Embed Size (px)
Citation preview

MARINE ECOLOGY PROGRESS SERIESMar Ecol Prog Ser
Vol. 379: 197–211, 2009doi: 10.3354/meps07892
Published March 30
INTRODUCTION
In recent years, many coastal fish resources havebeen overexploited (Castilla 2000), raising doubtsabout the long-term sustainability of certain fisheries(Pauly et al. 2002). Unless changes are implementedimmediately, many fisheries may collapse in the nextfew decades (Worm et al. 2006). The solutions pro-posed by managers for this critical problem are numer-ous (Pauly et al. 2002) and rely on: (1) reducing fishing
capacity through ‘traditional’ fisheries measures (e.g.quotas, reducing fishing effort, regulating fishingequipment); and (2) creating marine protected areas(MPAs). The first option has not always provided theanticipated effects, leading many to adopt MPAs as afishery management strategy (Roberts et al. 2003).
MPAs potentially facilitate the long-term sustain-ability of many fisheries (Gell & Roberts 2003, Ramos-Esplá et al. 2004). Closing areas allows animals to livelonger and grow to maturity, which is important for
© Inter-Research 2009 · www.int-res.com*Email: [email protected]
Effects of habitat on spillover from marineprotected areas to artisanal fisheries
Aitor Forcada1,*, Carlos Valle1, Patrick Bonhomme2, Géraldine Criquet3, Gwenaël Cadiou2, Philippe Lenfant3, José L. Sánchez-Lizaso1
1Unidad de Biología Marina, Departamento de Ciencias del Mar y Biología Aplicada, Universidad de Alicante, POB 99,03080 Alicante, Spain
2GIS Posidonie, Parc Scientifique & Technologique de Luminy, Case 901, 13288 Marseille Cedex 09, France3Laboratoire Ecosystèmes Aquatiques Tropicaux et Méditerranéens, UMR 5244 CNRS–EPHE–UPVD ‘Biologie et Ecologie
Tropicale et Méditerranéenne’, Université de Perpignan, 52 avenue Paul Alduy, 66860 Perpignan Cedex, France
ABSTRACT: Marine protected areas (MPAs) potentially enhance the long-term sustainability ofcoastal fish resources that have been overexploited. The types and quality of habitats, both inside andoutside the MPAs, may determine the likelihood of migration by fish to surrounding unprotectedareas where spillover to fisheries occurs. We assessed whether MPAs enhanced catches of artisanalfisheries, using an experimental fishing study with the same fishing gear as that used by local fishers.This approach allowed us to test the hypothesis of increased catches along the borders of MPAs incomparison with those in other fishing grounds located at medium and far distances from 3 Medi-terranean MPAs: Tabarca Marine Reserve, Carry-le-Rouet Marine Reserve and Cerbère-BanyulsMarine Reserve. Surveys were done over 2 homogeneous habitats (Posidonia oceanica meadow andsand), in 3 different seasons. Catches were significantly higher for some species near the borders ofthe MPAs when fishing on P. oceanica meadows, but not when fishing on sandy bottoms. Thespillover effect appears to be limited by a lack of continuous suitable habitat through the boundariesof the MPA. Some of the species that showed a significant response to protection and concurrenthigher catches near the MPA borders, such as Dentex dentex, Mullus surmuletus, Phycis phycis,Sciaena umbra and Scorpaena porcus, are target species of artisanal fisheries. Although we foundthat the spatial scale of the spillover-induced density gradient was localized, it was sufficient toprovide local benefits to artisanal fisheries. We conclude that spillover effects are not a universalconsequence of siting MPAs in temperate waters and that they are related to the distribution of habi-tats inside and around MPAs.
KEY WORDS: MPA · Export production · Artisanal fisheries · Fish · Habitat connectivity · Spillover ·Mediterranean Sea
Resale or republication not permitted without written consent of the publisher
OPENPEN ACCESSCCESS

Mar Ecol Prog Ser 379: 197–211, 2009
supporting fisheries because of the exponential rela-tionship between fecundity and body size (Bohnsack1990). MPAs are predicted to benefit adjacent fisheriesthrough 2 mechanisms: (1) net emigration of adults andjuveniles across borders, termed ‘spillover’; and (2)increased production and export of pelagic eggs andlarvae (Gell & Roberts 2003, Kaunda-Arara & Rose2004, Abesamis & Russ 2005, Sale et al. 2005). Spilloverof juvenile and adult fish to surrounding non-protectedareas could result from random movements of individ-uals from MPAs to outside their borders or by directedmovements over a large home range (Rakitin & Kramer1996, Kramer & Chapman 1999, Tremain et al. 2004).Emigration may also occur through ontogenetic habitatshifts (Cocheret de la Moriniere et al. 2002, Nagel-kerken & van der Velde 2002). All these cases areconsidered density-independent movements. Anothermechanism that could lead to export is the occurrenceof density-dependent movements of competitivelysubordinate individuals from preferred habitats insideMPAs to suboptimal habitats outside (Sánchez-Lizasoet al. 2000, Abesamis & Russ 2005). Both density-independent and density-dependent processes wouldproduce a gradient of abundance across MPAs bordersand should have an influence on the yields and qualityof the catches in the surrounding fishing grounds (Russ& Alcala 1996, Gell & Roberts 2003). However, thepatchy nature of the marine environment might act asa barrier for the movement of fish, although habitatdiscontinuities might not be perceived in the samemanner by all animals (Wiens et al. 1993). Many fishesare habitat specific and are reluctant to disperse across‘foreign’ habitats (Chapman & Kramer 2000). For thisreason, spillover will also be influenced by the habitatbordering a reserve (Rowley 1994).
Some of the best evidence for spillover comes fromlandings data that have demonstrated increased cap-tures in fisheries adjacent to MPAs in many parts ofthe world: Kenya (McClanahan & Kaunda-Arara 1996,McClanahan & Mangi 2000), Florida and St Lucia(Roberts et al. 2001a), New England (Murawski et al.2000, 2005), the Egyptian Red Sea (Galal et al. 2002)and Apo Island in the Philippines (Russ et al. 2003).However, only a few studies have experimentallytested for spillover through increased captures in adja-cent fished areas, often along density gradients: Bar-bados (Rakitin & Kramer 1996), Kenya (McClanahan& Mangi 2000, Kaunda-Arara & Rose 2004) and ApoIsland in the Philippines (Abesamis & Russ 2005). Inthe Mediterranean, few data are available to assess thevalue of MPAs as a source of biomass to surroundingfisheries (Goñi et al. 2006).
Encouraged by the recent findings on spillover re-sults, many countries and states have started initiativesto establish networks of marine reserves. However,
too little evidence exists to define the basic responsesof fish populations to reserve protection (Willis et al.2003) and their potential for improving fisheries yields(Hilborn et al. 2004). Moreover, reserves remain highlycontroversial among fishers and the fishing industry,who argue that fishery benefits remain unproven.
Protection of breeding stock, enhancement of re-cruitment to neighbouring areas, and restocking ofexploited marine species were the initial goals of the3 Mediterranean MPAs studied here. The main objec-tive of this work was to estimate fisheries enhancementaround these MPAs and to assess the influence ofhabitat on this process. We adopted an experimentalfishing approach with commercial trammel nets, themost common fishing gear used in Mediterraneanartisanal fisheries. This approach allows for testingthe hypothesis that catches will be increased alongthe border of MPAs in comparison with other fishinggrounds. We incorporated 2 habitats in the presentstudy (Posidonia oceanica seagrass meadow and sandybottom), with the aim of testing whether habitat typealters the degree of spillover from MPAs.
MATERIALS AND METHODS
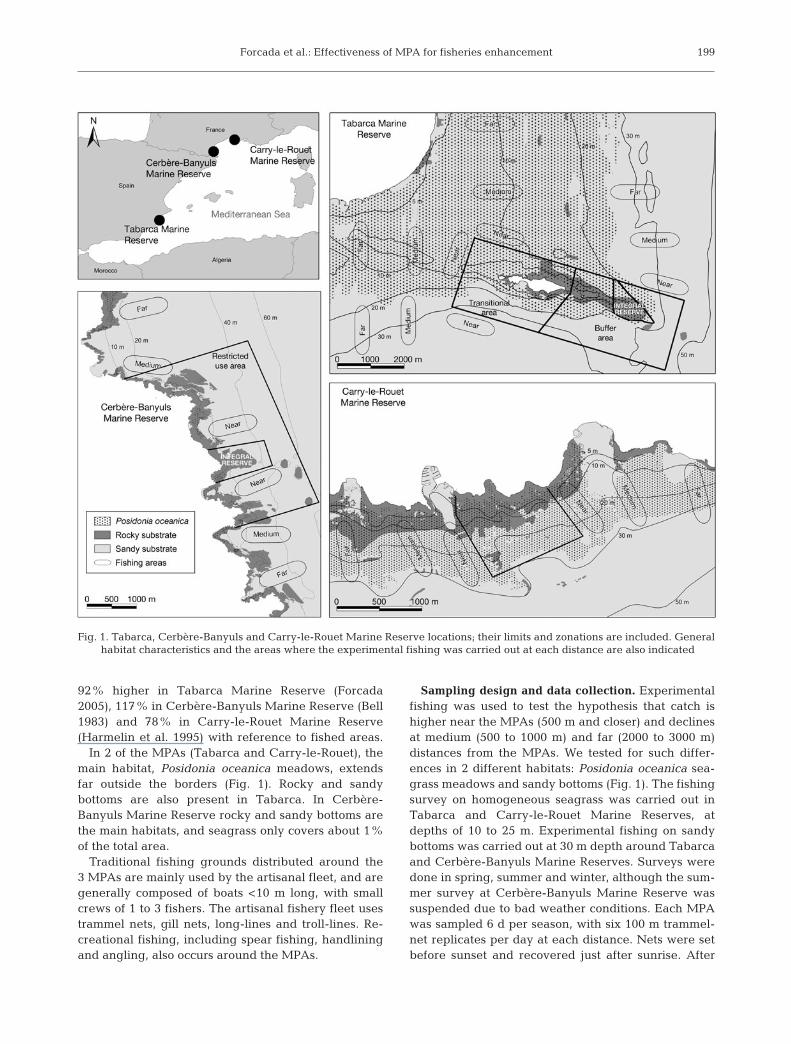
Study area. This study was conducted July 2003 toJune 2005 in 3 MPAs in the western MediterraneanSea, off the coasts of Spain and France, encompassinga geographic range >1000 km (Fig. 1). Tabarca MarineReserve covers 1400 ha, and is substantially largerthan Cerbère-Banyuls Marine Reserve (650 ha) andCarry-le-Rouet Marine Reserve (85 ha). TabarcaMarine Reserve, which is only 4 km from the mainland,contains 3 management zones with different levels ofprotection: the integral reserve area, where all activi-ties except scientific research are forbidden; the bufferarea, in which selective artisanal fishing gear (trapnets that target on pelagic species) is allowed; and thetransitional area, in which the prior selective fishinggear and recreational activities (swimming, diving,mooring of yachts) are permitted. However, the entiremarine reserve acts as a no-take-zone for the speciestargeted by trammel nets, because this gear is bannedinside the MPA. Cerbère-Banyuls Marine Reserve has2 management zones: the integral reserve, where allactivities except scientific surveys are forbidden, andthe restricted use area, in which commercial and re-creational fishing, swimming and diving activities areregulated and spear fishing is forbidden. In contrast,the Carry-le-Rouet Marine Reserve has no zoning andis entirely a no-take marine reserve.
