Embed Size (px)
Citation preview

Direct and correlated responses to upward and downward selection foroutcrossing in Vicia faba
Marıa Jose Suso a,*, Fouad Maalouf b
a Instituto de Agricultura Sostenible, CSIC, Apdo. 4084, Cordoba, Spainb Biodiversity and Integrated Gene Management, ICARDA, Box 5466, Aleppo, Syria
Field Crops Research 116 (2010) 116–126
A R T I C L E I N F O
Article history:
Received 5 May 2009
Received in revised form 1 December 2009
Accepted 2 December 2009
Keywords:
Mating system
Floral traits
Faba bean breeding
Pollination
Sustainable agriculture
Crop improvement
A B S T R A C T
Upward and downward artificial selection for outcrossing were performed in faba bean open-pollinated
populations: (1) to characterize the mating systems of the populations resulting from upward/
downward selection of allogamy; (2) to assess the magnitude and direction of differentiation in floral
phenotypes among selected and control populations; and (3) to determine its relationship to
experimentally imposed selection.
Field experiments were conducted using two original synthetic populations derived from different
genetic pools; the control populations (C) and two upward (S+), and two downward (S�) selected
populations. In these populations, design and display of floral traits grouped into functional categories
was studied. Outcrossing rates were estimated using codominant isozyme markers, and the mixed
mating and the multilocus methods. The simultaneous responses of functional floral traits to selection
for allogamy/autogamy were analyzed by using multivariate Discriminant Analysis.
Whether selection was for increased or decreased outcrossing, the four selected groups shifted in the
opposite direction of the selected type. Discriminant Analysis revealed how the patterns of increased or
decreased outcrossing in the selected populations were due to a concordant multidimensional change in
floral traits, thus limiting or contra-balancing the effects of artificial selection. Indirect selection does not
act on traits individually, but on combinations of traits that are functionally integrated. Our results
provide evidence that evolution towards self-fertilization has occurred by different genetic pathways
after upward selection for outcrossing. This occurrence was by autogamous and by geitonogamous
selfing.
For the improvement of faba bean populations in the context of sustainable agriculture, direct
selection on outcrossing cannot be a selection criterion. However, floral traits, in combination with
pollinator behaviour, should be used as indirect selection criteria to increase allogamy. This approach, in
turn, will allow the development of both pollinator-friendly varieties and enhance the environmental
services of faba beans.
� 2009 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Field Crops Research
journal homepage: www.e lsev ier .com/ locate / fc r
1. Introduction
Vicia faba L. has been considered as a partially allogamousspecies in all genetic and breeding studies carried out to date. Thelevel of allogamy can be both considerable and variable, rangingfrom 4 to 84% with a mean around 30–60% (Bond and Pope, 1974;Link, 1990; Link et al., 1994a; Suso and Moreno, 1999; Suso et al.,2001; Gasim et al., 2004). Outcrossing may vary widely geograph-ically (Link et al., 1994a; Suso and Moreno, 1999) depending onlocal environmental conditions, particularly the composition of thepollinator fauna (Bond and Kirby, 1999, 2001; Pierre et al., 1996,
* Corresponding author. Tel.: +34 957499237.
E-mail address: [email protected] (M.J. Suso).
0378-4290/$ – see front matter � 2009 Elsevier B.V. All rights reserved.
doi:10.1016/j.fcr.2009.12.001
1999). Among different European agro-ecological conditions, Susoet al. (2008) observed substantial variation in the outcrossing rateof faba bean; southwestern cultivars showing a consistently higheroutcrossing rate than northeastern cultivars. Additionally, undersouthwestern conditions, a tendency for higher selfing rates wasevident in the local cultivars when compared to the more widelyadapted cultivars.
The level of outcrossing potentially determines the level ofheterozygosity and the amount of realized heterosis. Therefore, themating system has an important impact on the performance ofimproved open-pollinated populations and synthetic varieties.Previous studies indicated that yield and yield stability andresistance to biotic and abiotic stresses were mediated by heterosisin faba bean cultivars and could be improved by increasing thelevel of heterozygosity (Link et al., 1994b, 1996; Abdelmula et al.,

M.J. Suso, F. Maalouf / Field Crops Research 116 (2010) 116–126 117
1999; Arbaoui and Link, 2008; Gasim and Link, 2007; Terzopouloset al., 2008; Maalouf et al., 2008), which could be achieved throughincreased outcrossing. This knowledge-based approach wasproposed by Suso et al. (2005) specifically for faba bean, and isbeing employed profitably to improve yield of wind-pollinatedcereals (Virmani, 1996; Abdel-Ghani et al., 2003, 2004; Parzies etal., 2008).
Cross-fertilization rates are at a high enough level (Metz et al.,1993, 1994) for breeding programmes to improve open-pollinatedpopulations and to develop synthetic varieties. Therefore, it isimportant to know whether it is possible to select for highoutcrossing rates in segregating populations. This would necessi-tate clear evidence that selection for a high level of outcrossingactually causes an increase in outcrossing rates.
Faba bean has a mixed mating system, and several authors haveproposed in their breeding programmes to transform the matingsystem of faba beans towards autogamy (Kambal et al., 1976;Adcock and Lawes, 1976; Bozzini and Chiaretti, 1999a). Recently,some authors consider that it is questionable whether the apparentsuperiority of synthetics over inbred lines holds true in organicagriculture. Inbred lines are uniform, thus giving the option to bespecifically adapted and, in addition, uniformity was apparentlyappreciated by organic farmers (Ghaouti et al., 2008). Thereforeselection for autogamy should be considered (Bozzini andChiaretti, 1999b).
One important prerequisite for the change after selection forhigh outcrossing or selfing is heritable genetic variation. Theinheritance of allogamy in faba bean was polygenic showing a highlevel of heritability and significant additive effects (Link, 1990;Metz et al., 1993, 1994). The feasibility of changing the faba beanmating system, either for increasing allogamy or autogamy, inpopulations never has been analyzed through selection experi-ments. There is no clear evidence of (1) to what extent the mixedmating system of faba bean populations could be altered byselection for autogamy or allogamy and (2) whether selection forincreasing or decreasing shift in the direction of selection, e.g., ifupward selection for outcrossing causes increased outcrossingrates in the resulting populations. There are not reports that showwhich floral traits are involved in the response for selection forallogamy/autogamy and which traits change as correlatedresponses to selection.
Another important prerequisite for the change in outcrossing isthe simultaneous answer of floral variation. Even if there isheritable variation in selection for outcrossing/selfing, their changemay be constrained if it cannot vary independently as aconsequence of phenotypic or genetic correlations (Lande andArnold, 1983).
Selection acting on one trait causes change not just in this onetrait but also in traits that are genetically correlated. This occursbecause genes are pleiotropic (affect several traits) or becausethey are linked (Falconer and Mackay, 1996). Under fieldconditions, the mating system of populations is affected by theavailability of pollinators, environment, and floral traits thateither facilitate selfing or outcrossing (Charlesworth, 2006). Theselection for allogamy may thus be accompanied by changes inphenology, design and display floral traits. Timing and structuralseparation of male and female stages of flowering i.e., synchrony,flower size, degree of self-incompatibility and heterostyly, andpollen-ovule ratio, appear to have accompanied changes in themating system (Barrett et al., 1996; Motten and Stone, 2000;Cruden, 2000).
Floral traits that are not under direct selection may still changesimultaneously and indirectly because of genetic correlations withselection for allogamy/autogamy. The details of how preciselyselecting for outcrossing/selfing can change floral traits areimportant, because they determine whether the necessary genetic
and phenotypic changes are likely to change, or difficult to change,thus affecting the rates of changes to allogamy/autogamy.
Legume breeding for sustainable agriculture is linked to thedevelopment of non-food services such as environmental services(Helenius and Stoddard, 2007). Breeders are encouraged to developa breeding approach that strives to integrate food production intothe healthy functioning of the agro-ecosystems (Allen-Wardellet al., 1998; Maria-Klein et al., 2007). Faba bean is visited by a greatnumber of solitary and social bees (Bond and Kirby, 1999, 2001;Pierre et al., 1996, 1999). The provision of floral resources withinthe crop for supporting beneficial insect pollinator populationscould be a promising strategy to enhance environmental functionof faba bean (Palmer et al., 2009). Selection for allogamy couldaffect faba bean floral attractiveness to pollinators. Thus the studyof the simultaneous response of floral traits to selection alsoprovides the opportunity to advance the required search for waysto develop the environmental services of faba beans. Moreover,there is a need for increasing the number of functional traits andfacilitating the interactions among crops and pollinators in thepollination network that can positively affect both services (Hajjaret al., 2008).
This study investigated floral trait patterns of variation incontrol and selected populations for autogamy/allogamy popula-tions of faba beans in order to elucidate the underlying processes.Specifically, we address the following questions: (1) is there adirect response to upward and downward selection on the level ofallogamy? (2) Are there simultaneous responses of floral traits toselection for allogamy/autogamy?
