Embed Size (px)
Citation preview

international journal of andrology, 18:23-34 (1 995)
Effect of androgens on the germ cell-depleted testes of prenatally irradiated rats
G. VERHOEVEN, L. DEBOEL, J. SWINNEN, L. ROMBAUTS, E VANDERHOYDONC, J. ROSSEELS, E. HOEBEN and W. HEYNS Laboratory for Experimental Medicine and Endocrinology, Onderwijs en Navorsing, Gasthuisberg, Catholic University of Leuven, B-3000 Leuven, Belgium
Summary To study the effect of androgens on somatic testicular cells, rats were rendered germ
cell depleted by prenatal irradiation (RX). Adult R X rats were treated with a desensitizing dose of a GnRH agonist (GnRHa; Zoladex), combined with an antiandrogen (Nilutamide) to preclude all androgen effects, or combined with testosterone or hCG to restore androgen action. The effect of these treatments for 3 weeks on the weight of testes and accessory sex glands, hormones (LH, FSH, testosterone, inhibin), testicular proteins, the pattern of incorporation of [ 35S]-methionine into testicular proteins (studied by two dimensional gel electrophoresis) and steady state mRNA levels for transferrin and androgen-binding protein (ABP) were evaluated. Combined treatment with GnRHa and antiandrogen virtually eliminated gonadotrophins, androgens and androgen effects. Testicular weight was reduced to 50% of that observed in R X controls. Treatment with GnRHa and testosterone resulted in supraphysiological levels of testosterone and testicular weights comparable to those observed in R X controls. FSH levels in these animals, however, were in the normal range. A low dose of hCG also restored testicular weight in the presence of low concentrations of serum testosterone and low normal levels of FSH. Neither polyacrylamide gel electrophoresis of total testicular proteins nor two dimensional gel electrophoresis of [ S]-methionine labelled proteins revealed striking changes in distinct testicular proteins as a result of androgen withdrawal or androgen treatment. Dot blot hybridization showed a three-fold increase in the mRNA level for ABP (expressed per pg total RNA) in the Sertoli cell enriched testes of R X rats. This level was barely influenced by androgen withdrawal or androgen administration. The mRNA level for transferrin was increased six-fold in R X rats. A 50% reduction of this level was observed after combined treatment with GnRHa and antiandrogen. It is concluded that, in the germ cell-depleted testis, the major effect of androgens is an overall increase in protein and RNA synthesis rather than a very important and selective increase of a few gene products.
35
Keywords: GnRH, antiandrogens, transferrin, ABP, androgen action, Sertoli cell- enriched testis
Correspondence: G. Verhoeven, Legendo, Ondenvijs en Navorsing, Gasthuisberg, B-3000 Leuven, Belgium.
0 1995 Ulackwrll Scirricr Ltd

24 G. Verhoeven eta/.
Introduction Initiation and maintenance of spermatogenesis depend
both on LH and on FSH. FSH acts directly on the tubular compartment whereas LH exerts its effects via testosterone produced by the Leydig cells (Steinberger, 1971; Parvinen, 1972; Hansson et al., 1976). I t is generally assumed that both androgens and FSH act via somatic cells, and probably niain- ly via Sertoli cells, to influence germ cell proliferation and differentiation (Fritz, 1978). During the last decade the activity of these hormones has been studied extensively in isolated and cultured cells. These studies have revealed that the effects of androgens on isolated Sertoli cells are less pronounced than expected Louis & Fritz, 1979; Sharpe, 1986; Verhoeven, 1992). Moreover, evidence has been pre- sented that some effects of androgens on Sertoli cells niay be mediated o r at least modulated by paracrine factors produced by the underlying peritubular cells (Skinner & Fritz, 1985; Skinner et al., 1988; Verhoeven & Cailleau, 1988a). Attempts to study the primary effects of androgens in the intact testis are hampered by the indirect but over- whelming effects of androgens o n germ cell development. Hypophysectoniized, germ cell-depleted animals have been used to study the effects of androgens on soniatic cells of the testis (Mills, 1990). O n e drawback of this experimental model, however, is that hypophysectoniy renioves not only gonadotrophins but also other pituitary hormones that may influence androgen action in the testis directly or indirectly. In the present study w e have explored the use of prenatally irradiated rats treated with a desensitizing dose of a G n K H agonist to study the effects of androgens on testicular soniat- ic cells irz vivo. O n e of the aims was to evaluate whether such animals might provide useful starting material to identify key somatic androgen-regulated genes by subtraction cloning.
Materials and methods
Aiiinials Wistar rats were supplied by the breeding center of the
Catholic University of Leuven. T h e animals were kept under standard conditions of light, teniperatu're and humid- ity and had free access to a standard pellet diet and water. Rat? were killed by exsanguination under ether anaesthesia. T h e serum was divided into aliquots and stored at -2OOC for hormone measurements. T h e organs (testes, prostate, senii- nal vesicles) were removed and weighed ininiediately. Testes were decapsulated, pooled, frozen and stored in liquid nitrogen for K N A extraction. From some animals testicular fragments were saved for histological examination, polyacrylamide gel electrophoresis or protein labelling experiments. Kats with the testicular feminization syndronie were kindly provided to our colleague Dr Vanderschueren by Dr. K . Chung (llepartnient of Anatomical Sciences, Oklahoma City, O.K., U.S.A.). Testes were removed at the end of their experiments at the age of approximately 4 months (Vanderschueren ct ul., 1993).
Experimental protocol Time-bred female rats were irradiated on the 20th day of
pregnancy to obtain male offspring with Sertoli cell- enriched testes. Irradiation was performed as described by Fakunding et al. (1976). Pregnant control animals were not irradiated. Twenty-one days after birth the animals were weaned and male rats were selected and divided into groups (n=10) for further experiments.
In the first experiment, animals were subjected to five different treatment protocols. Group I consisted of non-irra- diated controls. Group I1 consisted of irradiated controls. In group I11 irradiated, Sertoli cell-enriched males were treated with a desensitizing dose of G n R H agonist (Goserilin, Zoladex) and an antiandrogen (Nilutamide, RU 23908). Group IV contained irradiated animals treated with the G n R H agonist (GnRHa) and a high dose of testosterone and Group V were irradiated animals treated with testos- terone only. All the indicated treatments were started at the age of 82 days and were continued for 3 weeks. Zoladex was provided kindly by ICI-Pharma (Destellbergen, Belgium) and was administered as a single subcutaneous injection (0.9 &animal) of the depot formulation in the neck region. T h e nonsteroidal antiandrogen Nilutamide (Raynaud et al., 1979) was supplied kindly by Roussel Uclaf (Paris, France) and was administered subcutaneously (5 mg/animal/day in 0.2 ml sesame oil containing 5% benzyl alcohol). Testosterone was administered using silastic capsules (inter- nal diameter: 1.98 nini, external diameter: 3.18 mm; 4 x 5 cni/animal).
In the second experiment all treatments were started at the age of 150 days. T h e same controls ( Group I and 11) were included as in the first experiment. Group 111 was treated with G n K H a only, Group IV with G n K H a and the antiandrogen, and Group V with G n R H a and h C G (Pregnyl, 1.5 U/day administered subcutaneously in 0.1 ml saline). All animals were killed 3 weeks after the start of the indicated treatments.
Hormone wieasurements LH and FSH were measured in seruni by double-anti-
body radioimniunoassays using reagents supplied by Dr. A.F. Parlow and the NIDDK (Bethesda, M.D., U.S.A.). T h e standard preparations used were LH-RP-3 and FSH- KP-2. Tracers were prepared froni rLH-1-9 and rFSH-1-8 and the antisera were anti rLH S-10 and anti rFSH-S-11 respectively. Using 0.2 nd serum in the assay the detection limits were 0.5 n g / d for LH and 0.75 n g / d for FSH. All of the final hormone measurements were performed in a single assay. T h e intra-assay coeficient of variation was 7.3% for LH and 3.8% for FSH.
