biopesticides affect the
demographic traits of a parasitoid wasp and its biocontrol services through sublethal
effects? Plos One, 8(9): e76548.
Blüthgen, N. and Fiedler, K. (2004). Preferences for sugars and amino acids and their
conditionality in a diverse nectar-feeding ant community. Journal of Animal
Ecology, 73: 155-166.
Blüthgen, N., Gottsberger, G. and Fiedler, K. (2004). Sugar and amino acid composition
of ant-attended nectar and honeydew sources from an Australian rainforest. Austral
Ecology, 29: 418-429.
Burke, C.J. and Waddell, S. (2011). Remembering nutrient quality of sugar in
Drosophila. Current Biology, 21: 746-750.
Carter, C., Shafir, S., Yehonatan, L., Palmer, R.G. and Thornburg, R. (2006). A novel
role for proline in plant floral nectars. Naturwissenschaften, 93: 72-79.
Cicero, L., Sivinski, J. and Aluja, M. (2012). Effect of host diet and adult parasitoid diet
on egg load dynamics and egg size of braconid parasitoids attacking Anastrepha
ludens. Physiological Entomology, 37: 177-184.
Coskun, M. and Emre, I. (2015). Role of Lipids, Amino Acids, and Sucrose on the Total
Adult and Female Emergence, and Content of Glycogen and Protein in Pimpla
turionellae (Hymenoptera: Ichneumonidae). Annals of the Entomological Society
of America, 108(5): 820-826.
Daane, K.M. and Johnson, M.W. (2010). Olive Fruit Fly: Managing an Ancient Pest in
Modern Times. Annual Review of Entomology, 55(1):151-169.
Desneux, N., Decourtye, A. and Delpuech J-M. (2007). The Sublethal Effects of
Pesticides on Beneficial Arthropods. Annual Review of Entomology, 52: 81-106.
DGAV (2018). Direção-Geral de Alimentação e Veterinária. Official phytosanitary
products.
http://www.dgav.pt/fitofarmaceuticos/guia/finalidades_guia/Insec&Fung/Culturas/oli
veira.htm and http://www.drapc.minagricultura.pt/drapc/servicos/
fitossanidade/files/herbicidas_olival_2017.pdf. Cited 10 September 2018 (In
Portugal).

xlvii
Ellers, J., Sevenster, J.G. and Driessen, G. (2000). Egg Load Evolution in Parasitoids.
The American Naturalist, 156(6): 650-665.
Faria, C.A., Wäckers, F.L. and Turlings, T.C.J. (2008). The nutritional value of aphid
honeydew for non-aphid parasitoids. Basic and Applied Ecology, 9: 286-297.
Giron, D. Pincebourde, S. and Casas, J. (2004). Lifetime gains of host-feeding in a
synovigenic parasitic wasp. Physiological Entomology, 29(5): 436-442.
Grenier, S. (2012). Artificial Rearing of Entomophagous Insects, with Emphasis on
Nutrition and Parasitoids - General Outlines from Personal Experience. Karaelmas
Science and Engineering Journal, 2 (2): 1-12.
Hagen, K.S., Greany, P., Sawall Jr., E.F. and Tassain, R.L. (1976). Tryptophan in
artificial honeydews as a source of an attractant for adult Chrysopa carnea.
Environmental Entomology 5(3): 458-468.
Hajirajabi, N., Fazel, M.M., Harvey, J.A., Arbab, A. and Asgari, S. (2016). Dietary sugars
and proline influence biological parameters of adult Trissolcus grandis, an egg
parasitoid of Sunn pest, Eurygaster integriceps. Biological Control, 96: 21-27.
Harvey, J.A., Cloutier, J., Visser, B., Ellers, J., Wäckers, F.L., and Gols, R. (2012). The
effect of different dietary sugars and honey on longevity and fecundity in two
hyperparasitoid wasps. Journal of Insect Physiology, 58(6): 816-823.
Hausmann, C.H., Wäckers, F.L. and Dorn, S. (2005). Sugar convertibility in the
parasitoid Cotesia glomerata (Hymenoptera: Braconidae). Archives of Insect
Biochemistry and Physiology, 60: 223-229.
Hawkes, N.J., Janes, R.W., Hemingway, J. and Vontas, J. (2005). Detection of resistance-
associated point mutations of organophosphate-insensitive acetylcholinesterase in
the olive fruit fly, Bactrocera oleae (Gmelin). Pesticide Biochemistry and
Physiology, 81(3), 154-163.
Hidalgo, K., Mouline, K., Mamai, W., Foucreau, N., Dabire, K.R., Bouchereau, A.,
Simard, F. and Renault, D. (2014). Novel insights into the metabolic and
biochemical underpinnings assisting dry-season survival in female malaria
mosquitoes of the Anopheles gambiae complex. Journal of Insect Physiology 70:
102-116.
Heimpel, G.E. and Rosenheim, J.A. (1998). Egg limitation in parasitoids: a review of the
evidence and a case study. Biological Control, 11(2): 160-168.
Human, H. and Nicolson, S.W. (2006). Nutritional content of fresh, bee-collected and
stored pollen of Aloe greatheadii var. davyana (Asphodelaceae). Phytochemistry,
67(14): 1486-1492.
Jacas, J.A. and Viñuela, E. (1994a). Side-effects of pesticides on Opius concolor Szèp.
(Hymenoptera:Braconidae), a parasitoid of the olive fruit fly. Bull. OILB/IOBC Bull.,
17(10): 143-146.
Jacas, J. A. and Viñuela, E. (1994b). Analysis of a laboratory method to test the effects
of pesticides on adult females of Opius concolor (Hym., Braconidae), a parasitoid of
the olive fruit fly, Bactrocera oleae (Dip., Tephritidae). Biocontrol Science and
Technology, 4: 147-154.
Kishinevsky, M., Cohen, N., Chiel, E., Wajnberg, E. and Keasar, T. (2017). Sugar feeding
of parasitoids in an agroecosystem: effects of community composition, habitat and
vegetation. Insect Conservation and Diversity, 11: 50-57.
Lee, J.C. and Heimpel, G.E. (2008). Floral resources impact longevity and oviposition
rate of a parasitoid in the field. Journal of Animal Ecology, 77: 565-572.
Leroy, P. D., Wathelet, B., Sabri, A., Francis, F., Verheggen, F. J., Capella. Q., Thonart,
P. and Haubruge, E. (2011). Aphid-host plant interactions: does aphid honeydew

xlviii
exactly reflect the host plant amino acid composition? Arthropod-Plant Interactions,
5(3): 193-199.
Li, Y., Zhang, L., Zhang, Q., Chen, H. and Denlinger, D.L. (2014). Host diapause status
and host diets augmented with cryoprotectants enhance cold hardiness in the
parasitoid Nasonia vitripennis. Journal of Insect Physiology, 70:8-14.
Loumou, A. and Giourga, C. (2003). Olive groves: The life and identity of the
Mediterranean. Agriculture and Human Values, 20(1): 87-95.
MAPAMA, 2018. Ministry of Agriculture and Fisheries Food and Environment. Official
phytosanitary products entry.
http://www.mapama.gob.es/es/agricultura/temas/sanidad-vegetal/productos-
fitosanitarios/registro/menu.asp Cited 25 May 2017 (In Spanish).
Micheu, S., Crailsheim, K., and Leonhard, B. (2000). Importance of proline and other
amino acids during honeybee flight. Amino Acids, 18(2), 157-175.
McEwen, P., Jervis, M.A. and Kidd, N.A.C. (1994). Use of sprayed L-tryptophan solution
to concentrate numbers of the green lacewing Chrysoperla carnea in olive tree
canopy. Entomologia Experimentalis et Applicata, 70(1):97-99.
Nave, A., Gonçalves, F., Nunes, F.M., Campos, M. and Torres, L. (2017a). Evaluating
potential sugar food sources from the olive grove agroecosystems for Prays oleae
parasitoid Chelonus elaeaphilus. Biocontrol Science and Technology, 27: 686-695.
Nave, A., Crespí, A.L., Gonçalves, F., Campos, M. and Torres, L. (2017b). Native
Mediterranean plants as potential food sources for natural enemies of insect pests in
olive groves. Ecological Research, 32(4): 459-459.
Nicolson S.W and Thornburg R.W. (2007). Nectar Chemistry. In., Nicolson, M. Nepi &
E. Pacini, (eds.), Nectaries and Nectar. Springer, Dordrecht. pp. 215-263.
Pavlidi, N., Kampouraki, A., Tseliou, V., Wybouw, N., Dermauw, W., Roditakis, E.,
Nauen, R. Van Leeuwen, T., Vontas, J. (2018). Molecular characterization of
pyrethroid resistance in the olive fruit fly Bactrocera oleae. Pesticide Biochemistry
and Physiology, 148: 1-7.
Petanidou, T. (2005). Sugars in Mediterranean floral nectars: an ecological and
evolutionary approach. Journal of Chemical Ecology, 31: 1065-1088.
Petanidou, T., Van Laere, A., Ellis, W.N. and Smets, E. (2006). What shapes amino acid
and sugar composition in Mediterranean floral nectars? Oikos, 115: 155-169.
Pinheiro, L.A., Medina, P., Bento, A., Pereira, J.A and Santos, S.A.P. 2017. Bioecology
of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and Management
Strategies to Increase Biological Control of the Olive Fruit Fly. In Natural Enemies:
Identification, Protection Strategies and Ecological Impacts, ed. Santos, S.A.P. 141-
172. New York: Nova Science Publishers.
R core team 2014. R: a language and environment for statistical computing. R Foundation
for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/
Rivero, A. and Casas, J. (1999) Incorporating physiology into parasitoid behavioral
ecology: the allocation of nutritional resource. Research in Population Ecology,
41(1): 39-45.
Ruiz-Torres M. and Montiel-Bueno A. (2007). Efecto de los tratamientos-cebo aereos
con spinosad contra mosca del olivo (Bactrocera oleae, Gmel.; Diptera: Tephritidae)
sobre la entomofauna del olivar en la provincia de Jaén. Boletín de
sanidad vegetal. Plagas, 33: 267-284.
Segoli, M. and Rosenheim, J.A. (2013). Spatial and temporal variation in sugar
availability for insect parasitoids in agricultural fields and consequences for
reproductive success. Biological Control, 67: 163-169.

xlix
SPSS Inc., IBM Company (2016) IBM, SPSS statistic for windows, version 24.0. New
York, USA.
Soares, M.E.; Pereira, J.A. and Bastos, M.L. (2004). Copper residues in olives after olive
tree treatments with three different copper formulations. 2nd International IUPAC
Symposium Trace Elements in Food (TEF 2). Bruxelas (Bélgica), 7-8 October 2004.
Book of Abstracts Ed. By Kortsen, B.; Bickel, M.; Grobecker, K.-H. P. 92.
Sterk, G. et al., (1999). Results of the Seventh Joint Pesticide Testing Programme Carried
Out by the IOBC/WPRSWorking Group “Pesticides and Beneficial Organisms”.
BioControl, Vol.44, No.1, (March 1999), pp.99-117
Talebi, K., Kavousi A. and Sabahi Q. (2008). Impacts of Pesticides on Artropod
Biological Control. Pest Technology, 2(2): 87-97.
Tena, A., Pekas, A., Cano, D., Wäckers, F.L. and Urbaneja, A. (2015). Sugar provisioning
maximizes the biocontrol service of parasitoids. Journal of Applied Ecology, 52:
795-804.
Tena, A., Wäckers, F.L., Heimpel, G.E., Urbaneja, A. and Pekas, A. (2016). Parasitoid
nutritional ecology in a community context: the importance of honeydew and
implications for biological control. Current Opinion in Insect Science, 14: 100-104.
Tena, A., Senft, M., Desneux, N., Dregni, J. and Heimpel, G.E. (2018). The influence of
aphid-produced honeydew on parasitoid fitness and nutritional state: a comparative
study. Basic and Applied Ecology, 29: 55-68.
Therneau T (2014). _A Package for Survival Analysis in S_. R package version 2.37-7,
URL: http://CRAN.R-project.org/package=survival.[accesed: 11/08/2016].
Therneau T.M., Grambsch P.M. (2000) Modeling Survival Data: Extending the Cox
Model. Statistics for biology and health. Springer. USA.
Tompkins, J.M.L., Wratten, S.D. and Wäckers, F.L. (2010). Nectar to improve parasitoid
fitness in biological control: does the sucrose:hexose ratio matter? Basic and Applied
Ecology, 11: 264-271.
Villa, M., Santos, S.A.P., Mexia, A., Bento, A. and Pereira, J.A. (2017). Wild flower
resources and insect honeydew are potential food items for Elasmus flabellatus.
Agronomy for Sustainable Development, 37:15.
Wäckers, F.L. (1999). Gustatory response by the hymenopteran parasitoids Cotesia
glomerata to a range of nectar and honeydew sugars. Journal of Chemical Ecology,
25(12): 2863-2877.
Wäckers, F.L. (2000). Do oligosaccharides reduce the suitability of honeydew for
predators and parasitoids? A further facet to the function of insect-synthesized
honeydew sugars. Oikos, 90: 197-201.
Wäckers, F.L. (2001). A comparison of nectar- and honeydew sugars with respect to their
utilization by the hymenopteran parasitoid Cotesia glomeratae. Journal of Insect
Physiology, 47: 1077-1084.
Wäckers, F. L. (2005). Suitability of (extra-)floral nectar, pollen, and honeydew as insect
food sources. F.L. Wäckers, P.C.J. van Rijn and J. Bruin (Eds.). Plant-Provided Food
for Carnivorous Insects, A Protective Mutualism and its Applications (pp. 17-74).
Cambridge: Cambridge University Press.
Wäckers, F.L., Lee, J., Heimpel, G., Winkler, K. and Wagenaar, R. (2006).
Hymenopteran parasitoids synthesize honeydew-specific’ oligosaccharides.
Functional Ecology, 20: 790-798.
Wäckers, F.L., van Rijn, P.C.J. and Heimpel, G.E. (2008). Honeydew as a food source
for natural enemies: Making the best of a bad meal? Biological Control: 45, 176-184.

l
Wade, M.R., Hopkinson, J.E. and Zalucki, M.P. (2008a). Influence of food
supplementation on the fitness of two biological control agents: a predatory nabid
bug and a bollworm pupal parasitoid. Journal of Pest Science, 81: 99-107.
Wade, M.R., Zalucki, M.P., Wratten, S.D. and Robinson, K.A. (2008b). Conservation
biological control of arthropods using artificial food sprays: current status and future
challenges. Biological Control, 45: 185-199.
Walters, A.S. (2011). Weed management systems for no-tillage vegetable production. In
Soloneski S. and Larramendy M.L. (Eds.), Herbicides: theory and applications (pp.
2-40). Rijeka: IntechOpen.
Wang, X.-G., Johnson, M.W., Opp, S.B., Krugner, R. and Daane, K.M.
(2011). Honeydew and insecticide bait as competing food resources for a fruit fly
and common natural enemies in the olive agroecosystem. Entomologia
Experimentalis et Applicata, 139: 128-137.
Williams III, L., Deschodt, P., Pointurier, O. and Wyckhuys, K.A.G. (2015). Sugar
concentration and timing of feeding affect feeding characteristics and survival of a
parasitic wasp. Journal of Insect Physiology, 79: 10-18.
Winkler, K., Wackers, F.L., Stingli, A. and van Lenteren, J.C. (2005). Plutella xylostella
(diamondback moth) and its parasitoid Diadegma semiclausum show different
gustatory and longevity responses to a range of nectar and honeydew sugars.
Entomologia Experimentalis et Applicata, 115(1):187-192.
Winkler, K., Wackers, F.L., Bukovinszkine-Kiss, G. and van Lenteren, J.C. (2006).
Nectar resources are vital for Diadegma semiclausum fecundity under field
conditions. Basic and Applied Ecology, 7(2): 133-140.
Zhang, Y., Yang, N., Wang, J. and Wan, F. (2014). Effect of six carbohydrate sources on
the longevity of a whitefly parasitoid Eretmocerus hayati (Hymenoptera:
Aphelinidae). Journal of Asia-Pacific Entomology, 17: 723-728.

li
PHOTOGRAPHS OF THE SPECIES STUDIED IN THIS THESIS
The loving mother
Psyttalia concolor
# The parasitoid
The baby wasp home
Larvae of Bactrocera oleae
# The victim

52
Chapter 1
Introduction

53
Chapter 1: GENERAL INTRODUCTION
1. INTRODUCTION
Olive growing is an important land-use in the Mediterranean region, with significant
environmental, social and economic roles (Loumou and Giourga, 2003). However, the
olive tree is attacked by diverse pests that reduce the yield, the olive oil quality and
growers’ income. Among those pests, the olive fruit fly, Bactrocera oleae (Rossi)
(Diptera: Tephritidae), is the major pest of commercial olives worldwide (Daane and
Johnson, 2010). This pest is primarily controlled by synthetic pesticides, which have
considerable drawbacks for the quality of environment and human health as well as
notably negative effects on biodiversity and biological control potential. Furthermore,
after chemical application, severe out-breaks of the pest population may occur due to the
reduction of its natural enemies or to the development of insecticide resistance (Soares,
Pereira and Bastos, 2004; Hawkes et al., 2005). During the last few years, B. oleae has
gained economic importance in the northern part of Portugal, where the risk of attack
caused by this pest was traditionally low. This increase in the level of pest attack is an
aspect of main concern. In sustainable olive groves, olive fruit fly control should be
achieved by maximizing the action of its natural enemies. In this context, innovative
aspects deriving from pest-parasitoid interactions are needed and should be explored.
Among the complex of natural enemies, the braconid wasp Psyttalia concolor (Szépligeti)
(Hymenoptera: Braconidae) is considered an important endoparasitoid of the olive fruit
fly as well as of other tephritids of economic importance (Benelli and Canale, 2013).
Furthermore, in laboratory, P. concolor is easily mass reared on an alternative host, the
medfly Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). As consequence, plant-
pest interactions have been studied in order to establish the parasitoid in the olive grove
and the ability to complete its life cycle. Adult parasitoids require floral resources (pollen
and nectar) and honeydew to fulfill its energetic needs and for the reproductive system
maturation (Winkler et al., 2006). Thus, conservation or enhancement of P. concolor
populations hinges on the maintenance of non-crop plants (e.g. vegetation strips and
bushes) within or around the olive groves. This involves the study of the effect of non-
crop plants as sources of supplementary resources (sugars, honeydew and amino acids),
on survival, reproduction and behaviour of P. concolor, and understand how they
contribute to the conservation of the parasitoid in the olive grove. Understanding these

Chapter 1: General introduction
54
interactions ultimately provide information to developing integrated techniques for
sustainable pest control.
1.1. Dietary requirements
To life parameters (e.g., longevity, maintain oogenesis) and foraging activity, adult
parasitoids need to obtain different nutrients, such as carbohydrates, proteins, amino
acids, lipids, vitamins and minerals (Lundgren, 2009). The nutrients of the diet should be
properly balanced, especially in the ratio of proteins (amino acids)/carbohydrates (Panizzi
and Parra, 2012). In the agroecosystems, the available composition of food source
influences the food dietary and parasitoid fitness. The majority of adult parasitoid explore
a range of sugary-rich foods such as (extra-)floral nectars and honeydew (e.g., Wäckers,
2001; 2005; Tena et al., 2015). Similarly, pollen provide nutrients that benefit parasitoid
fitness (Zhang et al. 2004). To locate these food resources, parasitoids use visual (e.g
colors) and tactile stimuli (e.g. shapes), chemical cues (e.g., scents or plants infested by
herbivores emit specific volatiles) or its combination, which can be innate attractive
stimuli or learned ones (Wäckers, 1994; Giunti et al. 2015).
1.1.1. Carbohydrates
Carbohydrates are the main source of energy, being used for the synthesis of fats and
glycogen (Yamada et al., 2018). Moreover, sugar-rich foods are indicative of parasitoid
nutritional status (Fadamiro and Heimpel, 2001; Lee, Heimpel and Leibee, 2004). The
suitability of carbohydrates as a source of energy depends on detectability, accessibility
and efficacy by which the sugars can be converted into energy (Wäckers, 2000). The
concentration of sugar in nectar vary considerably, normally ranging between 20% and
40% on most plants, mainly consisting in three main nectar sugars: one disaccharide
(sucrose) and two monosaccharides (fructose and glucose) (Wäckers, 2005). However,
some nectar sugars can be toxic for parasitoids (Wäckers, 2001).
Longevity studies indicate that distinct sugars increased parasitoid longevity (e.g.,
Wäckers, 2001; Kishinevsky et al., 2017). The effect of sugars and honeydew for
parasitoid P. concolor are studied in chapter 4. The experiments demonstrated the
potential for carbohydrates to positively affect biological control.

Chapter 1: General introduction
55
1.1.2. Amino acids
Pollen is rich in amino acids described as essential for development or reproduction of
parasitoids (Wäckers, 2005). Amino acids (essential and non-essential amino acids and
some non-protein amino acids) are the most abundant nectar solutes after sugars
(Petanidou et al., 2006). Furthermore, proline and phenylalanine are the most abundant
nectar amino acids (Petanidou, 2007). According to Nepi (2014), non-protein amino acids
in nectar benefit parasitoid in different ways: affecting the physiology of the nervous
system of the insect, regulating nectar intake through phagostimulation and promoting
muscle function during flight. In synovigenic parasitoids, the importance of amino acids
sources for egg production and survival seems to vary from species to species (Rivero
and Casas, 1999). For parasitoid P. concolor, the effects are discussed in Chapter 5.
2. IMPACTS OF CLIMATE CHANGE ON CROP-PEST-PARASITOID
INTERACTION: IMPLICATIONS ON BIOLOGICAL CONTROL
The response of food safety and biodiversity to under different scenarios of global
climatic change has been an active subject at research (Miraglia et al., 2009). Climate
changes frequency increase crop losses (by impacting crop growth and production) due
to climatic variability and frequency of extreme events (such as droughts and floods) or
changes in precipitation, temperature and CO2 concentration (Olesen and Bindi, 2002;
Altieri et al., 2015; Johnson and Züst, 2018). It is expected that this change affect agro-
ecosystems in several ways, including impact on crop-pest-natural enemy interaction,
disease systems and induced changes in vegetation (Chapin III et al., 2000; Bellard et al.,
2012).
Temperature and available moisture are two of the most important abiotic variables
limiting the distribution and abundance of many species. Increased temperature and/or
precipitation can potentially affect (positively or negatively) insect survival, reproduction
development, change gender ratios, existence of hosts, population size and geographic
range (Kambrekar et al., 2015). For example, increased temperatures can accelerate the
development of a pest, resulting in more generations per year, increasing crop damage
and consequently leading to higher costs on crop protection (Kambrekar et al., 2015).
Several models were already performed to predict the impact of climate change on
biodiversity, the majority indicating alarming consequences on biodiversity (see the
review Bellard et al., 2012).

Chapter 1: General introduction
56
Although Mediterranean Basin have an ideal climatic agroecosystem for the olive tree
(Olea europaea L.), the climate changes have an important impact not only on this culture
and on olive fruit fly (Bactrocera oleae), but also on its interaction (Ponti et al., 2014).
Temperature interfere with some aspects of olive biology, such as rates of development,
photosynthesis, respiration, subunit production rates (leaves, stem, root) initiation,
maximum number of flower buds, period of flowering and fruit maturation, or even cause
its death (Gutierrez, Ponti and Cossu, 2009). Mortality of immature stages and adults of
olive fruit fly is also promoted by temperature, which is also responsible for a reduction
of its reproduction and an induction of reproductive dormancy (Gutierrez, Ponti and
Cossu, 2009; Ponti, Cossu and Gutierrez, 2009). The models used to analyse the effects
of climate change on olive tree and olive fly in the Mediterranean Basin showed different
tolerances to temperature (Ponti et al., 2014). Gutierrez, Ponti and Cossu (2009) and more
specifically that olive tree has a much wider temperature range of favourability than olive
fly. As an example, an average climate warming of 1.8 °C will benefit some olive-
producing areas, adversely affect others, and some will remain relatively unchanged
(Ponti et al., 2014). According to Ponti et al., 2014, these changes might lead to declines
of 18% for the Iberian Peninsula, 21% for Italy and France, 23% for Greece, Turkey, and
the Balkans, 2% for North Africa and 80% for the Middle East. (Ponti et al., 2014).
Moreover, according to model of Gutierrez, Ponti and Cossu (2009) in California is
expected that olive expand northward and along coastal areas due to global warming in
southern desert areas. Consequently, it is expected an increase on the range of olive fly
on these areas Currently in Italy, only northern regions and the mountains of central Italy
are unfavourable to olive and olive fruit fly, but it is expected that unfavourable areas
(e.g. colder areas at higher elevation) become favourable and increased the range of olive
fruit fly due to climate warming (Gutierrez, Ponti and Cossu, 2009; Ponti, Cossu
Gutierrez, 2009). The model assumes that, while geographic distribution of olive bloom
phenology and yield will by affected by climate warming, water availability will only
affect this culture in very severe conditions (Gutierrez, Ponti and Cossu, 2009). These
changes anticipate negative consequences on rural development and poverty alleviation
in marginal areas across (Ponti et al., 2014).
Global warming influences also parasitoids fitness, activity and parasitoid-hosts
interaction, as well as food source of parasitoids (e.g. biology of flowers: timing and
abundance of flowering) (Stange and Ayres, 2010). For example, Wetherington et al.
(2017) found that climate variation alters the synchrony of host-parasitoid interactions

Chapter 1: General introduction
57
between Oobius agrili Zhang and Huang (Hymenoptera: Encyrtidae) and solitary egg
parasitoid of emerald ash borer (EAB), Agrilus planipennis Fairmaire (Coleoptera:
Buprestidae). The success of parasitoids used in biological programs depends of
understanding the consequences of climate variability (low or high temperatures) on crop-
pest-parasitoid interaction.
Parasitoids adults have morphological adaptations to winter conditions such as
variability in their colour morphs (clear individuals are reflective whereas dark colours
contribute to more efficient heat absorption). Also, in case of koinobiont parasitoids, it
conditions the mobility of the parasitized host, leading it to locations where survival is
more probable. Considering idiobiont parasitoids, it affects the distribution of oviposition
sites to decrease the expected stochastic mortality (Hance et al., 2007). Additionally,
some parasitoids diapause as pupae became more resistant to variable winter conditions,
while parasitoids immatures have a set of strategies to protect against freezing not only
themselves but also the body of their hosts (Hance et al., 2007).
A higher metabolic activity of parasitoids has been registered under higher
temperatures, possibly being more receptive to plant-release chemicals (Yuan et al.,
2009). Plants volatiles depends on plant tissues (e.g. leaves and flowers), varieties and
cultivars and on plant signals emitted by the presence of herbivore insect or by damaged
tissues) (Yuan et al., 2009). The capacity of parasitoids to distinguish signals of host
from non-host on different plants implies a level of behavioural plasticity of parasitoids
that involves learning and context-dependent analysis of chemical cues (De Moraes et al.,
1998). The success of parasitism depends of (1) capacity to adult parasitoids to locate and
evaluate its host and (2) the larva ability to overcome the host immune response and adapt
to a constantly changing host environment, which could be compromised by changes in
environmental conditions (Hance et al., 2007). It is possible that, with higher
temperatures, the parasitoids exert a strong effect on their hosts even when the
development time of the host is shortened and reduced the timeframe of hosts to attack
(Stange and Ayres, 2010). Moreover, it has an impact on population dynamics and the
distribution of host/parasitoids enables the extinction of one or both species and
impossibility of release of the parasitoids. Also, alterations on parasitism rates have
higher implication for parasitoids specialists, whose larvae feed exclusively on a single
arthropod pest.

Chapter 1: General introduction
58
Overall, the climate change could affect the pest-parasitoid interaction and have
implications for consumer-floral resources by parasitoids and consequently the
effectiveness of biological control.
3. REFERENCES
Altieri, M.A., Nicholls, C.I., Henao, A. and Lana, M.A. (2015). Agroecology and the
design of climate change-resilient farming systems. Agronomy for Sustainable
Development, 35(3): 869-890.
Bellard, C., Bertelsmeier, C., Leadley, P., Thuiller, W. and Courchamp, F. (2012).
Impacts of climate change on the future of biodiversity. Ecology Letters, 15(4): 365-
377.
Benelli, G. and Canale, A. (2013). Do tephritid-induced fruit volatiles attract males of the
fruit flies parasitoid Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae)?
Chemoecology, 23(3):191-199.
Chapin III, F.S., Zavaleta, E.S., Eviner, V.T., Naylor, R.L., Vitousek, P.M., Reynolds,
H.L., Hooper, D.U., Lavorel, S., Sala, O.E., Hobbie, S.E., Mack, M.C. and Díaz, S.
(2000). Consequences of changing biodiversity. Nature, 405: 234-242.
Daane, K.M. and Johnson, M.W. (2010). Olive Fruit Fly: Managing an Ancient Pest in
Modern Times. Annual Review of Entomology, 55(1):151-169.
De Moraes, C.M., Lewis, W.J., Paré, P.W., Alborn, H.T. and Tumlinson, J.H. (1998).
Herbivore-infested plants selectively attract parasitoids. Nature, 393(6685): 570-573.
Fadamiro, H.Y. and Heimpel, G.E. (2001). Effects of partial sugar deprivation on lifespan
and carbohydrate mobilization in the parasitoid Macrocentrus grandii
(Hymenoptera: Braconidae). Annals of the Entomological. Society of America, 94:
909-916.
Giunti, G., Canale, A., Messing, R.H., Donati, E., Stefanini, C., Michaud, J.P. and
Benelli, G. (2015). Parasitoid learning: Current knowledge and implications for
biological control. Biological Control, 90: 208-219.
Gutierrez, A.P., Ponti, L. and Cossu, Q.A. (2009). Effects of climate warming on Olive
and olive fly (Bactrocera oleae (Gmelin)) in California and Italy. Climatic Change,
95(1-2), 195-217.
Hance, T., van Baaren, J., Vernon, P. and Boivin, G. (2007). Impact of Extreme
Temperatures on Parasitoids in a Climate Change Perspective. Annual Review of
Entomology, 52(1), 107-126.
Hawkes, N.J., Janes, R.W., Hemingway, J. and Vontas, J. (2005). Detection of resistance-
associated point mutations of organophosphate-insensitive acetylcholinesterase in
the olive fruit fly, Bactrocera oleae (Gmelin). Pesticide Biochemistry and
Physiology, 81(3), 154-163.
Johnson, S.N. and Züst, T. (2018). Climate Change and Insect Pests: Resistance Is Not
Futile? Trends in Plant Science, 23(5): 367-369.
Kambrekar, D.N., Guledgudda, S.S., katti, A. and Mohankumar (2015). Impact of climate
change on insect pests and their natural enemies. Journal of Agricultural Science, spl.
28(5): 814-816.
Kishinevsky, M., Cohen, N., Chiel, E., Wajnberg, E. and Keasar, T. (2017). Sugar feeding
of parasitoids in an agroecosystem: effects of community composition, habitat and
vegetation. Insect Conservation and Diversity, 11(1): 50-57.

Chapter 1: General introduction
59
Lee, J.C., Heimpel, G.E. and Leibee, G.L. (2004). Comparing floral nectar and aphid
honeydew diets on the longevity and nutrient levels of a parasitoid wasp.
Entomologia Experimentalis et Applicata 111: 189–199.
Loumou, A. and Giourga, C. (2003). Olive groves: The life and identity of the
Mediterranean. Agriculture and Human Values, 20(1): 87-95.
Lundgren, J. (2009). Relationships of natural enemies and non-prey foods. Springer
International, Dordrech.
Miraglia, M. et al. (2009). Climate change and food safety: An emerging issue with
special focus on Europe. Food and Chemical Toxicology, 47(5): 1009-1021.
Nepi, M. (2014). Beyond nectar sweetness: the hidden ecological role of non-protein
amino acids in nectar. Journal of Ecology, 102(1): 108-115.
Olesen, J.E. and Bindi, M. (2002). Consequences of climate change for European
agricultural productivity, land use and policy. European Journal of Agronomy, 16(4):
239-262.
Parra, J.R.P (2012). The Evolution of Artificial Diets and Their Interactions in Science
and Technology. In Panizzi, A.R. and Parra, J.R.P. (eds.). Insect Bioecology and
Nutrition for Integrated Pest Management. CRC Press - Taylor & Francis Group,
Boca Raton, pp 51-92.
Petanidou, T. (2007) Ecological and evolutionary aspects of floral nectars in
Mediterranean habitats. In Nicolson, S.W. Nepi, M. and Pacini, E. (eds). Nectaries
and nectar. Springer, Dordrecht, pp. 343-375.
Petanidou, T., van Laere, A., Ellis, W.N. and Smets, E. (2006). What shapes amino acid
and sugar composition in Mediterranean floral nectars? Oikos, 115: 155-169.
Ponti, L., Gutierrez, A.P., Ruti, P.M. and Dell’Aquila, A. (2014). Fine-scale ecological
and economic assessment of climate change on olive in the Mediterranean Basin
reveals winners and losers. Proceedings of the National Academy of Sciences,
111(15): 5598-5603.
Ponti, L., Cossu, Q.A. and Gutierrez, A.P. (2009). Climate warming effects on the Olea
europaea-Bactrocera oleae system in Mediterranean islands: Sardinia as an example.
Global Change Biology, 15(12): 2874-2884.
Rivero, A. and Casas, J. (1999). Incorporating physiology into parasitoid behavioral
ecology: the allocation of nutritional resources. Researches on Population Ecology,
41(1), 39-45.
Soares, M.E.; Pereira, J.A. and Bastos, M.L. (2004). Copper residues in olives after olive
tree treatments with three different copper formulations. 2nd International IUPAC
Symposium Trace Elements in Food (TEF 2). Bruxelas (Bélgica), 7-8 October 2004.
Book of Abstracts Ed. By Kortsen, B.; Bickel, M.; Grobecker, K.-H. P. 92.
Stange, E.E. and Ayres, M.P. (2010). Climate Change Impacts: Insects. In: Encyclopedia
of Life Sciences (ELS). John Wiley and Sons, Ltd: Chichester.
Wäckers, F.L. (1994). The effect of food deprivation on the innate visual and olfactory
preferences in the parasitoid Cotesia rubecula. Journal of Insect Physiology, 40(8),
641-649.
Wäckers, F.L. (2000). Do oligosaccharides reduce the suitability of honeydew for
predators and parasitoids? A further facet to the function of insect-synthesized
honeydew sugars. Oikos, 90(1): 197-201.

Chapter 1: General introduction
60
Wäckers, F.L. (2001). A comparison of nectar- and honeydew sugars with respect to their
utilization by the hymenopteran parasitoid Cotesia glomerata. Journal of Insect
Physiology, 47(9):1077-1084.
Wäckers, F.L. (2005). Suitability of (extra-) floral nectar, pollen, and honeydew as insect
food sources. In: Wäckers, F.L., van Rijn, P.C.J. and Bruin, J. (eds). Plant-provided
food for carnivorous insects: A protective mutualism and its applications. Cambridge
University, Cambridge, pp 17-74.
Wetherington, M.T., Jennings, D.E., Shrewsbury, P.M. and Duan, J.J. (2017). Climate
variation alters the synchrony of host-parasitoid interactions. Ecology and Evolution,
7(20): 8578-8587
Winkler, K., Wackers, F.L., Bukovinszkine-Kiss, G. and van Lenteren, J.C. (2006).
Nectar resources are vital for Diadegma semiclausum fecundity under field
conditions. Basic and Applied Ecology, 7(2): 133-140.
Yamada, T., Habara, O., Kubo, H. and Nishimura, T. (2018). Fat body glycogen serves
as a metabolic safeguard for the maintenance of sugar levels in Drosophila.
Development, 145(6), dev158865.
Yuan, J.S., Himanen, S.J., Holopainen, J.K., Chen, F. and Stewart, C.N. (2009). Smelling
global climate change: mitigation of function for plant volatile organic compounds.
Trends in Ecology and Evolution, 24(6): 323-331.
Zhang, G., Zimmermann, O. and Hassan, S.A. (2004). Pollen as a source of food for egg
parasitoids of the genus Trichogramma (Hymenoptera: Trichogrammatidae).
Biocontrol Science and Technology, 14(2): 201-209.

61
Chapter 2
Objectives

62
Chapter 2:
OBJECTIVES
The olive fruit fly, Bactrocera oleae, is the main pest of the olive tree worldwide. In
Trás-os-Montes, the levels of pest attack have increased significantly in the last years. To
face this, the action of its natural enemies should be promoted under a sustainable pest
control system. Among the complex of natural enemies, the braconid wasp Psyttalia
concolor is an important endoparasitoid of the pest. Although, for an effective pest
control, several factors that affect the success of the parasitoid need to be clarified. Thus,
the aim of this work will be to study the different components of the interaction P.
concolor-olive fruit fly as well as the effect of non-crop plants on the fitness of P.
concolor, in order to, promote biological control in olive groves. To accomplish this aim,
the working program is organized in three parts with different objectives:
Literature Review: “BIOECOLOGY OF PSYTTALIA CONCOLOR (SZÉPLIGETI)
(HYMENOPTERA: BRACONIDAE) AND MANAGEMENT STRATEGIES TO INCREASE
BIOLOGICAL CONTROL OF THE OLIVE FRUIT FLY” (Chapter 3).
To study the effect of spontaneous non-crop plants, as sources of supplementary
resources, on survival, reproduction and behaviour of P. concolor and understand how
they attract and contribute to the conservation of the parasitoid in the olive grove;
Laboratory experiments: P. concolor specimens will be tested with:
• Sugars: the monosaccharides, glucose and fructose, the disaccharides, sucrose,
trehalose and melibiose and the trisaccharide, melezitose, as well as, the sugar
alcohol, sorbitol (Chapter 4);
• Honeydews: obtained from two olive pests, the black-scale, Saissetia oleae, and
the olive psyllid, Euphyllura olivine (Chapter 4);
• Amino acids: tryptophan, phenylalanine and proline were tested singly and mixed
with glucose (Chapter 5);

Chapter 2: Objectives
63
To evaluate the susceptibility of the adults of Psyttalia concolor, to some fresh
residues of insecticides, pesticides and herbicides (used in Mediterranean olive groves),
under conditions of maximum exposure (inert substrate) on plant and under conditions of
laboratory, following the standardized method proposed by the OILB (Chapter 6).
Integration of results and establishment of management strategies that promoting the
abundance of P. concolor and contributing to the sustainability of the olive grove
(Chapter 7).

64
Chapter 3
Literature
Review
Part of this chapter is an adapted version of the published book chapter:
Pinheiro, Lara A; Medina, P.; Bento, Albino; Pereira, José A. and Santos, Sónia A.,
2017. Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and
Management Strategies to Increase Biological Control of the Olive Fruit Fly. In
Natural Enemies: Identification, Protection Strategies and Ecological Impacts, ed.
Santos, S.A.P., 141-172. ISBN: 978-1-63485-921-9. New York: Nova Science
Publishers.

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
65
Chapter 3:
BIOECOLOGY OF PSYTTALIA CONCOLOR (SZÉPLIGETI)
(HYMENOPTERA: BRACONIDAE) AND MANAGEMENT
STRATEGIES TO INCREASE BIOLOGICAL CONTROL OF THE
OLIVE FRUIT FLY
1. ABSTRACT
Olive growing is an important land-use in the Mediterranean region, with significant
environmental, social and economic roles. However, the olive tree is attacked by several
pests that reduce the yield, the olive oil quality and growers’ income. Among those pests,
the olive fruit fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae), is the major pest of
commercial olives worldwide. In sustainable olive groves, the control of the olive fruit
fly should be achieved by the maximization of the action of its natural enemies. Among
the complex of natural enemies, Psyttalia concolor (Szépligeti) (Hymenoptera:
Braconidae) has been reported as an important endoparasitoid of the olive fruit fly as
well as of other tephritids of economic importance. However, conservation or
enhancement of P. concolor populations in agroecosystems hinges on the maintenance
of non-crop plants (e.g. vegetation strips and bushes) within or around the olive groves
since adult parasitoids require floral resources (pollen and nectar) or honeydew to fulfill
their energetic needs.
Keywords: Bactrocera oleae, biological control, Olea europaea, parasitoid, Psyttalia
concolor
2. INTRODUCTION
The olive tree, Olea europaea L. is an emblematic plant in Mediterranean countries
where it represents an important social, economic and environmental role. Olive groves
are complex agroecosystems harboring organisms belonging to different trophic levels,
some of them causing economic losses such as pathogens (e.g., Verticillium dahliae Kleb
and Colletotrichum acutatum Simmonds) or phytophagous insects [e.g., Bactrocera oleae
(Rossi) (Diptera: Tephitidae) and Prays oleae (Bernard) (Lepidoptera: Praydidae)]
whereas others exert limiting action on these olive enemies such as entomophagous
predators [e.g., Chrysoperla carnea (Stephens) and Chilocorus bipustulatus (Linnaeus)],
parasitoids such as Psyttalia lounsburyi (Silvestri), Ageniaspis fuscicollis (Dalman) or

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
66
Coccophagus lycimnia (Walker) and pathogen antagonists such as Bacillus subtilis
(Ehrenberg) (Daane et al., 2015; Ghribi et al.,2011; López-Escudero and Mercado-
Blanco, 2011; Mnif and Ghribi, 2015; Paredes et al., 2015; Santos et al., 2009).
Among pests, the olive fruit fly B. oleae is a monophagous frugivorous pest feeding
on Olea species (i.e. cultivated O. europaea and wild relatives) (Nardi et al., 2005), and
is considered the key pest with higher economic impact on olive production and product
quality (table olives and olive oil) (Malheiro et al., 2015). The economic losses are related
with the consumption of the fruit pulp by the newly hatched larvae, normally ranging
from 50 to 150 mg/larvae (Neuenschwander and Michelakis, 1978). Moreover, one
female can oviposit 200-250 eggs during its lifetime (Mavragani-Tsipidou, 2002).
Control methods traditionally used against the olive fruit fly rely on the insecticide
application. However, good agricultural practices (GAP) have become an important issue
in agricultural systems as a way to ensure environmental sustainability and food safety
and quality, which is not compatible with the excessive use of insecticides. Therefore, an
alternative agronomic practice to conventional management is the use of natural enemies
reported to attack the olive fruit fly. Some of the parasitoid species referred are the
ectoparasitoids Eupelmus urozonus Dalman (Hymenoptera: Eupelmidae) and Pnigalio
mediterraneus Walker (Hymenoptera: Eulophidae) and endoparasitoids such as Psyttalia
concolor (Szépigeti) and P. lounsburyi (Hymenoptera: Braconidae) (Daane et al., 2015;
Daane and Johnson, 2010).
Historically, P. concolor was almost the only exotic parasitoid used in European
biological control programs against the olive fruit fly, already tested in inoculative and
inundative mass releases (Raspi and Loni, 1994), or combined with other techniques, such
as mass trapping (Hepdurgun et al., 2009; Liaropoulos et al., 2002; 2005). This species
was originally described as Opius concolor by Szépligeti (1910), being the first
description based on samples collected from olives infested with olive fruit flies in
Tunisia. Afterwards, P. concolor was introduced in Italy to control the olive fruit fly
(Delucchi, 1957) being successively used in biocontrol programs.
This parasitoid can be easily reared in laboratory on the Mediterranean fruit fly,
Ceratitis capitata (Wiedemann) (Diptera: Tephritidae), not requiring great costs unlike
other parasitoids (González-Núñez, 1998). In addition, another advantage of the use of P.
concolor in biological control programs is its great capacity to survive to shipping
conditions during importation process (Yokoyama et al., 2006). In the field, the success

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
67
of a parasitoid species depends on its survival, dispersal, attack rate and multi-
generational persistence (Sivinski and Aluja, 2012).
Besides attacking a target pest, a parasitoid should also not attack other important
species in the ecosystem (namely non-pestiferous congeners). In fact, it is known that P.
concolor attacks at least 14 tephritid pests (Diptera) in its native range and in areas where
it has been introduced (Canale et al., 2014). Thus, for an effective pest control, several
factors affecting the success of the parasitoid need to be clarified. In this chapter, the
biology, reproduction, behavior, ecology of the parasitoid and components of the
interaction P. concolor-olive fruit fly are described and new insights to the conservation
of the parasitoid in olive groves are discussed.
3. DISTRIBUTION OF PSYTTALIA CONCOLOR
Psyttalia genus has 79 valid species (Wharton, 2009). Among them, P. concolor is
native to Africa (Figure 3.1) but its taxonomic situation is confusing, with some authors
associating this species to a species complex and others dealing with it as synonyms. For
a more detailed description of taxonomic problems associated with P. concolor see
Kimani-Njogu et al. (2001), Karam et al. (2008) and Rugman-Jones et al. (2009).
In the last decades, several efforts were attempted to introduce P. concolor in a
number of countries (Figure 3.1), allowing its spread, in order to control the population
of the olive fruit fly and of other pests (e.g., C. capitata). Several releases aiming the
biological control of the olive fruit fly by P. concolor occurred in Italy (Monastero, 1968;
Monastero and Delanoue, 1966), Greece (Kapatos et al., 1977), Spain (Jimenez et al.,
1990; Miranda et al., 2008), and California (Yokoyama et al., 2006; 2008). Although
numerous releases were attempted, only few were successful, such as those described by
Raspi and Loni (1994) in Italy and Miranda et al. (2008) in Majorca - Balearic Islands
(Spain). Several factors affected the effectiveness of P. concolor in olive groves, namely
the longevity (in the presence or absence of host) and the accessibility of food source. In
the present chapter these factors are exposed in detail. It is of great importance to create
strategies that will contribute to an adaptation and establishment of P. concolor under
different environmental conditions.