The objectives were therefore: (1) to characterize the matingsystems of the populations resulting from upward/downwardselection of allogamy; (2) to assess the magnitude and direction ofdifferentiation in floral phenotypes among selected and controlpopulations; and (3) to determine its relationship to experimen-tally imposed selection.
2. Materials and methods
2.1. Experimental design: selection protocol and source of seeds for
the experiments
Overview of the selection protocol is in Fig. 1. The original(‘‘base line’’) synthetic populations were described in detail byMaalouf et al. (1999). Briefly, seven lines, showing a wide range offloral variation and belonging to the four botanical groups of fababean were selected from the world germplasm collection of the IAS(Cordoba, Spain) to develop two experimental synthetic popula-tions, syn-4 and syn-5. Experimental populations were originallyconstructed by mixing identical numbers of seeds from theparental lines (six lines for syn-4, and seven lines for syn-5). Thesepopulations were not subject to artificial selection before beingused for this experiment.
Seeds of these synthetics were used to generate the populationsfor the artificial selection experiment. The level of outcrossing wasmeasured in syn-4 and syn-5 synthetics as described in detail bySuso et al., 2005. The families were ranked according to the level ofallogamy and the greatest allogamous and autogamous motherplants in syn-4 and syn-5 synthetics were identified. The motherplants from each syn with the most extreme values were used forthe artificial selection experiment.
‘‘Two-way’’ or ‘‘divergent’’ selection for high and low level ofoutcrossing was performed (Falconer and Mackay, 1996). Toperform artificial selection for high level of outcrossing, seeds from13 plants of the syn-4 and seeds from 12 plants from the syn-5with the most extreme values (level of outcrossing greater than0,70 in both synthetics) were bred to produce the selected upwardpopulations (S+). The size of the upward populations was 400

Fig. 1. Overview of the selection protocol.
M.J. Suso, F. Maalouf / Field Crops Research 116 (2010) 116–126118
seeds for syn-4 and 330 seeds from syn-5. The mean level ofoutcrossing of the selected seeds was 0.84 for syn-4 and 0.82 forsyn-5, respectively. The deviation between the mean of the entiremeasured population and that of the subset selected for breeding iscalled the directional selection differential (Lande and Arnold,1983), 0.84–0.51 = 0.33 in syn-4 and 0.82–0.48 = 0.34 in syn-5. Themother plants were weighted according to its seeds contribution.To perform artificial selection for low level of outcrossing, seedsfrom 11 plants of the syn-4 and seeds from 9 individuals from thesyn-5 with level of almost totally autogamy were bred to producethe downward populations (S�), 200 seeds for syn-4 and 290 fromsyn-5.
In order to minimize environmental differences due to climaticand general management, the obvious way of eliminatingenvironmental fluctuations from the assessment of the rate ofresponse of the selected population is by keeping unselectedcontrol synthetics (Falconer and Mackay, 1996). On the assump-tion that the environmental differences affect the selected andcontrol populations alike, and in order to avoid the effect ofenvironmental differences, two control synthetic populations, syn-4 (C) and syn-5 (C) were used as checks. By comparing traits inselected populations with those in the control synthetic popula-tions, the effects of selection acting in each direction could beassessed.
The trial was conducted at Instituto de Agricultura Sostenible(IAS), Cordoba, Spain, using standard agronomic practices for fababean. The environmental conditions correspond to a Mediterra-nean climate. In total, the experiment consisted of two groups ofthree field plots, upward and downward selected populations andcontrol populations for each synthetic. The synthetics were grownallowing for natural pollinators, mainly solitary bees of the speciesEucera numida Lep. (Pierre et al., 1999 and Suso, M.J. personalverification), to visit the flowering plants. Flowers of the syntheticspopulations were of the usual colour (the standard petal is whitewith anthocyanin veins and each of the two wing petals bears alarge dark spot). Every plot was completely surrounded by a 3 mstrip of Vicia narbonensis. V. narbonensis was used because of itssimilarity to V. faba, and because it is an autogamous crop, it woulddiscourage insect pollinators from leaving the faba bean plots.Mating system parameters were estimated and floral traits wereevaluated from plots at the following densities: space among rows0.7 m and among plants 0.1 m.
2.2. Floral traits measurements
Floral traits (Fig. 2) were recorded for 55 plants randomlychosen in each syn-4 and syn-5 groups and in each (S+), (S�), and(C) populations. The mean of two flowers from the fifth, sixth, or
seventh node and selected the same day was used as the trait valuefor each individual plant.
Floral traits were classified into three categories: (1) phenology,(2) design, and (3) display.
Flowering periods were determined on the basis of phenologi-cal states: beginning (FO), peak (PF), and end (FE) of flowering, andflowering duration (CICLO).
The floral design traits chosen for analysis included a variety ofsimple and composite measures of the standard and keel petals,and the ovary. For each plant, we measured two flowers betweenthe fifth and seventh nodes. We specifically chose flowers in the‘‘open flower’’ stage of development, as characterized by Osborneet al. (1997). Bees preferentially visit flowers in this stage,presumably because they are the easiest to manipulate, are themost attractive and offer suitable rewards. The mean measure-ment of the two flowers was used as the phenotypic value for eachplant. To measure flower size and shape, we used a digital scannerto record images of individual flowers and analyzed these imageswith the UTHSCSA Image Tool program (http//www.uthsca.edu/dig/itdesc.html). For each flower, we measured the followingtraits: maximum length of the standard petal, standard width,standard perimeter, length of the keel petal, keel width, keelopening, floral-tube length, style length, ovary length, the style-ovary angle, and secondary pollen presenter length.
We also measured floral display traits. Floral display traitsassociated with outcrossing may change over the plant-life, thusinvestigations of outcrossing must be considered within adevelopmental context. Thus, once a plant’s first flower opened,we counted the number of open flowers and the number ofinflorescences with open flowers once a week for the next 6 weeks.
Additionally, following Cresswell (1998), we classified floraltraits into functional categories: advertisement (e.g., standarddimensions and floral display), female sexual dimensions (e.g.,style length, ovary length), and fit to pollinator (e.g., keeldimensions and floral-tube length, angle between the style andovary, and secondary pollen presenter length).
2.3. Mating system parameters
Mating systems were analyzed by examining the segregationpatterns of allozymes in the (S+), (S�), and (C) populations of thetwo groups of syn-4 and syn-5 synthetics. The outcrossing rate of50 plants collected from each of (S+), (S�), and (C) populations wasestimated based on 15 seeds/plant. Protocols for enzyme extrac-tions, PAG electrophoresis and stain recipes were described bySuso et al. (1993). No microsatellites were needed becauseadequate polymorphisms were found by using allozymes (Parkeret al., 1998). Seven polymorphic loci were scored: 6Pgd, Skd, Est,
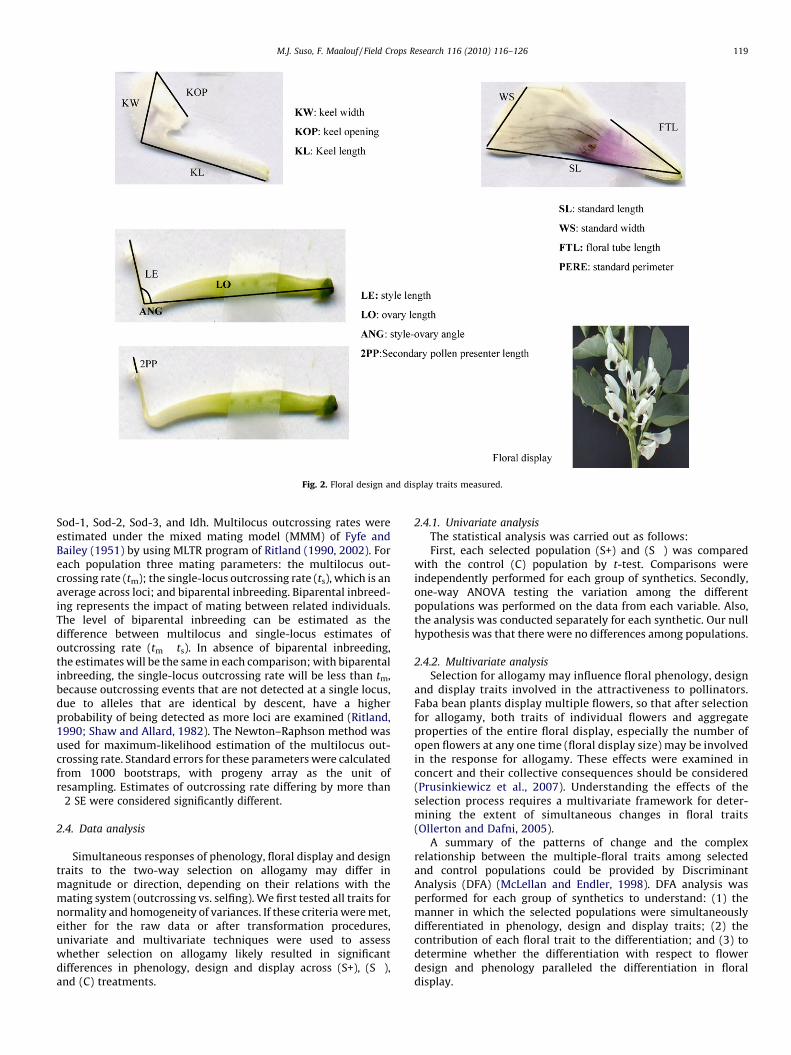
Fig. 2. Floral design and display traits measured.