Testosterone was nieasured by a radioininiunoassay as described previously (Verhoeven et al., 1982). lnhibin was assayed using a radioininiunoassay kit provided kindly by l l r . 1). Robertson and Professor 1). M. de Kretser (Monash University, Victoria, Melbourne, Australia). T h e kit

Androgens and somatic cells of the testis 25
includes a rabbit antiserum (number 1989) raised against purified bovine 31 kDa inhibin. The assay was performed essentially as described by the authors but a second antibody solid-phase system (SAC-cells, IDS-Ltd, Boldon, U.K.) was used to separate free and antibody-bound inhibin (Robertson et al., 1988; Rombauts et al., 1992). A pool of Sertoli cell conditioned medium was used as an operational standard and was assigned an arbitrary value of 1 U/ml. Dilution curves of this pool and male rat serum were paral- lel. The within- and between-assay coefficients of variation were 3.2% and 6.7% respectively. The limit of detection was 0.04 U/ml.
Sodium dodecyl scrlphate-~olyacrylamide gel electrophore- sis (SDS-PAGE) Testicular fragments from two animals from each group
in Experiment 1 were homogenized in 5 vols phosphate- buffered saline. The protein concentration was determined by use of the Pierce BCA protein assay reagent (Pierce, Rockford, M.D., U.S.A.). Aliquots corresponding to 25 pg protein were subjected to SDS-PAGE under reducing con- ditions with a 3% stacking and a 10%1 resolving gel (Laemmli, 1970). Proteins were visualized by staining with Cooniassie blue.
Protein labelling and two-dimensional gel electrophoresis [ 35S]-niethionine incorporation was studied in two ani-
mals from each treatment group in Experiment 1 and in animals from groups I1 and IV in Experiment 2. Aliquots corresponding to 25 mg testicular tissue were minced with scissors, suspended in 250 p1 niethionine-free RPMI-A and incubated for 24 h a t 32OC. Thereafter, fresh medium sup- plemented with 25 pCi [ 35S]-methionine (Tran35S-Label, ICN, Irvine, C.A., U.S.A.) was added and incubation was continued for another 24 h. The medium, containing secreted proteins, was centrifuged at 10,000 x g, dialysed against 0.1M NH,HC03 and aliquots corresponding to 1.5 x lo6 cpm were subjected to electrophoresis. The reniain- ing tissue was dissolved in O'Farrel lysis buffer and aliquots corresponding to lo6 cpni were processed further. Two- dimensional electrophoresis (O'Farell, 1975) was performed using Inimobiline DryStrips pH 4-7 (Pharniacia) for the first dimension and ExcelGel SDS, gradient 8-18 (Pharmacia, Brussels, Belgium) for the second dimension. After the run, gels were fixed in 7% acetic acid, pretreated with Amplify (Amersham International, Aniershani, Buckinghanishire, U.K.), dried and exposed to X-ray film (Hyperfilm MP, Aniersham).
RNA analysis To prepare RNA, testes from all animals in each treat-
ment group were pooled and pulverized in liquid nitrogen. Total RNA was prepared by the guanidine isothiocyanate- CsCl procedure (Chirguin et ul., 1979) using 1 g aliquots of the thoroughly mixed pulverized material.
Northern blot analysis was perfornied on RNA samples (20 &lane) separated on a 1% (w/v) agarose gel containing formaldehyde as described by Sambrook et al. (1989). The R N A was blotted onto nylon membranes (Hybond N') using a 0.01 M NaOH capillary blotting buffer. After the transfer, the blots were rinsed for 1 min in 5 x SSC (20 x SSC: 3 M NaCl and 0.3 M tri-sodium citrate). Blots were prehybridized in 50% (v/v) formamide, 50 mM sodium phosphate (pH 6.5), 5 x SSC, 5 x Denhardt's (100 x Denhardt's: 2% (w/v) Ficoll 400, 2% (w/v) polyvinylpyrrolidone, 2%(w/v) bovine serum albumin fraction V), 250 pg/ml boiled sonicated salmon sperm DNA and 0.1% (w/v) SDS at 42OC overnight. Three probes were used for hybridization: to transferrin, androgen binding pro- tein (ABP) and to 18s rRNA. A pBR 325 plasmid contain- ing a 1540 bp rat transferrin cDNA probe cloned in the Eco RI site was provided kindly by Professor G. Schreiber (Melbourne, Australia (Aldred et al., 1984). The ABP probe was prepared by reverse transcription and polymerase chain reaction (PCR). cDNA was generated by reverse transcrip- tion of poly A+ R N A derived from 7-day-old Sertoli cell cultures prepared and maintained as described previously. Oligo d T was used as a primer. With this DNA as a tem- plate a fragment of rat ABP corresponding to nucleotides 16G1121 was amplified using as primers: GTTCTGAC- CATTGACCTCACC and CTGTAAGCTTTGTCTAT- GTCCAGTCTC. After digestion with Bam HI and Hind 111, an 896 bp ABP fragment was inserted into the plasmid pUCl8. DNA sequencing confirmed that this fragment corresponded to the published sequence of ABP. A pUCl8 plasmid containing an approximately 4000bp Eco RI restriction fragment of the genomic DNA of the coelen- therate Anernonia sulcata with approximately 1800bp of the gene encoding 18s rRNA, was provided kindly by Dr L. Hendriks (Department of Biochemistry, Universitaire Instelling Antwerpen, Belgium) (Hendriks et al., 1990). A 1549 bp Eco RI-NdeI restriction fragment containing the major part of the 18s rRNA gene was used as a probe. The probes were labelled using (a-32P)dCTP (3000 Ci/mmole; New England Nuclear, Wilmington, DE, U.S.A.) and a random primed labelling kit provided by Boehringer Mannheim (Mannheini, Germany). Unincorporated nucleotides were removed by centrifugation of the reaction mixture through a Sephadex G50 column (Pharmacia, Uppsala, Sweden). The blots were incubated with radiola- belled probe at 106 cpm/ml of hybridization buffer consist- ing of 50% formamide, 20 mM sodium phosphate (pH 6.5), 5 x SSC, 5 x Denhardt's, 100 p g / d boiled sonicated salmon sperm DNA and 7.5% (w/v) dextran sulphate at 42OC for 48 h. After hybridization, the blots were washed first in 0.5 X SSC, 0.1% (w/v) SDS at room temperature, then in 0.5 x SSC, 0.1% SDS at 65OC (30 min) and finally in 0.2 x SSC, 0.1% SDS at 65OC (30 min). The blots were autoradiographed using intensifying screens. Probes were removed by placing the membranes in a boiling solution

26 0. Verhoeven et al.
containing 0.1% SDS and allowing the solution to cool down to room temperature.
Dot blot analysis was performed on samples of total RNA, denatured in the presence of formaldehyde for 10 min at 60°C and adjusted to 15 x SSC (15 x SSC: 2.25 M NaCl and 0.225 M tri-sodium citrate). Serial dilutions (1:2) were made in 15 x SSC in a microtitre plate and applied to a Minifold (Schleicher and Schuell Inc., Keene, NH, U.S.A.) using Hybond-C extra-fortified nitrocellulose membranes (Amershani). R N A was immobilized at 80OC for 2 h. Blots were hybridized with the transferrin and AE3P probes as described above. After autoradiography, the probes were removed by boiling in an aqueous solution containing 0.1% SDS and the same blots were rehybridized with the 18 S probe. Autoradiograms were scanned using an LKB Bromma Ultroscan XL Enhanced Laser Densitometer. Densitometric measurements were compared in the linear parts of the absorbance curves and expressed in relative den- sitometric units.
His tolugy Testicular fragments for histological examination were
removed from two animals in each treatment group. The tissue was fixed in Bouins, washed, dehydrated and embedded in paraffin. Sections (3pm) were stained with hematoxylin and eosin.
Statistical analysis Statistical significances were assessed using one-way
analysis of variance supplemented by Tuckey’s studentized range test.
