Embed Size (px)
Citation preview

nature biotechnology VOLUME 33 NUMBER 8 AUGUST 2015 853
A rt i c l e s
Protocols to generate tissue-specific cell types from human embryonic stem cells (hESCs) and induced pluripotent stem cells (iPSCs) have facilitated the development of novel in vitro models of human devel-opment and disease that have opened new avenues for drug discovery and cell therapy. Of the different lineages that can be studied with this approach, those of the liver are of special interest as hepatocytes and cholangiocytes, which make up the liver’s biliary system, are primary targets of the adverse effects of drugs and inherited and infectious diseases1. Biliary disorders such as cystic fibrosis (CF) and Alagille syndrome are common causes of liver disease that often necessitate liver transplantation2. More complex biliary diseases, such as primary sclerosing cholangitis and biliary atresia, lack appropriate models for understanding their pathophysiology and for screening pharmaco-logical agents.
Most studies to date have focused on generating hepatocytes from hPSCs3–6 and have yielded protocols that give rise to relatively mature cells suitable for predictive drug toxicology and modeling certain liver diseases in vitro5,6. Recently, cells with cholangiocyte characteristics were generated from hPSCs7, demonstrating that it is possible to derive this lineage in vitro as well. However, because these populations were established as monolayers, their functional capacity remains unclear, as cells cultured in this format often do not accurately recapitulate the normal physiological responses of complex tissues. The generation of functional three-dimensional (3D) biliary structures from hPSCs by following developmentally relevant signal-ing cues would overcome this limitation and provide an in vitro model
for studying biliary diseases and a foundation for developing tissue replacement strategies.
Cholangiocytes derive from a bipotential progenitor known as the hepatoblast that also gives rise to hepatocytes8. Targeting studies in mice have shown that cholangiocyte specification from the hepatoblast is mediated by the interaction of Notch2, expressed by the progeni-tors, and Jagged1, present on the developing portal mesenchyme9,10. Notch signaling also plays a critical role in intrahepatic bile duct morphogenesis in mice11,12. The discovery that the pediatric biliary disorder Alagille syndrome, characterized by bile duct paucity, is caused by mutations in JAG1 or NOTCH2 provides strong evidence that NOTCH signaling is also involved in cholangiocyte develop-ment in humans13–15. However, the role of the NOTCH pathway in regulating human cholangiocyte development from hPSCs has not been investigated.
Here we developed a serum-free protocol that directs the efficient differentiation and maturation of functional cholangiocytes from hPSCs by delivering developmentally relevant cues, including NOTCH signaling through co-culture with OP9 stromal cells, at the hepatoblast stage of development. We show that hPSC-derived cholangiocytes grown in 3D culture form epithelialized cystic and/or ductal structures that express markers found in mature bile ducts, including the CFTR. Cholangiocytes from normal hPSCs generate cystic and/or ductal structures and display epithelial functions, including CFTR-mediated fluid secretion, observed through the regulation of cyst swelling, after stimulation of the cAMP pathway with forskolin. Cysts generated
Directed differentiation of cholangiocytes from human pluripotent stem cellsMina Ogawa1,2,11, Shinichiro Ogawa1,11, Christine E Bear3, Saumel Ahmadi3, Stephanie Chin3, Bin Li4, Markus Grompe4, Gordon Keller1,5,6,12, Binita M Kamath7,8,12 & Anand Ghanekar2,9,10,12
Although bile duct disorders are well-recognized causes of liver disease, the molecular and cellular events leading to biliary dysfunction are poorly understood. To enable modeling and drug discovery for biliary disease, we describe a protocol that achieves efficient differentiation of biliary epithelial cells (cholangiocytes) from human pluripotent stem cells (hPSCs) through delivery of developmentally relevant cues, including NOTCH signaling. Using three-dimensional culture, the protocol yields cystic and/or ductal structures that express mature biliary markers, including apical sodium-dependent bile acid transporter, secretin receptor, cilia and cystic fibrosis transmembrane conductance regulator (CFTR). We demonstrate that hPSC-derived cholangiocytes possess epithelial functions, including rhodamine efflux and CFTR-mediated fluid secretion. Furthermore, we show that functionally impaired hPSC-derived cholangiocytes from cystic fibrosis patients are rescued by CFTR correctors. These findings demonstrate that mature cholangiocytes can be differentiated from hPSCs and used for studies of biliary development and disease.
1McEwen Centre for Regenerative Medicine, University Health Network, Toronto, Ontario, Canada. 2Toronto General Research Institute, University Health Network, Toronto, Ontario, Canada. 3Program in Molecular Structure & Function, The Hospital for Sick Children Research Institute, Toronto, Ontario, Canada. 4Department of Pediatrics, Oregon Health and Science University, Portland, Oregon, USA. 5Princess Margaret Cancer Centre, University Health Network, Toronto, Ontario, Canada. 6Department of Medical Biophysics, University of Toronto, Toronto, Ontario, Canada. 7Division of Gastroenterology, Hepatology and Nutrition, The Hospital for Sick Children, Toronto, Ontario, Canada. 8Department of Pediatrics, University of Toronto, Toronto, Ontario, Canada. 9Division of General Surgery, University Health Network, Toronto, Ontario, Canada. 10Department of Surgery, University of Toronto, Toronto, Ontario, Canada. 11These authors contributed equally to this work. 12These authors jointly directed this work. Correspondence should be addressed to A.G. ([email protected]) or S.O. ([email protected]).
Received 2 February; accepted 19 June; published online 13 July 2015; doi:10.1038/nbt.3294
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.
854 VOLUME 33 NUMBER 8 AUGUST 2015 nature biotechnology
A rt i c l e s
from CF patient iPSCs show impairment in forskolin-induced swelling that is rescued by addition of clinically relevant CFTR modu-lators. These findings demonstrate that delivery of NOTCH signaling to hPSC-derived hepatoblasts drives the generation of structurally and functionally mature cholangiocytes that can be used to model clinically relevant biliary diseases in vitro.
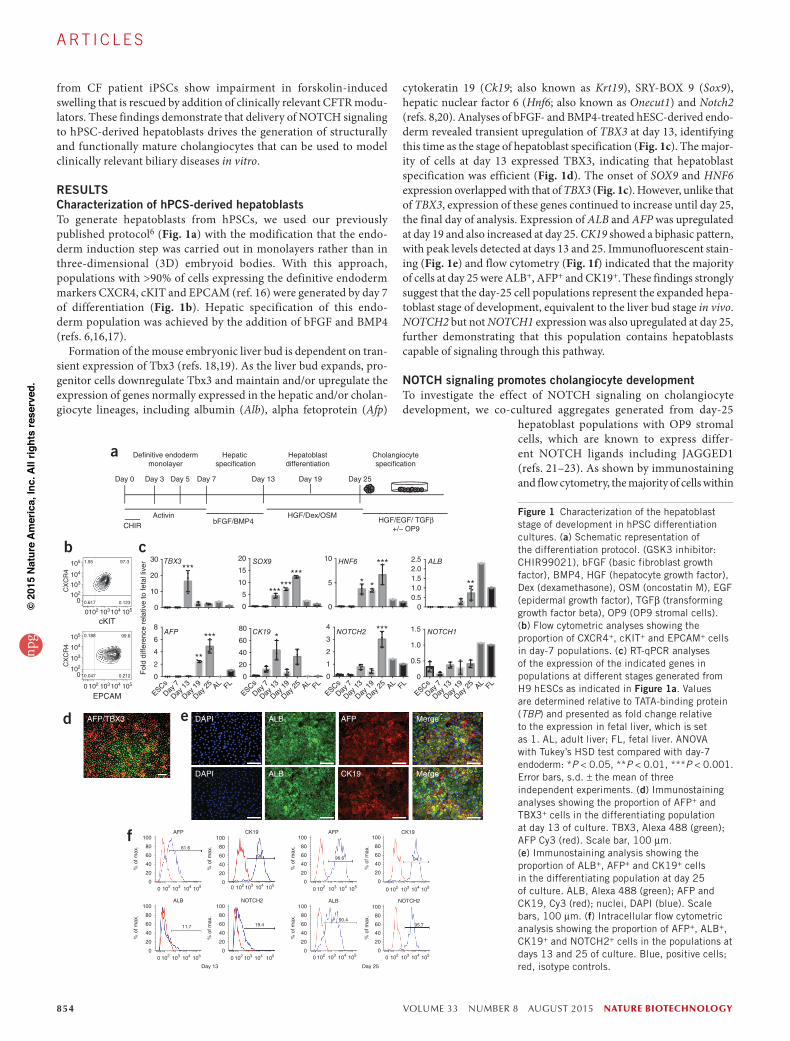
RESULTSCharacterization of hPCS-derived hepatoblastsTo generate hepatoblasts from hPSCs, we used our previously published protocol6 (Fig. 1a) with the modification that the endo-derm induction step was carried out in monolayers rather than in three-dimensional (3D) embryoid bodies. With this approach, populations with >90% of cells expressing the definitive endoderm markers CXCR4, cKIT and EPCAM (ref. 16) were generated by day 7 of differentiation (Fig. 1b). Hepatic specification of this endo-derm population was achieved by the addition of bFGF and BMP4 (refs. 6,16,17).