The 3 MPAs have all been established for at least20 yr, and they have yielded evidence of higher bio-mass within their borders. Overall fish abundance was
198
Forcada et al.: Effectiveness of MPA for fisheries enhancement
92% higher in Tabarca Marine Reserve (Forcada2005), 117% in Cerbère-Banyuls Marine Reserve (Bell1983) and 78% in Carry-le-Rouet Marine Reserve(Harmelin et al. 1995) with reference to fished areas.
In 2 of the MPAs (Tabarca and Carry-le-Rouet), themain habitat, Posidonia oceanica meadows, extendsfar outside the borders (Fig. 1). Rocky and sandybottoms are also present in Tabarca. In Cerbère-Banyuls Marine Reserve rocky and sandy bottoms arethe main habitats, and seagrass only covers about 1%of the total area.
Traditional fishing grounds distributed around the3 MPAs are mainly used by the artisanal fleet, and aregenerally composed of boats <10 m long, with smallcrews of 1 to 3 fishers. The artisanal fishery fleet usestrammel nets, gill nets, long-lines and troll-lines. Re-creational fishing, including spear fishing, handliningand angling, also occurs around the MPAs.
Sampling design and data collection. Experimentalfishing was used to test the hypothesis that catch ishigher near the MPAs (500 m and closer) and declinesat medium (500 to 1000 m) and far (2000 to 3000 m)distances from the MPAs. We tested for such differ-ences in 2 different habitats: Posidonia oceanica sea-grass meadows and sandy bottoms (Fig. 1). The fishingsurvey on homogeneous seagrass was carried out inTabarca and Carry-le-Rouet Marine Reserves, atdepths of 10 to 25 m. Experimental fishing on sandybottoms was carried out at 30 m depth around Tabarcaand Cerbère-Banyuls Marine Reserves. Surveys weredone in spring, summer and winter, although the sum-mer survey at Cerbère-Banyuls Marine Reserve wassuspended due to bad weather conditions. Each MPAwas sampled 6 d per season, with six 100 m trammel-net replicates per day at each distance. Nets were setbefore sunset and recovered just after sunrise. After
199
Fig. 1. Tabarca, Cerbère-Banyuls and Carry-le-Rouet Marine Reserve locations; their limits and zonations are included. Generalhabitat characteristics and the areas where the experimental fishing was carried out at each distance are also indicated

Mar Ecol Prog Ser 379: 197–211, 2009
fishing, all captured individuals were identified to spe-cies, and total length and wet weight were recorded.
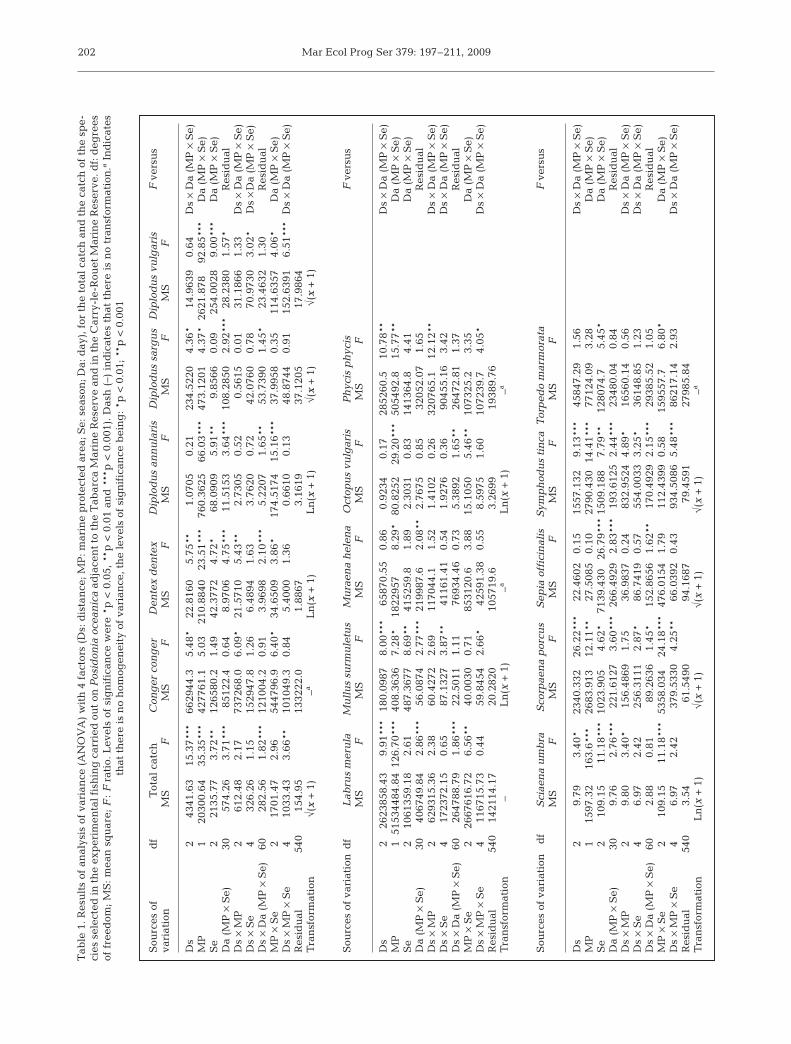
Data analysis. The experimental design was differ-ent for each habitat. On Posidonia oceanica meadowsthe experimental design consisted of 4 factors: Dis-tance (3 levels, fixed), MPA (2 levels, fixed and ortho-gonal), Season (3 levels, fixed and orthogonal) and Day(6 levels, random and nested in the interaction MPA ×Season). Thus, with n = 6 trammel nets, there was atotal of 648 observations.
On sand, the experimental design consisted of 3factors: Distance (3 levels, fixed), Season (3 levels inTabarca Marine Reserve and 2 levels in Cerbère-Banyuls Marine Reserve, fixed and orthogonal) andDay (6 levels, random and nested in Season). There-fore, with n = 6 trammel nets, there was a total of 324observations for the Tabarca Marine Reserve and216 for the Cerbère-Banyuls Marine Reserve.
Analysis of variance (ANOVA) was used to test forsignificant differences in biomass of total catch andof the abundant species (Underwood 1997). When theANOVA F-test was significant, post hoc analyses wereconducted using Student-Newman-Keuls (SNK) mul-tiple comparisons (Underwood 1981). Before analysis,Cochran’s test (Cochran 1951) was used to test forhomogeneity of variance. When significant hete-rogeneity was found, the data were transformed by√(x + 1) or ln(x + 1). When transformations did notremove heterogeneity, analyses were performed onthe untransformed data, but with the F-test α-valueset at 0.01, since ANOVA is robust to departures fromthis assumption, especially when the design is bal-anced and contains a large number of samples or treat-ments (Underwood 1997).
RESULTS
Posidonia oceanica meadows in Tabarca andCarry-le-Rouet Marine Reserves
In the survey, 6619 individuals of 76 species werecaptured (Table A1 in Appendix 1, available at www.int-res.com/articles/suppl/m379p197_app.pdf). Totalweight of the catch was 1373.5 kg. The most commonfamily was Sparidae, followed by Labridae, with 14and 12 species, respectively. Although 57 species werefound in Tabarca Marine Reserve and 56 in Carry-le-Rouet Marine Reserve, the 2 MPAs differed in theircatch composition. Twenty of the species caught atTabarca did not appear at Carry-le-Rouet. In contrast,19 species were caught exclusively at Carry-le-Rouet.
Despite the large number of species caught in bothTabarca and Carry-le-Rouet Marine Reserves, around85% of the catch was represented by 15 species. The
most common species of the total catch in both MPAswas Scorpaena porcus. Other species common to bothMPAs were Labrus merula, Symphodus tinca andSepia officinalis. Some species were only common inone MPA — Sciaena umbra and Dentex dentex inTabarca and Diplodus annularis and Octopus vulgarisin Carry-le-Rouet.
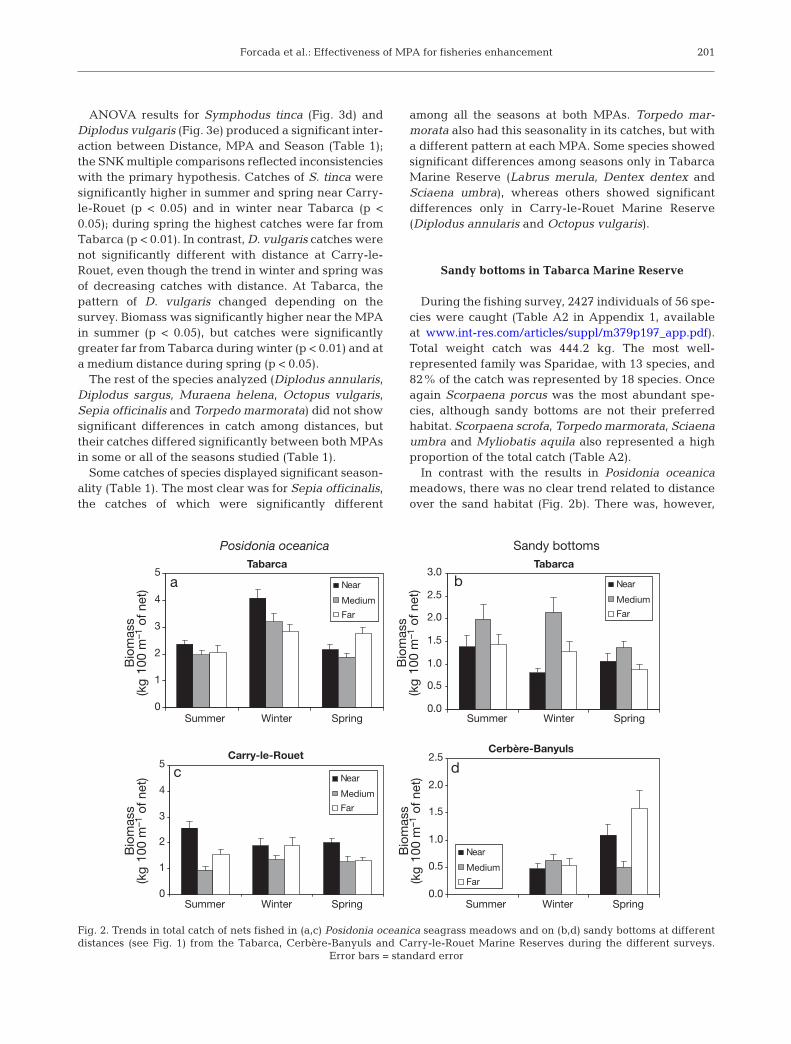
Total mean biomass was greater in Tabarca (2.59 ±0.08 kg per 100 m of net) than in Carry-le-Rouet (1.65 ±0.07 kg per 100 m of net). Differences between dis-tances were greater during winter in Tabarca andduring summer in Carry-le-Rouet (Fig. 2a,c). Highercatches usually occurred near the borders of bothMPAs, but there were exceptions. In spring, total meanbiomass was higher far from the border of Tabarca(Fig. 2a). In Carry-le-Rouet, catches were similar atnear and far distances in winter (Fig. 2c). With respectto seasonality, only Tabarca showed a clear pattern,with the greater values in winter.