Results
Body and organ weights Prenatal irradiation reduced testicular weight markedly
whereas prostate and seminal vesicle weight were main- tained (Table 1). Chemical castration of the irradiated ani- mals with GnRHa reduced the weight of the accessory sex glands significantly and tended to decrease testicular weight. The combination of GnRHa with an antiandrogen reduced testicular weight to 50% of that observed in irradiated controls but no significant further decrease in prostate and seminal vesicle weight was observed. Irradiated animals treated with testosterone implants either in the presence or in the absence of GnRHa had testicular weights indistin- guishable from those of irradiated c.ontrols. Prostate and seminal vesicle weights, however, increased to twice the control size. Substitution of GnRHa-treated animals with hCG restored the weights of testes, prostate and seminal vesicles to values observed in irradiated controls.
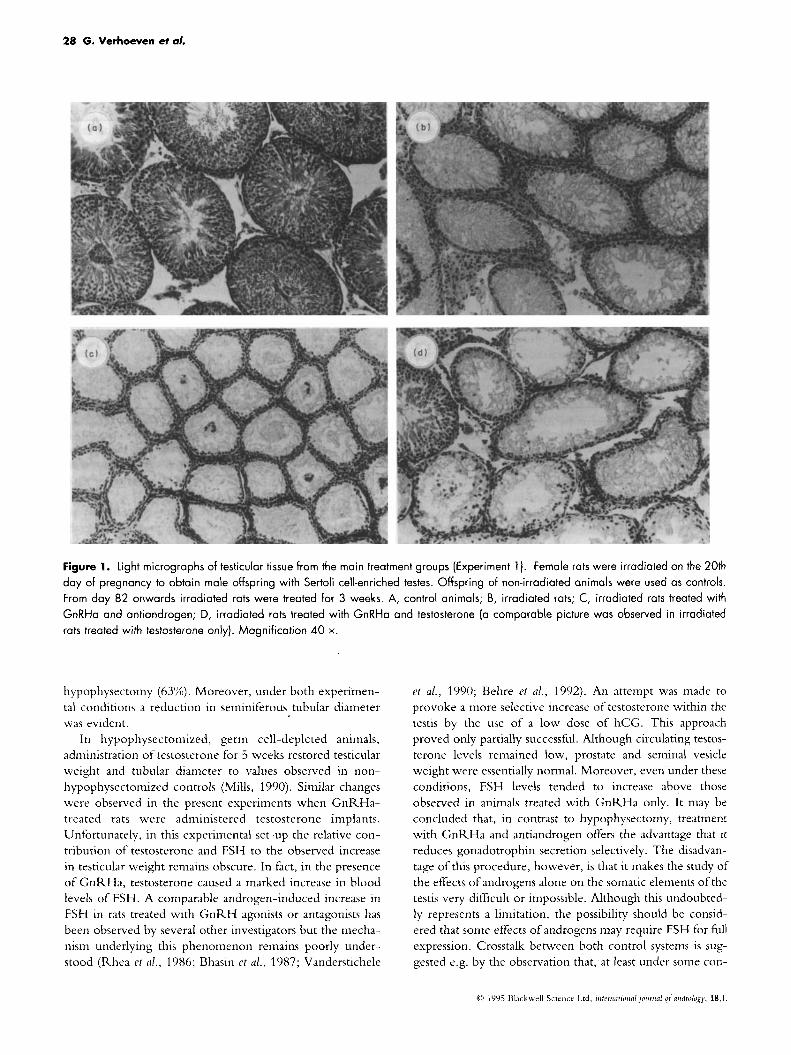
Histological examination confirmed that prenatal irradia- tion resulted in the production of Sertoli cell-enriched testes. Although no attempts were made to quantify changes in tubular diameter, it is evident from Fig.1 that irradiation
reduced this parameter. In accordance with the observed changes in testicular weight, the combination of GnRHa with antiandrogen caused a further reduction in tubular diameter, whereas no such changes were evident in animals treated with testosterone only (not shown) or with GnRHa plus testosterone.
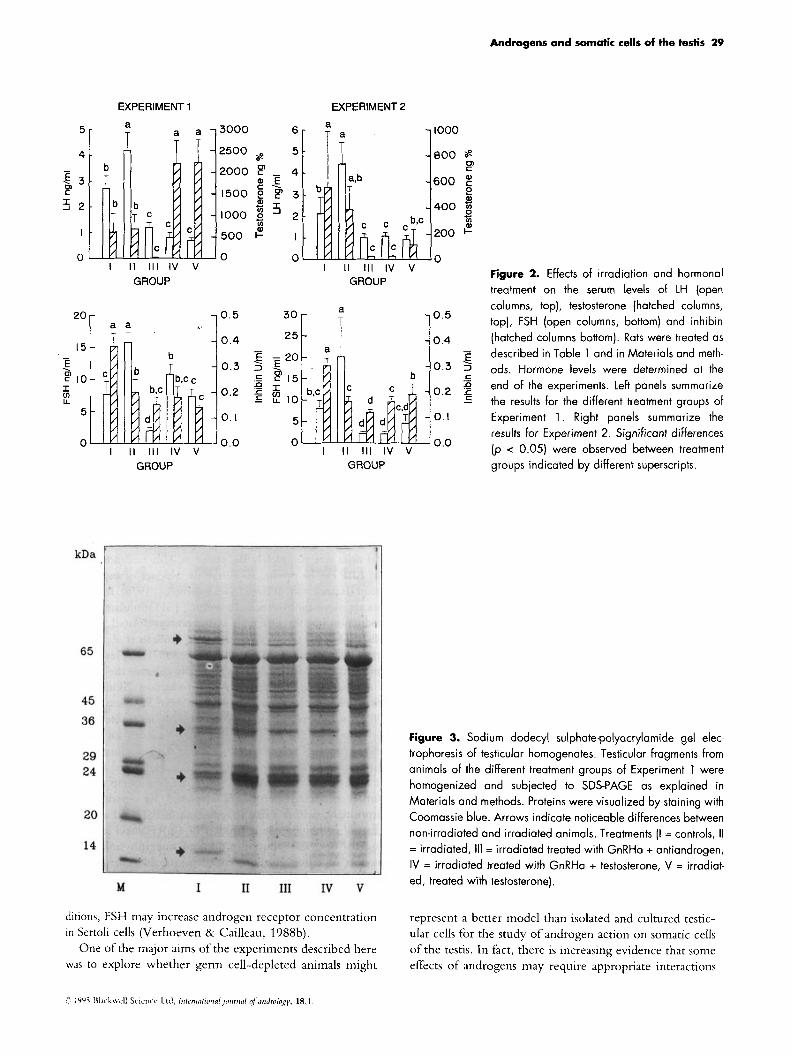
Hormone levels As expected, gonadotrophin levels were raised in the
irradiated animals (Fig. 2). Despite the increase in LH, testosterone levels were maintained in the physiological range suggesting a resetting of the testicular hypophyseal- hypothalamic feedback axis. GnRHa treatment decreased LH and FSH levels markedly and androgen levels became barely detectable; additional treatment with an androgen antagonist did not affect the low gonadotrophin and testos- terone levels further. The testosterone implants increased the circulating levels of testosterone to more than three times those observed in untreated controls. In irradiated animals, testosterone decreased LH levels markedly whereas FSH was reduced to levels observed in untreated controls. In irradiated animals treated with GnRHa plus testosterone, FSH was in the normal range. A tendency to increased FSH levels was even noted in GnRHa-treated animals supple- mented with hCG, a change that occurred despite the fact that circulating levels of androgen in these animals remained rather low. Levels of immunoreactive inhibin in blood dropped to 50% of their original value in irradiated germ- cell depleted animals. Chemical castration or androgen administration barely influenced inhibin levels, whereas a small increase was observed after administration of hCG.