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
68
Figure 3.1. Geographical distribution of Psyttalia concolor: In black are represented its
native countries and in gray are represented the countries where it was introduced.
4. BIOLOGY AND LIFE-HISTORY OF PSYTTALIA CONCOLOR
4.1. The life cycle of Psyttalia concolor
P. concolor is a synovigenic koinobiont larval-pupal endoparasitoid (Figure 3.2) that
fully depends on the host for its development until the adult stage. P. concolor life-cycle
has four developmental stages: egg, larva (with four stages: one initial, two intermediates
and one terminal), pupa (red-eye pupa or young pupa, and black-eyed pupa or old pupa)
and adult (González et al., 1996). Psyttalia concolor is a parasitoid of second- and third-
instar larvae of tephritids, although it might also parasitize freshly molted pupae. The life
cycle begins when a P. concolor adult female lays its eggs onto the host larvae, such as
the olive fruit fly (Figure 3.2), feeding the immature parasitoid on tissues (or hemolymph)
of the host body (Harvey and Strand, 2002). Therefore, the fitness of larvae is related to
the quality of the host and host selection decision by the oviposition female.
Figure 3.2. Psyttalia concolor laying one egg on an olive fruit fly larvae.
Normally, P. concolor life-cycle takes around 17.87 to 21.52 (at 23 and 25ºC,
respectively) from egg to the new adult born, with the eclosion taking around 6 - 7 minutes
(Biliotti and Delanoue, 1959; Loni, 1997). At 24 - 25ºC, egg incubation takes generally

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
69
one day and larval and pupal stages take about 5 - 9 days and 6 - 15 days, respectively
(Bartlett et al., 1978).
4.2. Longevity of Psyttalia concolor adults
P. concolor males emerge between 2 - 3 days before females at 25 ± 2ºC, i.e., 18 - 19
days after host parasitization. Females have a higher survival rate than males, living about
30 and 17 days (respectively) under laboratory conditions (25 ± 2ºC) (González-Núñez,
1998). This reveals a sexual discrepancy in developmental time probably due to sexual
dimorphism. Thus, a faster development of males might represent an increment in mating
success. In females, potential fecundity and oviposition rates are correlated with body
size (Berger et al., 2012), likely representing an investment in reproduction potential that
ensures future survival of the specie.
In the field, longevity is influenced by climatic conditions (temperature and relative
humidity), availability of food sources and costs of sexual interaction. On the other hand,
according to Yokoyama et al. (2006), longevity of P. concolor adults does not depend on
the presence of infested and non-infested olive fruits, which only increases the parasitism
rate and the stability of the parasitoid population. Sime et al. (2006) also showed that P.
concolor life span decreases significantly when food resources are scarce, even in the
presence of hosts. Moreover, in greenhouse experiments, adult parasitoids without food
and water lived 4 and 1 days, in simulated coastal climate and simulated inland valley
climate, respectively (Yokoyama et al., 2008). These results demonstrate that the
parasitoid can live longer in a colder and more humid climate than in a warmer and drier
environment and the presence of water and food sources is essential and increases survival
in the agroecosystems.
Costs of sexual interactions (courtship, mating and reproduction) are generally
assumed to be high both for males (that include gamete production, courtship and combat
and scramble competition) and females (that include physical damage caused by male
genitalia, toxic seminal fluid and offspring production and care) (Benelli et al., 2013a).
For both sexes, the presence of other males and females interferes on their longevity.
Male longevity is extremely reduced when they are kept with females or other males,
being female longevity reduced in the presence of males comparing with the presence of
females (Benelli et al., 2013a). High rates of progeny were produced when parasitoids
were about 12 - 16 days old (Sime et al., 2006; Yokoyama et al., 2006).

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
70
4.3. Courtship and mating behavior
Courtship and mating behavior are the keys for reproduction and knowledge of these
parameters is extremely important for the success of mass rearing and optimization of
biological control programs. Responsiveness and mating success were affected by the
periodicity and duration of mating, being longer and more successful in the early morning
than in the afternoon. However, in males, mating is not affected by the female mating
status (virgin or mated females) (Benelli et al., 2012). Some authors justify that mated
females continue to produce sex attaints after mating; or that the presence of visual signals
emitted by the female are sufficient to stimulate a sexual response from males (Benelli et
al., 2012; De Freitas et al., 2004). Concerning Aphidius ervi (Hymenoptera: Braconidae)
McClure et al. (2007) divided the courtship phase into three steps: (1) a courtship phase,
where male uses its antennas to interact with the female through visual and chemical cues;
(2) followed by a close-range phase, getting females attention through wing fanning; and
lastly (3), a contact phase, characterized by acoustic, mechanical and tactile movements
of the antennas leading to mating efforts. This sequence of courtship is similar to the one
described for P. concolor, with the courtship song being emitted at a frequency of 180 Hz
(Benelli et al., 2012; Canale et al., 2013). In experiments conducted by Benelli et al.
(2012), P. concolor spent 20.3 ± 3.49 seconds in pre-copulatory courtship and the male
approached the female with one or more (till a maximum of nine) copulation attempts.
Another interesting aspect concerning P. concolor courtship is related with male
sexual experience. Since a male-male interaction was observed before female emergence,
Benelli and Canale (2012a; 2013) hypothesized that this previous mating experience
would improve mating intensity. However, no differences were observed in mating
success and copula duration.
4.4. Host-stage preference
The interaction host-parasitoid is also an important factor in biological control
programs optimization, specifically in inundative/augmentative releases. Females spend
a large part of its life looking for potential hosts and the success of parasitoid development
depends on parameters such as female age, size of the host, host exposure to females and
learning experience.
P. concolor is able to discriminate the host instar, searching preferentially for the
second and third instars of the olive fruit fly, but revealing an increase in the reproduction
success when larvae are in the third instar (Canale and Loni, 2006; Raspi and Canale,

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
71
2000; Sime et al., 2006; Yokoyama et al., 2008). Canale and Loni (2006) showed the
difficulties of P. concolor in locating young host larvae. According to those authors, the
parasitoid identifies the larvae inside the fruit through vibrations emitted by the host,
being easier to detect higher vibrations produced by the third instar larvae. On the other
hand, Kouame and Mackauer (1991) suggested that the preference for a specific host
stage could also be the result of the ability of larvae to defend itself from the parasitoid,
being the cost of lost “opportunity time” lesser in larvae with a weaker immune response.
The size of the host larvae influences the parasitoid size, with greater host larvae
originating an increase in the offspring, wing size and the number of flagellomeres
(antennal segments excluding scape and pedicel) (Billah et al., 2005). In the same study,
the relationships between host size, host type and tibia lengths were analyzed, suggesting
that the size of the tibia increased with the host size. However, differences in the
ovipositor–tibia ratio and ovipositor sheath–tibia ratio were not found when P. concolor
was reared on bigger hosts.
In other experiments, when large hosts were exposed to P. concolor females, the sex
ratio was independent of female age (Avilla and Albajes, 1984). However, the sex ratio
was influenced by the size and number of hosts exposed to P. concolor females, emerging
more males when pupae were small but in few numbers (Avilla and Albajes, 1984; Loni,
2003). This can be explained due to the fact that P. concolor females mate only a single
time (monandrous females) and, contrastingly, the males mate multiple times, not
requiring many males in an offspring to fertilize all the females (Ragusa, 1974). P.
concolor seems to have a preference for non-parasitized larvae. However, if larvae have
already been parasitized, the parasitoid preferred hosts with fewer parasitizations due to
competition, resulting in a lower return in offspring number and quality (Benelli, Gennari
and Canale, 2013). However, and still according to the same authors, P. concolor was not
able to recognize the author of larvae parasitization (self-parasitized vs. conspecífic-
parasitized).
Furthermore, antennas are used as chemoreceptors (particularly the last
antennomeres) and play an important role in a first host location behaviour, being the
ovipositor important for exact host location and host discrimination (Barbarossa et al.,
1998; Canale and Raspi, 2000). After larvae approach, the oviposition is usually
successful after 30-40 seconds (Canale and Raspi, 2000). In addition to size, fruit

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
72
characteristics such as epicarp and pulp depth and firmness, and seed size and
morphology can also influence the accessibility to the host larvae (Sivinski, 1991; 2001).
4.5. Visual, olfactory and mechanical stimuli
The success of P concolor in finding one suitable tephritid host in the orchard (habitat)
or fruit (micro-habitat location) is the result of olfactory (volatile semiochemicals), visual
and/or mechanical cues (Benelli et al., 2013b; Quilici and Rousse, 2012). There are some
techniques (e.g., olfactometry and/or electroantennography) that allow studying the
influence of these factors on the ovipositor-probing response (Quilici and Rousse, 2012).
Acording to Stuhl et al. (2011), volatile organic compounds (VOCs) emitted by plants
(from leaves, flowers or fruits) and/or by tephritid larvae, while feeding on the olive pulp,
elicit a response in antennae and ovipositor sensillas.
Benelli et al. (2013b) reported that volatile compounds of healthy and infested fruits
may represent a first approximation to the host selection. Subsequently, VOCs derived
from short-range micro-habitat (e.g., derived from substrate fermentation) and/or
excretion (by the larvae) help in host location.
Color might also have an important role in P. concolor host searching behavior.
Benelli and Canale (2012b) demonstrated a color preference through laboratory trials,
associating larvae with a given color (after a previous training period). The authors also
showed a reaction to color hue, but not to differences in light intensity. However, no
preference for shapes and patterns were found. Transposing to field conditions, P.
concolor might use previous location experiences to reduce searching and decision time
for hosts.
Once P. concolor females are on the surface of the fruit, sounds or vibrations emitted
by the feeding larvae are crucial for host location. Canale (2003) also demonstrated that
interaction between mobility of larvae and host substrate elicited an ovipositor-probing
behavior due to a change of the chemical composition of parasitized fruits, when
compared with healthy fruits. In a situation of larvae immobilization, other factors such
as body temperature and CO2 radiation are recognized by the female, leading to an
oviposition behavior (Meyhöfer and Casas, 1999).
Beyond chemical, visual, and mechanical stimuli, factors such as learning, climatic
conditions and physiological state of the parasitoid influence host location (Quilici and

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
73
Rousse, 2012). During host search, data on female parasitoid response indicate that
localization and oviposition behaviors are synergistic.
4.6. Life-time fecundity
P. concolor female is a synovigenic species (i.e., depends on nutrients for egg
maturation) and each female produces around 60 mature eggs (Genduso, 1969). The egg
laying distribution pattern is associated with finding suitable or unsuitable larvae (Loni,
2003). If P. concolor female accepts the host, lays one egg on each parasitization (Canale
and Loni, 2006). However, Loni (2003) described that the highest percentage of host
larvae are superparasitized with two eggs in order to ensure the developed of the
parasitoid. Zhang and Raspi (1999) also demonstrated that self-superparasitism occurred
at a parasitoid: host ratio of 1:5 using both ”experienced” and “inexperienced” females,
after an exposure period of 6 h. Superparasitism refers to the oviposition behavior of
female when lays an egg in a host already parasitized and can be a strategy to maximize
the progeny development by reducing host larvae immune defense system. In fact, the
venom injected during the oviposition of each egg may be insufficient to weaken the
immune-system, in particular of the latest host instar larvae (Loni, 2003; Raspi and
Canale, 2000). However, only one larva remains alive after superparasitism.
The lifetime production of offspring observed per P. concolor female varied from
22.2 to 28.7 days with higher rates of progeny being produced during the first 12-14 days
under controlled conditions (22 ± 2°C, 40% RH fed on a solution of honey - 50%) (Sime
et al., 2006). Adult females of parasitoids search for nectar and honeydew during the first
few days of life instead of hosts (Jervis and Heimpel, 2005). When food resources are
limited and females do not have nutrients for somatic maintenance, they obtain them by
reabsorbing the eggs (Collier, 1995). This suggests that sources of nutrition are essential
for reproduction success, as adult performance involves high cost of energy and nutrient
accumulation, especially by the females. In this sense, a good strategy for a successful
biological control program would be to increase the potential fecundity and fertility with
e.g., supplemental pollen, since this will enhance P. concolor population (Russell et al.,
2015).

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
74
5. ECOLOGY AND HABITAT RELATIONSHIP
5.1. Psyttalia concolor as a biological control agent of Bactrocera oleae
The role of parasitoid-pest relationship in ecosystem functioning refers to the
importance of P. concolor in limiting B. oleae emergence through the parasitization of
infested olive fruits (Figure 3.3).
Figure 3.3. Interaction between female Psyttalia concolor and Bactrocera oleae.
Oviposition success and pest control can be evaluated through the parasitism rate that
shows the impact of the parasitoid on the pest population, being this parameter studied in
olive orchards for many years. In 1965, in Halki Island (Greece), 25,000 individuals of
P. concolor were released and the parasitoid was successfully established, reaching
parasitism rates of 50% at the end of the study (Stavraki-Pavlopoulou, 1967). In another
release, in Corfu, Kapatos et al. (1977) observed that 30-50% of the host larvae were
parasitized one week after the release. In Jaen (Spain), after three consecutive years of
releases, individuals of P. concolor were collected from olives, showing the winter
permanence of the parasitoid (Jimenez et al., 1990).
One of the most recent studies, reported in two organic olive orchards located in
Majorca (Balearic Islands, Spain), parasitism rates of P. concolor in B. oleae ranged from
6.0% to 40.0% with seasonal averages of 22.4% and 23.4% in each orchard. (Miranda et
al., 2008). Also, P. concolor releases performed in San José (coastal location) and
Grapevine Road (inland valley location), California, lead to parasitism rates from 0.5 to
4.0% and 1.5 to 30.0%, respectively (Yokoyama et al., 2006).

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
75
Mathé-Hubert et al. (2013) estimated parasitism rates through PCR method for P.
concolor and demonstrated an increase of parasitism rates with the time of exposure of 1
h and 8 h from 12.0% to 52.0%, respectively. However, P. concolor parasitism seems to
be low when compared to other parasitoids. Other studies considering factors that limit
the maintenance of the parasitoid in olive orchards need to be performed.
5.2. Thermal compatibility between Psyttalia concolor and Bactrocera oleae
Temperature is an extremely strong factor delimiting survival, reproduction,
emergence development and behavior of insects (e.g. feeding, migration) (González et
al., 1996; Mohamed et al., 2006). Loni (1997) reported a mean development time of P.
concolor ranging between 14.62 and 77.25 days for temperatures varying from 15ºC to
30ºC. Under extreme hot temperatures (> 30°C) or cold temperatures (< 13°C), P.
concolor development is restricted and the reproduction activity is practically inhibited
and often lethal (Loni, 1997), while B. oleae develops and survives from 10 to 30°C
(Wang et al., 2012). When compared with B. oleae, P. concolor presented a similar low
threshold (10.01ºC and 11.70ºC, respectively) (Loni, 1997). This statement shows a
positive match concerning biological thermal requirements. Therefore, temperature not
only affects behavior and performance of P. concolor but also survival of their host
species and might affect the success of biocontrol programs.
At optimal temperature (20 – 25°C), females quickly mate (5 – 10 seconds) after
emergence and host parasitization highly increase (Loni, 1997). When temperatures reach
27°C, P. concolor initiates the thermal stress (Loni, 1997). A curious fact is that P.
concolor cannot complete any developmental stage at 10ºC, but stays alive during 23 to
29 days as larva, 31 days as pupa, becoming lethal for eggs after 6 days (Genduso, 1970).
González et al. (1996) observed a similar lethality on eggs, but a different conclusion was
reported for the last larval stage that survived longer, registering then a higher emergence
than the first and second larval stages. Besides that, no losses were found in the emergence
beneficial capacity in pupae with 14 days (González et al., 1996). These results suggested
that the inability of a parasitoid species for establishing during biological programs may
be attributed to many factors beyond the temperature, such as, inhospitable environments
or seasonal gaps in alternative host availability or specific food resources (e.g., pollen or
nectar of spontaneous flowers).

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
76
5.3. Effect of pesticides on Psyttalia concolor
Insecticides are management tools intended to minimize the impact of pests on
farmer’s crops and economies. Concerning the olive fruit fly, traditional control of this
pest has been based in organophosphate insecticides (e.g., dimethoate), pyrethroids (e.g.,
lambda-cyhalothrin) or spinosad (Skouras et al., 2007; Margaritopoulos et al., 2008).
Although the efficacy of these insecticides in short-term mortality is a point to consider,
others like ecological (e.g. degradation of residues through time) and toxicological (non-
target species can be directly and indirectly affected) effects on natural enemies of pests
should also be evaluated (Guedes et al., 2016). The toxicity of an insecticide depends on
its class, as well as the species and developmental stage of the insect. Some of this
toxicological side effects in natural enemies are: (1) acute toxicity, driving to mortality
or (2) chronic toxicity, causing changes in reproduction (e.g. number of eggs), behavior
performance (e.g. parasitization activity), biochemical or physiological effects (e.g.
changes in enzyme activity or respiration) (Amano and Haseeb, 2001). A number of
studies showed the effect of pesticides on longevity and reproduction of P. concolor (e.g.,
Adán et al., 2007; Bengochea et al., 2012; González et al., 1998a; 1998b; Jacas
andViñuela, 1994; Medina et al., 2006,2008; Sterk et al. 1999, , Viñuela et al., 2000,2001;
Zapata et al., 2005). Thus, a significant decrease of the emergence of adults was observed
when P. concolor pupae were treated with dimethoate; but treatments with kaolin,
Bordeaux mixture and copper oxychloride did not affected emergence (Bengochea et al.,
2014). The mortality rate of females was also studied after 72 h of exposure to pesticides
applied on glass surfaces, on olive tree leaves and under semi-field conditions. Similarly,
to the previous experiment, only dimethoate showed deleterious effect, with 100% of
mortality after 24 h, except on the semi-field experiment (where 100% of mortality was
observed after 72 h). However, reproduction was not significantly affected (Bengochea
et al., 2014).
Other experiments showed that the ingestion of azadirachtin caused a negative impact
on longevity, number of attacked hosts and progeny size of P. concolor females, with
these parameters showing a decrease of 84.4, 15.6 and 32.5% respectively, when
compared with the control treatment (Viñuela et al., 2000). More studies concerning the
effect of pesticides on P. concolor are needed, which allow to harmonize the application
of pesticides and the maintenance of P. concolor in the olive grove.

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
77
6. CONCLUSION
This chapter focused the main factors that influence the abundance of P. concolor in
the olive grove. Food sources have an impact on the parasitoid longevity, but their
influence on pest/parasitoid interaction is still unknown, as well as how they affect
biological control and, consequently, management decisions.
The establishment of P. concolor in a new environment depends on a combination of
abiotic and biotic factors. Some factors such as fruit size, weight, color, fruit epicarp
hardness, phenological stage of the olive fruit and chemical factors may influence the
attractiveness of olive cultivars to P. concolor female. It is clear that landscape structure
plays an important role on parasitoid survival, nutrition and activity. Thus, it is imperative
to study P. concolor feeding habits and nutritional requirements, and their consequences
to longevity, development and reproduction. The abundance of alternative hosts it is
equally important, especially in the absence of olive fruits. Pointing this, it is essential to
find plants with alternative tephritid host flies in olive groves and conserve or enhance
the abundance of these plants.
Moreover, it is essential to understand host - P. concolor mechanisms, as well as host
- associated stimuli and response behavior of females (e.g., antennal search, ovipositor
search and vibrotaxis). Many questions about the interaction between P. concolor and B.
oleae still need to be clarified. It is also important to identify the volatile compounds
involved in these interactions, which can influence host identification and oviposition
behavior of P. concolor.
These studies will contribute to promote the best management practices in the olive
grove and provision of biological control of the olive fruit fly by attracting and conserving
P. concolor in the crop. Moreover, future research about increasing P. concolor longevity
leading to higher parasitism rates and lower B. oleae densities should be exploited. Other
natural enemies of the olive fruit fly should also be surveyed as agents that might be used
with P. concolor.
7. REFERENCES
Adán, A., González, T., Bastante, R., Budia, F., Medina, P., Del Estal, P. and Viñuela, E.
(2007). Efectos de diversos insecticidas aplicados en condiciones de laboratorio
extendido sobre Psyttalia concolor (Szèpligeti) (Hymenoptera: Braconidae). Boletín
de Sanidad Vegetal. Plagas, 33: 391-397.

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
78
Amano, H. and Haseeb, M. (2001). Recently-proposed methods and concepts of testing
the effects of pesticides on the beneficial mite and insect species: study limitations
and implications in IPM. Journal Applied Entomology, 36: 1-11.
Avilla, J. and Albajes, R. (1984). The influence of female age and host size on the sex
ratio of the parasitoid Opius concolor. Entomologia Experimentalis et Applicata, 35:
43-47.
Balzan, M.V. and Wackers, F.L. (2013). Flowers to selectively enhance the fitness of a
host-feeding parasitoid: adult feeding by Tuta absoluta and its parasitoid Necremnus
artynes. Biological Control, 67: 21-31.
Barbarossa, I.T., Muroni, P., Dardani, M., Casula, P. and Angioy, A.M. (1998). New
insight into the antennal chemosensory function of Opius concolor (Hymenoptera:
Braconidae). Italian Journal of Zoology, 65: 367-370.
Bartlett, B.R., Clausen, C.P., DeBach, P., Goeden, R.D., Legner, E.F., McMurtry, J.A.
and Oatman, E.R. (1978). In: C.P. Clausen (Ed.). Introduced parasites and predators
of arthropod pests and weeds: A world review. (pp. 329-331). Washington:
Department of Agriculture, Agricultural Research Service.
Begum, M., Gurr, G., Wratten, S.D., Hedberg P.R. and Nico, H.I. (2006). Using selective
food plants to maximize biological control of vineyard pests. Journal of Applied
Ecology, 43: 547-554.
Benelli, G., Bonsignori, G., Stefanini, C. and Canale, A. (2012). Courtship and mating
behaviour in the fruit fly parasitoid Psyttalia concolor (Szépligeti) (Hymenoptera:
Braconidae): the role of wing fanning. Journal of Pest Science, 85: 55-63
Benelli, G. and Canale, A. (2012a). Do Psyttalia concolor (Hymenoptera: Braconidae)
males gain in mating competitiveness from being courted by other males while still
young? Entomological Science, 15: 257-260.
Benelli, G. and Canale, A. (2012b). Learning of visual cues in the fruit fly parasitoid
Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae). BioControl, 57: 767-777.
Benelli, G. and Canale, A. (2013). Male-male sexual behavior in the parasitic wasp
Psyttalia concolor. Journal of Insect Science, 13:25.
Benelli, G., Gennari, G. and Canale. A. (2013). Host discrimination ability in the tephritid
parasitoid Psyttalia concolor (Hymenoptera: Braconidae). Journal of Pest Science,
86: 245-251.
Benelli, G., Gennari, G., Francini, A. and Canale, A. (2013a). Longevity costs of same-
sex interactions: first evidence from a parasitic wasp. Invertebrate Biology, 132: 156-
162.
Benelli, G., Revadi, S., Carpita, A., Giunti, G., Raspi, A., Anfora, G. and Canale, A.
(2013b). Behavioral and electrophysiological responses of the parasitic wasp
Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) to Ceratitis capitata-
induced fruit volatiles. Biological Control, 64: 116-124.
Bengochea, P., Christiaens, O., Amor, F., Viñuela, E., Rougé, P., Medina, P. and
Smagghe, G. (2012). Ecdysteroid receptor docking suggests that dibenzoylhydrazine-
based insecticides are devoid to any deleterious effect on the parasitic wasp Psyttalia
concolor (Hym. Braconidae). Pest Management Science, 68: 976-985.
Bengochea, P., Budia, F., Viñuela, E. and Medina, P. (2014). Are kaolin and copper
treatments safe to the olive fruit fly parasitoid Psyttalia concolor? Journal of Pest
Science, 87: 351-359.
Berger, D., Olofsson, M., Friberg, M. Karlsson, B., Wiklund, C. and Gotthard, K. (2012).
Intraspecific variation in body size and the rate of reproduction in female insects –
adaptive allometry or biophysical constraint? Journal of Animal Ecology, 8: 1244-
1258

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
79
Biliotti, E. and Delanoue, P. (1959). Contribution a l'étude biologique d' Opius concolor
Szepl. (Hym. Braconidae) en élevage de laboratoire. Entomophaga, 4: 7-14.
Billah, M.K., Kimani-Njogu, S., Overholt, W.A., Wharton, R.A., Wilson D.D. and
Cobblah, M.A. (2005). The effect of host larvae on three Psyttalia species
(Hymenoptera: Braconidae), parasitoids of fruit-infesting flies (Diptera: Tephritidae).
International Journal of Tropical Insect Science, 25: 168-175.
Canale, A. (2003). Psyttalia concolor (Hymenoptera Braconidae): role of host movement
and host substrate in ovipositor-probing behaviour. Bulletin of Insectology, 56: 211-
213.
Canale, A. Benelli, G., Lanzo, F., Giannotti, P. Mazzoni, V. and Lucchi, A. (2013). The
courtship song of fanning males in the fruit fly parasitoid Psyttalia concolor
(Szépligeti) (Hymenoptera: Braconidae). Bulletin of Entomological Research, 103:
303-309.
Canale, A., Geri, S. and Benelli, G. (2014). Associative learning for host-induced fruit
volatiles in Psyttalia concolor (Hymenoptera: Braconidae), a koinobiont parasitoid of
tephritid flies. Bulletin of Entomological Research, 104: 774-780.
Canale, A. and Loni, A. (2006). Host location and acceptance in Psyttalia concolor: role
of host instar. Bulletin of Insectology, 59: 7-10.
Canale, A. and Raspi, A. (2000). Host location and oviposition behaviour in Opius
concolor Szépligeti (Hymenoptera, Braconidae). Entomological Problems, 31: 25-
32.
Cobo, A., González-Núñez, M., Sánchez-Ramos, I. and Pascual, S. (2014). Selection of
non-target tephritids for risk evaluation in classical biocontrol programmes against
the olive fruit fly. Journal Applied Entomology, 139: 179-191.
Collier, T.R. (1995). Host Feeding, Egg Maturation, Resorption, and Longevity in the
Parasitoid Aphytis melinus (Hymenoptera: Aphelinidae). Annals of the Entomological
Society of America, 88: 206-214.
Copeland, R.S., Wharton, R.A., Luke, Q. and De Meyer, M. (2002). Indigenous hosts of
Ceratitis capitata (Diptera: Tephritidae) in Kenya. Annals of the Entomological
Society of America, 95: 672-694.
Daane, K.M. and Johnson, M.W. (2010). Olive Fruit Fly: Managing an Ancient Pest in
Modern Times. Annual Review of Entomology, 55: 151-169.
Daane, K.M., Wang, X., Nieto, D.J., Pickett, C.H., Hoelmer, K.A., Blanchet, A. and
Johnson, M.W. (2015). Classic biological control of olive fruit fly in California, USA:
release and recovery of introduced parasitoids. BioControl, 60: 317-330.
De Freitas, M.R.T., De Lima Medonça, A., Do Nascimento, R.R. and Sant’Ana, E.G.
(2004). Behavioural evidence for a female sex pheromone in Cotesia flavipes
(Hymenoptera: Braconidae). Physiological Entomology, 29: 183-187.
Delucchi, V. (1957). Les parasites de la mouche des olives. Entomophaga, 2: 107-118.
Farrell, B.D., Mitter, C. and Futuyma, D.J. (1992). Diversification at the insect-plant
interface, insights from phylogenetics. BioScience, 42: 34-42.
Furtado, C. Belo, A.F., Nunes, F.M., Ganhão, E., Müller, C.T., Torres, L. and Rei, F.T.
(2016). Evaluating potential olive orchard sugar food sources for the olive fly
parasitoid Psyttalia concolor. BioControl, 61(5): 473-483.
Genduso, P. (1970). Influenza della temperatura sulla durata e sulla resistenza degli stadi
preimmaginali dell' Opius c. siculus Mon. Bolletino dell’Instituto di Entomologia
Agraria e dell' Observatorio di Fitopatologia di Palermo, 8: 1-7
Ghribi, D., Mnif, I., Boukedi, H., Kammoun, R., Ellouze-Chaabouni, S., (2011).
Statistical optimization of low-cost medium for economical production of Bacillus

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
80
subtilis biosurfactant, a biocontrol agent for the olive moth Prays oleae. African
Journal of Microbiology Research, 5: 4927-4936.
González, M., Bahena, F. and Viñuela, E. (1998a). Efectos de distintos reguladores del
crecimiento de insectos (RCI) sobre el parasitoide Opius concolor Szèpligeti cuando
son ingeridos por la larva huésped. Boletín de Sanidad Vegetal. Plagas, 24: 193-200.
González, M., Bahena, F. and Viñuela, E. (1998b). Desarrollo de un método de
semicampo para el estudio de los efectos secundarios de los productos fitosanitarios
sobre el parasitoide Opius concolor Szèpligeti. Boletín de Sanidad Vegetal. Plagas,
24: 661-668.
González, M., Jacas, J.A., Jiménez, A. and Viñuela, E. (1996). Optimización de la cría de
Opius concolor Szepl. mediante la utilización de bajas temperaturas durante su
desarrollo pre-imaginal. Boletín de Sanidad Vegetal. Plagas, 22: 107-114.
González-Núñez, M. (1998). Uso conjunto de plaguicidas y enemigos naturales en el
olivar: Optimización del manejo de Opius concolor Szèpligeti, parasitoide de la
mosca del olivo, B. oleae (Gmelin). PhDThesis. Madid:. Universidad Politécnica de
Madrid. ETSI Agrónomos.
Guedes, R.N.C., Smagghe, G., Stark, J.D. and Desneux, N. (2016). Pesticide-induced
stress in arthropod pests for optimized integrated pest management programs. Annual
Review of Entomology, 61: 3.1-3.20.
Harvey J.A. and Stran, M.R. (2002). The developmental strategies of endoparasitoid
wasps vary with host feeding ecology. Ecology, 83: 2439-2451.
Hepdurgun, B. Turanli, T. and Zümreoğlu, A. (2009). Parasitism rate and sex ratio of
Psyttalia (=Opius) concolor (Hymenoptera: Braconidae) reared on irradiated
Ceratitis capitata larvae (Diptera: Tephritidae). Biocontrol Science and Technology,
19: 157-165.
Hernandez-Ortiz, V. and Perez-Alonso, R. (1993). The natural host plants of Anastrepha
(Diptera: Tephritidae) in a tropical rain forest of Mexico. The Florida Entomologist,
76: 447-460.
Jacas J.A. and Viñuela, E. (1994). Normalización del método para estudiar los efectos
secundarios de los fítosanitarios sobre Opius concolor Szèpligetti (Hym. Braconidae).
Boletín de Sanidad Vegetal. Plagas, 20: 409-417.
Jervis, M.A. and Heimpel, G.E. (2005). Phytophagy. In: M.A. Jervis (Ed.), Insects as
Natural Enemies: A Practical Perspective. (pp. 525-550). Dordrecht: Springer.
Jervis, M.A., Kidd, N.A.C., Fitton, M.G., Huddleston, T. and Dawah, H.A. (1993).
Flower-visiting by hymenopteran parasitoids. Journal of Natural History, 27: 67-105.
Jimenez, A., Castillo, E. and Lorite, P. (1990). Supervivencia del himenóptero bracónido
Opius concolor Szep. parásito de Dacus oleae Gmelin. en olivares de Jaén. Boletín
de Sanidad Vegetal. Plagas, 16: 97-103.
Kapatos, E., Fletcher, B.S., Pappas, S. and Laudeho, Y. (1977). The release of Opius
concolor and O. concolor (var. siculus [Hym.: Braconidae] against the spring
generation of Dacus oleae [Dipt.: Trypetidae] on Corfu. Entomophaga, 22: 265-270.
Karam, N., Guglielmino, C.R., Bertin, S., Gomulski, L.M., Bonomi A., Baldacchino, F.,
Simeone, V. and Malacrida, A.R. (2008). RAPD analysis in the parasitoid wasp
Psyttalia concolor reveals Mediterranean population structure and provides SCAR
markers. Biological Control, 47: 22-27.
Kimani-Njogu, S.W., Trostle, M.K., Wharton, R.A., Woolley, J.B. and Raspi, A. (2001).
Biosystematics of the Psyttalia concolor species complex (Hymenoptera: Braconidae:
Opiinae): the identity of populations attacking Ceratitis capitata (Diptera:
Tephritidae) in coffee in Kenya. Biological Control, 20: 167-174.

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
81
Kouame, K.L. and Mackauer, M. (1991). Influence of aphid size, age and behaviour on
host choice by the parasitoid wasp Ephedrus californicus: a test of host-size models.
Oecologia, 88: 197-203.
Leblanc, L., Vueti, E.T., Drew, R.A.I. and Allwood, A.J. (2012). Host plant records for
fruit flies (Diptera: Tephritidae: Dacini) in the Pacific Islands. Proceedings of the
Hawaiian Entomological Society, 44: 11-53.
Liaropoulos, C., Mavraganis, V.G., Broumas, T. and Ragoussis, N. (2002). Field tests on
the efficacy of mass trapping combined with the release of Opius concolor
(Hymenoptera: Braconidae) parasites for the control of the olive fruit fly Bactrocera
oleae (Diptera: Tephritidae). VIIth European Congress of Entomology, October 7-13,
Thessaloniki (Greece).
Liaropoulos, C., Mavraganis, V.G., Broumas, T. and Ragoussis, N. (2005). Field tests on
the combination of mass trapping with the release of the parasite Opius concolor
(Hymenoptera: Braconidae), for the control of the olive fruit fly Bactrocera oleae
(Diptera: Tephritidae). In Proceedings of the Working Group on Integrated Protection
of Olive Crops, Florence, Italy: IOBC/WPRS Bulletin, 30: 77-81.
Lombardo, N. (2003). Aspetti generali dell´olivicoltura. In: P. Fiorino (ed.) Olea, Tratatto
di Olivicoltura. (pp. 3-12). Bologna: Ed. Edagricole.
Loni, A. (1997). Developmental rate of Opius concolor (Hym.: Braconidae) at various
constant temperatures. Entomophaga, 42: 359-366.
Loni, A. (2003). Impact of host exposure time on mass-rearing of Psyttalia concolor
(Hymenoptera Braconidae) on Ceratitis capitata (Diptera Tephritidae). Bulletin of
Insectology, 56: 277-282.
López-Escudero, F.J., and Mercado-Blanco, J. (2011). Verticillium wilt of olive: a case
study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil,
344: 1-50.
Malheiro, R., Casal, S., Baptista, P. and Pereira, J.A. (2015). A review of Bactrocera
oleae (Rossi) impact in olive products: From the tree to the table. Trends in Food
Science and Technology, 44: 226-242.
Margaritopoulos, J.T., Skavdis, G., Kalogiannis, N., Nikou, D., Morou, E., Skouras,P.J.,
Tsitsipis, J.A. and Vontas, J. (2008). Efficacy of the pyrethroid alpha-cypermethrin
against Bactrocera oleae populations from Greece, and improved diagnostic for an
iAChE mutation. Pest Management Science, 64: 900-908.
Mathé-Hubert, H., Gatti, J-L., Poirié, M. and Malausa, T. (2013). A PCR-based method
for estimating parasitism rates in the olive fly parasitoids Psyttalia concolor and P.
lounsburyi (Hymenoptera: Braconidae). Biological Control, 67: 44-50.
Mavragani-Tsipidou, P. (2002). Genetic and cytogenetic analysis of the olive fruit fly
Bactrocera oleae (Diptera: Tephritidae). Genetica, 116: 45-57.
McClure, M., Whistlecraft. J. and McNeil, J.N. (2007). Courtship behaviour in relation
to the female sex pheromone in the parasitoid Aphidius ervi (Hymenoptera:
Braconidae). Journal of Chemical Ecology, 33: 1946-1959.
Medina, P., Corrales, E., González-Núñez , M., Smagghe, G. and Viñuela, E. (2008).
Effects of Beauveria bassiana, Heterorhabditis bacteriophora, H. megidis and
Sterinernema feltiae on the Mediterranean fruit fly Ceratitis capitata and the very
sensitive braconid Psyttalia concolor. IOBC/wprs Bulletin. 35: 113-121.
Medina, P., Budia, F., Rodriguez, B., González, M., Diaz, A., Huerta, A., Zapata, N. and
Viñuela, E. (2006) Effects of botanical insecticides on two natural enemies of
importance in Spain: Chrysoperla carnea (Stephens) and Psyttalia concolor
(Szépligeti). IOBC/wprs Bulletin. 29: 85-93.

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
82
Meyhöfer, R. and Casas, J. (1999). Vibratory stimuli in host location by parasitic wasps.
Journal of Insect Physiology, 45: 967-971.
Miranda, M.A., Miquel, M., Terrassa, J., Melis, N. and Monerris, M. (2008). Parasitism
of Bactrocera oleae (Diptera, Tephritidae) by Psyttalia concolor (Hymenoptera,
Braconidae) in the Balearic Islands (Spain). Journal of Applied Entomology, 132:
798-805.
Mnif, I. and Ghribi, G. (2015). Potential of bacterial derived biopesticides in pest
management. Crop Protection, 77: 52-64.
Mohamed, S.A., Overholt, W.A., Wharton, R.A. and Lux, S.A. (2006). Effect of
temperature on developmental time and longevity of Psyttalia cosyrae (Hymenoptera:
Braconidae). Biocontrol Science and Technology, 16 : 717-726.
Monastero, S. (1968). Nouvelle expérimentation à grande échelle de la lutte biologique
contre la mouche de l'olive (Dacus oleae Gmel.) au moyen d' Opius concolor siculus
Mon. en Sicile en 1967. Entomophaga, 13 : 251-261.
Monastero, S. and Delanoue P. (1966). Lutte biologique expérimentale contre la mouche
de l'olive (Dacus oleae Gmel.) au moyen d' Opius concolor Szepl. siculus Mon. dans
les iles éoliennes (Sicile) en 1965. Entomophaga, 11: 411-432.
Nardi, F., Carapelli, A., Dallai, R., Roderick, G.K. and Frati, F. (2005). Population
structure and colonization history of the olive fly, Bactrocera oleae
(Diptera:,Tephritidae). Molecular Ecology, 14: 2729-2738.
Neuenschwander, P. and Michelakis, S. (1978). Infestation of Dacus oleae (Gmel.)
(Diptera, Tephritidae) at harvest time and its influence on yield and quality of olive
oil in Crete. Journal of Applied Entomology, 86: 420-433.
Ortega, M. and Pascual, S. (2014). Spatio-temporal analysis of the relationship between
landscape structure and the olive fruit fly Bactrocera oleae (Diptera: Tephritidae)
Agricultural and Forest Entomology, 16: 14-23.
Paredes, D., Cayuela, L., Gurr, G.M. and Campos, M. (2015). Single best species or
natural enemy assemblages? A correlational approach to investigating ecosystem
function. BioControl, 60: 37-45.
Quilici, S. and Rousse, P. (2012). Location of host and host habitat by fruit fly parasitoids.
Insects, 3: 1220-1235.
Ragusa, S. (1974). Influence of temperature on the oviposition rate and longevity of Opius
concolor siculus [Hymenoptera: Braconidae]. Entomophaga, 19: 61-66.
Raspi, A. and Canale, A. (2000). Effect of superparasitism on Ceratitis capitata
(Wiedemann) (Diptera Tephritidae) second instar larvae by Psyttalia concolor
(Szépligeti) (Hymenoptera, Braconidae). Redia, 83: 123-131.
Raspi, A. and Loni, A. (1994). Alcune note sull’allevamento massale di Opius concolor
Szépligeti (Hym.: Braconidae) e su recenti tentative d’introduzione della specie in
Toscana e Liguria. Frustula Entomologica, 30: 135-145.
Rugman-Jones, P.F. Wharton, R., van Noort, T. and Stouthamer, R. (2009). Molecular
differentiation of the Psyttalia concolor (Szépligeti) species complex (Hymenoptera:
Braconidae) associated with olive fly, Bactrocera oleae (Rossi) (Diptera:
Tephritidae), in Africa. Biological Control, 49: 17-26.
Russell, M. (2015). A meta-analysis of physiological and behavioral responses of
parasitoid wasps to flowers of individual plant species. Biological Control, 82: 96-
103.
Santos, S.A.P., Pereira, J.A., Torres, L.M. and Nogueira, A.J.A. (2009). Voracity of
coccinellid species on different phenological stages of the olive pest Saissetia oleae
(Homoptera, Coccidae). Applied Ecology and Environmental Research, 7: 359-365.