In ANOVAs for catch, the 3-way interaction of Dis-tance, MPA and Season was significant (Table 1). InSNK comparisons, catch was significantly higher nearthe border in Tabarca during winter and in Carry-le-Rouet during spring. Moreover, during summer inCarry-le-Rouet, total catch was highest near thereserve.
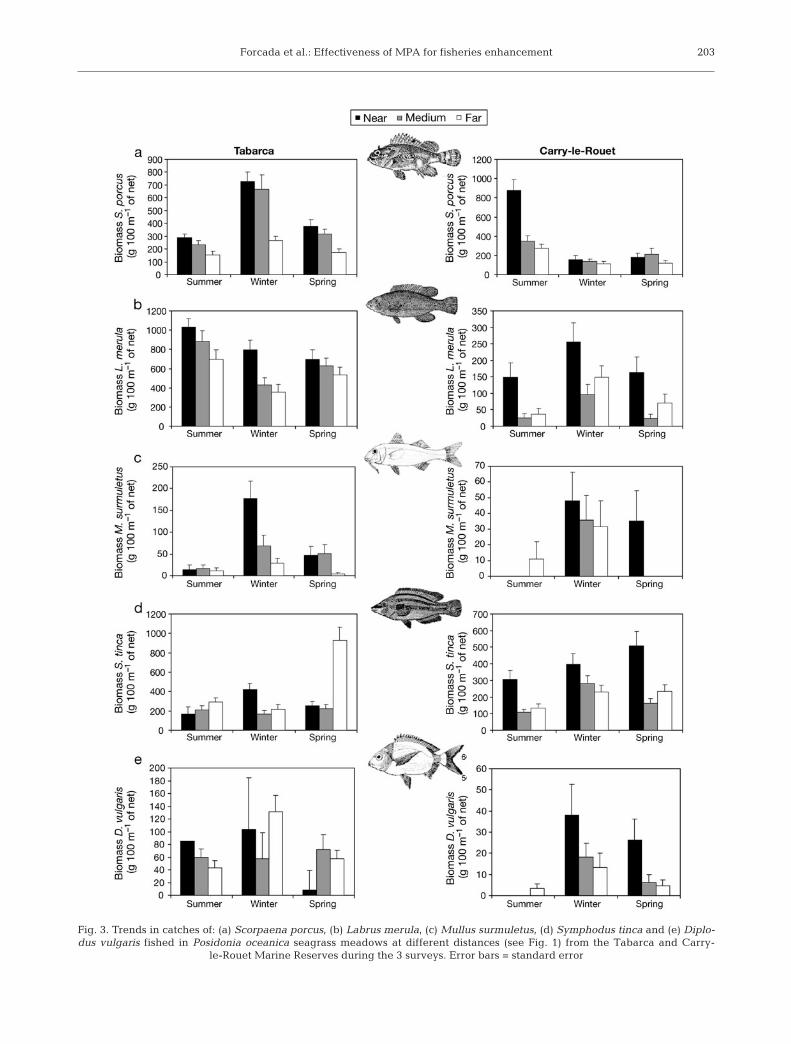
At the species level, catches of Scorpaena porcusdecreased away from the boundaries at both MPAs(Fig. 3a), and the interaction between Distance, MPAand Season was significant (Table 1). SNK test resultsindicated that far from Tabarca catches were lowest.However in Carry-le-Rouet, higher catches wereobtained near the MPA border only in summer (p <0.01). Labrus merula also had a decreasing trend withdistance at both MPAs (Fig. 3b), but catches were onlysignificantly higher close to Tabarca. Furthermore,catches of this species were significantly higher inTabarca than in Carry-le-Rouet during all seasons (p <0.01). Catches of Mullus surmuletus declined withdistance in both MPAs, but only in winter (Fig. 3c).However, these differences were significant only inTabarca, with higher catches near the boundaries (p <0.01).
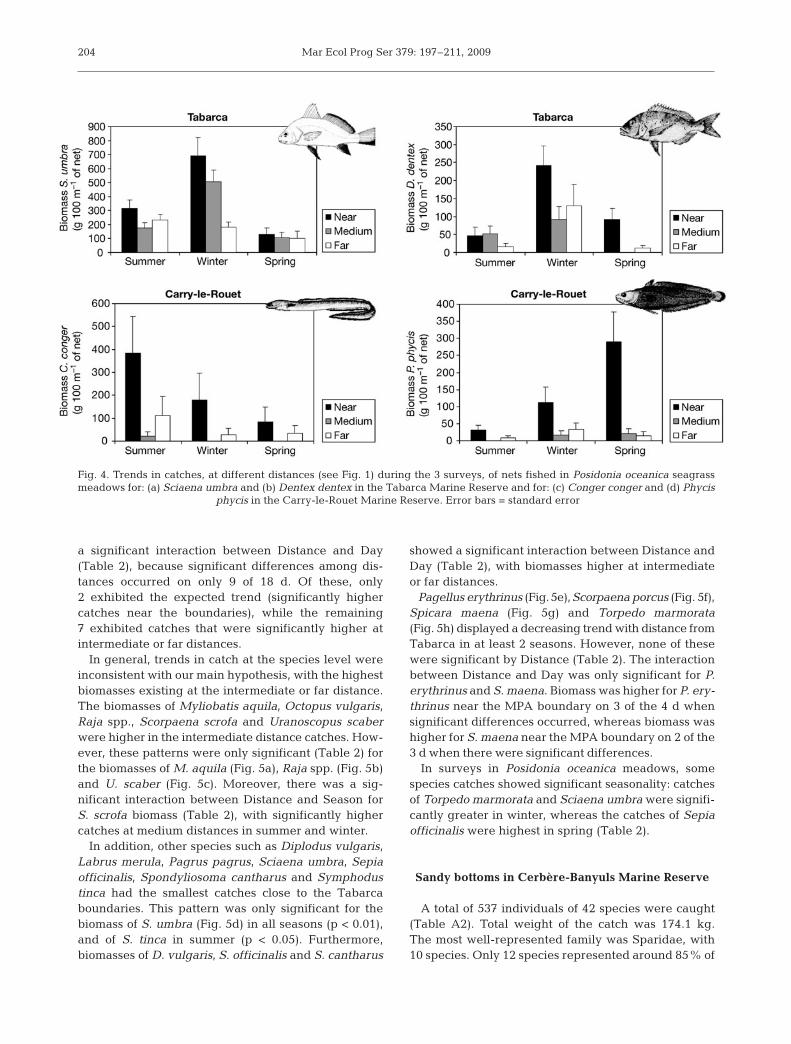
Other important species, such as Sciaena umbra(Fig. 4a) and Dentex dentex (Fig. 4b), showed catchesdecreasing with distance, but only in Tabarca MarineReserve (p < 0.01); these species were rare or absentat the other MPAs. In contrast, decreasing trendswith distance of Conger conger (Fig. 4c) and Phycisphycis (Fig. 4d) were observed only in Carry-le-Rouet;catches around Tabarca were low. C. conger wasalways caught more frequently (p < 0.05) close toCarry-le-Rouet boundaries, while catches of P. phyciswere significantly higher (p < 0.01) only in winter andspring.
200
Forcada et al.: Effectiveness of MPA for fisheries enhancement
ANOVA results for Symphodus tinca (Fig. 3d) andDiplodus vulgaris (Fig. 3e) produced a significant inter-action between Distance, MPA and Season (Table 1);the SNK multiple comparisons reflected inconsistencieswith the primary hypothesis. Catches of S. tinca weresignificantly higher in summer and spring near Carry-le-Rouet (p < 0.05) and in winter near Tabarca (p <0.05); during spring the highest catches were far fromTabarca (p < 0.01). In contrast, D. vulgaris catches werenot significantly different with distance at Carry-le-Rouet, even though the trend in winter and spring wasof decreasing catches with distance. At Tabarca, thepattern of D. vulgaris changed depending on thesurvey. Biomass was significantly higher near the MPAin summer (p < 0.05), but catches were significantlygreater far from Tabarca during winter (p < 0.01) and ata medium distance during spring (p < 0.05).
The rest of the species analyzed (Diplodus annularis,Diplodus sargus, Muraena helena, Octopus vulgaris,Sepia officinalis and Torpedo marmorata) did not showsignificant differences in catch among distances, buttheir catches differed significantly between both MPAsin some or all of the seasons studied (Table 1).
Some catches of species displayed significant season-ality (Table 1). The most clear was for Sepia officinalis,the catches of which were significantly different
among all the seasons at both MPAs. Torpedo mar-morata also had this seasonality in its catches, but witha different pattern at each MPA. Some species showedsignificant differences among seasons only in TabarcaMarine Reserve (Labrus merula, Dentex dentex andSciaena umbra), whereas others showed significantdifferences only in Carry-le-Rouet Marine Reserve(Diplodus annularis and Octopus vulgaris).
Sandy bottoms in Tabarca Marine Reserve
During the fishing survey, 2427 individuals of 56 spe-cies were caught (Table A2 in Appendix 1, availableat www.int-res.com/articles/suppl/m379p197_app.pdf).Total weight catch was 444.2 kg. The most well-represented family was Sparidae, with 13 species, and82% of the catch was represented by 18 species. Onceagain Scorpaena porcus was the most abundant spe-cies, although sandy bottoms are not their preferredhabitat. Scorpaena scrofa, Torpedo marmorata, Sciaenaumbra and Myliobatis aquila also represented a highproportion of the total catch (Table A2).
