Embed Size (px)
Citation preview

Environmental Research 96 (2004) 163–175
ARTICLE IN PRESS
*Correspond
E-mail addr
0013-9351/$ - se
doi:10.1016/j.en
DNA adducts, benzo(a)pyrene monooxygenase activity, andlysosomal membrane stability in Mytilus galloprovincialis from
different areas in Taranto coastal waters (Italy)
M. Pisoni,a,* L. Cogotzi,a A. Frigeri,a I. Corsi,b S. Bonacci,b A. Iacocca,b L. Lancini,b
F. Mastrototaro,c S. Focardi,b and M. Sveltoa
aDepartment of General and Environmental Physiology, University of Bari, Via Amendola, 165/A, Bari 70125, ItalybDepartment of Environmental Science, University of Siena, Via delle Cerchia 3, 53100 Siena, Italy
cDepartment of Zoology, University of Bari, Via Amendola 165/A, Bari 70125, Italy
Received 23 July 2003; received in revised form 13 February 2004; accepted 27 February 2004
Abstract
The aim of this study was to investigate the impact of environmental pollution at different stations along the Taranto coastline
(Ionian Sea, Puglia, Italy) using several biomarkers of exposure and the effect on mussels, Mytilus galloprovincialis, collected in
October 2001 and October 2002. Five sampling sites were compared with a ‘‘cleaner’’ reference site in the Aeronautics Area. In this
study we also investigated the differences between adduct levels in gills and digestive gland. This Taranto area is the most significant
industrial settlement on the Ionian Sea known to be contaminated by polycyclic aromatic hydrocarbons (PAHs), polychlorinated
biphenyls, heavy metals, etc. Exposure to PAHs was evaluated by measuring DNA adduct levels and benzo(a)pyrene
monooxygenase activity (B(a)PMO); DNA adducts were analyzed by 32P-postlabeling with nuclease P1 enhancement in both
gills and digestive glands to evaluate differences between DNA adduct levels in the two tissues. B(a)PMO was assayed in the
microsomal fraction of the digestive glands as a result of the high expression of P450-metabolizing enzymes in this tissue. Lysosomal
membrane stability, a potential biomarker of anthropogenic stress, was also evaluated in the digestive glands of mussels, by
measuring the latent activity of b-N-acetylhexosaminidase. Induction of DNA adducts was evident in both tissues, although the
results revealed large tissue differences in DNA adduct formation. In fact, gills showed higher DNA adduct levels than did digestive
gland. No significant differences were found in DNA adduct levels over time, with both tissues providing similar results in both
years. DNA adduct levels were correlated with B(a)PMO activity in digestive gland in both years (r ¼ 0:60 in 2001; r ¼ 0:73 in 2002).Increases were observed in B(a)PMO activity and DNA adduct levels at different stations; no statistical difference was observed in
B(a)PMO activity over the two monitoring campaigns. The membrane labilization period in mussels from some stations was
decreased in both years. No statistical differences were established in the membrane labilization times from 2001 to 2002. Our results
suggest the existence of different sources and amounts of environmental contaminants at the stations investigated. The formation of
DNA adducts confirms the existence of activation pathways in mussels and shows the importance of DNA adduct analysis in the gill
tissue in addition to the more commonly used digestive gland; these results confirm the utility of lysosomal membrane stability as a
biomarker of general stress. Overall, the integrated use of biomarkers of exposure and the effects of environmental contaminants on
living marine organisms may help to better interpret the impact of pollutants in a marine coastal environment.
r 2004 Elsevier Inc. All rights reserved.
Keywords: DNA adducts; B(a)PMO; Lysosomal membrane stability; Taranto sea; Mytilus galloprovincialis
1. Introduction
In recent years, levels of contaminants in the marineenvironment have increased as a consequence of human
ing author. Fax: +39-080-5443388.
ess: [email protected] (M. Pisoni).
e front matter r 2004 Elsevier Inc. All rights reserved.
vres.2004.02.011
activities. The deterioration in water quality can lead toa decrease in natural resources. As recognized byinternational organizations and environmental agencies,risk assessment based on chemical analysis of environ-mental samples provides no indication of deleteriouseffects by contaminants on the biota. The use ofbiological markers or biomarkers measured at the

ARTICLE IN PRESSM. Pisoni et al. / Environmental Research 96 (2004) 163–175164
cellular or molecular level has been suggested as asensitive ‘‘early warning’’ tool for pollution monitoringand environmental quality assessment (Bayne, 1989;Gray, 1992). The selected biomarkers should indicatewhether the organisms have been exposed to pollutants(exposure biomarkers) and/or the magnitude of theorganism’s response to the pollutant (effect biomarkers)(Cajaraville et al., 2000). Mussels, such as Mytilus
galloprovincialis and other marine bivalves, have beenwidely used as sentinel organisms in pollution monitor-ing programs, due to their ability to accumulate andtolerate high concentrations of many organic andinorganic pollutants (Widdows and Donkin, 1992). Asmonitoring organisms, they present many advantagesbecause they are widespread, sessile, easily accessible,and often available in high numbers. Additionally, theyare an important component of coastal ecosystems andeven commercially important in certain regions (Ericsonet al., 2002).In the present study, the coastal marine area of the
Mar Grande, on the Gulf of Taranto in the Ionian Sea,was chosen due to the presence of important naval andindustrial activities. This area is connected via twonarrow channels to the basin of the Mar Piccolo, amarine ecosystem where biological balances have beenmodified by development. Moreover, as the Mar Piccolois a semienclosed basin, it has marked problems of waterexchange due mainly to moderate tidal ranges. Thesource of pollution in the Mar Piccolo originates mainlyfrom urban effluents, discharges from local industry,spillages from vessels, harbor operations, and presum-ably atmospheric transport. In addition, the industrialarea of Taranto holds a large industrial settlement,which is the most important steel production plant inEurope, and a petroleum refinery. Both represent highsources of heavy metals and polycyclic aromatichydrocarbons (PAHs) in the marine environmentaccording to studies by Amodio-Cocchieri et al.(1993), Storelli et al. (2000), and Storelli and Marco-trigiano (2001).PAHs are known mutagens and/or carcinogens
(Akcha et al., 2000) that require metabolic activationto exert their genotoxicity. The amount of ultimatecarcinogen produced is the result of competing activa-tion and detoxification pathways. Inefficient detoxifica-tion or increased activation of these compounds isbelieved to modulate the biologically active doseavailable for the molecular end target (DNA) and thusinfluence the levels of biomarkers reflecting exposure toand damage by genotoxic agents (Brescia et al., 1999).Some of the metabolites produced are highly reactive tointracellular nucleophiles such as DNA, leading tocovalent binding and DNA adduct formation. Thepresence of adducts may cause critical mutations andtrigger a cascade of subsequent events leading to celldeath, uncontrolled cell replication, and other cell
dysfunctions through DNA replication or erroneousrepair (Venier, 2001). As suggested by Kurelec (1993),the genotoxic effect of such widespread pollutants inaquatic invertebrates may have a great importance atthe individual, population, and ecological levels, accord-ing to the definition of ‘‘genotoxic disease syndrome.’’DNA adducts are considered sensitive biomarkers ofboth PAH exposure and PAH effect (Stein et al., 1993;Van der Oost et al., 1994) and are considered to be acumulative index of current and past exposure togenotoxic compounds (Ericson et al., 2002). In marineenvironment biomonitoring, bulky DNA adducts havebeen widely studied using the 32P-postlabeling assay(Randerath et al., 1981; Reddy and Randerath, 1986;Harvey and Parry, 1997; Pfau, 1997; Ericson et al., 2002;Akcha et al., 2003). This technique can detect oneadduct in 109–1010 bases and is useful for studying theeffects of environmental genotoxic agents of unknownstructure and concentration (Beach and Gupta, 1992).Previously, only a limited number of experiments tocompare DNA adduct formation in gills and digestivegland tissues of mussels had been conducted. Bothtissues are important sites of xenobiotic metabolism andbiotransformation and show significant structural andfunctional differences. In fact, this type of information isvery important for designing and interpreting contami-nant monitoring studies.Two enzymes are of particular interest in the
biotransformation of PAHs: cytochrome P450 (CYP)1A(phase I) and glutathione S-transferase (GST) (phase II)involved in their activation and detoxification, respec-tively.The presence of cytochrome P450 monooxygenase or
mixed-function oxygenase (MFO) system and GSTactivity have been purified and characterized in mussels,although to a limited extent compared with fish andmammals (Fitzpatrick and Sheehan, 1993; Canova et al.,1998). A previous study indicated at least five CYPforms in digestive gland of Mytilus sp. (Peters et al.,1998); moreover, multiple GST isoenzymes from diges-tive gland of M. galloprovincialis and gill of Mytilus
edulis were separated by fast protein liquid chromato-graphy (Fitzpatrick and Sheehan, 1993).The existence of distinct cytochrome P450 genes and
GST isoenzymes has been recognized (Fitzpatrick et al.,1995a, b; Wootton et al., 1995). Enzyme activity relatedto the cytochrome P450 system such as benzo(a)pyrenemonooxygenase (B(a)PMO) is significantly induced inresponse to PAHs (Akcha et al., 2000). B(a)PMOactivity has been successfully used in assessing environ-mental pollution as a specific marker of exposure toPAHs in aquatic organisms with particular regard tobivalves such as mussels (Kurelec et al., 1977; Nasciet al., 1989; Michel et al., 1993; Livingstone et al., 1997).The digestive gland, and particularly the lysosomes of
the digestive cells, are the major accumulation sites of

ARTICLE IN PRESSM. Pisoni et al. / Environmental Research 96 (2004) 163–175 165
metallic and organic contaminants and other pollutantssuch as nutrients and detergents (Domouhtsidou andDimitriadis, 2001) and they play key roles in detoxifica-tion processes and further excretion of these compounds(Moore, 1985; Viarengo et al., 1987). The accumulationof xenobiotics and their related metabolic productsinside the lysosomes weakens their membrane stabilityand may induce diffusion of hydrolytic lysosomalenzymes into the cytosol. Thus, the measurement oflysosomal membrane stability was proposed as arational biomarker of general stress in marine bivalves(Moore, 1985; Viarengo et al., 1987; Petrovic et al.,2001). Lysosomal responses to pollutants are assessedusing histochemical tests for lysosomal membranestability (Moore, 1988; Viarengo et al., 1995). It wasconsidered the most reliable of the recommendedbiomarkers in water quality assessment, but its greatestvalue emanates from its prognostic value for tissueatrophy and subsequent pathology (Moore, 1980, 1985,1988). The use of this powerful biomarker has beenevaluated in transplanted mussels (Viarengo and Canesi,1991; Regoli and Orlando, 1994; Stien et al., 1998), inlaboratory studies (Viarengo et al., 1981,1987; Harrisonand Berger, 1982), and in in vitro studies (Winston et al.,1991).In the present study, the integrated use of DNA
adducts, B(a)PMO activity, and lysosomal membranedestabilization were investigated in mussels, M. gallo-
provincialis, as biomarkers of exposure for pollutionmonitoring and water quality assessment in Tarantocoastal waters. The levels of adduct in gills and digestive
Fig. 1. Map of the study area with the six sampling stations G1, area near t
Grande); P3, aeronautics area (second inlet, Mar Piccolo); P4, ‘‘Fronte’’ zon
inlet, Mar Piccolo); P5, area near Canal Citrello (first inlet, Mar Piccolo);
Piccolo).