Influence of irradiation and hormone treatment on protein and RNA synthesis T o study the effects of the mentioned treatments on pro-
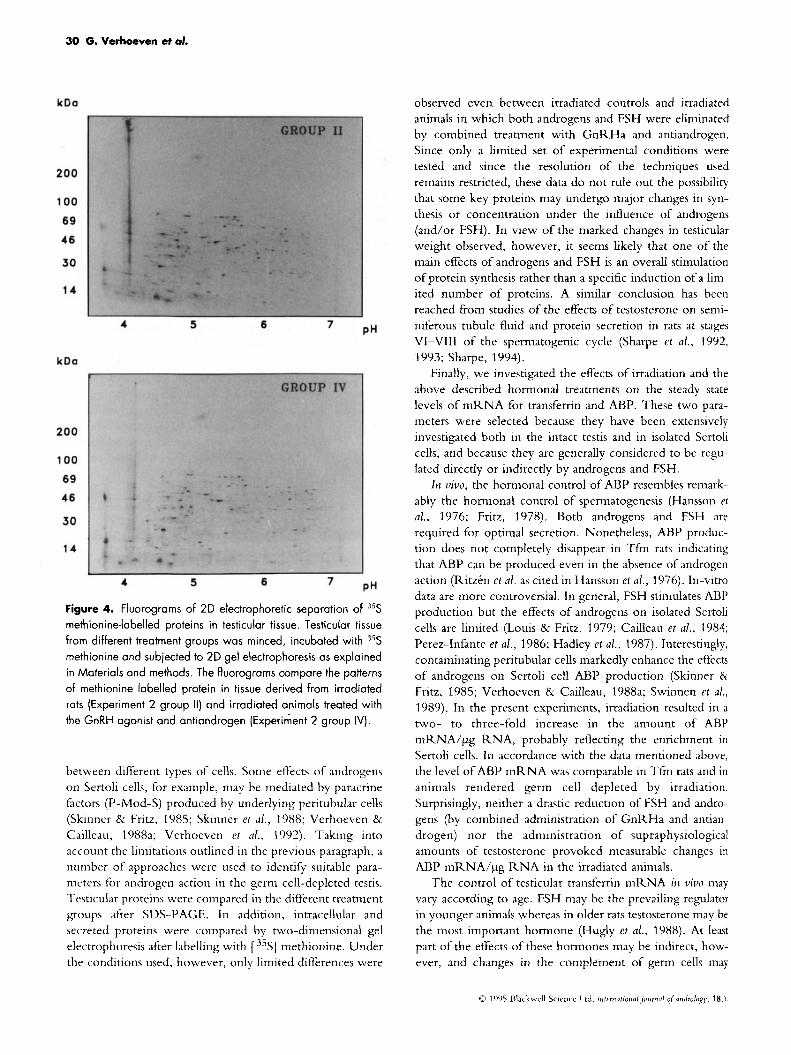
tein synthesis two approaches were explored. In the first, the protein profiles of the testes derived from animals of the dif- ferent treatment groups in Experiment 1 were compared by SDS-polyacrylamide gel electrcphoresis followed by Coomassie blue staining. As shown in Fig.3, major differ- ences were observed between non-irradiated and irradiated animals e.g. in the 68, 35, 24 and 15 kDa region, but only minor differences were noted between groups 11-V. In the second approach, tissue fragments of the same animals were incubated in the presence of [ 35S] methionine and proteins in the tissues or proteins secreted in the media were analysed by two-dimensional gel electrophoresis. Again, remarkably similar patterns were obtained for groups 11-V. Figure 4 shows the results for the tissues from irradiated animals (Experiment 2, group 11) and irradiated animals treated with GnRHa plus antiandrogens (Experiment 2, group IV).
The influence of the described treatments was then analysed at the level of the mRNA for AE3P and transferrin, two Sertoli cell proteins which are supposedly controlled in part by androgens (Hansson et al., 1976; Louis & Fritz, 1979;
0 I Y Y 5 Dlackwcll Scirncr Ltd, infrmattonal.,oumal ?f artdrolofy, 18.1.
Androgens and somatic cells of the testis 27
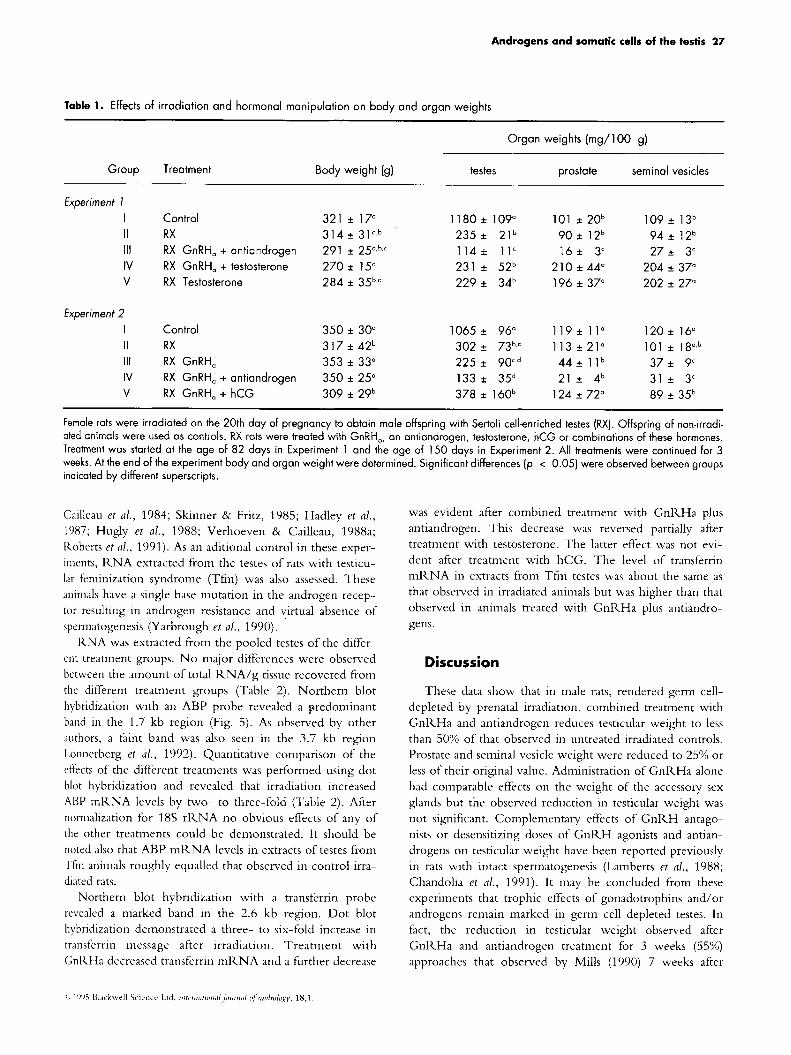
Table 1. Effects of irradiation and hormonal manipulation on body and organ weights
Organ weights (mg/100 g)
Group Treatment Body weight (9) testes prostate seminal vesicles
Experiment I I Control 321 * 17" 1180 * 109" 101 *20b 109 * 13b
111 RX GnRH, + antiandrogen 291 * 25".b,' 1 1 4 i 11' 16* 3' 27* 3' IV RX GnRH, + testosterone 270 f 15' 231 i 52b 210 f 44" 204 f 37"
II RX 314 * 31a.bb 235f 21b 9 0 * 12b 94 i 1 2 b
V RX Testosterone 284 * 35b,' 229* 34b 196 f 37" 202 f 27"
Experiment 2 1065 f 96" 119f 1 1 " 1 2 0 + 16" I Control 350 f 30"
II RX 317 f 42b 302f 73b,' 113 + 2 1 " 101 * 18°,b
IV RX GnRH, + antiandrogen 350 * 25" 133 f 35d 21 f 4b 31 * 3' V RX GnRH, + hCG 309 ~t 29b 378 i 160b 124 f 72" 89 f 35b
111 RX GnRH, 353 f 33" 225 f 90',d 44* l l b 37* 9'
Female rats were irradiated on the 20th day of pregnancy to obtain male offspring with Sertoli cell-enriched testes (RX). Offspring of non-irradi- ated animals were used as controls. RX rats were treated with GnRH,, an antiandrogen, testosterone, hCG or combinations of these hormones. Treatment was started at the age of 82 days in Experiment 1 and the age of 150 days in Experiment 2. All treatments were continued for 3 weeks. At the end of the experiment body and organ weight were determined. Significant differences (p < 0.05) were observed between groups indicated by different superscripts.