In contrast with the results in Posidonia oceanicameadows, there was no clear trend related to distanceover the sand habitat (Fig. 2b). There was, however,
201
Posidonia oceanica
Sandy bottoms Tabarca Tabarca
0
1
2
3
4
5
Bio
mas
s(k
g 1
00 m
–1 o
f net
)
Bio
mas
s(k
g 1
00 m
–1 o
f net
)
Bio
mas
s(k
g 1
00 m
–1 o
f net
)
Bio
mas
s(k
g 1
00 m
–1 o
f net
)
Near
Medium
Far
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Carry-le-Rouet
0
1
2
3
4
5Cerbère-Banyuls
0.0
0.5
1.0
1.5
2.0
2.5
Summer Winter Spring
Summer Winter Spring Summer Winter Spring
Summer Winter Spring
Near
Medium
Far
Near
Medium
Far
Near
Medium
Far
a b
c d
Fig. 2. Trends in total catch of nets fished in (a,c) Posidonia oceanica seagrass meadows and on (b,d) sandy bottoms at differentdistances (see Fig. 1) from the Tabarca, Cerbère-Banyuls and Carry-le-Rouet Marine Reserves during the different surveys.
Error bars = standard error
Mar Ecol Prog Ser 379: 197–211, 2009202
Tab
le 1
. Res
ult
s of
an
alys
is o
f va
rian
ce (
AN
OV
A)
wit
h 4
fac
tors
(D
s: d
ista
nce
; MP
: mar
ine
pro
tect
ed a
rea;
Se:
sea
son
; Da:
day
), f
or t
he
tota
l ca
tch
an
d t
he
catc
h o
f th
e sp
e-ci
es s
elec
ted
in t
he
exp
erim
enta
l fis
hin
g c
arri
ed o
ut
on P
osid
onia
oce
anic
aad
jace
nt
to t
he
Tab
arca
Mar
ine
Res
erve
an
d in
th
e C
arry
-le-
Rou
et M
arin
e R
eser
ve. d
f: d
egre
esof
fre
edom
; M
S:
mea
n s
qu
are;
F:
Fra
tio.
Lev
els
of s
ign
ific
ance
wer
e *p
< 0
.05,
**p
< 0
.01
and
***
p <
0.0
01).
Das
h (
–) i
nd
icat
es t
hat
th
ere
is n
o tr
ansf
orm
atio
n.a
Ind
icat
es
that
th
ere
is n
o h
omog
enei
ty o
f va
rian
ce, t
he
leve
ls o
f si
gn
ific
ance
bei
ng
: *p
< 0
.01;
**p
< 0
.001
Sou
rces
of
df
Tot
al c
atch
Con
ger
con
ger
Den
tex
den
tex
Dip
lod
us
ann
ula
ris
Dip
lod
us
sarg
us
Dip
lod
us
vulg
aris
Fve
rsu
sva
riat
ion
M
SF
MS
FM
SF
MS
FM
SF
MS
F
Ds
243
41.6
315
.37*
**66
2944
.35.
48*
22.8
160
5.75
**1.
0705
0.21
234.
5220
4.36
*14
.963
90.
64D
s ×
Da
(MP
×S
e)M
P1
2030
0.64
35.3
5***
4277
61.1
5.03
210.
8840
23.5
1***
760.
3625
66.0
3***
473.
1201
4.37
*26
21.8
7892
.85*
**D
a (M
P ×
Se)
Se
221
35.7
73.
72**
1265
80.2
1.49
42.3
772
4.72
*68
.090
95.
91**
9.85
660.
0925
4.00
289.
00**
*D
a (M
P ×
Se)
Da
(MP
×S
e)30
574.
263.
71**
*85
122.
40.
648.
9706
4.75
***
11.5
153
3.64
***
108.
2850
2.92
***
28.2
380
1.57
*R
esid
ual
Ds
×M
P2
612.
482.
1773
7268
.06.
09*
21.5
710
5.43
**2.
7305
0.52
0.56
150.
0131
.186
61.
33D
s ×
Da
(MP
×S
e)D
s ×
Se
432
6.26
1.15
1529
47.8
1.26
6.48
941.
633.
7620
0.72
42.0
760
0.78
70.9
730
3.02
*D
s ×
Da
(MP
×S
e)D
s ×
Da
(MP
×S
e)60
282.
561.
82**
*12
1004
.20.
913.
9698
2.10
***
5.22
071.
65**
53.7
390
1.45
*23
.463
21.
30R
esid
ual
MP
×S
e2
1701
.47
2.96
5447
96.9
6.40
*34
.650
93.
86*
174.
5174
15.1
6***
37.9
958
0.35
114.
6357
4.06
*D
a (M
P ×
Se)
Ds
×M
P ×
Se
410
33.4
33.
66**
1010
49.3
0.84
5.40
001.
360.
6610
0.13
48.8
744
0.91
152.
6391
6.51
***
Ds
×D
a (M
P ×
Se)
Res
idu
al54
015
4.95
1332
22.0
1.88
673.
1619
37.1
205
17.9
864
Tra
nsf
orm
atio
n√(
x+
1)
–aL
n(x
+ 1
)L
n(x
+ 1
)√(
x+
1)
√(x
+ 1
)
Sou
rces
of
vari
atio
nd
fL
abru
s m
eru
laM
ull
us
surm
ule
tus
Mu
raen
a h
elen
a
Oct
opu
s vu
lgar
isP
hyc
is p
hyc
isF
vers
us
MS
FM
SF
MS
FM
SF
MS
F
Ds
226
2385
8.43
9.91
***
180.
0987
8.00
***
6587
0.55
0.86
0.92
340.
1728
5260
.510
.78*
*D
s ×
Da
(MP
×S
e)M
P1
5153
4484
.84
126.
70**
*40
8.36
367.
28*
1822
957
8.29
*80
.825
229
.20*
**50
5492
.815
.77*
*D
a (M
P ×
Se)
Se
210
6135
9.18
2.61
487.
3677
8.69
**41
5259
.81.
892.
3031
0.83
1413
64.8
4.41
Da
(MP
×S
e)D
a (M
P ×
Se)
3040
6749
.84
2.86
***
56.0
874
2.77
***
2199
87.6
2.08
**2.
7675
0.85
3205
2.07
1.65
Res
idu
alD
s ×
MP
262
9315
.36
2.38
60.4
272
2.69
1170
44.1
1.52
1.41
020.
2632
0765
.112
.12*
*D
s ×
Da
(MP
×S
e)D
s ×
Se
417
2372
.15
0.65
87.1
327
3.87
**41
161.
410.
541.
9276
0.36
9045
5.16
3.42
Ds
×D
a (M
P ×
Se)
Ds
×D
a (M
P ×
Se)
6026
4788
.79
1.86
***
22.5
011
1.11
7693
4.46
0.73
5.38
921.
65**
2647
2.81
1.37
Res
idu
alM
P ×
Se
226
6761
6.72
6.56
**40
.003
00.
7185
3120
.63.
8815
.105
05.
46**
1073
25.2
3.35
Da
(MP
×S
e)D
s ×
MP
×S
e4
1167
15.7
30.
4459
.845
42.
66*
4259
1.38
0.55
8.59
751.
6010
7239
.74.
05*
Ds
×D
a (M
P ×
Se)
Res
idu
al54
014
2114
.17
20.2
820
1057
19.6
3.26
9919
389.
76T
ran
sfor
mat
ion
–L
n(x
+ 1
)–a
Ln
(x+
1)
–a
Sou
rces
of
vari
atio
nd
f
S
ciae
na
um
bra
Sco
rpae
na
por
cus
Sep
ia o
ffic
inal
isS
ymp
hod
us
tin
caT
orp
edo
mar
mor
ata
Fve
rsu
sM
SF
MS
FM
SF
MS
FM
SF
Ds
29.
793.
40*
2340
.332
26.2
2***
22.4
602
0.15
1557
.132
9.13
***
4584
7.29
1.56
Ds
×D
a (M
P ×
Se)
MP
115
97.3
216
3.6*
**26
83.9
1312
.11*
*27
.508
50.
1027
90.4
3014
.41*
**77
124.
093.
28D
a (M
P ×
Se)
Se
210
9.15
11.1
8***
1023
.905
4.62
*71
39.4
3026
.79*
**15
09.1
887.
79**
1280
74.7
5.45
*D
a (M
P ×
Se)
Da
(MP
×S
e)30
9.76
2.76
***
221.
6127
3.60
***
266.
4929
2.83
***
193.
6125
2.44
***
2348
0.04
0.84
Res
idu
alD
s ×
MP
29.
803.
40*
156.
4869
1.75
36.9
837
0.24
832.
9524
4.89
*16
560.
140.
56D
s ×
Da
(MP
×S
e)D
s ×
Se
46.
972.
4225
6.31
112.
87*
86.7
419
0.57
554.
0033
3.25
*36
148.
851.
23D
s ×
Da
(MP
×S
e)D
s ×
Da
(MP
×S
e)60
2.88
0.81
89.2
636
1.45
*15
2.86
561.
62**
170.
4929
2.15
***
2938
5.52
1.05
Res
idu
alM
P ×
Se
210
9.15
11.1
8***
5358
.034
24.1
8***
476.
0154
1.79
112.
4399
0.58
1595
57.7
6.80
*D
a (M
P ×
Se)
Ds
×M
P ×
Se
46.
972.
4237
9.53
304.
25**
66.0
392
0.43
934.
5086
5.48
***
8621
7.14
2.93
Ds
×D
a (M
P ×
Se)
Res
idu
al54
03.
5461
.549
094
.168
779
.459
127
985.
