Embed Size (px)
Citation preview

Dubreucq et al.
1
Ventral Tegmental Area Cannabinoid Type-1 Receptors Control Voluntary Exercise Performance
Supplemental Information
Supplemental Methods
Animals
C57BL/6N mice were from Janvier (France) whilst male CB1 receptor mutant and
wild-type animals were bred at the NeuroCentre Magendie. All mice were housed individually
with food and water ad libitum and exposed to a regular 12-h light/dark cycle with lights on at
07:00 h. Mutant and wild-type mice, bred in a mixed genetic background with a predominant
C57BL/6N contribution, were obtained, maintained, and genotyped/regenotyped as
described previously (1-6).
Muscle Mitochondrial Respiration
Soleus muscles from GABA-CB1+/+ and GABA-CB1
-/- mice were dissected out for
subsequent mitochondrial respiration assays according to Kuznetsov et al. (7). Oxygen
consumption assays were performed using a Clark electrode chamber (Hansatech, Eurosep
Instruments, France), respiratory rates being monitored by the addition of 10 mM pyruvate,
10 mM malate, and 40 µM ADP. Fibers were then dried and weighed, and oxygen
consumption rates calculated as ng atom/min/mg of fiber.
Genomic DNA Analyses
GABA-CB1+/+ and GABA-CB1
-/- mice were killed by cervical dislocation before the
removal of the brain (for the immediate dissection of the hippocampus, the brainstem, and
the cerebellum), and of white (gastrocnemius) and red (soleus) muscles, brown adipose
tissue, kidney, adrenal, heart, liver, spleen, pancreas, intestine, lung, gonads. Genomic DNA
was extracted by standard methods. The band corresponding to CB1 genomic deletion and
the CB1-floxed allele were detected by polymerase chain reaction (PCR) using the following

Dubreucq et al.
2
primers: 5’-GCTGTCTCTGGTCCTCTTAAA-3’, 5’-GGTGTCACCTCTGAAAACAGA-3’, 5’-
CTCCTGTATGCCATAGCTCTT-3’. The PCR was carried out using the following conditions:
95°C, 5 min; 37 x (95°C,30 sec; 55°C, 30 s; 72°C, 45 sec); 72°C, 5 min, with standard PCR
buffers.
Drug Infusion in the Ventral Tegmental Area (VTA)
For intra-VTA infusion of drugs (1 µg/side, except for JZL195 which was 3 µg/side) or
their vehicles, 4.7-mm-long injectors were connected to polyethylene tubing to Hamilton
syringes (10-µl volumes) and 250 nl/min of the drugs/vehicles were infused in each side for 2
min. This was followed by a 1-min period during which the injectors were left in place to allow
further diffusion. Thereafter, all mice were returned to their home cages. At the end of the
experiments, mice were bilaterally injected with Sky Blue before being sacrificed. Brains
were rapidly removed, placed in dry ice before storage at -80°C. Coronal sections (40 µm-
wide) were then cut using a Microm HM 500M cryostat (Microm Microtech, France), stained
with Neutral Red and observed under an Olympus SZX10 stereomicroscope (Olympus,
France). All animals with injection sites outside of the VTA were excluded from the study.
Locomotor Activity
In a first series of experiments, C57BL/6N mice were acutely injected with
rimonabant/vehicle whilst placed since one week into locomotor activity cages equipped with
infra-red sensors allowing the measurement of horizontal activity. All cages were under a
regular 12-h light/dark cycle with lights on at 07:00 h, rimonabant/vehicle being injected
between 18:00-18:30 h. Locomotor activities were then recorded from 19:00-22:00 h. In a
second series of experiments, GABA-CB1+/+ and GABA-CB1
-/- mice were housed 5 days
under the conditions described above, locomotor activities being recorded throughout whilst
in a third series of experiments, GABA-CB1+/+ mice housed under the aforementioned
conditions were acutely injected with JZL195/vehicle before activity recordings.