M.J. Suso, F. Maalouf / Field Crops Research 116 (2010) 116–126 119
Sod-1, Sod-2, Sod-3, and Idh. Multilocus outcrossing rates wereestimated under the mixed mating model (MMM) of Fyfe andBailey (1951) by using MLTR program of Ritland (1990, 2002). Foreach population three mating parameters: the multilocus out-crossing rate (tm); the single-locus outcrossing rate (ts), which is anaverage across loci; and biparental inbreeding. Biparental inbreed-ing represents the impact of mating between related individuals.The level of biparental inbreeding can be estimated as thedifference between multilocus and single-locus estimates ofoutcrossing rate (tm � ts). In absence of biparental inbreeding,the estimates will be the same in each comparison; with biparentalinbreeding, the single-locus outcrossing rate will be less than tm,because outcrossing events that are not detected at a single locus,due to alleles that are identical by descent, have a higherprobability of being detected as more loci are examined (Ritland,1990; Shaw and Allard, 1982). The Newton–Raphson method wasused for maximum-likelihood estimation of the multilocus out-crossing rate. Standard errors for these parameters were calculatedfrom 1000 bootstraps, with progeny array as the unit ofresampling. Estimates of outcrossing rate differing by more than�2 SE were considered significantly different.
2.4. Data analysis
Simultaneous responses of phenology, floral display and designtraits to the two-way selection on allogamy may differ inmagnitude or direction, depending on their relations with themating system (outcrossing vs. selfing). We first tested all traits fornormality and homogeneity of variances. If these criteria were met,either for the raw data or after transformation procedures,univariate and multivariate techniques were used to assesswhether selection on allogamy likely resulted in significantdifferences in phenology, design and display across (S+), (S�),and (C) treatments.
2.4.1. Univariate analysis
The statistical analysis was carried out as follows:First, each selected population (S+) and (S�) was compared
with the control (C) population by t-test. Comparisons wereindependently performed for each group of synthetics. Secondly,one-way ANOVA testing the variation among the differentpopulations was performed on the data from each variable. Also,the analysis was conducted separately for each synthetic. Our nullhypothesis was that there were no differences among populations.
2.4.2. Multivariate analysis
Selection for allogamy may influence floral phenology, designand display traits involved in the attractiveness to pollinators.Faba bean plants display multiple flowers, so that after selectionfor allogamy, both traits of individual flowers and aggregateproperties of the entire floral display, especially the number ofopen flowers at any one time (floral display size) may be involvedin the response for allogamy. These effects were examined inconcert and their collective consequences should be considered(Prusinkiewicz et al., 2007). Understanding the effects of theselection process requires a multivariate framework for deter-mining the extent of simultaneous changes in floral traits(Ollerton and Dafni, 2005).
A summary of the patterns of change and the complexrelationship between the multiple-floral traits among selectedand control populations could be provided by DiscriminantAnalysis (DFA) (McLellan and Endler, 1998). DFA analysis wasperformed for each group of synthetics to understand: (1) themanner in which the selected populations were simultaneouslydifferentiated in phenology, design and display traits; (2) thecontribution of each floral trait to the differentiation; and (3) todetermine whether the differentiation with respect to flowerdesign and phenology paralleled the differentiation in floraldisplay.

M.J. Suso, F. Maalouf / Field Crops Research 116 (2010) 116–126120
Pearson’s correlation coefficients among floral traits werecalculated, prior to the DFA. For the Discriminant Analysis, onlyfloral traits not highly correlated with each other and significantlydifferent among the populations (ANOVA and t-tests) wereincluded.
All statistical management was conducted using statisticalprogram package Statistica v.7 (StatSoft).
3. Results
3.1. Mating system of the two-way selected and control synthetic
populations
Table 1 shows the multilocus (tm), single locus average (ts)outcrossing rates, and biparental inbreeding for the selected andcontrol populations of the two groups of synthetics. Both syn-4 andsyn-5 controls retain evidence of their history of mating andexhibit a mixed mating system with about 50% of progenyresulting from outcrossing (0.46 and 0.49, respectively).
The pattern of responses to outcrossing selection showed atrend opposite to that expected. The expectation was thatoutcrossing would be higher in populations selected for highlevels of allogamy and it would be lower in populations selected forhigh levels of autogamy. However, in our two-way selectionexperiments, no agreement with the expected responses wasfound.
Responses to upward selection (S+) for allogamy of both twogroups of synthetics were in the same direction. Selection for highallogamy levels significantly reduced the outcrossing rate orincreased the selfing rate from 0.49 to 0.31 in synthetic 5:selection for high levels of allogamy significantly increased selfingrate by about one-half, from 0.47 to 0.27 in syn-4. In contrast,downward selection for outcrossing (S�), which is expected tocause a reduction in the level of allogamy, had no significant effecton selfing and selected populations returned to the level ofcontrols in both groups, syn-5 and syn-4. The level of allogamy of(S�) was not significantly different from the level of allogamy of(S+) in syn-4.
The single-locus outcrossing rate was lower than the multilocusrate for the controls and each selection in both groups ofsynthetics, but not significantly different. Estimates of biparentalinbreeding were not significantly altered by the experimentaltreatments.
3.2. Simultaneous responses of floral traits to two-way selection
3.2.1. Univariate analysis
Results of the t-tests performed for individual charactersshowed differences among the selected (S+), (S�), and controlsynthetics. However, these differences depended on the synthetics(phenotypic means � SE for floral traits entering in the DFA model inthe selected and control populations) and are shown in Table 2.
Table 1Multilocus (tm), single locus average (ts) outcrossing rates and biparental
inbreeding for the selected and control populations of the two groups of synthetics
syn-4 and syn-5 of faba bean.
Synthetic Selection
level
Outcrossing rate Biparental
inbreeding
tm (SE) ts (SE) tm� ts (SE)
4 (S+) 0.27 (0.03) 0.20 (0.03) 0.07 (0.01)
4 (S�) 0.34 (0.05) 0.24 (0.04) 0.10 (0.02)
4 (C) 0.46 (0.03) 0.37 (0.03) 0.09 (0.01)
5 (S+) 0.31 (0.05) 0.23 (0.04) 0.08 (0.01)
5 (S�) 0.51 (0.06) 0.45 (0.06) 0.06 (0.01)
5 (C) 0.49 (0.04) 0.44 (0.03) 0.05 (0.01)
Compared to controls, downward selection in syn-4 producedplants with shorter and smaller size of standard and keels andsecondary pollen presenter length and earlier end of flowering. Thestyle-ovary angle also changed. In addition, downward selectionincreased the number of open flowers in the third, fourth, and fiveweeks of flowering, and number of inflorescences in the sixth weekof flowering. These floral display traits usually are considered astypical of allogamous plants. Compared to controls, upwardselection in syn-4 produced plants with shorter size of standard,and earlier end of flowering accompanied by shorter floweringduration. In addition, upward selection was accompanied by areduction in the number of open flowers and inflorescences formost of the flowering time. These floral traits usually areconsidered typical of autogamous plants.
Compared to controls, downward selection in syn-5 producedplants with smaller standard width, ovary length, length ofsecondary pollen presenter length, and delay of beginning andpeak of flowering, and shorter flowering duration. In relation tofloral display, downward selection led to fewer numbers of openflowers in the first week of flowering and reduced number of openinflorescences in the first, second, and third weeks of flowering,and increased number of open flowers in the fourth week. In syn-5group, the consequences of upward selection were only marginalfor flower size, but compared to controls, upward selectionradically changed the phenology. It leads to plants that startedflowering later and ended flowering earlier, so the duration of theflowering time was reduced. Again, these floral traits usually areconsidered typical of autogamous plants. In addition, the displayfloral traits changed along the flowering time. The number offlowers and inflorescences in the first week of flowering and thenumber of inflorescences in the sixth week decreased, and flowersin the second, third, and fourth weeks of flowering increased.
The ANOVA reduced from 26 floral traits to 16 the traits thatsignificantly differed among the (S+), (S�), and control populationsin the syn-4 group (phenotypic means � SE for floral traits enteringin the DFA model in the selected and control populations) and aregiven in Table 2. In addition to number of flowers and inflorescencesin the fourth and fifth weeks of flowering and number ofinflorescences in the sixth week of flowering, the following displaytraits: ovary, style and floral-tube length did not show any associatedresponse to the two-way selection. Peak period of flowering did notchange among selected and control populations.