Formation of the mouse embryonic liver bud is dependent on tran-sient expression of Tbx3 (refs. 18,19). As the liver bud expands, pro-genitor cells downregulate Tbx3 and maintain and/or upregulate the expression of genes normally expressed in the hepatic and/or cholan-giocyte lineages, including albumin (Alb), alpha fetoprotein (Afp)
cytokeratin 19 (Ck19; also known as Krt19), SRY-BOX 9 (Sox9), hepatic nuclear factor 6 (Hnf6; also known as Onecut1) and Notch2 (refs. 8,20). Analyses of bFGF- and BMP4-treated hESC-derived endo-derm revealed transient upregulation of TBX3 at day 13, identifying this time as the stage of hepatoblast specification (Fig. 1c). The major-ity of cells at day 13 expressed TBX3, indicating that hepatoblast specification was efficient (Fig. 1d). The onset of SOX9 and HNF6 expression overlapped with that of TBX3 (Fig. 1c). However, unlike that of TBX3, expression of these genes continued to increase until day 25, the final day of analysis. Expression of ALB and AFP was upregulated at day 19 and also increased at day 25. CK19 showed a biphasic pattern, with peak levels detected at days 13 and 25. Immunofluorescent stain-ing (Fig. 1e) and flow cytometry (Fig. 1f) indicated that the majority of cells at day 25 were ALB+, AFP+ and CK19+. These findings strongly suggest that the day-25 cell populations represent the expanded hepa-toblast stage of development, equivalent to the liver bud stage in vivo. NOTCH2 but not NOTCH1 expression was also upregulated at day 25, further demonstrating that this population contains hepatoblasts capable of signaling through this pathway.
NOTCH signaling promotes cholangiocyte developmentTo investigate the effect of NOTCH signaling on cholangiocyte development, we co-cultured aggregates generated from day-25
hepatoblast populations with OP9 stromal cells, which are known to express differ-ent NOTCH ligands including JAGGED1 (refs. 21–23). As shown by immunostaining and flow cytometry, the majority of cells within
1.95
0.212
99.6
97.3
0.123
105104103102
105
104
103
102
0
105
104
103
102
0
0
1051041031020
1051041031020
1051041031020 1051041031020
1051041031020 1051041031020
1051041031020 1051041031020
1051041031020
0
5
10
15
20
0
10
20
30
Day 0 Day 3 Day 5 Day 7 Day 13 Day 19 Day 25
De�nitive endodermmonolayer
Hepatic speci�cation
Hepatoblastdifferentiation
Cholangiocytespeci�cation
ActivinCHIR
bFGF/BMP4HGF/Dex/OSM
HGF/EGF/ TGFβ+/– OP9
cKIT
EPCAM
AFP/TBX3 DAPI
DAPI
ALB
ALB
AFP
CK19
Merge
Merge
CX
CR
4C
XC
R4
Fol
d di
ffere
nce
rela
tive
to fe
tal l
iver
00.51.01.52.02.5
0
5
10TBX3 SOX9 HNF6 ALB***
******
**** *
***
**
0
0.5
1.0
1.5
0
1
2
3
4
0
20
40
60
80
0
2
4
6
8AFP CK19 NOTCH2 NOTCH1
**
*** ****
AFP CK19
ALB NOTCH2
Day 13
AFP CK19
NOTCH2ALB
Day 25
ESCs
Day 7
Day 1
3
Day 1
9
Day 2
5 AL FLESCs
Day 7
Day 1
3
Day 1
9
Day 2
5 AL FLESCs
Day 7
Day 1
3
Day 1
9
Day 2
5 AL FLESCs
Day 7
Day 1
3
Day 1
9
Day 2
5 AL FL
61.6
96
19.411.7
96.6 94.1
95.790.4
100
80
60
40
20
0
100
80
60
40
20
0
100
80
60
40
20
0
100
80
60
40
20
0
% o
f max
.
% o
f max
.%
of m
ax.
100
80
60
40
20
0
% o
f max
.
100
80
60
40
20
0
% o
f max
.
100
80
60
40
20
0
% o
f max
.
100
80
60
40
20
0
% o
f max
.
% o
f max
.
a
b c
d e
f
0.617
0.047
0.188
Figure 1 Characterization of the hepatoblast stage of development in hPSC differentiation cultures. (a) Schematic representation of the differentiation protocol. (GSK3 inhibitor: CHIR99021), bFGF (basic fibroblast growth factor), BMP4, HGF (hepatocyte growth factor), Dex (dexamethasone), OSM (oncostatin M), EGF (epidermal growth factor), TGFβ (transforming growth factor beta), OP9 (OP9 stromal cells). (b) Flow cytometric analyses showing the proportion of CXCR4+, cKIT+ and EPCAM+ cells in day-7 populations. (c) RT-qPCR analyses of the expression of the indicated genes in populations at different stages generated from H9 hESCs as indicated in Figure 1a. Values are determined relative to TATA-binding protein (TBP) and presented as fold change relative to the expression in fetal liver, which is set as 1. AL, adult liver; FL, fetal liver. ANOVA with Tukey’s HSD test compared with day-7 endoderm: *P < 0.05, **P < 0.01, ***P < 0.001. Error bars, s.d. ± the mean of three independent experiments. (d) Immunostaining analyses showing the proportion of AFP+ and TBX3+ cells in the differentiating population at day 13 of culture. TBX3, Alexa 488 (green); AFP Cy3 (red). Scale bar, 100 µm. (e) Immunostaining analysis showing the proportion of ALB+, AFP+ and CK19+ cells in the differentiating population at day 25 of culture. ALB, Alexa 488 (green); AFP and CK19, Cy3 (red); nuclei, DAPI (blue). Scale bars, 100 µm. (f) Intracellular flow cytometric analysis showing the proportion of AFP+, ALB+, CK19+ and NOTCH2+ cells in the populations at days 13 and 25 of culture. Blue, positive cells; red, isotype controls.
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature biotechnology VOLUME 33 NUMBER 8 AUGUST 2015 855
A rt i c l e s
DAPI ALB CK19 Merge
DAPI ALB AFP Merge
a
ALB CK19DAPI
CTRL EGF TGFβ1
EGF
HGF
TGFβ1 HGF EGFTGFβ1 GSI
HGFEGF
HGFEGF TGFβ1
b
SOX9 CK19
CFTR DAPI
ASBT DAPI
CK19α-tubulin
DAPI
c
d
0
1
2
3
4
5HES5 HEY1
Fol
d di
ffere
nce
rela
tive
to O
P9
(–)
HES1
OP9(–)
OP9(+)
OP9(+)GSI(+)
**
****
***
OP9(–)
OP9(+)
OP9(+)GSI(+)
OP9(–)
OP9(+)
OP9(+)GSI(+)
0
1
2
0
5
10
15
Fol
d ch
ange
rel
ativ
e to
hep
atob
last
Day 3
0 GSI(–
)
HB (day
27)
Day 3
3 GSI(–
)
Day 3
6 GSI(–
)
Day 3
0 GSI(+
)
Day 3
3 GSI(+
)
Day 3
6 GSI(+
)AL FL
ALB
CK19
e
f
Figure 2 NOTCH signaling promotes cholangiocyte differentiation from hPSC-derived hepatoblasts. (a) Confocal microscopic images of immunostained day-27 aggregates showing co-expression of ALB+, AFP+ and CK19+ cells. ALB, Alexa 488 (green); AFP and CK19, Cy3 (red); nuclei, DAPI (blue). Scale bars, 50 µm. (b) Immunostaining analysis showing the presence of ALB+ and CK19+ cells in hepatoblast populations co-cultured for 9 d (total culture time 36 d) on OP9 stromal cells in the indicated conditions. CTRL: control with no additional cytokines. ALB, Alexa 488 (green); CK19 Cy3 (red); nuclei, DAPI (blue). Scale bars, 100 µm. (c) Immunostaining analysis showing the presence of CK19+/SOX9+ cells (upper panel), CFTR+ cells (middle panel) and ASBT+ cells (lower panel) in hepatoblast populations co-cultured for 9 d (total culture time 36 d) on OP9 stromal cells in HGF, EGF and TGFβ1. SOX9, Alexa 488 (green); CK19, CFTR and ASBT, Cy3 (red); nuclei, DAPI (blue). Scale bars, 100 µm. (d) Confocal microscopy images for immunostaining of acetylated α-tubulin localized in primary cilia on H9-derived cholangiocytes cultured on OP9 stroma in HGF, EGF and TGFβ1. Acetylated α-tubulin, Cy3 (red); nuclei, DAPI (blue). Inset image with triple staining confirms colocalization of CK19 and primary cilia. CK19, Alexa 488 (green). Scale bar, 10 µm. (e) RT-qPCR-based analysis of the expression of NOTCH target genes in hepatoblast populations after 9 d of culture (total 36 d) on OP9 stroma (OP9+) with or without GSI or Matrigel-covered wells (OP9−) in the presence of HGF, EGF and TGFβ1. Values were determined relative to TBP and presented as fold-change relative to the levels in the cells cultured on Matrigel for 9 d (OP9−), which is set as 1. Bars in all graphs represent the s.d. ± the mean of three independent experiments. Student’s t-test compared with the culture on OP9 stroma (OP9+): **P < 0.01, ***P < 0.001 (n = 3). (f) RT-qPCR-based expression analysis of ALB and CK19 in hepatoblast populations cultured on OP9 stroma in the presence of HGF, EGF and TGFβ1 for 3 d (30 d total), 6 d (33 d total) and 9 d (36 d total). The addition of GSI is indicated. Hepatoblast (HB: day 27) represents the population before co-culture. Values were determined relative to TBP and presented as fold change relative to levels in the day 27 hepatoblast aggregates, which is set as 1.
the aggregates before co-culture were ALB+AFP+CK19+NOTCH2+, indicating that they maintained hepatoblast characteristics (Fig. 2a and Supplementary Fig. 1a). When co-cultured on OP9 cells for 9 d, the aggregates formed distinct clusters of CK19+ cells that downregu-lated ALB expression, suggesting that they had undergone the initial stage of cholangiocyte specification (Fig. 2b, CTRL). As studies in mice have shown that Hgf, Egf and Tgfβ1 signaling plays a role in bile duct development8,20,24, we next added combinations of these factors to the cultures to determine whether these pathways would promote further development of CK19+ clusters. The addition of EGF alone, TGFβ1 alone or the combination of EGF, TGFβ1 and HGF led to a 3.0- to 3.5-fold increase in cell number compared with nontreated controls after 9 d of culture (Fig. 2b and Supplementary Fig. 1b). HGF alone had little effect. The majority (90%) of the cells in the populations cultured under these conditions were CK19+ and ALB−, indicating efficient specification and development of the cholangio-cyte lineage (Supplementary Fig. 1c).