Dubreucq et al.
3
In Vivo Electrophysiology
Stereotaxic surgery was performed under halothane anesthesia as previously
described. Recording pipettes were inserted into the VTA with the skull flat, at the following
coordinates: -3.16 mm from bregma; 0.5 mm from midline. A glass micropipette (tip diameter
= 2-3 µm, 4-6 MΩ) filled with a 2% pontamine sky blue solution in 0.5 M NaCH3CO2 was
lowered into the VTA. Dopamine (DA) neurons were identified according to well-established
electrophysiological features (8). The extracellular potential was recorded with an
Axoclamp2B amplifier in the bridge mode. The extracellular potential amplified 10 times by
the Axoclamp2B amplifier was further amplified 100 times and filtered (low-pass filter at 300
Hz and high-pass filter at 0.5 kHz) via a differential AC amplifier (model 1700; A-M Systems,
Carlsborg, WA). Single neuron spikes were discriminated and digital pulses were collected
online using a laboratory interface and software (CED 1401, SPIKE 2; Cambridge Electronic
Design). At the end of each recording experiment, the electrode placement was marked with
an iontophoretic deposit of pontamine Sky Blue dye (-20 mA, continuous current for 12-15
min), the animals deeply anaesthetized with halothane (5%) and decapitated. Brains were
removed and snap-frozen in a solution of isopentane at -70°C. Four parameters of VTA DA
neuron impulse activity were computed over 200-second epochs after a 5-min stable
baseline period: 1) basal firing rate, 2) burst event frequency, 3) mean number of
spikes/burst, and 4) number of spontaneously active cells/track. The amount of bursting
activity was then calculated as the burst event frequency x number of spikes/burst. Bursts
were identified as discrete events consisting in a sequence of spikes such that their onset
was defined by two consecutive spikes within an interval lower than 80 msec whenever they
terminated with an interval greater than 160 msec (8).
Drugs
Except for Δ9-tetrahydrocannabinol (THC), which was stored in ethanol at -20°C, all
drugs were stocked in DMSO at -20°C before final preparation, which occured 30 min before
administration. For systemic injections, rimonabant, O-2050 or their vehicle (DMSO, final

Dubreucq et al.
4
concentration: 1.25%), and JZL195 or its vehicle (DMSO, final concentration: 3%) were
diluted in one droplet of Tween 80 and then in 0.9% NaCl (10 ml/kg). THC or its vehicle
(ethanol, final concentration: 3.7%) were diluted in two droplets of Tween 80 and then in
0.9% NaCl (10 ml/kg). For local infusions, rimonabant or its vehicle (DMSO, final
concentration: 10%) and JZL195 or its vehicle (DMSO, final concentration: 20%) were diluted
in Tween 80 (final concentration: 10%) and then in 0.9% NaCl. AM251, JWH133, and AM630
were prepared in a way similar to rimonabant, except that Chremophor EL (Sigma-Aldrich)
was used for Tween 80.