The ANOVA reduced from 26 floral traits to 14 the traits thatsignificantly differ among the (S+), (S�), and control populations inthe syn-5 group (phenotypic means � SE for floral traits entering inthe DFA model in the selected and control populations) are given inTable 2. Floral design traits, with the exception of standard width andsecondary pollen presenter length, did not show any correlatedresponse to the two-way selection. In relation to the flower display,the number of flowers in the fifth and sixth weeks of flowering andnumber of inflorescences in the fourth week of flowering, did notshow any associated response to the two-way selection.
3.2.2. Multivariate analysis
(S+), (S�), and the control synthetics were compared byDiscriminant Analysis in each group of synthetics. We reportresults on the discriminant functions (DFI and DFII axes) in eachgroup of synthetics because they contained highly significantamount of discriminatory power (Table 3). They accounted forcomparable amounts of variation (around 60 and 40%, respective-ly) in both groups indicating that upward and downward selectionfor outcrossing resulted in significant shifts in plant floral traits.Besides, the number of major floral traits needed to analyse anddistinguish the effect of the associated response of floral traits tothe two-way selection has been reduced from 16 to 9 in the syn-4and from 14 to 9 in the syn-5 synthetics.
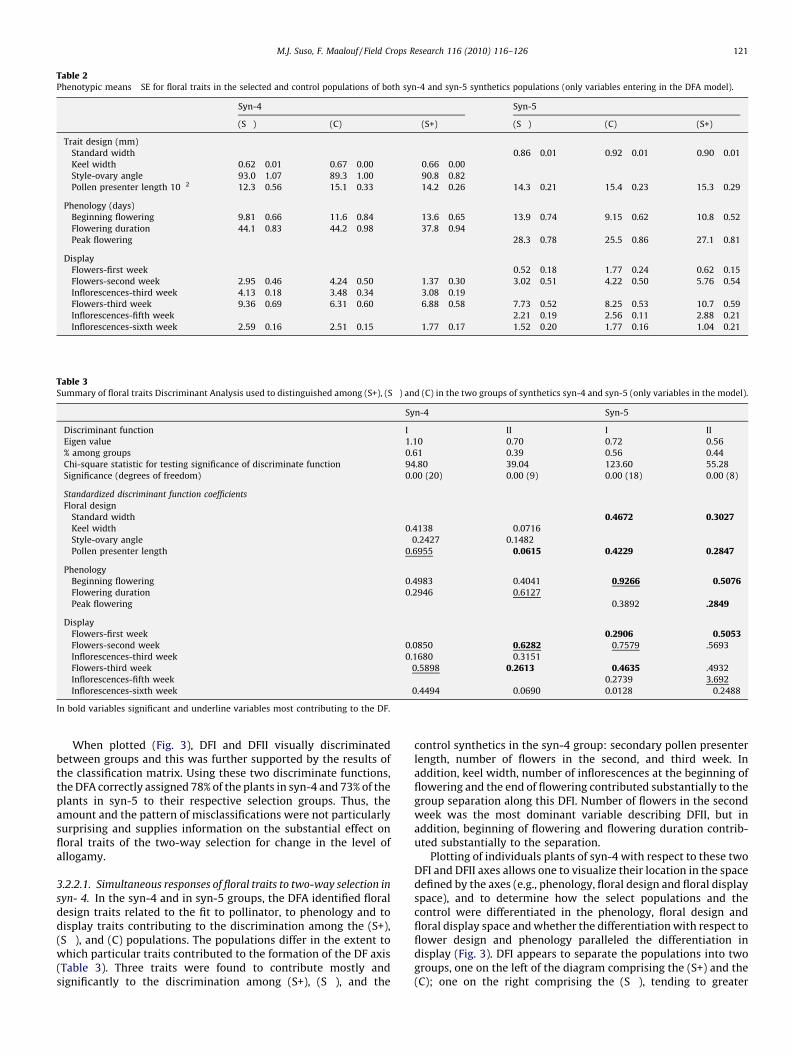
Table 2Phenotypic means� SE for floral traits in the selected and control populations of both syn-4 and syn-5 synthetics populations (only variables entering in the DFA model).
Syn-4 Syn-5
(S�) (C) (S+) (S�) (C) (S+)
Trait design (mm)
Standard width 0.86� 0.01 0.92� 0.01 0.90� 0.01
Keel width 0.62� 0.01 0.67� 0.00 0.66� 0.00
Style-ovary angle 93.0�1.07 89.3�1.00 90.8� 0.82
Pollen presenter length 10�2 12.3� 0.56 15.1� 0.33 14.2� 0.26 14.3� 0.21 15.4� 0.23 15.3� 0.29
Phenology (days)
Beginning flowering 9.81� 0.66 11.6� 0.84 13.6� 0.65 13.9� 0.74 9.15� 0.62 10.8� 0.52
Flowering duration 44.1� 0.83 44.2� 0.98 37.8� 0.94
Peak flowering 28.3� 0.78 25.5� 0.86 27.1� 0.81
Display
Flowers-first week 0.52� 0.18 1.77� 0.24 0.62� 0.15
Flowers-second week 2.95� 0.46 4.24� 0.50 1.37� 0.30 3.02� 0.51 4.22� 0.50 5.76� 0.54
Inflorescences-third week 4.13� 0.18 3.48� 0.34 3.08� 0.19
Flowers-third week 9.36� 0.69 6.31� 0.60 6.88� 0.58 7.73� 0.52 8.25� 0.53 10.7� 0.59
Inflorescences-fifth week 2.21� 0.19 2.56� 0.11 2.88� 0.21
Inflorescences-sixth week 2.59� 0.16 2.51� 0.15 1.77� 0.17 1.52� 0.20 1.77� 0.16 1.04� 0.21
Table 3Summary of floral traits Discriminant Analysis used to distinguished among (S+), (S�) and (C) in the two groups of synthetics syn-4 and syn-5 (only variables in the model).
Syn-4 Syn-5
Discriminant function I II I II
Eigen value 1.10 0.70 0.72 0.56
% among groups 0.61 0.39 0.56 0.44
Chi-square statistic for testing significance of discriminate function 94.80 39.04 123.60 55.28
Significance (degrees of freedom) 0.00 (20) 0.00 (9) 0.00 (18) 0.00 (8)
Standardized discriminant function coefficients
Floral design
Standard width 0.4672 0.3027Keel width 0.4138 �0.0716
Style-ovary angle �0.2427 0.1482
Pollen presenter length 0.6955 �0.0615 0.4229 0.2847
Phenology
Beginning flowering 0.4983 �0.4041 �0.9266 �0.5076Flowering duration 0.2946 �0.6127
Peak flowering �0.3892 .2849
Display
Flowers-first week 0.2906 �0.5053Flowers-second week 0.0850 �0.6282 �0.7579 .5693
Inflorescences-third week 0.1680 �0.3151
Flowers-third week �0.5898 0.2613 �0.4635 .4932
Inflorescences-fifth week 0.2739 3.692
Inflorescences-sixth week �0.4494 �0.0690 0.0128 �0.2488
In bold variables significant and underline variables most contributing to the DF.
M.J. Suso, F. Maalouf / Field Crops Research 116 (2010) 116–126 121
When plotted (Fig. 3), DFI and DFII visually discriminatedbetween groups and this was further supported by the results ofthe classification matrix. Using these two discriminate functions,the DFA correctly assigned 78% of the plants in syn-4 and 73% of theplants in syn-5 to their respective selection groups. Thus, theamount and the pattern of misclassifications were not particularlysurprising and supplies information on the substantial effect onfloral traits of the two-way selection for change in the level ofallogamy.
3.2.2.1. Simultaneous responses of floral traits to two-way selection in
syn- 4. In the syn-4 and in syn-5 groups, the DFA identified floraldesign traits related to the fit to pollinator, to phenology and todisplay traits contributing to the discrimination among the (S+),(S�), and (C) populations. The populations differ in the extent towhich particular traits contributed to the formation of the DF axis(Table 3). Three traits were found to contribute mostly andsignificantly to the discrimination among (S+), (S�), and the
control synthetics in the syn-4 group: secondary pollen presenterlength, number of flowers in the second, and third week. Inaddition, keel width, number of inflorescences at the beginning offlowering and the end of flowering contributed substantially to thegroup separation along this DFI. Number of flowers in the secondweek was the most dominant variable describing DFII, but inaddition, beginning of flowering and flowering duration contrib-uted substantially to the separation.