glands were also compared to evaluate the differentsusceptibilities of the two tissues.
2. Materials and methods
2.1. Animals and sampling
In the October 2001 and 2002 monitoring campaigns,400 wild mussels (M. galloprovincialis) were collectedfrom six stations (stations G1 and G2 on the MarGrande; stations P3, P4, P5, and P6 on the Mar Piccolo,respectively, in the second and first inlets) along theTaranto coast (southern Italy) (Fig. 1). For each station,15 pools of five entire organisms were set up accordingto size (length of valve) and sex.Gills and digestive glands were rapidly excised, frozen
in liquid nitrogen, and stored at �80�C until processedfor subsequent analyses of DNA adducts and enzymaticassay. Portions of digestive gland were also finely cut forcryosectioning as described by Moore (1988).Stations G1 and G2 were located in Mar Grande.
This is an area of considerable industrial and navalactivity and urban sources from the city of Taranto. TheMar Piccolo basin covers an area of 2176� 104m2, has awater volume of about 152� 106m3, and lies to thenorth of Taranto. It is divided by two peninsular stripsof land into the first and the second inlet and isconnected with the Mar Grande basin (Ionian Sea)through two narrow passages, the Porta Napoli Channeland the Navigabile Channel. Some surface rivers drain
he ‘‘Chiapparo’’ zone (Mar Grande); G2, area near cistern ships (Mar
e located in the vicinity of the Canal D’Aiedda sewage outlet (second
P6, ‘‘Pumping station’’ zone, steel production plant (first inlet, Mar

ARTICLE IN PRESSM. Pisoni et al. / Environmental Research 96 (2004) 163–175166
into the basin: Galeso, with a more significant flow, inthe upper part of the second inlet. Also, eight urbandischarges drain directly into the basin, the mostsignificant being Canale D’Aiedda (south, second inlet).The Mar Piccolo, being a semienclosed basin withmarked problems of water exchange due mainly tomoderate sea tides, is a marine ecosystem wherebiological balances have been modified due to urbandevelopment and, in particular, the large industrialsettlement (Storelli et al., 2000). Station P3, located inthe Aeronautics Area (Mar Piccolo, second inlet), wasconsidered to be a ‘‘cleaner’’ reference site based onprevious chemical analyses of marine sediment (L.Lopez, personal communication). Station P4 waslocated in the vicinity of the Canale D’Aiedda discharge.Sites P5 and P6 were situated in the first inlet, wherethere are different sources of PAHs, such as navaltraffic, the naval base, and the mouth of the GalesoRiver (near station P5). Station P6 was located near apumping station, along the west coast of the first inlet,that sucks about 150,000m3/h of water from the MarPiccolo to the steelworks for blast furnace cooling andgenerates an equalizing water stream from the MarGrande.
2.2. Chemicals
RNase A, RNase T1, a amylase, proteinase K,spermidine, dithiothreitol (DTT), Sevag (chloroform:i-soamylic alcohol 24:1), micrococcal nuclease, phospho-diesterase II, nuclease P1, potato apyrase, salmon spermDNA, B(a)P, disodium salt of reduced-form b-nicoti-namide adenine dinucleotide phosphate (NADPH),phenylmethylsulfonyl fluoride (PMSF), leupeptin, pep-statin, aprotinin, naphthol, polypep low viscosity, andfast violet B were purchased from Sigma-AldrichChemical; T4 polynucleotide kinase was from RocheDiagnostics; [g32]ATP was from Perkin–Elmer LifeScience; the standard adduct was from NCI ChemicalCarcinogen Standard Repository (Bethesda, MD,USA); polygram polyester sheet was from Trimitals.r.l; and aqueous mounting medium was from Glycer-gel–Daco. All other chemicals used were of analyticalgrade.
2.3. DNA extraction
DNA was purified as described in Venier and Canova(1996) with some modifications. Each pool of digestiveglands and gills was gently homogenized in 1mMEDTA, 1% SDS at pH 8 (10mL per g tissue) with aPolytron homogenizer (8–10 s) and thereafter brieflycentrifuged at 500g, at 4�C. The pellets were incubatedtwice with a mix of RNase A (600 mg/mL) and RNaseT1 (100U/mL) for 30min at 37�C. In the case ofdigestive gland samples, a-amylase was included during
the RNase treatment. Proteins were digested byproteinase K (200 mg/mL) at 45�C for 1 h. Samples werethen adjusted to 1.5% SDS and heated to 60�C for10min; then 125 mL of 5M sodium perchlorate wasimmediately added to precipitate proteins. DNA wasextracted once with Sevag (1 volume) by gently shakingat room temperature for 30min and centrifuging at14,000g, at 4�C for 15min. DNA was precipitated fromthe aqueous phase by addition of 2 volumes of cold100% ethanol with storage at �20�C overnight. Aftercentrifugation (4�C, 15min, 14,000g), ethanol wasremoved and the DNA pellet dried by evaporationunder low pressure. DNA pellets were resuspended in500 mL buffer (10mM Tris–HCl, 1mM EDTA, pH 7.4)and left to dissolve for 1 h at 37�C. Following additionof 2.5M sodium acetate (0.1 volume) and cold absoluteethanol (1 volume), DNA was precipitated again. Aftercentrifugation (4�C, 20min, 14,000g), the pellet wasdried and resuspended in 500 mL MilliQ water. TheDNA concentration was determined by measuring theabsorbance at 260 nm and the purity was checked byevaluating the absorbance ratios A260:A280 andA260:A230. The ratios were always within the range1.870.5 and B2, respectively.
2.4. 32P-postlabeling assay
Each DNA sample was precipitated as indicatedabove and resuspended in 20mM Tris–HCl, pH 7.5, toreach a final concentration of 2 mg DNA/mL. DNA washydrolyzed to deoxyribonucleoside 30-monophosphatesby treatment with micrococcal nuclease (0.32U/mgDNA) and spleen phosphodiesterase (0.003U/mgDNA) in 20mM sodium succinate, 10mM calciumchloride, pH 6 (31
2h at 37�C). Normal dNP was
dephosphorylated by incubation for 30min at 37�Cwith nuclease P1 (0.375 mg/mg DNA) in 8.5mM zincchloride, 70.8M sodium acetate, pH 5. The reaction wasstopped with 2.5 mL of 0.5M Tris–HCl, pH 9. AdducteddNP (B5 mg DNA digest), resistant to nuclease P1, waslabeled by incubation for 40min at 37�C with 2 mL of[g32]ATP (420 mCi) (specific activity 45000Ci/mmol)and 2.5U polynucleotide kinase T4 in 10mM bicine, pH9.5, 10mM magnesium chloride, 10mM DTT, 1mMspermidine. Free [g32]ATP was hydrolyzed with 0.05Uof potato apyrase (40min at 37�C). Purification andresolution of 32P-labeled adducts were carried out byTLC on polyethyleneimine–cellulose sheets.Chromatography was carried out according to a
published method (Venier and Canova, 1996) using thefollowing solvents: direction (D)1, 1M sodium phos-phate (pH 6.8); D2, 5.3M lithium formate, 8.5M urea(pH 3.5) (opposite D1); D3, 1.2M lithium chloride,0.5M Tris–HCl, 8.5M urea (pH 8) (perpendicular toD2); and D4, 1.7M sodium phosphate (pH 6) (directedas D3). TLC plates were dipped and gently shaken for

ARTICLE IN PRESSM. Pisoni et al. / Environmental Research 96 (2004) 163–175 167
15min in deionized water after each run (D1–D4). Awick of Whatman paper was stapled to the top of theplates for the D1 and D4 developments. The chromato-grams were visualized by autoradiography at �80�C for72 h, using intensifying screens. Adduct levels weredetermined by excising areas from chromatograms andmeasuring their radioactivity by Cherenkov counting.Relative adduct labeling (RAL) was calculated from theamount of radioactivity on the chromatograms, fromthe DNA amount loaded on the TLC sheet, and fromthe specific activity of the [g-32P] ATP used for labeling.A positive control of [3H](anti)B(a)PDE-DNA stan-dard, a negative control of salmon sperm DNA, and asample without DNA were routinely assayed along withexperimental samples. Samples were tested in threereplicates at least.