Dubreucq et al.
5
Figure S1. Polymerase chain reaction (PCR) on genomic DNA extracted from different brain
regions and peripheral tissues of wild-type GABA-CB1
+/+ mice (W) and mutant GABA-CB1
-/-
mice (M). C1, negative PCR control; C2, positive control for the floxed CB1 allele; C3,
positive control for both the CB1-floxed allele (lower band) and the excised CB1 allele (upper
band). WM: white muscle (gastrocnemius); RM: red muscle (soleus); BAT: brown adipose
tissue; KID: kidney; ADR: adrenal; HRT: heart; LIV: liver; SPL: spleen; PA: pancreas; INT:
intestine; LUN: lung; GON: gonads; HPC: hippocampus; BS: brainstem; CER: cerebellum.
CB1 flox/flox
WM RM BAT WAT KID ADR
HRT LIV SPL PA INT LUN
W M W M W M W M W M W M
CB1 flox/flox CB1 ∆flox
W M W W M W M W M W M M
W M
GON
HPC BS CER
W M W M W M
C1 C2 C3
C1 C2 C3
C1 C2 C3
CB1 ∆flox
CB1 flox/flox CB1 ∆flox
Dubreucq et al.
6
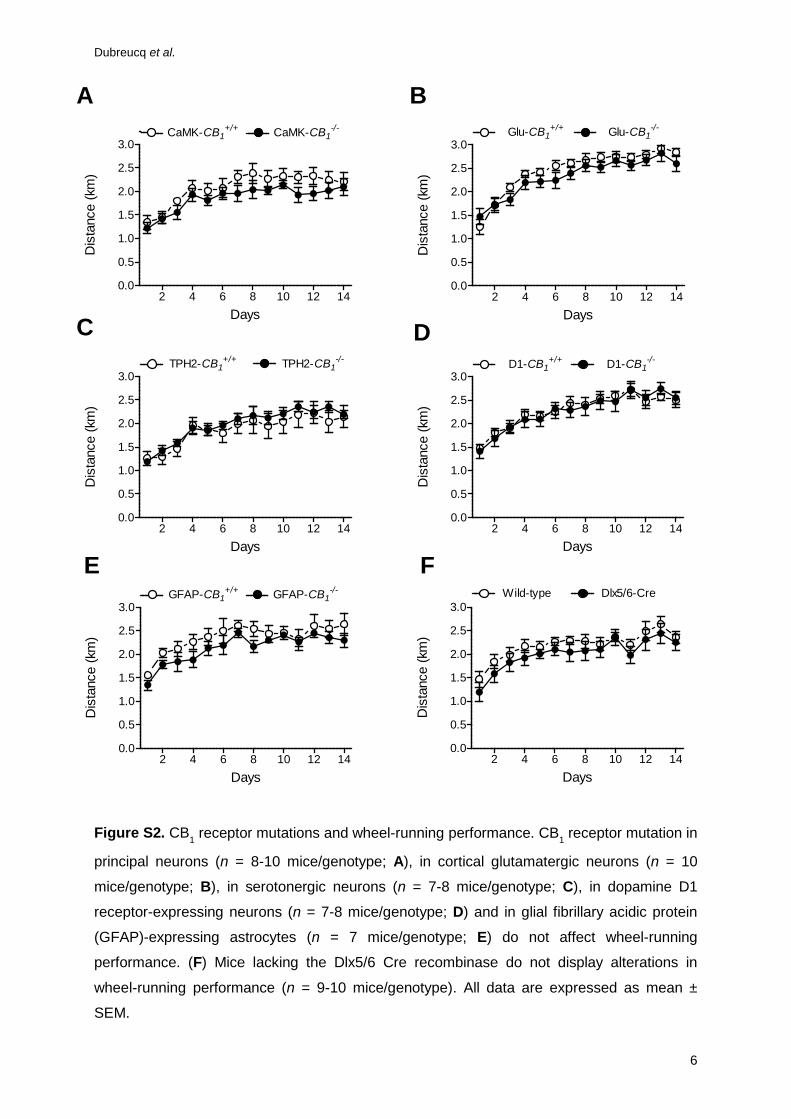
Figure S2. CB1 receptor mutations and wheel-running performance. CB1 receptor mutation in
principal neurons (n = 8-10 mice/genotype; A), in cortical glutamatergic neurons (n = 10
mice/genotype; B), in serotonergic neurons (n = 7-8 mice/genotype; C), in dopamine D1
receptor-expressing neurons (n = 7-8 mice/genotype; D) and in glial fibrillary acidic protein
(GFAP)-expressing astrocytes (n = 7 mice/genotype; E) do not affect wheel-running
performance. (F) Mice lacking the Dlx5/6 Cre recombinase do not display alterations in
wheel-running performance (n = 9-10 mice/genotype). All data are expressed as mean ±
SEM.
2 4 6 8 10 12 140.0
0.5
1.0
1.5
2.0
2.5
3.0CaMK-CB1
+/+ CaMK-CB1-/-
Days
Dis
tanc
e (k
m)
2 4 6 8 10 12 140.0
0.5
1.0
1.5
2.0
2.5
3.0Glu-CB1
+/+ Glu-CB1-/-
Days
Dis
tanc
e (k
m)
2 4 6 8 10 12 140.0
0.5
1.0
1.5
2.0
2.5
3.0TPH2-CB1
+/+ TPH2-CB1-/-
Days
Dis
tanc
e (k
m)
2 4 6 8 10 12 140.0
0.5
1.0
1.5
2.0
2.5
3.0GFAP-CB1
+/+ GFAP-CB1-/-
Days
Dis
tanc
e (k
m)
2 4 6 8 10 12 140.0
0.5
1.0
1.5
2.0
2.5
3.0Wild-type Dlx5/6-Cre
Days
Dis
tanc
e (k
m)
A B
C D
FE2 4 6 8 10 12 14
0.0
0.5
1.0
1.5
2.0
2.5
3.0D1-CB1
+/+ D1-CB1-/-
Days
Dis
tanc
e (k
m)

Dubreucq et al.
7
Figure S3. CB1 receptors in gamma-aminobutyric acid (GABA) neurons and wheel-running
performance. (A) GABA-CB1-/-
mutant mice display a decrease in their daily wheel-running
duration. (B) Maximal running speed is not affected in GABA-CB1-/-
mutant mice. All data are
expressed as mean ± SEM of 8 mice/genotype and the p value refers to the overall genotype
influence.
A B
2 4 6 8 10 12 140
30
60
90
120
150
p = .005
GABA-CB1+/+ GABA-CB1
-/-
Days
Run
ning
tim
e (m
in)
2 4 6 8 10 12 140
30
60
90
120
150GABA-CB1
+/+ GABA-CB1-/-
Days
Max
imal
spe
ed(re
volu
tions
per
min
)