84T
ran
sfor
mat
ion
Ln
(x+
1)
√(x
+ 1
)√(
x+
1)
√(x
+ 1
)–a
Forcada et al.: Effectiveness of MPA for fisheries enhancement 203
Fig. 3. Trends in catches of: (a) Scorpaena porcus, (b) Labrus merula, (c) Mullus surmuletus, (d) Symphodus tinca and (e) Diplo-dus vulgaris fished in Posidonia oceanica seagrass meadows at different distances (see Fig. 1) from the Tabarca and Carry-
le-Rouet Marine Reserves during the 3 surveys. Error bars = standard error
Mar Ecol Prog Ser 379: 197–211, 2009
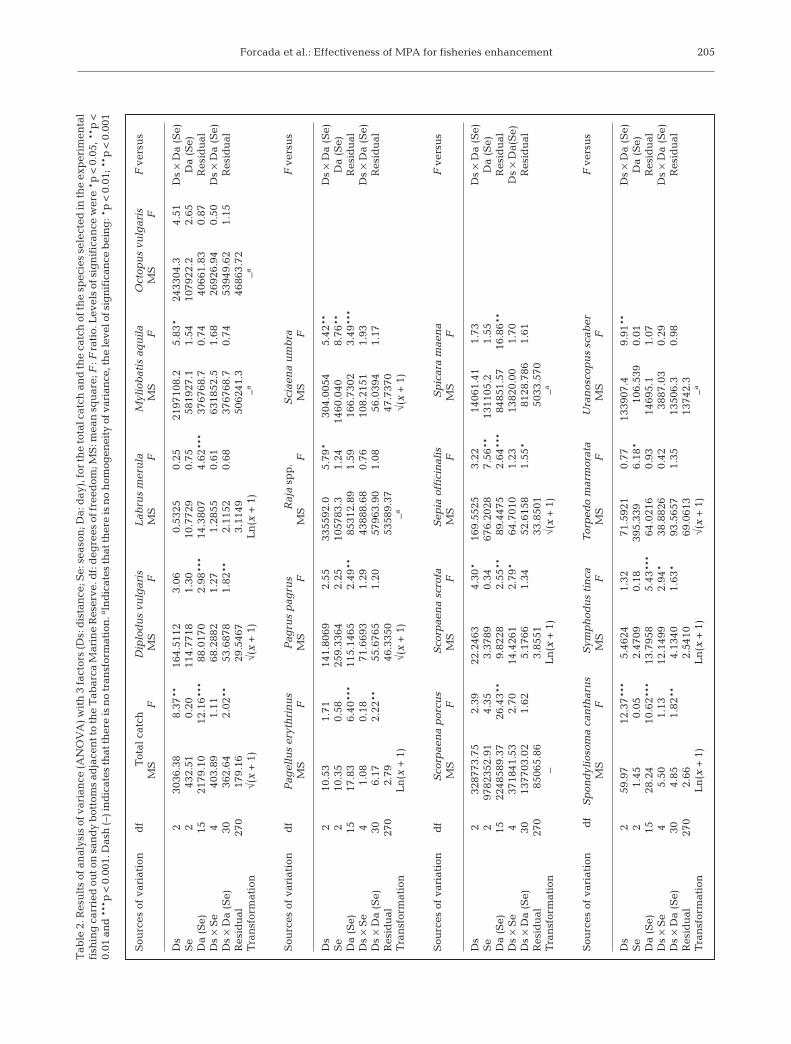
a significant interaction between Distance and Day(Table 2), because significant differences among dis-tances occurred on only 9 of 18 d. Of these, only2 exhibited the expected trend (significantly highercatches near the boundaries), while the remaining7 exhibited catches that were significantly higher atintermediate or far distances.
In general, trends in catch at the species level wereinconsistent with our main hypothesis, with the highestbiomasses existing at the intermediate or far distance.The biomasses of Myliobatis aquila, Octopus vulgaris,Raja spp., Scorpaena scrofa and Uranoscopus scaberwere higher in the intermediate distance catches. How-ever, these patterns were only significant (Table 2) forthe biomasses of M. aquila (Fig. 5a), Raja spp. (Fig. 5b)and U. scaber (Fig. 5c). Moreover, there was a sig-nificant interaction between Distance and Season forS. scrofa biomass (Table 2), with significantly highercatches at medium distances in summer and winter.
In addition, other species such as Diplodus vulgaris,Labrus merula, Pagrus pagrus, Sciaena umbra, Sepiaofficinalis, Spondyliosoma cantharus and Symphodustinca had the smallest catches close to the Tabarcaboundaries. This pattern was only significant for thebiomass of S. umbra (Fig. 5d) in all seasons (p < 0.01),and of S. tinca in summer (p < 0.05). Furthermore,biomasses of D. vulgaris, S. officinalis and S. cantharus
showed a significant interaction between Distance andDay (Table 2), with biomasses higher at intermediateor far distances.
Pagellus erythrinus (Fig.5e),Scorpaena porcus (Fig. 5f),Spicara maena (Fig. 5g) and Torpedo marmorata(Fig. 5h) displayed a decreasing trend with distance fromTabarca in at least 2 seasons. However, none of thesewere significant by Distance (Table 2). The interactionbetween Distance and Day was only significant for P.erythrinus and S. maena. Biomass was higher for P. ery-thrinus near the MPA boundary on 3 of the 4 d whensignificant differences occurred, whereas biomass washigher for S. maena near the MPA boundary on 2 of the3 d when there were significant differences.
In surveys in Posidonia oceanica meadows, somespecies catches showed significant seasonality: catchesof Torpedo marmorata and Sciaena umbra were signifi-cantly greater in winter, whereas the catches of Sepiaofficinalis were highest in spring (Table 2).
Sandy bottoms in Cerbère-Banyuls Marine Reserve
A total of 537 individuals of 42 species were caught(Table A2). Total weight of the catch was 174.1 kg.The most well-represented family was Sparidae, with10 species. Only 12 species represented around 85% of
204
Fig. 4. Trends in catches, at different distances (see Fig. 1) during the 3 surveys, of nets fished in Posidonia oceanica seagrassmeadows for: (a) Sciaena umbra and (b) Dentex dentex in the Tabarca Marine Reserve and for: (c) Conger conger and (d) Phycis
phycis in the Carry-le-Rouet Marine Reserve. Error bars = standard error
Forcada et al.: Effectiveness of MPA for fisheries enhancement 205
Tab
le 2
. Res
ult
s of
an
alys
is o
f var
ian
ce (A
NO
VA
) wit
h 3
fact
ors
(Ds:
dis
tan
ce; S
e: s
easo
n; D
a: d
ay),
for
the
tota
l cat
ch a
nd
the
catc
h o
f th
e sp
ecie
s se
lect
ed in
the
exp
erim
enta
lfi
shin
g c
arri
ed o
ut o
n s
and
y b
otto
ms
adja
cen
t to
the
Tab
arca
Mar
ine
Res
erve
. df:
deg
rees
of f
reed
om; M
S: m
ean
sq
uar
e; F
: Fra
tio.
Lev
els
of s
ign
ific
ance
wer
e *p
<0.
05, *
*p<
0.01
an
d *
**p
< 0
.001
. Das
h (–
) in
dic
ates
that
ther
e is
no
tran
sfor
mat
ion
. a In
dic
ates
that
ther
e is
no
hom
ogen
eity
of v
aria
nce
, th
e le
vel o
f sig
nif
ican
ce b
ein
g: *
p <
0.0
1; *
*p <
0.0
01
Sou
rces
of
vari
atio
nd
fT
otal
cat
chD
iplo
du
s vu
lgar
isL
abru
s m
eru
laM
ylio
bat
is a
qu
ila
Oct
opu
s vu
lgar
isF
vers
us
MS
FM
SF
MS
FM
SF
MS
F
Ds
230
36.3
88.
37**
164.
5112
3.06
0.53
250.
2521
9710
8.2
5.83
*24
3304
.34.
51D
s ×
Da
(Se)
Se
243
2.51
0.20
114.
7718
1.30
10.7
729
0.75
5819
27.1
1.54
1079
22.2
2.65
Da
(Se)
Da
(Se)
1521
79.1
012
.16*
**88
.017
02.
98**
*14
.380
74.
62**
*37
6768
.70.
7440
661.
830.
87R
esid
ual
Ds
×S
e4
403.
891.
1168
.288
21.
271.
2855
0.61
6318
52.5
1.68
2692
6.94
0.50
Ds
×D
a (S
e)D
s ×
Da
(Se)
3036
2.64
2.02
**53
.687
81.
82**
2.11
520.
6837
6768
.70.
7453
949.
621.
15R
esid
ual
Res
idu
al27
017
9.16
29.5
467
3.11
4950
6241
.346
863.
72T
ran
sfor
mat
ion
√(x
+ 1
)√(
x+
1)
Ln
(x+
1)
–a–a
Sou
rces
of
vari
atio
nd
fP
agel
lus
eryt
hri
nu
sP
agru
s p
agru
sR
aja
spp
.S
ciae
na
um
bra
Fve
rsu
sM
SF
MS
FM
SF
MS
F
Ds
210
.53
1.71
141.
8069
2.55
3355
92.0
5.79
*30
4.00
545.
42**
Ds
×D
a (S
e)S
e2
10.3
50.
5825
9.33
642.
2510
5783
.31.
2414
60.0
408.
76**
Da
(Se)
Da
(Se)
1517
.83
6.40
***
115.
1465
2.49
**85
312.
891.
5916
6.73
023.
49**
*R
esid
ual
Ds
×S
e4
1.08
0.18
71.6
693
1.29
4388
8.68
0.76
108.
2151
1.93
Ds
×D
a (S
e)D
s ×
Da
(Se)
306.
172.
22**
55.6
765
1.20
5796
3.90
1.08
56.0
394
1.17
Res
idu
alR
esid
ual
270
2.79
46.3
350
5358
9.37
47.7
370
Tra
nsf
orm
atio
nL
n(x
+ 1
)√(
x+
1)
–a√(
x+
1)
Sou
rces
of
vari
atio
nd
fS
corp
aen
a p
orcu
sS
corp
aen
a sc
rofa
Sep
ia o
ffic
inal
isS
pic
ara
mae
na
Fve
rsu
sM
SF
MS
FM
SF
MS
F
Ds
232
8773
.75
2.39
22.2
463
4.30
*16
9.55
253.
2214
061.
411.
73D
s ×
Da
(Se)
Se
297
8235
2.91
4.35
3.37
890.
3467
6.20
287.
56**
1311
05.2
1.55
Da
(Se)
Da
(Se)
1522
4858
9.37
26.4
3**
9.82
282.
55**
89.4
475
2.64
***
8485
1.57
16.8
6**
Res
idu
alD
s ×
Se
437
1841
.53
2.70
14.4
261
2.79
*64
.701
01.
2313
820.
001.
70D
s ×
Da(
Se)
Ds
×D
a (S
e)30
1377
03.0
21.
625.
1766
1.34
52.6
158
1.55
*81
28.7
861.
61R
esid
ual
Res
idu
al27
085
065.
863.
8551
33.8
501
5033
.570
Tra
nsf
orm
atio
n–
Ln
(x+
1)
√(x
+ 1
)–a
Sou
rces
of
vari
atio
nd
f
Sp
ond
ylio
som
a ca
nth
aru
sS
ymp
hod
us
tin
caT
orp
edo
mar
mor
ata
Ura
nos
cop
us
scab
erF
vers
us
MS
FM
SF
MS
FM
SF
Ds
259
.97
12.3
7***
5.46
241.
3271
.592
10.
7713
3907
.49.
91**
Ds
×D
a (S
e)S
e2
1.45
0.05
2.47
090.
1839
5.33
96.
18*
106.
539
0.01
Da
(Se)
Da
(Se)
1528
.24
10.6
2***
13.7
958
5.43
***
64.0
216
0.93
1469
5.1
1.07
Res
idu
alD
s ×
Se
45.
501.
1312
.149
92.
94*
38.8
826
0.42
3887
.03
0.29
Ds
×D
a (S
e)D
s ×
Da
(Se)
304.
851.
82**
4.13
401.
63*
93.5
657
1.35
1350
6.3
0.98
Res
idu
alR
esid
ual
270
2.66
2.54
1069
.061
313
742.
3T
ran
sfor
mat
ion
Ln
(x+
1)
Ln
(x+
1)
√(x
+ 1
)–a
Mar Ecol Prog Ser 379: 197–211, 2009
the catches, with the main species being Phycis phycisand Pagellus acarne. Other important species in thetotal catch were Scorpaena scrofa, Scorpaena porcusand Sepia officinalis.
