Embed Size (px)
Citation preview

172 Journal of Vector Ecology June2012
Energy-state dependent responses of Anopheles gambiae (Diptera: Culicidae) to simulated bednet-protected hosts
SimonP.W.ZappiaandBernardD.Roitberg
Department of Biology, Simon Fraser University, Burnaby, BC V5A 1S6, Canada, [email protected]
Received 21 September 2011; Accepted 3 February 2012
ABStRAct: Innature,Anopheles gambiaemosquitoesarefoundatvariousenergylevelsandsuchfemalesmustchoosebetweenseekingsomaticenergyfromsugarsourcesandobtainingbothsomaticandgameticenergyfrombloodhosts.Weusedastraight-tubeolfactometercontainingasimulatedunobtainablebloodhost(humanfootsmellprotectedbyanet)aswellasasugarsource(honeyodor).Weassessedfemaleprobingrateandresidencetimeatthenetasafunctionofenergystate (0,24,48,72-hstarved). Inour trials,0-hstarved femalesshowed lowresponse tohumanodor, lowprobingrate,andresidencetimeatthehumanodorsite.Bycontrast,both48and72-hindividualsshowedhighresponsetofootodor,longerresidencetime,andhigherprobingrates.Seventy-two-hfemalesalsoflewtowardsthehoneysourcelessoftenthanothergroups.Ourfindingssuggestthatmanagingsugarsourcesmightbeaviablestrategyforinfluencingmosquitobitingbehavior.Journal of Vector Ecology 37 (1): 172-178. 2012.
Keyword Index: Anopheles gambiae, behavior,energystate,bitingrate,foragingtheory,olfactometer.
INtRODUctION
An. gambiae is the main vector of Plasmodium spp.,thecausativeagentofmalaria inSub-SaharanAfrica.Thedisease causes both mortality (~800,000 people yearly)andmorbidityinthemillions,anditisasignificantsocial-economicburdeninallendemiccountries(WHO2010).tocombatthespreadofmalaria,itisimportanttounderstandboththelife-historyandbehaviorofitsvector(chavesandKoenraadt2010).
Upon emergence, adult An. gambiae have lowteneral reserves and, generally, will readily feed on sugarsources (Stone et al. 2011). After two days, females beginresponding to human odor stimuli (Foster and takken2004) and are faced with two possible feeding choiceswhich are often mutually exclusive due to the mosquito’ssize-constrained crop (Fernandes and Briegel 2004). Anindividualfemalemosquitomaychoosetocontinuefeedingonreadilyavailable sugar to support somatic function,orfeed from a blood host. The latter would provide energyforbothsomaticmaintenanceandreproductionbutislessreadily obtainable and more dangerous to acquire. Sugaravailability has been linked to longer lifespan (Straif andBeier1996,GaryandFoster2001,GaryandFoster2004),increased inseminationratesbymales (Stoneetal.2009),andisrelevanttomanagementofmalaria(Guetal.2011).
to maximize lifetime fitness, the choice betweensomaticandgameticenergysources(Stoneetal.2011)maybemediatedbybothexternalandinternalfactors(MaandRoitberg2008,RoitbergandMangel2010).Externalfactors,suchastheriskofdeathduetoadefensivehost,presenceofpotentialpredators,andabednetmaylowerthemosquito'sattackpersistencewhenattemptingtoobtainabloodmealfromaparticularhost(WalkerandEdman1985).Likewise,internalfactorssuchasenergylevel,age,andlifeexpectancy
oftheindividualmayaffectthispersistence(Roitbergetal.1993).Bothexternalandinternalfactorsmaybeassessedbythefemalepriortoseekingasourceofenergy.
For years, insecticide-treated bednets (ItNs) havebeen the most effective tools against Anopheles bites(D’Alessandro2001).Whilemanystudieshaveinvestigatedmosquito interactions with bednets (Lines et al. 1987,takken2002),toourknowledge,nostudyhastestedhowenergystateaffectstheseinteractions,althoughtheoreticalstudieshaveexaminedthisrelationship(MaandRoitberg2008,RoitbergandMangel2010).