The combination of either EGF and HGF or EGF, HGF and TGFβ1 induced substantial morphological changes and promoted the forma-tion of CK19+ branched structures (Fig. 2b). CK19+ cells in structures treated with all three factors expressed other proteins indicative of the cholangiocyte lineage, including SOX9, the CFTR and the apical sodium-dependent bile acid transporter (ASBT) (Fig. 2c). They also displayed primary cilia as detected with acetylated α-tubulin (Fig. 2d). CFTR, ASBT and α-tubulin were not expressed in aggregates before OP9 cell co-culture (Supplementary Fig. 2). These findings demon-strate that co-culture of hepatoblasts with OP9 cells in the presence
of EGF, HGF and TGFβ1 promotes the development of a population that expresses cholangiocyte markers. With the efficiency of lineage specification in the co-culture model, we generated, on average, 4.16 ± 0.64 CK19+ALB− cholangiocyte-like cells per day-25 hepato-blast and 6.42 ± 0.53 cholangiocyte-like cells per input hESC.
Expression of NOTCH targets HES1, HES5 and HEY1 was upregu-lated after 9 d of culture on OP9 cells in the presence of EGF, HGF and TGFβ1 (Fig. 2e). This increase in expression was blocked by gamma secretase inhibitor (GSI), a NOTCH pathway antagonist, demonstrat-ing that co-culture with OP9 cells effectively activated the NOTCH pathway. GSI also blocked downregulation of ALB, reduced the proportion of CK19+ cells and inhibited the development of branched structures in the cultures, indicating that these effects were medi-ated by NOTCH signaling (Fig. 2b and Supplementary Fig. 1c). RT-qPCR analyses (Fig. 2f) confirmed the immunostaining and flow cytometric findings and demonstrated a dramatic reduction in ALB and an upregulation of CK19 after co-culture with OP9 cells in the presence of EGF, HGF and TGFβ1. These expression patterns were reversed with GSI. Collectively, these findings indicate that activa-tion of NOTCH signaling in the hepatoblast population induces the initial stages of cholangiocyte development and the combination of HGF, EGF and TGFβ1 signaling promotes the formation of branched structures reminiscent of early duct morphogenesis.
Differentiation of two other hPSC lines (ESC HES2 and MSC-iPSC1) revealed similar temporal patterns of TBX3 and hepatoblast marker expression (Supplementary Fig. 3a). Aggregates of hepatoblasts from both generated branched structures consisting of CK19+ALB− cells
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

856 VOLUME 33 NUMBER 8 AUGUST 2015 nature biotechnology
A rt i c l e s
following co-culture with OP9 stroma (Supplementary Fig. 3b), dem-onstrating the applicability of the approach to different hPSC lines.
3D culture promotes cholangiocyte maturationTo determine whether growth in 3D structures would promote further maturation of the hPSC-derived cholangiocytes, we next generated chimeric aggregates consisting of day-25 hepatoblasts and OP9 (GFP+) stromal cells (Supplementary Fig. 4a) and cultured them in gels composed of 1.2 mg/ml collagen type-1, 40% Matrigel and containing HGF, EGF and TGFβ1. Within 2 weeks of culture, the aggregates gave rise to 3D structures that displayed either a tubular and ductal morphology, a hollow cyst morphol-ogy or a mixture of both (Fig. 3a–c). Expression of NOTCH target genes HES1, HES5 and HEY1 was significantly higher in structures derived from chimeric aggregates than in those aggregates without OP9 cells, indicating that NOTCH signaling was active under these conditions (Supplementary Fig. 4b). Approximately 17 (17.1 ± 6.06) structures developed per 105 day-25 input hepatoblasts, the majority (62.9 ± 30.7%) displaying the cyst morphology. The generation of these cyst and duct structures was dependent on NOTCH signaling, as they did not develop in the presence of GSI. Rather, under these conditions, we observed the formation of dense colonies that we refer to as spheres.
Histological analyses revealed that tubular and cystic structures at day 44 had a lumen and contained epithelial-like cells that expressed
CK19 and E-CADHERIN but not ALB (Fig. 3d and Supplementary Fig. 4c). These cells also expressed the tight junction marker ZO-1 (Zonula occludens 1) CFTR, SCTR (secretin receptor) and ASBT and contained primary cilia (Fig. 3d,e). Most of these markers were restricted to the apical side of the structures, suggesting that the cells acquired apicobasal polarity, a feature of mature epithelial ducts. ASBT was also detected on the apical side of branching 3D struc-tures (Supplementary Fig. 4d). The presence of only small numbers of Ki-67+ cells in CK19+ duct structures at this stage suggests limited proliferative capacity (Supplementary Fig. 4e). Similar patterns of CK19, CFTR and α-tubulin staining were observed in bile ducts of normal adult liver (Supplementary Fig. 5).
Western blot analysis, RT-qPCR and flow cytometric analyses confirmed the presence of CFTR in cells generated from chimeric aggregates (Fig. 3f and Supplementary Fig. 6a,b). The levels of CFTR mRNA and protein were considerably higher in cells derived from chi-meric aggregates than in those cultured without OP9 cells, indicating that CFTR expression was NOTCH-dependent.
The spheres that developed in the presence of GSI expressed ALB and low levels of CK19, indicating that, in the absence of NOTCH signaling, cells with hepatoblast characteristics persist (Fig. 3d). RT-qPCR analyses showed similar changes as expression levels of hepatocyte markers (ALB, AFP and CYP3A7) increased whereas cholangiocyte-associated markers (CK19, SOX9 and CFTR) decreased (Supplementary Fig. 6c) after addition of GSI.
Mix Cyst
Tubular Sphere
a b
3D GSI(–)
3D GSI(+)
% o
f eac
h ty
pe o
f str
uctu
re
MixCystTubularSphere
0102030405060708090
100c
GSI(+)
GSI(–)
DAPI CK19 CFTR Merge
DAPI MergeCK19 ZO-1
DAPI SCTR CK19 Merge
DAPI ASBT CK19 Merge
ALB CK19DAPI Merge
d
CK19α-tubulin
DAPI
e f
250
130
95
α CFTR
α CNX
B and C
OP9(–)
OP9(+)
ES cells(kD)
Figure 3 3D culture promotes cholangiocyte maturation. (a) Photomicrographs of duct-like and cyst-like structures that develop in the collagen/Matrigel cultures after 17 d of culture of chimeric aggregates consisting of H9-derived hepatoblasts and OP9 stromal cells (4:1 ratio) in the absence of GSI. Spheres are dense colonies that develop in the presence of GSI. Scale bar, 100 µm. (b) Low-magnification photomicrograph showing cyst-like structures that develop from chimeric aggregates consisting of H9-derived hepatoblasts and OP9 stromal cells (4:1 ratio) after 17 d of culture in the collagen/Matrigel cultures. Scale bar, 100 µm. (c) Proportion of structures with tubular, cystic, mixed and sphere morphology that developed per well from chimeric aggregates cultured in the absence or presence of GSI. Values represent averages from five independent experiments. (d) Confocal microscopy images of immunostained histological sections of 3D ductal and/or cyst structures after 17 d (total 44 d) of culture in the presence of HGF, EGF and TGFβ1 without GSI (GSI(−)). First and second rows: CK19, Alexa 488 (green); ZO-1 and CFTR, Cy3 (red). Third and fourth rows: SCTR and ASBT, Alexa 488 (green); CK19, Cy3 (red). Bottom row: confocal microscopic images of immunostained histological sections of 3D sphere structures after 17 d (total 44 d) of culture in the presence of HGF, EGF and TGFβ1 with GSI (GSI(+)); ALB, Alexa 488 (green); CK19, Cy3 (red). In all images, the nuclei are stained by DAPI (blue). Scale bars, 50 µm. (e) Confocal microscopy images of immunostaining of acetylated α-tubulin localized in primary cilia present in a 3D ductal structure after 17 d (total 44 d) of co-culture with OP9 stromal cells in the presence of HGF, EGF and TGFβ1. Acetylated α-tubulin, Alexa 488 (green); CK19, Cy3 (red); nuclei, DAPI (blue). Scale bar, 10 µm. (f) Western blot analysis shows the presence of the mature glycosylated form of the CFTR protein (band C) in MSC-iPSC1-derived ductal and/or cyst structures generated after 17 d of culture of chimeric aggregates (OP9+) or aggregates of hepatoblasts without OP9 (OP9−). Undifferentiated iPSCs were used as negative control and calnexin (CNX) was measured as a loading control.
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature biotechnology VOLUME 33 NUMBER 8 AUGUST 2015 857
A rt i c l e s
To further validate the utility of the 3D protocol, we next applied it to the human iPSC line MSC-iPSC1. A distribution of CK19+/CFTR+/ASBT+ tubular and/or cystic structures similar to that observed in the H9 cultures developed from MSC-iPSC1-derived hepatoblast aggregates (Supplementary Fig. 7a). The efficiency of tubular and/or duct/cyst formation was also similar to that of the H9 hESCs, with 105 day-25 iPSC-derived hepatoblasts giving rise to 20.0 ± 1.64 structures. Primary cilia were also detected in these duct structures (Supplementary Fig. 7b).