Cailleau et a/ . , 1984; Skinner & Fritz, 1985; Hadley et a/.,
1987; Hugly et a/ . , 1988; Verhoeven & Cailleau, 1988a; Koberts et a/., 1991). As an aditional control in these exper- iments, KNA extracted from the testes of rats with testicu- lar feminization syndrome (Tfm) was also assessed. These animals have a single base mutation in the androgen recep- tor resulting in androgen resistance and virtual absence of cpermatogenesis (Yarbrough et al., 1990).
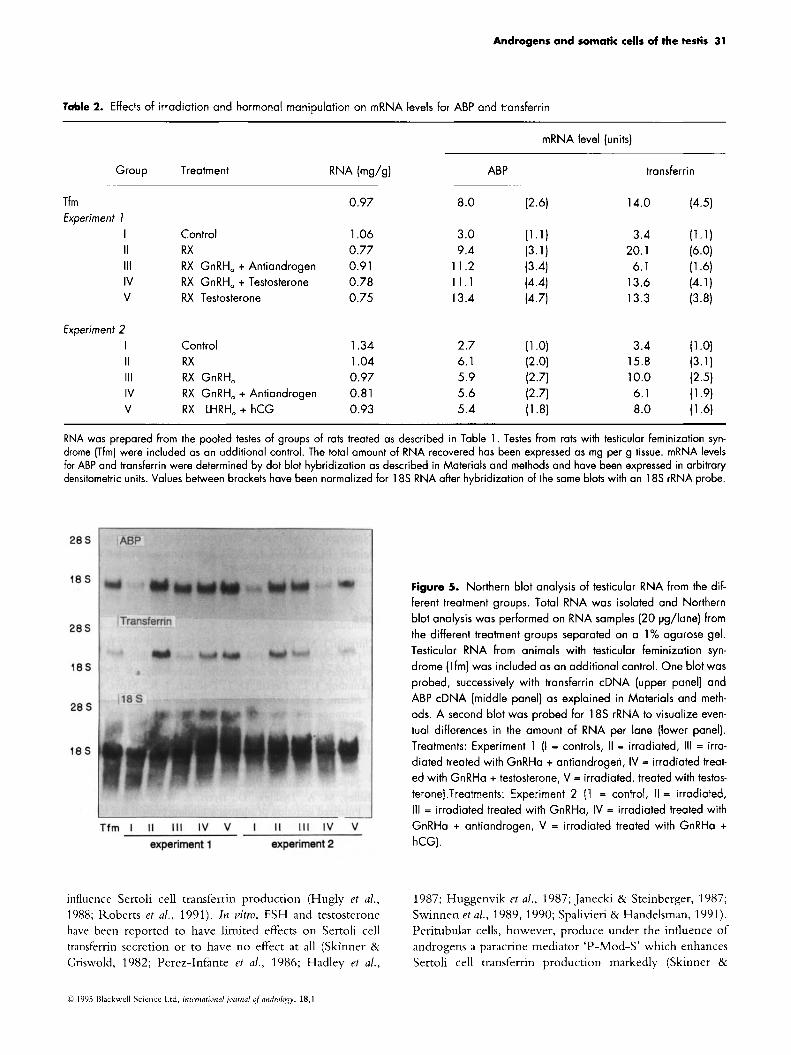
KNA was extracted from the pooled testes of the differ- ent treatment groups. N o major differences were observed between the amount of total KNA/g tissue recovered from the different treatment groups (Table 2). Northern blot hybridization with an ABP probe revealed a predominant band in the 1.7 kb region (Fig. 5). As observed by other duthors, a faint band was also seen in the 3.7 kb region Lolinerberg et al., 1992). Quantitative coniparison of the effects of the different treatments was perfornied using dot blot hybridization and revealed that irradiation increased ABP mRNA levels by two- to three-fold (Table 2). Aiter normalization for 18s rKNA no obvious effects of any of the other treatments could be demonstrated. It should be noted also that ABP niRNA levels in extracts of testes from Tfni animals roughly equalled that observed in control irra- diated rats.
Northern blot hybridization with a transferrin probe revealed a niarked band in the 2.6 kb region. Dot blot hybridization demonstrated a three- to six-fold increase in trmsferrin message after irradiation. Treatment with GnKHa decreased transferrin mKNA and a further decrease
was evident after combined treatment with GnKHa plus antiandrogen. This decrease was reversed partially after treatment with testosterone. The latter effect was not evi- dent after treatment with hCG. The level of transfernn niRNA in extracts from Tfni testes was about the same as that observed in irradiated animals but was higher than that observed in animals treated with GnKHa plus antiandro- gens.
Discussion
These data show that in male rats, rendered germ cell- depleted by prenatal irradiation, combined treatment with GnRHa and antiandrogen reduces testicular weight to less than 50% of that observed in untreated irradiated controls. Prostate and seminal vesicle weight were reduced to 25%) or less of their original value. Administration of GnRHa alone had comparable effects on the weight of the accessory sex glands but the observed reduction in testicular weight was not significant. Complementary effects of GnRH antago- nists or desensitizing doses of GnKH agonists and antian- drogens on testicular weight have been reported previously in rats with intact spermatogenesis (Lamberts et a/., 1988; Chandolia et a/., 1991). It may be concluded from these experiments that trophic effects of gonadotrophins and/or androgens reniain marked in germ cell depleted testes. In fact, the reduction in testicular weight observed after GnKHa and antiandrogen treatment for 3 weeks (55%) approaches that observed by Mills (1990) 7 weeks after
28 G. Verhoeven et al.
Figure 1. Light micrographs of testicular tissue from the main treatment groups (Experiment 1). Female rats were irradiated on the 20th day of pregnancy to obtain male offspring with Sertoli cell-enriched testes. Offspring of non-irradiated animals were used as controls. From day 82 onwards irradiated rats were treated for 3 weeks. A, control animals; B, irradiated rats; C, irradiated rats treated with GnRHa and ontiandrogen; D, irradiated rats treated with GnRHa and testosterone (a comparable picture was observed in irradiated rats treated with testosterone only). Magnification 40 x.
hypophysectoniy (63%). Moreover, under both experimen- tal conditions a reduction in seminiferous tubular diameter was evident.
In hypophysectomized, germ cell-depleted animals, administration of testosterone for 5 weeks restored testicular weight and tubular diameter to values observed in non- hypophysectomized controls (Mills, 1990). Similar changes were observed in the present experiments when GnRHa- treated rats were administered testosterone implants. Unfortunately, in this experimental set-up the relative con- tribution of testosterone and FSH to the observed increase in testicular weight reniains obscure. In fact, in the presence of GnRHa, testosterone caused a niarked increase in blood levels of FSH. A coniparable androgen-induced increase in FSH in rats treated with G n R H agonists or antagonists has been observed by several other investigators but the mecha- nism underlying this phenomenon remains poorly under- stood (Rhea et a!., 1986; Bhasin et d., 1987; Vanderstichele
et al., 1990; Behre et al., 1992). An attempt was made to provoke a more selective increase of testosterone within the testis by the use of a low dose of hCG. This approach proved only partially successful. Although circulating testos- terone levels remained low, prostate and seminal vesicle weight were essentially normal. Moreover, even under these conditions, FSH levels tended to increase above those observed in animals treated with GnRHa only. I t may be concluded that, in contrast to hypophysectomy, treatment with GnRHa and antiandrogen offers the advantage that i t reduces gonadotrophin secretion selectively. The disadvan- tage of this procedure, however, is that it makes the study of the effects of androgens alone on the somatic elements of the testis very difficult or impossible. Although this undoubted- ly represents a limitation, the possibility should be consid- ered that sonie effects of androgens may require FSH for full expression. Crosstalk between both control systems is sug- gested e.g. by the observation that, at least under some con-
Androgens and somatic cells of the testis 29
. E -20
5
0
5
4 - 5 3 cn
3 2
I
0
EXPERIMENT 1 EXPERIMENT 2
li I II Ill IV v GROUP
2or a a
15 - E
I
. F 10 E?