Giventhatadultshavelimitedabilitytomaintainenergyatconstanthighlevels,ourinterestliesinunderstandingthepointatwhichamosquitogivesuponapotentialbloodmealandhowenergystateaffectstheprobingrateandtimespentwith the potential host as well as the kind of interactionswithanunobtainablehost.Similarstudieshaveaddressedthisissue(WalkerandEdman1985,Nasci1991withAedes aegyptiandRoitbergetal.2010awithAn. gambiae),butthemosquitoes were being actively dislocated from the hostandnoalternativefoodsourcewasprovided.Inthisstudy,wepredictedthatenergy-deprivedindividualswouldhaveincreasedprobingratestowardsanunobtainablebloodhost,givinguplessreadilythanmosquitoeswhoseenergylevelsarehighandthushavelowerriskofstarvation.Incontrast,individualswithahigherenergylevelwouldhaveawiderrange of options: they could wait for the host to becomeavailableormoveontoanothersourceofenergy,seekingamoreobtainablebloodhostorsugarsource.
to test these predictions, we employed a straight-tubeolfactometer inwhichwe simulatedahuman that isbeingprotectedbyabednetwithinadomicile.We testedindividualfemalemosquitoesatvariouslevelsofstarvationandgavethemtwopotentialfoodoptions:anunobtainablebloodmealandasugarsource.

Vol.37,no.1 Journal of Vector Ecology 173
MAtERIALSANDMEtHODS
ColonyAnopheles gambiae s.s. was reared from a colony
originatingfromNjagi,tanzania, in1997.Ourlaboratoryculture was kept at 28 ± 2° c, 83 ± 3% RH under theL:D 12:12 h photoperiod with 1 h of twilight transition(conviron).Eggswerecollectedonmoistfilterpaperfromovipositingadultsthatwereblood-fedbyoneoftheauthors(SZ). Eggs hatched within 72 h and 1st instar larvae wereplaced in largewhiteplasticcontainers (30cmx45cmx6cm)with3.5litersofwater.Thelarvaewerefedfishflakefood(Nutrafin-Hagen)ad lib untilpupaeweretransferredin water-filled containers to Perspex cages with screeningon three sides (30 cm x 30 cm x 30 cm) under the sameRH and L:D conditions. Following eclosion, adults wereprovided with 5% sucrose solution ad lib from a medicalgrade cotton wick (Richmond Dental) contained in anErlenmeyerflask.All145experimentalfemaleswerelarge(wing length of 2.70-3.50 mm), 3-9 days post-eclosion,not previously blood-fed, presumed mated (mating wasobserved inside the cages), and without visibly swollenabdomenstoincreasethelikelihoodoffemaleresponsetoodorstimuli(FernandesandBriegel2004).
Experimental apparatusWeusedalarge,custom-madeglasstube(olfactometer)
(Figure1),145cminlengthwithdiameterof17cmwhichallowedunconstrainedflightofthemosquito.Anairpump(PetceteraAP5000doubletype)pushedambientairthroughanactivatedcarbonfilterat0.025liters/sandfunnelledittothetubeatmid-heightatoneendofthechamber.Acottonball containing approximately 5 g of honey (Kidd bros.unpasteurizedalfalfaandcloverhoney),whichwasfoundtobeattractiveat fullstrength(pers.obs.),wasplacedona metallic tray suspended midway into the tube. A semi-rigid,plastic,cylindrical,tancolorednet(10cminheight,2.5cmindiameter,1x1mmmeshholes),whichservedasa simulated bednet containing a conditioned nylon sock(Secretbrand,kneehigh),wasplaceddownwindfromthehoney,atadistanceof73cmandwassuspendedfromthetop(Figure1).ThesockwaswornbySZfortwoconsecutivedays tocondition it tobeattractive to femalemosquitoes(Braksandtakken1999,Smallegangeetal.2010);on the
secondday,SZperformedastrenuoussport.Thesockwasthen placed in a sealed plastic bag and incubated in theexperimentalchamberforonefulldaypriortobeingusedin the olfactometer. The sock was substituted every fourtofivedaysand itwasrandomlyassigned to thedifferentstarvationtreatments.Avinylpipewasplacedattheupperextremityofthesimulatedbednettoallowtheexperimentertoexhaletosimulatethepresenceofalivebloodhostandstimulate host-searching by the mosquitoes (Healy andcopland 1995). At the end of the tube (60 cm from thesimulatedbednet)weplacedaplasticreleasechamberwithamovablemetallicscreen.It isimportanttonotethattheexperiment was arranged such that released mosquitoesencounteredthesimulatedbloodhostpriortothesimulatednectar source in a linear arrangement. The experimentalruns were observed under dim red light (average of 1.79μmol/(s*m2)). The olfactometer was aerated and flusheddailywithplainwatertominimizeresidualodors.