2.5. Isolation of microsomal fraction
Digestive gland pools of about 1 g were used forenzyme analysis prior to microsome extraction. Thetissue was finely cut and homogenized with a Potter–Elvehjem homogenizer in a sucrose buffer in 1:4 (wv�1)proportions (50mM K2HPO4, 75mM sucrose, 1mMEDTA, 0.5mM DTT, 400 mL PMSF, 10 mM leupeptin,1 mM pepstatin, 1mgmL�1 aprotin, pH 7.5) at 4�C. Thehomogenate obtained was centrifuged first at 9000g for20min at 4�C and second at 100,000g for 2 h at 4�C. Thepellet was resuspended in 10mM Tris buffer, 20% wv�1
glycerol, 0.5mM DTT, 1 mM pepstatin, 10 mM leupep-tin, 1mgmL�1 aprotinin, and 400 mM PMSF (pH 7.5,4�C) and stored at �80�C until analysis. The isolatedmicrosomal fraction was then used to determineB(a)PMO activity by Kurelec et al.’s (1977) fluorimetricmethod.
2.6. B(a)PMO activity
The B(a)PMO assay was performed using 200 mL ofmicrosomes in an incubating mixture of buffer made upof 0.11M Tris–HCl, 15mM magnesium chloride,1.8mM NADPH, and 2mM B(a)P, left to run for 1 hat 30�C, and stopped with cool n-hexane and acetone.The fluorimetric assay was carried out according to theNebert and Gelboin (1968) method, as modified byWalters et al. (1979). The activity was expressed as unitsof fluorescence min�1mg proteins�1. A Perkin–ElmerLS 50 B spectrophotofluorimeter was used; each samplewas dosed in triplicate.
2.7. Lysosomal membrane stability
The evaluation of lysosomal membrane stability wasperformed by the method of Moore (1988), measuringthe latent activity of b-N-acetylhexosaminidase. Cryo-stat tissue sections (10 mm) of digestive gland were cut in
a CM 1900 (Leica) cryostat with a cabinet temperatureof �27�C. Sections were transferred to glass slides atroom temperature and allowed to flash dry on them for1–2min. Serial cryostat sections in duplicate series weresubjected to acid labilization at room temperature in0.1M citrate buffer (pH 4.5) containing 2.5% NaCl(w:v). The acid labilization sequence involved treatingserial sections for 0–35min in 3- to 5-min intervals.Sections were then transferred to the substrate incuba-tion medium containing 20mg naphthol AS-BI-N-acetyl–b-d-glucosaminide dissolved in 2.5mL 2-meth-oxyethanol and made up to 50mL with 0.1M citratebuffer (pH 4.5) containing 2.5% NaCl (w:v) and 3.5 g oflow-viscosity polypeptide to act as a section stabilizer.Sections were incubated for 20min at 37�C in a waterbath, rinsed in 3% NaCl at 37�C for 2min, and stainedwith 1mg/mL fast violet B in 0.1M phosphate buffer(pH 7.4) at room temperature for 10min. Slides wererinsed in running tap water for 5min, rinsed in distilledwater, and mounted in aqueous mounting medium.The determination of lysosomal membrane stability
was based on the time of acid labilization treatmentrequired to produce the maximum staining intensity,according to Moore (1976). The maximum reactionproduct was assessed using an image processingprogram such as Scion Image. For statistical interpreta-tion, every section was examined by quarters in setsfrom three different animals. A mean value was thenderived for each section, corresponding to an individualdigestive gland.
2.8. Statistical analysis
Statistically significant differences were established atPo0:05 using the Student t test. Data from differentsites were compared statistically by one-way ANOVAfollowed by the Tukey test when appropriate. Correla-tion matrix was also calculated to study the relationshipbetween DNA adduct levels and B(a)PMO activity.
3. Results
3.1. DNA adduct analysis
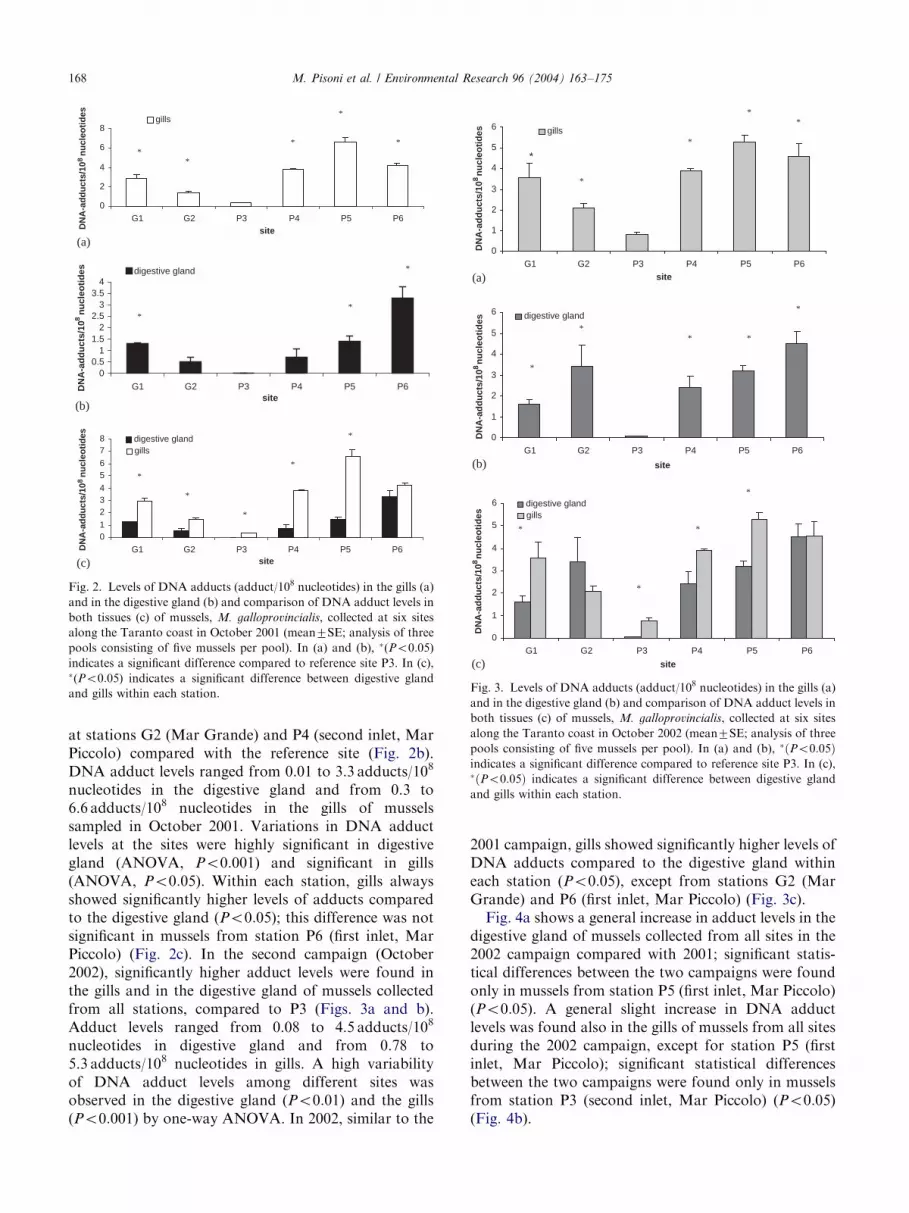
Adduct levels of mussels collected in the five sites inTaranto coastal waters were compared, in October 2001,with the reference site P3 at the second inlet (MarPiccolo), considered a cleaner area on the basis ofanalyses of PAH marine sediment (L. Lopez, personalcommunication).During the 2001 campaign, adduct levels were
significantly higher in gills from mussels collected at allstations compared to those collected at the reference site(P3) (second inlet, Mar Piccolo) (Fig. 2a) (Po0:05); inthe digestive gland, this increase was not significant only
ARTICLE IN PRESS
0
2
4
6
8
G1 G2 P3 P4 P5 P6siteD
NA
-ad
du
cts/
108
nu
cleo
tid
es
gills
00.5
11.5
22.5
33.5
4
G1 G2 P3 P4 P5 P6site
DN
A-a
dd
uct
s/10
8 nu
cleo
tid
es digestive gland
01
234
56
78
G1 G2 P3 P4 P5 P6site
DN
A-a
dd
uct
s/10
8 n
ucl
eoti
des digestive gland
gills
*
**
*
*
*
*
*
*
*
*
**
(a)
(b)
(c)
Fig. 2. Levels of DNA adducts (adduct/108 nucleotides) in the gills (a)
and in the digestive gland (b) and comparison of DNA adduct levels in
both tissues (c) of mussels, M. galloprovincialis, collected at six sites
along the Taranto coast in October 2001 (mean7SE; analysis of three
pools consisting of five mussels per pool). In (a) and (b), �(Po0:05)
indicates a significant difference compared to reference site P3. In (c),�(Po0:05) indicates a significant difference between digestive gland
and gills within each station.
0
1
2
3
4
5
6
G1 G2 P3 P4 P5 P6
site
DN
A-a
dd
uct
s/10
8n
ucl
eoti
des gills
*
0
1
2
3
4
5
6
G1 G2 P3 P4 P5 P6
site
DN
A-a
dd
uct
s/10
8n
ucl
eoti
des
digestive gland
0
1
2
3
4
5
6
G1 G2 P3 P4 P5 P6
site
DN
A-a
dd
uct
s/10
8n
ucl
eoti
des
digestive glandgills
*
*
*
*
*
*
**
*
**
*
*
(a)
(b)
(c)
Fig. 3. Levels of DNA adducts (adduct/108 nucleotides) in the gills (a)
and in the digestive gland (b) and comparison of DNA adduct levels in
both tissues (c) of mussels, M. galloprovincialis, collected at six sites
along the Taranto coast in October 2002 (mean7SE; analysis of three
pools consisting of five mussels per pool). In (a) and (b), �ðPo0:05Þindicates a significant difference compared to reference site P3. In (c),�ðPo0:05Þ indicates a significant difference between digestive gland
and gills within each station.