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
83
Sime, K.R., Daane, K.M., Messing, R.H and Johnson, M.W. (2006). Comparison of two
laboratory cultures of Psyttalia concolor. (Hymenoptera: Braconidae), as a parasitoid
of the olive fruit fly. Biological Control, 39: 248-255.
Sivinski, J. (1991). The influence of host fruit morphology on parasitization rates in the
Caribbean fruit fly. Anastrepha suspensa. Entomophaga, 36: 447-454.
Sivinski, J., Vulinec, K. and Aluja, M. (2001). Ovipositor length in a guild of parasitoids
(Hymenoptera: Braconidae) attacking Anastrepha spp. fruit flies (Diptera:
Tephritidae) in southern Mexico. Annals of the Entomological Society of America,
94: 886-895.
Sivinski, J., Wahl, D., Holler, T., Dobai, S.A. and Sivinski, R. (2011). Conserving natural
enemies with flowering plants: Estimating floral attractiveness to parasitic
Hymenoptera and attraction’s relationship to flower and plant morphology. Biological
Control, 58: 208-214.
Skouras, P.J., Margaritopoulos, J.T., Seraphides, N.A., Ioannides, I.M., Kakani, E.G.,
Mathiopoulos, K.D. and Tsitsipis, J.A. (2007). Organophosphate resistance in olive
fruit fly, Bactrocera oleae, populations in Greece and Cyprus. Pest Management
Science, 63: 42-48.
SPSS Inc., IBM Company (2010). IBM, SPSS Statistic for Windows, version 19.0.0. New
York
Stavraki-Pavlopoulou, H. (1967). Essais préliminaires de lâchers d’ Opius concolor
Szepl. (Hymen. : Braconidae) parasite du Dacus oleae Gmel. (Dipt. : Trypedidae)
dans l’île de Chalki 1965. Annales de l’lnstitut Phytopathologique Benaki, 8 : 23-31.
Sterk, G., Hassan. S.A., Baillod, M., Bakker, F., Bigler, F., Blümel, S., Bogenschütz, H.,
Boller, E., Bromand, B., Brun, J., Calis, J.N.M., Coremans-Pelseneer, J., Duso, C.,
Garrido, A., Grove, A., Heimbach, U., Hokkanen, H., Jacas, J., Lewis, G., Moreth,
L., Polgar, L., Rovesti, L., Samsoe-Petersen, L., Sauphanor, B., Schaub, L., Staübli,
A., Tuset, J.J., Vainio, A., Van De Veire, M., Viggiani, G., Viñuela, E. and Vogt, H.
(1999). Results of the seventh joint pesticide testing programme carried out by The
IOBC/wprs Working Group Pesticides and Beneficials. BioControl, 44: 99-117.
Stuhl, C., Sivinski, J., Teal, P. Paranhos, B. and Martin, A. (2011). A compound produced
by fruigivorous tephritidae (Diptera) larvae promotes oviposition behavior by the
biological control agent Diachasmimorpha longicaudata (Hymenoptera:
Braconidae). Environmental Entomology, 40: 727-736.
Szèpligeti, G. (1910). Sur un Braconide (Hym.) nouveau, parasite du Dacus oleae.
Bulletin de la Societe Entomologique de France, 13 : 243-244.
Vayssières, J-F., Adandonon, A., N'Diaye, O., Sinzogan, A., Kooymann, C., Badji, K.,
Rey, J-Y. and Wharton, R.A. (2012). Native parasitoids associated with fruit flies
(Diptera: Tephritidae) in cultivated and wild fruit crops in Casamance, Senegal.
African Entomology, 20: 308-315.
Viñuela, E., Adán, A., Smagghe, G., González, M., Medina, M.P., Budia, F., Vogt, H.
and Del Estal, P. (2000). Laboratory effects of ingestion of azadirachtin by two pests
(Ceratitis capitata and Spodoptera exigua) and three natural enemies (Chrysoperla
carnea, Opius concolor and Podisus maculiventris). Biocontrol Science and
Technology, 10: 165-177.
Viñuela, E., Medina, M.P., Schneider, M., González, M., Budía, F., Adán, A. and Del
Estal, P. (2001). Comparison of side-effects of spinosad, tebufenozide and
azadirachtin on the predators Chrysoperla carnea and Podisus maculiventris and the
parasitoids Opius concolor and Hyposoter didymator under laboratory conditions.
IOBC/wprs Bulletin. 24: 25-34.

Chapter 3: Bioecology of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and management strategies to increase
biological control of the olive fruit fly
84
Yokoyama, V.Y., Rendon, P.A. and Sivinski, J. (2006). Fruit flies of economic
importance: from basic to applied knowledge. Proceedings of the 7th International
Symposium on Fruit Flies of Economic Importance (pp. 157-164), 10-15 September,
Salvador (Brazil).
Yokoyama, V.Y., Rendón, P.A. and Sivinski, J. (2008). Psyttalia cf. concolor
(Hymenoptera: Braconidae) for biological control of olive fruit fly (Diptera:
Tephritidae) in California. Environmental Entomology, 37: 764-773.
Wäckers, F. L. (2005). Suitability of (extra-)floral nectar, pollen, and honeydew as insect
food sources. F.L. Wäckers, P.C.J. van Rijn and J. Bruin (Eds.). Plant-Provided Food
for Carnivorous Insects, A Protective Mutualism and its Applications. (pp 17-74).
Cambridge: Cambridge University Press,
Wang, X-G, Levy, K., Son, Y., Johnson, M.W. and Daane, K.M. (2012). Comparison of
the thermal performance between a population of the olive fruit fly and its co-adapted
parasitoids. Biological Control, 60: 247-254.
Wharton, R.A. (2009). Two new species of Psyttalia Walker (Hymenoptera, Braconidae,
Opiinae) reared from fruit-infesting tephritid (Diptera) hosts in Kenya. ZooKeys, 20:
349-377.
Zapata, N., Medina, P., Viñuela, E. and Budia, F. (2005). Toxicidad de malation,
pimetrocina, piretrinas naturales + PBO y triflumuron en adultos del parasitoide
Psyttalia concolor (Szépligeti) (Hym.: Braconidae) según el modo de aplicación.
Boletín de Sanidad Vegetal. Plagas, 31: 111-118.
Zhang, L. and Raspi, A. (1999). Learning behaviour of Opius concolor host
discrimination Szlèpligeti (Hymenoptera: Braconidae) in host discrimination.
Entomologia Sinica, 6: 259-266.

85
Chapter 4
Sugars and honeydew increase
longevity and parasitism
capacity of Psyttalia
concolor (Hymenoptera:
Braconidae)

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
86
Chapter 4:
SUGARS AND HONEYDEW INCREASE LONGEVITY AND
PARASITISM CAPACITY OF PSYTTALIA CONCOLOR
(HYMENOPTERA: BRACONIDAE)
1. ABSTRACT
The potential of Psyttalia concolor Szépligeti as a biological control agent of
Bactrocera oleae (Rossi) (Diptera: Tephritidae) is expected to increase when different
sugar sources are supplied. Sugars are an important food source required by adults this
parasitoid species to enhance its survival, fecundity and fitness. The objective of this
study was to test the effect of seven sugars (glucose, fructose, sucrose, trehalose,
melibiose, melezitose and sorbitol) on longevity and parasitism capacity of P. concolor.
Moreover, we evaluated the effect of two types of honeydews excreted by hemipteran
pests present in olive groves, Saissetia oleae Olivier (Coccidae) and Euphyllura olivina
(Costa) (Psyllidae) on the longevity of the parasitoid.
Our results showed a positive relationship between carbohydrate (single sugars and
honeydew) and parasitoid survival. Females longevity increased when fed on sorbitol and
melibiose, however for males, glucose and fructose were the best food resources,
suggesting that hexose-nectars would benefit this sex. Interestingly, sucrose increased
the percentage of attacked hosts and parasitism rate while melezitose decreased
significantly these percentages, comparatively to the other sugars offered. Psyttalia
concolor benefited more from feeding on honeydew than on sugars, representing this food
item an important source of sugar in olive groves. This information is important for
biological control, indicating the importance of certain nutrients for promoting the
population of P. concolor in agroecosystems.
Keywords: Honeydew, longevity, nutrition, Olea europaea, parasitoid, sugars
2. INTRODUCTION
Adult parasitoids are one important group of natural enemies of insect pests that
depend on non-host food (nectar, pollen or honeydew excreted by hemiptera to survive
and reproduce (Benilli et al., 2017). The consumption of carbohydrates, either present in

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
87
(extra)-floral nectar or in honeydew seems to be the main food resource commonly
exploited by parasitoids (Wäckers, 2005; Wäckers, van Rijn and Heimpel, 2008).
Although grains of pollen have been found in the guts of some parasitoids (e.g., Zhang,
Zimmermann and Hassan, 2004), according to Jervis (1998), pollen-feeders appeared to
be rare.
Nectar is mainly composed by quickly digestible carbohydrates with high-energy
content, being two hexoses (glucose and fructose) and one disaccharide (sucrose) the
principal sugars, with concentration ranges from 7% to 70% w/w (Baker and Baker,
1983). After sugars, amino acids (essential, non-essential and some non-protein amino
acids) are the most abundant components in nectar (Petanidou et al., 2006). Several
studies demonstrated the greatest positive effect of nectar, especially floral nectar, on
parasitoid longevity due to the fact that these sugars are more easily converted into energy
(Hausmann, Wäckers and Dorn, 2005; Rivero and Casas, 1999).
Although, according to the literature, honeydew is considered an inferior source of
sugar compared to nectar, there is a variation in the chemical profile due to honeydew
producer and its host plant (Hogervorst, Wäckers and Romeis, 2007a, b). In agricultural
systems where flowering plants are frequently scarce, honeydew is probably the only
available source of carbohydrates (Wäckers, 2005). On the other hand, all plant-derived
food is not available in the same way for the parasitoids, depending on accessibility,
quality (e.g., concentration and composition), quantity, distribution and detectability
(Lewis et al., 1998). For example, the quantity and quality of nectar depends (1) on the
plant species available in the foraging area or/and (2) the production of nectar, being
variable with time of day, climatic conditions and competition with other nectar-foragers
(see Lucchetta et al., 2008).
Under laboratory conditions, parasitoids fed on floral resources (especially nectar)
increased its longevity, fecundity (e.g., can expand the reproductive period, mature more
eggs, attack more hosts) and foraging ability when compared with starved parasitoids,
(e.g., Wäckers, 2001; Russell, 2015; Nave et al., 2017a). In the field, several studies also
confirmed that parasitoids benefited from sugar feeding (Winkler et al., 2006; Lee and
Heimpel, 2008; Tena et al., 2015). Moreover, the effects of sugar feeding on nutrients
levels of parasitoids (sugars, glycogen and lipids) have also been reported (e.g., Olson et
al., 2000; Lee, Heimpel and Leibee, 2004; Nafziger Jr. and Fadamiro, 2011; Snart et al.,
2018).

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
88
The olive tree (Olea europaea L.) is among the oldest and most widespread crops in
the Mediterranean region, having relevant social, economic, cultural and landscape
importance (Loumou and Giourga, 2003). The olive tree is attacked by different pest
species, being the olive fruit fly Bactrocera oleae (Rossi) (Diptera: Tephritidae) the key
pest in many olive crop areas, affecting both quality and quantity of oil and table olives
(Daane and Johnson, 2010). Control methods traditionally used against the olive fruit fly
rely on insecticide applications. However, more sustainable practices to control this pest
are pursued such as the use of the parasitoid wasp species, Psyttalia concolor (Szépligeti)
(Hymenoptera: Braconidae), that parasitizes larvae of B. oleae developing in the olive
fruit. Psyttalia concolor has been released in the aim of some biocontrol programs
developed against B. oleae in European countries (e.g., Italy, Greece or Spain) and in the
USA to control the olive fruit fly (Raspi and Loni, 1994). However, only few actions were
successfully described (Raspi and Loni, 1994; Miranda et al., 2008) and more studies on
potential strategies that could be adopted in order to promote the maintenance of P.
concolor in olive groves are needed. One of those strategies regards the availability of
sugary food items that can maintain adult specimens in the field. Thus, the objective of
this work is to study the effect of different sugars on longevity and parasitism capacity of
P. concolor as a way to improve biocontrol services against B. oleae. Moreover, the effect
of two types of honeydew excreted by two secondary olive tree pests, the black- scale,
Saissetia oleae (Olivier) (Hemiptera: Coccidae) and the olive psyllid, Euphyllura olivina
(Costa) (Hemiptera: Psyllidae) on longevity of P. concolor was tested.
3. MATERIAL AND METHODS
3.1. Insect rearing
Psyttalia concolor was reared on Ceratitis capitata (Wiedemann, 1824) (Diptera:
Tephritidae) in the Crop Protection Unit, ETSIAAB-UPM, following the methodology
described by Jacas and Viñuela (1994). Both insects were reared at 25 ± 2ºC temperature,
75±5% humidity and 16:8h light: dark photoperiod. Parasitoids were neither exposed to
food or water prior to the experiments. C. capitata was used as a model host to test the
effect of different food sources on parasitism capacity of P. concolor.

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
89
3.2. Food sources
A set of seven sugars were tested in the experiment 1: two monosaccharides (D(+)-
Glucose 1-hydrate (RFE, USP, BP, Ph. Eur., DAB) PRS-CODEX, PanReac AppliChem
and D(+)-Fructose, 99%, ACROS Organics), three disaccharides ((D+)-Sucrose, + 99%
ACROS Organics; D(+)-Trehalose dihydrate, 99%, ACROS Organics and Melibiose, 98
% Sigma-Aldrich)), one trisaccharide (D(+)-Melezitose monohydrate, 99+%, Acros
Organics) and one sugar alcohol (Sorbitol BioUltra, ≥99.5% (HPLC), Sigma-Aldrich).
These sugars were selected based on the described major compounds of plant-derived-
sugars (floral nectars and honeydew - see Wäckers, 2001). The concentration used for all
the sugar solutions was 1 M, representing the upper limit of sugars found in floral nectar
and honeydew (Wäckers, 2001).
For the honeydew treatment (experiment 2), S. oleae and E. olivina honeydew were
collected from 2-year olive trees maintained in a climate chamber at 24 ± 1ºC, 70 ± 5%
relative humidity (RH) and a photoperiod of 16:8 h (L:D) at School of Agriculture,
Polytechnic Institute of Bragança. A piece of Parafilm® (5 cm x 5 cm) was placed under
olive tree branches infested with each honeydew producer, respectively, for 24 hours.
After 24 hours, droplets of honeydew accumulated on the Parafilm® were offered to the
parasitoids.
3.3. Longevity and parasitism capacity of Psyttalia concolor fed on different sugars
3.3.1. Longevity
For longevity experiments, five recently emerged (< 24 h) females and males of P.
concolor (Fig. 4.1. A) were placed in a plexiglass cage (12 cm in diameter x 5 cm height)
with a hole on the top (5.5 cm in diameter) covered with a net for ventilation (Fig. 4.1.
B). A total of six replicate cages were tested for each treatment.
Fig. 4.1. A: Psyttalia concolor adult; B: Plexiglass cage used in the
longevity tests; C: Detail of feeding glass vials.
A B C

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
90
Each sugar solution was offered individually in a glass vial (2 ml) with a strip of
sponge and overlaid with a strip of Parafilm® (Fig. 4.1. C). Glass vials were glued to the
base of the plexiglass cages with a piece of sticky putty and a similar vial with distilled
water was also provided. Distilled water and solid diet - mixture of Brewer`s yeast and
icing sugar (1:4) - were used ad libitum as negative and positive controls, respectively.
Sugars and solid diet were refilled on a daily basis and changed every week, while
melezitose was renewed daily due to its propensity to crystallize. Parasitoid survival was
assessed on a daily basis.
3.3.2. Parasitism capacity
The parasitism capacity parameters, i.e percentage of attacked hosts (percentage of
puparia without C. capitata emergence – induced parasitized hosts and naturally died C.
capitate pupae) and percentage of progeny size (percentage of parasitoids emerged from
parasitised puparia) were evaluated using similar cages to the longevity experiments, but
with an additional hole (5.5 cm in diameter) on the bottom of the cage covered with a net
(Fig. 4.2. A). One mated female was placed in the cage and fed with a single sugar or
solid diet (control).
Fig. 4.2. A: Plexiglass cage used in the parasitation tests; B: 3rd instar larvae
of C. capitata were offered to P. concolor; C: Detail of parasitism.
After seven days, 3rd instar larvae of C. capitata were offered to P. concolor females
at a proportion of 20 larvae/female by placing them on the top of a glass bottle covered
with a piece of Parafilm® (4 cm in diameter) (Fig. 4.2. B) for one hour (according to
Bengochea et al. (2014) (Fig. 4.2. C) and this procedure was repeated during five
consecutive days. Subsequently, C. capitata larvae were transferred into Petri dishes and
placed at 25 ± 2ºC, 75 ± 5% and 16: 8 h light: dark until emergence. The time elapsed

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
91
from the pupal stage until emergence of C. capitate adults was recorded. Since previous
studies showed that females need one day to learn to parasite, only 4 days of parasitization
were used in data analysis. A total of 25 females were tested for each treatment.
3.4. Longevity of Psyttalia concolor fed on honeydew
Groups of 10 recently emerged males or females (12-24 h old) were placed in plastic
cages (diameter 7.5 cm x height 4.5 cm) with a hole on the top covered with a net for
ventilation. Each cage was provided with (1) honeydew (S. oleae or E. olivina) + distilled
water or (2) 10% Mountain multifloral honey (Bragança Mountain) + distilled water
(positive control) or (3) only distilled water (food deprived parasitoids - negative control).
Water and honey were provided in 1.5 ml microcentrifuge tubes with a strip of sponge
and plugged with a strip of Parafilm®. Honeydew was supplied as described in paragraph
3.2.
The diet offered to adults was replaced every day. The cages with parasitoids were
kept in a climate chamber (25 ± 2ºC, 75 ± 5% and 16:8 h) and the survival of adults in
each cage was recorded daily until all adults were dead. Each treatment included 30
parasitoids of each sex.
3.5. Data analysis
3.5.1. Longevity
Longevity means, and standard errors were calculated separately by P. concolor sexes
when provided with different foods. All analyses and graphics were performed with R
software (R core team 2014). For survival analysis, the survfit function
[in survival package] was used to calculate estimates of the different survival curves
using the Kaplan–Meier method. The Kaplan–Meier survival curves of individual
treatments were compared with the log-rank test survdiff. A non-parametric Cox
proportional hazards model (coxph function [in survival package]) was used to analyze
the effect of the food source treatment on parasitoid survival. The assumption of
proportional hazards was tested by visual inspections using the cox.zph function of the
same package. Cox proportional hazards model fitted using “food resource” the
proportional hazard assumption was accepted, and nonlinearity was not detected. No
influential observations were detected (all dfbetas <1).
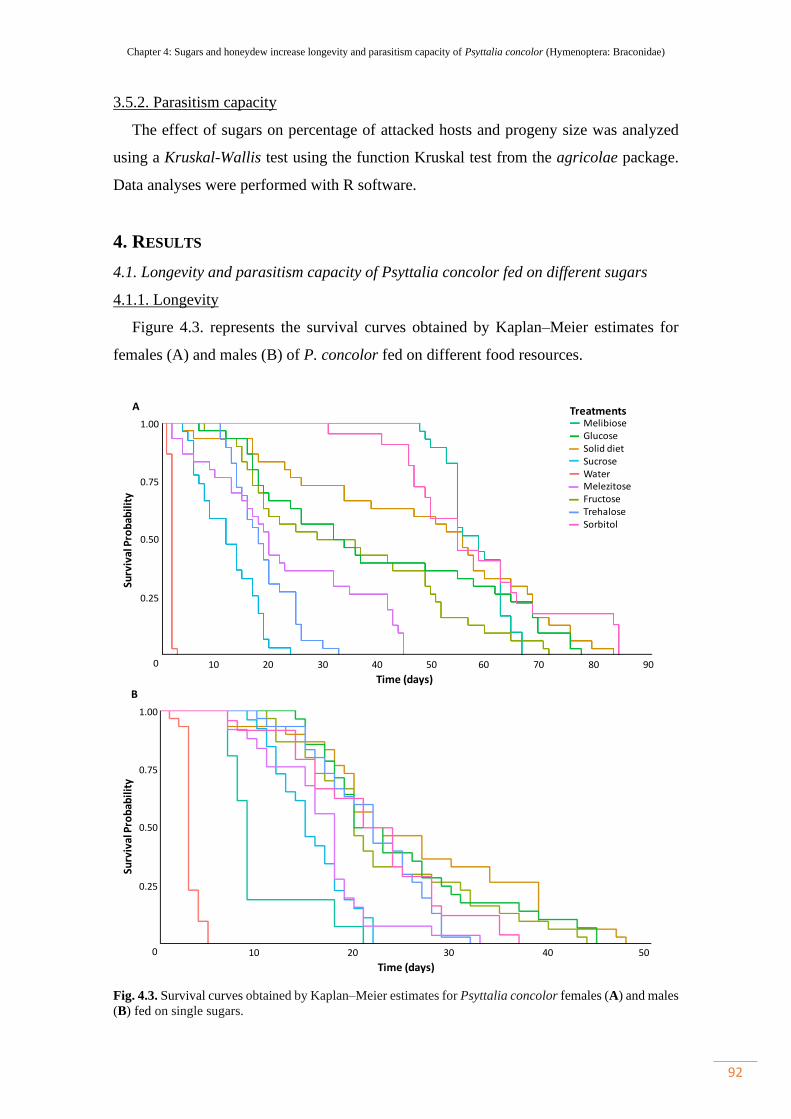
Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
92
3.5.2. Parasitism capacity
The effect of sugars on percentage of attacked hosts and progeny size was analyzed
using a Kruskal-Wallis test using the function Kruskal test from the agricolae package.
Data analyses were performed with R software.
4. RESULTS
4.1. Longevity and parasitism capacity of Psyttalia concolor fed on different sugars
4.1.1. Longevity
Figure 4.3. represents the survival curves obtained by Kaplan–Meier estimates for
females (A) and males (B) of P. concolor fed on different food resources.
Fig. 4.3. Survival curves obtained by Kaplan–Meier estimates for Psyttalia concolor females (A) and males
(B) fed on single sugars.
Surv
ival
Pro
bab
ilit
y
1.00
0.75
0.50
0.25
Time (days)
A TreatmentsMelibioseGlucoseSolid dietSucroseWaterMelezitoseFructoseTrehaloseSorbitol
10 20 30 40 50 60 70 80 900
10 20 30 40 50
Surv
ival
Pro
bab
ilit
y
1.00
0.75
0.50
0.25
Time (days)
B
0

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
93
The Cox proportional hazards model showed that death hazards were significantly
different among diet treatments for females (χ2 = 412, df=8, p < 0.001) and males (χ2 =
409 df = 8, p < 0.001). For females, sorbitol reduced the risk of death when compared
with all the sugars and solid diet but not significantly, with the exception of melibiose
(35% reduction, p < 0.01). Likewise, melibiose does not significantly reduced the risk of
death when compared with all remaining sugars and solid diet. However, it reduced the
risk of death when compared with only water (>99 % reduction, p < 0.05). For solid diet,
the results indicated that this treatment makes a smaller contribution to the difference in
daily hazard of death when compared with other food sources by a factor of exp(beta) ≈
1%, which is not a significant contribution. On the other hand, glucose reduced
significantly the risk of death when compared with sucrose (2% reduction, p < 0.05) (Fig.
4.3. A).
Considering males, solid diet reduced the risk of death when compared with all sugars
but not significantly. Glucose reduced the risk of death when compared with all the food
sources but only significantly when compared with sucrose (31% reduction, p < 0.001)
and melibiose (< 1% reduction, p < 0.01). Similarly, fructose reduced significantly the
risk of death when compared with trehalose (18% reduction, p < 0.01) and melezitose
(17% reduction, p< 0.01) (Fig. 4.3. B).
The highest longevity of females fed on sugars solution was observed on sorbitol
(58.91 ± 3.26 days), melibiose (58.59 ± 1.04 days), solid diet (positive control, 48.87 ±
4.25 days) and glucose (40.13 ± 4.29 days) (Fig. 4.4. A). Males fed with diet (positive
control, 26.33 ± 2.03 days), glucose (24.82 ± 1.77 days) and fructose (23.23 ± 1.72 days)
presented the highest longevity (Fig. 4.4. B).
4.1.2. Parasitism capacity
The effects on parasitism capacity parameters resulted from each food tested are
summarized in Table 4.1. The percentage of attacked hosts was higher than 83% except
for parasitoids feed on melezitose (48.90 %). Sucrose originated the highest percentage
of attacked hosts (94.95 %), being only significantly different from melezitose (Table
4.1.). The results obtained for the percentage of progeny size were more variable. Solid
diet (control treatment) showed a higher value for the percentage of parasitism (74.45 %)
being only significantly different from the sugar Melezitose, which was not significantly
different from Melibiose (Table 4.1.).
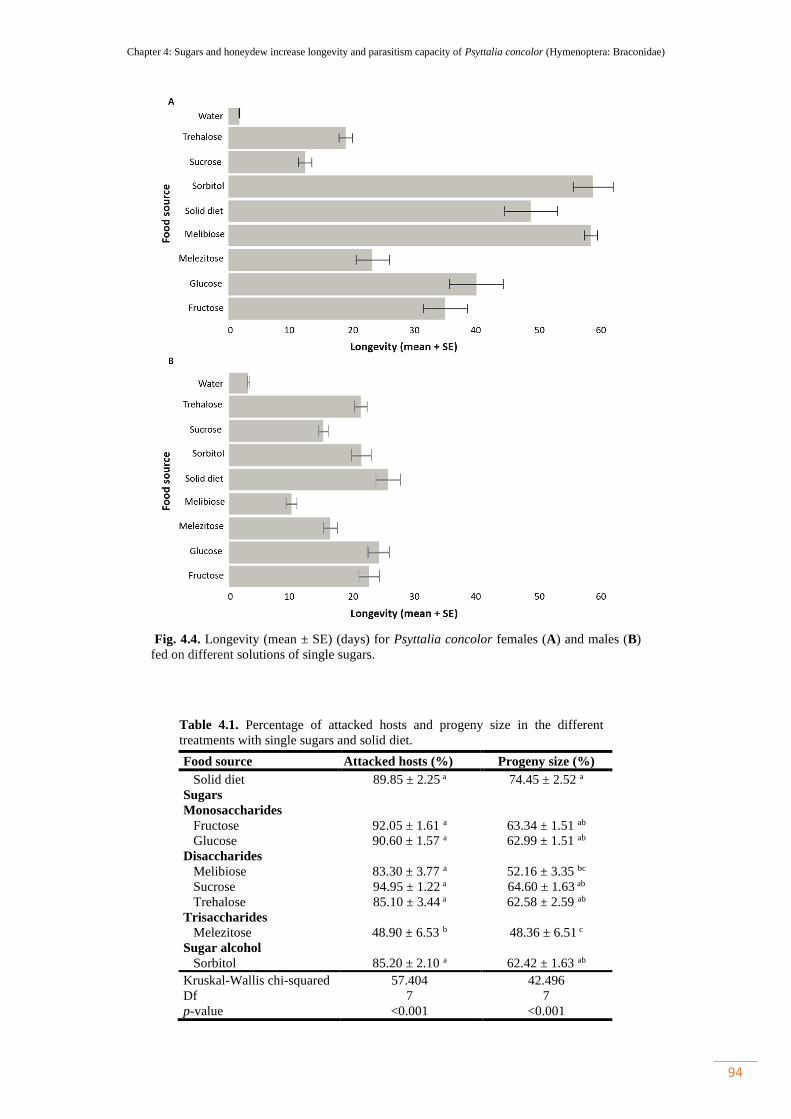
Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
94
Fig. 4.4. Longevity (mean ± SE) (days) for Psyttalia concolor females (A) and males (B)
fed on different solutions of single sugars.
Table 4.1. Percentage of attacked hosts and progeny size in the different
treatments with single sugars and solid diet.
Food source Attacked hosts (%) Progeny size (%)
Solid diet 89.85 ± 2.25 a 74.45 ± 2.52 a
Sugars Monosaccharides
Fructose 92.05 ± 1.61 a 63.34 ± 1.51 ab
Glucose 90.60 ± 1.57 a 62.99 ± 1.51 ab
Disaccharides
Melibiose 83.30 ± 3.77 a 52.16 ± 3.35 bc
Sucrose 94.95 ± 1.22 a 64.60 ± 1.63 ab
Trehalose 85.10 ± 3.44 a 62.58 ± 2.59 ab
Trisaccharides
Melezitose 48.90 ± 6.53 b 48.36 ± 6.51 c
Sugar alcohol
Sorbitol 85.20 ± 2.10 a 62.42 ± 1.63 ab
Kruskal-Wallis chi-squared 57.404 42.496
Df 7 7
p-value <0.001 <0.001
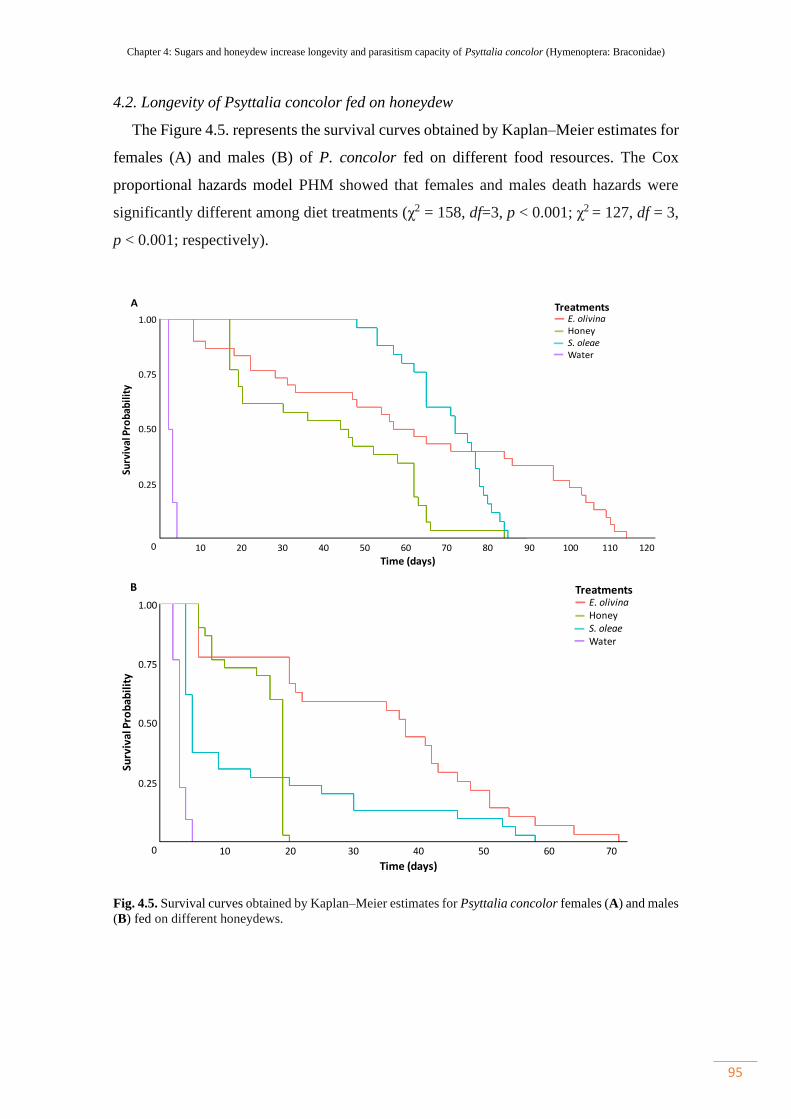
Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
95
4.2. Longevity of Psyttalia concolor fed on honeydew
The Figure 4.5. represents the survival curves obtained by Kaplan–Meier estimates for
females (A) and males (B) of P. concolor fed on different food resources. The Cox
proportional hazards model PHM showed that females and males death hazards were
significantly different among diet treatments (χ2 = 158, df=3, p < 0.001; χ2 = 127, df = 3,
p < 0.001; respectively).
Fig. 4.5. Survival curves obtained by Kaplan–Meier estimates for Psyttalia concolor females (A) and males
(B) fed on different honeydews.
Surv
ival
Pro
bab
ility
1.00
0.75
0.50
0.25
Time (days)
A TreatmentsE. olivinaHoneyS. oleaeWater
10 20 30 40 50 60 70 80 90 100 110 120 0
Surv
ival
Pro
bab
ilit
y
10 20 30 40 50 60 70
1.00
0.75
0.50
0.25
Time (days)
B
0
TreatmentsE. olivinaHoneyS. oleaeWater
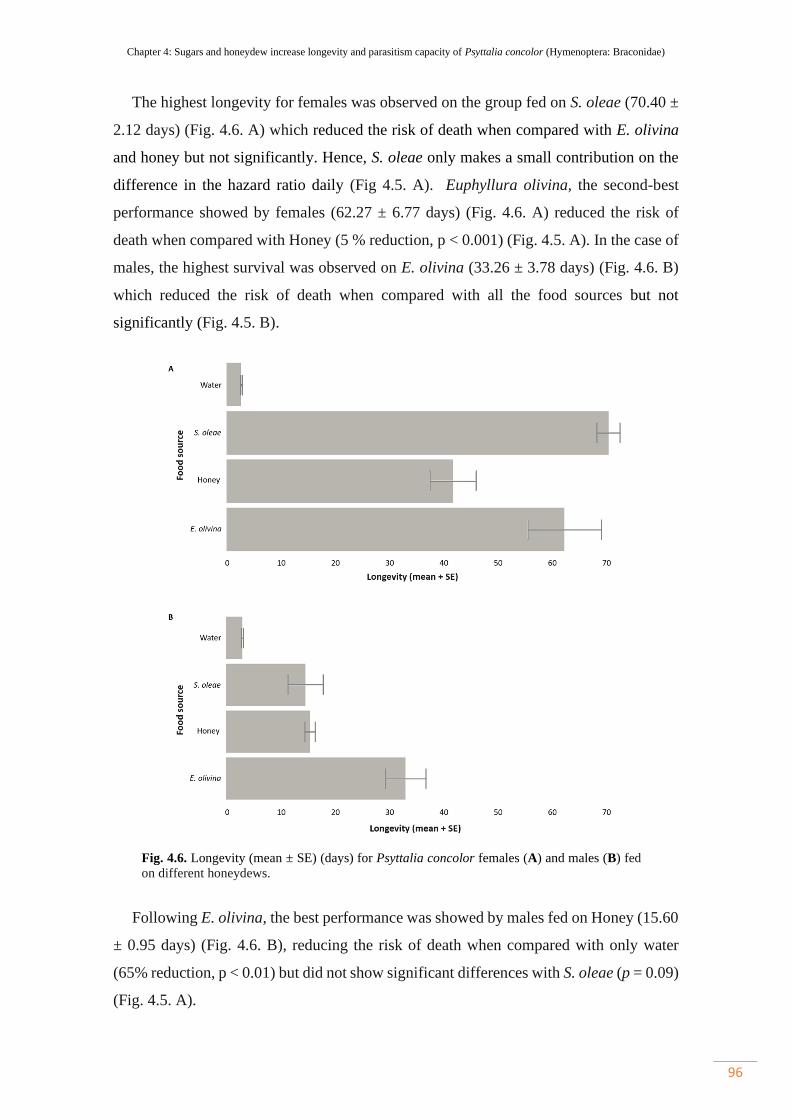
Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
96
The highest longevity for females was observed on the group fed on S. oleae (70.40 ±
2.12 days) (Fig. 4.6. A) which reduced the risk of death when compared with E. olivina
and honey but not significantly. Hence, S. oleae only makes a small contribution on the
difference in the hazard ratio daily (Fig 4.5. A). Euphyllura olivina, the second-best
performance showed by females (62.27 ± 6.77 days) (Fig. 4.6. A) reduced the risk of
death when compared with Honey (5 % reduction, p < 0.001) (Fig. 4.5. A). In the case of
males, the highest survival was observed on E. olivina (33.26 ± 3.78 days) (Fig. 4.6. B)
which reduced the risk of death when compared with all the food sources but not
significantly (Fig. 4.5. B).
Fig. 4.6. Longevity (mean ± SE) (days) for Psyttalia concolor females (A) and males (B) fed
on different honeydews.
Following E. olivina, the best performance was showed by males fed on Honey (15.60
± 0.95 days) (Fig. 4.6. B), reducing the risk of death when compared with only water
(65% reduction, p < 0.01) but did not show significant differences with S. oleae (p = 0.09)
(Fig. 4.5. A).

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
97
5. DISCUSSION
Psyttalia concolor have been widely used for biological control of a diversity of fruit
fly pests associated to a mass rearing. The sugars and honeydew food sources tested in
this work presented a variation with respect to their effect on females and males parasitoid
life span. These results showed that: (1) females and males of P. concolor had different
preferences or needs for sugars, and (2) sugars benefited more females than males when
the same sugar was supplied. Similarly, Williams III et al. (2015) found that females of
P. lounsburyi (Silvestri) (Hymenoptera: Braconidae) presented higher longevity than
males. It is possible that this higher longevity observed on females is due to a higher
nutritional requirement than males in order to provide energy for host forage and
oviposition (Zhang et al., 2014).
Females of P. concolor lived more when fed on sugars typically present in
Mediterranean nectars (and only sporadically found in honeydew) (Petanidou, 2005), i.e.,
sorbitol and melibiose. In addition to its nutritional value, Burke and Waddell (2011)
showed that sorbitol is a sugar which provides memory persistence, an important ability
to the reinforcement of appetitive memory, even though it is not considered as sweet
sugar. Moreover, the accumulation of sorbitol in parasitoids is also associated with a
tolerance to low temperatures (Bale, 2002), which benefits the presence of parasitoids in
agroecosystems when winter is more severe. Following these findings, flowers with
nectar containing sorbitol and melibiose should be present or preserved in olive grove,
such as Asparagus acutifolius L. (Asparagaceae) and Convolvulus arvensis L.
(Convolvulaceae) (Petanidou, 2005; Nave et al., 2017b)
In this study, P. concolor fed on sucrose showed lower longevity when compared to the
other tested sugars and the positive control. In contrast, other studies evidenced the
positive effect of sucrose-feeding in other braconids, such as Cotesia glomeratae (L.)
(Lee and Heimpel, 2008), Chelonus elaeaphilus Silvestri (Nave et al., 2017a) and P.
lounsburyi (Williams III et al., 2015). This finding showed that longevity responses to
sugars vary between species, as well as, the ability to digest and synthesize sugars (see
Wäckers, 1999; Wäckers et al., 2006; Tompkins, Wratten and Wäckers, 2010).
In synovigenic species, such as P. concolor, the percentage of attacked hosts is an
important factor for biological control efficiency. In our study, females fed on sucrose
increased the percentage of attacked hosts and parasitism rate comparatively to the other
sugars offered, suggesting that this sugar might quickly replace the energy spent on

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
98
reproduction. Interestingly, the sugar melezitose decreased the percentage of attacked
hosts (significantly) and the parasitism rate. This effect can be ascribed to the fact that
melezitose is a sugar that quickly crystallizes, which difficult sugar uptake by the
parasitoid (Wäckers, 2000).
In general, parasitism rate observed in P. concolor fed on sugars averaged 50-60%,
which indicates a risk of egg limitation in the field by P. concolor, as similarly observed
in other synovigenic parasitoids (see Heimpel and Rosenheim, 1998). The nutrients
necessary for egg production are partially acquired by proteins and lipids, while
carbohydrates enhance the lifespan fecundity of parasitoids (Benelli et al., 2017). It is
important to consider that parasitoid needs other basic nutrients such as vitamins, mineral
salts, steroids, nucleic acids for normal development and reproduction.
Glucose and fructose, two of the main sugars present in various floral nectars and
honeydews, provided the best results for longevity on males. These results indicated that
males of P. concolor appeared to be a more generalist feeder than females, suggesting
that hexoses-dominant nectars would benefit this gender. These sugars can be
offered/preserved in the olive groves through wild flowering plant with hexose-dominant
nectars such as Raphanus raphanistrum (Brassicaceae) and Malva neglecta (Malvaceae)
or hexose-rich nectars, such as Daucus carota (Apiaceae), Conopodium majus
(Apiaceae), Foeniculum vulgare (Apiaceae), Asparagus acutifolius (Asparagaceae) and
Anarrhinum bellidifolium (Scrophulariaceae) (Nave et al., 2017a).
Interestingly, males fed on melibiose had its longevity reduced: males survived on
average 10 days on melibiose, while on glucose and fructose, the longevity was more than
doubled. This is consistent with a study on Cotesia glomerata (L.) (Hymenoptera:
Braconidae) about quantifying food intake which suggested that, contrarily to melibiose,
glucose and fructose were immediately converted into energy, fulfilling the nutritional
needs of parasitoids (Hausmann, Wäckers and Dorn, 2005).
Currently, the available research on parasitoid longevity when feeding on honeydew
have inconsistent results since some studies indicated that honeydew did not increase
parasitoid survival, while others showed that honeydew increased its survival but was an
inferior food source when compared to nectars or honey solutions (reviewed by Wäckers
and van Rijn, 2005). However, there is a high variation of honeydew types with different
carbohydrate contents which provides different longevities on parasitoids (Tena et al.,
2018). Also, lower longevity may be attributed to high viscosity and to the tendency of
some honeydew sugars to rapidly crystallize (Faria, Wäckers and Turlings, 2008). In our

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
99
study, honeydew was not an inferior food source from the perspective of enhancing
survival of females and males of P. concolor, presenting even higher values than single
sugars. Honeydew from S. oleae and E. olivina was a good food source for females,
presenting superior longevity than honey solution (positive control). However, for males,
E. olivina exhibited the best longevity performance, with S. oleae and honey showing
similar results. In contrast, Villa et al. (2017) found that E. olivina and S. oleae honeydew
reduced the death hazard of Elasmus flabellatus (Fonscolombe) (Hymenoptera:
Eulophidae) females comparatively to the other tested flowers but not to honey solution
(positive control).
Results from Wang et al. (2011) indicated that S. oleae honeydew also increased the
longevities of parasitoids P. humilis (Silvestri) (Hymenoptera: Braconidae) and
Scutellista caerulea (Fonsc.) (Hymenoptera: Pteromalidae) with a single meal.
In the field, when flowers produce little, if any, nectar, E. olivina and S. oleae honeydew
can be the primary exogenous carbohydrate source available (Wäckers, 2005). However,
these food sources are usually only present for a few months and often at low densities
(Williams III et al., 2015). On the other hand, one of the advantages of these food sources
is related to the community of plants in this environment, since honeydew producers may
be associated to the crop itself, the weeds close to this culture or even the surrounding
vegetation (Wäckers, van Rijn and Heimpel, 2008). Thus, the presence of honeydew
when nectar is rare extends the survival of P. concolor (even as a single meal) and
potentially enhances the efficiency of biological control on B. oleae population.
A manipulation tactic that has been proposed as alternative of flowering plants
management involves the application of solutions rich in carbohydrates and/or proteins
to crops, to act as artificial food supplements also known as artificial food sprays or
artificial honeydews (Wade et al. 2008a, b; Tena et al. 2015). This strategy acquires
primary importance in crops where vegetation has been removed through farming
practices such as tillage and herbicide application, leading to a reduction of floral
resources (Walters, 2011). Furthermore, low density of plants during the Mediterranean
summer can eventually provide limited sugars resources for a limited period (due to low
quantity and quality of resources and detectability by the insects) (Segoli and Rosenheim,
2013; Tena et al., 2016; Kishinevsky et al., 2017).
In conclusion, it is worth to notice that experiments with caged individuals are not
necessarily representative of basic needs in the field, being an indication of the
importance of certain food sources for life span of parasitoids (Wäckers, 2001). Under

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
100
more realistic conditions, nutritional requirements of P. concolor are expected to be
higher due to their foraging behaviour and parasitizing hosts. Our findings suggest that a
good source of carbohydrates (sugar or honeydew) is important on sustainability of P.
concolor in the field, especially in the life and oviposition of female parasitoid. Having
this, it is important to provide sustainably food through flowering plants management or
artificial food supplementation.
Considering the benefits of sugar supplements to released parasitoids, it is important
to develop not only an effective field delivery system, like feeding containers, smears on
wooden stakes, foliage sprays, but also dedicate to the formulation of methodologies that
ensure the conservation and availability of these sugars in the field for a longer period of
time.
6. REFERENCES
Baker, H.G. and Baker, I. (1983). Floral nectar sugar constituents in relation to pollinator
type. In: C.E. Jones, and R.J. Little (Eds.), Handbook of pollination biology (pp. 117–
141). New York: Van Nostrand Reinhold.
Bale, J.S. (2002). Insects and low temperatures: from molecular biology to distributions
and abundance. Philosophical Transactions of the Royal Society B: Biological
Sciences, 357(1423): 849-862.
Benelli, G., Giunti, G., Tena, A., Desneux, N., Caselli, A. and Canale, A. (2017). The
impact of adult diet on parasitoid reproductive performance. Journal of Pest Science,
90: 807-823.
Bengochea, P., Budia, F., Viñuela, E. and Medina, P. (2014). Are kaolin and copper
treatments safe to the olive fruit fly parasitoid Psyttalia concolor? Journal of Pest
Science, 87: 351-359.
Burke, C.J. and Waddell, S. (2011). Remembering nutrient quality of sugar in
Drosophila. Current Biology, 21: 746-750.
Daane, K.M., Johnson, M.W. (2010). Olive fruit fly: managing an ancient pest in modern
times. Annual Review of Entomology 55: 151-169.
Faria, C.A., Wäckers, F.L. and Turlings, T.C.J. (2008). The nutritional value of aphid
honeydew for non-aphid parasitoids. Basic and Applied Ecology, 9: 286-297.
Hausmann, C.H., Wäckers, F.L. and Dorn, S. (2005). Sugar convertibility in the
parasitoid Cotesia glomerata (Hymenoptera: Braconidae). Archives of Insect
Biochemistry and Physiology, 60: 223-229.
Heimpel, G.E. and Rosenheim, J.A. (1998). Egg limitation in parasitoids: a review of the
evidence and a case study. Biological Control, 11(2): 160-168.
Hogervorst, P.A.M., Wäckers, F.L. and Romeis, J. (2007a). Effects of honeydew sugar
composition on the longevity of Aphidius ervi. Entomologia Experimentalis et
Applicata, 122: 223-232.
Hogervorst, P.A.M., Wäckers, F.L. and Romeis, J. (2007b). Detecting nutritional state
and food source use in field-collected insects that synthesize honeydew
oligosaccharides. Functional Ecology, 21: 936-946.