5
0 I I1
b T
I
Ill IV v GROUP
2500
2000
I500
1000
500
0
2 iil 0 I I
1 0.5 i 0.4 301 25 i
6 IV
GROUP
0.5
40.4 - E
0.3 3
0.2
0. I
0.0
c .-
-
II Ill IV v GROUP
Figure 2. Effects of irradiation and hormonal treatment on the serum levels of LH (open columns, top), testosterone (hatched columns, top], FSH (open columns, bottom) and inhibin (hatched columns bottom). Rats were treated as described in Table 1 and in Materials and meth- ods. Hormone levels were determined at the end of the experiments. Left panels summarize the results for the different treatment groups of Experiment 1 . Right panels summarize the results for Experiment 2. Significant differences (p < 0.05) were observed between treatment groups indicated by different superscripts.
Figure 3. Sodium dodecyl sulphate-polyacrylamide gel elec- trophoresis of testicular homogenates. Testicular fragments from animals of the different treatment groups of Experiment 1 were homogenized and subjected to SDS-PAGE as explained in Materials and methods. Proteins were visualized by staining with Coomassie blue. Arrows indicate noticeable differences between non-irradiated and irradiated animals. Treatments (I = controls, II = irradiated, 111 = irradiated treated with GnRHa + antiandrogen, IV = irradiated treated with GnRHa + testosterone, V = irradiat- ed, treated with testosterone).
ditions, FSH may increase androgen receptor concentration in Sertoli cells (Verhoeven & Cailleau, 1988b).
One of the major aims of the experiments described here was to explore whether germ cell-depleted animals might
represent a better model than isolated and cultured testic- ular cells for the study of androgen action on somatic cells of the testis. In fact, there is increasing evidence that some effects of androgens may require appropriate interactions
C 1995 Blackwell Sciencr Ltd, i,ireniatiurinl joirnral ~ f f a n d r o l o ~ y y , 18.1
30 G. Verhoeven er a/.
Figure 4. Fluorograms of 20 electrophoretic separation of 35S methionine-labelled proteins in testicular tissue. Testicular tissue from different treatment groups was minced, incubated with 35S methionine and subjected to 2D gel electrophoresis as explained in Materials and methods. The fluorograms compare the patterns of methionine labelled protein in tissue derived from irradiated rats (Experiment 2 group 11) and irradiated animals treated with the GnRH agonist and antiandrogen (Experiment 2 group IV).
between different types of cells. Some effects of androgens on Sertoli cells, for example, may be mediated by paracrine factors (P-Mod-S) produced by underlying peritubular cells (Skinner & Fritz, 1985; Skinner et al., 1988; Verhoeven 81 Cailleau, 1988a; Verhoeven et al., 1992). Taking into account the limitations outlined in the previous paragraph, a number of approaches were used to identify suitable para- meters for androgen action in the germ cell-depleted testis. Testicular proteins were compared in the different treatment groups after SDS-PAGE. In addition, intracellular and secreted proteins were compared by two-dimensional gel electrophoresis after labelling with [ 35S] methionine. Under the conditions used, however, only limited differences were
observed even between irradiated controls and irradiated animals in which both androgens and FSH were eliminated by combined treatment with GnRHa and antiandrogen. Since only a limited set of experimental conditions were tested and since the resolution of the techniques used remains restricted, these data do not rule out the possibility that some key proteins may undergo major changes in syn- thesis or concentration under the influence of androgens (and/or FSH). In view of the marked changes in testicular weight observed, however, it seems likely that one of the main effects of androgens and FSH is an overall stimulation of protein synthesis rather than a specific induction of a lim- ited number of proteins. A similar conclusion has been reached from studies of the effects of testosterone on semi- niferous tubule fluid and protein secretion in rats at stages VI-VIII of the spermatogenic cycle (Sharpe et al., 1992, 1993; Sharpe, 1994).
Finally, we investigated the effects of irradiation and the above described hormonal treatments on the steady state levels of mRNA for transferrin and ABP. These two para- meters were selected because they have been extensively investigated both in the intact testis and in isolated Sertoli cells, and because they are generally considered to be regu- lated directly or indirectly by androgens and FSH.
In vivo, the hormonal control of ABP resembles remark- ably the hormonal control of spermatogenesis (Hansson et al., 1976; Fritz, 1978). Both androgens and FSH are required for optimal secretion. Nonetheless, ABP produc- tion does not completely disappear in Tfm rats indicating that ABP can be produced even in the absence of androgen action (Ritzkn et al . as cited in Hansson et al., 1976). In-vitro data are more controversial. In general, FSH stimulates ABP production but the effects of androgens on isolated Sertoli cells are limited (Louis & Fritz, 1979; Cailleau et al., 1984; Perez-Infante et al., 1986; Hadley et al., 1987). Interestingly, contaminating peritubular cells markedly enhance the effects of androgens on Sertoli cell ABP production (Skinner & Fritz, 1985; Verhoeven 81 Cailleau, 1988a; Swinnen et al., 1989). In the present experiments, irradiation resulted in a two- to three-fold increase in the amount of ABP niRNA/pg RNA, probably reflecting the enrichment in Sertoli cells. In accordance with the data mentioned above, the level of ABP mRNA was comparable in Tfm rats and in animals rendered germ cell depleted by irradiation. Surprisingly, neither a drastic reduction of FSH and andro- gens (by combined administration of GnRHa and antian- drogen) nor the administration of supraphysiological amounts of testosterone provoked measurable changes in ABP mRNA/pg R N A in the irradiated animals.
The control of testicular transferrin mRNA in vivo may vary according to age. FSH may be the prevailing regulator in younger animals whereas in older rats testosterone may be the most important hormone (Hugly et al., 1988). At least part of the effects of these hormones may be indirect, how- ever, and changes in the complement of germ cells may
0 1 Y Y 5 Ulackwrll Science Ltd, rnf<,rnafional journal ofandruloyy, l 8 , l
Androgens and somatic cells of the testis 31
Table 2. Effects of irradiation and hormonal manipulation on mRNA levels for ABP and transferrin
mRNA level (units)
Group Treatment ABP transferrin
Tfm Experiment I
I II 111 IV V
Experiment 2 I II 111 IV V
Control RX RX GnRH, + Antiandrogen RX GnRH, + Testosterone RX Testosterone
Control RX RX GnRH, RX GnRH, + Antiandrogen RX LHRH, + hCG
0.97
1.06 0.77 0.91 0.78 0.75
1.34 1.04 0.97 0.8 1 0.93
8.0 (2.6)
3 .O (1.1) 9.4 (3.11
11.2 (3.41 11.1 (4.4) 13.4 (4.71
14.0 (4.51
RNA was prepared from the pooled testes of groups of rats treated as described in Table 1. Testes from rats with testicular feminization syn- drome (Tfm) were included as an additional control. The total amount of RNA recovered has been expressed as mg per g tissue. mRNA levels for ABP and transferrin were determined by dot blot hybridization as described in Materials and methods and have been expressed in arbitrary densitometric units. Values between brackets have been normalized for 18s RNA after hybridization of h e same blots with an 18s rRNA probe.
Figunt 5. Northern blot analysis of testicular RNA from the dif- ferent treatment groups. Total RNA was isolated and Northern blot analysis was performed on RNA samples (20 pg/lane) from the different treatment groups separated an a 1% agarose gel. Testicular RNA from animals with testicular feminization syn- drome (Tfm) was included as an additional control. One blot was probed, successively with transferrin cDNA (upper panel) and ABP cDNA (middle panel) as explained in Materials and meth- ods. A second blot was probed for 18s rRNA to visualize even- tual differences in the amount of RNA per lane (lower panel). Treatments: Experiment 1 (I = controls, II = irradiated, Ill = irra- diated treated with GnRHa + antiandrogen, IV = irradiated treat- ed with GnRHa + testosterone, V = irradiated, treated with testos- terone).Treatments: Experiment 2 (1 = control, I I = irradiated, 111 = irradiated treated with GnRHa, IV = irradiated treated with GnRHa + antiandrogen, V = irradiated treated with GnRHa + hCG).
influence Sertoli cell transferrin production (Hugly et a/. ,
1988; Roberts et a/., 1991). Ivl vitro, FSH and testosterone have been reported to have limited effects on Sertoli cell transferrin secretion or to have no effect at all (Skinner & Griswold, 1982; Perez-Infante et a/., 1986; Hadley et a/. ,
1987; Huggenvik et a/., 1987; Janecki & Steinberger, 1987; Swinnen e ta / . , 1989, 1990; Spalivieri & Handelsman, 1991). Peritubular cells, however, produce under the influence of androgens a paracrine mediator ‘P-Mod-S’ which enhances Sertoli cell transferrin production markedly (Skinner &

32 0. Verhoeven eta/.