As with the results for sandy bottoms in Tabarca, therewas no clear trend with distance in total catch (Fig. 2d),though there was a significant interaction between Dis-tance and Day (Table 3). On only 2 of the 12 samplingdays were there significant differences among distances.Season was periodically important (Table 3); total bio-mass was significantly greater in spring than in winter.
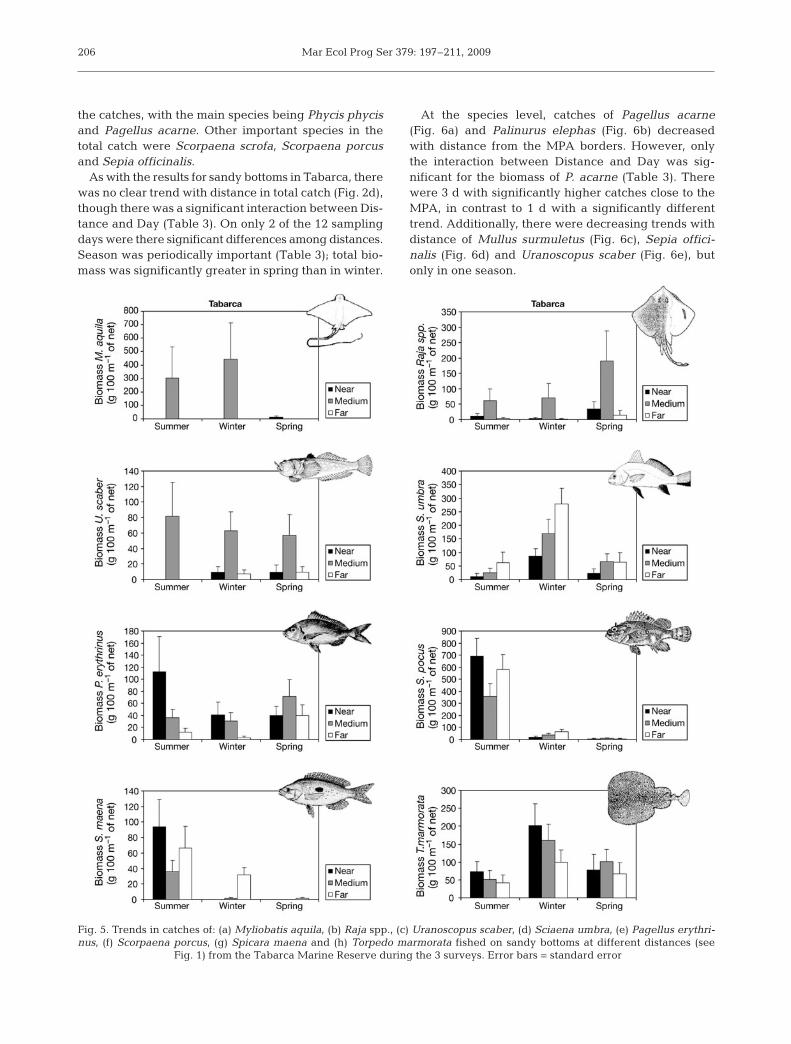
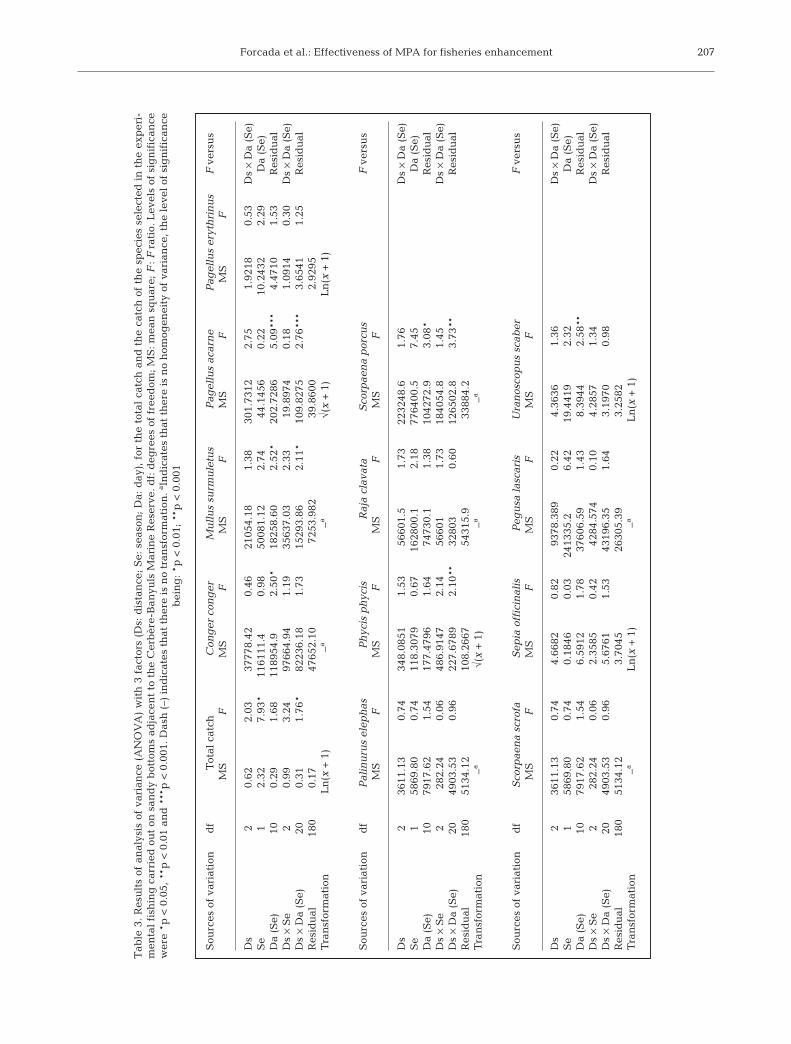
At the species level, catches of Pagellus acarne(Fig. 6a) and Palinurus elephas (Fig. 6b) decreasedwith distance from the MPA borders. However, onlythe interaction between Distance and Day was sig-nificant for the biomass of P. acarne (Table 3). Therewere 3 d with significantly higher catches close to theMPA, in contrast to 1 d with a significantly differenttrend. Additionally, there were decreasing trends withdistance of Mullus surmuletus (Fig. 6c), Sepia offici-nalis (Fig. 6d) and Uranoscopus scaber (Fig. 6e), butonly in one season.
206
Fig. 5. Trends in catches of: (a) Myliobatis aquila, (b) Raja spp., (c) Uranoscopus scaber, (d) Sciaena umbra, (e) Pagellus erythri-nus, (f) Scorpaena porcus, (g) Spicara maena and (h) Torpedo marmorata fished on sandy bottoms at different distances (see
Fig. 1) from the Tabarca Marine Reserve during the 3 surveys. Error bars = standard error
Forcada et al.: Effectiveness of MPA for fisheries enhancement 207
Tab
le 3
. R
esu
lts
of a
nal
ysis
of
vari
ance
(A
NO
VA
) w
ith
3 f
acto
rs (
Ds:
dis
tan
ce;
Se:
sea
son
; D
a: d
ay),
for
th
e to
tal
catc
h a
nd
th
e ca
tch
of
the
spec
ies
sele
cted
in
th
e ex
per
i-m
enta
l fi
shin
g c
arri
ed o
ut
on s
and
y b
otto
ms
adja
cen
t to
th
e C
erb
ère-
Ban
yuls
Mar
ine
Res
erve
. df:
deg
rees
of
free
dom
; MS
: mea
n s
qu
are;
F: F
rati
o. L
evel
s of
sig
nif
ican
cew
ere
*p <
0.0
5, *
*p <
0.0
1 an
d *
**p
< 0
.001
. Das
h (
–) i
nd
icat
es t
hat
th
ere
is n
o tr
ansf
orm
atio
n. a I
nd
icat
es t
hat
th
ere
is n
o h
omog
enei
ty o
f va
rian
ce, t
he
leve
l of
sig
nif
ican
ce
bei
ng
: *p
< 0
.01;
**p
< 0
.001
Sou
rces
of
vari
atio
nd
fT
otal
cat
chC
ong
er c
ong
erM
ull
us
surm
ule
tus
Pag
ellu
s ac
arn
eP
agel
lus
eryt
hri
nu
sF
vers
us
MS
FM
SF
MS
FM
SF
MS
F
Ds
20.
622.
0337
778.
420.
4621
054.
181.
3830
1.73
122.
751.
9218
0.53
Ds
×D
a (S
e)S
e1
2.32
7.93
*11
6111
.40.
9850
081.
122.
7444
.145
60.
2210
.243
22.
29D
a (S
e)D
a (S
e)10
0.29
1.68
1189
54.9
2.50
*18
258.
602.
52*
202.
7286
5.09
***
4.47
101.
53R
esid
ual
Ds
×S
e2
0.99
3.24
9766
4.94
1.19
3563
7.03
2.33
19.8
974
0.18
1.09
140.
30D
s ×
Da
(Se)
Ds
×D
a (S
e)20
0.31
1.76
*82
236.
181.
7315
293.
862.
11*
109.
8275
2.76
***
3.65
411.
25R
esid
ual
Res
idu
al18
00.
1747
652.
1072
53.9
8239
.860
02.
9295
Tra
nsf
orm
atio
nL
n(x
+ 1
)–a
–a√(
x+
1)
Ln
(x+
1)
Sou
rces
of
vari
atio
nd
fP
alin
uru
s el
eph
asP
hyc
is p
hyc
isR
aja
clav
ata
Sco
rpae
na
por
cus
Fve
rsu
sM
SF
MS
FM
SF
MS
F
Ds
236
11.1
30.
7434
8.08
511.
5356
601.
51.
7322
3248
.61.
76D
s ×
Da
(Se)
Se
158
69.8
00.
7411
8.30
790.
6716
2800
.12.
1877
6400
.57.
45D
a (S
e)D
a (S
e)10
7917
.62
1.54
177.
4796
1.64
7473
0.1
1.38
1042
72.9
3.08
*R
esid
ual
Ds
×S
e2
282.
240.
0648
6.91
472.
1456
601
1.73
1840
54.8
1.45
Ds
×D
a (S
e)D
s ×
Da
(Se)
2049
03.5
30.
9622
7.67
892.
10**
3280
30.
6012
6502
.83.
73**
Res
idu
alR
esid
ual
180
5134
.12
108.
2667
5431
5.9
3388
4.2
Tra
nsf
orm
atio
n–a
√(x
+ 1
)–a
–a
Sou
rces
of
vari
atio
nd
fS
corp
aen
a sc
rofa
Sep
ia o
ffic
inal
isP
egu
sa l
asca
ris
Ura
nos
cop
us
scab
erF
vers
us
MS
FM
SF
MS
FM
SF
Ds
236
11.1
30.
744.
6682
0.82
9378
.389
0.22
4.36
361.
36D
s ×
Da
(Se)
Se
158
69.8
00.