Experimental proceduretreatments of unstarved (0-h) and 24, 48, and 72-h
starvation were randomly assigned to adults aged threeto nine days post-pupal-eclosion to avoid any age effect.The sugar source was then removed and substituted withdeionizedwater(Knolsetal.1994).Femalemosquitoeswererandomly selected and individually placed into a releasechamber.Aftera3-minrestingperiod, thewire screenofthe release chamber was opened, allowing the mosquitointotheolfactometer.Theexperimenterthenbegantimingthestartoftherunandstartedblowingquickpulsesofairevery6-8 svia thevinyl tube,whichwere sustaineduntilthe end of the experimental run. If the mosquito wasunresponsivefor3mintheexperimenterwouldgentlytaponthewallsofthereleasechamberuntilthefemaleexitedintotheolfactometeratwhichpointtheexperimentstarted.We monitored the female’s probing rate by counting thenumberofattemptedlandings/probesonthesockandnet(Nasci1991)perunittime.Weconsideredalandingtobecompletedoncethefemaletouchedthenetanddidnotflymorethan5cmawayfromthenet.
We also recorded the residence time at the net withsockcalculatedas the timespent inphysicalcontactwiththenet.Theexperimentwasterminatedifthefemaleflewupwindpastaline5cminfrontofthecottonballcontaining
Figure 1. Experimental olfactometer setup. A. Mosquito release chamber. B. Simulated bed net containing aconditionedsockandavinyltubethroughwhichtheexperimentexhaledair.c.Positionofhoneyattractant.D.carbonfilter.E.Airpump.

174 Journal of Vector Ecology June2012
honeyorrestedonthesidesoftheglasstubeformorethanfiveconsecutiveminutes.Onceareplicatewasconcluded,thefemalewasremoved,dried,anditswingsremovedandmeasured using an ocular micrometer and the softwareAnalyzingDigitalImages,Version11-2008.
Data analysisAllstatisticsandgraphswereprocessedusingJMP9and
GraphPadPrism5.Weusedaχ2testtoassesssignificancefor thenumberofAn. gambiae responding to thehumanodor stimulus as well as the number of mosquitoes thatactivelyflewtothehoneysourceorrested.Kruskall-Wallistest was used for analyzing significant differences in theprobingrate,residencetimeatsockandnet,aswellastheaveragetimeperprobingeventbetweentreatments,duetothezeroinflateddistributionofthevalues.Lastly,Log-Rank(Mantel-cox)(i.e.,survivalanalysis)testwasusedtoanalyzethe differences in sustained activity towards obtaining abloodmealasafunctionofenergystate.Agestratification(number of days after emergence) was included in eachanalysis.
RESULtS
A total replicate size of 145 females met our criteriaof evaluation. Single experimental runs lasted from a fewsecondstoamaximumof27min(average~3min).