RT-qPCR analyses revealed that expression of genes encoding proteins found in functional cholangiocytes in vivo, including SOX9, HNF1B, HNF6, CK19, CFTR, SCTR, aquaporin-1 (AQP1), the Cl−/HCO3− anion exchanger 2 (SLC4A2/AE2), ASBT, gamma-glutamyltransferase (GGT), TGR5 and the somatostatin receptor 2 (SSTR2) was significantly upregulated in hPSC-derived cystic structures compared to day-25 hepatoblasts (Fig. 4). Notably, the levels found in these structures were equivalent to or greater than those detected in primary cholangiocytes isolated by fluorescence- activated cell sorting (FACS) from human liver using the ductal anti-body DHIC5-4D9 (ref. 25; Supplementary Figs. 5 and 8). In contrast, HNF4A and ALB were expressed at higher levels in primary cultured hepatocytes and hPSC-derived hepatoblasts than in the cysts or primary cholangiocytes (Fig. 4 and Supplementary Table 3). Together, these findings show that when cultured in a mixture of Matrigel and collagen, the hepatoblast population can generate duct-like structures that express mature biliary markers.
hPSC-derived cholangiocytes form ductal structures in vivoTo evaluate the developmental potential of hPSC-derived cholan-giocytes in vivo, we transplanted 106 cells in a Matrigel plug into mammary fat pads of nonobese diabetic–severe combined immunodeficient–interleukin 2rγ−/− (NOD-SCID-IL2rγ−/−; NSG) mice. For these studies, we used dissociated cells from branched structures generated by co-culture of day-25 hepatoblasts derived
from HES2-RFP hESCs26 with OP9 stroma. Six to 8 weeks after transplantation, multiple duct-like structures were detected in the Matrigel plug (Supplementary Fig. 9a). Cells within the ducts were RFP+, demonstrating that they were of human origin (Supplementary Fig. 9b,c). Additionally they expressed CK19, CFTR and primary cilia, indicating that they displayed characteristics of cholangiocytes (Supplementary Fig. 9d,e). Teratomas were not observed in any of the animals receiving transplants.
hPSC-derived cholangiocytes are functionalTo assess function of the hPSC-derived cholangiocytes, we evaluated their ability to efflux rhodamine 123, a dye used to measure activity of the multidrug resistance protein 1 (MDR1) transporter in normal bile duct cells. The cystic structures derived from H9 hESCs and normal iPSCs transported dye to the luminal space, indicative of transporter activity (Fig. 5a). In the presence of 20 µM verapamil, an inhibitor of MDR1, rhodamine did not accumulate in the lumen, confirming that dye movement reflected active transport through MDR1.
To demonstrate CFTR functional activity, we performed a forskolin- induced swelling assay on cystic structures (Fig. 5b). With this assay, activation of the cAMP pathway by addition of forskolin/IBMX increases CFTR function, resulting in fluid transport and cyst swelling27 that can be visualized after staining with the cell- permeable fluorescent dye calcein green. Addition of forskolin/IBMX to the cultures induced a fold increase of 1.23 ± 0.09 and 1.40 ± 0.17 in the size of H9- and iPSC-derived cysts, respectively, when measured 1 h later. Addition of the CFTR inhibitor CFTRinh-172 blocked the forskolin/IBMX-induced swelling, indicating that the increase
0
0.5
1.0
1.5
2.0
0
0.5
1.0
1.5
0
0.5
1.0
1.5
0
5
10
15
20
0
1
2
3
0
5
10
15
20
25
0
0.5
1.0
1.5
0
1
2
3
4
0
2
4
6
0
5
10
15
20
0
0.5
1.0
1.5
0
0.5
1.0
1.5
2.0
2.5
Fol
d di
ffere
nce
rela
tive
to s
orte
d ch
olan
gioc
yte
SOX9 HNF1B HNF6
CK19
******
****
***** SCTR
*
** CFTR
****
AQP1
**
SLC4A2/AE2
*** ASBT *
**
GGT * TGR5
**
*SSTR2
**
****
0
1
2
3
0
50
100HNF4A
****
0
2,000
4,000
6,000
8,000
10,000 AFP***
******
**ALB
Sorte
d ch
olang
iocyte
Primar
y hep
atoc
yte
Cholan
giocy
te (i
PSC)
Cholan
giocy
te (H
9)
Hepat
oblas
t (iP
SC)
Hepat
oblas
t (H9)
iPSC
hESC
Sorte
d ch
olang
iocyte
Primar
y hep
atoc
yte
Cholan
giocy
te (i
PSC)
Cholan
giocy
te (H
9)
Hepat
oblas
t (iP
SC)
Hepat
oblas
t (H9)
iPSC
hESC
Sorte
d ch
olang
iocyte
Primar
y hep
atoc
yte
Cholan
giocy
te (i
PSC)
Cholan
giocy
te (H
9)
Hepat
oblas
t (iP
SC)
Hepat
oblas
t (H9)
iPSC
hESC
Figure 4 Gene expression profile of hPSCs-derived cholangiocytes compared to sorted cholangiocytes. RT-qPCR analyses of the expression of the indicated genes in the indicated populations. hESC: undifferentiated H9 cells; iPSC: undifferentiated MSC-iPSCs; hepatoblasts: day-25 populations; cholangiocytes: cystic structures generated from chimeric aggregates of either H9 or MSC-iPSCs-derived hepatoblasts; primary cultured hepatocytes; sorted cholangiocytes: cholangiocytes isolated from human liver samples by FACS, using the ductal-specific monoclonal antibody DHIC5-4D9 (ref. 25). Data are expressed as the fold change relative to sorted human cholangiocytes, which is set as 1. Student’s t-test: *P < 0.05, **P < 0.01, ***P < 0.001 (n = 3). Error bars in all graphs represent the s.d. ± the mean of three independent experiments. SCTR (secretin receptor), AQP1 (aquaporin 1), SLC4A2/AE2 (Anion exchange protein 2), ASBT, GGT (gamma-glutamyl transpeptidase), TGR5 (G protein- coupled bile acid receptor 1), SSTR2 (somatostatin receptor 2), HNF4A, AFP, (α-fetoprotein), ALB.
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

858 VOLUME 33 NUMBER 8 AUGUST 2015 nature biotechnology
A rt i c l e s
in cyst size was CFTR-dependent (Supplementary Fig. 10). When measured 24 h later, the swelling of H9- and iPSC-derived cysts increased 2.09 ± 0.21- and 2.65 ± 0.31-fold, respectively (Fig. 5c and Supplementary Movies 1 and 2). This swelling was also blocked by addition of CFTRinh-172. These findings demonstrate that cells in hPSC-derived cyst and/or duct-like structures display properties of functional cholangiocytes.
Analyses of cholangiocytes generated from CF patient iPSCsTo demonstrate the utility of this system to model disease in vitro, we analyzed cyst formation from iPSCs generated from two different
CF patients carrying the common F508del mutation (C1-iPSCs and GM00997-iPSCs). Both patient-derived iPSC lines generated defini-tive endoderm and hepatoblasts with kinetics similar to that of nor-mal iPSCs (Fig. 6a,b and data not shown). Although hepatoblast development was not altered, cyst formation from the CF-iPSCs was clearly impaired as only branched structures were observed in gels after 17 d of culture (Fig. 6c). Cyst formation could be induced by addition of forskolin for the first week of the 2-week culture period (Fig. 6d). However, even under these conditions many cysts that developed from patient iPSCs were not completely hollow and con-tained branched ductal structures (Fig. 6e). As with the wild-type
0
100
200
300
400
500
0
100
200
300
400
H9-
deriv
ed
chol
angi
ocyt
es
% s
wel
ling
rela
tive
to
noni
nduc
ed c
ysts
Ver
apam
ilD
MS
O
H9 MSC-iPSC1
*** * ***
(–) (+)
MS
C-iP
SC
1-de
rived
ch
olan
gioc
ytes
(+)(+) (+)(+) (+)(+) (+) (+)(–) (–) (–)
(+)
MSC-iPSC1H9
*** *** ***
FSK/IBMXCFTR inhibitor (+) (+) (+)(–) (–) (–)
(+)(+) (+)(+) (+) (+)
b ca
Figure 5 Function of hPSC-derived cholangiocytes in vitro. (a) Phase contrast images showing uptake of rhodamine 123 dye in 17-d-old cysts generated from cholangiocytes derived from either H9 hESCs or MSC-iPSC1 cells. To demonstrate functional transport, cysts were incubated with verapamil for 30 min before the addition of rhodamine 123. Images were taken 10 min after the addition of rhodamine (green). Scale bars, 100 µm. (b,c) Forskolin-induced swelling assay demonstrating functional CFTR activity in hPSC-derived cysts. (b) Representative epifluorescence microscopy images of hESC and MSC-iPSC1-derived cysts before (−) and 24 h after (+) stimulation with forskolin/IBMX (F/I). Cells were labeled with calcein green and images were taken 24 h after F/I stimulation. Scale bars, 500 µm. (c) Quantification of the degree of cyst swelling 24 h after F/I stimulation in the absence or presence of the CFTR inhibitor CFTR Inh-172 (30 µM). Cyst swelling was quantified using velocity imaging software. Values shown are percent swelling relative to the cyst size before stimulation. Analyses from three independent experiments are shown. Student’s t-test: *P < 0.05, ***P < 0.001 (n = 3).