740.
1846
0.03
2413
35.2
6.42
19.4
419
2.32
Da
(Se)
Da
(Se)
1079
17.6
21.
546.
5912
1.78
3760
6.59
1.43
8.39
442.
58**
Res
idu
alD
s ×
Se
228
2.24
0.06
2.35
850.
4242
84.5
740.
104.
2857
1.34
Ds
×D
a (S
e)D
s ×
Da
(Se)
2049
03.5
30.
965.
6761
1.53
4319
6.35
1.64
3.19
700.
98R
esid
ual
Res
idu
al18
051
34.1
23.
7045
2630
5.39
3.25
82T
ran
sfor
mat
ion
–aL
n(x
+ 1
)–a
Ln
(x+
1)
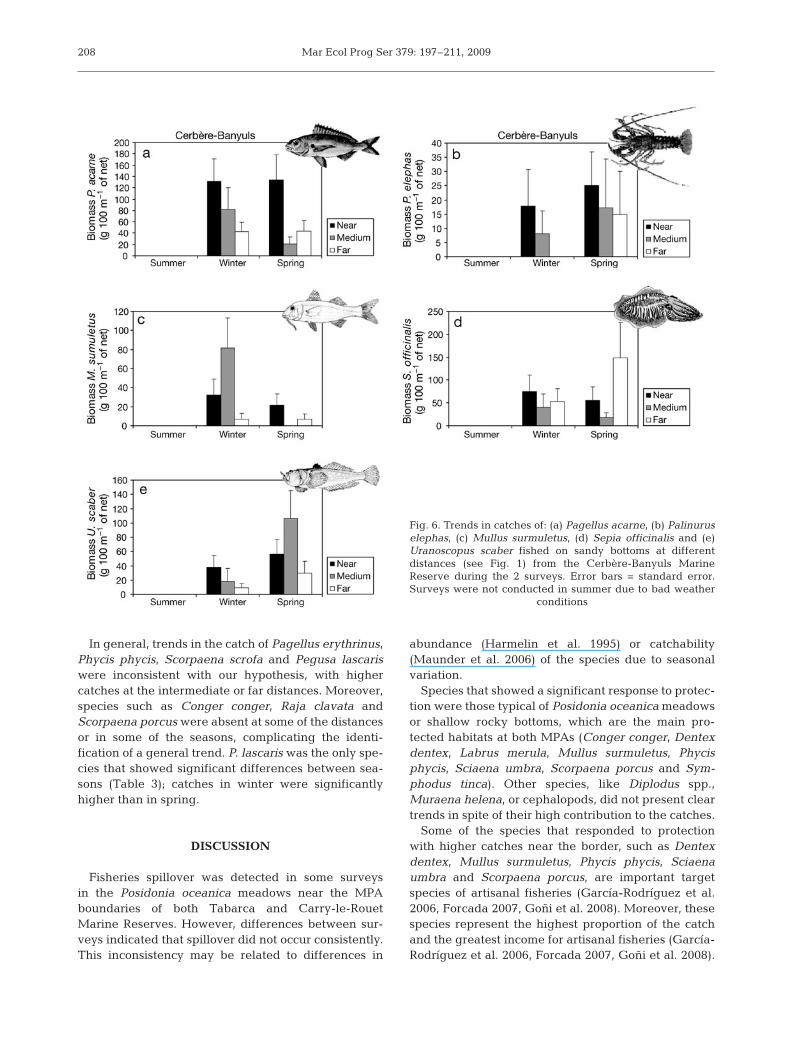
Mar Ecol Prog Ser 379: 197–211, 2009
In general, trends in the catch of Pagellus erythrinus,Phycis phycis, Scorpaena scrofa and Pegusa lascariswere inconsistent with our hypothesis, with highercatches at the intermediate or far distances. Moreover,species such as Conger conger, Raja clavata andScorpaena porcus were absent at some of the distancesor in some of the seasons, complicating the identi-fication of a general trend. P. lascaris was the only spe-cies that showed significant differences between sea-sons (Table 3); catches in winter were significantlyhigher than in spring.
DISCUSSION
Fisheries spillover was detected in some surveysin the Posidonia oceanica meadows near the MPAboundaries of both Tabarca and Carry-le-RouetMarine Reserves. However, differences between sur-veys indicated that spillover did not occur consistently.This inconsistency may be related to differences in
abundance (Harmelin et al. 1995) or catchability(Maunder et al. 2006) of the species due to seasonalvariation.
Species that showed a significant response to protec-tion were those typical of Posidonia oceanica meadowsor shallow rocky bottoms, which are the main pro-tected habitats at both MPAs (Conger conger, Dentexdentex, Labrus merula, Mullus surmuletus, Phycisphycis, Sciaena umbra, Scorpaena porcus and Sym-phodus tinca). Other species, like Diplodus spp.,Muraena helena, or cephalopods, did not present cleartrends in spite of their high contribution to the catches.
Some of the species that responded to protectionwith higher catches near the border, such as Dentexdentex, Mullus surmuletus, Phycis phycis, Sciaenaumbra and Scorpaena porcus, are important targetspecies of artisanal fisheries (García-Rodríguez et al.2006, Forcada 2007, Goñi et al. 2008). Moreover, thesespecies represent the highest proportion of the catchand the greatest income for artisanal fisheries (García-Rodríguez et al. 2006, Forcada 2007, Goñi et al. 2008).
208
Fig. 6. Trends in catches of: (a) Pagellus acarne, (b) Palinuruselephas, (c) Mullus surmuletus, (d) Sepia officinalis and (e)Uranoscopus scaber fished on sandy bottoms at differentdistances (see Fig. 1) from the Cerbère-Banyuls MarineReserve during the 2 surveys. Error bars = standard error.Surveys were not conducted in summer due to bad weather
conditions

Forcada et al.: Effectiveness of MPA for fisheries enhancement
Therefore, export of fish from the MPAs, even if it islimited, should provide economic benefits to artisanalfleets.
Although fisheries enhancement was detected veryclose to MPA borders in general, there were some dif-ferences between the Tabarca and Carry-le-RouetMarine Reserves. They are located in different biogeo-graphical sectors (Garibaldi & Caddy 1998, Bianchi &Morri 2000), and one of the most important differencesis that of sea temperature, which ranges at the surfacefrom 13 to 27°C in Tabarca and from 12 to 24°C inCarry-le-Rouet. This accounts for the higher abun-dance at Tabarca of certain species typical of warmerwaters (Sciaena umbra and Dentex dentex), which arerare in the northern Mediterranean (Harmelin 1991,Ramos-Esplá & Bayle 1991, Francour et al. 1994).
The other difference between the MPAs is that atCarry-le-Rouet the increase of the catches was onlydetected near the boundaries, whereas at Tabarcasome effects, like the increase of Scorpaena porcusand Sciaena umbra catches, reached an intermediatedistance, only declining at the fishing ground furthestfrom the MPA. This may be interpreted in relation tothe size of each MPA. Carry-le-Rouet is a very smallreserve (85 ha) that has a very limited influence overthe surrounding fished area, while Tabarca is a largerone (1400 ha) that seems to have a wider exportationeffect. The effect of protection of juveniles and adultsis edge dependent. The probability of fish leavinga given reserve, and consequently increasing theirvulnerability, will decrease in proportion to the in-crease in size of the MPA (Kramer & Chapman 1999,Chapman & Kramer 2000, Roberts et al. 2001b). There-fore, small MPAs may not support populations effi-ciently (Edgar & Barrett 1999), especially for mobilespecies that often cross reserve boundaries. However,small MPAs can have an important protective role inthe case of sedentary species or if the the reserves aresituated at crucial locations, such as aggregation sitesfor spawning (Beets & Friedlander 1999). There are noupper limits on reserve size that are relevant to conser-vation goals, but to achieve an export of fishable stocksthey should not be too large (Roberts et al. 2003). Ifreserves are made too large, then spillover to fisherieswill be prevented, while making reserves too small willyield no benefits. It is difficult to be precise about whatconstitutes ‘too large’ because it depends on the mobil-ity of the species involved and local oceanographicconditions (Roberts et al. 2003). To determine whatreserve size is too small more empirical study is re-quired (Edgar & Barrett 1999, Halpern 2003, Gui-detti et al. 2005). From the results obtained, TabarcaMarine Reserve and Carry-le-Rouet Marine Reserveseem large enough to promote population recovery,and yet are small enough to permit some spillover for
the benefit of local fisheries. Even the small MPAappears capable of generating increases in catches nearits boundaries; the larger one shows the same effects,but proportional to its size. For that reason, theabsolute impacts of small and large reserves will bevery different. Although the observations at each MPAsize were not replicated, these results offer someempirical evidence suggesting that the amount of pro-tected area is important.
In contrast, there was no clear trend in catches onsand in Tabarca and Cerbère-Banyuls Marine Reser-ves. Only a few of the surveys showed higher catchesnear the boundaries, and only 2 of the species, Pagel-lus acarne and Palinurus elephas, evidenced decreas-ing trends with distance in the Cerbère-BanyulsMarine Reserve, but neither of these was significant.
It appears that biomass is exported through Posido-nia oceanica meadows (at least for some species andseasons), while little export occurs through sandybottoms. At both Tabarca and Carry-le-Rouet MarineReserves, the main habitat, P. oceanica meadow,extends outside the borders. Rocky and sandy bottomsare also present in Tabarca Marine Reserve, but arevery limited, ~10 and ~20%, respectively, of the sur-face protected. Something similar occurs in Cerbère-Banyuls Marine Reserve, which protects mainly rockybottoms, and sandy bottoms start just near the MPAboundaries.
The types and qualities of habitats, both inside andoutside the reserve, determine how a species respondsto reserve protection (Agardy 1995, Nilsson 1998). It isdesirable that reserves protect habitats where speciesfeed and reproduce and where they spend a consider-able portion of time (Kramer & Chapman 1999). In thissense, habitats can act as surrogates for species inreserve planning, simplifying the task of decidingwhat to protect (Roberts et al. 2001b). On the otherhand, the relative mobility of some species (Chapman& Kramer 2000) suggests that movement across theboundary would be much greater if the boundarydivides an area of continuous habitat. It has beenpointed out (Roberts 2000) that habitat continuitythrough MPA boundaries is important for biomassexport to open fished areas, and our results confirmthis assessment.
Although we found that the spatial scale of thespillover-induced density gradient is very localized, itis, nevertheless, sufficient to provide local benefits toartisanal fisheries (through juvenile and adultspillover) and possibly more regional benefits (throughgreater larval export). We conclude that spillovereffects are not a universal consequence of marine pro-tected areas in temperate waters, and that they arerelated to the distribution of habitat inside and aroundthe reserve.