M. Pisoni et al. / Environmental Research 96 (2004) 163–175168
at stations G2 (Mar Grande) and P4 (second inlet, MarPiccolo) compared with the reference site (Fig. 2b).DNA adduct levels ranged from 0.01 to 3.3 adducts/108
nucleotides in the digestive gland and from 0.3 to6.6 adducts/108 nucleotides in the gills of musselssampled in October 2001. Variations in DNA adductlevels at the sites were highly significant in digestivegland (ANOVA, Po0:001) and significant in gills(ANOVA, Po0:05). Within each station, gills alwaysshowed significantly higher levels of adducts comparedto the digestive gland (Po0:05); this difference was notsignificant in mussels from station P6 (first inlet, MarPiccolo) (Fig. 2c). In the second campaign (October2002), significantly higher adduct levels were found inthe gills and in the digestive gland of mussels collectedfrom all stations, compared to P3 (Figs. 3a and b).Adduct levels ranged from 0.08 to 4.5 adducts/108
nucleotides in digestive gland and from 0.78 to5.3 adducts/108 nucleotides in gills. A high variabilityof DNA adduct levels among different sites wasobserved in the digestive gland (Po0:01) and the gills(Po0:001) by one-way ANOVA. In 2002, similar to the
2001 campaign, gills showed significantly higher levels ofDNA adducts compared to the digestive gland withineach station (Po0:05), except from stations G2 (MarGrande) and P6 (first inlet, Mar Piccolo) (Fig. 3c).Fig. 4a shows a general increase in adduct levels in the
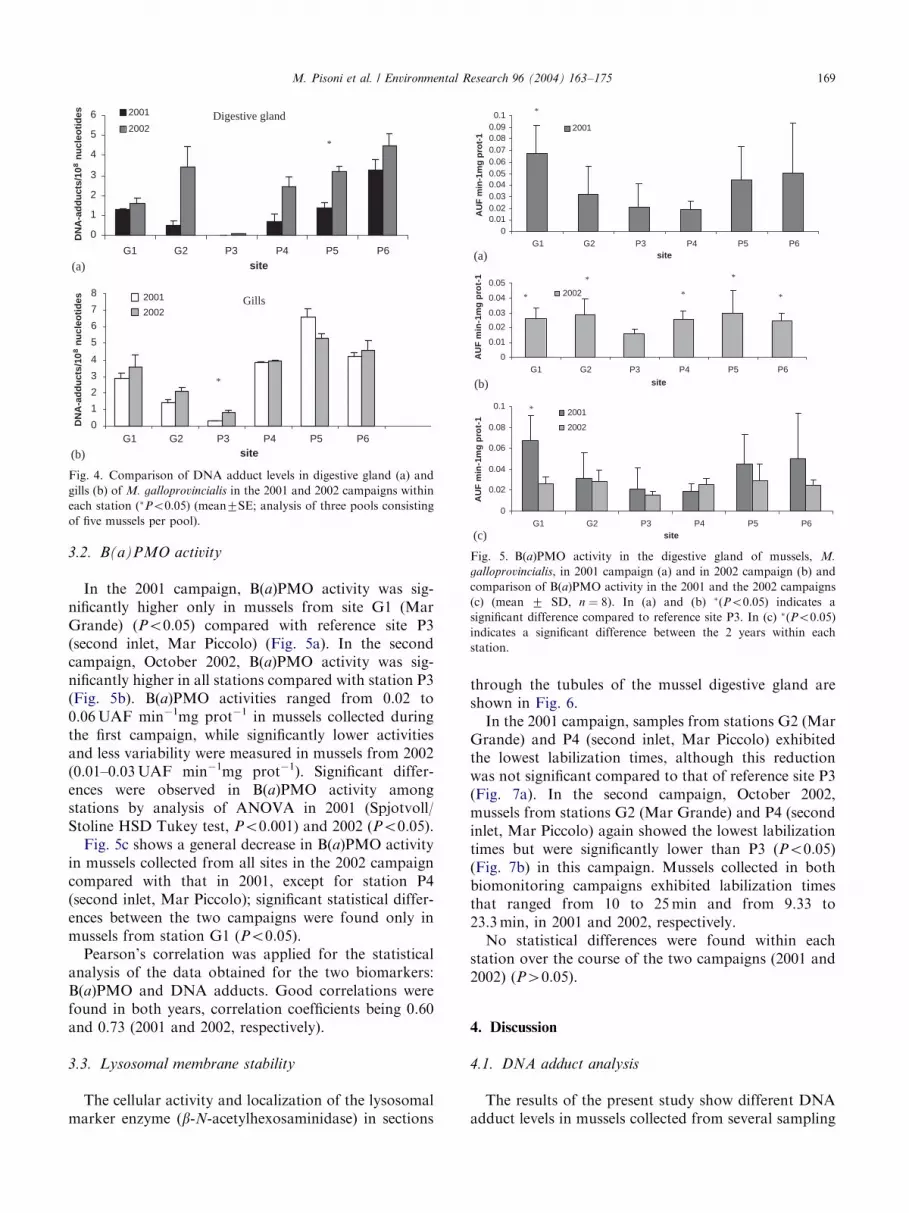
digestive gland of mussels collected from all sites in the2002 campaign compared with 2001; significant statis-tical differences between the two campaigns were foundonly in mussels from station P5 (first inlet, Mar Piccolo)(Po0:05). A general slight increase in DNA adductlevels was found also in the gills of mussels from all sitesduring the 2002 campaign, except for station P5 (firstinlet, Mar Piccolo); significant statistical differencesbetween the two campaigns were found only in musselsfrom station P3 (second inlet, Mar Piccolo) (Po0:05)(Fig. 4b).
ARTICLE IN PRESS
0
1
2
3
4
5
6
G1 G2 P3 P4 P5 P6
site
2001
2002
2001
2002
0
1
2
3
4
5
6
7
8
G1 G2 P3 P4 P5 P6site
DN
A-a
dd
uct
s/10
8n
ucl
eoti
des
*
*
Digestive gland
Gills
DN
A-a
dd
uct
s/10
8n
ucl
eoti
des
(a)
(b)
Fig. 4. Comparison of DNA adduct levels in digestive gland (a) and
gills (b) of M. galloprovincialis in the 2001 and 2002 campaigns within
each station (�Po0:05) (mean7SE; analysis of three pools consisting
of five mussels per pool).
00.010.020.030.040.050.060.070.080.090.1
G1 G2 P3 P4 P5 P6site
AU
F m
in-1
mg
pro
t-1
2001
0
0.01
0.02
0.03
0.04
0.05
G1 G2 P3 P4 P5 P6
site
AU
F m
in-1
mg
pro
t-1
2002
0
0.02
0.04
0.06
0.08
0.1
G1 G2 P3 P4 P5 P6site
AU
F m
in-1
mg
pro
t-1
2001
2002
*
*
*
*
*
*
*
(a)
(b)
(c)
Fig. 5. B(a)PMO activity in the digestive gland of mussels, M.
galloprovincialis, in 2001 campaign (a) and in 2002 campaign (b) and
comparison of B(a)PMO activity in the 2001 and the 2002 campaigns
(c) (mean 7 SD, n ¼ 8). In (a) and (b) �(Po0:05) indicates a
significant difference compared to reference site P3. In (c) �(Po0:05)
indicates a significant difference between the 2 years within each
station.
M. Pisoni et al. / Environmental Research 96 (2004) 163–175 169
3.2. B(a)PMO activity
In the 2001 campaign, B(a)PMO activity was sig-nificantly higher only in mussels from site G1 (MarGrande) (Po0:05) compared with reference site P3(second inlet, Mar Piccolo) (Fig. 5a). In the secondcampaign, October 2002, B(a)PMO activity was sig-nificantly higher in all stations compared with station P3(Fig. 5b). B(a)PMO activities ranged from 0.02 to0.06UAF min�1mg prot�1 in mussels collected duringthe first campaign, while significantly lower activitiesand less variability were measured in mussels from 2002(0.01–0.03UAF min�1mg prot�1). Significant differ-ences were observed in B(a)PMO activity amongstations by analysis of ANOVA in 2001 (Spjotvoll/Stoline HSD Tukey test, Po0:001) and 2002 (Po0:05).Fig. 5c shows a general decrease in B(a)PMO activity
in mussels collected from all sites in the 2002 campaigncompared with that in 2001, except for station P4(second inlet, Mar Piccolo); significant statistical differ-ences between the two campaigns were found only inmussels from station G1 (Po0:05).Pearson’s correlation was applied for the statistical
analysis of the data obtained for the two biomarkers:B(a)PMO and DNA adducts. Good correlations werefound in both years, correlation coefficients being 0.60and 0.73 (2001 and 2002, respectively).