Plotting of individuals plants of syn-4 with respect to these twoDFI and DFII axes allows one to visualize their location in the spacedefined by the axes (e.g., phenology, floral design and floral displayspace), and to determine how the select populations and thecontrol were differentiated in the phenology, floral design andfloral display space and whether the differentiation with respect toflower design and phenology paralleled the differentiation indisplay (Fig. 3). DFI appears to separate the populations into twogroups, one on the left of the diagram comprising the (S+) and the(C); one on the right comprising the (S�), tending to greater
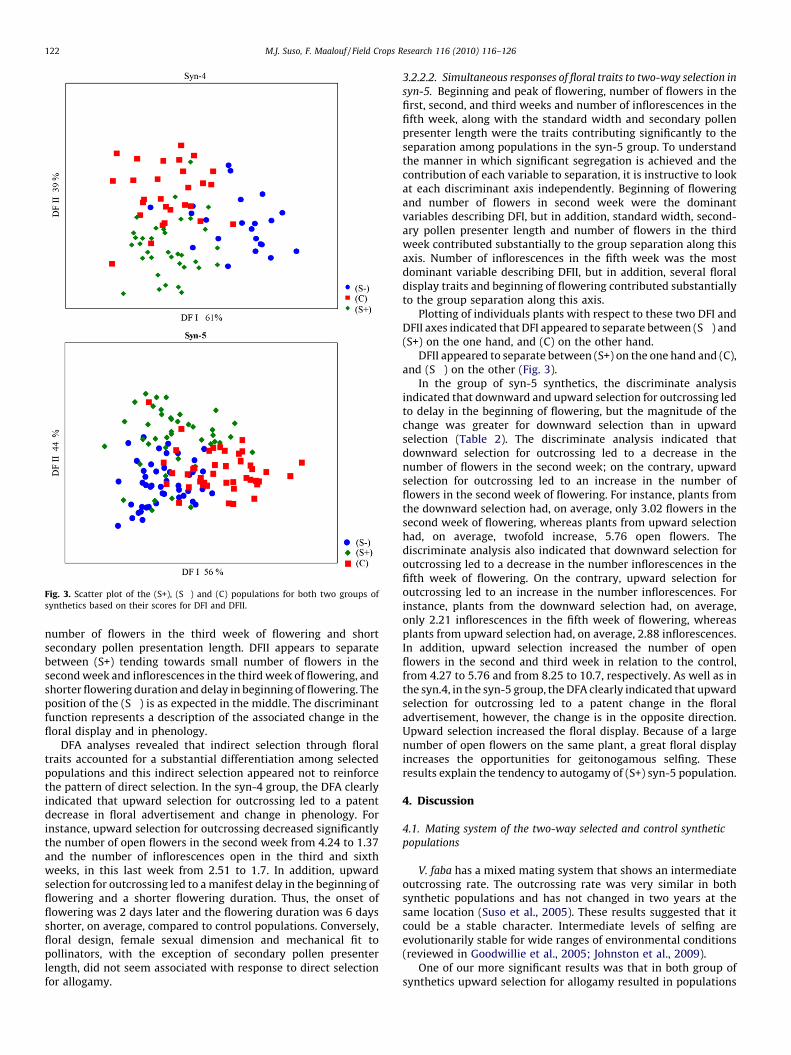
Fig. 3. Scatter plot of the (S+), (S�) and (C) populations for both two groups of
synthetics based on their scores for DFI and DFII.
M.J. Suso, F. Maalouf / Field Crops Research 116 (2010) 116–126122
number of flowers in the third week of flowering and shortsecondary pollen presentation length. DFII appears to separatebetween (S+) tending towards small number of flowers in thesecond week and inflorescences in the third week of flowering, andshorter flowering duration and delay in beginning of flowering. Theposition of the (S�) is as expected in the middle. The discriminantfunction represents a description of the associated change in thefloral display and in phenology.
DFA analyses revealed that indirect selection through floraltraits accounted for a substantial differentiation among selectedpopulations and this indirect selection appeared not to reinforcethe pattern of direct selection. In the syn-4 group, the DFA clearlyindicated that upward selection for outcrossing led to a patentdecrease in floral advertisement and change in phenology. Forinstance, upward selection for outcrossing decreased significantlythe number of open flowers in the second week from 4.24 to 1.37and the number of inflorescences open in the third and sixthweeks, in this last week from 2.51 to 1.7. In addition, upwardselection for outcrossing led to a manifest delay in the beginning offlowering and a shorter flowering duration. Thus, the onset offlowering was 2 days later and the flowering duration was 6 daysshorter, on average, compared to control populations. Conversely,floral design, female sexual dimension and mechanical fit topollinators, with the exception of secondary pollen presenterlength, did not seem associated with response to direct selectionfor allogamy.
3.2.2.2. Simultaneous responses of floral traits to two-way selection in
syn-5. Beginning and peak of flowering, number of flowers in thefirst, second, and third weeks and number of inflorescences in thefifth week, along with the standard width and secondary pollenpresenter length were the traits contributing significantly to theseparation among populations in the syn-5 group. To understandthe manner in which significant segregation is achieved and thecontribution of each variable to separation, it is instructive to lookat each discriminant axis independently. Beginning of floweringand number of flowers in second week were the dominantvariables describing DFI, but in addition, standard width, second-ary pollen presenter length and number of flowers in the thirdweek contributed substantially to the group separation along thisaxis. Number of inflorescences in the fifth week was the mostdominant variable describing DFII, but in addition, several floraldisplay traits and beginning of flowering contributed substantiallyto the group separation along this axis.
Plotting of individuals plants with respect to these two DFI andDFII axes indicated that DFI appeared to separate between (S�) and(S+) on the one hand, and (C) on the other hand.
DFII appeared to separate between (S+) on the one hand and (C),and (S�) on the other (Fig. 3).
In the group of syn-5 synthetics, the discriminate analysisindicated that downward and upward selection for outcrossing ledto delay in the beginning of flowering, but the magnitude of thechange was greater for downward selection than in upwardselection (Table 2). The discriminate analysis indicated thatdownward selection for outcrossing led to a decrease in thenumber of flowers in the second week; on the contrary, upwardselection for outcrossing led to an increase in the number offlowers in the second week of flowering. For instance, plants fromthe downward selection had, on average, only 3.02 flowers in thesecond week of flowering, whereas plants from upward selectionhad, on average, twofold increase, 5.76 open flowers. Thediscriminate analysis also indicated that downward selection foroutcrossing led to a decrease in the number inflorescences in thefifth week of flowering. On the contrary, upward selection foroutcrossing led to an increase in the number inflorescences. Forinstance, plants from the downward selection had, on average,only 2.21 inflorescences in the fifth week of flowering, whereasplants from upward selection had, on average, 2.88 inflorescences.In addition, upward selection increased the number of openflowers in the second and third week in relation to the control,from 4.27 to 5.76 and from 8.25 to 10.7, respectively. As well as inthe syn.4, in the syn-5 group, the DFA clearly indicated that upwardselection for outcrossing led to a patent change in the floraladvertisement, however, the change is in the opposite direction.Upward selection increased the floral display. Because of a largenumber of open flowers on the same plant, a great floral displayincreases the opportunities for geitonogamous selfing. Theseresults explain the tendency to autogamy of (S+) syn-5 population.
4. Discussion
4.1. Mating system of the two-way selected and control synthetic
populations
V. faba has a mixed mating system that shows an intermediateoutcrossing rate. The outcrossing rate was very similar in bothsynthetic populations and has not changed in two years at thesame location (Suso et al., 2005). These results suggested that itcould be a stable character. Intermediate levels of selfing areevolutionarily stable for wide ranges of environmental conditions(reviewed in Goodwillie et al., 2005; Johnston et al., 2009).
One of our more significant results was that in both group ofsynthetics upward selection for allogamy resulted in populations

M.J. Suso, F. Maalouf / Field Crops Research 116 (2010) 116–126 123
with increased selfing rates compared with the control, in one ofthe populations by as much as twofold. However, selection forautogamy increased the level of outcrossing in the descendentprogeny considerably and it did not change compared to thecontrol synthetic populations.
Whether selection was for increasing or decreasing allogamy,all the selected four groups shifted in the opposite direction of theselected type. We did achieve a significant increase on autogamybut when selecting for allogamy and in the process of selection forselfing (t almost zero), the progeny returned to the intermediatevalues of allogamy.
The following is a possible reason for the unexpected results.The mixed mating system of V. faba might result from a negativeassociation/genetic correlation between the level of allogamy andthe level of heterozygosis of the individual. If upward selection iscarried out, descendents of individuals with high levels ofoutcrossing must be heterozygous at more loci than descendentsof individuals with low levels of outcrossing. Thus they will show agreater tendency for selfing and vice versa if downward selection iscarried out with descendents of individuals with high levels ofselfing, which must be heterozygous at fewer loci than descen-dents of individuals with high levels of outcrossing. They will showa greater tendency to outcross.
The tendency of heterotic plants arising from outcrossing with ahigher selfing rate and vice versa, and plants arising from selfing tooutcrossing have been reported for other crops (Damgaard andLoeschcke, 1994), as well as for faba beans (Drayner, 1956, 1959;Link, 1990; Metz et al., 1993). Drayner (1956) indicated that theoverall effect was to maintain the population in a stable state ofintermediate hetererozygosity despite disturbing external pres-sures. This regulating mechanism represents a means of main-taining a moderate degree of heterozygosis without strongdependence on insect pollinators.