209

Mar Ecol Prog Ser 379: 197–211, 2009
Acknowledgements. We very much appreciated the helpreceived from A. Delauney, K. Kawahara, M. Capoulade andB. Luna during field data collection. We acknowledge thefriendly cooperation of the fishermen of Tabarca, Carry-le-Rouet and Cerbère-Banyuls. This research was financed byUE-DG Fisheries through the project BIOMEX (QLRT-2001-00891), WP5. A.F. was supported by an FPI grant from theGeneralitat Valenciana (CTBPRB/2003/146). We thank theFAO for providing species drawings for Figs. 3, 4, 5 & 6. Wealso thank the anonymous reviewers and the editor for theiruseful comments on the manuscript.
LITERATURE CITED
Abesamis RA, Russ GR (2005) Density-dependent spilloverfrom a marine reserve: long-term evidence. Ecol Appl 15:1798–1812
Agardy TE (1995) The science of conservation in the coastalzone: new insight on how to design, implement, and mon-itor marine protected areas. A marine conservation anddevelopment report. IUCN, Gland
Beets J, Friedlander A (1999) Evaluation of a conservationstrategy: a spawning aggregation closure for red hind,Epinephelus guttatus, in the US Virgin Islands. EnvironBiol Fishes 55:91–98
Bell JD (1983) Effects of depth and marine reserve fishingrestrictions on the structure of a rocky reef fish assem-blage in the north-western Mediterranean Sea. J ApplEcol 20:357–369
Bianchi CN, Morri C (2000) Marine biodiversity of the Medi-terranean Sea: situation, problems and prospects for futureresearch. Mar Pollut Bull 40:367–376
Bohnsack JA (1990) The potential of marine fishery reservesfor reef fish management in the US southern Atlantic.NOAA Tech Memo NMFS-SEFC-261, National Oceanicand Atmospheric Administration, Miami, FL
Castilla JC (2000) Roles of experimental marine ecology incoastal management and conservation. J Exp Mar BiolEcol 250:3–21
Chapman MR, Kramer DL (2000) Movements of fishes withinand among fringing coral reefs in Barbados. Environ BiolFishes 57:11–24
Cocheret de la Moriniere E, Pollux BJA, Nagelkerken I, vander Velde G (2002) Post-settlement life cycle migrationpatterns and habitat preference of coral reef fish that useseagrass and mangrove habitats as nurseries. Estuar CoastShelf Sci 55:309–321
Cochran WG (1951) Testing a linear relation among vari-ances. Biometrics 7:17–32
Edgar GJ, Barrett NS (1999) Effects of the declaration ofmarine reserves on Tasmanian reef fishes, invertebratesand plants. J Exp Mar Biol Ecol 242:107–144
Forcada A (2005) Ictiofauna en reservas marinas: influenciade la estructura del hábitat, efecto de la protección ytamaño del área protegida. Instituto Alicantino de Cul-tura Juan Gil-Albert, Publicaciones de la Diputación deAlicante, Alicante
Forcada A (2007) Evaluación de las Áreas Marinas Protegidasy su efecto en pesquerías artesanales del MediterráneoOccidental. PhD thesis, University of Alicante, Alicante
Francour P, Boudouresque CF, Harmelin-Vivien M, HarmelinJG, Quignard JP (1994) Are the Mediterranean watersbecoming warmer? Information from biological indicators.Mar Pollut Bull 28:523–526
Galal N, Ormond RFG, Hassan O (2002) Effect of a network ofno-take reserves in increasing catch per unit effort and
stocks of exploited reef fish at Nabq, South Sinai, Egypt.Mar Freshw Res 53:199–205
García-Rodríguez MA, Fernández M, Esteban A (2006) Char-acterisation, analysis and catch rates of the small-scalefisheries of the Alicante Gulf (SE Spain) over a 10 yearstime series. Fish Res 77:226–238
Garibaldi L, Caddy JF (1998) Biogeographic characterizationof Mediterranean and Black Seas faunal provinces usingGIS procedures — a need for concerted internationalaction. Ocean Coast Manag 39:211–227
Gell FR, Roberts CM (2003) Benefits beyond boundaries: thefishery effects of marine reserves. Trends Ecol Evol 18:448–455
Goñi R, Quetglas A, Reñones O (2006) Spillover of spinylobsters Palinurus elephas from a marine reserve to anadjoining fishery. Mar Ecol Prog Ser 308:207–219
Goñi R, Adlerstein S, Alvarez-Berastegui D, Forcada A andothers (2008) Spillover from six western Mediterraneanmarine protected areas: evidence from artisanal fisheries.Mar Ecol Prog Ser 366:159–174
Guidetti P, Verginella L, Viva C, Odorico R, Boero F (2005)Protection effects on fish assemblages, and comparison oftwo visual-census techniques in shallow artificial rockyhabitats in the northern Adriatic Sea. J Mar Biol Assoc UK85:247–255
Halpern BS (2003) The impact of marine reserves: Doreserves work and does size matter? Ecol Appl 13:117–137
Harmelin JG (1991) Statut du corb (Sciaena umbra) enMéditerranée. In: Boudouresque CF, Avon M, Gravez V(eds) Les espèces marines à protéger en Méditerranée.GIS Posidonie Publications, Marseille, p 219–227
Harmelin JG, Bachet F, García F (1995) Mediterranean marinereserves: fish indices as tests of protection efficiency.PSZN I: Mar Ecol 16:233–250
Hilborn R, Stokes K, Maguire JJ, Smith T and others (2004)When can marine reserves improve fisheries manage-ment? Ocean Coast Manag 47:197–205
Kaunda-Arara B, Rose GA (2004) Effects of marine reefNational Parks on fishery CPUE in coastal Kenya. BiolConserv 118:1–13
Kramer DL, Chapman MR (1999) Implications of fish homerange size and relocation for marine reserve function.Environ Biol Fishes 55:65–79
Maunder MN, Sibert JR, Fonteneau A, Hampton J, Kleiber P,Harley SJ (2006) Interpreting catch per unit effort data toassess the status of individual stocks and communities.ICES J Mar Sci 63:1373–1385
McClanahan TR, Kaunda-Arara B (1996) Fishery recovery ina coral-reef marine park and its effect on the adjacent fish-ery. Conserv Biol 10:1187–1199
McClanahan TR, Mangi S (2000) Spillover of exploitablefishes from a marine park and its effect on the adjacentfishery. Ecol Appl 10:1792–1805
Murawski SA, Brown R, Lai HL, Rago PJ, Hendrickson L(2000) Large-scale closed areas as a fishery-managementtool in temperate marine systems: the Georges Bankexperience. Bull Mar Sci 66:775–798
Murawski SA, Wigley SE, Fogarty MJ, Rago PJ, Mountain DG(2005) Effort distribution and catch patterns adjacent totemperate MPAs. ICES J Mar Sci 62:1150–1167
Nagelkerken I, van der Velde G (2002) Do non-estuarinemangroves harbour higher densities of juvenile fish thanadjacent shallow-water and coral reef habitats in Curaçao(Netherlands Antilles)? Mar Ecol Prog Ser 245:191–204
Nilsson P (1998) Criteria for the selection of marine protectedareas. Report 4834, Swedish Environmental ProtectionAgency, Stockholm
210

Forcada et al.: Effectiveness of MPA for fisheries enhancement
Pauly D, Christensen V, Guénette S, Pitcher TJ and others(2002) Towards sustainability in world fisheries. Nature418:689–695
Rakitin A, Kramer DL (1996) Effect of a marine reserve on thedistribution of coral reef fishes in Barbados. Mar Ecol ProgSer 131:97–113
Ramos-Esplá AA, Bayle J (1991) Estatuto del Dentex dentex(Linneaus, 1758) en el Meiterráneo. In: Boudouresque CF,Avon M, Gravez V (eds) Les espèces marines à protégeren Méditerranée. GIS Posidonie Publications, Marseille,p 237–244
Ramos-Esplá AA, Valle-Pérez C, Bayle-Sempere JT, Sánchez-Lizaso JL (2004) Áreas Marinas Protegidas como her-ramientas de Gestión Pesquera en el Mediterráneo (ÁreaCOPEMED). Serie Informes y Estudios COPEMED No. 11,Proyecto FAO-COPEMED, Madrid
Roberts CM (2000) Selecting marine reserve locations: opti-mality versus opportunism. Bull Mar Sci 66:581–592
Roberts CM, Bohnsack JA, Gell F, Hawkins JP, Goodridge R(2001a) Effects of marine reserves on adjacent fisheries.Science 294:1920–1923
Roberts CM, Halpern BS, Palumbi SR, Warner RR (2001b)Reserve networks. Why small, isolated protected areas arenot enough. Biol Conserv Pract 2:10–17
Roberts CM, Branch G, Bustamante RH, Castilla JC andothers (2003) Applications of ecological criteria in select-ing marine reserves and developing reserve networks.Ecol Appl 13:215–228
Rowley RJ (1994) Marine reserves in fisheries management.Aquat Conserv: Mar Freshw Ecosyst 4:233–254
Russ GR, Alcala AC (1996) Do marine reserves export adult
fish biomass? Evidence from Apo Island, central Philip-pines. Mar Ecol Prog Ser 132:1–9
Russ GR, Alcala AC, Maypa AP (2003) Spillover from marinereserves: the case of Naso vlamingii at Apo Island, thePhilippines. Mar Ecol Prog Ser 264:15–20
Sale PF, Cowen RK, Danilowicz BS, Jones GP and others(2005) Critical science gaps impede use of no-take fisheryreserves. Trends Ecol Evol 20:74–80
Sánchez-Lizaso JL, Goñi R, Reñones O, García Chartón JAand others (2000) Density dependence in marine pro-tected populations: a review. Environ Conserv 27:114–158
Tremain DM, Harnden CW, Adams DH (2004) Multidirec-tional movements of sportfish species between an estuar-ine no-take zone and surrounding waters of the IndianRiver Lagoon, Florida. Fish Bull (Wash DC) 102:533–544
Underwood AJ (1981) Techniques of analysis of variance inexperimental marine biology and ecology. Oceanogr MarBiol Annu Rev 19:513–605
Underwood AJ (1997) Experiments in ecology: their logicaldesign and interpretation using analysis of variance. Cam-bridge University Press, Cambridge
Wiens JA, Stenseth NC, Van Horne B, Ims RA (1993) Eco-logical mechanisms and landscape ecology. Oikos 66:369–380
Willis TJ, Millar RB, Babcock RC, Tolimiery N (2003) Burdensof evidence and the benefits of marine reserves: PuttingDescartes before des (sic) horse? Environ Conserv 30:97–103
Worm B, Barbier EB, Beaumont N, Duffi E and others (2006)Impacts of bbiodiversity loss on ocean ecosystem services.Science 314:787–790
211
Editorial responsibility: Romuald Lipcius,Gloucester Point, Virginia, USA
Submitted: November 23, 2007; Accepted: December 16, 2008Proofs received from author(s): March 13, 2009





