3.3. Lysosomal membrane stability
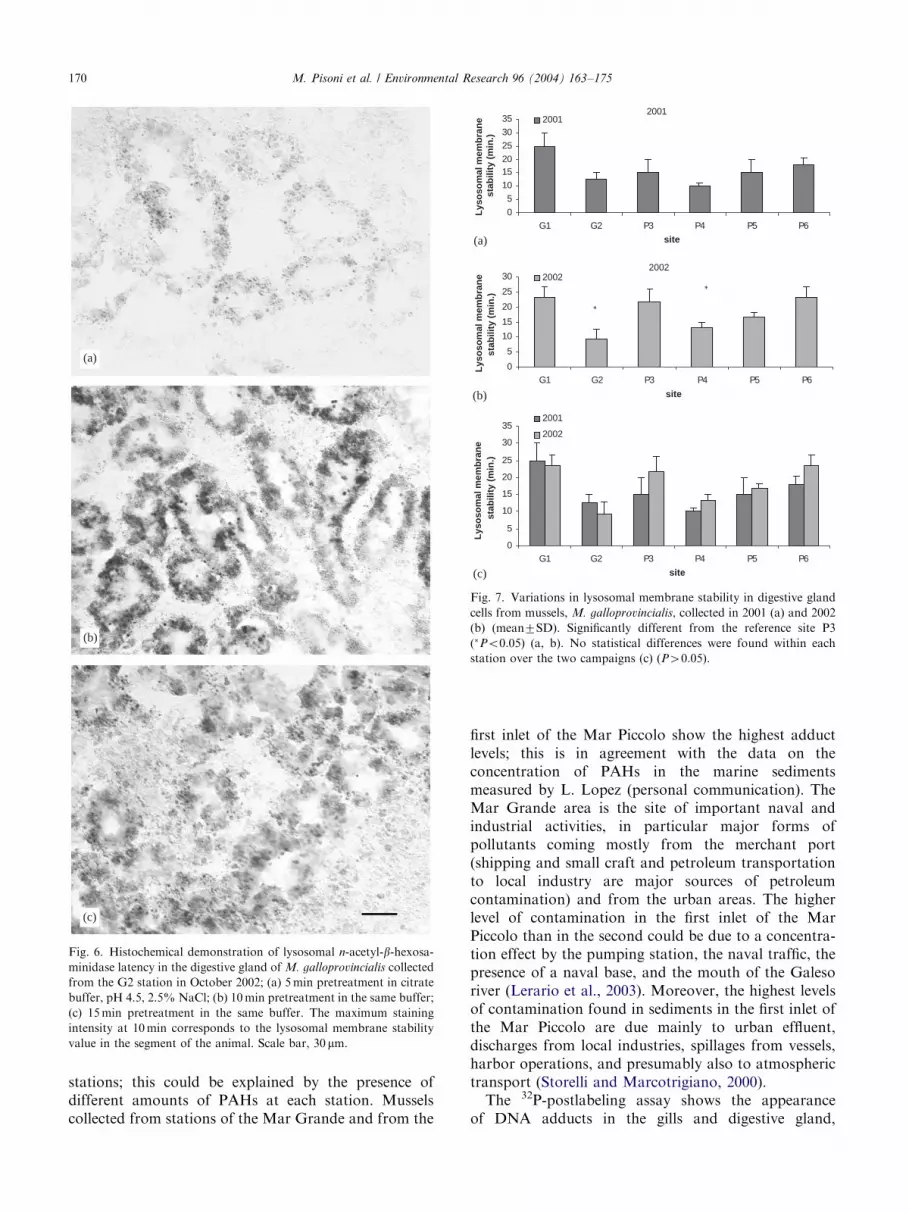
The cellular activity and localization of the lysosomalmarker enzyme (b-N-acetylhexosaminidase) in sections
through the tubules of the mussel digestive gland areshown in Fig. 6.In the 2001 campaign, samples from stations G2 (Mar
Grande) and P4 (second inlet, Mar Piccolo) exhibitedthe lowest labilization times, although this reductionwas not significant compared to that of reference site P3(Fig. 7a). In the second campaign, October 2002,mussels from stations G2 (Mar Grande) and P4 (secondinlet, Mar Piccolo) again showed the lowest labilizationtimes but were significantly lower than P3 (Po0:05)(Fig. 7b) in this campaign. Mussels collected in bothbiomonitoring campaigns exhibited labilization timesthat ranged from 10 to 25min and from 9.33 to23.3min, in 2001 and 2002, respectively.No statistical differences were found within each
station over the course of the two campaigns (2001 and2002) (P40:05).
4. Discussion
4.1. DNA adduct analysis
The results of the present study show different DNAadduct levels in mussels collected from several sampling
ARTICLE IN PRESS
(a)
(b)
(c)
Fig. 6. Histochemical demonstration of lysosomal n-acetyl-b-hexosa-minidase latency in the digestive gland of M. galloprovincialis collected
from the G2 station in October 2002; (a) 5min pretreatment in citrate
buffer, pH 4.5, 2.5% NaCl; (b) 10min pretreatment in the same buffer;
(c) 15min pretreatment in the same buffer. The maximum staining
intensity at 10min corresponds to the lysosomal membrane stability
value in the segment of the animal. Scale bar, 30 mm.
2001
0
5
10
1520
25
30
35
G1 G2 P3 P4 P5 P6
site
site
site
Lys
oso
mal
mem
bra
ne
stab
ility
(m
in.)
2001
2002
0
5
10
15
20
25
30
G1 G2 P3 P4 P5 P6
Lys
oso
mal
mem
bra
ne
stab
ility
(m
in.)
2002
0
5
10
15
20
25
30
35
G1 G2 P3 P4 P5 P6
Lys
oso
mal
mem
bra
ne
stab
ility
(m
in.)
2001
2002
*
*
(a)
(b)
(c)
Fig. 7. Variations in lysosomal membrane stability in digestive gland
cells from mussels, M. galloprovincialis, collected in 2001 (a) and 2002
(b) (mean7SD). Significantly different from the reference site P3
(�Po0:05) (a, b). No statistical differences were found within each
station over the two campaigns (c) (P40:05).
M. Pisoni et al. / Environmental Research 96 (2004) 163–175170
stations; this could be explained by the presence ofdifferent amounts of PAHs at each station. Musselscollected from stations of the Mar Grande and from the
first inlet of the Mar Piccolo show the highest adductlevels; this is in agreement with the data on theconcentration of PAHs in the marine sedimentsmeasured by L. Lopez (personal communication). TheMar Grande area is the site of important naval andindustrial activities, in particular major forms ofpollutants coming mostly from the merchant port(shipping and small craft and petroleum transportationto local industry are major sources of petroleumcontamination) and from the urban areas. The higherlevel of contamination in the first inlet of the MarPiccolo than in the second could be due to a concentra-tion effect by the pumping station, the naval traffic, thepresence of a naval base, and the mouth of the Galesoriver (Lerario et al., 2003). Moreover, the highest levelsof contamination found in sediments in the first inlet ofthe Mar Piccolo are due mainly to urban effluent,discharges from local industries, spillages from vessels,harbor operations, and presumably also to atmospherictransport (Storelli and Marcotrigiano, 2000).The 32P-postlabeling assay shows the appearance
of DNA adducts in the gills and digestive gland,

ARTICLE IN PRESSM. Pisoni et al. / Environmental Research 96 (2004) 163–175 171
confirming M. galloprovincialis’s ability to biotransformand detoxify genotoxic compounds (Venier and Canova,1996; Akcha et al., 2000). Many studies have investi-gated DNA adduct formation in mussels following fieldand intentional exposure to genotoxic compounds: asignificant increase in adduct levels was shown, althoughnever exceeding 1 RAL, in the gills of mussels from anindustrial district compared with adduct levels from lesspolluted sites in the Venice lagoon, and a markedpresence of DNA adducts was detected in the digestivegland of mussels from contaminated sites on theGalician coast (La Coruna, North Spain) (Venier,2001); negative findings have been described for dabliver, where DNA adducts did not appear to beassociated with PAH exposure (Akcha et al., 2003).Pollution-related DNA adducts have only occasionallybeen reported in mussels environmentally exposed to oilrefinery waste-waters (Venier and Canova, 1996). Withregard to studies in mussels following laboratoryexposure to genotoxic compounds, the majority showa positive relationship between adducts and contamina-tion levels (Canova et al., 1998; Akcha et al., 2000;Ching et al., 2001; Skarphethinsdottir et al., 2003).
4.2. Tissue differences of DNA adducts
Our results revealed tissue-specific differences inadduct formation; in fact, higher DNA adduct levelswere found in gills than in digestive glands at all sites inboth years, except for G2, in 2002. This study is the firstto report on the tissue-specific differences in DNAadduct formation in mussels, M. galloprovincialis, notexposed in laboratory to B(a)P. Our results are inagreement with the studies presented by Ericson et al.(2002) on mussels, M. edulis, collected from contami-nated sites and by Skarphethinsdottir et al. (2003) onmussels exposed experimentally to B(a)P. The gillsprovide a large adsorption surface and filter largeamounts of water containing dissolved contaminantsand suspended particulate matter, whereas the digestivegland is a major site of organic xenobiotic metabolism.Significant increases in DNA adducts proportional tothe dose received have been detected in B(a)P-treatedgills of Mediterranean mussels (Venier and Canova,1996), in the hepatopancreas of Perna viridis (Chinget al., 2001), in the digestive glands of M. galloprovin-
cialis (Harvey and Parry, 1997; Akcha et al., 2000), andin the gills but not in the digestive glands of Baltic Seablue mussels (M. edulis) (Skarphethinsdottir et al.,2003). Only slight differences in adduct levels have beenfound in the gills and digestive glands of Mediterraneanmussels (Canova et al., 1998). Disagreements betweenstudies could be explained by differences in the rates ofdistinct metabolic reactions and DNA repair in the twotissues.