Our sampling scheme, though modest, is the only one to use anopen-pollinated population design. There have not been studies inwhich selection for both increase and decrease on outcrossing wasattempted with the same open-pollinated population. Theseattributes lend greater confidence to our identification of theeffect of selection for outcrossing. Despite the lack of replication,the consistency of the results across synthetics suggests that theresponse on populations to outcrossing selection is not straight-forward and it indicates that it is not going to be easy to move themating system of faba bean populations. We demonstrated withfaba beans that a shift from intermediate values of selfing towardshigh or low outcrossing could not be readily achieved. This patternthat has been showed in faba bean might be representative in otherpartially allogamous crops. Species with intermediate levels ofoutcrossing may not move readily toward outcrossing or selfing(Escalante et al., 1994).
4.2. Simultaneous responses of floral traits to two-way selection
Population genetic theory shows that if there are geneticcorrelations between the selected trait and other traits, then acorrelated response to selection can occur. Thus, genetic correla-tions among traits are important in the process of selection, asthey can slow or constrain the change, and the efficacy of artificialselection could be limited. Correlated responses between twophenotypic traits can be caused either by pleiotropy, in which onelocus affects both traits, or by linkage disequilibrium, in which thetwo traits are affected by distinct gene loci but some evolutionaryforce creates and maintains a nonrandom association between thealleles present at these loci. The genetic correlation determinesthe magnitude and direction of correlated response to selection ona correlated trait (Falconer and Mackay, 1996; Conner, 2002,2003).
V. faba plants exhibit spectacular variation in flower phenology,floral design and display, and much of the functional basis of thisdiversity is associated with levels of outcrossing (Suso et al., 2005).Flowering characteristics might influence the extent of outcrossingchange that can be expected. Our study showed that they wereconsistent changes in floral traits which could account for thealteration of the mating system after selection for outcrossing.Patterns of increase and decrease in outcrossing in the selectedpopulations were due to a concordant change in floral traits. Ourresults also illustrate and provide evidence that floral traits mayevolve in response to changes in mating patterns, thus limiting orcontra-balancing the effects of selection. Indirect selection in thiscase does not act on traits individually, but on combinations oftraits that are functionally integrated, that is, those that worktogether to affect plant floral display schedule.
Floral display schedule is a multivariate trait composed by thearrangement of open flowers and inflorescences on each week ofthe flowering time season. In animal-pollinated species, floraldisplay advertises rewards to potential pollinators, and therefore,floral displays have an important influence on mating patterns(reviews by Harder and Barrett, 1996; Harder et al., 2001).
Univariate and multivariate analyses showed that selection forallogamy has caused multiple-floral traits to change. It appearedthat the somewhat stronger selection to decrease or increaseoutcrossing produced the overriding indirect effect on the floraltraits. Thus, the level of outcrossing on the selected resultingpopulations arose as a by-product of associated selection for floraltraits. DFA analyses revealed that indirect selection through floraltraits accounted for a substantial differentiation among selectedpopulations (S+), (S�), and (C). However, this indirect selectionappeared not to reinforce the pattern of direct selection inconcordance with the results of the mating system estimation.
Upward and downward selection for outcrossing altered thefloral display schedule. The beginning of flowering, floweringduration and peak of flowering, and the number of flowers andinflorescences open each week of the flowering season were themajor traits that changed indirectly, after upward and downwardselection for outcrossing. Besides, discriminant analyses alsoindicated that some floral shape traits, standard and keel widthand mechanical fit to pollinator traits, the style-ovary angle andthe secondary pollen presenter length, responded strongly toselection for outcrossing. In contrast, a few floral size traits relatedto the advertisement, the sexual dimension traits and a fewmechanical fit to pollinators traits such as floral-tube length, keelopening and length did not change extensively in response toselection for outcrossing. Although the size, shape and symmetryof flowers (Barrett, 2003) are thought to be important for matingsystem evolution, changes on these traits were not significant inV. faba.
There, nevertheless, exist intra-specific variations in thecorrelated response of floral traits in which selection for out-crossing can evolve towards selfing. Selection for outcrossinginfluences the floral display schedule by changing the availablepool of flowers. The types of design and display changes that mightallow self-fertilization vary depending on the synthetic. Thepresent data suggested that phenological displacement tends todelay beginning of flowering and reduce flowering duration as aresult of upward selection in syn-4. However, syn-5 showed acontrasting pattern; downward selection delayed the beginningand the peak of flowering in syn-5. Outcrossing rates, are a productof floral design, display and phenology as well as pollinatoractivity. Flowering phenology is particularly important because itdetermines synchrony with or attractiveness to pollinators. Thus,differences in flowering phenologies among syn-populationsrelated to timing of pollinator availability during the seasonmay be a major factor affecting the responses to selection for

M.J. Suso, F. Maalouf / Field Crops Research 116 (2010) 116–126124
outcrossing. Faba beans have showed temporally fluctuatingenvironments in terms of pollinators visitation rate dynamics.During the flowering period, the gender of the main flower visitorchanges. Early in the season mainly E. numida males visit faba beanflowers. However, as flowering season progress, they are soonreplaced by E. numida females (Pierre et al., 1999). Plants of syn-5could have flowered over the relatively short period during whichE. numida males were visiting faba bean. We could hypothesizethat this seasonal change in visitors may be related to thedifferences in the response to selection for outcrossing.
Upward selection also was associated with changes in thearrangement of open flowers and inflorescences during theflowering time. Although, again, the mechanisms involved werevaried and depended on the synthetic. The number of open flowersand inflorescences decreased, floral traits that were clearly relatedto selfing, in syn-4 after upward selection for outcrossing, however,they increased in syn-5. The results of syn-4 were not necessarilyincompatible with those of syn-5, but provided evidence that theshift towards self-fertilization after selection for outcrossing hasoccurred by different genetic pathways. Whereas, a small floraldisplay was likely to increase autogamous selfing in syn-4, greaterfloral display could increase the geitonogamous selfing (selfingoccurs by pollinators moving from flower to flower on the sameplant) in syn-5. Self-pollination was not a single unvarying processthat always occurred in the same manner. On the contrary, self-pollination occurred in several fundamentally different ways or‘‘modes’’ (Lloyd and Schoen, 1992). Insects often visited moreflowers on plants with larger floral displays, which could increaseselfing rates via geitonogamy (Karron et al., 2004). The amount ofgeitonogamy may be varied by altering the number or dispositionof flowers or their individual attractiveness, which alters thenumber of successive visits that a pollinator makes to the flowersof a plant. All these changes, however, alter not only the amount ofgeitonogamy but also that of cross-pollination. Notice that floraldisplay traits are major traits loading in both DFI and DF II. A largenumber of open flowers on the same plant increase the probabilityof geitonogamous selfing, whereas a small number of open flowerswere likely related to autogamous selfing. Our results indicatedthat both modes of selfing were important in the faba beanpopulations.
Patterns of floral variation suggested that the mating systemwas rather firmly controlled by floral architecture and not, whichis commonly believed among breeders, more or less controlled bythe stochastic behaviour of pollinators and the general environ-ment. The occurrence of these different schemes providedevidence for multiple forms of self-fertilization in faba beanand also suggested that different genetic factors were involvedwith floral modification. The genetic architecture of floral traitsassociated with selfing/outcrossing maintaining outcrossing waslikely to be polygenic and it can presumably influence the tempoof the transition from outcrossing to selfing (Schoen and Busch,2008).
4.3. Relevance for practical breeding
Our results likely reflected the combined effects of variation ingenes controlling the outcrossing rate and genes with opposingeffects on multi-floral traits and suggested that the short-termevolution of outcrossing rate was necessarily constrained by floraltraits.
Correlations between characters seriously complicate pheno-typic selection, because selection on a particular trait producesnot only a direct effect on the distribution of that trait in apopulation, but also produces indirect effects on the distributionof correlated characters (Falconer and Mackay, 1996). Theproblem of the associated response of floral traits must not be
ignored in current methods for selection on outcrossing. Selectionfor high or low outcrossing should not be treated as if it affectsonly single characters. This is obviously a tremendous oversim-plification.
Studies to increase the level of outcrossing therefore should notbe conducted in isolation from other changes for phenotypesinvolving floral traits. According to Falconer and Mackay (1996),the consideration of correlated responses suggest that it mightsometimes be possible to achieve more rapid progress underselection for correlated response than from selection for thedesired trait itself. In other words, if we want to increase ordecrease outcrossing, we might select for determined floral traitsand achieve progress through the correlated response of traits,through indirect multiple-floral traits selection. The conditionunder which indirect selection could be expected to be advanta-geous over direct selection is when the trait has substantiallyhigher heritability and the genetic correlation is high. Besides,there are practical considerations that make indirect selectionpreferable, when the desired trait is difficult and costly to measureas it is in the present case. Davis (2001) reviewed floral structuralfeatures and nectar characteristics that could be considered forindirect selection to enhanced attractiveness of flowering speciesto nectar-gathering insects which may lead to improvements inpollination. Knapp and Teuber (1993) determined that selectionfor ease of tripping was an easily utilized indirect selection methodwhich may improve alfalfa seed yield, but did not alter theoutcrossing rate of the resulting populations. However, matingsystem depends on the frequency and quality of the interactionbetween pollen vectors and floral traits (Harder et al., 2004) andaddressing only outcrossing and floral traits would not be anefficient approach. Although indirect selection for floral traitscould be presented as an alternative to direct selection foroutcrossing, the most effective method is neither one nor the otherbut a combination of the two and an additional source ofinformation about the pollen vectors. In order to develop improvedtechniques for managing pollinator agents as effective tools forcrossing, future studies on the constraints to increase allogamy, forobtaining heterosis-mediated resistance to biotic and abioticstresses, should be focused on the analysis of the interactionbetween floral traits and pollinator behaviour. Furthermore,experiments that aim to disentangle the functional significanceof floral traits are justified, if the strategy is incorporated intobreeding schemes for the development of pollinator-friendlyvarieties and development of the environmental services of fababeans (Palmer et al., 2009).