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
101
Jacas, J.A., Viñuela, E. (1994). Analysis of a lab method to test the effects of pesticides
on adult females of Opius concolor, a parasitoid of the olive fruit fly Bactrocera
oleae. Biocontrol Science and Technology, 4: 147-154.
Jervis, M. (1998). Functional and evolutionary aspects of mouthpart structure in
parasitoid wasps. Biological Journal of the Linnean Society, 63: 461-493.
Kishinevsky, M., Cohen, N., Chiel, E., Wajnberg, E. and Keasar, T. (2017). Sugar feeding
of parasitoids in an agroecosystem: effects of community composition, habitat and
vegetation. Insect Conservation and Diversity, 11: 50-57.
Lee, J.C. and Heimpel, G.E. (2008). Floral resources impact longevity and oviposition
rate of a parasitoid in the field. Journal of Animal Ecology, 77: 565-572.
Lee, J. C., Heimpel, G. E. and Leibee, G.L. (2004). Comparing floral nectar and aphid
honeydew diets on the longevity and nutrient levels of a parasitoid wasp.
Entomologia Experimentalis et Applicata, 111: 189-199.
Lewis, W.J., Stapel, J.O, Cortesero, A.M. and Takasu, K. (1998). Understanding how
parasitoids balance food and host needs: importance to biological control. Biological
Control, 11: 175-183.
Lucchetta, P., Bernstein, C., Théry, M., Lazzari, C. and Desouhant, E. (2008). Foraging
and associative learning of visual signals in a parasitic wasp. Animal Cognition, 11:
525-533.
Loumou, A. Giourga, C. (2003). Olive groves: The life and identity of the Mediterranean.
Agriculture and Human Values, 20: 87–95.
Miranda, M.A., Miquel, M., Terrassa, J., Melis, N. and Monerris, M. (2008). Parasitism
of Bactrocera oleae (Diptera; Tephritidae) by Psyttalia concolor (Hymenoptera;
Braconidae) in the Balearic Islands (Spain). Journal of Applied Entomology, 132(9-
10): 798-805.
Nafziger, T.D. and Fadamiro, H.Y. (2011). Suitability of some farmscaping plants as
nectar sources for the parasitoid wasp, Microplitis croceipes (Hymenoptera:
Braconidae): effects on longevity and body nutrients. Biological Control, 56: 225-
229.
Nave, A., Gonçalves, F., Nunes, F.M., Campos, M. and Torres, L. (2017a). Evaluating
potential sugar food sources from the olive grove agroecosystems for Prays oleae
parasitoid Chelonus elaeaphilus. Biocontrol Science and Technology, 27: 686-695.
Nave, A., Crespí, A.L., Gonçalves, F., Campos, M. and Torres, L. (2017b). Native
Mediterranean plants as potential food sources for natural enemies of insect pests in
olive groves. Ecological Research, 32(4): 459-459.
Olson, D.M., Fadamiro, H., Lundgren, J.G. and Heimpel, G.E. (2000). Effects of sugar
feeding on carbohydrate and lipid metabolism in a parasitoid wasp. Physiological
Entomology, 25: 17-26.
Petanidou, T. (2005). Sugars in Mediterranean floral nectars: an ecological and
evolutionary approach. Journal of Chemical Ecology, 31: 1065-1088.
Petanidou, T., Van Laere, A., Ellis, W.N. and Smets, E. (2006). What shapes amino acid
and sugar composition in Mediterranean floral nectars? Oikos, 115: 155-169.
R core team 2014. R: a language and environment for statistical computing. R Foundation
for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/
Raspi, A. and Loni, A. (1994). Alcune note sull’allevamento massale di Opius concolor
Szépligeti (Hym.: Braconidae) e su recenti tentative d’introduzione della specie in
Toscana e Liguria [Some notes on the breeding of massale Opius concolor Szépligeti
(Hym.: Braconidae) and recent tentative introduction of the species in Tuscany and
Liguria]. Frustula Entomologica, 30, 135-145.

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
102
Rivero, A. and Casas, J. (1999). Incorporating physiology into parasitoid behavioral
ecology: the allocation of nutritional resources. Researches on Population Ecology,
41: 39-45.
Russell, M. (2015). A meta-analysis of physiological and behavioral responses of
parasitoid wasps to flowers of individual plant species. Biological Control, 82: 96-
103.
Segoli, M. and Rosenheim, J.A. (2013). Spatial and temporal variation in sugar
availability for insect parasitoids in agricultural fields and consequences for
reproductive success. Biological Control, 67: 163-169.
Snart, C.J.P., Kapranas, A., Williams, H., Barrett, D.A. and Hardy, I.C.W.
(2018). Sustenance and Performance: Nutritional Reserves, Longevity, and Contest
Outcomes of Fed and Starved Adult Parasitoid Wasps. Frontiers in Ecology and
Evolution, 6.
Tena, A., Pekas, A., Cano, D., Wäckers, F.L. and Urbaneja, A. (2015). Sugar provisioning
maximizes the biocontrol service of parasitoids. Journal of Applied Ecology, 52:
795-804.
Tena, A., Wäckers, F.L., Heimpel, G.E., Urbaneja, A. and Pekas, A. (2016). Parasitoid
nutritional ecology in a community context: the importance of honeydew and
implications for biological control. Current Opinion in Insect Science, 14: 100-104.
Tena, A., Senft, M., Desneux, N., Dregni, J. and Heimpel, G.E. (2018). The influence of
aphid-produced honeydew on parasitoid fitness and nutritional state: a comparative
study. Basic and Applied Ecology, 29: 55-68.
Tompkins, J.M.L., Wratten, S.D. and Wäckers, F.L. (2010). Nectar to improve parasitoid
fitness in biological control: does the sucrose:hexose ratio matter? Basic and Applied
Ecology, 11: 264-271.
Villa, M., Santos, S.A.P., Mexia, A., Bento, A. and Pereira, J.A. (2017). Wild flower
resources and insect honeydew are potential food items for Elasmus flabellatus.
Agronomy for Sustainable Development, 37:15.
Wäckers, F.L. (1999). Gustatory response by the hymenopteran parasitoids Cotesia
glomerata to a range of nectar and honeydew sugars. Journal of Chemical Ecology,
25(12): 2863-2877.
Wäckers, F.L. (2000). Do oligosaccharides reduce the suitability of honeydew for
predators and parasitoids? A further facet to the function of insect-synthesized
honeydew sugars. Oikos, 90: 197-201.
Wäckers, F.L. (2001). A comparison of nectar- and honeydew sugars with respect to their
utilization by the hymenopteran parasitoid Cotesia glomeratae. Journal of Insect
Physiology, 47: 1077-1084.
Wäckers, F. L. (2005). Suitability of (extra-)floral nectar, pollen, and honeydew as insect
food sources. F.L. Wäckers, P.C.J. van Rijn and J. Bruin (Eds.). Plant-Provided Food
for Carnivorous Insects, A Protective Mutualism and its Applications (pp. 17-74).
Cambridge: Cambridge University Press.
Wäckers, F.L., Lee, J., Heimpel, G., Winkler, K. and Wagenaar, R. (2006).
Hymenopteran parasitoids synthesize honeydew-specific’ oligosaccharides.
Functional Ecology, 20: 790-798.
Wäckers, F.L., van Rijn, P.C.J. and Heimpel, G.E. (2008). Honeydew as a food source
for natural enemies: Making the best of a bad meal? Biological Control: 45, 176-184.
Wade, M.R., Hopkinson, J.E. and Zalucki, M.P. (2008a). Influence of food
supplementation on the fitness of two biological control agents: a predatory nabid
bug and a bollworm pupal parasitoid. Journal of Pest Science, 81: 99-107.

Chapter 4: Sugars and honeydew increase longevity and parasitism capacity of Psyttalia concolor (Hymenoptera: Braconidae)
103
Wade, M.R., Zalucki, M.P., Wratten, S.D. and Robinson, K.A. (2008b). Conservation
biological control of arthropods using artificial food sprays: current status and future
challenges. Biological Control, 45: 185-199.
Walters, A.S. (2011). Weed management systems for no-tillage vegetable production. In
Soloneski S. and Larramendy M.L. (Eds.), Herbicides: theory and applications (pp.
2-40). Rijeka: IntechOpen.
Wang, X.-G., Johnson, M.W., Opp, S.B., Krugner, R. and Daane, K.M.
(2011). Honeydew and insecticide bait as competing food resources for a fruit fly
and common natural enemies in the olive agroecosystem. Entomologia
Experimentalis et Applicata, 139: 128-137.
Williams III, L., Deschodt, P., Pointurier, O. and Wyckhuys, K.A.G. (2015). Sugar
concentration and timing of feeding affect feeding characteristics and survival of a
parasitic wasp. Journal of Insect Physiology, 79: 10-18.
Winkler, K., Wäckers, F., Bukovinszkine-Kiss, G. and van Lenteren, J. (2006). Sugar
resources are vital for Diadegma semiclausum fecundity under field conditions.
Basic and Applied Ecology, 7: 133-140.
Zhang, G., Zimmermann, O. and Hassan S.A. (2004). Pollen as a source of food for egg
parasitoids of the genus Trichogramma (Hymenoptera: Trichogrammatidae),
Biocontrol Science and Technology, 14: 201-209.
Zhang, Y., Yang, N., Wang, J. and Wan, F. (2014). Effect of six carbohydrate sources on
the longevity of a whitefly parasitoid Eretmocerus hayati (Hymenoptera:
Aphelinidae). Journal of Asia-Pacific Entomology, 17: 723-728.

104
Chapter 5
Amino acids diet influence
survival and reproductive
capacity of Psyttalia concolor
(Hymenoptera: Braconidae), a
parasitoid of tephritid flies?

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
105
Chapter 5:
AMINO ACIDS DIET INFLUENCE SURVIVAL AND
REPRODUCTIVE CAPACITY OF PSYTTALIA CONCOLOR
(HYMENOPTERA: BRACONIDAE), A PARASITOID OF
TEPHRITID FLIES?
1. ABSTRACT
Insects differ in their nutritive preferences, generally concerning amino acids. The
composition of food sources, usually in form of pollen, nectar and honeydew, interfere
with adult parasitoid fitness. Psyttalia concolor (Szépligeti, 1910) (Hymenoptera:
Braconidae) is one of the major natural enemy associated with the olive fruit fly,
Bactrocera oleae (Rossi) (Diptera: Tephritidae). In the olive grove, while adults are non-
predatory and consume plant-derived foods and honeydew, larvae of P. concolor parasite
the eggs of an olive fruit fly. The effect of the constituents of these food source exploited
in olive orchards by adults in survival and reproductive success are not precisely known.
The present study aims to determine the effect of three amino acids (tryptophan,
phenylalanine and proline) and a mixture of the same amino acids on longevity and
reproductive parameters of parasitoid P. concolor, in order to analyze their suitability as
potential food sources. Cox’s proportional hazard regression models were used to analyse
the differences between parasitoid survivals. Females fed with phenylalanine and proline
showed a significant reduction of death hazard, while for males the risk of dying was
reduced with the consumption of Mix and phenylalanine. We also found that sugar
solution increased the reproductive performance more than amino acids.
Our results indicate that an exact combination of sugars and amino acids is important
for the success of parasitoid as a biological control agent. However, adult food is not the
only important resource for P. concolor; alternative hosts and refuges from adverse biotic
and abiotic conditions are also important for parasitoid-pest interaction.
Keywords: nectar, parasitoids, phenylalanine, proline, protein intake, tryptophan
2. INTRODUCTION
The olive fruit fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae) is a monophagous
frugivorous pest which feeds exclusively from Olea species and is a key pest of the olive

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
106
tree (Olea europaea L. (Lamiales: Oleaceae)). Losses caused by this pest goes from
productivity losses due to fruit drop and fruit rot to a decrease on olive products quality
and composition (see review, Malheiro et al., 2015). Traditionally, control is performed
with resource to pesticides; however, more sustainable management programs for this
pest have been employed, such as indigenous natural enemies (e.g. Eupelmus urozonus
Dalm. (Eupelmidae), Pnigalio mediterraneus Walk. (Eulophidae) and Eurytoma martellii
Dom) (Daane et al., 2015). Also, inundative and inoculative releases have been widely
recommended (Daane and Johnson 2010). Among these, Psyttalia concolor (Szépligeti,
1910) (Hymenoptera: Braconidae) has been used for the biological control of olive fruit
fly in Mediterranean basin (e.g. Spain (Miranda et al., 2008), Italy (Raspi and Loni, 1994)
and California (Yokoyama, Rendón and Sivinski, 2008a; b). This endoparasitoid is a
synovigenic koinobiont larval-pupal reported to attack 14 tephritids host species
(Diptera), including B. oleae (Benelli and Canale, 2012).
While parasitoids larvae are strictly carnivorous, adults require nectar, pollen and
honeydew as source of energy and protein for reproduction (Rivero and Casas, 1999). In
fact, synovigenic females, which emerge with some immature eggs, have high protein
demands associated to egg production and maturation during their reproductive lifetime
(Zhang et al., 2004; Mondy et al., 2006). Having this, amino acids-rich food resources
would be particularly important for egg production by parasitoid P. concolor.
There is increasing evidence that the provision of flowering plants in agroecosystems
enhance the fitness of parasitoids and consequently support biological control (Arnó,
Oveja and Gabarra, 2018). Amino acids, as well as sugars, have been identified in various
floral nectars, pollens and honeydews. In nectar, amino acids are the second component
more abundant being basically composed by essential, non-essential and non-protein
amino acid (Petanidou et al., 2006). The most abundant amino acids of the nectars are
alanine, serine, proline, tyrosine, phenylalanine, valine and acid aspartic. However, other
amino acids have also been identified (Gardener and Gillman, 2001a; Carter et al., 2006).
In floral nectars, certain amino acids are found more frequently than others when
compared with extra-floral nectars even for the same plant. Cysteine group (cysteine,
cystine and cysteic acid), lysine, asparagine, and tyrosine occur with more frequency in
extra-floral nectars, while glutamic acid appears in floral nectar (Baker, Opler and Baker,
1978). In addition, non-protein amino acids are found with more frequency in extra-floral
nectars (Baker, Opler and Baker, 1978). Besides nectar, honeydew offers amino acids

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
107
but, here, the composition depends on the sap-feeders and it is normally composed of
non-essential amino acids (Sandström and Moran, 2001). Particularly, these compounds
are glutamine, glutamic acid, asparagine, aspartic acid and serine (Fischer et al., 2002).
In pollen, unlike nectar and honeydew, proteins and free amino acids are the principal
components (Roulston and Cane, 2000). Pollen is particularly rich in glutamic acid,
aspartic acid, and proline (Lundgren and Wiedenmann, 2004; Szczesna, 2006). However,
other amino acids are present in equal or higher amounts (see Human and Nicolson, 2006;
Komosinska-Vassev et al., 2015). The nutritional value of proteins depends on the quality
and quantity of amino acid content present in floral resources. Therefore, the composition
of amino acids depends on plants species, population, age, time of day and the climate,
and in the same plant is determined by the type of secretion, floral or extra-floral nectar
(e.g. Petanidou, van Laere and Smets,1996; Petanidou, Goethals and Smets,2000). Also,
the production of amino acids differs in time and space (environmental and seasonal
condition), as well as according to agricultural practices (Baker and Baker, 1986). As an
example, amino acids concentration varies significantly with soil fertilizer (Gardene and
Gillman, 2001b). Moreover, the effect of different amino acids on insect life has been
analysed by many researchers. The composition of amino acids is important to insect taste
receptors (e.g., Wada et al., 2001). Indeed, several studies showed that insects are able to
distinguish between sugar-only composed nectars from sugar-amino acid mixtures (Alm
et al.,1990; Mevi-Schutz and Erhardt, 2003; Carter et al., 2006).
The aim of this study is to investigate the effect of three amino acids (phenylalanine,
proline, and tryptophan) on fitness of P. concolor. While tryptophan and phenylalanine
are essential amino acid for insects, proline is a common non-essential amino acid. All
these amino acids have been reported to occur in nectar, pollen, and honeydew (Blüthgen,
Gottberger and Fiedler, 2004; Human and Nicolson, 2006; Petanidou et al., 2006; Leroy
et al., 2011). We hypothesized that dietary supplementation with one or a mixture of
amino acids may be beneficial de life cycle of P. concolor for (1) increase its longevity;
and (2) enhance the efficacy of parasitation on B. oleae larvae.
3. MATERIAL AND METHODS
3.1. Insect rearing
Psyttalia concolor was reared on Ceratitis capitata (Wiedemann, 1824) (Diptera:
Tephritidae) in the Crop Protection Unit, ETSIA, Technical University of Madrid

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
108
following the methodology described by Jacas and Viñuela (1994). Both insects were
reared at 25 ± 2ºC temperature, 75±5% R. H. and 16:8h light: dark cycle.
3.2.Food sources
Three amino acids, L-Tryptophan (Trp) (Sigma-Aldrich; ≥ 98%), L-Phenylalanine
(Phe) (Sigma-Aldrich; ≥ 98%), and L-Proline (Pro) (Sigma-Aldrich; ≥ 99%) – were tested
singly and mixed (Mix) at a concentration of 0.1 mM. Al the amino acids (tested singly
or mixed) were dissolved in D(+)-Glucose (Sigma-Aldrich; ≥ 99.5%) at 1 M.
The amino acid concentration was selected based on experiments conducted by
Blüthgen and Fiedler (2004). Distilled water and D(+)-Glucose 1M were used as negative
and positive control, respectively.
3.3. Longevity
For longevity experiments, five females and five males of recently emerged (< 24 h)
P. concolor (Fig. 5.1. A) were placed in a plexiglass cage (12 cm in diameter x 5 cm
height) with a hole on the top (5.5 cm in diameter) covered with a net for ventilation. A
total of six replicate cages were tested in each treatment.
Figure 5.1. Detail of plexiglass cage used in the longevity tests:
A: Psyttalia concolor adults; B: feeding glass vials.
The amino acid + glucose solutions were offered in a glass vial (2 ml) with a strip of
sponge and overlaid with a strip of Parafilm® (Fig. 5.1. B). Glass vials were glued to the
base of the plexiglass containers with a piece of sticky putty and a similar vial with
distilled water was provided. The solutions were refilled on a daily basis and changed
A
B

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
109
every week. Mortality of P. concolor individuals was registered on a daily basis and dead
animals were removed from the cages and stored at –20ºC.
3.4. Reproduction
Thirty recently emerged (< 24 h) couples were placed individually in plexiglas cages,
similar to those used in longevity experiments, but with an additional hole (5.5 cm in
diameter) on the bottom of the cage covered with a net (Fig. 5.2. A). After seven days,
3rd instar larvae of C. capitata were offered to P. concolor at a proportion of 20
larvae/female (Fig. 5.2. B) by placing them on the top of a glass bottle covered with a
piece of Parafilm® (4 cm in diameter). The larvae were offered for one hour, according
to Bengochea et al. (2014) and this procedure was repeated every five days. Subsequently,
these larvae were transferred into Petri dishes and placed at 25 ± 2ºC, 75 ± 5% and 16: 8
h light: dark until emergence. The time elapsed from the pupal stage until emergence was
recorded. The percentage of attacked hosts (percentage of puparia without medfly
emergence) and the percentage of progeny size (percentage of parasitoids emerged from
parasitised puparia) were evaluated. Only 4 days of parasitization were used in data
analysis due to a need of parasitization earn by the female. A total of 30 females were
tested individually in each treatment.
Figure 5.2. A: Plexiglass cage used in the parasitation tests; B: 3rd instar larvae
of C. capitata offered to P. concolor;
A
B

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
110
3.5. Data analysis
Survival curves were represented for each treatment with Kaplan-Meier estimates
using the Surv and survfit functions from the survival package (Therneau, 2014) in R (R
core team, 2014. Two separated Cox Proportional Hazard Models (Cox PHM), using the
coxph function from the same package, were fitted for females and males in order to
analyze the effect of different food resources on the survival of adults. The proportional
hazard assumption (PHA) of the Cox regression was confirmed testing the no correlation
between the Schoenfeld’s residuals and the survival time using the cox.zph function of
the same package and plotting the smoothed Schoenfeld’s residuals against the different
levels of the categorical variable (Therneau and Grambsch, 2000). When the PHA was
not achieved, firstly the specific moment in which the PHA was violated was identified
in the smoothed Schoenfeld’s residual plots. Then an extra categorical variable (age at
death) with two levels (Y: parasitoids dying younger, O: parasitoids dying older) was
created in order to obtain different hazard ratios for parasitoids dying before and after the
specific point when the PHA was violated. Thus, different survival behaviors may be
identified and analyzed for new born and older parasitoids. The cluster function in model
formulas was used to indicate the cage in order to account with non-independent
observations.
The reproductive parameters calculated were: (a) mean of emerged parasitoid [±
Standard Error (SE)], (b) percentage of the attacked host [mean± Standard Error (SE)]
and (c) the percentage of progeny size [mean± Standard Error (SE)]. ANOVA function
was used to analyze the differences between amino acid treatments, followed by pairwise
comparison with Tukey’s Studentized Range Test (HSD) at p = 0.05.
4. RESULTS
4.1. Survival
Survival curves are represented in Fig. 5.3.. Amino acids feeding increased survival,
but the positive effect of feeding on a glucose solution was considerably higher, especially
in the case of males (Fig. 5.3.). Moreover, in general, female parasitoids lived longer than
males.
All females fed only on water dyed before any other female fed on the remaining
treatments and in the Cox PHM fitted for females the death hazard ratio was infinitive at
comparing the death risk with water and with the other treatments. For that reason, the
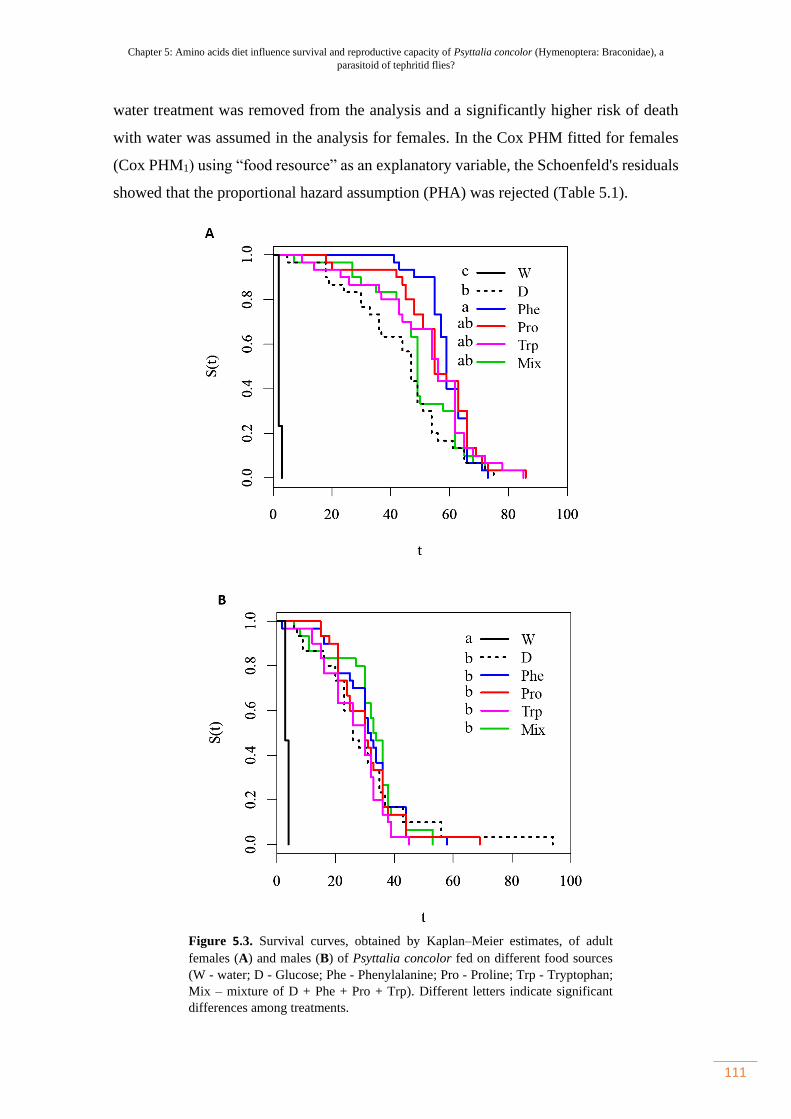
Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
111
water treatment was removed from the analysis and a significantly higher risk of death
with water was assumed in the analysis for females. In the Cox PHM fitted for females
(Cox PHM1) using “food resource” as an explanatory variable, the Schoenfeld's residuals
showed that the proportional hazard assumption (PHA) was rejected (Table 5.1).
Figure 5.3. Survival curves, obtained by Kaplan–Meier estimates, of adult
females (A) and males (B) of Psyttalia concolor fed on different food sources
(W - water; D - Glucose; Phe - Phenylalanine; Pro - Proline; Trp - Tryptophan;
Mix – mixture of D + Phe + Pro + Trp). Different letters indicate significant
differences among treatments.
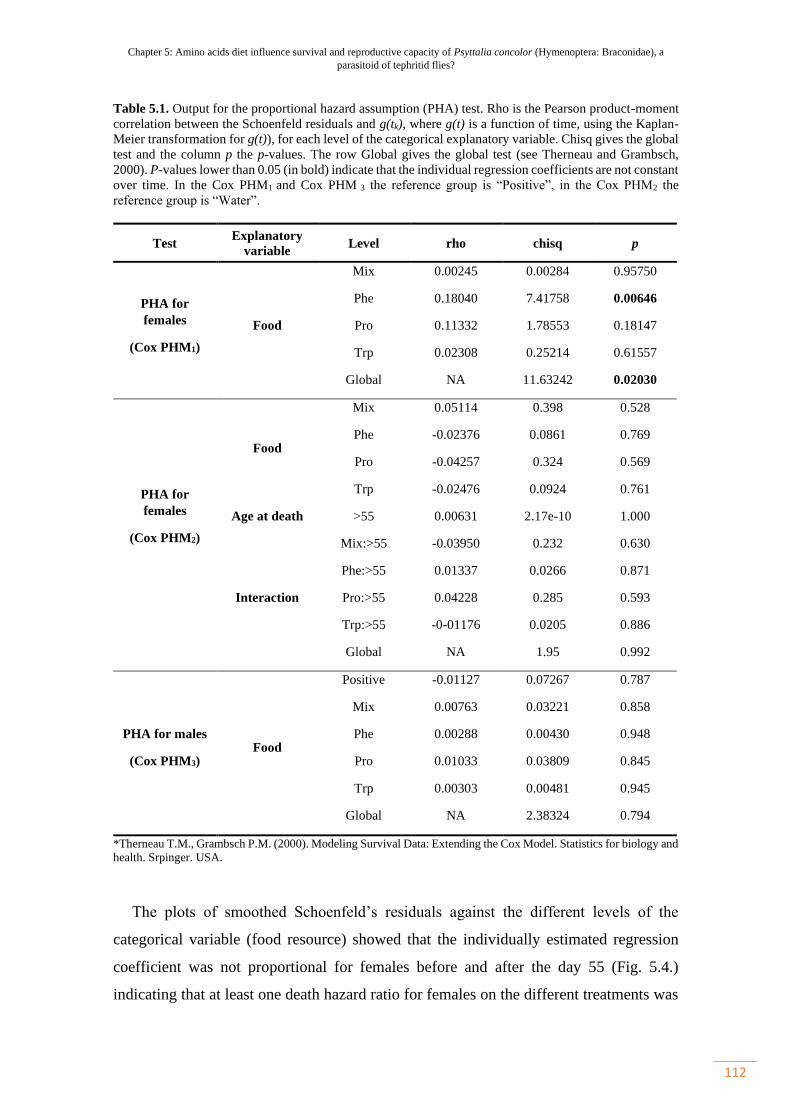
Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
112
Table 5.1. Output for the proportional hazard assumption (PHA) test. Rho is the Pearson product-moment
correlation between the Schoenfeld residuals and g(tk), where g(t) is a function of time, using the Kaplan-
Meier transformation for g(t)), for each level of the categorical explanatory variable. Chisq gives the global
test and the column p the p-values. The row Global gives the global test (see Therneau and Grambsch,
2000). P-values lower than 0.05 (in bold) indicate that the individual regression coefficients are not constant
over time. In the Cox PHM1 and Cox PHM 3 the reference group is “Positive”, in the Cox PHM2 the
reference group is “Water”.
Test Explanatory
variable Level rho chisq p
PHA for
females
(Cox PHM1)
Food
Mix 0.00245 0.00284 0.95750
Phe 0.18040 7.41758 0.00646
Pro 0.11332 1.78553 0.18147
Trp 0.02308 0.25214 0.61557
Global NA 11.63242 0.02030
PHA for
females
(Cox PHM2)
Food
Mix 0.05114 0.398 0.528
Phe -0.02376 0.0861 0.769
Pro -0.04257 0.324 0.569
Trp -0.02476 0.0924 0.761
Age at death >55 0.00631 2.17e-10 1.000
Interaction
Mix:>55 -0.03950 0.232 0.630
Phe:>55 0.01337 0.0266 0.871
Pro:>55 0.04228 0.285 0.593
Trp:>55 -0-01176 0.0205 0.886
Global NA 1.95 0.992
PHA for males
(Cox PHM3) Food
Positive -0.01127 0.07267 0.787
Mix 0.00763 0.03221 0.858
Phe 0.00288 0.00430 0.948
Pro 0.01033 0.03809 0.845
Trp 0.00303 0.00481 0.945
Global NA 2.38324 0.794
*Therneau T.M., Grambsch P.M. (2000). Modeling Survival Data: Extending the Cox Model. Statistics for biology and
health. Srpinger. USA.
The plots of smoothed Schoenfeld’s residuals against the different levels of the
categorical variable (food resource) showed that the individually estimated regression
coefficient was not proportional for females before and after the day 55 (Fig. 5.4.)
indicating that at least one death hazard ratio for females on the different treatments was

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
113
different before and after that day. Then, a new Cox PHM was fitted for females (Cox
PHM2) with an extra categorical variable (age at death) with two levels: females dying
before (level Young: Y) and after (level Old: O) the day 55. PHAs for the Cox PHM2
fitted for females using “food resource” and “age at death” as explanatory variables and
the Cox PHM fitted for males (Cox PHM3) using “food resource” as explanatory variable
were accepted (Table 5.1. and Fig. 5.5). The Cox PHM2 showed that the death hazard
ratio for females was significantly different when fed on different food resources
(likelihood ratio = 227.3, df = 9, p < 0.001).
The risk of dying at any time was reduced in females fed on Phe and Pro, revealing
significant differences when compared with the remaining treatments (Fig. S5.1 A), but
it did not differ between Phe and Pro (p = 0.5448). The risk also did not differ between
Glucose and Mix (p = 0.8694), Glucose and Trp (p = 0.7001) and Mix and Trp (p =
0.7309). The risk of dying for females after the day 55 compared with the risk of dying
before highly increased with all treatments (Glucose: 1.139e+10%, p < 2e-16; Mix:
1.002e+10%, p < 2e-16; Phe: 1.424e+09%, p < 2e-16; Pro: 4.161e+09%, p < 2e-16). This
increase was strongly higher with Phe than with the other treatments (Fig. S5.1. B) with
the exception of Pro which did not differ from Phe (p = 0.0994). Also, the risk with Pro
increased compared with Glucose (Fig. S5.1. B). The increase with the other treatments
did not differ between them (between Pro and Mix (p = 0.2070), between Pro and Trp (p
= 0.3297), between Mix and Glucose (p = 0.8520), between Mix and Trp (p = 0.789),
between Glucose and Trp (p = 0.6841).
The Cox PHM3 showed that the death hazard ratio for males fed on different food
resources was significantly different (likelihood ratio = 141.3, df = 5, p < 0.001). The risk
of dying at any time was reduced when males were fed on any treatment when compared
to the water treatment (negative control) (Fig. S5.1. C). The Mix and Phe foods reduced
the death hazard for males when compared with Trp treatment by 37.31% (p = 0.0199)
and 31.13% (p = 0.0027) respectively. The risk of death did not differ between the other
treatments (Mix and Glucose: p = 0.495; Mix and Phe: p = 0.7375; Mix and Pro: p =
0.6512; Glucose and Phe: p = 0.850; Glucose and Pro: p = 0.940; Glucose and Trp: p =
0.118; Phe and Pro: p = 0.8191; Pro and Trp: p = 0.339).

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a parasitoid of tephritid flies?
114
Figure 5.4. Schoenfeld’s residuals plotted against the different levels of the categorical variable “food resource” of the Cox PHMs fitted for adults of Psyttalia concolor (Cox
PHM1 for females: A, B, C, D; Cox PHM3 for males: E, F, G, H, I). An arrow in panel B shows the specific point when the PHA was violated.
Time
Bet
a(t)
fo
r F
oo
d_
trp
23 43 48 54 56 62 65 72
-15
-10
-50
51
01
5
Time
Bet
a(t)
fo
r F
oo
d_
pro
23 43 48 54 56 62 65 72
-15
-10
-50
51
01
5
Time
Bet
a(t)
fo
r F
oo
d_
ph
e
23 43 48 54 56 62 65 72
-15
-10
-50
51
01
5
Time
Bet
a(t)
fo
r F
oo
d_
mix
23 43 48 54 56 62 65 72
-15
-10
-50
51
01
5
A B C D
Time
Bet
a(t)
fo
r F
oo
d_
po
siti
ve
3.6 6.8 21 26 31 34 37 45
-10
0-5
00
50
10
0
Time
Bet
a(t)
fo
r F
oo
d_
mix
3.6 6.8 21 26 31 34 37 45
-10
0-5
00
50
10
0
TimeB
eta(
t) f
or
Fo
od
_p
he
3.6 6.8 21 26 31 34 37 45
-10
0-5
00
50
10
0
Time
Bet
a(t)
fo
r F
oo
d_
pro
3.6 6.8 21 26 31 34 37 45
-10
0-5
00
50
10
0
Time
Bet
a(t)
fo
r F
oo
d_
trp
3.6 6.8 21 26 31 34 37 45
-10
0-5
00
50
10
0
E F G H I
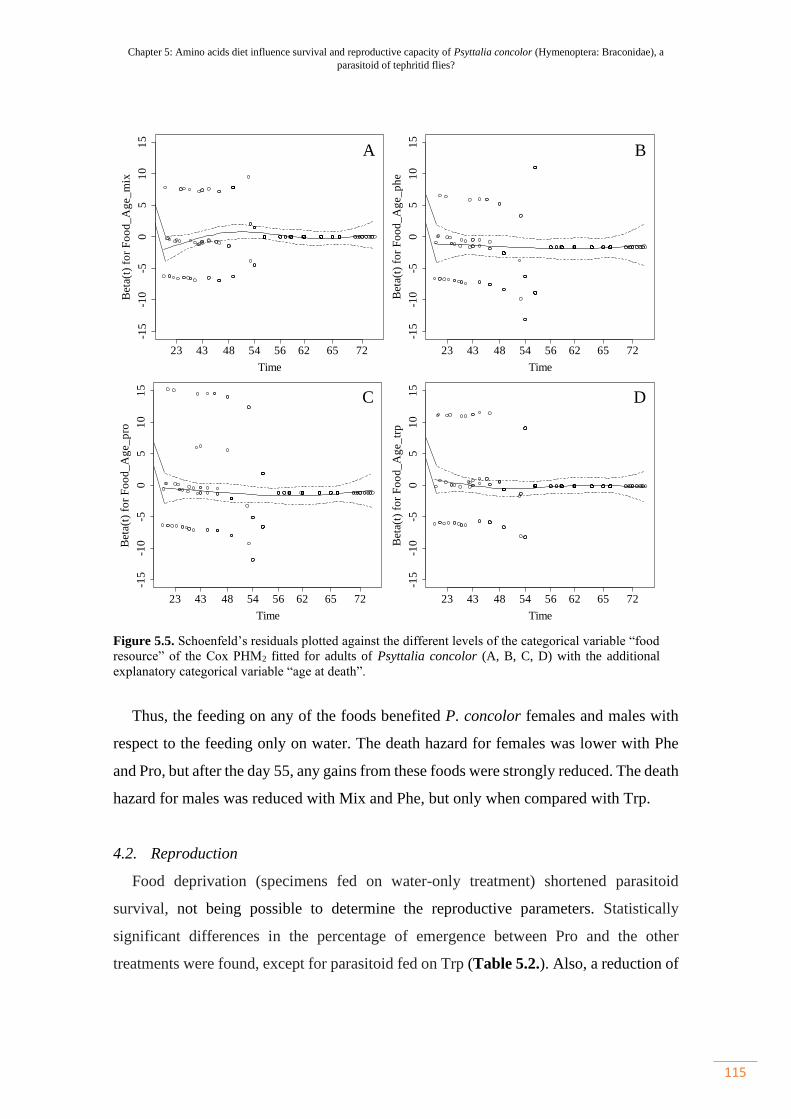
Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
115
Figure 5.5. Schoenfeld’s residuals plotted against the different levels of the categorical variable “food
resource” of the Cox PHM2 fitted for adults of Psyttalia concolor (A, B, C, D) with the additional
explanatory categorical variable “age at death”.
Thus, the feeding on any of the foods benefited P. concolor females and males with
respect to the feeding only on water. The death hazard for females was lower with Phe
and Pro, but after the day 55, any gains from these foods were strongly reduced. The death
hazard for males was reduced with Mix and Phe, but only when compared with Trp.
4.2. Reproduction
Food deprivation (specimens fed on water-only treatment) shortened parasitoid
survival, not being possible to determine the reproductive parameters. Statistically
significant differences in the percentage of emergence between Pro and the other
treatments were found, except for parasitoid fed on Trp (Table 5.2.). Also, a reduction of
Time
Bet
a(t)
fo
r F
oo
d_
Ag
e_tr
p
23 43 48 54 56 62 65 72
-15
-10
-50
51
01
5
Time
Bet
a(t)
fo
r F
oo
d_
Ag
e_p
ro
23 43 48 54 56 62 65 72
-15
-10
-50
51
01
5
Time
Bet
a(t)
fo
r F
oo
d_
Ag
e_p
he
23 43 48 54 56 62 65 72
-15
-10
-50
51
01
5
Time
Bet
a(t)
fo
r F
oo
d_
Ag
e_m
ix
23 43 48 54 56 62 65 72
-15
-10
-50
51
01
5A B
C D

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
116
17% on the emergence was scored for Pro when compared with Glucose (positive
control).
Table 5.2. Percentages of parasitoid emergence, attacked hosts and progeny size in the different
experiments with amino acids (mean ± Standard Error of the mean-SE).
Data followed by the same letter on the same column are not significantly different.
* on the same line, no differences were detected between females and males
The percentage of the attacked host was higher than 75% but the percentage of progeny
size is variable (Table 5.2.). Glucose and the mixture of three amino acids had a
statistically significant effect on the percentage of progeny when compared with the other
amino acids. For females fed on Pro, the percentage of attacked hosts was significantly
reduced when compared with the other food resources (Table 5.2.).
5. DISCUSSION
Various studies have reported the positive effect of different sugars in Hymenoptera
species (Wäckers, 2001; Winkler et al., 2005; Harvey et al., 2012). However, nutrients
such as proteins, amino acids and vitamins may also be important for fitness of parasitoids
(Harvey et al., 2012). Our results present the first report of the effects of amino acid
consumption on survival and reproduction of P. concolor, and this knowledge may help
to explain the need for certain food sources (pollen, nectar or/and honeydew) by
parasitoid and to improve pest control. Amino acids-rich food is extremely important in
biological control programmes ensuring the generation of productive females,
prolongation of the period of oviposition, increasing of the egg loads/maturation ratio and
the proportion of sex ratio (Coskun and Emre, 2015). Our data demonstrated that: (1)
food sources are fundamental to females and males of P. concolor and (2) the risk of
death associated with consumption of diets depends on sex.
Food source
Parasitoid emergence Attacked Hosts
(%) Progeny (%)
Female Male
Glucose 41.26 ± 2.36 a 17.04 ± 2.00 a * 92.13 ± 1.64 a 78.88 ± 1.26 a
Glucose + Phe 37.68 ± 1.97 a 11.00 ± 0.76 a * 90.58 ± 2.56 a 14.96 ± 1.13 b
Glucose + Pro 24.20 ± 3.22 b 15.93 ± 2.83 a 75.92 ± 4.66 b 61.80 ± 3.90 c
Glucose + Trp 32.86 ± 2.37 ab 13.86 ± 1.39 a * 89.69 ± 2.06 a 64.58 ± 2.61 c
Glucose Mix. (Phe+Pro+Trp) 40.69 ± 2.77 a 16.00 ± 2.53 a * 92.84 ± 2.04 a 76.16 ± 2.42 a

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
117
Not surprisingly, parasitoids fed on water had a shortest survival compared with
parasitoids well-fed. Addition of amino acids at a concentration of 0.1 mM to the diet did
not allow the survival to increase significantly. Also, while on males no differences
between amino acids and glucose (positive control) were observed on the survival curves,
Phe showed a significantly lower values when compared with glucose, but not to the other
sugar solutions with amino acids. Although these results indicate that sugars are necessary
for survival of P. concolor, the applied concentration of amino acids is possibly
insufficient to satisfy the nutritional needs of the parasitoids. The fact that no additional
benefit was conferred by the addition of amino acids to parasitoids survival does not
indicate that amino acids are not an important factor for the fitness of P. concolor. In fact,
considering the death hazard ratio (Cox PHM), the risk of dying at any time was reduced
in females and males fed with certain amino acids. Independently of its age, females fed
with Phe and Pro showed a significant reduction of death hazard, while for males the risk
of dying was reduced with the consumption of Mix, followed by Phe. The effect of Phe e
Pro in reducing death hazard may be a result of the strong phagostimulant activity of these
amino acids (Carter et al., 2006; Petanidou et al., 2006). Phenylalanine is an essential
amino acid for insect growth and development (Coskun and Emre, 2015). Moreover,
phenylalanine is one of the most common amino acids found in the floral nectars of
Mediterranean plant species (Petanidou et al., 2006) ensuring an optimal nutrition in the
field. On the other hand, Pro can potentially be used for flight metabolism and diapause
(Li et al., 2014; Micheu, Crailsheim and Leonhard, 2000). According to our results, Trp
did not reduce the risk of death of females and males of P. concolor. This amino acid
represents an important component of the exoskeleton of insects and is particularly
important for cuticle sclerotisation (see Hidalgo et al.,2014). Trp have also an important
role in insect chemosensory receptors (Nicolson and Thornburg, 2007) and was attractive
to, for example green lacewing adults (Chrysoperla carnea Stephens, Hagen et al., 1976).
Indeed, the use of spray solution with L-tryptophan on olive tree canopy increased the
number of C. carnea, a natural enemy of the olive moth pest, Prays oleae (Bernard) (see,
McEwen, Jervis and Kidd, 1994).
Moreover, our reproductive results showed that the addiction of amino acids (Phe, Pro,
Trp and Mix) did not increase the reproductive parameters compared to the control
(glucose). Interestingly, only parasitoid fed with Pro caused a significant reduction of the
percentage of female emergence and attacked hosts compared to glucose. Furthermore,

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
118
values of the percentage of progeny size were much more changeable since, depending
on the experiment, they ranked from the 14% up to the 78%. Thus, compared to glucose,
no statistical differences for progeny size were found only when parasitoid fed with Mix.
The effect of proline on reproductive parameters seem to be different depending not only
the concentration but also the type of diet (ratio of proteins (amino acids): carbohydrates)
and parasitoid species (Hajirajabi et al., 2016). According to Hajirajabi et al. (2016),
proline had positive effects on offspring sex ratio (more females produced), oviposition
period, fecundity, fertility and reproductive index for the parasitoid Trissolcus grandis
Thompson (Hymenoptera: Scelionidae). The Pro ingested by P. concolor may have been
invested into either for fuel and somatic functions (maintenance and locomotion) as
described in the literature (e.g. Arrese and Soulages, 2010). Thus, while Pro is an efficient
fuel for short run, glucose is pointed to extended flight (Carter et al., 2006).
There is also some evidence that phenylalanine is a possible oviposition stimulant for
parasitoids (Barrett and Schmidt, 1991). However, our results demonstrated that the
consumption of Phe does not increment the reproduction of parasitoids. According to
Grenier (2012), high concentrations of phenylalanine can be toxic for parasitoids and,
since in the present study it did not affect survival, we can conclude that probably
secondary metabolites were potentially toxic in the reproduction process. Likewise, Trp
does not supply all nutritional needs to reproduction, as observed on Chrysoperla carnea
(McEwen, Jervis and Kidd, 1994).
In synovigenic parasitoids as P.concolor, parasitoids have a lower number of mature
eggs (the initial egg load), but they can also mature eggs throughout their adult life (Ellers,
Sevenster and Driessen, 2000). Thus, females need to obtain proteins with higher amino
acids demands associated with egg production. However, some parasitoids produced eggs
while feeding on a sugar source (Rivero and Casas, 1999). This might suggest that for P.
concolor, sugars may also be a requirement for reproduction. It is also possible that the
reproductive success of female does not depend exclusively on the quality and availability
of amino acids. For example, for the synovigenic ectoparasitoid, Eupelmus vuilletti
(Crawford) (Hymenoptera, Eupelmidae), protein obtained via intake has no effect on
longevity and fecundity, but proteins obtained by host-feeding increased egg production
and survival (Giron, Pincebourde and Casas, 2004). Moreover, some parasitoids use
lipids and proteins reserves accumulated during the larval stage for eggs production and
maturation (Cicero, Sivinski and Aluja, 2012).