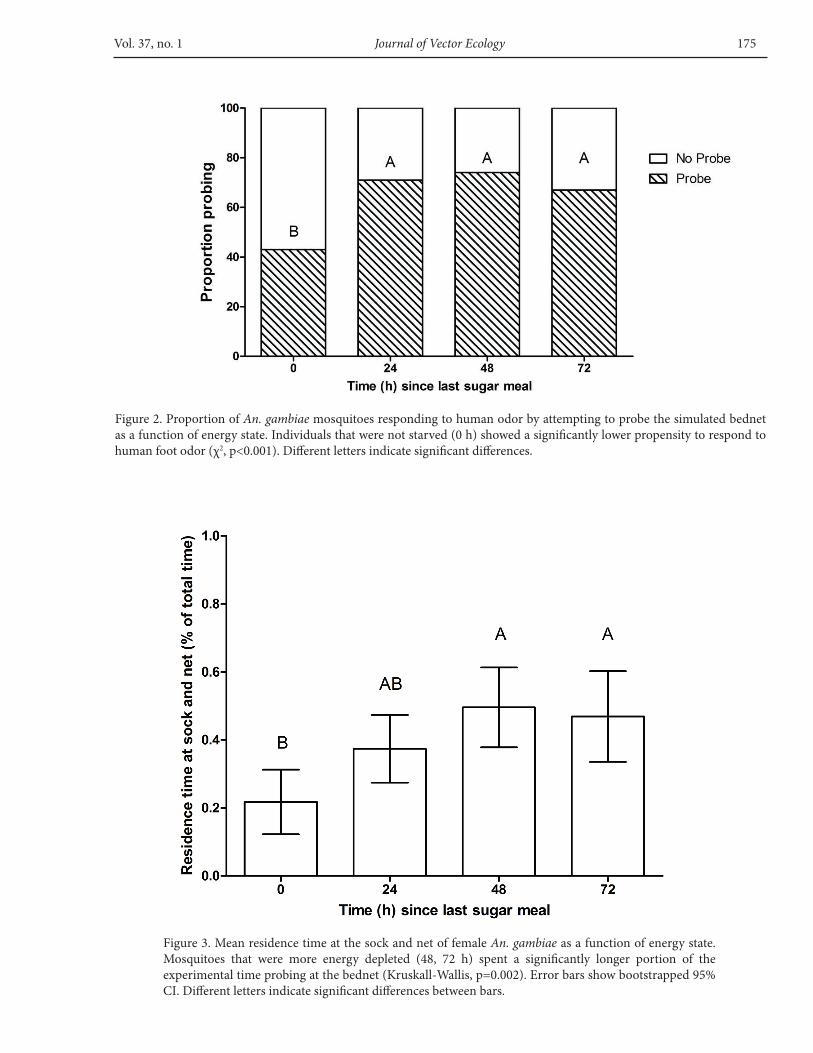
Non-starved mosquitoes (0-h) responded to thehumanodorbyprobingthesimulatedbednetsignificantlyfewer times than all the other treatments, as only 43% oftheindividualswererecordedtohaveprobedatleastonceduring the experiment. The other groups had a higherproportionofindividualsrespondingtothehumanodorat71%,74%,and67%for24,48,and72h,respectively(χ2df.=3,F=25.91,p<0.001)(Figure2).
The relative residence time at the sock and net (i.e.,the proportion of experimental time spent probing) ofnon-starved females was 22±0.1% and was significantlydifferent from the 48-h and 72-h groups (Kruskall-WallisF=15.45,p=0.002).Thesetreatmentsyieldedto50±0.1%and47±0.1%relativeresidencetime,respectively,whilethe24-hstarvationtreatmentaveragedat37±0.1%(Figure3).
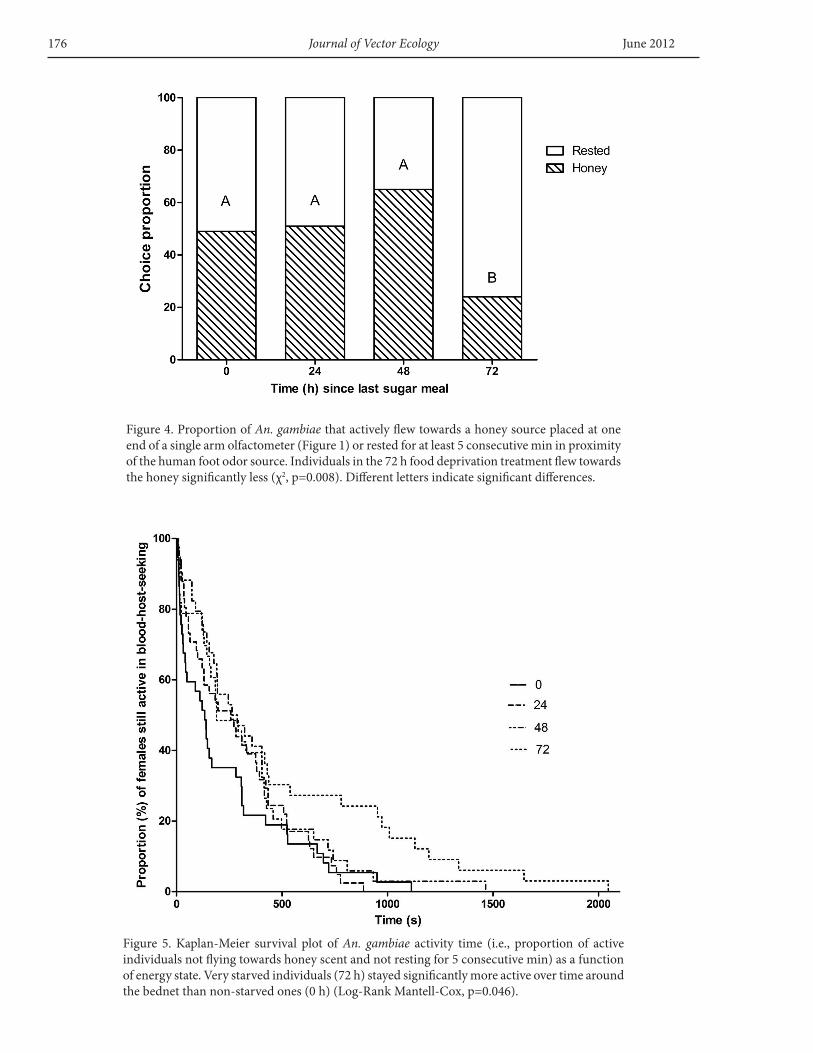
When considering the proportion of individuals thatactivelyleftthe“hut”byflyingtothehoneysourceatoneend of the olfactometer vs the proportion that rested inthe immediate surroundingsof thehumanodor, theonlysignificantly different group was the 72-h treatment inwhich24%oftheindividualsmovedtothehoney(χ2,df.=3,F=35.02,p<0.001).Theothergroupsscored48%,51%,and65%for0,24,and48h,respectively(Figure4).
The proportion of females still active in host-seekingafter a given time, measured as total experimental time,yielded significant differences between the two extremetreatments: 0 and 72 h. The latter showed a much longeroverallactivehost-seekingtimelength(Log-RankMantell-coxF=7.985,p=0.046)(Figure5).
Probingrate(totalprobes/totalexperimentaltime)wassignificantlydifferentbetween0-h(0.239±0.001bites/min)
and 48-h (0.481±0.001 bites/min) treatments (Kruskal-Wallis,F=9.563,p=0.023).24hand72hwererecordedat0.407±0.001and0.430±0.001bites/min,respectively.