Although DNA repair systems have previously beenreported in mussels (Accomando et al., 1999), virtuallynothing is known of specific enzymes or other systemsthat may be involved in the repair or removal ofcovalently bound DNA adducts.With regard to oxidative lesion (8-OHdG) quantifica-
tion, basal levels of oxidative DNA lesions were higherin the digestive glands than in the gills, emphasizing theexistence of structural and functional differences be-tween the two tissues (e.g., lipid content, endogenousperoxidation rate, array of metabolic reaction). Higheramounts of endogenous oxidation and an overestima-tion of 8-OHdG levels in the lipid-rich tissue (digestivegland) could explain the above evidence (Canova et al.,1998). Additionally, it depends on a large degree on thecomplicated balance between activation (phase I) anddetoxification (phase II) processes (Akcha et al., 2000).For example, GST activity (phase II) in gills appears tobe indirectly inhibited by B(a)P exposure, whereas thesame activity remains unchanged in the digestive gland.Competition between endogenous metabolites for GSTand those produced by B(a)P biotransformation couldbe responsible for the inhibition of GST that results in abioaccumulation of reactive electrophilic intermediatesin the tissue, instead of it being bound to glutathioneand then removed. This may explain the increase inB(a)P-related DNA adduct formation in gills comparedwith digestive gland. A possible variation in the levels ofparticular GST isoenzymes was investigated in thedigestive gland and gills of mussels in sampling sitesrelative to pollution status (Fitzpatrick et al., 1995a, b).Moreover, it is interesting to note that field studies
indicate at least five different digestive gland CYP formsin mussels (Peters et al., 1998). Whether this property ofthe digestive gland MFO system also exists in musselgills is not known. Moreover, in vitro data indicate asaturation of B(a)P metabolism by digestive glandmicrosomes (Vmax) at 60–100 mM B(a)P, since in bothtissues the internal dose detected may have exceeded thelimiting rate of B(a)P-activating reactions (Canova et al.,1998). Several studies indicate seasonal variation inP450 and GST enzyme activities, both in the digestivegland and in the gills, even though this is not the mainfactor for the observed tissue differences (Skarphethins-dottir et al., 2003).The absence of statistical differences between the
adduct levels of mussels collected in 2001 and those in2002 in both tissues suggests that the amount ofenvironmental pollution in the Taranto area does notchange over the course of 1 year.
4.3. Response of the enzymatic biomarker
P450 monooxygenase is a multigene enzyme super-family involved in the oxidative metabolism of bothendogenous and exogenous compounds. CYP-dependent

ARTICLE IN PRESSM. Pisoni et al. / Environmental Research 96 (2004) 163–175172
monooxygenase has been identified in at least 21 speciesof mollusks and has attracted particular attention due toits key role in the biotransformation of many foreigncompounds, including dioxins, furans, polychlorinatedbiphenyls (PCBs), and PAHs (Livingstone, 1991).Among the biotransformation enzymes used in pollu-tion monitoring in aquatic invertebrates, B(a)PMO hasalready been used as a biomarker of exposure to thoseclasses of toxic compounds, and the existence of aCYP1A-like gene in mussels (Porte et al., 1991; Woottonet al., 1995) justifies the investigation of B(a)PMOenzyme activation pathways previously identified invertebrates (Kurelec et al., 1977; Akcha et al., 2000).The ability of M. galloprovincialis to metabolize B(a)Phas already been discussed in previous studies and is nolonger under discussion, though its use as an exposurebiomarker in pollution monitoring programs has stillnot been completely addressed (Michel et al., 1994).Many studies have in fact reported weak correlations
between B(a)PMO activity in mussels and levels ofPAHs and PCBs in the sediment from highly contami-nated sites (Michel et al., 1994; Sole et al., 1996). Withregard to studies conducted in vivo, a correlation hasbeen observed between CYP1A-immunopositive proteinand B(a)PMO activity in digestive gland microsomes ofMytilus sp. exposed to PAHs and PCBs underlaboratory conditions (Livingstone et al., 1985; Living-stone, 1988; Michel et al., 1993, 1994; Canova et al.,1998). Similarly, recent field studies have shown positivecorrelations between this B(a)PMO activity or othercytochrome P450-related measurements and environ-mental exposure to PAHs (Livingstone, 1988; Narbonneet al., 1991; Porte et al., 1991; Michel et al., 1994; Soleet al., 1996; Corsi et al., 2003).In our study, the high B(a)PMO activity levels
reported in digestive gland microsomes of mussels fromstations G1 (Mar Grande) and P5 and P6 (first inlet,Mar Piccolo) are in agreement with high DNA adductlevels found in these sites. Significantly higher levelswere detected in the first inlet than in the second and thiscould be explained by the presence of a limited source ofPAHs and high levels of potent inhibitors of P450, suchas TBTO and tin in sediments and in mussels, similar tothose reported by Morcillo and Porte (1997) and byStorelli and Marcotrigiano (2001).We reported a good correlation between B(a)PMO
activity and DNA adduct levels in the digestive gland inthe 2001 and 2002 campaigns (r ¼ 0:60 and 0.73,respectively). This result suggests an induction ofenzyme activity by exposure to B(a)P or B(a)P-likechemicals that leads to an increase in the phase Ibiotransformation process and then to higher produc-tion of phase I metabolites directly involved in bulkyDNA adduct formation (Akcha et al., 2000). Because ofits good correlation to adduct levels, B(a)PMO activityappears in this work as a relevant biomarker of B(a)P
exposure in pollution monitoring using this musselspecies as a sentinel organism. Our observations are inagreement with those found in the works of Varanasiet al. (1987) and Myers et al. (1991).We also observed no significant differences in
B(a)PMO activity in mussels sampled from six stationsbetween 2001 and 2002, in accordance with the adductlevel results.
4.4. Lysosomal membrane stability
Lysosomal membrane stability is considered the mostreliable of the recommended biomarkers in waterquality assessment (UNEP, 1997). In field studies,evaluation of the lysosomal membrane labilizationperiod using the lysosomal membrane stability test isconsidered to be a very sensitive biomarker of generalstress (Moore et al., 1982, 1986, 1987; Viarengo et al.,1987; Moore, 1988; Regoli, 1992; Krishnakumar et al.,1994) due to the impact of pollutants. The use of thispowerful biomarker has been evaluated in transplantedmussels (Viarengo and Canesi, 1991; Regoli andOrlando, 1994; Stien et al., 1998), in laboratory studies(Viarengo et al., 1981, 1987; Harrison and Berger, 1982),and in in vitro studies (Winston et al., 1991).Our data showed significantly lower lysosomal
membrane labilization period values in the musselssampled from stations G2 (Mar Grande) and P4 (secondinlet, Mar Piccolo) in both campaigns compared to thereference site. These results are indicative of a higherdegree of general stress in these two sites, due probablyto pollution of various origins. Since only PAHschemical data have so far been available in thesediments, we can only speculate on the nature andorigin of these pollutants. In agreement with the presentstudy, eight urban sewage outlets were found to draindirectly into the second inlet of the Mar Piccolo, ofwhich the most significant is Canal D’Aiedda (Storelliand Marcotrigiano, 2000). Moreover, high levels of Cdand Zn have been recently reported in samples of M.
galloprovincialis from the Mar Piccolo (Storelli et al.,2000). Indeed, in the Mar Grande, in the vicinity ofTaranto harbor, intense activity is carried out: 48% ofthe movement of goods is made up of metallic mineralsdestined for the metallurgic industry, 27% oil and itsderivates, 24% solid combustibles, and 1% bitumen(Storelli and Marcotrigiano, 2000). The waste waters inthis area contain toxic pollutants such as ammonia,phenols, mineral oil, heavy metals, etc. (Storelli andMarcotrigiano, 2000). Previous field studies performedin the Mediterranean and other coastal regions haveconfirmed a corresponding correlation with the presenceof various contaminants (Moore, 1988; Viarengo et al.,1988; Regoli, 1992; Krishnakumar et al., 1994; Loweet al., 1995, Ringwood et al., 1998; Domouhtsidou andDimitriadis, 2001; Petrovic et al., 2001). In the present

ARTICLE IN PRESSM. Pisoni et al. / Environmental Research 96 (2004) 163–175 173
study, lysosomal stability showed values similar to thosefound by Petrovic et al. (2001) in samples obtained fromRijeka Harbor (on the eastern coast of the NorthernAdriatic), one of the most heavily polluted areas due topresence of municipal, industrial, and commercialharbor waste waters.No statistical differences between samples collected
during the October 2001 and 2002 campaigns werefound at each station, indicating no major differences inpollutant content.
5. Conclusions
The results of this study suggest the presence ofdifferent sources and amounts of environmental pollu-tion in the coastal waters around Taranto. The exposureof M. galloprovincialis to these contaminants leads toseveral changes at the cellular and molecular levels. Theformation of PAH–DNA adducts confirms the bio-transformational capacity of PAHs by M. galloprovin-
cialis. PAH transformation leads to changes inB(a)PMO enzymatic biomarker measurements, to ahigher production of phase I metabolites, and, conse-quently, to DNA adduct formation. The correlationbetween the two biomarkers confirms the validity ofDNA adduct and B(a)PMO activity as suitable biomar-kers of PAH exposure. Moreover, higher DNA adductlevels were found in the gill tissue than in the digestivegland, although the MFO system is predominant in thelatter tissue and the digestive gland is the major site forhydrocarbon uptake, underlining the importance of alsoanalyzing gills in mussels. This work shows tissue-specific differences in DNA adduct formation inmussels, M. galloprovincialis, collected from some areasof coastal waters with different sources of pollutants,underlining that digestive gland and gills have differentsusceptibilities.Our results are in agreement with other field studies,
confirming the utility and the sensitivity of lysosomalmembrane stability as a biomarker of general stressfor detecting and monitoring sublethal changes inmarine organisms. Its greatest value emanates fromits prognostic value for tissue atrophy and sub-sequent pathology (Moore et al., 1978; Moore andAllen, 2002).In conclusion, our findings provide further indications
that there is a pressing need to analyze DNA adducts ingills together with digestive gland to facilitate theinterpretation of future field and experimental studieson mussels exposed to genotoxic compounds. Finally,the integrated use of biomarkers of exposure and of theeffects of environmental contaminants on living marineorganisms may help better interpret the impact ofpollutants on a marine coastal environment.
Acknowledgments
We gratefully thank Professor A. Tursi and ProfessorA. Matarrese for their scientific support and appreciatethe assistance of R. Chiea and G. Perra throughout thesampling campaign. Our special thanks go to ProfessorA. Viarengo and his lab personnel for introducing us tothe assay of lysosomal membrane stability. We thankProfessor L. Lopez for chemical analyses. We also thankDr. Anthony Green for reviewing the English. Thiswork was undertaken in the framework of the SPICA-MAR project, which was financially supported by theCoNISMa (Consorzio Interuniversitario per le Scienzedel Mare).