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
119
Overall, the consumption of amino acids affects the survival of P. concolor and
have important implications on host-parasitoid interaction and future investigations are
needed to confirm the importance of supplemental feeding for maximizing longevity and
reproduction of parasitoid in the field.
6. REFERENCES
Alm, J., Ohnmeiss, T.E., Lanza, J. and Vriesenga, L. (1990). Preference of cabbage white
butterflies and honey bees for nectar that contains amino acids. Oecologia, 84(1):
53-57.
Arnó, J., Oveja, M.F. and Gabarra, R. (2018). Selection of flowering plants to enhance
the biological control of Tuta absoluta using parasitoids. Biological Control, 122:
41-50.
Arrese, E.L. and Soulages, J.L. (2010). Insect Fat Body: Energy, Metabolism, and
Regulation. Annual Review of Entomology, 55(1): 207-225.
Baker, H.G. and Baker, I. (1986). The occurrence and significance of amino acids in floral
nectar. Plant Systematics and Evolution, 151(3-4): 175-186.
Baker, H.G., Opler, P.A. and Baker, I. (1978). A comparison of the amino acids
complements of floral and extrafloral nectars. Botanical Gazette, 139(3): 322-332.
Barrett M. and Schmidt, J.M. (1991). A comparison between the amino acid composition
of an egg parasitoid wasp and some of its hosts. Entomologia Experimentalis et
Applicata, 59(1): 29-41.
Benelli, G. and Canale, A. (2012). Learning of visual cues in the fruit fly parasitoid
Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae). Biocontrol, 57(6):767-
777.
Bengochea P., Budia F., Viñuela E. and Medina, P. (2014). Are kaolin and copper
treatments safe to the olive fruit fly parasitoid Psyttalia concolor? Journal of Pest
Science, 87(2): 351-359.
Blüthgen, N. and Fiedler, K. (2004). Preferences for sugars and amino acids and their
conditionality in a diverse nectar-feeding ant community. Journal of Animal
Ecology, 73: 155-166.
Blüthgen, N., Gottsberger, G. and Fiedler, K. (2004). Sugar and amino acid composition
of ant-attended nectar and honeydew sources from an Australian rainforest. Austral
Ecology, 29: 418-429.
Carter, C., Shafir, S., Yehonatan, L., Palmer, R.G. and Thornburg, R. (2006). A novel
role for proline in plant floral nectars. Naturwissenschaften, 93: 72-79.
Cicero, L., Sivinski, J. and Aluja, M. (2012). Effect of host diet and adult parasitoid diet
on egg load dynamics and egg size of braconid parasitoids attacking Anastrepha
ludens. Physiological Entomology, 37: 177-184.
Coskun, M. and Emre, I. (2015). Role of Lipids, Amino Acids, and Sucrose on the Total
Adult and Female Emergence, and Content of Glycogen and Protein in Pimpla
turionellae (Hymenoptera: Ichneumonidae). Annals of the Entomological Society
of America, 108(5): 820-826.
Daane, K.M., and Johnson, M.W. (2010). Olive Fruit Fly: Managing an Ancient Pest in
Modern Times. Annual Review of Entomology, 55(1):151-169.
Daane, K.M., Wang, X., Nieto, D.J., Pickett, C.H., Hoelmer, K.A., Blanchet, A. and
Johnson, M.W. (2015). Classic biological control of olive fruit fly in California,
USA: release and recovery of introduced parasitoids. BioControl, 60(3): 317-330.

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
120
Ellers, J., Sevenster, J.G. and Driessen, G. (2000). Egg Load Evolution in Parasitoids.
The American Naturalist, 156(6): 650-665.
Fischer, M.K., Völkl, W., Schopf R. and Hoffmann, K.H. (2002). Age-specific patterns
in honeydew production and honeydew composition in the aphid Metopeurum
fuscoviride: implications for ant-attendance. Journal of Insect Physiology, 48(3):
319-326.
Gardener, M.C. and Gillman, M.P. (2001a). Analyzing variability in nectar amino acids:
composition is less variable than concentration. Journal of Chemical Ecology,
27(12): 2545-2558.
Gardener, M. C. and Gillman, M.P. (2001b). The effects of soil fertilizer on amino acids
in the floral nectar of corncockle, Agrostemma githago (Caryophyllaceae). Oikos,
92(1): 101-106.
Giron, D. Pincebourde, S. and Casas, J. (2004). Lifetime gains of host-feeding in a
synovigenic parasitic wasp. Physiological Entomology, 29(5): 436-442.
Grenier, S. (2012). Artificial Rearing of Entomophagous Insects, with Emphasis on
Nutrition and Parasitoids - General Outlines from Personal Experience. Karaelmas
Science and Engineering Journal, 2 (2): 1-12.
Hagen, K.S., Greany, P., Sawall Jr., E.F. and Tassain, R.L. (1976). Tryptophan in
artificial honeydews as a source of an attractant for adult Chrysopa carnea.
Environmental Entomology 5(3): 458-468.
Hajirajabi, N., Fazel, M.M., Harvey, J.A., Arbab, A. and Asgari, S. (2016). Dietary sugars
and proline influence biological parameters of adult Trissolcus grandis, an egg
parasitoid of Sunn pest, Eurygaster integriceps. Biological Control, 96: 21-27.
Harvey, J.A., Cloutier, J., Visser, B., Ellers, J., Wäckers, F.L., and Gols, R. (2012). The
effect of different dietary sugars and honey on longevity and fecundity in two
hyperparasitoid wasps. Journal of Insect Physiology, 58(6): 816-823.
Hidalgo, K., Mouline, K., Mamai, W., Foucreau, N., Dabire, K.R., Bouchereau, A.,
Simard, F. and Renault, D. (2014). Novel insights into the metabolic and
biochemical underpinnings assisting dry-season survival in female malaria
mosquitoes of the Anopheles gambiae complex. Journal of Insect Physiology 70:
102-116.
Human, H. and Nicolson, S.W. (2006). Nutritional content of fresh, bee-collected and
stored pollen of Aloe greatheadii var. davyana (Asphodelaceae). Phytochemistry,
67(14): 1486-1492.
Jacas, J. A. and Viñuela, E. (1994). Analysis of a lab method to test the effects of
pesticides on adult females of Opius concolor, a parasitoid of the olive fruit fly
Bactrocera oleae. Biocontrol Science and Technology, 4(2): 147-154.
Komosinska-Vassev, K., Olczyk, P. Kaźmierczak, J., Mencner, L. and Olczyk1, K.
(2015). Bee Pollen: Chemical Composition and Therapeutic Application. Evidence-
Based Complementary and Alternative Medicine, 15: 1-6.
Leroy, P. D., Wathelet, B., Sabri, A., Francis, F., Verheggen, F. J., Capella. Q., Thonart,
P. and Haubruge, E. (2011). Aphid-host plant interactions: does aphid honeydew
exactly reflect the host plant amino acid composition? Arthropod-Plant Interactions,
5(3): 193-199.
Li, Y., Zhang, L., Zhang, Q., Chen, H. and Denlinger, D.L. (2014). Host diapause status
and host diets augmented with cryoprotectants enhance cold hardiness in the
parasitoid Nasonia vitripennis. Journal of Insect Physiology, 70:8-14.
Lundgren, J.G. and Wiedenmann R.N. (2004). Nutritional suitability of corn pollen for
the predator Coleomegilla maculata (Coleoptera: Coccinellidae). Journal of Insect
Physiology, 50(6): 567-575.

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
121
Malheiro, R., Casal, S., Baptista, P. and Pereira, J.A. (2015). A review of Bactrocera
oleae (Rossi) impact in olive products: From the tree to the table. Trends in Food
Science & Technology, 44(2): 226-242.
Micheu, S., Crailsheim, K., and Leonhard, B. (2000). Importance of proline and other
amino acids during honeybee flight. Amino Acids, 18(2), 157-175.
Miranda, M.A., Miquel, M., Terrassa, J. Melis N. and Monerris M. (2008). Parasitism of
Bactrocera oleae (Diptera; Tephritidae) by Psyttalia concolor (Hymenoptera;
Braconidae) in the Balearic Islands (Spain). Journal of Applied Entomology, 132(9-
10): 798-805.
McEwen, P., Jervis, M.A. and Kidd, N.A.C. (1994). Use of sprayed L-tryptophan solution
to concentrate numbers of the green lacewing Chrysoperla carnea in olive tree
canopy. Entomologia Experimentalis et Applicata, 70(1):97-99.
Mevi-Schutz J. and Erhardt, A. (2003). Effects of nectar amino acids on fecundity of the
wall brown butterfly (Lasiommata megera L.). Basic and Applied Ecology, 4(5):
413-421.
Mondy, N. Corio-Costet, M.-F., Bodin, A., Mandon, N., Vannier, F. and Monge, J.-P.
(2006). Importance of sterols acquired through host feeding in synovigenic
parasitoid oogenesis. Journal of Insect Physiology, 52(9): 897-904.
Nicolson S.W and Thornburg R.W. (2007). Nectar Chemistry. In., Nicolson, M. Nepi &
E. Pacini, (eds.), Nectaries and Nectar. Springer, Dordrecht. pp. 215-263.
Petanidou, T., Goethals, V. and Smets, E. (2000). Nectary structure of Labiatae in relation
to their nectar secretion and characteristics in a Mediterranean shrub community -
Does flowering time matter?. Plant Systematics and Evolution, 225(1-4):103-118.
Petanidou, T., Van Laere, A.J., and Smets, E. (1996). Change in floral nectar components
from fresh to senescent flowers of Capparis spinosa L. (Capparidaceae), a
nocturnally flowering Mediterranean shrub. Plant Systematics and Evolution,
199(1-2): 79-92.
Petanidou, T., Van Laere, A.J., Ellis, W.N. and Smets, E. (2006). What shapes amino acid
and sugar composition in Mediterranean floral nectars? Oikos, 115(1): 155-169.
R Core Team (2014). R: A language and environment for statistical computing. R
Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-
project.org/.[accesed: 11/08/2016].
Raspi, A. and Loni, A. (1994). Alcune note sull’allevamento massale di Opius concolor
Szepligeti (Hym.: Braconidae) e su recnti tentative d’introduzione della specie in
Toscana e Liguria. Frustula Entomologia, 30:135-145.
Rivero, A. and Casas, J. (1999) Incorporating physiology into parasitoid behavioral
ecology: the allocation of nutritional resource. Research in Population Ecology,
41(1): 39-45.
Roulston, T.H. and Cane. J.H. (2000). Pollen nutritional content and digestibility for
animals. Plant Systematics Evolution, 222:187-209.
Sandström, J.P. and Moran N. (2001). Amino acid budgets in three aphid species using
the same host plant. Physiological Entomology, 26: 202-211.
Szczesna, T. (2006). Protein content and amino acid composition of bee-collected pollen
from selected botanical origins. Journal of Apicultural Science, 50: 81-90.
Therneau T (2014). _A Package for Survival Analysis in S_. R package version 2.37-7,
URL: http://CRAN.R-project.org/package=survival.[accesed: 11/08/2016].
Therneau T.M., Grambsch P.M. (2000) Modeling Survival Data: Extending the Cox
Model. Statistics for biology and health. Springer. USA.

Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
122
Wäckers, F.L. (2001). A comparison of nectar- and honeydew sugars with respect to their
utilization by the hymenopteran parasitoid Cotesia glomerata. Journal of Insect
Physiology, 47(9):1077-1084.
Wada, A., Isobe, Y., Yamaguchi, S., Yamaoka, R. and Ozaki, M. (2001). Taste-enhancing
effects of glycine on the sweetness of glucose: a gustatory aspect of symbiosis
between the ant Camponotus japonicus and the larvae of the lycaenid butterfly
Niphanda fusca. Chemical Senses, 26: 983-992.
Winkler, K., Wackers, F.L., Stingli, A. and van Lenteren, J.C. (2005). Plutella xylostella
(diamondback moth) and its parasitoid Diadegma semiclausum show different
gustatory and longevity responses to a range of nectar and honeydew sugars.
Entomologia Experimentalis et Applicata, 115(1):187-192.
Yokoyama, V.Y., Rendón, P.A. and Sivinski, J. (2008a). Psyttalia cf.
concolor (Hymenoptera: Braconidae) for Biological Control of Olive Fruit Fly
(Diptera: Tephritidae) in California. Environmental Entomology, 37(3): 764-773.
Yokoyama, V.Y., Rendón, P.A. and Sivinski, J. (2008b). Biological control of olive fruit
fly (Diptera: Tephritidae) by releases of Psyttalia cf. concolor (Hymenoptera:
Braconidae) in California, parasitoid longevity in presence of the host, and host
status of walnut husk fly. In: Proceedings of the 7th international symposium on
fruit flies of economic importance, 10-15.
Zhang, G., Zimmermann, O. and Hassan, S.A. (2004). Pollen as a source of food for egg
parasitoids of the genus Trichogramma (Hymenoptera: Trichogrammatidae).
Biocontrol Science and Technology, 14(2):201-209.
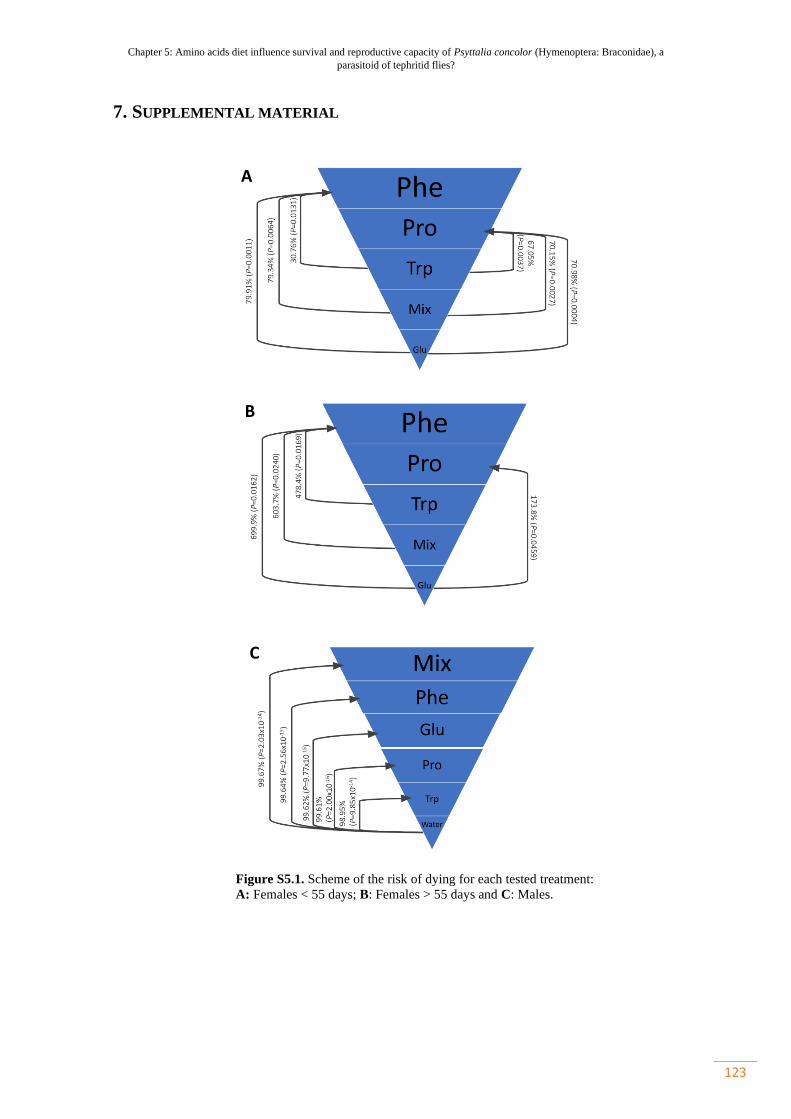
Chapter 5: Amino acids diet influence survival and reproductive capacity of Psyttalia concolor (Hymenoptera: Braconidae), a
parasitoid of tephritid flies?
123
7. SUPPLEMENTAL MATERIAL
Figure S5.1. Scheme of the risk of dying for each tested treatment:
A: Females < 55 days; B: Females > 55 days and C: Males.
A
B
C

124
Chapter 6
Side effects of pesticides
on the parasitoid Psyttalia
concolor (Szépligeti): a
review

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
125
Chapter 6:
SIDE EFFECTS OF PESTICIDES ON THE PARASITOID
PSYTTALIA CONCOLOR (SZÉPLIGETI): A REVIEW
1. ABSTRACT
Pesticides application for pest management in olive orchards might change parasitoids
Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) efficacy in biological control
of Bactrocera oleae (Rossi) (Diptera: Tephritidae). When pesticides are sprayed in olive
groves, P. concolor can be mainly contaminated by direct exposure to spray droplets, by
contact with treated surfaces and by oral uptake from contaminated food sources (e.g.
feeding on contaminated water, pollen, nectar or honeydew available on plant parts). The
application of a pesticide not only reduces pest but also the parasitoid populations,
because they need certain population of the pest to survive and depends on its ability to
recover from pesticide toxic effects. Identifying selective pesticides with low effects on
P. concolor and an appropriate timing of application/parasitoid releases provide an
opportunity to use the parasitoid in IPM, improving the control of B. oleae as well as
reducing the environmental impact of repetitive application of pesticides.
This manuscript attempts to review the lethal and sublethal effects reported in literature
on the parasitoid which have revealed, as expected, that insecticides are generally more
toxic to parasitoid, above all organophosphates and pyrethroids, while fungicides,
herbicides and biopesticides have, in general, less effects on mortality and reproductive
parameters of P. concolor. However, some fungicides proved to be quite harmful for
parasitoid. Most of the studies were conducted in laboratory and focused on mortality
(lethal effects) and the reproduction as unique sublethal effect evaluated, only consider
the effect of a single pesticide and persistence was hardly explored. Field studies are quite
scarce and absolutely needed to satisfactorily assessing the impacts of pesticides when
combined with biological control.
Keywords: Bactrocera oleae, biopesticides, fungicides, herbicides, insecticides, pest
control

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
126
2. INTRODUCTION
Agricultural production must be increased considerably in the foreseeable future to
meet the food demands of a rising human population, being at risk due to the incidence
of pests and plant pathogens. These organisms, that can cause substantial losses
(quantitative and/or qualitative), can be reduced or eliminated using several methods,
namely biological, chemical physical and cultural control methods (Oerke, 2006).
However, modern agriculture has extensively relied on pesticides for pest control (Casida
and Quistad, 1998; Damalas, 2009).
For growers, the application of pesticides takes several advantages, concerning its
efficacy, simplicity and economical costs (minimizing time and money and reducing
other costly inputs as labour or fuel). Adverse effects of pesticides can include the killing
of non-target organisms, contamination of water supplies and persistence of unwanted
residues of foods and animal feed (Weinzierl, 2008). Moreover, resistance to one or more
pesticides, secondary pest outbreaks and/or target pest resurgence have been also a focus
of several studies (Denholm, Pickett and Devonshire, 2001; Cloyd, 2012).
In recent years, the increasing number of agricultural producers consciously alerted
for a sustainable use of pesticides have contributed to a growing implementation of
Integrated Pest Management (IPM) programs (Kenis et al., 2017; Stenberg, 2017),
becoming mandatory in all European Union (EU) countries (OJEU 2009) since the EU
directive 2009/128/EC, on sustainable use of pesticides, came into force (Fernandez et
al., 2017). According to this directive, it is essential to understand the risks of pesticides
and adopt pest control strategies that include more effective and selective substances
(Cooper and Dobson, 2007; Damalas, 2009). Moreover, it encourages the use of more
environmentally friendly alternatives to pesticides such as biological control methods. In
this context, an essential component of IPM programs is the use of “macrobial” (predators
or parasitoids) or “microbial” (bacteria, viruses, fungi and nematodes) agents targeted
against insect and mite pests. Although the biological control approaches minimize the
disruption to agroecosystems and effectively combat pests, they are rarely so robust as to
provide all the pest suppression needed and therefore, the best strategy is the optimal use
of natural enemies, supplemented with selective insecticides when necessary, which
integrates biological and chemical control (Medina et al., 2007; Gentz, Murdoch and
King, 2010). Therefore, the first step for developing a successful IPM implementation is

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
127
an accurate evaluation of the potential side effects of pesticides on biological control
agents (Garzón et al., 2015).
2.1. Effects of pesticides on natural enemies: methods for testing compatibility
The side effects of pesticides on biological control agents has been extensively studied
in the last thirty years. For a long time, the classical laboratory method was to determine
a median lethal dose or concentration (LD50/LC50) of both pest and natural enemies
(Desneux, Decourtye and Delpuech, 2007). When this ratio is ˃1, the pesticide is more
toxic to pest insect than natural enemies. In Europe, the Working Group “Pesticides and
Beneficial Organisms” of the International Organization for the Biological Control/West
Palearctic Regional Section (IOBC/WPRS) was established in 1974 with the major aim
at encouraging “the development of standard methods for testing the side-effects of
pesticides on natural enemies” with the goal of harmonization in order to compare the
effects obtained by different tests (in similar environmental conditions) for the same or
different natural enemies, in different countries. A further aim of the group was therefore
to test the side-effects of commonly used pesticides on the most important natural enemies
and to provide an “information service” to the other IOBC groups (Hassan, 1998).
All IOBC test methods are based on a sequential scheme of three steps that include
laboratory, semi-field and field studies (Fig. 6.1.). This sequence assumes that pesticides
that are harmless in the laboratory will also be safe in semi-field and field conditions, and
do not need to be evaluated in further steps. When a chemical is, however, categorized as
harmful in one step, its effect in the next step cannot be inferred, and the sequence must
be followed until it finishes at field conditions or displays no negative effects. Pesticides
are tested with a residual test at the maximum recommended field rate, based on
commercial formulations, starting by a laboratory test. Overall, laboratory methods
evaluate the level of mortality (lethal effect) as well as sublethal effects of the pesticides
on the reproductive parameters.
The semi-field tests are performed in more natural condition, as direct application of
pesticide on plants and field studies are carried out in semi-natural field conditions (see
Hassan, 1998 for details; Amano and Haseeb, 2001; Macfadyen et al., 2014). In practice,
pesticides are finally ranked, according to their effects on natural enemies, for (a)
laboratory studies in 1 (harmless, ˂30% mortality to non-target organisms), 2 (slightly
harmful, 30-79%), 3 (moderately harmful, 80-98%) and 4 (harmful, >99%); and (b) semi-
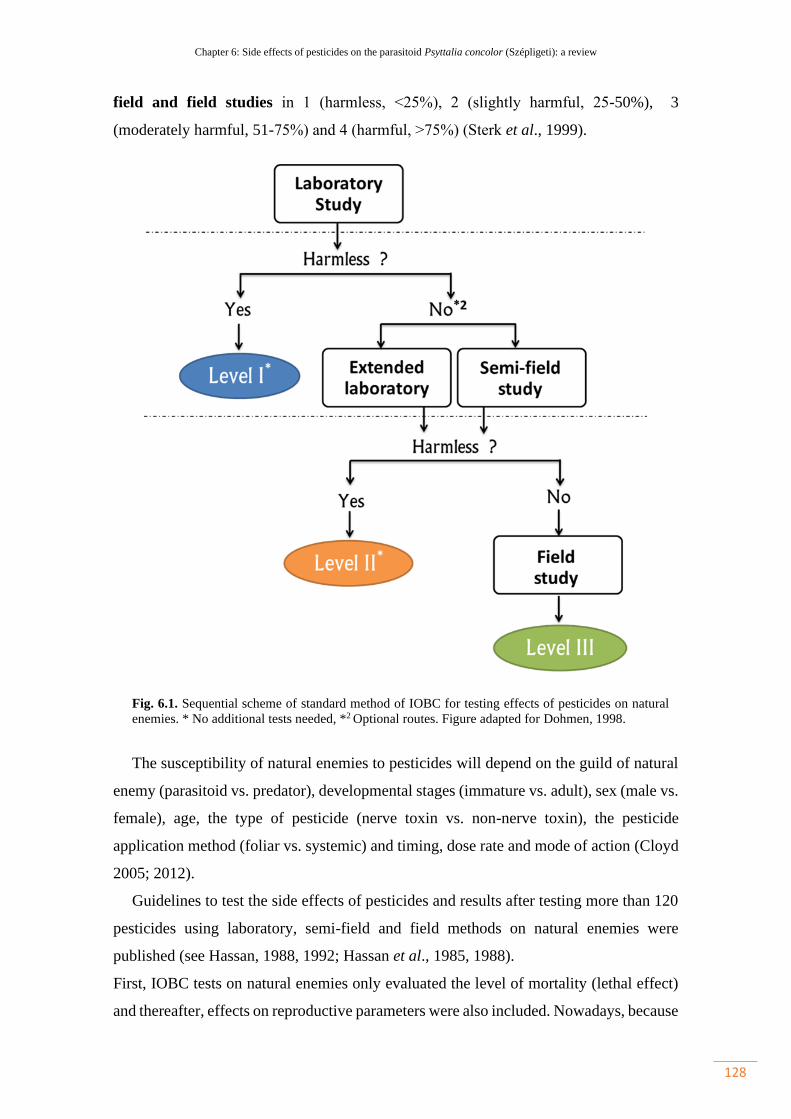
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
128
field and field studies in 1 (harmless, ˂25%), 2 (slightly harmful, 25-50%), 3
(moderately harmful, 51-75%) and 4 (harmful, ˃75%) (Sterk et al., 1999).
Fig. 6.1. Sequential scheme of standard method of IOBC for testing effects of pesticides on natural
enemies. * No additional tests needed, *2 Optional routes. Figure adapted for Dohmen, 1998.
The susceptibility of natural enemies to pesticides will depend on the guild of natural
enemy (parasitoid vs. predator), developmental stages (immature vs. adult), sex (male vs.
female), age, the type of pesticide (nerve toxin vs. non-nerve toxin), the pesticide
application method (foliar vs. systemic) and timing, dose rate and mode of action (Cloyd
2005; 2012).
Guidelines to test the side effects of pesticides and results after testing more than 120
pesticides using laboratory, semi-field and field methods on natural enemies were
published (see Hassan, 1988, 1992; Hassan et al., 1985, 1988).
First, IOBC tests on natural enemies only evaluated the level of mortality (lethal effect)
and thereafter, effects on reproductive parameters were also included. Nowadays, because

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
129
of greater relevance given to beneficial arthropods, the increasing important of beneficial
arthropods, sublethal effects have been more extensively studied. Although sublethal
doses/concentrations do not kill directly the exposed natural enemies, there may be a
reduction of their populations and or an interference on physiological and behavioural
traits which can significantly affect their fitness and then compromise the success of
biological control programs (Desneux, Decourtye and Delpuech, 2007; Geiger et al.,
2010).
2.2. Impact of pesticides on Bactrocera oleae-Psyttalia concolor interaction
The olive fruit fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae), is a monophagous
frugivorous pest which females cause damages through the oviposition of their eggs in
the olive fruits (Olea europaea L. cultivated and wild) and is considered to be the key
pest of the Mediterranean Basin olive tree orchards. Females lay eggs in olive fruits and
larvae feed the pulp and finally pupate inside the olive or exit to pupate on the ground
(Daane and Johnson, 2010). However, not all olive tree cultivars are equally susceptible
to olive fruit fly infestation, being the severity of the damages influenced by olive
characteristics (e.g., fruit size, colour, fruit exocarp hardness or/and chemical
composition of olive fruits), geographical area and weather conditions (Gonçalves et al.,
2012; Malheiro et al., 2015). This pest causes a reduction in yield owing to a premature
fruit drop or a loss of weight of the fruit caused by the feeding larvae. Moreover,
microorganism growth inside the fruit increases the acidity of olive oils, reducing their
price (Malheiro et al., 2015).
Olive fruit fly control methods has been based on the use of (a) insecticides: bait
applications (insecticides mixed with an attractant) and cover sprays, which are applied
in some cases, (b) traps and (c) Sterile Insect Technique (SIT) (Kampouraki et al., 2018).
Although spray applications have been efficient and economical methods used against
the olive fruit fly, this pest has evolved resistance to some of the most commonly used
insecticides (Kampouraki, et al., 2018; Vontas et al., 2011).
Apart from chemical control, biological control of B. oleae with natural enemies can
be an alternative method not involving insecticides (Hladnik, 2017; Saour and Makee,
2004). In Mediterranean area, biological control has mainly focused on chalcididae
ectoparasitoids, Eupelmus urozonus Dalman (Eupelmidae), Pnigalio mediterraneus
Ferriere and Delucchi (Eulophidae), Eurytoma martellii Domenichini (Eurytomidae),
Cyrtoptyx latipes (Rondani) (Pteromalidae), and the braconid endoparasitoid Psyttalia

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
130
concolor (Szépligeti) cited by Túnez, in 1910, for the first time. In this work, we focus
on P. concolor, a synovigenic koinobiont larval-pupal endoparasitoid of many
Tephritidae (Diptera), released in several Mediterranean and Californian olive groves for
the biological control of B. oleae with varying levels of success (Miranda et al., 2008;
Yokoyama, Rendón and Sivinski, 2008; Hepdurgun, Turanli and Zümreoğlu, 2009;
Daane and Johnson, 2010). Indeed, the rate of success of parasitoid introduction depends
of several factors including the field-establishment of ecological infrastructures in olive
groves. The presence of some landscape elements such as flowering strips, banker plants
and hedgerows provide (1) food (pollen, nectar and hemipteran honeydew) (2) alternative
hosts and (3) refuges (e.g., from tillage or pesticides), important for the persistence of
natural enemies in agroecosystems and enhance the efficacy of biological control (Fiedler
Landis and Wratten, 2008; Gillespie, Gurr and Wratten, 2016). However, these landscape
structures should also be free of pesticides because of the possible risk of contamination
by parasitoids, both by contact and oral-intoxication. Some studies showed that residues
of pesticides persist in plant tissues long enough to contaminate the pollen and nectar
(Botías et al., 2016; Cowles and Eitzer, 2017), essential source of energy (carbohydrates)
and protein for survival, host foraging and reproduction of parasitoids (Benelli et al.,
2017) (Fig. 6.2.). Moreover, parasitoids can also be exposed to pesticides per contact with
leaves during their search for hosts, feeding, mating and resting (Stapel, Cortesero and
Lewis 2000; Krischik, Landmark and Heimpel, 2007), not only in the landscape but most
often in the crop. Furthermore, parasitoids may feed on liquid exuded from host for food
(host feeding) (Jervis and Kidd, 1996). This type of food is particularly important for
synovigenic females, such as Psyttalia concolor, which often needs to feed for eggs
maturation (Genduso, 1970), thus increasing the risk of exposure to insecticides and
fungicides. On the other hand, pesticide application induces changes in the chemical
constituents of the flowering plants and/or host plants which may decrease the nutritional
value and/or became less attractive, e.g., pollen viability, changes in phagostimulating
metabolites (sucrose, serine, alanine) and in the essential branched-chain amino acids
(Hervé et al., 2014; Qian et al., 2015a,b; Serra et al., 2015a,b). Moreover, it assumes that
semiochemical diversity is enhanced, and that also this interferes with olfactory
orientation of insects in oviposition behaviours (reduced capacity to find the host and the
responses to host kairomones) change foraging patterns and sex pheromonal
communication (Longley and Jepson, 1996; Stapel, Cortesero and Lewis, 2000; Desneux,
Pham-Delègue and Kaiser, 2004; Tappert et al., 2017).

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
131
Fig. 6.2. Biological relationship between olive fruit fly and beneficial insects and pesticides application.
P. concolor can be contaminated by (1) contact after direct exposure to spray droplets
or residues and (2) oral uptake from contaminated food sources (e.g. feeding on
contaminated water, pollen, nectar or honeydew available on plant parts). In such
scenario, the application of a pesticide not only reduces the pest but also the parasitoid
population, because (1) the parasitoid population needs certain levels of the population of
the pest to survive, and (2) the parasitoid depends on its ability to recover from pesticide
toxic effects. Many studies, most of them reviewed herein with special emphasis on
insecticides applications, have demonstrated some side effects on P. concolor, whereas
fungicides treatments were less studied. Studies with herbicides were traditionally
neglected as direct contact after a treatment, especially in the case of pre-emergence
herbicides, due to its uncommon presence in traditional exposure methodology proposed
by IOBC. Nevertheless, from a broader perspective, all herbicides can indirectly affect P.
concolor population as they limit the growth of plants providing food, and post-
emergence herbicides, in addition, potentially might reduce the survival and reproduction
of P. concolor adults.
This work aimed at reviewing the lethal and some sublethal effects (those related to
reproduction) of pesticides already registered or that potentially might be applied on olive
groves in Europe, discussing the implications of those effects when both, parasitoids and
pesticides must coexist within a pest management program. The first part focused in a