Nosignificantresultswereobtainedwhencomparingthe average length of time per landing (~67 s across alllevels) on the simulated bednet between energy levels(Kruskall-WallisF=4.232,p=0.238).
DIScUSSION
Inthisinvestigation,mosquitoeswithdifferentenergylevels were offered a choice between an unobtainable(simulated)hostunderabednetandanalternativesourceof energy,honey, aproxy foranectar sourceoutsideof adomicile. The experiment was designed spatially suchthat themosquitowouldencounter thebloodhostbeforethehoney(Figure1).Thegoalwastoseeiffemaleswouldseek the unobtainable blood meal indefinitely or give upthis potential source of energy to actively fly towards thealternativesugarsource.
We found that An. gambiae female mosquitoesmodulate their behavior toward trying to bite anunobtainable host based on their energy state. In nature,femalesarefoundwithabroadrangeofenergystates,andour research helps elucidate how they might react to thepresenceoftwopotentialenergysourceswithverydifferentfitnessadvantagesandcostsfortheindividualfemale.
Mosquitoes that were granted ad libitum access tosugarwateruntilthestartoftheexperiment(0hstarved)wereoveralllessinclinedtorespondtoanyofthetwoodorstimuli (Figure 2). This absence of response suggests thatthehighenergylevel,perhapscoupledwithaperceivedfullcrop, reduces meal-seeking activity (Jones and Madhukar1976).ThesefindingsareinlinewithRoitbergetal.(2010a)whereitwasshownthatwell-fedfemaleshadlowerattackrates and were predicted by Ma and Roitberg (2008). Inthe few instances where these females probed for blood,theirresidencetimeatthesockandnetwasshortandtheytendedtohavelowerprobingratesandtoterminateblood-host-seekingsignificantlyearlierthanothergroups(Figures3,4,and5).