Dubreucq et al.
8
Figure S4. Locations of the cannulae through which rimonabant (and its vehicle; A), AM251
(and its vehicle; B), and JZL 195 (and its vehicle; C) were perfused in the ventral tegmental
area. (D) Histology of bilateral perfusion sites in one representative mouse. Images in panels
A-C are adapted with permission from Franklin and Paxinos (9).
A B
C D

Dubreucq et al.
9
Figure S5. Numbers of dopamine (DA) neurons recorded/track (n = 28-37 tracks/mouse
group; A), DA neuronal firing rates (n = 31-39 cells/mouse group; B), and bursting scores (n
= 31-39 cells/mouse group; C) in GABA-CB1
-/- mice housed 8 days with locked wheels
(controls) or 7 days with wheels set free 3-h per day before being locked for one day
(runners). Data expressed as mean ± SEM. * p < .05 for the difference with control animals.

Dubreucq et al.
10
Figure S6. The intraperitoneal administration of rimonabant (3 mg/kg, 30 min beforehand)
affects wheel-running profile (A) and performance (B) in mice of the CD1 strain. Data are
from n = 7 mice/group, and are expressed as mean ± SEM. The p value refers to the overall
treatment effect; * p < .05 for the difference with vehicle.

Dubreucq et al.
11
Figure S7. Respective effects of the intraperitoneal administration of O-2050 (1 mg/kg, 30
min beforehand; n = 9 mice/group; A), JWH133 (10 mg/kg, 30 min beforehand; n = 7
mice/group; B) and AM630 (10 mg/kg, 30 min beforehand; n = 10 mice/group; C) on wheel-
running profile. * p < .05 for the difference with vehicle.

Dubreucq et al.
12
Figure S8. Dose-dependent effects of intraperitoneal injections with Δ9-tetrahydrocannabinol
(THC, 30 min beforehand) on wheel-running profile (A) and general running performance
(B). Data are from n = 9 mice/group, and are expressed as mean ± SEM. * p < .05 and ** p <
0.01 for the differences with vehicle.

Dubreucq et al.
13
Supplemental References
1. Marsicano G, Goodenough S, Monory K, Hermann H, Eder M, Cannich A, et al. (2003):
CB1 cannabinoid receptors and on-demand defense against excitotoxicity. Science
302:84-88.
2. Monory K, Massa F, Egertová M, Eder M, Blaudzun H, Westenbroek R, et al. (2006): The
endocannabinoid system controls key epileptogenic circuits in the hippocampus. Neuron
51:455-466.
3. Monory K, Blaudzun H, Massa F, Kaiser N, Lemberger T, Schütz G, et al. (2007) Genetic
dissection of behavioural and autonomic effects of delta9-tetrahydrocannabinol in mice.
PLoS-Biol 5, e269.
4. Bellocchio L, Lafenêtre P, Cannich A, Cota D, Puente N, Grandes P, et al. (2010):
Bimodal control of stimulated food intake by the endocannabinoid system. Nat Neurosci
13:281-283.
5. Dubreucq S, Matias I, Cardinal P, Häring M, Lutz B, Marsicano M, Chaouloff F (2012):
Genetic dissection of the role of cannabinoid type-1 receptors in the emotional
consequences of repeated social stress in mice. Neuropsychopharmacology 37:1885-
1900.
6. Han J, Kesner P, Metna-Laurent M, Duan T, Xu L, Georges F, et al. (2012): Acute
cannabinoids impair working memory through astroglial CB1 receptor modulation of
hippocampal LTD. Cell 148:1039-1050.
7. Kuznetsov, AV Veksler V, Gellerich FN, Saks V, Margreiter R, Kunz WS (2008): Analysis
of mitochondrial function in situ in permeabilized muscle fibers, tissues and cells. Nat
Protoc 3:965-976.
8. Grace AA (1988): In vivo and in vitro intracellular recordings from rat midbrain dopamine
neurons. Ann NY Acad Sci 537:51-76.
9. Franklin KBJ, Paxinos G (2008): The mouse brain in stereotaxic coordinates (3rd
Edition). New York, NY: Academic Press.





