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
132
revision of the available literature about side effects of pesticides currently used in Europe
against olive pests on this parasitoid. The second is dedicated to the effects of different
pesticides on P. concolor females mortality and reproduction (laboratory case study),
with two routes investigated: contact and oral uptake (representing a realistic scenario in
olive groves). In the third part an integrated discussion of all results obtained in the review
and laboratory case study is given. Finally, the conclusions assembled from all the results
obtained.
3. IMPACT OF PESTICIDES ON PARASITOID PSYTTALIA CONCOLOR: A
REVIEW
The impact caused by pesticides is important for the development of future
management strategies that conserve and/or increase parasitoid population levels in
olive groves. Having this, a compilation of pesticides already tested against Psyttalia
concolor is presented as an overview in Table 6.1. (insecticides), Table 6.2. (fungicides
and herbicides) and Table 6.3. (biopesticides). Only pesticides currently registered in the
Europe Union were considered in Tables 6.1. and 6.2.
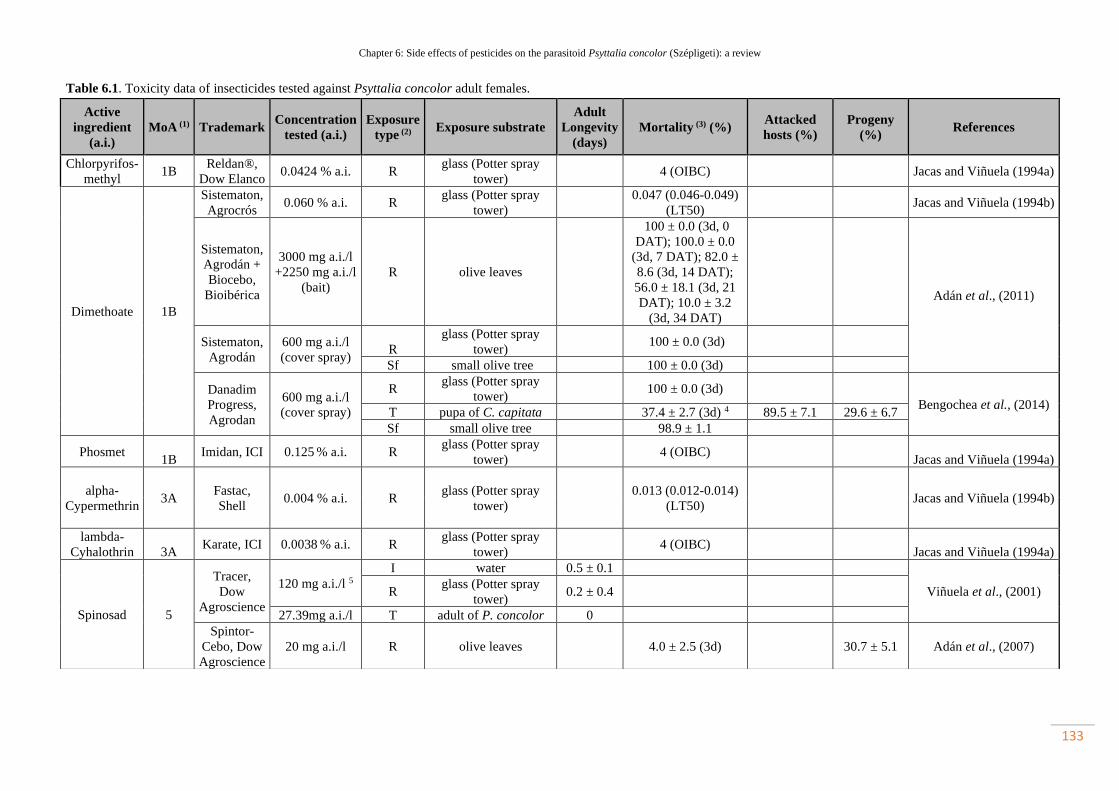
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
133
Table 6.1. Toxicity data of insecticides tested against Psyttalia concolor adult females.
Active
ingredient
(a.i.)
MoA (1) Trademark Concentration
tested (a.i.)
Exposure
type (2) Exposure substrate
Adult
Longevity
(days)
Mortality (3) (%) Attacked
hosts (%)
Progeny
(%) References
Chlorpyrifos-
methyl 1B
Reldan®,
Dow Elanco 0.0424 % a.i. R
glass (Potter spray
tower) 4 (OIBC) Jacas and Viñuela (1994a)
Dimethoate 1B
Sistematon,
Agrocrós 0.060 % a.i. R
glass (Potter spray
tower)
0.047 (0.046-0.049)
(LT50) Jacas and Viñuela (1994b)
Sistematon,
Agrodán +
Biocebo,
Bioibérica
3000 mg a.i./l
+2250 mg a.i./l
(bait)
R olive leaves
100 ± 0.0 (3d, 0
DAT); 100.0 ± 0.0
(3d, 7 DAT); 82.0 ±
8.6 (3d, 14 DAT);
56.0 ± 18.1 (3d, 21
DAT); 10.0 ± 3.2
(3d, 34 DAT)
Adán et al., (2011)
Sistematon,
Agrodán
600 mg a.i./l
(cover spray) R
glass (Potter spray
tower) 100 ± 0.0 (3d)
Sf small olive tree 100 ± 0.0 (3d)
Danadim
Progress,
Agrodan
600 mg a.i./l
(cover spray)
R glass (Potter spray
tower) 100 ± 0.0 (3d)
Bengochea et al., (2014)
T pupa of C. capitata 37.4 ± 2.7 (3d) 4 89.5 ± 7.1 29.6 ± 6.7
Sf small olive tree 98.9 ± 1.1
Phosmet 1B
Imidan, ICI 0.125 % a.i. R glass (Potter spray
tower) 4 (OIBC)
Jacas and Viñuela (1994a)
alpha-
Cypermethrin 3A
Fastac,
Shell 0.004 % a.i. R
glass (Potter spray
tower)
0.013 (0.012-0.014)
(LT50) Jacas and Viñuela (1994b)
lambda-
Cyhalothrin 3A Karate, ICI 0.0038 % a.i. R
glass (Potter spray
tower) 4 (OIBC)
Jacas and Viñuela (1994a)
Spinosad 5
Tracer,
Dow
Agroscience
120 mg a.i./l 5
I water 0.5 ± 0.1
Viñuela et al., (2001) R glass (Potter spray
tower) 0.2 ± 0.4
27.39mg a.i./l T adult of P. concolor 0
Spintor-
Cebo, Dow
Agroscience
20 mg a.i./l R olive leaves
4.0 ± 2.5 (3d) 30.7 ± 5.1 Adán et al., (2007)
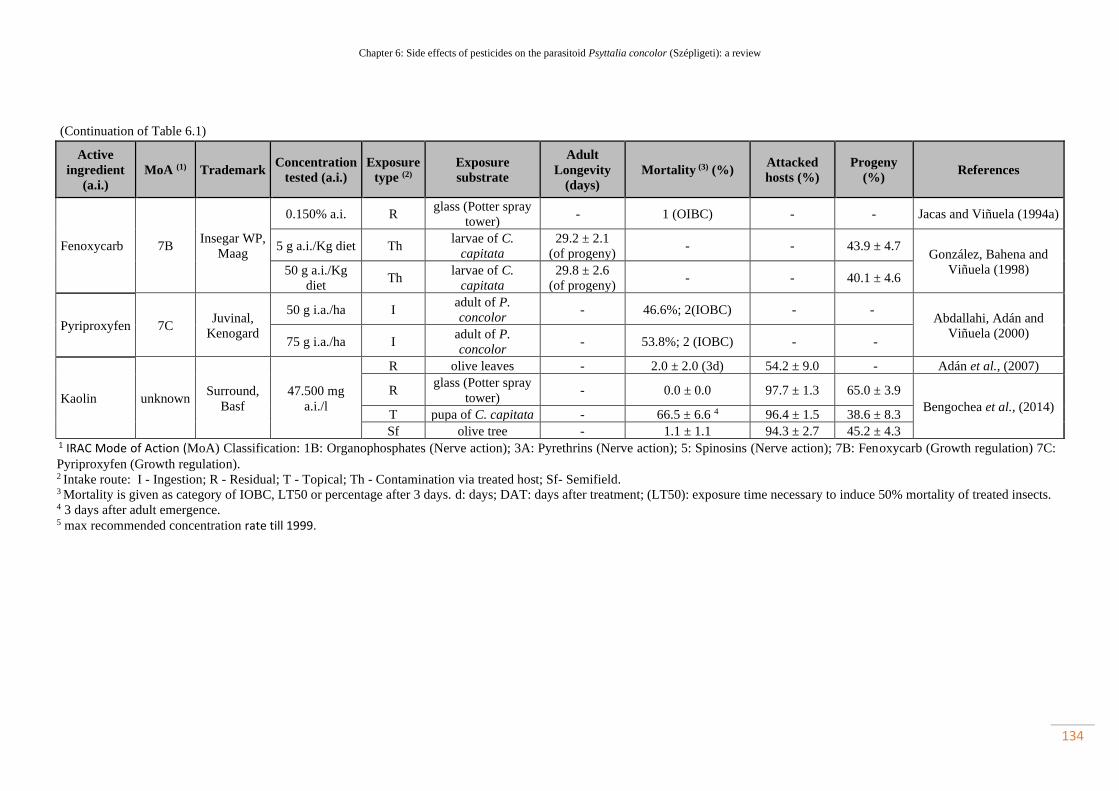
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
134
(Continuation of Table 6.1)
1 IRAC Mode of Action (MoA) Classification: 1B: Organophosphates (Nerve action); 3A: Pyrethrins (Nerve action); 5: Spinosins (Nerve action); 7B: Fenoxycarb (Growth regulation) 7C:
Pyriproxyfen (Growth regulation). 2 Intake route: I - Ingestion; R - Residual; T - Topical; Th - Contamination via treated host; Sf- Semifield. 3 Mortality is given as category of IOBC, LT50 or percentage after 3 days. d: days; DAT: days after treatment; (LT50): exposure time necessary to induce 50% mortality of treated insects. 4 3 days after adult emergence. 5 max recommended concentration rate till 1999.
Active
ingredient
(a.i.)
MoA (1) Trademark Concentration
tested (a.i.)
Exposure
type (2)
Exposure
substrate
Adult
Longevity
(days)
Mortality (3) (%) Attacked
hosts (%)
Progeny
(%) References
Fenoxycarb 7B Insegar WP,
Maag
0.150% a.i. R glass (Potter spray
tower) - 1 (OIBC) - - Jacas and Viñuela (1994a)
5 g a.i./Kg diet Th larvae of C.
capitata
29.2 ± 2.1
(of progeny) - - 43.9 ± 4.7
González, Bahena and
Viñuela (1998) 50 g a.i./Kg
diet Th
larvae of C.
capitata
29.8 ± 2.6
(of progeny) - - 40.1 ± 4.6
Pyriproxyfen 7C Juvinal,
Kenogard
50 g i.a./ha I adult of P.
concolor - 46.6%; 2(IOBC) - -
Abdallahi, Adán and
Viñuela (2000) 75 g i.a./ha I
adult of P.
concolor - 53.8%; 2 (IOBC) - -
Kaolin unknown Surround,
Basf
47.500 mg
a.i./l
R olive leaves - 2.0 ± 2.0 (3d) 54.2 ± 9.0 - Adán et al., (2007)
R glass (Potter spray
tower) - 0.0 ± 0.0 97.7 ± 1.3 65.0 ± 3.9
Bengochea et al., (2014) T pupa of C. capitata - 66.5 ± 6.6 4 96.4 ± 1.5 38.6 ± 8.3
Sf olive tree - 1.1 ± 1.1 94.3 ± 2.7 45.2 ± 4.3
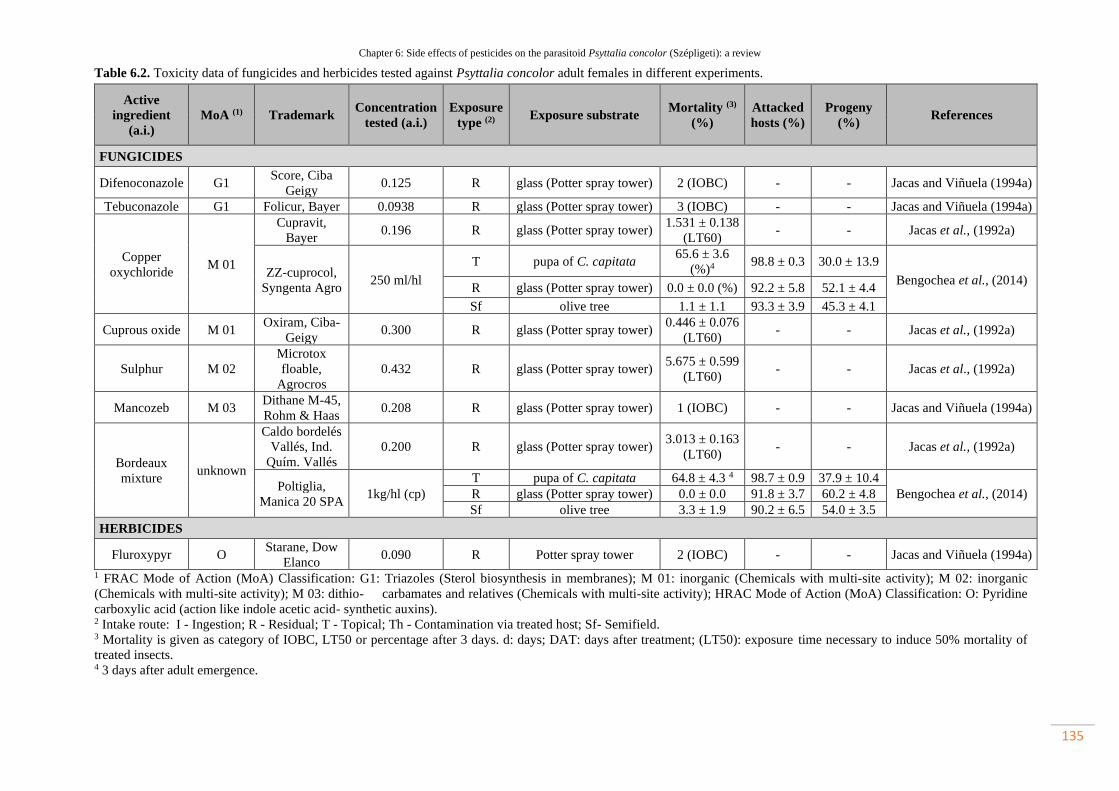
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
135
Table 6.2. Toxicity data of fungicides and herbicides tested against Psyttalia concolor adult females in different experiments.
1 FRAC Mode of Action (MoA) Classification: G1: Triazoles (Sterol biosynthesis in membranes); M 01: inorganic (Chemicals with multi-site activity); M 02: inorganic
(Chemicals with multi-site activity); M 03: dithio- carbamates and relatives (Chemicals with multi-site activity); HRAC Mode of Action (MoA) Classification: O: Pyridine
carboxylic acid (action like indole acetic acid- synthetic auxins). 2 Intake route: I - Ingestion; R - Residual; T - Topical; Th - Contamination via treated host; Sf- Semifield. 3 Mortality is given as category of IOBC, LT50 or percentage after 3 days. d: days; DAT: days after treatment; (LT50): exposure time necessary to induce 50% mortality of
treated insects. 4 3 days after adult emergence.
Active
ingredient
(a.i.)
MoA (1) Trademark Concentration
tested (a.i.)
Exposure
type (2) Exposure substrate
Mortality (3)
(%)
Attacked
hosts (%)
Progeny
(%) References
FUNGICIDES
Difenoconazole G1 Score, Ciba
Geigy 0.125 R glass (Potter spray tower) 2 (IOBC) - - Jacas and Viñuela (1994a)
Tebuconazole G1 Folicur, Bayer 0.0938 R glass (Potter spray tower) 3 (IOBC) - - Jacas and Viñuela (1994a)
Copper
oxychloride M 01
Cupravit,
Bayer 0.196 R glass (Potter spray tower)
1.531 ± 0.138
(LT60) - - Jacas et al., (1992a)
ZZ-cuprocol,
Syngenta Agro 250 ml/hl
T pupa of C. capitata 65.6 ± 3.6
(%)4 98.8 ± 0.3 30.0 ± 13.9
Bengochea et al., (2014) R glass (Potter spray tower) 0.0 ± 0.0 (%) 92.2 ± 5.8 52.1 ± 4.4
Sf olive tree 1.1 ± 1.1 93.3 ± 3.9 45.3 ± 4.1
Cuprous oxide M 01 Oxiram, Ciba-
Geigy 0.300 R glass (Potter spray tower)
0.446 ± 0.076
(LT60) - - Jacas et al., (1992a)
Sulphur M 02
Microtox
floable,
Agrocros
0.432 R glass (Potter spray tower) 5.675 ± 0.599
(LT60) - - Jacas et al., (1992a)
Mancozeb M 03 Dithane M-45,
Rohm & Haas 0.208 R glass (Potter spray tower) 1 (IOBC) - - Jacas and Viñuela (1994a)
Bordeaux
mixture unknown
Caldo bordelés
Vallés, Ind.
Quím. Vallés
0.200 R glass (Potter spray tower) 3.013 ± 0.163
(LT60) - - Jacas et al., (1992a)
Poltiglia,
Manica 20 SPA 1kg/hl (cp)
T pupa of C. capitata 64.8 ± 4.3 4 98.7 ± 0.9 37.9 ± 10.4
Bengochea et al., (2014) R glass (Potter spray tower) 0.0 ± 0.0 91.8 ± 3.7 60.2 ± 4.8
Sf olive tree 3.3 ± 1.9 90.2 ± 6.5 54.0 ± 3.5
HERBICIDES
Fluroxypyr O Starane, Dow
Elanco 0.090 R Potter spray tower 2 (IOBC) - - Jacas and Viñuela (1994a)
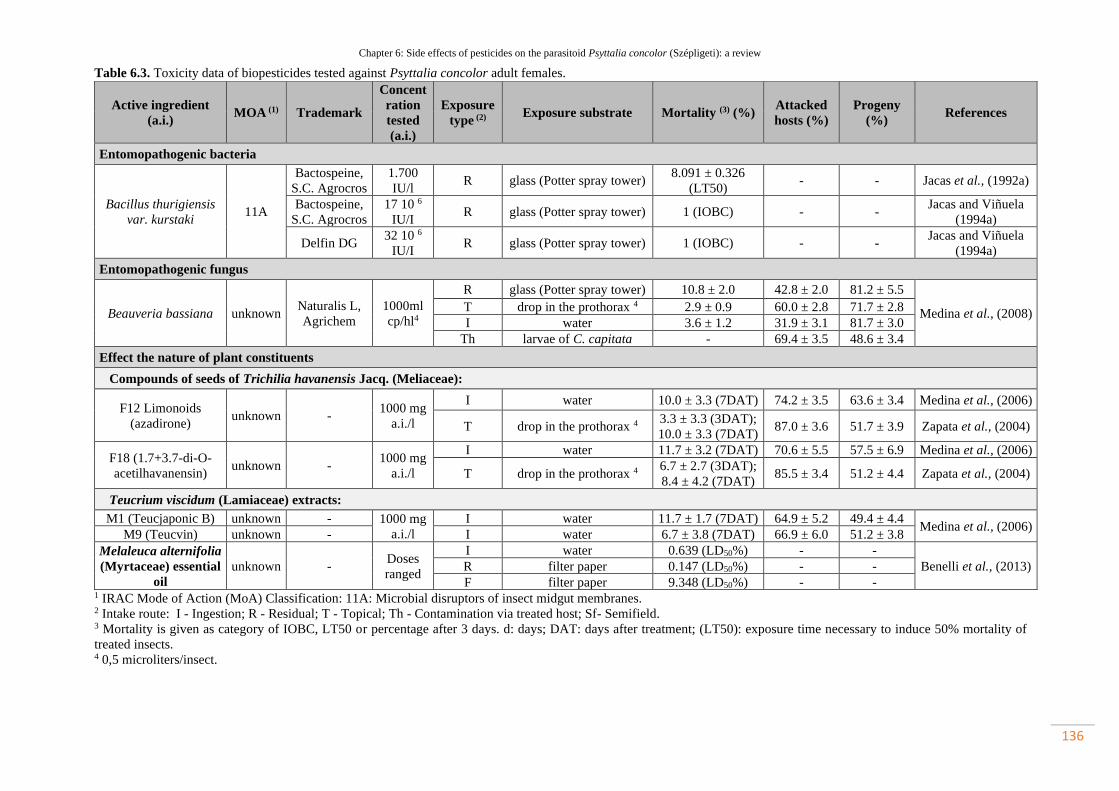
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
136
Table 6.3. Toxicity data of biopesticides tested against Psyttalia concolor adult females.
1 IRAC Mode of Action (MoA) Classification: 11A: Microbial disruptors of insect midgut membranes. 2 Intake route: I - Ingestion; R - Residual; T - Topical; Th - Contamination via treated host; Sf- Semifield. 3 Mortality is given as category of IOBC, LT50 or percentage after 3 days. d: days; DAT: days after treatment; (LT50): exposure time necessary to induce 50% mortality of
treated insects. 4 0,5 microliters/insect.
Active ingredient
(a.i.) MOA (1) Trademark
Concent
ration
tested
(a.i.)
Exposure
type (2) Exposure substrate Mortality (3) (%)
Attacked
hosts (%)
Progeny
(%) References
Entomopathogenic bacteria
Bacillus thurigiensis
var. kurstaki 11A
Bactospeine,
S.C. Agrocros
1.700
IU/l R glass (Potter spray tower)
8.091 ± 0.326
(LT50) - - Jacas et al., (1992a)
Bactospeine,
S.C. Agrocros
17 10 6
IU/I R glass (Potter spray tower) 1 (IOBC) - -
Jacas and Viñuela
(1994a)
Delfin DG 32 10 6
IU/I R glass (Potter spray tower) 1 (IOBC) - -
Jacas and Viñuela
(1994a)
Entomopathogenic fungus
Beauveria bassiana unknown Naturalis L,
Agrichem
1000ml
cp/hl4
R glass (Potter spray tower) 10.8 ± 2.0 42.8 ± 2.0 81.2 ± 5.5
Medina et al., (2008) T drop in the prothorax 4 2.9 ± 0.9 60.0 ± 2.8 71.7 ± 2.8
I water 3.6 ± 1.2 31.9 ± 3.1 81.7 ± 3.0
Th larvae of C. capitata - 69.4 ± 3.5 48.6 ± 3.4
Effect the nature of plant constituents
Compounds of seeds of Trichilia havanensis Jacq. (Meliaceae):
F12 Limonoids
(azadirone) unknown -
1000 mg
a.i./l
I water 10.0 ± 3.3 (7DAT) 74.2 ± 3.5 63.6 ± 3.4 Medina et al., (2006)
T drop in the prothorax 4 3.3 ± 3.3 (3DAT);
10.0 ± 3.3 (7DAT) 87.0 ± 3.6 51.7 ± 3.9 Zapata et al., (2004)
F18 (1.7+3.7-di-O-
acetilhavanensin) unknown -
1000 mg
a.i./l
I water 11.7 ± 3.2 (7DAT) 70.6 ± 5.5 57.5 ± 6.9 Medina et al., (2006)
T drop in the prothorax 4 6.7 ± 2.7 (3DAT);
8.4 ± 4.2 (7DAT) 85.5 ± 3.4 51.2 ± 4.4 Zapata et al., (2004)
Teucrium viscidum (Lamiaceae) extracts:
M1 (Teucjaponic B) unknown - 1000 mg
a.i./l
I water 11.7 ± 1.7 (7DAT) 64.9 ± 5.2 49.4 ± 4.4 Medina et al., (2006)
M9 (Teucvin) unknown - I water 6.7 ± 3.8 (7DAT) 66.9 ± 6.0 51.2 ± 3.8
Melaleuca alternifolia
(Myrtaceae) essential
oil
unknown - Doses
ranged
I water 0.639 (LD50%) - -
Benelli et al., (2013) R filter paper 0.147 (LD50%) - -
F filter paper 9.348 (LD50%) - -

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
137
4. LABORATORY CASE STUDY: EFFECTS OF DIFFERENT PESTICIDES ON
MORTALITY AND REPRODUCTION OF PSYTTALIA CONCOLOR FEMALES
4.1. MATERIAL AND METHODS
4.1.1. Psyttalia concolor and Ceratitis capitata rearing conditions
Psyttalia concolor adults were obtained from a mass-rearing culture on the medfly,
Ceratitis capitata (Wiedemann) (Dip. Tephritidae) installed in a laboratory (Crop
Protection Unit, ETSIAAB-UPM, Madrid, Spain). C. capitata was used as a substitution
host of B. oleae. Mass-rearing cultures of both insects were maintained in a chamber with
controlled environmental conditions (25±2◦C, 75±10% relative humidity and 16:8 L:D
photoperiod). For rearing details see Jacas and Viñuela (1994b) and Bengochea et al.
(2014).
The choice of females of P. concolor for the experiments was due to its longer life
span compared to than males (Pinheiro et al., 2017), its higher tolerance to pesticides and
only mated females can parasitise a host in the field (Jacas and Viñuela, 1994a).
4.1.2. Pesticides
Eight pesticides registered for different uses against important olive pests in Portugal
and/or in Spain were tested (DGAV, 2018; MAPAMA, 2018): one herbicide
(glyphosate), three insecticides (beta-cyfluthrin, deltamethrin and zeta-cypermethrin) and
four fungicides (difenoconazole, dodine, tebuconazole and trifloxystrobin). Data on trade
name, chemical group and site of action according to HRAC, FRAC and IRAC
classification, maximum application rate following MAPAMA (2018) and potential
target are summarised in Table 6.4.
Dimethoate, a systemic insecticide, was used as positive control due to its efficacy
controlling various olive-attacking pests and because it is the most commonly applied
insecticide against B. oleae in Mediterranean areas, including Iberian olive groves (Ruiz-
Torres and Montiel-Bueno 2007; Pavlidi et al., 2018). Distilled water was used as
negative control.
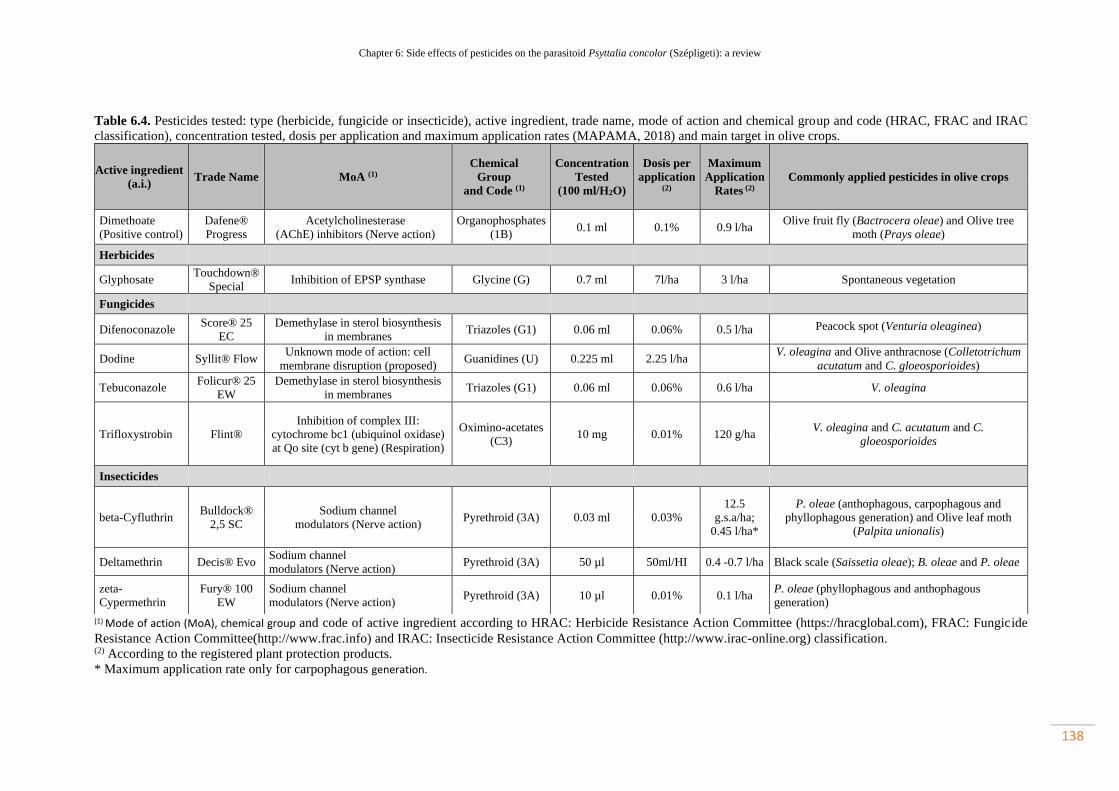
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
138
Table 6.4. Pesticides tested: type (herbicide, fungicide or insecticide), active ingredient, trade name, mode of action and chemical group and code (HRAC, FRAC and IRAC
classification), concentration tested, dosis per application and maximum application rates (MAPAMA, 2018) and main target in olive crops.
(1) Mode of action (MoA), chemical group and code of active ingredient according to HRAC: Herbicide Resistance Action Committee (https://hracglobal.com), FRAC: Fungicide
Resistance Action Committee(http://www.frac.info) and IRAC: Insecticide Resistance Action Committee (http://www.irac-online.org) classification. (2) According to the registered plant protection products.
* Maximum application rate only for carpophagous generation.
Active ingredient
(a.i.) Trade Name MoA (1)
Chemical
Group
and Code (1)
Concentration
Tested
(100 ml/H2O)
Dosis per
application (2)
Maximum
Application
Rates (2)
Commonly applied pesticides in olive crops
Dimethoate
(Positive control)
Dafene®
Progress
Acetylcholinesterase
(AChE) inhibitors (Nerve action)
Organophosphates
(1B) 0.1 ml 0.1% 0.9 l/ha
Olive fruit fly (Bactrocera oleae) and Olive tree
moth (Prays oleae)
Herbicides
Glyphosate Touchdown®
Special Inhibition of EPSP synthase Glycine (G) 0.7 ml 7l/ha 3 l/ha Spontaneous vegetation
Fungicides
Difenoconazole Score® 25
EC
Demethylase in sterol biosynthesis
in membranes Triazoles (G1) 0.06 ml 0.06% 0.5 l/ha Peacock spot (Venturia oleaginea)
Dodine Syllit® Flow Unknown mode of action: cell
membrane disruption (proposed) Guanidines (U) 0.225 ml 2.25 l/ha V. oleagina and Olive anthracnose (Colletotrichum
acutatum and C. gloeosporioides)
Tebuconazole Folicur® 25
EW
Demethylase in sterol biosynthesis
in membranes Triazoles (G1) 0.06 ml 0.06% 0.6 l/ha V. oleagina
Trifloxystrobin Flint®
Inhibition of complex III:
cytochrome bc1 (ubiquinol oxidase)
at Qo site (cyt b gene) (Respiration)
Oximino-acetates
(C3) 10 mg 0.01% 120 g/ha
V. oleagina and C. acutatum and C.
gloeosporioides
Insecticides
beta-Cyfluthrin Bulldock®
2,5 SC
Sodium channel
modulators (Nerve action) Pyrethroid (3A) 0.03 ml 0.03%
12.5
g.s.a/ha;
0.45 l/ha*
P. oleae (anthophagous, carpophagous and
phyllophagous generation) and Olive leaf moth
(Palpita unionalis)
Deltamethrin Decis® Evo Sodium channel
modulators (Nerve action) Pyrethroid (3A) 50 µl 50ml/HI 0.4 -0.7 l/ha Black scale (Saissetia oleae); B. oleae and P. oleae
zeta-
Cypermethrin
Fury® 100
EW
Sodium channel
modulators (Nerve action) Pyrethroid (3A) 10 µl 0.01% 0.1 l/ha
P. oleae (phyllophagous and anthophagous
generation)

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
139
4.1.3. Laboratory bioassays
Psyttalia concolor adults were exposed to pesticides by two pathways: contact using
olive tree leaves sprayed with the pesticide and through ingestion of the pesticide.
Parameters evaluated were: (1) mortality of the female parasitoid P. concolor (%) and (2)
reproduction, i.e., attacked hosts (%) (percentage of puparia without medfly emergence)
and progeny size (%) (percentage of parasitoids emerged from parasitized medfly
puparia) (see Bengochea et al., 2014 for details). The experiments were performed at the
same controlled laboratory conditions described above.
4.1.4. Contact bioassay
After adult P. concolor emergence, ten mated females (< 48-h-old) were placed in a
plexiglass cage (12 cm in diameter x 5 cm height) with a hole on the top (5.5 cm in
diameter) covered with a net for ventilation. In each test cage, one glass vial (30 x 35
mm2) with distilled water was provided as water and food (brewerʾs yeast and sugar in
the proportion of 1:4) was provided in plastic stoppers (24 x 6 mm2) ad libitum to P.
concolor females. One olive branch (cv. Picual; 7-y-old) with six leaves grown in the
greenhouses of the ETSIAAB-UPM was treated with the corresponding fresh solutions
of each commercial substance or water (negative control) using hand sprayers and offered
to the insects when dried (approximately, 1 hour after application) (Fig. 6.3.) Five
replicates per treatment were tested in mortality tests. The effect of the pesticides was
scored for 2, 4, 24, 48 and 72 hours after treatment (HAT). After 72 hours, 25 females in
total (five replicates per treatment) in treatments with a survival rate higher than 50%,
were chosen to test sublethal effects on reproduction for 5 days. Data from the first
parasitisation day were rejected because they can be very erratic as females need one day
at least to learn how to parasitise.
During those consecutive five days, every day for one hour, 3rd instar larvae of C.
capitata were offered to P. concolor in a 20 larvae/female proportion. The larvae of C.
capitata exposed to parasitisation were transferred to Petri dish to pupate and the
percentages of attacked host and progeny size were measured approximately one month
after parasitisation. The cage of parasitisation is similar to the one described for mortality
tests but with an additional hole (5.5 cm in diameter) on the bottom of the cage covered
with a net. Larvae were offered to P. concolor on a square of Parafilm® (10,2 cm2) placed

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
140
on the top of a glass pot and strapped with two rubber bands per cage of parasitisation in
order to immobilise the larvae (Fig. 6.4.) (see Bengochea et al., 2014 for details).
Fig. 6.3. Contact bioassay: A: Psyttalia concolor female; B: Experimental arena
used to evaluate the mortality with (1) distilled water, (2) diet and (3) treated olive
branch; C: Olive tree (cv. Picual) leaves treated with a pesticide using hand spray;
D and E: Olive branch totally covered with the pesticide to dry.
Fig. 6.4: A: Parasitisation cages used to evaluate the sublethal effects of pesticides
on Psyttalia concolor reproduction. (1) Female wasp individually to parasited, (2)
20 larvae (3rd instar) of C. capitata offered to P. concolor.

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
141
4.1.5. Ingestion bioassay
In this experiment, each experimental unit was formed by a cage containing a glass
vial (2 ml) with the corresponding solution of each pesticide and the solid diet was placed
on a plastic stopper cover ad libitum. No olive branches were introduced. Ten females (<
24h-old) per replicate and 5 replicates per treatment were used. Mortality and
reproduction were evaluated as described in the previous treatment.
4.1.6. Data analysis
Mortality rates failed the assumptions of ANOVA and a non-parametric Kruskal-
Wallis test was applied, with a pairwise comparison. Attacked hosts and progeny size
were analyzed using one-way analysis of variance (ANOVA) and Fisher's least
significant difference (LSD) test. Statistical analyses were performed with IBM-SPSS
statistics, version 24.0 (SPSS Inc. IBM Company, 2016). The normal distribution of the
residuals and the homogeneity of variances were evaluated by means of the Kolmogorov–
Smirnov and Levene’s tests, respectively. All statistical tests were performed at 5 %
significance level. Data are presented in the Tables 6.5., 6.6. and 6.7. (mean values ± SE).
Subsequently, data were interpreted according to the IOBC standards (Sterk et al.,
1999) in 1, harmless (< 30%); 2, slightly harmful (30-79%); 3, moderately harmful (80-
99%); and 4, harmful (>99%) for laboratory test (exposure via ingestion) and as 1,
harmless (< 25%); 2, slightly harmful (25-50%); 3, moderately harmful (51-75%); and 4
= harmful (>75%) for extended laboratory test (when females were exposed exposure to
pesticides residues on olive tree leaves).
4.2. RESULTS
4.2.1. Mortality of adult females
Mortality on exposure via contact and ingestion for P. concolor displayed different
levels of mortality (0-100%).
Mortality induced by insecticides was significantly different from the untreated control
72 HAT in the extended laboratory test (Table 6.5.). Beta-cyfluthrin, deltamethrin and
zeta-cypermethrin showed the highest impact on P. concolor adults resulting as harmful
(IOBC category 4). The remaining tested products were considered harmless (IOBC
category 1) (Table 6.5.).
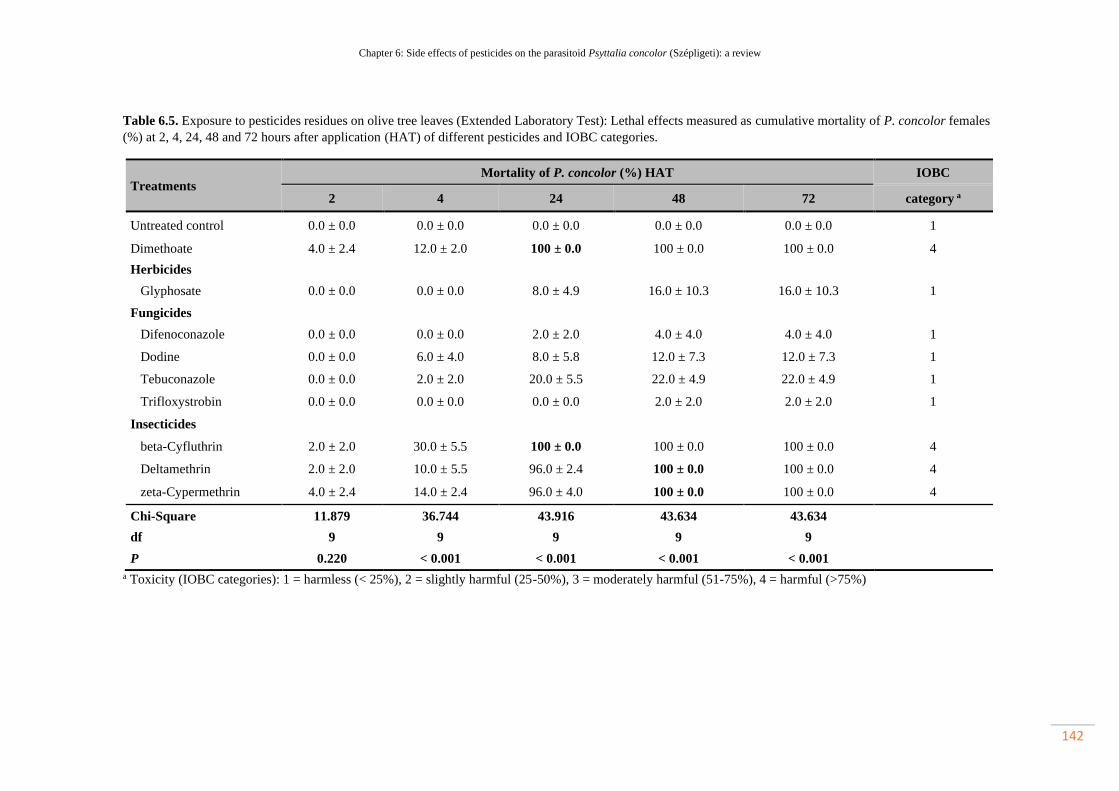
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
142
Table 6.5. Exposure to pesticides residues on olive tree leaves (Extended Laboratory Test): Lethal effects measured as cumulative mortality of P. concolor females
(%) at 2, 4, 24, 48 and 72 hours after application (HAT) of different pesticides and IOBC categories.
a Toxicity (IOBC categories): 1 = harmless (< 25%), 2 = slightly harmful (25-50%), 3 = moderately harmful (51-75%), 4 = harmful (>75%)
Treatments Mortality of P. concolor (%) HAT IOBC
2 4 24 48 72 category a
Untreated control 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 1
Dimethoate 4.0 ± 2.4 12.0 ± 2.0 100 ± 0.0 100 ± 0.0 100 ± 0.0 4
Herbicides
Glyphosate 0.0 ± 0.0 0.0 ± 0.0 8.0 ± 4.9 16.0 ± 10.3 16.0 ± 10.3 1
Fungicides
Difenoconazole 0.0 ± 0.0 0.0 ± 0.0 2.0 ± 2.0 4.0 ± 4.0 4.0 ± 4.0 1
Dodine 0.0 ± 0.0 6.0 ± 4.0 8.0 ± 5.8 12.0 ± 7.3 12.0 ± 7.3 1
Tebuconazole 0.0 ± 0.0 2.0 ± 2.0 20.0 ± 5.5 22.0 ± 4.9 22.0 ± 4.9 1
Trifloxystrobin 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 2.0 ± 2.0 2.0 ± 2.0 1
Insecticides
beta-Cyfluthrin 2.0 ± 2.0 30.0 ± 5.5 100 ± 0.0 100 ± 0.0 100 ± 0.0 4
Deltamethrin 2.0 ± 2.0 10.0 ± 5.5 96.0 ± 2.4 100 ± 0.0 100 ± 0.0 4
zeta-Cypermethrin 4.0 ± 2.4 14.0 ± 2.4 96.0 ± 4.0 100 ± 0.0 100 ± 0.0 4
Chi-Square 11.879 36.744 43.916 43.634 43.634
df 9 9 9 9 9
P 0.220 < 0.001 < 0.001 < 0.001 < 0.001

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
143
In exposure to pesticide via ingestion, difenoconazole and trifloxystrobin fungicides
did not cause any deleterious effect on the mortality of P. concolor even 72 hours after
treatment. Glyphosate, tebuconazole and zeta-cypermethrin caused low mortality (IOBC
category 2). Beta-cyfluthrin and dodine caused 80 and 88% mortality, respectively, being
classified as moderately harmful (IOBC category 3). Deltamethrin had the highest impact
on P. concolor, killing 100% of the adults 72 hours after treatment (IOBC category 4)
(Table 6.6.).
There were statistically significant differences between the different treatments at 4,
24, 48- and 72-hours HAT for both exposure modes (Table 6.5. and Table 6.6.). The
exposure via ingestion (p = 0.063) and contact (p = 0.220) to pesticides for 2 hours did
not have a significant impact on mortality of P. concolor. The significant pairwise
comparisons of treatments are presented in supplemental material.
4.2.2. Sublethal effects on reproduction: attacked hosts and progeny size
Non-significant differences were found between treatments and untreated control for
reproductive parameters: attacked hosts and progeny size in both modes of exposure
(Table 6.7.). The percentage of attacked hosts was higher than 95% in all cases. However,
the percentage of progeny was more variable, with a rate of 50 to 57% (Table 6.7.). These
lower percentages may be possibly related to fitness of the female and/or some specific
characteristics of medfly larvae used in experiments.
4.3. DISCUSSION
4.3.1. Insecticide exposure
The major classes of synthetic pesticides are organochlorines, organophosphates,
carbamates and pyrethroids. (Bora, Khanikor and Gogoi, 2012). The first three classes
were initially used in the control of the olive fruit fly, followed by the introduction of
pyrethroids, spinosad and other compounds (Vontas et al., 2011).
Carbamates and organophosphates (group 1B, IRAC) are considered broad spectrum
insecticides, with low selectivity and high acute toxicity. Both act in a similar way against
pests and natural enemies, inactivating acetylcholinesterase (AChE) and, consequently,
causing an over-stimulation of organs due to an excessive level of acetylcholine at the
nerve endings (Uma, Jacob and Lyla, 2014). However, the quantitative and qualitative
changes in AChE mechanism conferred resistance to insecticides (Vontas et al., 2001).
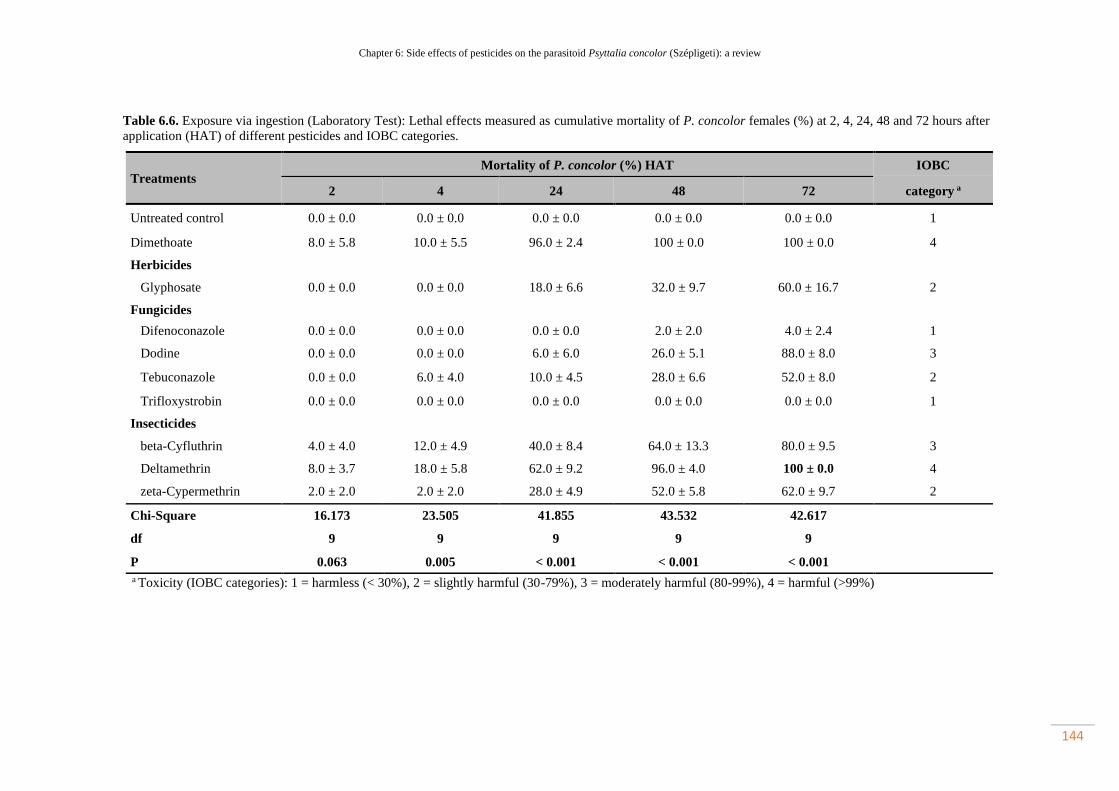
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
144
Table 6.6. Exposure via ingestion (Laboratory Test): Lethal effects measured as cumulative mortality of P. concolor females (%) at 2, 4, 24, 48 and 72 hours after
application (HAT) of different pesticides and IOBC categories.
a Toxicity (IOBC categories): 1 = harmless (< 30%), 2 = slightly harmful (30-79%), 3 = moderately harmful (80-99%), 4 = harmful (>99%)
Treatments Mortality of P. concolor (%) HAT IOBC
2 4 24 48 72 category a
Untreated control 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 1
Dimethoate 8.0 ± 5.8 10.0 ± 5.5 96.0 ± 2.4 100 ± 0.0 100 ± 0.0 4
Herbicides
Glyphosate 0.0 ± 0.0 0.0 ± 0.0 18.0 ± 6.6 32.0 ± 9.7 60.0 ± 16.7 2
Fungicides
Difenoconazole 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 2.0 ± 2.0 4.0 ± 2.4 1
Dodine 0.0 ± 0.0 0.0 ± 0.0 6.0 ± 6.0 26.0 ± 5.1 88.0 ± 8.0 3
Tebuconazole 0.0 ± 0.0 6.0 ± 4.0 10.0 ± 4.5 28.0 ± 6.6 52.0 ± 8.0 2
Trifloxystrobin 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 0.0 ± 0.0 1
Insecticides
beta-Cyfluthrin 4.0 ± 4.0 12.0 ± 4.9 40.0 ± 8.4 64.0 ± 13.3 80.0 ± 9.5 3
Deltamethrin 8.0 ± 3.7 18.0 ± 5.8 62.0 ± 9.2 96.0 ± 4.0 100 ± 0.0 4
zeta-Cypermethrin 2.0 ± 2.0 2.0 ± 2.0 28.0 ± 4.9 52.0 ± 5.8 62.0 ± 9.7 2
Chi-Square 16.173 23.505 41.855 43.532 42.617
df 9 9 9 9 9
P 0.063 0.005 < 0.001 < 0.001 < 0.001
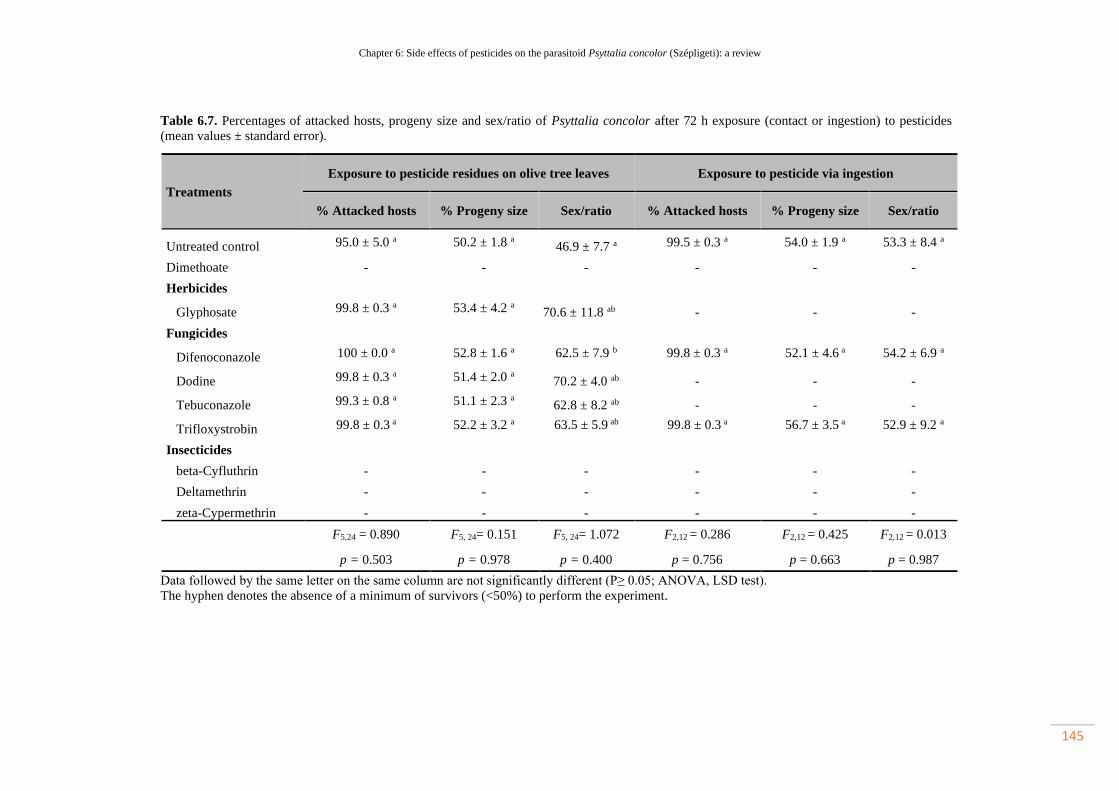
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
145
Table 6.7. Percentages of attacked hosts, progeny size and sex/ratio of Psyttalia concolor after 72 h exposure (contact or ingestion) to pesticides
(mean values ± standard error).
Data followed by the same letter on the same column are not significantly different (P≥ 0.05; ANOVA, LSD test).
The hyphen denotes the absence of a minimum of survivors (<50%) to perform the experiment.
Treatments
Exposure to pesticide residues on olive tree leaves Exposure to pesticide via ingestion
% Attacked hosts % Progeny size Sex/ratio % Attacked hosts % Progeny size Sex/ratio
Untreated control 95.0 ± 5.0 a 50.2 ± 1.8 a 46.9 ± 7.7 a 99.5 ± 0.3 a 54.0 ± 1.9 a 53.3 ± 8.4 a
Dimethoate - - - - - -
Herbicides
Glyphosate 99.8 ± 0.3 a 53.4 ± 4.2 a 70.6 ± 11.8 ab - - -
Fungicides
Difenoconazole 100 ± 0.0 a 52.8 ± 1.6 a 62.5 ± 7.9 b 99.8 ± 0.3 a 52.1 ± 4.6 a 54.2 ± 6.9 a
Dodine 99.8 ± 0.3 a 51.4 ± 2.0 a 70.2 ± 4.0 ab - - -
Tebuconazole 99.3 ± 0.8 a 51.1 ± 2.3 a 62.8 ± 8.2 ab - - -
Trifloxystrobin 99.8 ± 0.3 a 52.2 ± 3.2 a 63.5 ± 5.9 ab 99.8 ± 0.3 a 56.7 ± 3.5 a 52.9 ± 9.2 a
Insecticides
beta-Cyfluthrin - - - - - -
Deltamethrin - - - - - -
zeta-Cypermethrin - - - - - -
F5,24 = 0.890 F5, 24= 0.151 F5, 24= 1.072 F2,12 = 0.286 F2,12 = 0.425 F2,12 = 0.013
p = 0.503 p = 0.978 p = 0.400 p = 0.756 p = 0.663 p = 0.987