Within the agricultural ecosystem, the services provided bypollinators and services provided by crops are interdependent.There is a need to increase the number of functional traits and tofacilitate the interactions among crop and pollinators in thepollination network that can positively affect both services (Hajjaret al., 2008). Our results emphasize the need to recover andimprove functional floral traits that may have been lost throughextended breeding for conventional systems. These traits arerelated to the ability of crops to benefit from associativerelationships with beneficial species such as insect pollinators.The shift of paradigm in agriculture not only happens through theuse of sophisticated techniques but through the development ofnatural tools. In legumes cases, which are visited by a great numberof bees (Free, 1993; Delaplane and Mayer, 2000), bees have afundamental function in the agro-ecosystem that can be veryuseful for the breeder to improve yield and yield stability. Thisfunction is related to the bee role as agents of pollination andagents of crossing, moving pollen from one plant to another, thusincreasing the level of heterozygosis and heterosis potential ofcultivars, which in turn will improve resistance to biotic andabiotic stresses. In parallel, legumes preserve bee fauna by

M.J. Suso, F. Maalouf / Field Crops Research 116 (2010) 116–126 125
providing foraging places and nesting sites for solitary and socialbees. Sustainable agriculture is not just production oriented butrelies largely on locally available resources, such as bees. This studyhas stressed the importance for further research on the interactiveand combined agro-ecological services provided by the two-wayinteraction of plant and pollinator for underpinning sustainableproduction systems. Besides, this breeding approach might givesignificant economic benefit to growers by promoting wild beepopulations that enhance seed production and yield stability(Morandin and Winston, 2006).
Acknowledgements
The authors thank Dr. Reid Palmer for his unquestionable helpin reviewing and commenting the different versions of themanuscript. The technical assistance of M. Santaella and R. delRio are also gratefully acknowledged. This research was supportedby AGL2005-07497-CO2-02 project.
References
Abdel-Ghani, A.H., Parzies, H.K., Ceccarelli, S., Grando, S., Geiger, H.H., 2003.Evaluation of floral characteristics of barley in the semi-arid climate of northSyria. Plant Breed. 122, 273–275.
Abdel-Ghani, A.H., Parzies, H.K., Omary, A., Geiger, H.H., 2004. Estimating theoutcrossing rate of barley landraces and wild barley populations collected fromecologically different regions of Jordan. Theor. Appl. Genet. 109, 588–595.
Abdelmula, A.A., Link, W., von Kittlitz, E., Stelling, D., 1999. Heterosis and inheri-tance of drought tolerance in faba bean (Vicia faba L). Plant Breed. 118, 485–490.
Adcock, M.E., Lawes, D.A., 1976. Self-fertility and the distribution of seed yield inVicia faba L. Euphytica 25, 89–96.
Allen-Wardell, G., Bernhardt, P., Bitner, R., Burquez, A., Buchmann, S., Cane, J., Cox,P.A., Dalton, V., Feinsinger, P., Ingram, M., Inouye, D., Jones, C.E., Kennedy, K.,Kevan, P., Koopowitz, H., Medellin, R., Medellin-Morales, S., Nabhan, G.P., Pavlik,B., Tepedino, V., Torchio, P., Walker, S., 1998. The potential consequences ofpollinator declines on the conservation of biodiversity and stability of food cropyields. Consers. Biol. 12, 8–17.
Arbaoui, M., Link, W., 2008. Effect of hardening on frost tolerance and fatty acidcomposition of leaves and stems of a set of faba bean (Vicia faba L.) genotypes.Euphytica 162, 211–219.
Barrett, S.C.H., Harder, L.D., Worley, A.C., 1996. The comparative biology of pollina-tion and mating in flowering plants. Philos. Trans. R. Soc. B: Biol. Sci. 351, 1271–1280.
Barrett, S.C.H., 2003. Mating strategies in flowering plants: the outcrossing-selfingparadigm and beyond. Philos. Trans. R. Soc. B: Biol. Sci. 358, 991–1004.
Bond, D.A., Pope, M., 1974. Factors affecting the proportions of cross-bred and selfedseed obtained from field bean (Vicia faba L.) crops. J. Agric. Sci. Camb. 83, 343–351.
Bond, D.A., Kirby, E.J.M., 1999. Anthophora plumipes (Hymenoptera: Anthophoridae)as a pollinator of broad bean (Vicia faba major). J. Apic. Res. 38, 199–203.
Bond, D.A., Kirby, E.J.M., 2001. Further observations of Anthophora plumipes visitingautumn-sown broad bean (Vicia faba major) in the United Kingdom. J. Apic. Res.40, 113–114.
Bozzini, A., Chiaretti, D., 1999a. The genetic improvement of the MediterraneanFaba bean (Vicia faba L.). II. Transfer into Mediterranean lines of progressivetraits found in other genetic pools: seed quality associated with pure whiteflower. J. Gent. Breed. 53, 201–206.
Bozzini, A., Chiaretti, D., 1999b. The genetic improvement of the MediterraneanFaba bean (Vicia faba L.). III. Development of obligate self fertile lines. J. Genet.Breed. 53, 207–213.
Charlesworth, D., 2006. Evolution of plant breeding systems. Curr. Biol. 16, R726–R735.
Conner, J.K., 2002. Genetic mechanisms of floral trait correlations in a naturalpopulation. Nature 420, 407–410.
Conner, J.K., 2003. Artificial selection: a powerful tool for ecologists. Ecology 84,1650–1660.
Cresswell, J.E., 1998. Stabilizing selection and the structural variability of flowerswithin species. Ann. Bot. 81, 463–473.
Cruden, R.W., 2000. Pollen grains: why so many. Plant Syst. Evol. 222, 143–165.Damgaard, C., Loeschcke, V., 1994. Genetic variation for selfing rate and the
dependence of selfing rate on mating history in Brassica napus (rape seed).Heredity 72, 570–573.
Davis, A.R., 2001. Searching and breeding for structural features of flowers corre-lated with high nectar-carbohydrate production. In: Benedek, P., Richards, K.W.(Eds.), 8th Pollination Symposium. pp. 107–121.
Delaplane, K.S., Mayer, D.F., 2000. Crop Pollination by Bees. CABI Publishing, NewYork.
Drayner, J.M., 1956. Regulation of outbreeding in field beans (Vicia faba). Nature177, 489–490.
Drayner, J.M., 1959. Self- and cross-fertility in field beans (Vicia faba L.). J. Agric. Sci.Camb. 53, 387–403.
Escalante, A.M., Coello, G., Eguiarte, L.E., Pinero, D., 1994. Genetic structure andmating systems in wild and cultivated populations of Phaseolus coccineus and P.vulgaris (Fabaceae). Am. J. Bot. 81, 1096–1103.
Falconer, D.S., Mackay, T.F.C., 1996. Introduction to Quantitative Genetics. PrenticeHall, London.
Free, J.B., 1993. Insect Pollination of Crops. Academic Press, San Diego, CA.Fyfe, J.L., Bailey, P.J., 1951. Plant breeding studies in leguminous forage crops. J.
Agric. Sci. 41, 371–378.Gasim, S., Abel, S., Link, W., 2004. Extent, variation and breeding impact of natural
cross-fertilization in German winter faba beans using hilum colour as marker.Euphytica 136, 193–200.
Gasim, S., Link, W., 2007. Agronomic performance and the effect of self-fertilizationon German winter faba beans. J. Cen. Eur. Agric. 8, 121–128.
Ghaouti, L., Vogt-Kaute, W., Link, W., 2008. Development of locally-adapted fababean cultivars for organic conditions in Germany through a participatorybreeding approach. Euphytica 162, 257–268.
Goodwillie, C., Kalisz, S., Eckert, C.G., 2005. The evolutionary enigma of mixedmating systems in plants: occurrence, theoretical explanations, and empiricalevidence. Ann. Rev. Ecol. Evol. Syst. 36, 47–79.
Hajjar, R., Jarvis, D.I., Gemmill-Herren, B., 2008. The utility of crop genetic diversityin maintaining ecosystem services. Agric. Ecosyst. Environ. 123, 261–270.
Harder, L.D., Barrett, S.C.H., 1996. Pollen dispersal and mating patterns in animal-pollinated plants. In: Lloyd, D.G., Barrett, S.C.H. (Eds.), Floral Biology: Studies onFloral Evolution in Animal-Pollinated Plants. Chapman & Hall, New York, pp.140–190.