0
20
40
60
80
100
0
20
40
60
80
100
–FSK
MSC-iPSC1
Inh (+) Inh (–)
7 d
14 d
Cys
t for
mat
ion
(%)
C1-iPSC
FSK (+)FSK (–)
+FSK
EPCAM
CX
CR
4C
XC
R4
Fol
d di
ffere
nce
rela
tive
to fe
tal l
iver
7 d
14 d 7
d14
d 7 d
14 d
0
0.5
1.0
1.5
2.0
2.5
05
10152025
0
20
40
60
0
2
4
6
012345
0
2
4
6
8
C1-iP
SCDay
7
Day 1
3
Day 1
9
Day 2
5 AL FL
C1-iP
SCDay
7
Day 1
3
Day 1
9
Day 2
5 AL FL
C1-iP
SCDay
7
Day 1
3
Day 1
9
Day 2
5 AL FL
ALB AFP
TBX3 SOX9
CK19
NOTCH2
* ***
****
* *
******
*** ***
******
1.04 98.8
00.2
0 102 103 104 105
0102
103
104
105
0102
103
104
105
cKIT
0 102 103 104 105
0 99.7
0.280
*
a b c d
MSC-iPSC1 C1-iPSCeFigure 6 Generation of cholangiocytes from CF patient iPSCs. (a) Flow cytometric analyses showing the proportion of CXCR4+, cKIT+ and EPCAM+ cells in day-7 monolayer populations generated from a CF-iPSC line (C1 CFTR del). (b) RT-qPCR-based analyses of the expression of the indicated genes in populations at different stages generated from C1-iPSC. Cells were cultured as indicated in Figure 1a. Values shown are relative to TBP and presented as fold change relative to expression in fetal liver, which is set as one. AL: adult liver, FL: fetal liver. ANOVA with Tukey’s HSD test compared with day-7 endoderm: *P < 0.05, ***P < 0.001 (n = 3). Bars in all graphs represent the s.d. ± the mean of three independent experiments. (c) Photomicrographs of branched and cyst-like structures that develop in the collagen/Matrigel cultures after 17 d of culture of chimeric aggregates generated with cholangiocytes from CFTR F508del iPSCs (CF-iPSCs: C1-iPSC). Images are shown for structures that developed in the absence or presence of forskolin (FSK, 10 µM). Forskolin was added for the first week of the 17-d culture period (right). Scale bars, 100 µm. (d) Quantification of the number of cyst structures that developed from CF-iPSCs (C1-iPSC)-derived cholangiocytes cultured in the absence and presence of forskolin and from wild-type iPSCs (MSC-iPSC1) cultured in the absence and presence of the CFTR inhibitor CFTR Inh-172 (30 µM). Cyst structures were counted at 7 d and 14 d of culture. Forskolin was added to the CF-iPSC-derived aggregates and the CFTR inhibitor was added to the iPSC-derived aggregates for the first week of the 17-d-culture period (left graph). Bars represent the s.d. of the mean of three independent experiments. (e) Histological analyses showing the structure of iPSC- and CF-iPSC-derived cysts after 17 d of culture in the collagen/Matrigel gels (H&E staining; scale bars, 100 µm). Forskolin was included for the first week of the 17-d culture period.
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature biotechnology VOLUME 33 NUMBER 8 AUGUST 2015 859
A rt i c l e s
cells, CF iPSC-derived cysts contained primary cilia (Supplementary Fig. 11). A higher frequency of hollow cysts was detected following longer periods of culture, suggesting that maturation of the mutant cells was delayed. Addition of CFTR inhibitors to cultures of normal iPSC-derived cholangiocytes also delayed cyst formation, indicating that generation of these structures was dependent, to some degree, on functional CFTR at the cell surface (Fig. 6d).
We assessed functional restoration of F508del CFTR in cholangio-cytes from CF-patient iPSCs by adding the chemical correctors VX-809 and Corr-4a to cultures 2 d before the CFTR functional assay27–30. Both molecules correct folding defects of mutant CFTR protein. The addition of these correctors did not improve cyst formation but did result in accumulation of detectable levels of CFTR on the apical side of the lumen. Unlike the homogeneous distribution of CFTR in wild-type cysts, the mutant protein localized to the plasma membrane in some areas and exhibited perinuclear distribution in other regions (Fig. 7a). This pattern may reflect incomplete rescue of the trafficking defect of the mutant protein. Western blot analyses also revealed the effect of correctors. Wild-type CFTR normally migrates as two bands, one ~130–150 kD (band B) and the other ~170–190 kD (band C). This typical CFTR protein migration pattern is shown for lysates from human bronchial epithelial (HBE) cells (Fig. 7b). Band C represents the mature, Golgi-modified CFTR and band B, the imma-ture, ER-localized CFTR protein. The F508del-CFTR protein migrated as a 130-150 kD protein in the iPSC-derived cholangiocytes in the absence of correctors consistent with retention in the endoplasmic reticulum (ER) and failure to traffic to the cell surface. Treatment with correctors strongly increased the proportion of mature protein in these cells, demonstrating that these molecules are functional in CF iPSC-derived cholangiocytes.
To determine whether the correctors rescued functional expres-sion of F508del-CFTR, we subjected treated and untreated cysts to the forskolin and IBMX-induced swelling assay (Fig. 7c,d). For these
studies, the small-molecule potentiator VX-770 was added along with forskolin and IBMX to the media. CF cysts showed little swelling in the absence of the correctors. However, with addition of both cor-rectors, cysts generated from patient C1 increased by ~2.18 ± 0.52-fold, whereas those from patient GM00997 increased by 1.64 ± 0.08-fold 24 h after induction with forskolin, IBMX and VX-770 (Fig. 7c,d and Supplementary Movies 3 and 4). Although swelling over 24 h may reflect activities in addition to that of CFTR, sensitivity of swell-ing to CFTRinh-172 and other modulators of CFTR (cAMP agonists and VX-770) supports a major role for CFTR in this function. These findings show that it is possible to model CFTR defects in iPSC-derived cholangiocyte-like cells and to use these populations to test patient-specific response to CFTR modulatory compounds.
DISCUSSIONProgress in uncovering the causes of biliary disorders and developing new therapies has been hampered by inaccessibility of the target tissue in patients and a lack of experimental model systems. Our study demonstrates an approach to model human biliary development and disease through a directed differentiation protocol that efficiently generates functional cholangiocyte cells from hPSCs that self-organize into duct-like structures in vitro and ectopically in vivo. This protocol was designed to recapitulate key aspects of cholangiocyte develop-ment in vivo, including the generation of hepatoblasts, specification of this population to a cholangiocyte fate, and induction and matu-ration of epithelialized duct-like structures. Through co-culture of OP9 cells with hPSC-derived hepatoblasts, we were able to mimic the Jagged1-Notch2 interaction that regulates mouse cholangiocyte devel-opment9–12 and demonstrate that generation of the human cholangio-cyte lineage is also dependent on NOTCH signaling. Cholangiocyte specification with the co-culture approach appears to be very efficient as >90% of the structures that formed in 3D-cultures consisted of cholangiocyte-like cells and displayed bile duct characteristics.
DAPI CK19 CFTR
(+) Correction
C1-iPSC
GM00997-iPSC
250
130
95
(MW)(kD)
VX-809 + Corr-4a
HBE
α CFTRC
BA
α CNX
(–)
(–) Correction
a
(+) Correction
C1-iPSC GM00997–
iPSC
(–) (+)
0
100
200
300
400
0
50
100
150
200
250
VX-809 + Corr-4a(+)(+) (+)(+) (+) (+)(–) (–) (–)(+) (+) (+)
FSK/IBMXplus VX-770
*** *** *
*** *** ***
C1-iPSC
% s
wel
ling
rela
tive
tono
nind
uced
cys
ts
(+)(–) (+)
GM00997-iPSC
b
c d
Figure 7 Functional analysis of cholangiocytes from CF patient iPSCs. (a) Confocal images of histological sections showing the presence of the mutant CFTR protein in 16-d-old cyst structures generated from CF-iPSC-derived cholangiocytes cultured as chimeric aggregates in the presence or absence of the chemical correctors VX-809 (3 µM) and Corr-4a (5 µM). The correctors were added at 2 weeks of culture. Forty-eight hours after the addition of the correctors, the structures were fixed and analyzed for the presence of CFTR. Lower left panel shows the presence CFTR protein (Cy3, red) in the cysts treated with the correctors. Membrane localized CFTR was not detected in the nontreated cysts (upper left panel). Right panel shows co-expression of CFTR and CK19 in a treated cyst. CK19, Alexa 488 (green); CFTR, Cy3 (red); nuclei, DAPI (blue). Scale bars, 50 µm. Arrows indicate a perinuclear distribution pattern of CFTR protein. (b) Western blot analysis showing the presence of the mature glycosylated (band C) and core glycosylated (band B) form of the CFTR protein in the nontreated and VX-809/Corr-4a-treated 16-d-old cysts. HBE were used as a positive control. (c) Representative epifluorescence microscopy images of calcein-green-labeled cyst structures generated from cholangiocytes derived from CF-iPSC from two different patients (C1-iPSC and GM00997-iPSC). Images shown are before (−) and 24 h after (+) stimulation with forskolin, IBMX and the CFTR potentiator VX-770 (1 µM). Chimeric aggregates were generated in collagen/Matrigel gels in the presence of forskolin for the first week of the 16-d culture period. The correctors were added to the 2-week-old cultures for 48 h. At this stage, the cultures were stimulated with the combination of F/I and the potentiator VX-770. Cysts were measured 24 h later. Scale bars, 500 µm. (d) Quantification of the degree of swelling of the cysts derived from two different CF-iPSC lines. Cysts were cultured in the presence and absence of correctors, as indicated. Cyst swelling was quantified using velocity imaging software. The total size of the cyst was normalized to that before F/I stimulation from each of three individual experiments. Student’s t-test: *P < 0.05, *** P < 0.001 (n = 3).
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

860 VOLUME 33 NUMBER 8 AUGUST 2015 nature biotechnology
A rt i c l e s
Whereas Notch functions to specify the cholangiocyte lineage from the hepatoblast31, the Egf, Hgf and Tgfβ pathways play a role in the formation and maturation of the biliary ductal system in the develop-ing mouse8. EGF and HGF are thought to be required for stimulation of hepatoblast proliferation32, whereas TGFβ signaling is required to induce differentiation of biliary cells from periportal hepatoblasts24,33. We found that these pathways also influence cholangiocyte and duc-tal development in hPSC differentiation cultures. Whereas EGF and TGFβ signaling promoted proliferation of the CK19+ population, addition of HGF with these factors initiated formation of branched structures in two-dimensional (2D) monolayers. In 3D co-cultures, these factors promoted development of ductal and cystic struc-tures with lumens comprising cells that displayed morphologic and functional characteristics of mature biliary epithelial cells.