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
146
The resistance of the olive fruit fly to carbamates and organophosphates (usually
dimethoate) were reported in laboratory and field populations (Vontas et al., 2001;
Skouras et al., 2007; Vontas et al., 2011). The applications of these insecticides do not
just kill the olive fruit fly larvae (which live inside the drupe) but also have a negative
effect on arthropod populations (e.g., Santos et al., 2007).
To the best of our knowledge, and to what concerns P. concolor, only carbamate
insecticide (Carbaryl) showed to be harmful (Jacas et al., 1992b; Jacas and Viñuela,
1994a) and it is currently banned (not in Tables). In contrast, organophosphates, the most
studied insecticide group for the parasitoid, exhibited highly toxicity to P. concolor in
laboratory and semi-field trials (Table 6.1.). Chlorpyrifos-methyl and phosmet resulted to
be harmful to adults of P. concolor in laboratory tests, while dimethoate is considered
harmful at the laboratory and semi-field levels with mortalities above 98% at 72h after
treatments for all available studies (see references in the Table 6.1., 6.5. and 6.6.). This
percentage of mortality compromises the emergence of new individuals in the field due
to the lifespan of females (live long enough to oviposit; Bengochea et al., 2014). Some
adult parasitoids may have emerged from treated pupae (after dimethoate application)
contributing to the repopulation of P. concolor, but in a reduced percentage when
compared to untreated pupae (Bengochea et al., 2014). This toxicity of organophosphates
has been reported for other braconid parasitoids, e.g. Aphidius gifuensis Ashmead (Ohta
and Takeda, 2015) and Diachasmimorpha longicaudata Ashmed (Harbi et al., 2017).
According to Longley (1999), organophosphates are probably more toxic for the
development of parasitoids than pyrethroids or carbamates. Furthermore,
organophosphates have higher environmental impact and contaminating olive oils,
especially dimethoate. Pyrethroids, pyrethrins (group 3A, IRAC) are neurotoxic agents
that are rapidly absorbed by the insect integument in contact with the treated surface and
alter the function of voltage-sensitive sodium channels, interrupting the normal
transmission of nerve impulses. Thus, they have a quick action, causing hyperactivity,
convulsions and an immediate “knockdown” paralysis in insects (El-Wakeil, 2013;
Morais et al., 2016). They are commonly used against B. oleae as substitute for
dimethoate. The resistance to pyrethroid in B. oleae is more recent and with limited
number of resistances cases documented (Vontas et al., 2011), being referenced resistance
levels to alpha-cypermethrin (Margaritopoulos et al., 2008). For P. concolor, it has been
demonstrated that pyrethroids, alpha-cypermethrin and lambda-cyhalothrin have a
negative effect on survival (Table 6.1.). In general, we found similarly results for

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
147
deltamethrin, beta-cyfluthrin and zeta-cypermethrin in ingestion and contact bioassay
(see Table 6.5. and 6.6.). Specifically, only zeta-cypermethrin in ingestion test were
slightly harmful (Table 6.6.). The effect of pyrethroids in adults of other parasitoid species
were well documented by several authors as causing short-term mortality (e.g. Prabhaker
et al., 2007; Bayram et al., 2010; Morais et al., 2016; Garantonakis, Varikou and
Birouraki, 2016). Additionally, the effects on larval and pupal development, fecundity,
sex-ratio and oviposition behavior have been observed (Youssef et al., 2004; Ergin et al.,
2007; Vianna et al., 2009; Bayram et al., 2010). Moreover, pyrethroids interfere with the
mobility and orientation of parasitoids. It is well known that parasitoids spend a
significant proportion of their adult stage foraging and searching for hosts. This process
involves orientation to food source or host and plant odours or host-plant odours
performing the nervous transmissions an important role (target of pyrethroids) (Desneux,
Rafalimanana and Kaiser, 2004; Schoonhoven, Van Loon and Dicke, 2005). After
intoxication, insects present symptoms of loss of coordinated movement, periods of
convulsive activity and paralysis (Soderlund and Bloomquist, 1989). Some authors
defend that foraging and orientation behaviour induced by sublethal effects doses of some
pyrethroids in parasitoids can be normalized over a 12 to 24 hours after exposure to
insecticide (Longley and Jepson, 1996; Desneux, Pham-Delègue, and Kaiser, 2004;
Desneux, Denoyelle and Kaiser, 2006). We think that due to the knowledge of the
mechanism of action of these types of insecticide, together with the knowledge that
predicts a high risk of mortality at the field and interfere with multiple vital functions in
parasitoids (including reproduction, mobility, feeding, mating, host searching; Desneux,
Decourtye and Delpuech, 2007) pyrethroids should not be recommended in IPM
programs.
According to Kirst (2010), spinosad (group 5, IRAC) is a defined combination of the
two main fermentation factors, spinosyns A and D, produced by the actinomycete
Saccharopolyspora spinosa. Spinosyns have a unique mechanism of action (MoA)
involving disruption of nicotinic acetylcholine receptors and presenting a high selectivity
against target insects, harmless for most natural enemies and with reduced risks to the
environment (Kirst, 2010; Williams, Valle and Viñuela, 2003). Recently, spinosad was
introduced in bait sprays against olive fruit fly (Gonçalves et al., 2012) as an insecticide
that rapidly degrades in the field, with little residual toxicity at 3-7 days post application
(Williams, Valle and Viñuela, 2003). For these reasons, spinosad is recommended as an
important option for pest control namely for Integrated Pest Management (IPM) and

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
148
organic production systems (Biondi et al., 2012). However, long-term use of this
insecticide may lead to resistance problems in the olive fruit fly (Kakani et al., 2010).
There is some controversy regarding its effects on the natural enemies. Williams, Valle
and Viñuela (2003), in a review over the impact of spinosad on natural enemies, showed
that 71% of laboratory studies and 79% of field-type studies resulted as harmless for
predators and moderately harmful or harmful for parasitoids (with 78% of laboratory
studies and 86% of field-type studies). In a field trial of 2 years in olive groves located
in Trás-os-Montes (Portugal), baited spinosad treatments showed to be effective against
the olive fruit fly without harmful effects on the natural enemies community (Gonçalves
et al., 2012). Also, Pascual et al. (2014) in a field study conducted in olive trees treated
with spinosad bait sprays in Villarejo de Salvanés (Madrid, Spain) found compatibility
between the insecticide and most of the natural enemies, except for Orius spp. and
Aphelinidae. In contrast, Ruiz Torres and Montiel Bueno (2007) reported slight effects
on natural enemies in olive orchards of Jaén's province (Andalusia, Spain). For P.
concolor, when Spinosad is applied under laboratory condition, at the maximum field
recommended rates with different techniques (via Ingestion, residual and topical
application), spinosad was very harmful (Table 6.1.). Due to the high mortality (almost
100%), the percentage of attacked hosts and the progeny size was not determined
(Viñuela et al., 2001; Table 6.1.). Similarly, higher acute lethal toxicity to the parasitoid
Aphidius colemani Viereck (Hymenoptera: Braconidae) was found, not being possible to
subject the parasitoids to experiments to evaluate the effects on parasitism and progeny
production (see D’Ávila et al., 2018), concluding that “bioinsecticide products like
spinosad can be more toxic to biological control agents than certain conventional
insecticides”. In contrast, Stark, Vargas and Miller (2004) demonstrated that spinosad,
when used in bait-sprays do not present risks for two parasitoids of tephritid fruit fly
species, Fopius arisanus (Sonan) and Pyttalia fletcheri (Silvestri).
The insecticide fenoxycarb (group 7B, IRAC) is a carbamate Insect Growth Regulator
(IGR). Is used against various Lepidoptera and scale insects of fruits citrus, vineyards and
olives and different species of Diptera (Arzone et al., 1995; Bengochea et al., 2012). More
recently, it was applied as bait treatments for the control of olive fruit fly (Sánchez-Ramos
et al., 2013). Several authors refer that IGRs have contact and stomach action and
interfere with the normal development and/or reproduction of insects (Michel, Krause
and Buszewski, 2001; Sánchez-Ramos et al., 2013). The potential toxicity and directly
effects of fenoxycarb on different pest and non-target insects were reviewed by Grenier

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
149
and Grenier (1993). More recently, it was found that fenoxycarb can be compatible with
parasitoids (Hoddle et al., 2001). Results with P. concolor indicate that fenoxycarb does
not affect the longevity and emergence of parasitoid (Table 6.1., for more details see Jacas
and Viñuela, 1994a; González, Bahena and Viñuela, 1998). Other studies obtained similar
result and found that fenoxycarb was slightly harmful for other parasitoids, such as,
Encarsia formosa (Hymenoptera: Aphelinidae) larvae (Wang and Liu, 2016). Another
problematic feature of fenoxycarb is its low soil mobility, non-accumulation, and its quick
breaking down in the environment (Michel, Krause and Buszewski, 2001). Although it is
important to increase the knowledge on how fenoxycarb affects P. concolor, apparently
this insecticide could be compatible with the parasitoid and a rational candidate for IPM.
Considering pyriproxyfen (group 7C, IRAC), another IGR, Abdallahi, Adán and Viñuela
(2000) obtained slightly harmful effects in P. concolor mortality but harmless for the
parasitoids reproduction (Table 6.1.). In contrast, pyriproxyfen caused high levels of
mortality on Eretmocerus eremicus (Hymenoptera: Aphelinidae) (Hoddle et al., 2001)
and was harmful to Encarsia formosa (Hymenoptera: Aphelinidae) larvae and moderately
harmful to pupae (Wang and Liu, 2016). However, the application of pyriproxyfen highly
decreases the rates of the mature and emerged parasitoids (Wang and Liu, 2016).
Moreover, recent studies about the interference of IGRs on reproduction showed a strong
evidence of occurring vertical transmission of pyriproxyfen from the treated parent to the
egg (Penca and Hodges, 2017). Therefore, it is important to clarify the compatibility of
IGRs with biological control agents under field conditions.
Kaolin is a white aluminosilicate mineral clay, non-porous, low-abrasive that easily
disperses in water and is chemically inert over a wide pH range (Glenn and Puterka,
2005). When applied against insects, it works as a physical barrier and reduces the
oviposition rate of several pests (Bengochea et al., 2013). In olive groves is reported as
an alternative to insecticides that control the olive fly (Bengochea et al., 2014) and the
olive moth, Prays oleae (Bernard) (Lepidopteta: Praydidae) (De la Roca, 2003). Effects
of kaolin and copper salts on natural enemies have been studied at laboratory and field
level. In olive groves, some authors refer a reduction of abundance and diversity of the
arthropod community (e.g. Benhadi-Marín, Pereira and Santos, 2016; Bengochea et al.,
2013; Pascual et al., 2010) probably to its repellent/deterrent effect (Glenn and Puterka,
2005). For parasitoid P. concolor, Kaolin showed to be harmless and no lethal toxicity
or effects on reproduction were observed at laboratory and semi-field conditions (Table
6.1., for details see Bengochea et al., 2010; 2014). However, a slight reduction in the

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
150
percentage of parasitoids hosts was recorded in Kaolin-treated surface (Bengochea et al.,
2014) due to the facts pointed above.
As a general conclusion, Kaolin is a recommendable pest control product in olive
groves because it reduces the attack of important pest species with a lower impact when
compared by other products commonly used, such as dimethoate (Bengochea et al., 2013;
Pascual et al., 2010).
4.3.2. Fungicide exposure
Anthracnose (Colletotrichum spp. complex) is an important disease of olive fruits,
causing significant yield losses and poor fruit and oil quality (Talhinhas et al., 2011). The
control of the disease is usually associated to copper‐based fungicides, but trifloxystrobin
and dodine can also be used (Talhinhas, Loureiro and Oliveira, 2018; Cacciola et al.,
2012). Another important disease of Olea europaea L. is olive leaf spot, also known as
olive scab or peacock's eye disease (Venturia oleaginea (Castagne)) Rossman & Crous
(Romero et al., 2018). Traditionally, fungicides containing copper (Bordeaux mixture,
copper hydroxide, copper oxide and copper oxychlorides) are used in the control of this
pathogen. However, some long-persisting preventive fungicides, such as, difenoconazole
(Group G1, FRAC), dodine (Group U, FRAC), tebuconazole (Group G1, FRAC) and
trifloxystrobin (Group C3, FRAC), have been applied (D'Ascenzo, Crivelli, and Camillo,
2014; Iannotta, Monardo and Perri, 2002; Obanor et al., 2008; Romero et al., 2018).
According to our results, difenoconazole, tebuconazole, trifloxystrobin and dodine are
harmless by contact assays (Extended Laboratory Test) (Table 6.5.). When applied via
ingestion, dodine was moderately harmful and tebuconazole was slightly harmful (Table
6.6.). Moreover, no negative effects were reported for attacked host and progeny size in
both experiments (Table 6.7.). On the other hand, Jacas and Viñuela (1994a) showed that
difenoconazole was slightly harmful and tebuconazole was moderately harmful for P.
concolor adult females exposed to a fresh residue on a glass surface (Table 6.2.).
Concerning Bordeaux mixture and copper oxychloride, the first was slightly harmful
and copper oxide was moderately harmful (Jacas et al., 1992a). Bengochea et al. (2014)
showed that Bordeaux mixture and copper oxychloride were not toxic to P. concolor in
the laboratory and semi-field experiments. In this context, copper-based compounds ca
be considered compatible with parasitoids and can be used in IPM or organic farming
programmes, not only for controlling fungal diseases but also as a possible alternative to
traditionally insecticides for controlling the olive fruit fly (Bengochea et al., 2014).

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
151
Sulphur (group M2, FRAC) is a contact fungicide commonly used in pest management
programs. In the olive grove, it reduced the incidence of anthracnose (had only protective
activity) (Nigro et al., 2018). However, there is some evidence that sulphur had negative
effects on natural enemies, namely on parasitoids (e.g. Hassan et al., 1998; Jepsen,
Rosenheim and Matthews, 2007). A different result showed by Jacas and Viñuela (1992a;
Table 6.2.) demonstrated that sulphur could be considered as harmless to P. concolor.
However, according to Dugravot et al. (2002) sulphur volatiles can be toxic and influence
the metabolism or the activity of the nervous system.
Similarly, mancozeb (group M3, FRAC), other protectant fungicide of anthracnose
(Cacciola et al., 2012), was harmless for P. concolor (Jacas and Viñuela., 1994a; Table
6.2.). Results from other studies indicated mancozeb as non-toxic to other parasitoids,
such as Anagyrus species near pseudococci (Girault) and Coccidoxenoides perminutus
(Timberlake) (Hymenoptera: Encyrtidae) (Mgocheki and Addison, 2016).
The consequences of the application of fungicides on parasitoids should be further
studied, not only the direct mortality but also the sublethal effects on P. concolor. It is
also important to consider that fungicide application can reduce diversity and richness of
nectar microorganisms (fungus and bacteria) in exposed flowers, which influences nectar
quality, sugar concentration or pH with possible consequences for the feeding of insects
(Schaeffer et al., 2017).
4.3.3. Herbicide exposure
From an integrated weed control strategy perspective, it is important to select
herbicides compatible with parasitoids. Jacas and Viñuela (1994) performed the only
study on the effects of herbicides on P. concolor adult females. As a result, four active
ingredients (haloxyfop, metamitron, ethofumesate and fluroxypyr: Group O; HRAC)
were tested: three were class 2 and one was class 1 according to the IOBC testing scheme.
Only fluroxypyr remain authorized (Table 6.2.).
In our tests, the survival of females and reproductive parameters (progeny size and
attracted hosts) was evaluated After treatment with glyphosate (Group G; HRAC).
Nowadays, glyphosate is one of the most widely used herbicides over the world, being a
non-selective and post-emergent herbicide that affects plant growth (De Aguiar et al.,
2016).
The survival of females was classified as slightly harmful and harmless for exposure
via residual contact and ingestion respectively (Tables 6.5. and 6.6.). Apparently,

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
152
glyphosate does not interfere with the parasitism of P. concolor (Table 6.7.). Research
seems to show that glyphosate cause genotoxicity effects in Drosophila melanogaster
(Kaya et al., 2000). Also, have been demonstrated that have negative effects on other
natural enemies. For example, longevity, fecundity and fertility of Chrysoperla externa
Hagen (Neuroptera: Chrysopidae) were deeply reduced when exposed to glyphosate
treatment. Furthermore, adults developed tumours in the abdomen region at 20 d after
emergence (Schneider et al., 2009).
In conclusion, although these herbicides did not prove to be very toxic for P. concolor,
further studies are needed to understand the persistence and flow pathway of herbicides
on olive groves and, consequently, the real implications on parasitoids and in the pest
management programs. In addition, as previously mentioned, the toxicity of some
herbicides can decrease pollen production and viability (Qian et al., 2015a) and make
natural foods often out of reach and/or indigestible. Besides that, the indiscriminate
application of herbicides reduced floral community (plant cover and diversity), removes
refuges and plants with alternative hosts and with implication for environment (soil,
water, etc.) (Le Bellec et al., 2015).
4.3.4. Microbial and botanical exposure
Microorganisms (bacteria, fungi, viruses and others) are widely used and are
considered to have a good potential as biopesticides in IPM. The bacterium Bacillus
thuringiensis (Bt) is probably the most studied and the most widely used and successful
microorganism ‘insecticide’. The crystal proteins of Bt produced during its sporulation
phase need to be ingested by pest species to be effective (Andrews, Cripps and Edwards,
2011). This biopesticide have a relatively broad spectrum of activity, being used for the
management of coleopterans (Bt tenebrinonis, Bt lentimorbus), dipterans (Bt israelensis)
and lepidopterans (Bt kurstaki, Bt aizawai) (Usta, 2013). In olive groves, some isolates
showed to be toxic only to adults or larvae when used against the olive fruit fly and to
both stages of the olive fruit fly (Alberola et al., 1999). For example, in an experiment
for three successive years, with four to six sprayings per year of Bt 114A isolate, the
longevity of olive fruit fly, the oviposition period, number of eggs and percent egg hatch
decreased (Navrozidis et al., 2000). In addition, results from other study indicate that only
3 out of 14 isolates that produced significant mortality on adults and/or larvae of B. oleae
were found to be toxic for P. concolor (Alberola et al., 1999). Other authors demonstrated
that Bacillus thuringiensis var. kurstaki appears to be safe for parasitoid P. concolor

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
153
(Jacas et al., 1992a; Jacas and Viñuela, 1994a) (Table 6.3.). Similar, results were found
for adults and pupae of the parasitoid Bracon nigricans (Hymenoptera: Braconidae)
(Biondi et al. 2013). On the other hand, other studies showed that B. thuringiensis affect
the longevity and the time spent on host eggs patches of Trichogramma chilonis
(Hymenoptera: Trichogrammatidae) (Amichot et al., 2015).
Fungal agent Beauveria bassiana is another biopesticide used in the control of the olive
fruit fly (Mahmoud, 2009). Oliveira et al. (2012) reported the first presence of B. bassiana
on Prays oleae and open new strategies for the biocontrol of this pest. The mechanism of
infection has been described as starting with the conidia attachment, followed by
germination and penetration of host cuticle using hydrolytic enzymes (e.g., proteases,
lipases, chitinases), mechanical pressure and other factors (Ortiz-Urquiza and Keyhani,
2013). Medina et al. (2008) found that B. bassiana did not cause any mortality on P.
concolor. However, progeny size diminished after a residual contact treatment or the
number of attacked hosts when B. bassiana was ingested (Table 6.3.). Also, Oreste et al.
(2015), showed that P. concolor emergence is affected by fungal treatments, mainly when
applied 2 days after parasitized puparia. There is some evidence that Beauveria bassiana
is a safe biocontrol agent for environment, human health and non-target organisms
(Zimmermann, 2007). Although some studies evidenced that some predators and
parasitoids were susceptible in laboratory, the impact under field conditions was
minimum (Jaronski et al., 1998).
The entomopathogenic nematodes (EPN) species, Heterorhabditis bacteriophora and
Steinernema carpocapsae are effective and safe biological control agents for several
insect pests, including olive fruit fly (Han and Ehlers, 2000; Sirjani, Lewis and Kaya,
2009). Sirjani, Lewis and Kaya (2009) was the only study that evaluated the susceptibility
of the third instar larvae of olive fruit fly to EPNs, including H. bacteriophora and S.
carpocapsae. Recently Torrini et al. (2017) demonstrated that H. bacteriophora and S.
carpocapsae caused mortality in olive fruit fly pupae and adults, proposing a new
perspective in the control olive fruit flies. However, these biopesticides are not entirely
safe for natural enemy P. concolor. Although it does not cause mortality on parasitoids,
it decreases the beneficial capacity by residual contact or via treated host larvae (lower
progeny size) or by ingestion (lower number of attacked hosts) (Medina et al. 2008)
(Table 6.3.).
For a safer use of microorganism-based bioinsecticides it is important more realistic
studies about efficacy on the pest as well as on side-effects on natural enemies, namely

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
154
for parasitoid P. concolor. Also, the method of application of biopesticides in the field
should be further studied. Additionally, landscape structure is crucial for survival of
arthropod community, constituted a reservoir of genetic diversity for microorganisms
with potential for biological control.
Plant extracts and oils might be also used as alternative to insecticides. Compounds of
seeds of Trichilia havanensis Jacq. (Meliaceae): F12 Limonoids (azadirone) and F18
(1.7+3.7-di-O-acetilhavanensin) and the Teucrium viscidum (Lamiaceae) extracts: M1
(Teucjaponic B) and M9 (Teucvin) did not cause mortality on P. concolor neither reduced
the parasitoid beneficial capacity in laboratory conditions (Zapata et al., 2004; Medina et
al., 2006) (see Table 6.3.). Similar results were obtained by Rehman et al. (2009) on
infested olive fruit with 2% Peganum harmala (Zygophllaceae) seed extract for P.
concolor adults’ mortality and parasitism. Furthermore, Melaleuca alternifolia
(Myrtaceae) essential oil was considered more toxic for C. capitata than for P. concolor,
being indicated as an alternative to broad-spectrum insecticides (Benelli et al., 2013).
Overall, microbial and botanical pesticides are harmless for parasitoid P. concolor and
should be used to increase the sustainability of agricultural systems. Other advantage of
these “products” is their environmental safety, target-specificity, efficacy and
biodegradability (Kumar and Singh, 2015).
5. EMERGENT PROBLEM: A RISK FOR PSYTTALIA CONCOLOR SURVIVAL
Xylella fastidiosa, a xylem-limited bacterium that causes Pierce’s disease (PD) in
many crop species, is considered an emergent infirmity in olive groves (Saponari et al.,
2013; 2017). This pathogen has an extensive natural host range, which includes many
herbaceous and woody plants, cultivated crops and weeds (Denancé et al., 2017).
Transmission by xylem sap-feeding insects (mainly sharpshooters and froghoppers or
spittlebugs (Cicadellidae)) favours the natural spread of the disease (Janse and Obradovic,
2010). Generally, the insecticidal control of vectors has been unsuccessful in halting
disease spread but can be effective against local vector (for example, imidacloprid are
particularly active against sharpshooters), dramatically reducing its population (Jackson,
2008; Cornara et al., 2016). According to Adán et al. (2011), imidacloprid [Sub-group
4A: Neonicotinoids] toxicity depends on the mode of exposure. Imidacloprid was
harmless for females of P. concolor in bait sprays application (on olive branches and
Cebo) but caused high mortality or sublethal effects on progeny when applied in cover

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
155
sprays and via treated host (Adán et al., 2007; 2011). Moreover, this insecticide interferes
with the transmission of stimuli in the insect’s nervous system by causing a blockage in
the nicotinergic neuronal pathway, leading to an impair of sexual communication of
parasitoids and host finding with consequences for biological control (Tappert et al.,
2017). Pointing this, the use of imidacloprid or other insecticides (pyrethroids) that can
be used to combat the vector, as well as the possible tendency of the farmer to remove the
vegetation with herbicides (in order to eliminate the host plant species), can compromise
the availability of food sources for P. concolor and consequently their survival.
6. OVERVIEW
After reviewing the literature available, it can be concluded that:
1) In general, fungicides, herbicides and biopesticides have less effects on mortality
and reproductive parameters than insecticides, although some fungicides are also very
harmful to P. concolor. Although there is a significant variability in response of P.
concolor to insecticides, there are many evidences that neurotoxic insecticides are more
harmful than other compounds as insect growth regulators. In fact, the most used
compounds against B. oleae, organophosphates and pyrethroids, are quite often very toxic
insecticides to adult’s females of P. concolor. Moreover, although not much studied, it
has been reported that neurotoxic insecticides affect progeny sex-ratio and causes a
significant increase in the proportion of males in most braconid species. As described by
Biondi et al., (2013), since braconids have arrhenotokous parthenogenesis (i.e.,
unfertilized females produce only males) possibly the higher male progeny may be due
to male sterility or mating behavior alterations after wasps are exposed to neurotoxic
insecticides.
2) Despite the impact of fungicides on P. concolor is lesser than insecticides, they
should also be studied because they are not negligible when applied by foliar application
on the olive canopy. There are no data on the joint use of fungicides+insecticides
mixtures,
3) Most of the studies have been conducted in laboratory (under “standard
conditions”), with different uptake routes (residual, ingestion…) and focused only on
mortality after 3 days (lethal effects) and reproduction (attacked hosts and progeny size)
(sublethal effects). Other sublethal effects should be considered as well such as longevity,

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
156
learning performance, behavior, neurophysiology, physiological and immunology
(Desneux, Decourtye and Delpuech, 2007; Talebi, Kavousi and Sabahi, 2008).
4) Most of the laboratory studies only consider the effect of a single pesticide but not
the synergistic effects of various pesticides that can be applied at the same time in the
field.
5) Persistence of plant production products has not been explored with few
exceptions.
6) Results from laboratory studies should be supplemented with supportive data from
field studies and there are no references about this in the literature.
This ecotoxicological studies associated to the risks of pesticides application help to
promote changes in application patterns (e.g., the switch from a broad-spectrum to a
selective or optimum dose-rates) (Amano and Haseeb, 2001). Moreover, these studies
will allow the optimization of the release timing of this parasitoid after pesticide
applications (when levels maximize survival of parasitoid) and it will possible lead to
sustainable chemical control approaches, establishing the use of pesticides less toxic for
P. concolor as a pre-requisite for the control of B. oleae.
7. REFERENCES
Abdallahi, E., Adán, A. and Viñuela, E. (2000). Estudio de la actividad del Piriproxifen
y fipronil sobre Opius concolor Szèpligeti (Hymenoptera: Braconidae) y su huésped
de substitución Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Boletín de
sanidad vegetal Plagas, 26 (4): 503-512.
Adán, A., González, T., Bastante, R., Budia, F., Medina, P., Del Estal, P. and Viñuela, E.
(2007). Efectos de diversos insecticidas aplicados en condiciones de laboratorio
extendido sobre Psyttalia concolor Szèpligeti (Hymenoptera: Braconidae). Boletín de
sanidad vegetal Plagas, 33: 391-397.
Adán A., Viñuela, E., Bengochea, P., Budia, F., Del Estal, P. Aguado, P. and Medina, P.
(2011). Lethal and sublethal toxicity of fipronil and imidacloprid on Psyttalia concolor
(Hymenoptera: Braconidae). Journal of Economic Entomology, 104(5): 1541-1549.
Alberola, T.M., Aptosoglou, S., Arsenakis, M., Bel, Y., Delrio, G., Ellar, D.J. Ferré J.,
Granero, F., Guttmann, D.M., Koliais, S., Martínez-Sebastián, M. J., Prota, R., Rubino,
S., Satta, A., Scarpellini, G., Sivropoulou, A. and Vasara, E. (1999). Insecticidal
Activity of Strains of Bacillus thuringiensis on Larvae and Adults of Bactrocera oleae
Gmelin (Dipt. Tephritidae). Journal of Invertebrate Pathology, 74(2): 127-136.
Amano, H. and Haseeb, M. (2001). Recently-proposed methods and concepts of testing
the effects of pesticides on the beneficial mite and insect species: study limitations and
implications in IPM. Applied Entomology and Zoology, 36 (1): 1-11.
Amichot, M., Curty, C., Benguettat-Magliano, O., Gallet, A. and Wajnberg, E. (2015).
Side effects of Bacillus thuringiensis var. kurstaki on the hymenopterous parasitic
wasp Trichogramma chilonis. Environmental Science and Pollution Research, 23(4):
3097-3103.

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
157
Andrews, M., Cripps, M.G. and Edwards, G.R. (2011). The potential of beneficial
microorganisms in agricultural Systems. Annals of Applied Biology, 160: 1-5.
Arzone, A. Dolci, M., Marletto, F. and Minero, C. (1995). Translocation of Fenoxycarb
in the Agro-ecosystem, Bioscience, Biotechnology, and Biochemistry, 59(7): 1318-
1319.
Bayram, A., Salerno, G., Onofri, A. and Conti, E. (2010). Sub-lethal effects of two
pyrethroids on biological parameters and behavioural responses to host cues in the egg
parasitoid Telenomus busseolae. Biological Control, 53:153-160.
Benelli, G., Canale, A., Flamini, G., Cioni, P. L., Demi, F., Ceccarini, L., Macchia,
M. and Conti, B. (2013). Biotoxicity of Melaleuca alternifolia (Myrtaceae) essential
oil against the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae), and
its parasitoid Psyttalia concolor (Hymenoptera: Braconidae). Industrial Crops and
Products, 50: 596-603.
Benelli, G., Giunti, G., Tena, A., Desneux, N., Caselli, A. and Canale A. (2017). The
impact of adult diet on parasitoid reproductive performance. Journal of Pest Science,
90: 807-823.
Bengochea, P., Amor, F., Saelices, R., Hernando, S., Budia, F., Adán, A. and Medina, P.
(2013). Kaolin and copper-based products applications: Ecotoxicology on four natural
enemies. Chemosphere, 91(8): 1189-1195.
Bengochea, P., Budia, F., Viñuela, E. and Medina, P. (2014). Are kaolin and copper
treatments safe to the olive fruit fly parasitoid Psyttalia concolor? Journal of Pest
Science, 87(2): 351-359.
Bengochea, P., Christiaens, O., Amor, F., Viñuela, E., Rouge, P., Medina, P. and
Smagghe, G. (2012). Insect growth regulators as potencial insecticidas to control olive
fruit fly (Bactrocera oleae Rossi): insect toxicity bioassays and molecular docking
approach. Pest Management Science, 69: 1-34.
Bengochea, P., Hernando, S., Saelices, R., Adán, A., Budia, F., González-Núñez, M.,
Viñuela, E. and Medina, P. (2010). Side effects of kaolin on natural enemies found on
olive crops. Pesticides and Beneficial Organisms. IOBC/wprs Bulletin, 55: 61-67.
Benhadi-Marín, J., Pereira, J.A. and Santos, S.A.P. (2016). Effects of kaolin particle films
on the life span of an orb-weaver spider. Chemosphere, 144: 918-924.
Biondi, A., Mommaerts, V., Smagghe, G., Viñuela, E., Zappalà, L. and Desneux, N.
(2012). The non-target impact of spinosyns on beneficial arthropods. Pest
Management Science, 68(12): 1523-1536.
Biondi, A., Zappalà, L., Stark, J.D. and Desneux, N. (2013). Do biopesticides affect the
demographic traits of a parasitoid wasp and its biocontrol services through sublethal
effects? Plos One, 8(9): e76548.
Bora, D., Khanikor B. and Gogoi, H. (2012). Plant Based Pesticides: Green
Environment with Special Reference to Silk Worms, Pesticides, R.P. Soundararajan,
IntechOpen, DOI: 10.5772/47832. Available from:
https://www.intechopen.com/books/pesticides-advances-in-chemical-and-botanical-
pesticides/plant-based-pesticides-green-environment-with-special-reference-to-silk-
worms.
Botías, C., David, A., Hill, E.M. and Goulson, D. (2017). Contamination of wild plants
near neonicotinoid seed-treated crops, and implications for non-target insects. Science
of the Total Environment, 566-567: 269-278.
Cacciola, S.O., Faedda, R., Sinatra, F., Agosteo, G.E., Schena, L., Frisullo, S. and
Magnano di San Lio, G. (2012). Olive Anthracnose. Journal of Plant Pathology, 94
(1): 29-44.

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
158
Casida, J.E. and Quistad, G.B. (1998). Golden Age of Insecticide Research: Past, Present,
or Future? Annual Review of Entomology, 43: 1-16.
Cloyd, R. (2005). Compatibility conflict: is the use of biological control agents with
pesiticides a viable management strategy? Second International Symposium on
Biological Control of Arthropods, 546-554.
Cloyd, R. (2012). Indirect Effects of Pesticides on Natural Enemies, Pesticides R.P.
Soundararajan, IntechOpen, DOI: 10.5772/48649. Available from:
https://www.intechopen.com/books/pesticides-advances-in-chemical-and-botanical-
pesticides/indirect-effects-of-pesticides-on-natural-enemies.
Cooper, J. and Dobson H. (2007). The benefits of pesticides to mankind and the
environment. Crop Protection, 26: 1337-1348.
Cornara, D., Cavalieri, V., Dongiovanni, C., Altamura, G., Palmisano, F., Bosco, D.,
Porcelli, F., Almeida, R.P.P. and Saponari, M. (2016). Transmission of Xylella
fastidiosa by naturally infected Philaenus spumarius (Hemiptera, Aphrophoridae) to
different host plants. Journal of Applied Entomology, 141 (1-2): 80-87.
Cowles R.S. and Eitzer, B.D. (2017). Residues of neonicotinoid insecticides in pollen and
nectar from model plants. Journal of Environmental Horticulture, 35(1):24-34.
D'Ascenzo, D., Crivelli, L. and Camillo, L. di (2014). Nuove strategie di difesa nei
confronti del cicloconio dell'olivo in Italia centrale. Atti, Giornate Fitopatologiche,
Chianciano Terme (Siena), 2: 139-146
D’Ávila, V.A., Barbosa, W.F., Guedes, R.N.C. and Cutler, G.C. (2018). Effects of
Spinosad, Imidacloprid, and Lambda-cyhalothrin on Survival, Parasitism, and
Reproduction of the Aphid Parasitoid Aphidius colemani. Journal of Economic
Entomology, 111(3): 1096-1103.
Daane K.M. and Johnson, M.W. (2010). Olive fruit fly: managing an ancient pest in
modern times. Annual Review of Entomology, 55:151-169.
Damalas, C.A (2009). Understanding benefits and risks of pesticide use. Scientific
Research and Essay, 4 (10): 945-949.
De Aguiar, L.M., Figueira, F.H., Gottschalk, M. S. and da Rosa, C.E. (2016). Glyphosate-
based herbicide exposure causes antioxidant defence responses in the fruit fly
Drosophila melanogaster. Comparative Biochemistry and Physiology Part C:
Toxicology & Pharmacology, 185-186, 94-101.
De la Roca, M. (2003). Surround® Crop Protectant: La capa protectora natural para
cultivos como el olivar. Phytoma España, 148: 82-85.
Denancé, N., Legendre, B., Briand, M., Olivier, V., de Boisseson, C., Poliakoff, F. and
Jacques, M.-A. (2017). Several subspecies and sequence types are associated with the
emergence of Xylella fastidiosa in natural settings in France. Plant Pathology, 66(7):
1054-1064.
Denholm, I.; Pickett, J.A. and Devonshire, A.L. (2001). Insecticide Resistance: From
Mechanisms to Management, CABI and The Royal Society publishing, ISBN 0-
85199-367-2, UK.
Desneux, N., Decourtye, A. and Delpuech J-M. (2007). The Sublethal Effects of
Pesticides on Beneficial Arthropods. Annual Review of Entomology, 52: 81-106.
Desneux, N., Denoyelle, R. and Kaiser, L. (2006). A multi-step bioassay to assess the
effect of the deltamethrin on the parasitic wasp Aphidius ervi. Chemosphere, 65: 1697-
1706.
Desneux, N., Pham-Delègue, M.H. and Kaiser, L. (2004). Effect of sub-lethal and lethal
dose of lambda-cyhalothrin on oviposition experience and host searching behaviour of
a parasitic wasp, Aphidius ervi. Pest Management Science, 60: 381-389.

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
159
Desneux, N., Rafalimanana, H. and Kaiser, L. (2004). Dose–response relationship in
lethal and behavioural effects of different insecticides on the parasitic wasp Aphidius
ervi. Chemosphere, 54: 619-627.
DGAV (2018). Direção-Geral de Alimentação e Veterinária. Official phytosanitary
products.
http://www.dgav.pt/fitofarmaceuticos/guia/finalidades_guia/Insec&Fung/Culturas/oli
veira.htm and http://www.drapc.minagricultura.pt/drapc/servicos/
fitossanidade/files/herbicidas_olival_2017.pdf. Cited 10 September 2018 (In
Portugal).
Dugravot, S., Sanon, A., Thibout, E. and Huignard, J. (2002). Susceptibility of
Callosobruchus maculatus (Coleoptera: Bruchidae) and its Parasitoid Dinarmus
basalis (Hymenoptera: Pteromalidae) to Sulphur-Containing Compounds:
Consequences on Biological Control. Environmental Entomology, 31(3): 550-557.
El-Wakeil, N.E. (2013). Botanical Pesticides and Their Mode of Action. Gesunde
Pflanzen, 65:125-149.
Ergin, E., Er, A., Uçkan, F. and Rivers, D.B. (2007). Effect of Cypermethrin Exposed
Hosts on Egg-Adult Development Time, Number of Offspring, Sex Ratio, Longevity,
and Size of Apanteles galleriae Wilkinson (Hymenoptera: Braconidae). Belgian
Journal of Zoology, 137 (1): 27-31.
Fernandez, M., Medina, P., Wanumen, A.C., Del Estal, P., Smagghe, G. and Viñuela, E.
(2017). Compatibility of sulfoxaflor and other modern pesticides with adults of the
predatory mite Amblyseius swirskii. Residual contact and persistence studies.
Biocontrol, 62: 197-208.
Fiedler, A.K., Landis, D.A. and Wratten, S.D. (2008). Maximizing ecosystem services
from conservation biological control: the role of habitat management. Biological
Control, 45: 254-271.
Garantonakis, N., Varikou, K. and Birouraki, A. (2016). Comparative selectivity of
pesticides used in greenhouses, on the aphid parasitoids Aphidius colemani
(Hymenoptera: Braconidae). Biocontrol Science and Technology, 26(5): 678-690.
Garzón, A., Medina, P., Amor, F., Viñuela, E. and Budia, E. (2015). Toxicity and
sublethal effects of six insecticides to last instar larvae and adults of the biocontrol
agents Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae) and Adalia
bipunctata (L.) (Coleoptera: Coccinellidae). Chemosphere, 132: 87-93.
Geiger et al., (2010). Persistent negative effects of pesticides on biodiversity and
biological control potential on European farmland. Basic and Applied Ecology, 11:97-
105.
Genduso, P. (1970). Influenza della temperatura sulla durata e sulla resistenza degli stadi
preimmaginali dell'Opius c. siculus Mon. [Influence of temperature on the duration
and strength of the stadium preimaginal’ Opius c. siculus Mon.]. Bolletino
dell’Instituto di Entomologia Agraria e dell’ Observatorio di Fitopatologia di
Palermo & Bulletin of the Institute of Agricultural Entomology and Observatorio
Phytopathology Palermo, 8: 1-7.
Gentz, M., Murdoch, G. and King, G.F. (2010). Tandem use of selective insecticides and
natural enemies for effective, reduced-risk pest management. Biological Control, 52:
208-215.
Gillespie, M.A.K., Gurr, G.M. and Wratten, S.D. (2016). Beyond nectar provision: the
other resource requirements of parasitoid biological control agentes. Entomologia
Experimentalis et Applicata, 159: 207-221.
Glenn, M. and Puterka, G.J. (2005). Particle films: a new technology for agriculture.
Horticultural Reviews, 31: 1-44.