Harder, L.D., Williams, N.M., Jordan, C.Y., Nelson, W.A., 2001. The effects offloral design and display on pollinator economics and pollen dispersal. In:Chittka, L., Thomson, J.D. (Eds.), Cognitive Ecology of Pollination. CambridgeUniversity Press, Cambridge, pp. 297–317.
Harder, L.D., Jordan, C.Y., Gross, W.E., Routley, M.B., 2004. Beyond floricentrism: thepollination function of inflorescences. Plant Spec. Biol. 19, 137–148.
Helenius, J., Stoddard, F.L., 2007. Agro-ecosystem services from increased usage andnovel applications of legumes. Integrating legume biology for sustainableagriculture. Book of Abstracts. In: Proceeding of the 6th European Conferenceon Grain Legumes, Lisbon, Portugal, 12–16 November.
Johnston, M.O., Porcher, E., Cheptou, P.O., Eckert, C.G., Elle, E., Geber, M.A., Kalisz, S.,Kelly, J.K., Moeller, D.A., Vallejo-Marin, M., Winn, A.A., 2009. Correlations amongfertility components can maintain mixed mating in plants. Am. Nat. 173, 1–11.
Knapp, E.E., Teuber, L.R., 1993. Outcrossing rates of alfalfa populations differing inease of floret tripping. Crop Sci. 33, 1181–1185.
Karron, J.D., Mitchell, R.J., Holmquist, K.G., Bell, J.M., Funk, B., 2004. The influenceof floral display size on selfing rates in Mimulus ringens. Heredity 92, 242–248.
Kambal, A.E., Bond, D.A., Toynbee-Clarke, G., 1976. A study on the pollinationmechanism in field beans. J. Agric. Sci. Camb. 87, 519–526.
Lande, R., Arnold, S.J., 1983. The measurement of selection on correlated characters.Evolution 37, 1210–1226.
Link, W., 1990. Autofertility and rate of cross-fertilization: crucial characters forbreeding synthetic varieties in faba bean. (Vicia faba L.). Theor. Appl. Genet. 79,713–717.
Link, W., Ederer, W., Metz, P., Buiel, H., 1994a. Genotypic and environmentalvariation for degree of cross-fertilization in faba bean. Crop Sci. 34, 960–964.
Link, W., Stelling, D., Ebmeyer, E., 1994b. Factors determining the performance ofsynthetics in Vicia faba L. 1. Heterogeneity, heterozygosity, and degree of cross-fertilization. Euphytica 75, 77–84.
Link, W., Schill, B., von Kittlitz, E., 1996. Breeding for wide adaptation in faba bean.Euphytica 92, 185–190.
Lloyd, D.G., Schoen, D.J., 1992. Self-fertilization and cross-fertilization in plants. 1.Functional dimensions. Intl. J. Plant Sci. 153, 358–369.
Maria-Klein, A., Vaissiere, B.E., Cane, J.H., Steffan-Dewenter, I., Cunningham, S.A.,Kremen, C., Tscharntke, T., 2007. Importance of pollinators in changing land-scapes for world crops. Proc. Roy. Soc. B 274, 303–313.
Maalouf, F.S., Suso, M.J., Moreno, M.T., 1999. Choice of methods and indices foridentifying the best parentals for synthetic varieties in faba bean. Agronomie19, 705–712.
Maalouf, F., Ahmed, K.S., Munzir, K., Khalil, S., 2008. The effect of mating system fordeveloping combined resistance to chocolate spot and Ascochyta blight in fababean. In: Prohens, J., Badenes, M.L. (Eds.), Modern Variety Breeding for Presentand Future Needs, Proceedings of the 18th Eucarpia General Congress, Uni-versidad Politecnica de Valencia, Valencia, Spain, p. 16.
McLellan, T., Endler, J.A., 1998. The relative success of some methods for measuringand describing the shape of complex objects. Syst. Biol. 47, 264–281.
Metz, P.L.J., Buiel, A.A.M., Van Norel, A., Helsper, J.P.F.G., 1993. Rate and inheritanceof cross-fertilization in faba bean (Vicia faba). Euphytica 66, 127–133.
Metz, P.L.J., Buiel, A.A.M., Van Norel, A., Helsper, J.P.F.G., 1994. Genetic factorscontrolling outcrossing in faba bean (Vicia faba): effects of pollen donor andreceptor genotypes and estimates of inter-receptor cross-fertilization. J. Agric.Sci. 122, 249–254.
Morandin, L.A., Winston, M.L., 2006. Pollinators provide economic incentiveto preserve natural land in agroecosystems. Agric. Ecosyst. Environ. 116,289–292.
Motten, A.F., Stone, J.L., 2000. Heritability of stigma position and the effect ofstigma-anther separation on outcrossing in a predominantly self-fertilizingweed, Datura stramonium (Solanaceae). Am. J. Bot. 87, 339–347.

M.J. Suso, F. Maalouf / Field Crops Research 116 (2010) 116–126126
Ollerton, J., Dafni, A., 2005. Functional floral morphology and phenology. In: Dafni,A., Kevan, P.G., Husband, B.C. (Eds.), Practical Pollination Biology. Enviroquest,Cambridge, Ontario, Canada, pp. 1–26.
Osborne, J.L., Awmack, C.S., Clark, S.J., Williams, I.H., Mills, V.C., 1997. Nectar andflower production in Vicia faba L (field bean) at ambient and elevated carbondioxide. Apidologie 28, 43–55.
Palmer, R.G., Perez, P.T., Ortiz-Perez, E., Maalouf, F., Suso, M.J., 2009. The role of crop-pollinator relationships in breeding for pollinator-friendly legumes; from abreeding perspective. Euphytica 170, 35–52.
Parker, P.G., Snow, A.A., Schug, M.D., Booton, G.C., Fuerst, P.A., 1998. What moleculescan tell us about populations: Choosing and using a molecular marker. Ecology79, 361–382.
Parzies, H.K., Nke, C.F., Abdel-Ghani, A.H., Geiger, H.H., 2008. Outcrossing rate ofbarley genotypes with different floral characteristics in drought-stressed envir-onments in Jordan. Plant Breed. 127, 536–538.
Pierre, J., Le Guen, J., Pham Delegue, M.H., Tanguy, X., Renard, M., 1996. Comparativestudy of nectar secretion and attractivity to bees of two lines of spring-type Fababean (Vicia faba L. var equina Steudel). Apidologie 27, 65–75.
Pierre, J., Suso, M.J., Moreno, M.T., Esnault, R., Le Guen, J., 1999. Diversite et efficacitede l’entomofaune pollinisatrice (Hymenoptera: Apidae) de la feverole (Viciafaba L.) sur deux sites, en France et en Espagne. Ann. Soc. Entomol. Fr. 35(Suppl.), 312–318.
Prusinkiewicz, P., Erasmus, Y., Lane, B., Harder, L.D., Coen, E., 2007. Evolution anddevelopment of inflorescence architectures. Science 316, 1452–1456.
Ritland, K., 1990. A series of FORTRAN computer programs for estimating plantmating systems. J. Hered. 81, 235–237.
Ritland, K., 2002. Extensions of models for the estimation of mating systems using nindependent loci. Heredity 88, 221–228.
Shaw, D.V., Allard, R.W., 1982. Estimation of outcrossing rates in douglas-fir usingisozyme markers. Theor. Appl. Genet. 62, 113–120.
Schoen, D.J., Busch, J.W., 2008. On the evolution of self-fertilization in a metapo-pulation. Int. J. Plant Sci. 169, 119–127.
Suso, M.J., Moreno, M.T., Cubero, J.I., 1993. New isozyme markers in Vicia faba:inheritance and linkage. Plant Breed. 111, 170–172.
Suso, M.J., Moreno, M.T., 1999. Variation in outcrossing rate and genetic structureon six cultivars of Vicia faba L. as affected by geographic location and year. PlantBreed. 118, 347–350.
Suso, M.J., Pierre, J., Moreno, M.T., Esnault, R., Le Guen, J., 2001. Variation inoutcrossing levels in faba bean cultivars: role of ecological factors. J. Agric.Sci. Camb. 136, 399–405.
Suso, M.J., Harder, L.D., Moreno, M.T., Maalouf, F., 2005. New strategies for increas-ing heterozygosity in crops: Vicia faba mating system as a study case. Euphytica143, 51–65.
Suso, M.J., Hunady, I., Solis, I., Mondragao-Rodrigues, F., Winkler, J., 2008. Germ-plasm management of Vicia faba L.: comparative study of the mating system oflocal and common cultivars growing under different agro-ecological conditions.Plant Genet. Res. Newsl. 155, 46–51.
Terzopoulos, P.J., Kaltsikes, P.J., Bebeli, P.J., 2008. Determining the sources ofheterogeneity in Greek faba bean local populations. Field Crops Res. 105,124–130.
Virmani, S.S., 1996. Hybrid rice. Adv. Agron. 57, 377–462.





