The presence of functional CFTR protein in hPSC-derived epithelial cells within 3D duct and/or cyst structures is strong evidence that they represent physiologically relevant biliary cells, as this trans-porter is known to be expressed on the apical membrane of mature bile ducts34,35. Patients with CF often develop liver dysfunction due to biliary cirrhosis36. Although there are no approved targeted therapies to treat CF patients, two correctors,VX-809 and VX-661, and a poten-tiator, VX-770, are currently in clinical trials. These molecules restore mutant plasma membrane CFTR expression and function to <15% of wild-type levels in assays using HBE cells37. Whereas HBE cell–based assays have been used to identify potential therapeutic compounds, it is not known whether the rescue effect observed predicts clinical efficacy as the response in different patient-derived HBE cultures is variable. Additionally, it remains to be determined whether compound efficacy is tissue-dependent. The observation that impaired swelling of cholangiocyte cysts from two different CF patients could be enhanced using a combination of CFTR correctors and potentiators demon-strates the potential of this hPSC-based assay for validating novel therapeutic drugs in both a tissue- and patient-specific manner.
Previous studies have identified cells with cholangiocyte character-istics in hepatic populations generated from hPSCs24,38,39. The most recent report7 used a protocol that involved activation of the EGF, GH (growth hormone) and IL-6 pathways to generate a 2D monolayer population with characteristics of cholangiocytes. As the NOTCH pathway was not directly manipulated in this study, it is unclear whether endogenous levels of NOTCH signaling were responsible for specification of these cells. Given that the overall efficiency of cholan-giocyte development was not reported, comparisons to our protocol are not possible. As cells in 2D monolayers often fail to display func-tional properties of their counterparts in complex tissues, we focused our efforts on developing a protocol that generates 3D structures that reflect the architecture and physiology of the human biliary system. The ability to derive such structures provides an in vitro system for modeling human biliary development and disease.
In summary, we established a developmentally relevant strategy for the efficient generation of functional cholangiocytes in monolayer and 3D culture from hESCs and iPSCs. With this approach, we were able to model biliary CF in vitro and demonstrate that the efficacy of potential therapeutic agents could be measured using a cyst-swelling assay. As this protocol provides access to different stages of cholangio-cyte development, it should enable modeling of other biliary diseases that will likely affect different aspects of bile duct morphogenesis and function. Finally, establishment of a protocol to generate hPSC-derived cholangiocytes presents an opportunity to further develop this technology for the evaluation of novel therapeutic agents and for tissue replacement applications.
METHODSMethods and any associated references are available in the online version of the paper.
Note: Any Supplementary Information and Source Data files are available in the online version of the paper.
ACKnOwLEdGMEntSWe would like to thank J. Rossant, J. Ellis and A.P. Wong (Hospital for Sick Children, Toronto, ON, Canada) for providing CF patient iPSCs (C1 and GM00997). We thank members of the G.K. and A.G. laboratories for discussion and feedback on the manuscript. We would like to thank F. Xu (Advanced Optical Microscopy Facility, University Health Network) for technical assistance with the time-lapse video. In addition, we would also like to thank O. Adeyi and the members of Department Pathology, University Health Network for technical assistance with immunohistochemistry. H9 hESC was obtained from The Wicell Research Institute (Madison, WI, USA), and MSC-iPSC1 cells were obtained from G.Q. Daley (Harvard Stem Cell Institute). This work was supported by funding from the McEwen Centre for Regenerative Medicine and the Canadian Institutes of Health Research MOP133620, (G.K.), the University Health Network Multi-Organ Transplant Program Academic Enrichment Fund (sponsored by Astellas Pharma Canada), Alagille Syndrome Alliance, SickKids Research Institute, Rare Disease Foundation and the Childhood Liver Disease Research and Education Network, U01 DK062453 (Sokol) from the National Institute of Diabetes, Digestive and Kidney Diseases (B.M.K. and A.G.) and the Canadian Institutes of Health Research (MOP:97954 and GPG:102171 to C.E.B.).
AUtHOR COntRIBUtIOnSS.O., M.O., G.K., B.M.K. and A.G. conceptualized the study. S.O. and M.O. led the experimental design and development of the differentiation protocol with input from all authors. S.O. and M.O. performed the hPSC differentiation and characterization of hPSC-derived cholangiocytes. M.O., C.E.B., S.A. and S.C. developed the cyst swelling assay for hPSC-derived cholangiocytes. B.L. and M.G. developed and used the ductal antibody for isolation of primary cholangiocytes from human liver. All authors analyzed and interpreted the data. M.O., S.O., G.K., B.M.K. and A.G. prepared the manuscript with contributions and critical revision from all authors.
COMPEtInG FInAnCIAL IntEREStSThe authors declare no competing financial interests.
reprints and permissions information is available online at http://www.nature.com/reprints/index.html.
1. O’Hara, S.P., Tabibian, J.H., Splinter, P.L. & LaRusso, N.F. The dynamic biliary epithelia: molecules, pathways, and disease. J. Hepatol. 58, 575–582 (2013).
2. Kamath, B.M. et al. Outcomes of liver transplantation for patients with Alagille syndrome: the studies of pediatric liver transplantation experience. Liver Transpl. 18, 940–948 (2012).
3. Si-Tayeb, K. et al. Highly efficient generation of human hepatocyte-like cells from induced pluripotent stem cells. Hepatology 51, 297–305 (2010).
4. Touboul, T. et al. Generation of functional hepatocytes from human embryonic stem cells under chemically defined conditions that recapitulate liver development. Hepatology 51, 1754–1765 (2010).
5. Takayama, K. et al. Generation of metabolically functioning hepatocytes from human pluripotent stem cells by FOXA2 and HNF1alpha transduction. J. Hepatol. 57, 628–636 (2012).
6. Ogawa, S. et al. Three-dimensional culture and cAMP signaling promote the maturation of human pluripotent stem cell-derived hepatocytes. Development 140, 3285–3296 (2013).
7. Dianat, N. et al. Generation of functional cholangiocyte-like cells from human pluripotent stem cells and HepaRG cells. Hepatology 60, 700–714 (2014).
8. Si-Tayeb, K., Lemaigre, F.P. & Duncan, S.A. Organogenesis and development of the liver. Dev. Cell 18, 175–189 (2010).
9. Zong, Y. et al. Notch signaling controls liver development by regulating biliary differentiation. Development 136, 1727–1739 (2009).
10. Hofmann, J.J. et al. Jagged1 in the portal vein mesenchyme regulates intrahepatic bile duct development: insights into Alagille syndrome. Development 137, 4061–4072 (2010).
11. Kodama, Y., Hijikata, M., Kageyama, R., Shimotohno, K. & Chiba, T. The role of notch signaling in the development of intrahepatic bile ducts. Gastroenterology 127, 1775–1786 (2004).
12. Geisler, F. et al. Liver-specific inactivation of Notch2, but not Notch1, compromises intrahepatic bile duct development in mice. Hepatology 48, 607–616 (2008).
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature biotechnology VOLUME 33 NUMBER 8 AUGUST 2015 861
A rt i c l e s
13. Li, L. et al. Alagille syndrome is caused by mutations in human Jagged1, which encodes a ligand for Notch1. Nat. Genet. 16, 243–251 (1997).
14. McDaniell, R. et al. NOTCH2 mutations cause Alagille syndrome, a heterogeneous disorder of the notch signaling pathway. Am. J. Hum. Genet. 79, 169–173 (2006).
15. Kamath, B.M. et al. NOTCH2 mutations in Alagille syndrome. J. Med. Genet. 49, 138–144 (2012).
16. Gouon-Evans, V. et al. BMP-4 is required for hepatic specification of mouse embryonic stem cell-derived definitive endoderm. Nat. Biotechnol. 24, 1402–1411 (2006).
17. Zaret, K.S. & Grompe, M. Generation and regeneration of cells of the liver and pancreas. Science 322, 1490–1494 (2008).
18. Suzuki, A., Sekiya, S., Buscher, D., Izpisua Belmonte, J.C. & Taniguchi, H. Tbx3 controls the fate of hepatic progenitor cells in liver development by suppressing p19ARF expression. Development 135, 1589–1595 (2008).
19. Lüdtke, T.H., Christoffels, V.M., Petry, M. & Kispert, A. Tbx3 promotes liver bud expansion during mouse development by suppression of cholangiocyte differentiation. Hepatology 49, 969–978 (2009).
20. Raynaud, P., Carpentier, R., Antoniou, A. & Lemaigre, F.P. Biliary differentiation and bile duct morphogenesis in development and disease. Int. J. Biochem. Cell Biol. 43, 245–256 (2011).
21. Schmitt, T.M. & Zuniga-Pflucker, J.C. Induction of T cell development from hematopoietic progenitor cells by delta-like-1 in vitro. Immunity 17, 749–756 (2002).
22. Lehar, S.M., Dooley, J., Farr, A.G. & Bevan, M.J. Notch ligands Delta 1 and Jagged1 transmit distinct signals to T-cell precursors. Blood 105, 1440–1447 (2005).
23. Fernández-Sánchez, V. et al. In vitro effects of stromal cells expressing different levels of Jagged-1 and Delta-1 on the growth of primitive and intermediate CD34(+) cell subsets from human cord blood. Blood Cells Mol. Dis. 47, 205–213 (2011).
24. Takayama, K. et al. CCAAT/enhancer binding protein-mediated regulation of TGFbeta receptor 2 expression determines the hepatoblast fate decision. Development 141, 91–100 (2014).
25. Dorrell, C. et al. Transcriptomes of the major human pancreatic cell types. Diabetologia 54, 2832–2844 (2011).
26. Irion, S. et al. Identification and targeting of the ROSA26 locus in human embryonic stem cells. Nat. Biotechnol. 25, 1477–1482 (2007).
27. Dekkers, J.F. et al. A functional CFTR assay using primary cystic fibrosis intestinal organoids. Nat. Med. 19, 939–945 (2013).
28. Molinski, S. et al. Functional rescue of F508del-CFTR using small molecule correctors. Front. Pharmacol. 3, 160 (2012).