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
160
Gonçalves, M.F., Malheiro, R., Casal, S., Torres, L. and Pereira, J.A. (2012). Influence
of fruit traits on oviposition preference of the olive fly, Bactrocera oleae (Rossi)
(Diptera: Tephritidae), on three Portuguese olive varieties (Cobrançosa, Madural and
Verdeal Transmontana). Scientia Horticulturae, 145: 127-135.
González, M., Bahena, F. and Viñuela, E. (1998). Efectos de distintos Reguladores del
Crecimiento de Insectos (RCI) sobre el parasitoide Opius concolor Szèpligeti cuando
son ingeridos por la larva huésped. Boletín de sanidad vegetal Plagas, 24: 193-200.
Grenier S. and Grenier, A.-M., (1993). Fenoxycarb, a fairly new Insect Growth Regulator:
a review of its effects on insects. Annals Applied Biology, 122: 369-403.
Han, R. and Ehlers, R.-U. (2000). Pathogenicity, Development, and Reproduction of
Heterorhabditis bacteriophora and Steinernema carpocapsae under Axenic in Vivo
Conditions. Journal of Invertebrate Pathology: 75(1), 55-58.
Harbi, A., Abbes, K., Sabater-Muñoz, B., Beiti, F. and Chermiti, B. (2017). Residual
toxicity of insecticides used in Tunisian citrus orchards on the imported parasitoid
Diachasmimorpha longicaudata (Hymenoptera: Braconidae): Implications for IPM
program of Ceratitis capitata (Diptera: Tephritidae). Spanish Journal of Agricultural
Research, 15 (3): e1008.
Hassan, S.A. (1988). Guidelines for testing the effects of pesticides on beneficials
IOBC/WPRS Bulletin, XI(4), 143 pp.
Hassan S.A. (1998). The initiative of the IOBC/WPRS Working Group on Pesticides
and Beneficial Organisms. In: Haskell P.T., McEwen P. (eds), Ecotoxicology.
Springer, Boston, MA.
Hassan, S.A. (1992). Guidelines for testing the effect of pesticides on beneficial
organisms. IOBC/WPRS Bulletin XV (3), 186 pp.
Hassan, S.A. et al. (1988), Results of the fourth joint pesticide testing programme carried
out by the IOBCIWPRS-Working Group “Pesticides and Beneficial Organisms”.
Journal of Applied Entomology, 105: 321-329.
Hassan, S.A., et al. (1985). Standard methods to test the side-effects of pesticides on
natural enemies of insects and mites developed by the IOBC/WPRS Working Group
“Pesticides and Beneficial Organisms.” EPPO Bulletin, 15(2), 214-255.
Hepdurgun, B. Turanli T. and Zümreoğlu A. (2009). Control of the olive fruit fly,
Bactrocera oleae, (Diptera: Tephritidae) through mass trapping and mass releases of
the parasitoid Psyttalia concolor (Hymenoptera: Braconidae) reared on irradiated
Mediterranean fruit fly, Biocontrol Science and Technology, 19(1): 211-224.
Hervé, M. R., Delourme, R., Gravot, A., Marnet, N., Berardocco, S. and Cortesero, A.M.
(2014). Manipulating Feeding Stimulation to Protect Crops Against Insect Pests?
Journal of Chemical Ecology, 40(11-12): 1220-1231.
Hladnik, M.A (2017). Review of plant protection against the olive fly (Bactrocera
oleae (Rossi, 1790) Gmelin) and molecular methods to monitor the insecticide
resistance alleles. Acta Agriculturae Slovenica, 109 (1): 135-146.
Hoddle, M. S., Van Driesche, R.G., Lyon S.M. and Sanderson. J.P. (2001). Compatibility
of insect growth regulators with Eretmocerus eremicus for whitefly control on
poinsettias: I. Laboratory Assays. Biological Control, 20: 122-131.
Iannotta, N., Monardo, D. and Perri, L. (2002). Effects of different treatments against
Spilocaea oleagina (cast.) hugh. Acta Horticulturae, (586): 741-744.
Jacas, J.A. and Viñuela, E. (1994a). Side-effects of pesticides on Opius concolor Szèp.
(Hymenoptera:Braconidae), a parasitoid of the olive fruit fly. Bull. OILB/IOBC Bull.,
17(10): 143-146.
Jacas, J. A. and Viñuela, E. (1994b). Analysis of a laboratory method to test the effects
of pesticides on adult females of Opius concolor (Hym., Braconidae), a parasitoid of

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
161
the olive fruit fly, Bactrocera oleae (Dip., Tephritidae). Biocontrol Science and
Technology, 4: 147-154.
Jacas, J., Viñuela, E., Adán, A., Budia, F., Del Estal, P. and Marco, V. (1992b) Efectos
secundarios de los plaguicidas utilizados en el olivar español sobre adultos de Opius
concolor, Szèp. (Hymenoptera:Braconidae), parasitoide de la mosca de la aceituna
Bactrocera oleae (Gmel.) (Dip. Tephritidae). Boletín de sanidad vegetal Plagas, 18:
315-321.
Jacas, J., Viñuela, E., Adán, A., Budia, F., Del Estal, P. and Marco, V. (1992a).
Laboratory evaluation of selected pesticides against Opius concolor Szèpl.
(Hymenoptera:Braconidae). Annals of Applied Biology, 13(120): 140-141.
Jackson, R.S. (2008). Vineyard Practice, in: Wine Science (Third Edition), Academic
Press, San Diego, 108-238 pp.
Janse J.D. and Obradovic, A. (2010). Xylella fastidiosa: its biology, diagnosis, control
and risks. Journal of plant pathology, 92 (1) supplement), S1.35-S1.48.
Jaronski, S.T., Lord, J., Rosinska, J., Bradley, C., Hoelmer, K., Simmons, G., Osterlind,
R., Brown, C., Staten, R. and Antilla, L. (1998) Effect of Beauveria bassiana-based
mycoinsecticide on beneficial insects under field conditions. The 1998 Brighton
Conference - pests and diseases, 651-656.
Jepsen, S.J., Rosenheim, J.A. and Matthews, C.E. (2007). The impact of sulfur on the
reproductive success of Anagrus spp. parasitoids in the field. BioControl, 52: 599-612.
Jervis, M.A. and Kidd, N.A.C. (1996). Host feeding strategies in hymenopteran
parasitoids. Biology Reviews, 61: 395-434.
Kakani, E.G., Zygouridis, N.E., Tsoumani, K.T., Seraphides, N., Zalom, F.G. and
Mathiopoulos, K.D. (2010). Spinosad resistance development in wild olive fruit fly
Bactrocera oleae (Diptera: Tephritidae) populations in California. Pest Management
Science, 66: 447-453.
Kampouraki, A. et al. (2018). Recent evolution and operational impact of insecticide
resistance in olive fruit fly Bactrocera oleae populations from Greece. Journal of Pest
Science, 91(4): 1429-1439.
Kaya, B., Creus, A., Yanikoğlu, A., Cabré, O. and Marcos, R. (2000). Use of the
Drosophila wing spot test in the genotoxicity testing of different herbicides.
Environmental and Molecular Mutagenesis, 36: 40-46.
Kenis, M., Hurley, B.P., Hajek, A.E. and Cock, M.J.W. (2017). Classical biological
control of insect pests of trees: facts and figures. Biological Invasions, 19: 3401-3417.
Kirst, A.H. (2010). The spinosyn family of insecticides: realizing the potential of natural
products research. The Journal of Antibiotics, 63: 101-111.
Krischik, V.A., Landmark, A.L. and Heimpel, G.E. (2007). Soil-Applied Imidacloprid Is
Translocated to Nectar and Kills Nectar-Feeding Anagyrus pseudococci (Girault)
(Hymenoptera: Encyrtidae). Environmental Entomology, 36(5): 1238-1245.
Kumar, S. and Singh, A. (2015). Biopesticides: Present Status and the Future Prospects.
Journal of Fertilizers & Pesticides, 6(2): 1000e129.
Le Bellec, F., Vélua, A., Fournier, P., Le Squinc, S., Michels, T., Tendero, A. and
Bockstaller, C. (2015). Helping farmers to reduce herbicide environmental impacts.
Ecological Indicators, 54: 207-216.
Longley M. (1999). A review of pesticide effects upon immature aphid parasitoids within
mummified hosts. International Journal of Pest Management, 45:(2)139-145.
Longley, M. and Jepson, P.C. (1996). Effects of honeydew and insecticide residues on
the distribution of foraging aphid parasitoids under glasshouse and field conditions.
Entomologta Experimentahs et Applicata, 81:189-198.

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
162
Macfadyen, S., Banks, J.E., Stark, J.D. and Davies, A.P. (2014). Using Semifield Studies
to Examine the Effects of Pesticides on Mobile Terrestrial Invertebrates. Annual
Review of Entomology, 59: 383-404.
Mahmoud, M.F. (2009). Pathogenicity of Three Commercial Products of
Entomopathogenic Fungi, Beauveria bassiana, Metarhizum anisopilae and
Lecanicillium lecanii against Adults of Olive Fly, Bactrocera oleae (Gmelin) (Diptera:
Tephritidae) in the Laboratory. Plant Protection Science, 45(3): 98-102.
Malheiro, R. Casal, S., Baptista, P. and Pereira, J.A. (2015). Review of Bactrocera oleae
(Rossi) impact in olive products: From the tree to the table. Trends in Food Science &
Technology, 44: 226-242.
MAPAMA, 2018. Ministry of Agriculture and Fisheries Food and Environment. Official
phytosanitary products entry.
http://www.mapama.gob.es/es/agricultura/temas/sanidad-vegetal/productos-
fitosanitarios/registro/menu.asp Cited 25 May 2017 (In Spanish).
Margaritopoulos, J.T., Skavdis, G., Kalogiannis, N., Nikou, D., Morou, E., Skouras, P.J.,
Tsitsipis, J.A. and Vontas, J. (2008). Efficacy of the pyrethroid alpha-cypermethrin
against Bactrocera oleae populations from Greece and improved diagnostic for an
iAChE mutation. Pest Management Science, 64: 900-908.
Medina, P., Budia, F., González, M., Rodríguez, B., Díaz, A., Huerta, A., Zapata, N. and
Viñuela, E. (2006). Effects of botanical insecticides on two natural enemies of
importance in Spain: Chrysoperla carnea (Stephens) and Psyttalia concolor
(Szépligeti). Pesticides and Beneficial Organisms. IOBC/wprs Bulletin, 29(10): 85-93.
Medina, P., Corrales, E., González-Nuñez, M., Smagghe, G. and Viñuela, E. (2008).
Effects of Beauveria bassiana, Heterorhabditis bacteriophora, H. megidis and
Steinernema feltiae on the Mediterranean fruit fly Ceratitis capitata and the very
sensitive braconid Psyttalia concolor in the lab. Pesticides and Beneficial Organisms.
IOBC/wprs Bulletin, 35: 113-121.
Medina, P., Morales, J.J., Budia, F., Adán, A., Del Estal, P. and Viñuela, E. (2007).
Compatibility of the endoparasitoid Hyposoter didymator (Thunberg) (Hymenoptera:
Ichneumonidae) protected stages with five selected insecticidas. Journal of Economic
Entomology, 100(6): 1789-1796.
Mgocheki, N. and Addison, P. (2016). Effect of contact pesticides on vine mealybug
parasitoids, Anagyrus sp. near pseudococci (Girault) and Coccidoxenoides perminutus
(Timberlake) (Hymenoptera: Encyrtidae). South African Journal of Enology and
Viticulture, 30(2): 110-116.
Michel, M. Krause, A. and Buszewski, B. (2001). Column switching and liquid
chromatographic technique for the rapid determination of fenoxycarb insecticide
residues in apples. Polish Journal of Environmental Studies, 10 (4). 283-287.
Miranda, M.A., Miquel, M., Terrassa, J., Melis, N. and Monerris, M. (2008). Parasitism
of Bactrocera oleae (Diptera; Tephritidae) by Psyttalia concolor (Hymenoptera;
Braconidae) in the Balearic Islands (Spain). Journal of Applied Entomology,132: 798-
805.
Morais, M.R., Zanardi, O.Z., Rugno, G.R. and Yamamoto, P.T. (2016). Impact of five
insecticides used to control citrus pests on the parasitoid Ageniaspis citricola
Longvinovskaya (Hymenoptera: Encyrtidae). Ecotoxicology, 25: 1011-1020.
Navrozidis, E.I., Vasara, E., Karamanlidou, G., Salpiggidis, G.K. and Koliais, S.I. (2000).
Biological Control of Bactocera oleae (Diptera: Tephritidae) Using a Greek Bacillus
thuringiensis Isolate. Journal of Economic Entomology, 93(6): 1657-1661.
Nigro, F., Antelmi, I., Labarile, R., Sion, V. and Pentimone, I. (2018). Biological control
of olive anthracnose. Acta Horticulturae, 1199: 439-444.

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
163
Obanor, F.O., Jaspers, M.V., Jones, E. E. and Walter, M. (2008) Greenhouse and field
evaluation of fungicides for control of olive leaf spot in New Zealand. Crop Protection,
27: 1335-1342.
Oerke, E.-C. (2006). Crop losses to pests. Journal of Agricultural Science, 144: 31-43.
Ohta, I. and Takeda, M. (2015). Acute toxicities of 42 pesticides used for green peppers
to an aphid parasitoid, Aphidius gifuensis (Hymenoptera: Braconidae), in adult and
mummy stages. Applied Entomology and Zoology, 50: 207-212.
OJEU (Oficial Journal of the European Union) (2009). Directive 2009/128/EC of the
European Parliament and of the Council of 21 October 2009, establishing a framework
for Community action to achive the sustainable use of pesticides OJEU 209: 71-86.
Oliveira, I., Pereira, J. A., Lino-Neto, T., Bento, A. and Baptista, P. (2012). Fungal
Diversity Associated to the Olive Moth, Prays Oleae Bernard: A Survey for Potential
Entomopathogenic Fungi. Microbial Ecology, 63(4): 964-974.
Oreste, M., Baser, N., Bubici, G., Tarasco, E. (2015). Effect of Beauveria bassiana strains
on the Ceratitis capitata - Psyttalia concolor system. Bulletin of Insectology, 68 (2):
265-272.
Ortiz-Urquiza, A. and Keyhani, N. (2013). Action on the Surface: Entomopathogenic
Fungi versus the Insect Cuticle. Insects: 4(3): 357-374.
Pascual, S., Cobos, G., Seris, E. and González-Núñez, M. (2010). Effects of processed
kaolin on pests and non-target arthropods in a Spanish olive grove. Journal
of Pest Science, 83: 121-133
Pascual, S., Cobos, G., Seris E., Sánchez-Ramos, I. and González-Núñez, M. (2014).
Spinosad bait sprays against the olive fruit fly (Bactrocera oleae (Rossi)): effect on
the canopy non-target arthropod fauna, International. Journal of Pest Management,
60(4): 258-268.
Pavlidi, N., Kampouraki, A., Tseliou, V., Wybouw, N., Dermauw, W., Roditakis, E.,
Nauen, R. Van Leeuwen, T., Vontas, J. (2018). Molecular characterization of
pyrethroid resistance in the olive fruit fly Bactrocera oleae. Pesticide Biochemistry
and Physiology, 148: 1-7.
Penca, C. and Hodges, A.C. (2017). Pyriproxyfen treatment terminates Halyomorpha
halys reproductive diapause, with an indirect mortality effect on its egg parasitoid
Trissolcus japonicus. Journal of Pest Science,90: 1269-1276.
Pinheiro, L.A., Medina, P., Bento, A., Pereira, J.A and Santos, S.A.P. 2017. Bioecology
of Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) and Management
Strategies to Increase Biological Control of the Olive Fruit Fly. In Natural Enemies:
Identification, Protection Strategies and Ecological Impacts, ed. Santos, S.A.P. 141-
172. New York: Nova Science Publishers.
Prabhaker, N. Morse, J.G, Castle, S.J., Naranjo, S.E., Henneberry, T. J. and Toscano,
N.C. (2007). Journal of Economic Entomology, 100(4): 1053-1061.
Qian, H., Li, Y., Sun, C., Lavoie, M., Xie, J., Bai, X. and Fu, Z. (2015a.) Trace
concentrations of imazethapyr (IM) affect floral organs development and reproduction
in Arabidopsis thaliana: IM-induced inhibition of key genes regulating anther and
pollen biosynthesis. Ecotoxicology, 24: 163-171.
Qian, H., Lu, H., Ding, H., Lavoie, M., Li, Y., Liu, W. and Fu, Z. (2015b). Analyzing
Arabidopsis thaliana root proteome provides insights into the molecular bases of
enantioselective imazethapyr toxicity. Scientific Reports, 5: 11975.
Rehman, J. ur, Wang, X., Johnson, M.W., Daane, K.M., Jilani, G., Khan, M.A. and
Zalom, F. G. (2009). Effects of Peganum harmala (Zygophyllaceae) Seed Extract on
the Olive Fruit Fly (Diptera: Tephritidae) and Its Larval Parasitoid Psyttalia concolor
(Hymenoptera: Braconidae). Journal of Economic Entomology, 102(6): 2233-2240.

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
164
Romero, J., Agustí-Brisach, C., Roca, L.F., Moral, J., Gonzalez-Dominguez, E., Rossi,
V., and Trapero, A. (2018). A long-term study on the effect of agroclimatic variables
on olive scab in Spain. Crop Protection, 114: 39-43.
Ruiz-Torres M. and Montiel-Bueno A. (2007). Efecto de los tratamientos-cebo aereos
con spinosad contra mosca del olivo (Bactrocera oleae, Gmel.; Diptera: Tephritidae)
sobre la entomofauna del olivar en la provincia de Jaén. Boletín de
sanidad vegetal. Plagas, 33: 267-284.
Sánchez-Ramos, I., Fernández, C.E., González-Núñez, M. and Pascual, S. (2013).
Laboratory tests of insect growth regulators as bait sprays for the control of the olive
fruit fly, Bactrocera oleae (Diptera: Tephritidae). Pest Management Science, 69: 520-
526.
Santos, S.A.P., Pereira, J.A., Torres, L.M., Nogueira, A.J.A (2007). Evaluation of the
effects, on canopy arthropods, of two agricultural management systems to control pests
in olive groves from north-east of Portugal. Chemosphere, 67: 31-139.
Saour, G. and Makee, H.A. (2004). Kaolin-based particle film for suppression of the olive
fruit fly Bactrocera oleae Gmelin (Dip., Tephritidae) in olive groves. Journal of
Applied Entomology, 128: 28-31.
Saponari, M., Boscia, D., Altamura, G., Loconsole, G., Zicca, S., D’Attoma, G., Morelli,
M., Palmisano, F., Saponari, A., Tavano, D., Savino, V.N., Dongiovanni, C. and
Martelli, G.P. (2017). Isolation and pathogenicity of Xylella fastidiosa associated to
the olive quick decline syndrome in southern Italy. Scientific Reports, 7:17723.
Saponari, M., Boscia, D., Nigro, F. and Martelli, G.P. (2013). Identification of DNA
sequences related to Xylella fastidiosa in oleander, almond and olive trees exhibiting
leaf scorch symptoms in Apulia (southern Italy). Journal of Plant Pathology, 95(3):
668.
Schaeffer, R.N., Vannette, R.L., Brittain, C., Williams, N.M. and Fukami, T. (2017).
Non-target effects of fungicides on nectar-inhabiting fungi of almond flowers.
Environmental Microbiology Reports, 9(2): 79-84.
Schneider, M.I., Sanchez, N., Pineda S., Chi H. and Ronco, A. (2009). Impact of
glyphosate on the development, fertility and demography of Chrysoperla externa
(Neuroptera: Chrysopidae): Ecological approach. Chemosphere, 76: 1451-1455.
Schoonhoven, L.M., Van Loon, J.J.A. and Dicke, M. (2005). Insect-Plant Biology. 2nd
ed. Oxford University Press. 421 pp.
Serra, A.A., Couée, I., Renault, D., Gouesbet, G. and Sulmon, C. (2015a). Metabolic
profiling of Lolium perenne shows functional integration of metabolic responses to
diverse subtoxic conditions of chemical stress. Journal of Experimental Botany, 66:
1801-1816.
Serra, A.A., Couée, I., Heijnen, D., Michon-Coudouel, S., Sulmon, C. and Gouesbet, G.
(2015b). Genome-wide transcriptional profiling and metabolic analysis uncover
multiple molecular responses of the grass species Lolium perenne under low-intensity
xenobiotic stress. Frontiers in Plant Science, 6: 1124.
Sirjani, F.O., Lewis, E.E. and Kaya, H.K. (2009). Evaluation of entomopathogenic
nematodes against the olive fruit fly, Bactrocera oleae (Diptera: Tephritidae).
Biological Control, 48(3): 274-280.
Skouras, P.J., Margaritopoulos, J.T., Seraphides, N.A., Ioannides, I. M., Kakani, E.G.,
Mathiopoulos, K.D. and Tsitsipis, J.A. (2006). Organophosphate resistance in olive
fruit fly, Bactrocera oleae, populations in Greece and Cyprus. Pest Management
Science, 63(1): 42-48.
Soderlund, D.M. and Bloomquist, J.R., (1989). Neurotoxic actions of pyrethroid
insecticides. Annul Review of Entomology, 34: 77-96.

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
165
SPSS Inc., IBM Company (2016) IBM, SPSS statistic for windows, version 24.0. New
York, USA.
Stapel, J.O., Cortesero, A.M. and Lewis, W.J. (2000). Disruptive Sublethal Effects of
Insecticides on Biological Control: Altered Foraging Ability and Life Span of a
Parasitoid after Feeding on Extrafloral Nectar of Cotton Treated with Systemic
Insecticides. Biological Control, 17: 243-249.
Stark, J.D., Vargas, R. and Miller, N. (2004). Toxicity of Spinosad in Protein Bait to
Three Economically Important Tephritid Fruit Fly Species (Diptera: Tephritidae) and
Their Parasitoids (Hymenoptera: Braconidae). Journal of Economic Entomology,
97(3): 911-915.
Stenberg, J.A. (2017). A Conceptual Framework for Integrated Pest Management. Trends
in Plant Science, 22(9): 759-769.
Sterk, G. et al., (1999). Results of the Seventh Joint Pesticide Testing Programme Carried
Out by the IOBC/WPRSWorking Group “Pesticides and Beneficial Organisms”.
BioControl, Vol.44, No.1, (March 1999), pp.99-117.
Talebi, K., Kavousi A. and Sabahi Q. (2008). Impacts of Pesticides on Artropod
Biological Control. Pest Technology, 2(2): 87-97.
Talhinhas, P., Loureiro, A. and Oliveira, H. (2018). Olive anthracnose: a yield- and oil
quality-degrading disease caused by several species of Colletotrichum that differ in
virulence, host preference and geographical distribution. Molecular Plant Pathology,
19(8): 1797-1807.
Talhinhas, P., Mota-Capitão, C., Martins, S., Ramos, A.P., Neves-Martins, J., Guerra-
Guimarães, L., Várzea, V., Silva, M.C., Sreenivasaprasad S. and Oliveira, H.
(2011). Epidemiology, histopathology and aetiology of olive anthracnose caused by
Colletotrichum acutatum and C. gloeosporioides in Portugal. Plant Pathology, 60(3):
483-495.
Tappert, L. Pokorny, T., Hofferberth, J. and Ruther, J. (2017). Sublethal doses of
imidacloprid disrupt sexual communication and host finding in a parasitoid wasp.
Scientific Reports, 7: 42756.
Torrini, G., Mazza, G., Benvenuti, C. and Roversi, P.F. (2017). Susceptibility of olive
fruit fly, Bactrocera oleae (Diptera: Tephritidae) pupae to entomopathogenic
nematodes. Journal of Plant Protection Research, 57(3): 318-320.
Uma, S. Jacob, S. and Lyla, K.R. (2014). Acute contact toxicity of selected conventional
and novel insecticides to Trichogramma japonicum Ashmead (Hymenoptera:
Trichogrammatidae). Journal of Biopesticides, 7(Supp.): 133-136.
Usta, C. (2013). Microorganisms in Biological Pest Control - A Review (Bacterial Toxin
Application and Effect of Environmental Factors) in Current Progress in Biological
Research (ed. Silva-Opps, M.), 287-317.
Vianna, U.R., Pratissoli, D., Zanuncio, J.C., Lima, E.R., Brunner, J., Pereira, F.F. and
Serra, J.E. (2009). Insecticide toxicity to Trichogramma pretiosum (Hymenoptera:
Trichogrammatidae) females and effect on descendant generation. Ecotoxicology, 18:
180-186.
Viñuela, E., Medina, Mª.P., Schneider, M., González, M., Budia, F., Adán, A. and Del
Estal. P. (2001). Comparison of side-effects of spinosad, tebufenozide and
azadirachtin on the predators Chrysoperla carnea and Podisus maculiventris and the
parasitoids Opius concolor and Hyposoter didymator under laboratory conditions.
OILB/IOBC Bull., 24(4): 25-34.
Vontas, J. Hernández-Crespo, P., Margaritopoulos, J.T, Ortego, F., Feng, H.-T.,
Mathiopoulos, K.D. and Hsu, J.-C. (2011). Insecticide resistance in Tephritid flies.
Pesticide Biochemistry and Physiology, 100: 199-205.

Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
166
Vontas, J.G., Cosmidis, N., Loukas,M. Tsakas, S., Hejazi, M.J., Ayoutanti, A. and
Hemingway, J. (2001). Altered Acetylcholinesterase Confers Organophosphate
Resistance in the Olive Fruit Fly Bactrocera oleae. Pesticide Biochemistry and
Physiology, 71: 124-132.
Wang, Q. L. and Liu, T.-X. (2016). Effects of Three Insect Growth Regulators on
Encarsia formosa (Hymenoptera: Aphelinidae), an Endoparasitoid of Bemisia tabaci
(Hemiptera: Aleyrodidae). Journal of Economic Entomology, 1-8.
Weinzierl, R.A. (2008). Integrating pesticides with biotic and biological control for
arthropod pest management. In Integrated Pest Management: Concepts, Tactics,
Strategies and Case Studies (pp. 179-191). Cambridge University Press.
Williams, T. Javier Valle, J. and Viñuela, E. (2003). Is the Naturally Derived Insecticide
Spinosad® Compatible with Insect Natural Enemies? Biocontrol Science and
Technology, 13: (5) 459-475.
Yokoyama, V.Y., Rendón, P.A. and Sivinski, J. (2008). Psyttalia cf. concolor
(Hymenoptera: Braconidae) for Biological Control of Olive Fruit Fly (Diptera:
Tephritidae) in California. Environmental Entomology, 37(3): 764-773.
Youssef, A.I., Nasr, F.N., Stefanos, S.S., Elkhair, S.S.A., Shehata, W.A., Agamy, E.,
Herz, A. and Hassan, S.A. (2004). The side-effects of plant protection products used
in olive cultivation on the hymenopterous egg parasitoid Trichogramma cacoeciae
Marchal. Journal of Applied Entomology, 128(9-10): 593-599.
Zapata, N., Medina, P., González, M., Budia, F., Rodríguez, B. and Viñuela, E. (2004).
E. Toxicidad de azadirona y 1,7 + 3,7-di-O-acetilhavanensina (4:1) sobre adultos de
Psyttalia concolor (Szépligeti) (Hym.: Braconidae). Boletín de sanidad vegetal Plagas,
30: 783-789.
Zimmermann, G. (2007). Review on safety of the entomopathogenic fungi Beauveria
bassiana and Beauveria brongniartii. Biocontrol Science and Technology, 17: 553-
596
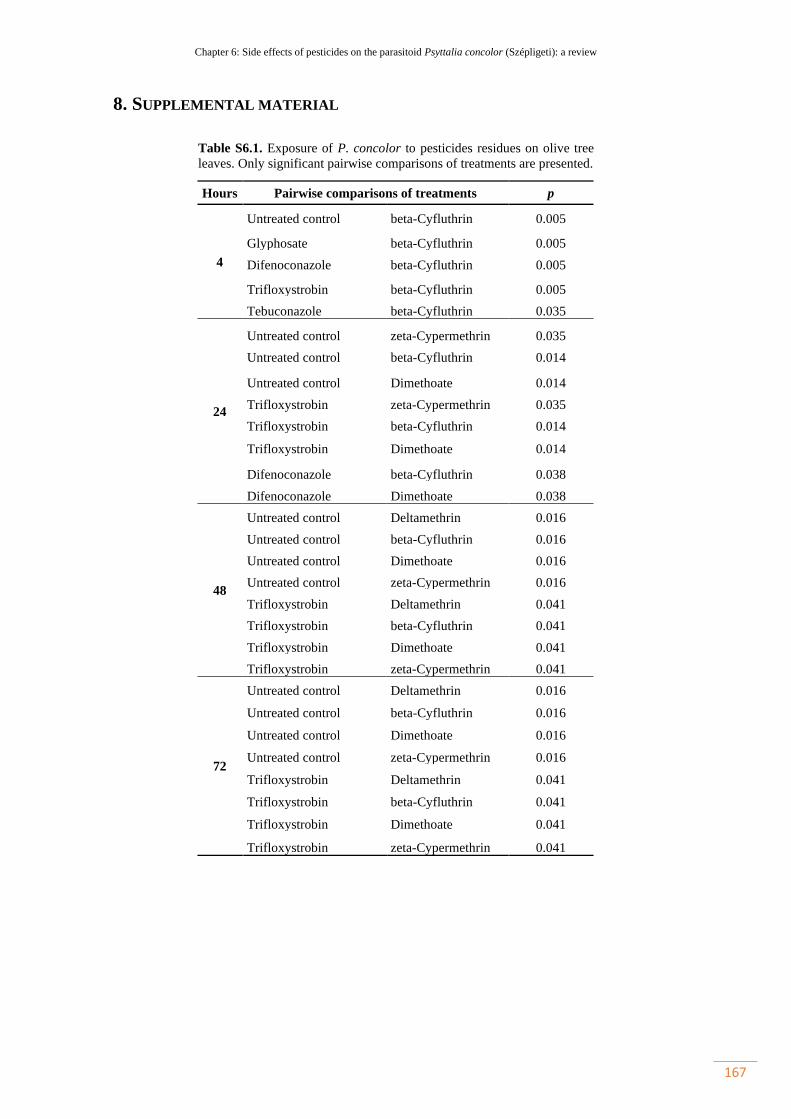
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
167
8. SUPPLEMENTAL MATERIAL
Table S6.1. Exposure of P. concolor to pesticides residues on olive tree
leaves. Only significant pairwise comparisons of treatments are presented.
Hours Pairwise comparisons of treatments p
4
Untreated control beta-Cyfluthrin 0.005
Glyphosate beta-Cyfluthrin 0.005
Difenoconazole beta-Cyfluthrin 0.005
Trifloxystrobin beta-Cyfluthrin 0.005
Tebuconazole beta-Cyfluthrin 0.035
24
Untreated control zeta-Cypermethrin 0.035
Untreated control beta-Cyfluthrin 0.014
Untreated control Dimethoate 0.014
Trifloxystrobin zeta-Cypermethrin 0.035
Trifloxystrobin beta-Cyfluthrin 0.014
Trifloxystrobin Dimethoate 0.014
Difenoconazole beta-Cyfluthrin 0.038
Difenoconazole Dimethoate 0.038
48
Untreated control Deltamethrin 0.016
Untreated control beta-Cyfluthrin 0.016
Untreated control Dimethoate 0.016
Untreated control zeta-Cypermethrin 0.016
Trifloxystrobin Deltamethrin 0.041
Trifloxystrobin beta-Cyfluthrin 0.041
Trifloxystrobin Dimethoate 0.041
Trifloxystrobin zeta-Cypermethrin 0.041
72
Untreated control Deltamethrin 0.016
Untreated control beta-Cyfluthrin 0.016
Untreated control Dimethoate 0.016
Untreated control zeta-Cypermethrin 0.016
Trifloxystrobin Deltamethrin 0.041
Trifloxystrobin beta-Cyfluthrin 0.041
Trifloxystrobin Dimethoate 0.041
Trifloxystrobin zeta-Cypermethrin 0.041
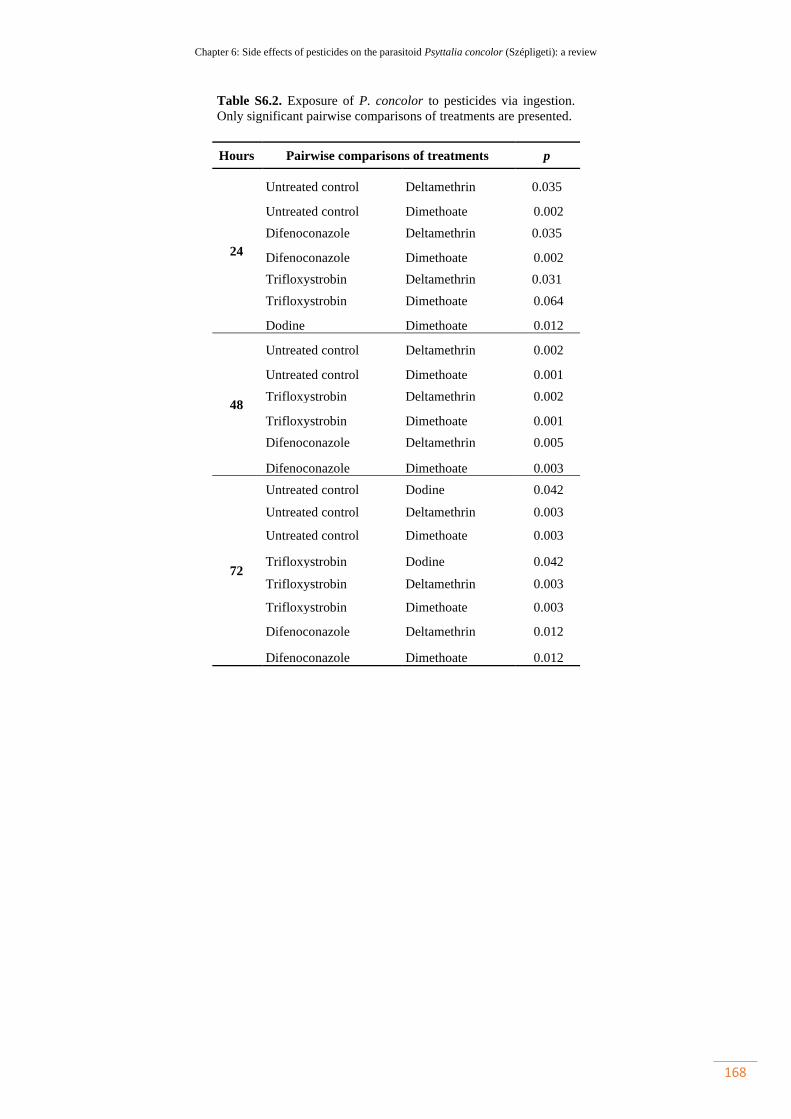
Chapter 6: Side effects of pesticides on the parasitoid Psyttalia concolor (Szépligeti): a review
168
Table S6.2. Exposure of P. concolor to pesticides via ingestion.
Only significant pairwise comparisons of treatments are presented.
Hours Pairwise comparisons of treatments p
24
Untreated control Deltamethrin 0.035
Untreated control Dimethoate 0.002
Difenoconazole Deltamethrin 0.035
Difenoconazole Dimethoate 0.002
Trifloxystrobin Deltamethrin 0.031
Trifloxystrobin Dimethoate 0.064
Dodine Dimethoate 0.012
48
Untreated control Deltamethrin 0.002
Untreated control Dimethoate 0.001
Trifloxystrobin Deltamethrin 0.002
Trifloxystrobin Dimethoate 0.001
Difenoconazole Deltamethrin 0.005
Difenoconazole Dimethoate 0.003
72
Untreated control Dodine 0.042
Untreated control Deltamethrin 0.003
Untreated control Dimethoate 0.003
Trifloxystrobin Dodine 0.042
Trifloxystrobin Deltamethrin 0.003
Trifloxystrobin Dimethoate 0.003
Difenoconazole Deltamethrin 0.012
Difenoconazole Dimethoate 0.012

169
Chapter 7
General
conclusions

Chapter 7: General conclusions
170
1. General conclusions
In order to investigate the effect of non-crop plants on the fitness of P. concolor and
(b) evaluate the susceptibility of parasitoid to some pesticides used in Mediterranean olive
groves, a number of hypotheses were proposed:
✓ Management strategies to increase biological control of B. oleae
✓ Sugars-resources enhance fitness in P. concolor by increased longevity and
reproduction
✓ Amino acids enhance fitness in P. concolor, particularly focusing female
reproduction
✓ Flowering plants improve the impact of P. concolor over B. oleae population
✓ Pesticides application for pest management in olive orchards change parasitoids
P. concolor efficacy in biological control of B. oleae
Existing literature, discused in chapter 3, showed may exemples of reserches to suggest
management strategies to increase biological control of the olive fruit fly.
The laboratory research examined in the chapter 4 showed a positive relationship
between carbohydrate (single sugars and honeydew) and parasitoid survival.
Additionally, results from amino acids research experiments showed that sugar solution
increased the reproductive performance more than amino acids (Chapter 5). These results
agree with several studies revealing the importance of sugars to parasitoids (Benelli et al.,
2017; Russell, 2015). These include the importance of feeding on nectar floral resource
and honeydew on adult parasitoid survival and reproduction increase. This knowledge is
important for biological control programmes in the form of habitat management through
planting nectar-species-rich wildflower strips and hedgerows or application of artificial
sugar sources. In his detailed work on sugar food sources for P. concolor, Furtado et al.
(2016) demonstrated a positive effect of the nectars of Anchusa azurea Mill., Rosmarinus
officinalis L., Lavatera cretica L. and Calamintha nepeta (L.) Savi on parasitoid survival.
Also, when offering flowers directly to parasitoid, Anchusa azurea, Lavatera cretica, and
Foeniculum vulgare L., Malva sylvestris L. and Veronica persica Poir. increased the
survival (Furtado et al., 2016 and Pinheiro et al., unpublished data). Overall, parasitoid
longevity can increase with flowering plants but depends on access to floral resources and

Chapter 7: General conclusions
171
floral architecture, since some plant species do not produce nectar and others have
nectaries that are difficult or impossible for parasitoid to access.
The presence of flowering plants can may create a conflict with herbicides-based
management activities (in conventional agriculture) that would kill the parasitoids. The
presence of flowering plants might create a conflict with herbicides-based management
activities (in conventional agriculture) that would kill the parasitoids. Moreover, P.
concolor can be also contaminated (and killed) when insecticides and fungicides are
sprayed in olive groves, by direct exposure (to spray droplets), by contact (with treated
surfaces) and by oral uptake from contaminated food sources (e.g. feeding on
contaminated water, pollen, nectar or honeydew available on plant parts), as demonstrated
in the review and laboratory experiments of chapter 6. As expected, this study focused on
survival and reproduction of P. concolor, insecticides (especially organophosphates and
pyrethroids) are more toxic when compared to fungicides, herbicides and biopesticides.
A solution to minimize impact of pesticides on P. concolor should be the implementation
of flowering plants adjacent to crop fields.
Overall, this study contributed to increase the current knowledge on possible
nutritional feeding of P. concolor, aiming its application as a possible biological control
against B. oleae.
2. References
Benelli, G., Giunti, G., Tena, A., Desneux, N., Caselli, A. and Canale, A. (2017). The
impact of adult diet on parasitoid reproductive performance. Journal of Pest Science,
90(3): 807-823.
Furtado, C., Belo, A.F., Nunes, F.M., Ganhão, E., Müller, C.T., Torres, L. and Rei, F.T.
(2016). Evaluating potential olive orchard sugar food sources for the olive fly
parasitoid Psyttalia concolor. BioControl, 61(5): 473-483.
Russell, M. (2015). A meta-analysis of physiological and behavioral responses of
parasitoid wasps to flowers of individual plant species. Biological Control, 82: 96-103.

Chapter 7: General conclusions
172
1. Conclusiones generales
Con el fin de investigar (a) la influencia de plantas no cultivadas en el desarrollo y
comportamiento reproductor de P. concolor y (b) evaluar la susceptibilidad del
parasitoide a determinados productos fitosanitarios utilizados en olivares mediterráneos,
se propusieron las siguientes hipótesis:
✓ Diferentes estrategias de manejo incrementan el control biológico de B. oleae
✓ Las fuentes de alimentación ricas en azúcares mejoran la sanidad de P. concolor
al incrementar la longevidad y su capacidad reproductora.
✓ Los aminoácidos mejoran la sanidad de P. concolor, por su particular influencia
positiva en la reproducción de las hembras
✓ Las plantas con flores aumentan la acción parasítica de P. concolor sobre las
poblaciones de B. oleae
✓ La aplicación de productos fitosanitarios para el manejo del olivar interfieren con
la eficacia del parasitoide P. concolor en el control biológico de B. oleae
La literatura previa, discutida en el capítulo 3, muestra muchos ejemplos de trabajos
que sugieren que diferentes estrategias de manejo del olivar pueden favorecer el control
de la mosca del olivo.
El trabajo de laboratorio desarrollado en el capítulo 4 muestra una relación positiva
entre los carbohidratos (azúcares simples y melazas) y la supervivencia del parasitoide.
Además, los resultados obtenidos con experimentos donde se aportan distintos
aminoácidos como fuente de alimento mostraron que las soluciones azucaradas
aumentaban la reproducción de parasitoide en mayor medida que los aminoácidos
(Capítulo 5). Estos resultados concuerdan con diferentes estudios que confirman la
importancia de los azúcares en la supervivencia y reproducción de los parasitoides
(Benelli et al., 2017; Russell, 2015), ya sea porque se alimenten de néctar floral o melazas.
Este conocimiento es importante en programas de control biológico que introducen el
manejo del hábitat, ya sea mediante la plantación de márgenes florales o setos con
especies ricas en néctar o por la aplicación de fuentes de azúcares artificiales. En el
estudio de Furtado et al. (2016) con el parasitoide P. concolor se demostró el efecto
positivo de los néctares de Anchusa azurea Mill., Rosmarinus officinalis L., Lavatera

Chapter 7: General conclusions
173
cretica L. y Calamintha nepeta (L.) Savi en la supervivencia del parasitoide. Además,
cuando se ofrecieron flores directamente al parasitoide, Anchusa azurea, Lavatera
cretica, Foeniculum vulgare L., Malva sylvestris L. y Veronica persica Poir. se
incrementó su supervivencia (Furtado et al., 2016 y Pinheiro et al., datos no publicados).
En resumen, la longevidad del parasitoide puede aumentar si se alimenta de néctar de
plantas en flor, pero dependerá del acceso a los recursos florales y de la arquitectura floral
de la planta, ya que algunas especies no producen néctar y otras tienen nectarios a los que
el parasitoide no puede acceder. La presencia de plantas con flores es incompatible con
actividades de manejo de control de malas hierbas basadas en herbicidas (agricultura
convencional), que podrían matar, además, al parasitoide. Además, P. concolor se puede
contaminar (y por tanto morir) cuando insecticidas y fungicidas son pulverizados en los
cultivos, ya sea de forma directa (mediante gotas), por contacto (con superficies tratadas
y ya secas) y/o por alimentación en recursos alimenticios contaminados (por ejemplo,
beber agua contaminada o alimentarse de polen, néctar o melazas disponibles en partes
de la planta tratadas, como se demostró en la revisión y experimentos del capítulo 6).
Como era de esperar, los insecticidas (especialmente organofosforados y piretroides) son
más tóxicos que los fungicidas, herbicidas y bioplaguicidas. Una propuesta para
minimizar el impacto de los productos fitosanitarios en P. concolor sería la
implementación de márgenes florales adyacentes a las parcelas cultivadas. Como
conclusión, este estudio contribuyó a aumentar el conocimiento actual de los
requerimientos nutritivos de P. concolor, con el objetivo de aplicarlos para mejorar el
control biológico contra la mosca del olivo B. oleae.
2. References
Benelli, G., Giunti, G., Tena, A., Desneux, N., Caselli, A. and Canale, A. (2017). The
impact of adult diet on parasitoid reproductive performance. Journal of Pest Science,
90(3): 807-823.
Furtado, C., Belo, A.F., Nunes, F.M., Ganhão, E., Müller, C.T., Torres, L. and Rei, F.T.
(2016). Evaluating potential olive orchard sugar food sources for the olive fly
parasitoid Psyttalia concolor. BioControl, 61(5): 473-483.
Russell, M. (2015). A meta-analysis of physiological and behavioral responses of
parasitoid wasps to flowers of individual plant species. Biological Control, 82: 96-103.





