Fritz, 1985; Skinner et al., 1988, 1989; Swinnen et al., 1989, 1990). In addition, round spermatids produce a paracrine agonist that also stimulate Sertoli cell transferrin secretion (Stallard & Griswold, 1990; Onoda & Djakiew, 1993). In the experiments described here irradiation increased testicu- lar transferrin mRNA by three- to six-fold. The relatively higher impact of irradiation on transferrin mRNA than on ABP mRNA is remarkable since both mRNAs are suppos- edly produced by Sertoli cells and accordingly the relative enrichment should be identical. This suggests that elimina- tion of the germ cells affects one of the mRNAs more than the other one or that the hormonal changes after irradiation affect both parameters differently. In contrast to ABP, com- bined treatment with GnRHa and antiandrogen reduced transferrin mRNA markedly and this effect was reversed partially by administration of high amounts of androgens. These data indicate that, in germ cell-depleted animals, transferrin mRNA is a better parameter of androgen and FSH action than is ABP mRNA. Nonetheless, it should be evident that even for transferrin the observed changes in steady state mRNA induced by androgens and FSH are maximally only two- to three-fold.
In conclusion, in germ cell-depleted rats withdrawal or restoration of androgens and FSH induces parallel changes in testicular weight and R N A content. Nonetheless, for two niRNAs that are obviously affected by these hormones in intact rats we could demonstrate only a limited (transferrin) or no (ABP) selective change in mRNA levels. At least three conclusions seem to be warranted from these data. First, the effects of these hormones on ABP and transferrin mRNA
may be mediated or modulated by germ cells. Second, although there may obviously be other messengers that exhibit more direct or more pronounced responses to androgens and/or FSH it is clear that one of the major effects of these hormones on the somatic component of the testis is an overall increase in R N A synthesis. This confirms the impression derived from the above discussed studies on testicular protein synthesis and is in accordance with data from other laboratories showing that both androgens and germ cells may cause a substantial overall increase in protein secretion by seminiferous tubules (Sharpe et al., 1992, 1993; Sharpe, 1994). Third, it is obvious that the absence of prominent changes in selected mRNAs or proteins and the intermingling of effects of androgens and FSH limit the attractiveness of germ cell-depleted rats treated with GnRHa as a starting material for subtraction cloning exper- iments aiming to isolate androgen-induced niRNAs in somatic cells.
Acknowledgments We thank Dr A. F. Parlow and the NIDDK for LH and
FSH RIA reagents. The help of Professor Dr E. van der Schueren and Mr W . Landuyt with the radiation experi- ments is kindly appreciated. Part of this research was sup- ported by a grant 'Geconcerteerde Onderzoeksactie van de Vlaamse Gemeenschap' and by a grant 3.0048.94 from the 'Nationaal Fonds voor Wetenschappelijk Onderzoek van Belgie' (N.F.W.O.). Dr J. V. Swinnen is a senior research assistant of the N.F.W.O.
References
Aldred, A. R., Howlett, G. J . & Schreiber, G. (1984) Synthesis of rat transferrin in Eschericliia coli containing a recombinant bacte- riophage. Bioheniical arid Biophysical Research Conrmrrriicatiows, 122, 960-965.
Behre, H. M., Nashan, D., Hubert, W. & Nieschlag, E. (1992) Depot gonadotropin-releasing hormone agonist blunts the androgen-induced suppression of spemiatogenesis in a clinical trial of male contraception. J o u r d qf Cliriical Eirdocririology arid
Bhasin, S., Fielder, T. J. & Swerdloff, R. S. (1987) Testosterone selectively increases follicle-stimulating hormone (FSH) but not luteinizing hormone (LH) in gonadotropin-releasing honnone antagonist-treated male rats: evidence for differential regulation of LH and FSH secretion. Biology cf Rcprodircriorr, 37, 55-59.
Cailleau, J., Heyns, W. & Verhoeven, G. (1984) A filter disc assay for the measurement and characterization of androgen-binding protein in unconcentrated media from Sertoli cell-enriched cultures. Jxrrrial gf Steroid Biodiettiistry, 21, 691-696.
Chandolia, K. K., Weinbauer, G. F., Fingscheidt, U., Bartlett, J. M. S. & Nieschlag, E. (1991) Effects of flutaniide on testicular involution induced by an antagonist of gonadotropin-releasing hornione and on stimulation of spermatogenesis by follicle- stimulating hormone in rats. Joirmal qf Reprodircrioir atid Fertility,
hfeldJO[i5nl, 74, 84-90.
93, 31 3-323.
Chirguin, J. M., Przybyla, A. E., MacDonald, R. J. & Rutter, W. J. (1979) Isolation of biologcally active nbonucleic acid from sources enriched in ribonuclease. Biochemistry, 18, 5294-5299.
Fakunding, J. L., Tindall, D. J., Dedman, J. R., Mena, C. R. 81 Means, A. R. (1 976) Biochemical actions of follicle-stimulating hormone in the Sertoli cell of the rat testis. Endocrinology, 98,
Fritz, I . B. (1978) Sites of action of androgens and follicle stimu- lating hormone on cells of the seminiferous tubule. In: Biochemical Acrioris of Hormones, Vol. 5 (ed. G. Litwack), pp. 249-281. Academic Press, New York
Iadley, M. A,, Djakiew, D., Byers, S. W. & Dym, M. (1987) Polarized secretion of androgen-binding protein and transferrin by Sertoli cells grown in a bicameral culture system. Eridocririology, 120, 1097-1 103.
Iansson, V., Calandra, R., Purvis, K., Ritzen, M. & French, F. S. (1976) Hormonal regulation of spermatogenesis. Vitamiws arid Horniories, 34, 187-214.
Iendriks, L., Van de Peer, Y., Van Herck, M., Nee&, J.-M. & De Wachter, R. (1990) The 18s ribosomal RNA sequence of the sea anemone' Anemonia sulcata and its evolutionary position among other eukaryotes. FEBS Letters, 269, 445-449.
Huggenvik, J . I., Idzerda, R. L., Haywood, L., Lee, D. C., McNight, G. S . & Griswold, M. D. (1987) Transferrin messen-
392-402.

Androgens and somatic cells of the testis 33
ger ribonucleic acid: molecular cloning and hormonal regula- tion in rat Sertoli cells. Endocrinology, 120, 332-340.
Hugly, S . , Roberts, K. & Griswold, M. D. (1988) Transferrin and sulfated glycoprotein-2 messenger ribonucleic acid levels in the testis and isolated Sertoli cells of hypophysectoniized rats. Endocririology, 122, 1390-1396.
Janecki, A. & Steinberger, A. (1987) Bipolar secretion of androgen- binding protein and transferrin by Sertoli cells cultured in a two-compartment culture chamber. Endocrinology, 120, 291 -298.
Laemmli, U. K. (1970) Cleavage of structural proteins during the actembly of the head of bacteriophage T4. Nature, 227, 680-685.
Laniberts, S. W. J., Uitterlinden, P. & cie Jong, F. H . (1988) Rat prostatic weight regression in reaction to ketoconazole, cypro- terone acetate, and R U 23908 as adjucts to a depot formulation of gonadotropin-releasing hormone analogue. Cancer Research,
Louis, B. G. & Fritz, I. €3. (1979) Follicle stimulating hormone and testocterone independently increase the production of androgen binding protein by Sertoli cells in culture. Eridocrirzo/ogy, 104, 454-461.