Here,wesuggestthatrecentlyfedfemalesmaypostponetheriskofattackingabloodhostorventuringawayfromituntilcropvolumeandmassdrops,perhapsmaximizingthe size of the future engorged meal and survivorship(RoitbergandGordon2005,MaandRoitberg2008).Thiscanbebestunderstoodusingamarginalanalysiswhereinthe marginal returns from both space-constrained bloodmealsandsugarmealsaresmallforrecentlyfedindividuals,whereasthemarginalcostsviamortalityriskarehigh(seeRoitbergetal.2010bforanexamplefromparasiticwasps).Incontrast,themarginalreturnsforstarvedindividualsarehighineithercase,butoneofthetwomealoptionsmightbemoreaccessible.Thisstatementissupportedbyourdata.Very starved females (48 and 72 h) had longer residencetimes at the sock and net than other groups and 48 hfemales had a significantly higher probing rate. However,72 h starved females showed no significant difference in
Vol.37,no.1 Journal of Vector Ecology 175
Figure2.ProportionofAn. gambiae mosquitoesrespondingtohumanodorbyattemptingtoprobethesimulatedbednetasafunctionofenergystate.Individualsthatwerenotstarved(0h)showedasignificantlylowerpropensitytorespondtohumanfootodor(χ2,p<0.001).Differentlettersindicatesignificantdifferences.
Figure3.MeanresidencetimeatthesockandnetoffemaleAn. gambiaeasafunctionofenergystate.Mosquitoes that were more energy depleted (48, 72 h) spent a significantly longer portion of theexperimentaltimeprobingatthebednet(Kruskall-Wallis,p=0.002).Errorbarsshowbootstrapped95%cI.Differentlettersindicatesignificantdifferencesbetweenbars.
176 Journal of Vector Ecology June2012
Figure4.ProportionofAn. gambiaethatactivelyflewtowardsahoneysourceplacedatoneendofasinglearmolfactometer(Figure1)orrestedforatleast5consecutivemininproximityofthehumanfootodorsource.Individualsinthe72hfooddeprivationtreatmentflewtowardsthehoneysignificantlyless(χ2,p=0.008).Differentlettersindicatesignificantdifferences.
Figure 5. Kaplan-Meier survival plot of An. gambiae activity time (i.e., proportion of activeindividualsnotflyingtowardshoneyscentandnotrestingfor5consecutivemin)asafunctionofenergystate.Verystarvedindividuals(72h)stayedsignificantlymoreactiveovertimearoundthebednetthannon-starvedones(0h)(Log-RankMantell-cox,p=0.046).

Vol.37,no.1 Journal of Vector Ecology 177
probing rate. Perhaps females in this category are on theverge of starvation and are energetically constrained tooverall lowactivity levels.Thishypothesis issupportedbythe extremely high mortality in the 72 h treatment cagesandthefactthatthistreatmentyieldedfemalesthatrestedalongsidethesimulatedblood-hostmoreoftenthanothergroups;wehypothesizethatthelatterhadsufficientenergytoflytowardsthesugarsourceinsearchofalternativefood.However,wecannotexcludethepossibilitythatmosquitoesthat flew towards the honey source were simply flyingupwind.Althoughsome femaleswereobserved toreadilyfeed on the cotton bearing honey, no data were collectedregardinghoneyavidity.
Finally, the low probing rate and residence time atthe sockandnetbywell-fed individuals suggests that thefemale’s energy level might allow for a flight to anotherperhaps more readily accessible blood host (Lines et al.1987).
Very starved individuals (48 h, 72 h treatments) notonlyrespondedmoreoftentoodorstimulibutalsoshowedahigherpersistencerateandresidencetimeatthesockandnet.Thepropensitytostayincontactforalongerdurationoftimewiththebednetcouldbeadvantageousinthefielduse of insecticide treated bednets (ItNs), as increasedcontact time with the chemicals proportionally increasesthelikelihoodofdeathbypoisoning(RoitbergandMangel2010).