29. Eckford, P.D., Li, C., Ramjeesingh, M. & Bear, C.E. Cystic fibrosis transmembrane conductance regulator (CFTR) potentiator VX-770 (ivacaftor) opens the defective channel gate of mutant CFTR in a phosphorylation-dependent but ATP-independent manner. J. Biol. Chem. 287, 36639–36649 (2012).
30. Okiyoneda, T. et al. Mechanism-based corrector combination restores DeltaF508-CFTR folding and function. Nat. Chem. Biol. 9, 444–454 (2013).
31. Tanimizu, N. & Miyajima, A. Notch signaling controls hepatoblast differentiation by altering the expression of liver-enriched transcription factors. J. Cell Sci. 117, 3165–3174 (2004).
32. Tanimizu, N., Saito, H., Mostov, K. & Miyajima, A. Long-term culture of hepatic progenitors derived from mouse Dlk+ hepatoblasts. J. Cell Sci. 117, 6425–6434 (2004).
33. Clotman, F. et al. Control of liver cell fate decision by a gradient of TGF beta signaling modulated by Onecut transcription factors. Genes Dev. 19, 1849–1854 (2005).
34. Turner, R. et al. Human hepatic stem cell and maturational liver lineage biology. Hepatology 53, 1035–1045 (2011).
35. Cardinale, V. et al. Multipotent stem/progenitor cells in human biliary tree give rise to hepatocytes, cholangiocytes, and pancreatic islets. Hepatology 54, 2159–2172 (2011).
36. Leeuwen, L., Fitzgerald, D.A. & Gaskin, K.J. Liver disease in cystic fibrosis. Paediatr. Respir. Rev. 15, 69–74 (2014).
37. Van Goor, F. et al. Correction of the F508del-CFTR protein processing defect in vitro by the investigational drug VX-809. Proc. Natl. Acad. Sci. USA 108, 18843–18848 (2011).
38. Zhao, D. et al. Derivation and characterization of hepatic progenitor cells from human embryonic stem cells. PLoS ONE 4, e6468 (2009).
39. Yanagida, A., Ito, K., Chikada, H., Nakauchi, H. & Kamiya, A. An in vitro expansion system for generation of human iPS cell-derived hepatic progenitor-like cells exhibiting a bipotent differentiation potential. PLoS ONE 8, e67541 (2013).
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature biotechnology doi:10.1038/nbt.3294
ONLINE METHODShPSC maintenance and differentiation. hPSCs were maintained on irradi-ated mouse embryonic feeder cells in hESC media consisting of DMEM/F12 (50:50; Gibco) supplemented with 20% knock-out serum replacement (KSR) as described previously40. Prior to the induction of endoderm in the monolayer cultures, hPSCs were passaged onto a Matrigel coated surface (typically 12-well plates) for 1 d. To initiate differentiation, the cells were cultured for 1 d in RPMI-based medium supplemented with glutamine (2 mM), MTG (4.5 × 10−4 M; Sigma), activin A (100 ng/ml), CHIR99021 (2 µM). At day 1, CHIR99021 was removed and cells were cultured for the next 2 d in RPMI supplemented with glutamine (2 mM), ascorbic acid (50 µg/ml: Sigma), MTG (4.5 × 10−4 M: Sigma), basic fibroblast growth factor (bFGF; 5 ng/ml), activin A (100 ng/ml) and for the after 2 d in serum-free-differentiation (SFD)-based medium with the same supplements. The media was changed every other day. At day 7, the definitive endoderm was specified to a hepatic fate by culture in H16 DMEM containing bFGF (40 ng/ml) and bone morphogenic protein (BMP4: 50 ng/ml) and supplemented with 1% vol/vol B27 supplement (Invitrogen: A11576SA), ascorbic acid, MTG. The media was changed every 2 d from day 7 to day 13. To promote expansion and maturation of the hepatoblast population, the cells were cultured in a mixture of H16 DMEM/Ham’s F12 (3:1) media with 0.1% BSA, 1% vol/vol B27 supplement, ascorbic acid, glutamine, MTG, hepatocyte growth factor (HGF: 20 ng/ml), dexamethasone (Dex: 40 ng/ml) and oncosta-tin M (OSM: 20 ng/ml) for 9 d. At day 21, the cells were cultured in a mixture of H21 DMEM/Ham’s F12 (3:1) with 0.1% BSA, 1% vol/vol B27 supplement, ascorbic acid, glutamine, MTG, HGF (20 ng/ml), Dex (40 ng/ml) and OSM (20 ng/ml) for 4 d. To generate aggregates the cells from the day-25 population were dissociated to make a small clump with collagenase and then cultured in 6-well ultralow cluster dishes at a concentration of 6 × 105 cells per well as described previously6. All cytokines were human and purchased from R&D Systems, unless stated otherwise. Embryoid bodies and monolayer cultures were maintained in a 5% CO2, 5% O2, 90% N2 environment. Aggregation cultures were maintained in a 5% CO2 in ambient air.
Cholangiocyte differentiation co-culture with OP9 cells. OP9 cells were maintained as described previously22. 30 Gy-irradiated OP9 cells were plated on Matrigel-coated wells (12-well plates) at a concentration of 2 × 105 per well in alpha-modified minimum essential media (α-MEM) supplemented with glutamine (2 mM) and 20% FBS. To induce the cholangiocyte differentiation of the hPSCs-derived hepatoblast population, day-27 hepatic aggregates were plated onto the irradiated OP9 cells and cultured for 9 d in H21 DMEM/Ham’s F12 (3:1) media supplemented with 0.1% BSA, 1% vol/vol B27 supplement, ascorbic acid, glutamine, MTG, HGF (20 ng/ml), epidermal growth factor (EGF: 50 ng/ml) and transforming growth factor β1 (TGFβ1: 5 ng/ml). To inhibit notch signaling, gamma-secretase inhibitor (GSI) L-685,485 (10 µM: Tocris) was included in the medium throughout the culture period. The medium was changed every 3 d.
Generation of chimeric 3D aggregates. Day-25 monolayer populations were dissociated by a combination of enzymatic (collagenase type I: Sigma C0130) and mechanical dissociation as previously described6. For the generation of chimeric aggregates, irradiated OP9 cells were mixed with the dissociated hepatic cells at a ratio of 1:4. To promote the formation of aggregates, the cells were cultured in 6-well ultralow cluster dishes at a concentration of 6 × 105 cells per well for 2 d.
Generation of 3D gel culture. The chimeric aggregates were suspended in a mixture of type I collagen (1.2 mg/ml: Nitta gelatin) and Matrigel (40%: BD) and plated at a volume of 200 µl in 12-well Millicell transwell culture inserts. Following a 2-h incubation at 37 °C to solidify the gel, 2 ml of H21 DMEM/Ham’s F12 (3:1) with 0.1% BSA, 1% vol/vol B27 supplement, ascorbic acid, glutamine, monothioglycerol, HGF (20 ng/ml), EGF (50 ng/ml) and TGFβ1 (5 ng/ml) was added to the chamber below the insert. Medium was changed every 2 d.
Flow cytometry. Flow cytometric analyses were done as described previ-ously41. Differentiated cells were dissociated into a single-cell suspensions. The cells in 3D aggregate and duct and/or cyst structure in collagen and Matrigel culture were collected by the dissociation with Hank’s solution containing
1 mg/ml collagenase type II (Worthington#LS004176) overnight at room temperature, as described previously6. Dead cells were excluded during flow cytometry analyses and gating was determined using isotype control. For cell surface marker analyses, staining was carried out in PBS with 10% FCS. For detection of intracellular proteins, staining was performed on cells fixed with 4% paraformaldehyde (PFA: Electron Microscopy Science, Hatfield, PA, USA) in PBS. Cells were permeabilized with 90% ice-cold methanol for 20 min for ALB, AFP, CK19 and Notch2 staining as previously described6. For the detec-tion of CFTR, after single-cell suspension, the cells were fixed with 4% PFA for 10 min and permeabilized with 0.1% Tween 20 in PBS for 20 min and blocked with 2% BSA in PBS for 10 min. After being washed twice in PBS with 10% FSC, the cells were subsequently incubated with anti-CFTR antibody (CF3: ab2784) for 1 h at room temperature and then with the secondary antibody IgM goat anti-mouse APC (Jackson ImmunoResearch) for 30 min at room temperature7. The stained cells were analyzed using an LSRII flow cytometer (BD). FACS antibodies and their dilution ratio are listed in Supplementary Table 1.
Immunostaining. Antibodies and immunostaining procedures are sum-marized in Supplementary Table 1. The staining with nonspecific isotypes matching the primary antibodies was carried out for control purposes. For staining of monolayers, cells were fixed with 4% PFA for 15 min and per-meabilized with 0.2% Triton X-100 or cold 100% methanol (TBX3, CFTR, ASBT) before blocking. Cells were washed three times with PBS for 10 min at room temperature (RT) before and after each staining step. All antibod-ies for monolayer were diluted in DPBS + 0.1% BSA + 0.1% Triton X-100. DAPI was used to counterstain the nuclei and staining was visualized using a fluorescent microscope (Leica CTR 6000) and images captured using Leica Application Suite software. For the detection of α tubulin/CK19 in monolayer cultures, the differentiation was carried out on Matrigel-coated cover slips, staining was analyzed using a confocal fluorescence microscope (Olympus Fluo View 1000 B laser scanning confocal) and images captured using the Olympus Application software. For the staining of the 3D aggregates or duct/cyst structures in collagen/Matrigel, samples were fixed with 4% PFA at 37 °C overnight, washed with normal saline (0.85% NaCl) and then embedded in 2% agar. The agar block was fixed with 10% PFA for 24 h, embedded in paraf-fin and sectioned. Paraffin-embedded sections were dewaxed with xylene, rehydrated, placed in Tris-EGTA-buffer (TES: 10 mM Tris, 0.5 mM EGTA, pH 9.0) and subjected to heat-induced (Microwave) epitope retrieval for 20 min. Tissue sections were permeabilized with cold 100% methanol for ZO-1, CFTR, SCTR, ASBT, E-cadherin and Ki-67 before blocking. For staining of normal human liver (34-year-old healthy female living liver donor), tissues were fixed in 4% PFA and then paraffin-embedded or frozen tissue sections were made. ProLong Gold Antifade with DAPI (Invitrogen) was used to counterstain the nuclei. All antibodies for 3D tissues were diluted in DPBS + 0.3% BSA + 0.3% Triton X-100. Staining of 3D gel culture and normal liver was analyzed using a confocal fluorescence microscope (Olympus Fluo View 1000 B laser scanning confocal) and images captured using the Olympus Application software.