Lonnerberg, P., Soder, O., Parvinen, M., RitzCn, E. M. & Persson, H. (1992) Beta-nerve growth factor influences the expression of androgen-binding protein messenger ribonucleic acid in the rat testis. Biolqy of Rcpodrrctiorr, 47, 381-388.
Milk, N . C . (1990) Androgen effects on Strrtoli cells. Irzternatiowal Jorrrrial of Aridrology, 13, 123-134.
O’Farell, P. H. (1 975) High resolution two-dimensional elec- trophoresis of proteins. j o u r n a l of Bio/ogica/ Chemistry, 250,
Onoda, M. & L)jakiew, D. (1993) A 29,000 Mr protein derived froni round cpermatids regulates Sertoli cell secretion. Molecular arid Cellirlar Eridocritiolopy, 93, 53-61.
I’arvinen, M. (1 972) Regulation of the seminiferous epithelium. Ew/ocrim, Reviews, 3, 404-417.
Perez-Infante, V., Bardin, C. W., Gunsalus, G. L., Musto, N. A., Rich, K . A. & Mather, J. P. (1986) Differential regulation of testicular transferrin and androgen-binding protein secretion in primary cultures of rat Sertoli cells. Etid(~cviriology, 118, 383-392.
Ibynaud, J . P., Bonne, C., Bouton, M. M., Lagace, L. & Labire, F. (1 979) Action of a nun-steroid anti-androgen RU23908 in peripheral and central tissues. Jortmal ($Steroid Bior/ze??fktry, 11, 93-99.
llhea, M. A., Weinbauer, G. F., Marshall, G. R. & Nieschlag, E. (1 986) Testosterone stimulates pituitary and serum FSH in GnRH antagonist-suppressed rats. Acta Ewdocritiologica, 113, 487-492.
Kobertc, K. P., Awoniyi, C. A., Santulli, I<. & Zirkin, B. R. (199 I ) Kegulation of Sertoli cell transferrin and sulfated glyco- protein- 2 messenger ribonucleic acid levels during the restora- tion of spermatogenesis in the adult hypophycectoinized rat. G i d o c r i r i o k ~ p y , 129, 341 7-3423.
Robertson, D. M., Hayward, S., Jacobcen, I . J., Clarke, L., McLachlan, I<. I . & de Kretser, D. M. (1988) Kadioinimunoassay of rat serum inhibin: changes after PMSG ctiniulation and gonadectomy. Molecular arid Cellirlar Etidocririolq y , 58, 1-8.
Konibauts, L., Vanniontfort, D., Verhoeven, G. & Decuypere, E. (1992) Ininiunoreactive inhibin in plasma, amniotic fluid, and gonadal tissue of male and female chick embryos. Bioicy)’ of Rrpr(~dirc-tiori, 46, 121 1-1216.
48, 6063-6068.
4007-4031.
Sambrook, J.. Fritsch, E. F. & Maniatis, R. (1989) In: Molecular Cloning: A Ldboratory Manual. Cold Spring Harbor, NY:Cold Spring Harbor Laboratory.
Sharpe, K . M. (1986) Paracrine control of the testis. Clinics in Endocrinology arid Metabolirm, 15, 185-207.
Sharpe, R. M. (1994) Regulation of spermatogenesis. In: The Physiology of Reproduction (eds F. Ktiobil and J. P. Neill), pp. 1363-1434. Raven Press Ltd, New York.
Sharpe, R. M., Maddocks, S., Millar, M., Saunders, P. T. K., Kerr, J. B. & McKinell, C. (1992) Testosterone and spermatogenesis: Identification of stage-dependent, androgen-regulated proteins secreted by adult rat seminiferous tubules. jourrzal of Arrdrology,
Sharpe, R. M., Millar, M. & McKiiinell, C. (1993) Relative roles of testosterone and the germ cell complement in determining stage-dependent changes in protein secretion by isolated rat seminiferous tubules. International Journal of Arzdrolqyy, 16,
Skinner, M. K. & Fritz, I. B. (1985) Testicular peritubular cells secrete a protein under androgen control that modulates Sertoli cell functions. Proceedirigs uf National Academy of Sciences USA,
Skinner, M. K. & Griswold, M. D. (1982) Secretion of testicular transferrin by cultured Sertoli cells is regulated by hormones and retinoids. Biology uffeproduction, 27, 21 1-221.
Skinner, M. K., Fetterolf, P. M. & Anthony, C. T. (1988) Purification of a paracrine factor, P-Mod-S, produced by testic- ular peritubular cells that modulates Sertoli cell function. journal $Biological Chemistry, 263, 2884-2890.
Skinner, M., Schlitz, S. M . & Anthony, C. T . (1989) Regulation of Sertoli cell differentiated function: testicular transferrin and androgen-binding protein expression. Endocrinology, 124,
Spalivieri, J. A. & Handelsnian, D. J. (1991) Effect of epidermal and insulin-like growth factors on vectorial secretion of transferrin by rat Sertoli cells In vitro. Molecular arid Cellular Endocrinology,
Stallard, B. J. & Griswold, M. D. (1990) Germ cell regulation of Sertoli cell transferrin mRNA levels. Molecular Errdocrinology, 4,
Steinberger, E. (1971) Hormonal coiitrol of mammalian spermato- genesis. Physiolo,@cal Review, 51, 1-22.
Swinnen, K., Cailleau, J., Heyns, W. & Verhoeven, C. (1989) Stronial cells from the rat proctate secrete androgen-regulated factors which modulate Sertoli cell function. Molecular and Cellular Eridocririo/osp, 62, 147-1 52.
Swinnen, K., Cailleau, J., Heyns, W. & Verhoeven, G. (1990) Prostatic stromal cells and testicular peritubular cells produce siniilar paracrine mediators of androgen action. Eridorriinology,
Vanderschueren, D., Van Herck, E., Suiker, A. M. H., Visser, W. J., Schot, L. P. C., Chung, K., Lucas, R. S., Einhorn, A. & Bouillon, R. (1993) Bone and inineral metabolism in the androgen-resistant (testicular feminized) male rat. Joitrrial qf Bowe atrd Mirierd Research, 8, 801 -809.
Vanderstichele, H., Eechaute, W . & Lacroix, E. (1990) Regulation of the pituitary 5a-reductase activity by gonadotropin releasing hormone and testosterone in the adult male rat. ]ourrial of Steroid BioclzeiTiistry a i d Molecirlar Bi~~logy, 35, 575-581.
Verhoeven, G. (1 992) Androgens and the testis. Verlmidelirr,qeri v ~ i i de Kortiiik/@c Acudetizie uoor G e r i e e d a d e vmi Belgie, 54,
13, 172-184.
71-81.
82, 114-118.
3015-3024.
81, 05-104.
393-401.
126, 142-1 50.
299-327.

34 G. Verhoeven et a/.
Verhoeven, G. & Cailleau, J. (1988a) Testicular peritubular cells secrete a protein under androgen control that inhibits induction of aroniatase activity in Sertoli cells. Endocrinology, 123, 2 1 00-2 1 10.
Verhoeven, G. & Cailleau, J. (1988b) Follicle-stimulating hormone and androgens increase the concentration of the androgen receptor in Sertoli cells. Endocrinology, 122, 1541-1550.
Verhoeven, G., Koninckx, P. & De Moor, P. (1982) Androgen and progestogen production in cultured interstitial cells derived from immature rat testis. joumal of Steroid Biochemistry, 17, 319-330.
Verhoeven, G., Swinnen, K., Cailleau, J., Deboel, L., Kombauts, L. & Heyns, W. (1992) The role of cell-cell interactions in androgen action. journal of Steroid Biochemistry and Molecular Biology, 41, 487-494.
Yarbrough, W . G., Quamiby, V. E., Siniental, J. A,, Joseph, D. R., Sar, M., Lubahn, D. B., Olsen, K. L., French, F. S. & Wilson, E. M. (1990) A single base mutation in the androgen receptor gene causes androgen insensitivity in the testicular feminized rat. journal of Biological Chemistry, 15, 889343900,
Received 30 Augus t 1994; accepted 17 October 1994





