Thelackofsugarsourcesintheimmediatevicinityofhumanhabitationsmaycontributetotheoveralldepletionofenergyinfemales,increasingtheexposuretothechemicalson ItNs, and this fact should be considered by malariavector managers. Similar considerations are relevant forfemales that have undergone changes due to Plasmodiuminfection(shortenedoverall lifeexpectancyand increasedriskybehavior)astheymaybelinkedtodepletionofenergyinthemosquitohost(FergusonandReid2002,RiveroandFerguson2003).
We were expecting that starved individuals wouldspendonaveragealongertimeperprobebutthatdidnotoccur.Thereareat least twoexplanationsforthisfinding.First,An. gambiaecouldhaveevolvedaninnatemaximumlength of time allowed per unsuccessful probe (see fixedgivinguptimesinforagingtheory,KrebsandDavies1997),whichcouldhaveevolvedinresponsetotheriskofreactionbythehostandthereforepotentialdeathofthemosquito.Second,byprovidinganadditionalfeedingoption(sugar),the energy state effect at the unobtainable host waseffectively removed; i.e., starved and well-fed individualsgave up at the same time but for different reasons, withstarved individualsdemonstrating immediate attention toenergystateandwell-fedindividualsforgoingcurrentriskytactics for future opportunities (see Roitberg and Mangel2010).
Our findings suggest that the management of sugarsources within a village should be a priority. In fact, assuggested by Utzinger et al. (2001), the then incidentalmanagement of vegetation within breeding grounds leadtoloweringmalarialoccurrenceinaregionofZambia.We
hereadvocateextendingthemanagementtonectar-bearingplantswithinforagingareasinintegratedpestmanagementprograms and the introduction of poisonous sugar baitswhich have shown to be an efficient control strategy inIsrael(MüllerandSchlein2008).
Further research is needed in this field and we arecurrently assessing whether An. gambiae behaves in thesamemannerinsemi-fieldconditions.
Acknowledgments
WethankDr.DavidGillespieandDr.GerhardGriesfor their involvement in designing this experiment.Thanks to Regine Gries, Amber Hoi, Karen Mok, AmberRichmond,andMikaPhillipsforthetechnicalsupportandtheRoitberglabforusefulinsightsandforreviewingearlyversionsofthismanuscript.
REFERENcEScItED
Braks, M.A.H. and W. takken. 1999. Incubated humansweatbutnotfreshsweatattractsthemalariamosquitoAnopheles gambiae sensu stricto.J.chem.Ecol.25:663-672.
chaves, L.F. and c.J.M. Koenraadt. 2010. climate changeandhighlandmalaria:freshairforahotdebate.Quart.Rev.Biol.85:27-55.
D’Alessandro, U. 2001. Insecticide treated bed nets topreventmalaria–Thechallengeliesinimplementation.Br.Med.J.322:249-250.
Ferguson, H.M. and A.F. Read. 2002. Genetic andenvironmental determinants of malaria parasitevirulence in mosquitoes. Proc. R. Soc. Lond. B 269:1217-1224.
Fernandes,L.andH.Briegel.2004.Reproductivephysiologyof Anopheles gambiae and Anopheles atroparvus. J.VectorEcol.30:11-26.
Foster, W.A. and W. takken. 2004. Nectar-related vs.human-related volatiles: behavioural response andchoicebyfemaleandmaleAnopheles gambiae(Diptera:culicidae)betweenemergenceandfirst feeding.Bull.Entomol.Res.94:145-157.
Gary,R.E.andW.A.Foster.2001.EffectsofavailablesugaronthereproductivefitnessandvectorialcapacityofthemalariavectorAnopheles gambiae(Diptera:culicidae).J.Med.Entomol.38:22-28.
Gary,R.E.andW.A.Foster.2004.Anopheles gambiaefeedingand survival on honeydew and extra-floral nectar ofperidomesticplants.Med.Vet.Entomol.18:102-107.
Gu, W.D., G. Muller, Y. Schlein, R.J. Novak, and J.c.Beier. 2011. Natural plant sugar sources of Anophelesmosquitoes strongly impact malaria transmissionpotential.PLoSOne6:e15996.
Healy, t.P. and M.J.W. copland. 1995. Activation ofAnopheles gambiae mosquitos by carbon dioxide andhumanbreath.Med.Vet.Entomol.9:331-336.