Quantitative real-time PCR. Total RNA was prepared using RNAqueous Micro Kit (Ambion) and treated with RNase-free DNase (Ambion). 500 ng to 1 µg RNA was reverse-transcribed into cDNA using random hexamers and Oligo (dT) with Superscript III Reverse Transcriptase (Invitrogen). qPCR was performed on a MasterCycler EP RealPlex (Eppendorf) using a QuantiFast SYBR Green PCR Kit (Qiagen), as described previously42. Expression levels were normalized to the housekeeping gene TATA box binding protein (TBP). Oligonucleotide sequences are available in Supplementary Table 2. Control samples of RNA of adult liver (AL), fetal liver (FL), primary culture hepatocyte6 and FACS-sorted cholangiocyte are listed in Supplementary Table 3.
Isolation of human cholangiocyte from liver tissue. Human primary liver cells were perfused with EBSS (Gibco, CA) with 1 mM EGTA (Fisher Chemical, NJ) and then collagenase type 2 (Worthington, NJ). Hepatocytes were depleted with a gravity spin of 500 r.p.m. for 3 min. Supernatant was spun at a speed of 1,400 r.p.m. for 5 min, nonparenchymal cells (NPCs) were included in the pellets. NPC cells were labeled at 4 °C for 20 min with FITC conjugated anti-DHIC5-4D9, APC-H7 CD45 (BD Pharmingen, CA) and APC-H7 CD14
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.

nature biotechnologydoi:10.1038/nbt.3294
(BD Pharmingen), APC-Cy7 anti- CD34 (BioLegend). Cells were washed three times and resuspended in staining medium containing propidium iodide (PI) (1 µg/ml) (Sigma, St Louis, MO). Labeled cells were analyzed and sorted with a BD Influx sorter (BD Biosciences, CA) and Flowjo software (Flowjo, OR). Cholangiocytes were isolated in the DHIC5-4D9+CD45−CD14−CD34− fraction. All procedures and protocols were approved by OHSU Institutional Review Board.
Mice and mammary fat pad transplantation. NSG mice purchased from The Jackson Laboratory (Bar Harbor, ME) were kept in a specific pathogen–free mouse facility at the Toronto Medical Discovery Tower and used at 8–12 weeks of age. Experiments were performed in accordance with the protocol approved by University Health Network animal care committee. To generate cells for transplantation, day-25 hepatoblast populations derived from hESC2-RFP hESCs were cultured for 9 d on OP9 stroma in medium containing HGF, EGF and TGFβ1. Following culture, the cells were dissociated by trypsin-EDTA after collagenase treatment. One million cells of the mixture of cholangiocytes and OP9 cells were injected into the mammary fat pad of NSG mice. After 6–8 weeks, animals were euthanized and the mammary fat pad tissue isolated and fixed with 10% formalin. Fixed samples were embedded in paraffin, sec-tioned and stained with hematoxylin and eosin (H&E) stain for morphological analyses. For immunohistochemistry of RFP the paraffin-embedded sections were dewaxed with xylene, rehydrated, placed in Tris-EGTA-buffer (TES; 10 mM Tris, 0.5 mM EGTA, pH 9.0) and subjected to heat-induced (Microwave) epitope retrieval. For detection of RFP, the samples were first incubated with an anti-RFP antibody (Abcam ab34771) for 1 h at room temperature after three times wash with PBS and then with a biotinylated goat anti–rabbit IgG (H+L) secondary antibody (Vector Labs, BA-1000). The color was developed with DAB. Antibodies and procedures for CFTR/CK19 and α-tubulin/CK19 are summarized in Supplementary Table 1.
Assay for MDR1 transporter of rhodamine 123. The cystic structures used for these assays were generated from hPSC-derived day-27 chimeric aggregates by culture in the collagen/Matrigel gels for 17 d. Prior to performing the trans-porter assay, the 3D gel culture was washed three times with PBS and then incu-bated with phenol red–free Hank’s balanced salt solution containing 100 µM rhodamine 123 (Sigma-Aldrich) for 10 min. Following the incubation, the gel was washed with fresh Hank’s balanced salt solution three times. To inhibit MDR1 transporter activity, the cultures were incubated with 20 µM R-(+)-verapamil (Sigma-Aldrich) for 30 min before adding rhodamine 123. The stained cells were visualized using a fluorescent microscope (Leica CTR6000) and images captured using the Leica application suite software.
CFTR functional assay. The CFTR functional assay was designed as a modification of similar assays developed for the analyses of gut organoids27. 3D gels containing 16-d-old cysts were incubated for 30 min in Hank’s buffered solution containing the live cell stain, calcein-AM (10 µM). Thereafter, the stain-containing solution was replaced with stain-free Hank’s buffer and the cysts imaged using live-cell imaging at 37 °C (Nikon Epifluorescence microscope). The gels were subsequently incubated in Hank’s buffer containing the cAMP agonists–forskolin (10 µM) and the nonselective PDE inhibitor IBMX (3-isobutyl-1-methylxanthine, 100 µM) (F/I) for 1 h or 24 h. One transwell in each experiment was pre-incubated with 30 µM CFTR-specific
inhibitor–Inh-172 (Cystic Fibrosis Foundation Therapeutics-CFFT) or DMSO control for 3 h. After 1 h or 24 h of incubation with F/I in presence of CFTR Inh-172 or DMSO, the gels were restained with calcein-AM (calcein green) and the cysts imaged again using live-cell epifluorescence imaging. The total area of each cyst was calculated before and 1 h and 24 h after stimulation using ImageJ (NIH–version 1.47) after exportation of the images from Volocity image analysis software (PerkinElmer–version 6.3). The change in total area of each cyst after 24 h stimulation was calculated and plotted. 10–20 cystic structures from each transwell were measured for each experiment (n = 3).
Differentiation, analysis and functional correction of cholangiocytes from CF patient iPSCs. iPSCs from two different CF patients carrying the common F508del mutation (C1 and GM00997) were obtained from the Cystic Fibrosis Centre at the Hospital for Sick children in Toronto43,44. The cells were maintained and differentiated as described above with the exception that the initial induction with CHIR99021 (1 µM) was carried out for 2 d rather than 1 d. Cystic structures were generated by culture of chimeric aggregates in the collagen/Matrigel gels, as detailed above. For the CF patient iPSCs, cyst development in the gels was dependent on the addition of forskolin (10 µM) to the media for the first week of the 16-d-culture period. Media was changed every two d. The chemical correctors VX-809 (3 µM: Cystic Fibrosis Foundation Therapeutics-CFFT) and Corr-4a (5 µM: Cystic Fibrosis Foundation Therapeutics-CFFT) were added into the medium 2 d before per-forming CFTR function assay. For the stimulation of CFTR function, the CFTR potentiator VX-770 (1 µM: Cystic Fibrosis Foundation Therapeutics-CFFT) was added together with F/I. The degree of cyst swelling was determined as described above.
Western blot analysis. For detection of CFTR protein, the gels containing the cystic structures were dissociated by culture overnight in Hank’s solution containing 1 mg/ml collagenase type II (Worthington #LS004176) overnight as previously described6. The cysts were lysed in RIPA buffer supplemented with cOmplete protease inhibitor (Roche) and Laemmle buffer. Lysates were run on SDS-PAGE gel and transferred to nitrocellulose membranes. The membranes were blocked with 5% skim milk in 1× PBST (0.1% Tween, 10 mM sodium phosphate, pH 7.2 and 150 mM NaCl in ddH2O), incubated overnight at 4 °C with a mouse monoclonal anti-CFTR specific antibody, 596 (1:500, University of North Carolina at Chapel Hill) and then for 1 h with a goat anti-mouse sec-ondary antibody (1:1,000, Cedarlane Labs). The blot was developed by chemi-luminescence using the ECL reagent (GE Healthcare, RPN2106) and analyzed using Odyssey Fc dual-mode imaging system (LI-COR Biosciences).
40. Kennedy, M., D’Souza, S.L., Lynch-Kattman, M., Schwantz, S. & Keller, G. Development of the hemangioblast defines the onset of hematopoiesis in human ES cell differentiation cultures. Blood 109, 2679–2687 (2007).
41. Nostro, M.C., Cheng, X., Keller, G.M. & Gadue, P. Wnt, activin, and BMP signaling regulate distinct stages in the developmental pathway from embryonic stem cells to blood. Cell Stem Cell 2, 60–71 (2008).
42. Nostro, M.C. et al. Stage-specific signaling through TGFbeta family members and WNT regulates patterning and pancreatic specification of human pluripotent stem cells. Development 138, 861–871 (2011).
43. Wong, A.P. et al. Directed differentiation of human pluripotent stem cells into mature airway epithelia expressing functional CFTR protein. Nat. Biotechnol. 30, 876–882 (2012).
44. Hotta, A. et al. EOS lentiviral vector selection system for human induced pluripotent stem cells. Nat. Protoc. 4, 1828–1844 (2009).
npg
© 2
015
Nat
ure
Am
eric
a, In
c. A
ll rig
hts
rese
rved
.





