References
Accomando, R., Viarengo, A., Moore, M.N., Marchi, B., 1999. Effects
of ageing on nuclear DNA integrity and metabolism in mussel cells
(Mytilus edulis L.). Int. J. Biochem. Cell. Biol. 31, 443–450.
Akcha, C., Izuel, C., Venier, P., Hubinzki, H., Burgeot, T., Narbonne,
J-F., 2000. Enzymatic biomarker measurement and study of DNA
adduct formation in benzo[a]pyrene-contaminated mussels, Myti-
lus galloprovincialis. Aquat. Toxicol. 49, 269–287.
Akcha, F., Hubert, F.V., Pfohl-Leszkowicz, A., 2003. Potential value
of the comet assay and DNA adduct measurement in dab (Limanda
limanda) for assessment of in situ exposure to genotoxic
compounds. Mutat. Res. 534, 21–32.
Amodio-Cocchieri, R., del Prete, U., Arnese, A., Giuliano, M.,
Concioni, A., 1993. Heavy metals and PAHs in marine organisms
from the Ionian Sea (Italy). Bull. Environ. Contam. Toxicol. 50,
618–625.
Bayne, B.L., 1989. Measuring the biological effect of pollution: the
mussel watch approach. Water Sci. Technol. 21, 1089–1100.
Beach, A.C., Gupta, R.C., 1992. Human biomonitoring and the 32P-
postlabeling assay. Carcinogenesis 13, 1053–1074.
Brescia, G., Celotti, L., Clonfero, E., Neumann, H.G., Forni, A., Foa,
V., Pisoni, M., Ferri, G.M., Assennato, G., 1999. The influence of
cytochrome P450 1A1 and glutathione S-transferase M1 genotypes
on biomarker levels in coke-oven workers. Arch. Toxicol. 73,
431–439.
Cajaraville, M.P., Bebianno, M.J., Blasco, J., Porte, C., Sarasquete, C.,
Viarengo, A., 2000. The use of biomarkers to assess the impact of
pollution in coastal environments of the Iberian Peninsula. Sci.
Total Environ. 247, 295–311.
Canova, S., Degan, P., Peters, L.D., Livingstone, D.R., Voltan, R.,
Venier, P., 1998. Tissue dose, DNA adducts, oxidative DNA
damage and CYP1A-immunopositive proteins in mussels exposed
to waterborne benzo[a]pyrene. Mutat. Res. 399, 17–30.
Ching, E.W.K., Siu, W.H.L., Lam, P.K.S., Xu, L., Zhang, Y.,
Richardson, B.J., Wu, R.S.S., 2001. DNA adduct formation and
DNA strand breaks in green-lipped mussels (Perna viridis) exposed
to benzo[a]pyrene: dose- and time-dependent relationships. Mar.
Pollut. Bull. 42, 603–610.
Corsi, I., Mariottini, M., Sensini, C., Lancini, L., Focardi, S., 2003.
Cytochrome P450, acetylcholinesterase and general histology for
evaluating contaminant exposure levels in fishes from a highly
eutrophic brackish ecosystem: the Orbetello Lagoon, Italy. Mar.
Pollut. Bull. 46, 203–212.
Domouhtsidou, G.P., Dimitriadis, V.K., 2001. Lysosomal and
lipid alterations in the digestive gland of mussels,

ARTICLE IN PRESSM. Pisoni et al. / Environmental Research 96 (2004) 163–175174
Mytilus galloprovincialis (L.) as biomarkers of environmental
stress. Environ. Pollut. 15, 123–137.
Ericson, G., Skarphethinsdottir, H., Dalla Zuanna, L., Svavarsson, J.,
2002. DNA adducts as indicators of genotoxic exposure in
indigenous and transplanted mussels, Mytilus edulis L. from
Icelandic coastal sites. Mutat. Res. 516, 91–99.
Fitzpatrick, P.J., Sheehan, D., 1993. Separation of multiple forms of
glutathione S-transferase from the blue mussel, Mytilus edulis.
Xenobiotica 23, 851–861.
Fitzpatrick, P.J., Krag, T.O.B., Hojrup, P., Sheehan, D., 1995a.
Characterization of a glutathione S-transferase and a related
glutathione-binding protein from gill of the blue mussel, Mytilus
edulis. Biochem. J. 305, 145–150.
Fitzpatrick, P.J., Sheehan, D., Livingstone, D.R., 1995b. Studies on
isoenzymes of glutathione S-transferase in the digestive gland of
Mytilus galloprovincialis with exposure to pollution. Mar. Environ.
Res. 39, 241–244.
Gray, J.S., 1992. Biological and ecological effects of marine pollutants
and their detection. Mar. Pollut. Bull. 25, 48–50.
Harrison, F.L., Berger, R., 1982. Reactions of lysosomes to cell injury.
In: Trump, B.F., Arstila, A.V. (Eds.), Pathology of Cell
Membranes, Vol. 2. Academic Press, New York, San Francisco,
London, pp. 252–285.
Harvey, J.S, Parry, J.M., 1997. The detection of genotoxin-induced
DNA adducts in the common mussel Mytilus edulis. Mutagenesis
12, 153–158.
Krishnakumar, P.K., Casillas, E., Varanasi, U., 1994. Effect of
environmental contamination on health of Mytilus edulis from
Puget Sound, Washington: cytochemical measures of lysosomal
responses in the digestive cells using automatic image analysis.
Mar. Ecol. Prog. Ser. 106, 249–261.
Kurelec, B., 1993. The genotoxic disease syndrome. Mar. Environ.
Res. 35, 341–348.
Kurelec, B., Brivtic, S., Rijaved, M., Muller, W.E.G., Zhan, R.K.,
1977. Benzo(a)pyrene monooxygenase induction in marine fish.
Molecular response to oil pollution. Mar. Biol. 44, 211–216.
Lerario, V.L., Giandomenico, S., Lopez, L., Cardellicchio, N., 2003.
Sources and distribution of polycyclic aromatic hydrocarbons
(PAHs) in sediments from the Mar Piccolo of Taranto, Ionian Sea,
southern Italy. Ann. Chim. 93, 397–406.
Livingstone, D.R., 1988. Responses of microsomal NADPH-cyto-
chrome c reductase activity and cytochrome P-450 in digestive
glands of Mytilus edulis and Littorina littorea to environmental and
experimental exposure to pollutants. Mar. Ecol. Prog. Ser. 46,
37–43.
Livingstone, D.R., 1991. Organic xenobiotic metabolism in marine
invertebrates. In: Gilles, R. (Ed.), Advances in Comparative and
Environmental Physiology, Vol. 7. Springer, Berlin, pp. 45–185.
Livingstone, D.R., Moore, M.N., Lowe, D.M., Nasci, C., Farrar, S.V.,
1985. Responses of the cytochrome P-450 monooxygenase system
to diesel oil in the common mussel, Mytilis edulis L. and the
periwinkle. Littorina littorea L. Aquat. Toxicol. 7, 79–91.
Livingstone, D.R., Nasci, C., Sole, M., Da Ros, L., O’Hara, S.C.M.,
Peters, L.D., Fossato, V., Wooton, A.N., Goldfarb, P.S., 1997.
Apparent induction of a cytochrome P450 with immunochemical
similarities to CYP1A in digestive gland of the common mussel
(Mytilus galloprovincialis L.) with exposure to 2,20,3,4,40,50-hexa-
chlorobiphenyl and Arochlor 1254. Aquat. Toxicol. 38, 205–224.
Lowe, D.M., Fossato, V.U., Depledge, M.H., 1995. Contaminant-
induced lysosomal membrane damage in blood cells of mussel
Mytilus galloprovincialis from the Venice Lagoon: an in vitro study.
Mar. Ecol. Prog. Ser. 129, 189–196.
Michel, X.R., Suteau, P., Robertson, L.W., Narbonne, J.-F., 1993.
Effects of benzo(a)pyrene, 3,30,4,40-tetrachlorobiphenyl on the
xenobiotic-metabolizing enzymes in the mussel (Mytilus gallopro-
vincialis). Aquat. Toxicol. 27, 335–344.
Michel, X., Salaun, J.-P., Galgani, F., Narbonne, J.-F., 1994.
Benzo(a)pyrene hydroxylase activity in the marine mussel Mytilus
galloprovincialis: a potential marker of contamination by polycyclic
aromatic hydrocarbon-type compounds. Mar. Environ. Res. 38,
257–273.
Moore, M.N., 1976. Cytochemical demonstration of latency of
lysosomal hydrolases in digestive cells of the common mussel
Mytilus edulis, and changes induced by thermal stress. Cell Tissue
Res. 175, 279–287.
Moore, M.N., 1980. Cytochemical determination of cellular responses
to environmental stressors in marine organisms. Rapp-v Reun.
Cons. Int. Explor. Mer. 179, 7–15.
Moore, M.N., 1985. Cellular responses to pollutants. Mar. Pollut.
Bull. 16, 134–139.
Moore, M.N., 1988. Cytochemical responses of the lysosomal system
and NADPH-ferrihemoprotein reductase in molluscan digestive
cells to environmental and experimental exposure to xenobiotics.
Mar. Ecol. Prog. Ser. 46, 81–89.
Moore, M.N., Allen, J.I., 2002. A computational model of the
digestive gland epithelial cell of marine mussels and its simulated
responses to oil-derived aromatic hydrocarbons. Mar. Environ.
Res. 54, 579–584.
Moore, M.N., Lowe, D.M., Fieth, P.E.M., 1978. Lysosomal responses
to experimentally injected anthracene in the digestive cells of
Mytilus edulis. Mar. Biol. 48, 297–302.
Moore, M.N., Pipe, R.K., Farrar, S.V., 1982. Lysosomal and
microsomal responses to environmental factors in Littorina littorea
from Sullom Voe. Mar. Pollut. Bull. 13, 340–345.