Jones,J.c.andB.V.Madhukar.1976.Effectsofsucroseonbloodavidityinmosquitoes.J.InsectPhysiol.22:357-

178 Journal of Vector Ecology June2012
360.Knols,B.G.J.,R.De Jong,andW.takken.1994.trapping
system for testing olfactory responses of the malariamosquito Anopheles gambiae in a wind tunnel. Med.Vet.Entomol.8:386-388.
Krebs,J.R.andN.B.Davies.1997. Behavioural Ecology: An Evolutionary Approach,4thed.Oxford:Blackwell.
Lines,J.D.,J.Myamba,andc.F.curtis.1987.Experimentalhut trials of permethrin-impregnated mosquito netsandeavecurtainsagainstmalariavectorsintanzania.Med.Vet.Entomol.1:37-51.
Ma, B.O. and B.D. Roitberg. 2008. The role of resourceavailability and state-dependence in the foragingstrategyofblood-feedingmosquitoes.Evol.Ecol.Res.10:1111-1130.
Müller, G.c. and Y. Schlein. 2008. Efficacy of toxic sugarbaitsagainstadultcistern-dwellingAnopheles claviger.t.Roy.Soc.trop.Med.Hyg.102:480-484.
Nasci,R.S.1991.InfluenceoflarvalandadultnutritiononbitingpersistenceinAedes aegypti(Diptera:culicidae).J.Med.Entomol.28:522-526.
Rivero,A.andH.M.Ferguson.2003.Theenergeticbudgetof Anopheles stephensi infected with Plasmodium chabaudi: is energy depletion a mechanism forvirulence?Proc.R.Soc.Lond.B270:1365-1371.
Roitberg, B.D. and I. Gordon. 2005. Does the Anophelesbloodmeal-fecunditycurve,curve?J.VectorEcol.30:83-86.
Roitberg,B.D.andM.Mangel.2010.Mosquitobitingandmovement ratesasanemergentcommunitypropertyand the implications for malarial intervention. Isr. J.Entomol.Evol.56:InPress.
Roitberg,B.D.,S.Keiser,andt.Hoffmeister.2010a.State-dependentattacksinamosquito.Physiol.Entomol.35:46-51.
Roitberg,B.D.,K.Zimmerman,andt.Hoffmeister.2010b.Dynamic response to danger in a parasitoid wasp.Behav.Ecol.Sociobiol.64:627-37.
Roitberg,B.D.,J.Sircom,c.A.Roitberg,J.J.M.Vanalphen,andM.Mangel.1993.Lifeexpectancyandreproduction.Nature364:108.
Smallegange, R.c., B.G.J. Knols, and W. takken. 2010.Effectiveness of synthetic versus natural humanvolatilesasattractantsforAnopheles gambiae(Diptera:culicidae)sensustricto.J.Med.Entomol.47:338-344.
Stone, c.M., I.M. Hamilton, and W.A. Foster. 2011. Asurvival and reproduction trade-off is resolved inaccordancewithresourceavailabilitybyvirginfemalemosquitoes.Anim.Behav.81:765-774.
Stone,c.M.,R.M.taylor,B.D.Roitberg,andW.A.Foster.2009. Sugar deprivation reduces insemination ofAnopheles gambiae (Diptera: culicidae), despite dailyrecruitment of adults, and predicts decline in modelpopulations.J.Med.Entomol.46:1327-1337.
Straif,S.c.andJ.c.Beier.1996.Effectsofsugaravailabilityontheblood-feedingbehaviourofAnopheles gambiae(Diptera:culicidae).J.Med.Entomol.33:608-612.
takken, W. 2002. Do insecticide-treated bednets have aneffectonmalariavectors?trop.Med.Int.Hlth.7:1022-1030.
Utzinger, J.,Y.tozan,andB.H.Singer.2001.Efficacyandcost-effectiveness of environmental management formalarialcontrol.trop.Med.Int.Hlth.6:677-687.
Walker,E.D.andJ.D.Edman.1985.The influenceofhostdefensivebehaviouronmosquito(Diptera:culicidae)bitingpersistence.J.Med.Entomol.22:370-372.
World Health Organization. 2010. World Malaria Report2010.Geneva.WorldHealthOrganization.





