Moore, M.N., Lowe, D.M., Livingstone, D.R., Dixon, D.R., 1986.
Molecular and cellular indices of pollutants effects and their
use in environmental impact assessment. Water Sci. Technol. 18,
223–232.
Moore, M.N., Pipe, R.K., Farra, S.V., Thomas, S., Donkin, P., 1987.
Lysosomal and microsomal responses to oil-derived hydrocarbons
in Littorina littorea. In: Capuzzo, J.R., Kester, D.R. (Eds.),
Oceanic Processes in Marine Pollution. ER Krieger, Malabar,
F.L, pp. 89–96.
Morcillo, Y., Porte, C., 1997. Interaction of tributyl- and triphenyltin
with the microsomal monooxygenase system of molluscs and fish
from the western Mediterranean. Aquat. Toxicol. 38, 35–46.
Myers, M.S., Landahl, J.T., Krahn, M.M., McCain, B.B., 1991.
Relationships between hepatic neoplasms and related lesions and
exposure to toxic chemicals in marine fish from the US West Coast.
Environ. Health Perspect. 90, 7–15.
Narbonne, J.F., Garrigues, P., Ribera, D., Raoux, C., Mathieu, A.,
Lemaire, P., 1991. Mixed-function oxygenase enzymes as tools for
pollution monitoring: field studies on the French coast of the
Mediterranean Sea. Comp. Biochem. Physiol. 100C, 37–42.
Nasci, C., Campesan, G., Fossato, V.U., Dolci, F., Mesetto, A., 1989.
Hydrocarbon content and microsomal BPH and reductase activity
in mussel, Mytilus sp. from the Venice area, North-east Italy. Mar.
Environ. Res. 28, 109–112.
Nebert, D.W., Gelboin, H.V., 1968. Substrate-inducible microsomal
aryl hydroxylase in mammalian cell culture. J. Biol. Chem. 243,
6242–6249.
Peters, L.D., Nasci, C., Livingstone, D.R., 1998. Immunochemical
investigations of cytochrome P450 forms/epitopes (CYP1A, 2B,
2E, 3A and 4A) in digestive gland of Mytilus sp. Comp. Biochem.
Physiol. C. Pharmacol. Toxicol. Endocrinol. 121, 361–369.
Petrovic, S., Ozretic, B., Krajnovic’-Ozretic’, M., Bobinac, D., 2001.
Lysosomal membrane stability and metallothioneins in digestive
gland of mussels (Mytilus galloprovincialis Lam) as biomarkers in a
field study. Mar. Pollut. Bull. 42, 1373–1378.
Pfau, W., 1997. DNA adducts in marine and freshwater fish as
biomarkers of environmental contamination. Biomarkers 2,
145–151.

ARTICLE IN PRESSM. Pisoni et al. / Environmental Research 96 (2004) 163–175 175
Porte, C., Sole, M., Albaiges, J., Livingstone, D.R., 1991. Response of
mixed-function oxygenase and antioxidase enzyme system of
Mytilus sp. to organic pollution. Comp. Biochem. Physiol. 100C,
138–186.
Randerath, K., Reddy, M.V., Gupta, R.C., 1981. 32P-postlabeling test
for DNA damage. Proc. Natl. Acad. Sci. USA 78, 6126–6129.
Reddy, M.V., Randerath, K., 1986. Nuclease P1-mediated increase of
sensitivity of 32P-postlabeling test for structural diverse DNA
adducts. Carcinogen 7, 1543–1551.
Regoli, F., 1992. Lysosomal responses as a sensitive stress index in
biomonitoring heavy metal pollution. Mar. Ecol. Prog. Ser. 84,
63–69.
Regoli, F., Orlando, E., 1994. Accumulation and subcellular distribu-
tion of metals (Cu, Fe, Mn, Pb and Zn) in the Mediterranean
mussel Mytilus galloprovincialis during a field transplant experi-
ment. Mar. Pollut. Bull. 28, 592–600.
Ringwood, A.H., Coners, D.E., Hoguet, J., 1998. Effects of natural
and anthropogenic stressors on lysosomal destabilization in oysters
Crassostrea virginica. Mar. Ecol. Prog. Ser. 166, 163–171.
Skarphethinsdottir, H., Ericson, G., Dalla Zuanna, L., Gilek, M.,
2003. Tissue differences, dose-response relationship and persistence
of DNA adducts in blue mussels (Mytilus edulis L.) exposed to
benzo[a]pyrene. Aquat. Toxicol. 62, 165–177.
Sole, M., Porte, C., Biosca, X., 1996. Effects of the ‘‘Aegean Sea’’ oil
spill on biotransformation enzymes, oxidative stress and DNA
adducts in digestive gland of the mussel (Mytilus edulis L.) Comp.
Biochem. Physiol. C113, 257–265.
Stein, J.E., Reichert, W.L., French, B., Varanasi, B., Varanasi, B.U.,
1993. 32P-postlabeling analysis of DNA adduct formation and
persistence in English sole (Pleuronectes vetulus) exposed to
b(a)pyrene and 7H-dibenzo[c,g]carbazole. Chem. Biol. Interact.
88, 55–69.
Stien, X., Percic, P., Gnassia-Barelli, M., Romeo, M., Lafaurie, M.,
1998. Evaluation of biomarkers in caged fishes and mussels to
assess the quality of waters in a bay of the NWMediterranean Sea.
Environ. Pollut. 99, 339–345.
Storelli, M.M., Marcotrigiano, G.O., 2000. Polycyclic aromatic
hydrocarbon distributions in sediments from the Mar Piccolo,
Ionian Sea, Italy. Bull. Environ. Contam. Toxicol. 65, 537–544.
Storelli, M.M., Marcotrigiano, G.O., 2001. Polycyclic aromatic
hydrocarbons in mussels (Mytilus galloprovincialis) from the Ionian
Sea, Italy. J. Food Prot. 64, 405–409.
Storelli, M.M., Storelli, A., Marcotrigiano, G.O., 2000. Heavy metals
in mussels (Mytilus galloprovincialis) from the Ionian Sea, Italy. J.
Food Prot. 63, 273–276.
UNEP, 1997. Report of the meeting of experts to review the MEDPOL
biomonitoring programme. Athens, Greece: UNEP-(OCA)/MED
WG. 132/7.
Van der Oost, R., Van Schooten, F.J., Ariese, F., Heida, H.,
Satumalay, K., Vermaulen, N.P.E., 1994. Bioaccumulation,
biotransformation and DNA binding of PAHs in feral eel (Anguilla
anguilla) exposed to polluted sediments: a field survey. Environ.
Toxicol. Chem. 13, 421–425.
Varanasi, U., Stein, J.E., Nishimoto, M., Reichert, W.L., Collier,
T.K., 1987. Chemical carcinogenesis in feral fish: uptake, activa-
tion, and detoxification of organic xenobiotics. Environ. Health
Perspect. 71, 155–170.
Venier, P., 2001. DNA adduct detection in mussels exposed to bulky
aromatic compounds in laboratory and field conditions. Biomar-
kers in Marine Organisms: A Practical Approach. pp. 65–83
(Chapter 4).
Venier, P., Canova, S., 1996. Formation of DNA adducts in gill tissue
of Mytilus galloprovincialis treated with benzo[a]pyrene. Aquat.
Toxicol. 34, 119.133.
Viarengo, A., Canesi, L., 1991. Mussels as biological indicators of
pollution. Aquaculture 94, 225–243.
Viarengo, A., Pertica, M., Mancinelli, G., Palmero, S., Zanicchi, G.,
Orunesu, M., 1981. Synthesis of Cu-binding proteins in different
tissues of mussels exposed to the metal. Mar. Pollut. Bull. 12, 347–
350.
Viarengo, A., Moore, M.N., Mancinelli, G., Mazzucotelli, A., Pipe,
R.K., Farrar, S.V., 1987. Metallothioneins and lysosomes in metal
toxicity and accumulation in marine mussels: the effect of cadmium
in the presence and absence of phenanthrene. Mar. Biol. 94,
251–257.
Viarengo, A., Mancinelli, G., Martino, G., Pertica, M., Canesi, L.,
Mazzucotelli, A., 1988. Integrated cellular stress indices in trace
metal contamination: critical evaluation in a field study. Mar. Ecol.
Prog. Ser. 46, 65–70.
Viarengo, A., Canesi, L., Pertica, M., 1995. Stress on stress response: a
simple monitoring tool in the assessment of a general stress
syndrome in mussels. Mar. Environ. Res. 39, 245–248.
Walters, J.M., Cain, R.B., Higgins I, .J., Corner, E.D., 1979. Cell-free
benzo(a)hydroxylase activity in marine zooplankton. J. Mar. Biol.
Assoc. UK 59, 553–563.
Widdows, J., Donkin, P., 1992. Mussels and environmental con-
taminants: bioaccumulation and physiological aspects. In: Gosling,
E. (Ed.), The Mussel Mytilus: Ecology, Physiology, Genetics
and Culture, of developments in Aquaculture and Fisheries
Science, Vol. 25. Elsevier Science Publishers, Amsterdam,
pp. 383–424.
Winston, G.W., Moore, M.N., Stratsburg, I., Kirchin, M., 1991.
Decreased stability of digestive gland lysosomes from the common
mussel Mytilus edulis L. by in vitro generation of oxygen-free
radicals. Arch. Environ. Contam. Toxicol. 21, 401–408.
Wootton, A.N., Herring, C., Spry, J.A., Wiseman, A., Livingstone,
D.R., Goldfarb, P.S., 1995. Evidence for the existence of
cytochrome P450 gene families (CYP1A, 3A, 4A, 11A) and
modulation of gene expression (CYP1A) in the mussel sp. Mar.
Environ. Res. 39, 21–27.





























