Embed Size (px)
Citation preview

E L S E V I E R Molecular and Biochemical Parasitology 65 (1994) 213-224
MOLECULAR AND BIOCHEMICAL PARASITOIZ)GY
Detection of ubiquinone in parasitic and free-living protozoa, including species devoid of mitochondria
Jayne E. Ellis a, Kenneth D.R. Setchell b, Edna S. Kaneshiro a'* aDepartment of Biological Sciences (ML 006), University of Cincinnati, Cincinnati, OH 45221-0006, USA; bClinical Mass Spectrometry,
Department of Pediatrics, Children's Hospital Medical Center, Cincinnati, OH 45229, USA
Received 11 January 1994; accepted 19 February 1994
Abstract
Ubiquinone (coenzyme Q, CoQ) was analyzed and individual homologues quantified in 11 species of parasitic and free- living protozoa by a combination of thin-layer chromatography and high performance liquid chromatography. Fast atom bombardment ionization-mass spectrometry was used for the first time to confirm the identity of the fractionated CoQ homologues and proved to be a fast, gentle and convenient method for ubiquinone identification. Ubiquinone was detected in all organisms including those devoid of identifiable mitochondria. However, significantly lower levels of CoQ were present in those organisms lacking this respiratory organelle (5- to 50-fold lower in Entamoeba histolytica (CoQ9) and 15- to 350-fold for Giardia lamblia (CoQ9) and Tritrichomonas foetus (CoQi0)). Coenzyme Q9 was the predominant homologue in promastigotes of Leishmania donovani and Leishmania major. Lower amounts of CoQ8 and CoQ10 were also detected in L. donovani, and CoQ8 in L. major. Comparison of the in vitro cultivated promastigote and amastigote forms of Leishmania pifanoi and Leishmania amazonensis revealed CoQ9 to be the sole detectable ubiquinone homologue in the amastigote (macrophage) stage, whereas CoQs and CoQl0 were also present in the promastigotes (life cycle stage found in the insect gut) of L. pifanoi, and CoQ7 and CoQ8 in promastigotes of L. amazonensis. Interestingly, the total amounts of CoQ were similar in both forms of these organisms. The free-living ciliates, Tetrahymena thermophila and Paramecium tetraurelia contained CoQs as the predominant ubiquinone species and this homologue was also present in the isolated cilia from P. tetraurelia. The marine ciliate, Parauronema acutum contained CoQ7 as well as CoQ8. Comparison of xenosome-containing P. acutum with organisms lacking the symbiont revealed that twice the level of CoQ8 was present in cells grown with this cytoplasmic gram-negative bacterium. Results suggest that CoQ is ubiquitous amongst the protozoa, regardless of the presence of mitochondria, and may function in alternative roles to that of mitochondrial electron transport chain component.
Key words: Parasitic protozoon; Free-living protozoon; Cilia; Fast atom bombardment-ionisation mass spectrometry; High-performance liquid chromatography; Coenzyme Q; Endosymbiont
*Corresponding author. Tel.: (513) 556 9712; Fax: (513) 556 5280.
Abbreviations: HPLC, high performance liquid chromatogra- phy; TLC, thin layer chromatography; FAB-MS, fast atom bombardment-ionization mass spectrometry; CoQ, coenzyme Q.
0166-6851/94/$7.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0 1 6 6 - 6 8 5 1 ( 9 4 ) 0 0 0 4 6 - P
I. Introduction
Coenzyme Q (CoQ, ubiquinone) is widely dis- tributed in biological systems and carries out the

214 J.E. Ellis et al./Molecular and Biochemical Parasitology 65 (1994) 213-224
well characterized role as a redox component in- volved in the transfer of electrons and protons in mitochondrial and bacterial electron transport chains. Ubiquinone, however, is not restricted in mammalian cells to the mitochondria, and subcel- lular studies reveal that this lipophilic molecule accumulates in all endomembranes, especially con- centrated in the Golgi and lysosomal membranes [1]. The essential role of CoQ in biological energy transduction is well established. In addition to this role, however, ubiquinone has been implicated in a wide range of cellular functions including: influen- cing the fluidity and stability of membranes [2], non-mitochondrial redox pathways e.g. the Golgi apparatus and plasma membrane [3-5], roles in growth control and secretion-related membrane flow [6], control of cellular internal pH and cal- cium exchange [6] and membrane antioxidant in the reduced (ubiquinol) form [7].
The CoQ series of compounds consists of monounsaturated 5 carbon (isoprene) units which are attached covalently to an aromatic nucleus (2,3-dimethoxy-5-methyl- 1,4-benzoquinone). The polyprenyl side chain of CoQ varies from 6 to 12 isoprene units in different cell types and is often characteristic of a particular species. Higher ani- mals usually contain CoQ10 as the sole endogen- ous quinone [8]. The length of the polyprenyl chain has been used as a taxonomic marker for yeast [9] and for bacteria, where gram-negative organisms contain CoQ (and in some cases mena- quinone (MK)), and gram-positive organisms con- tain MK only [10]. The development of a predo- minant CoQ homolog which is specific for parti- cular species is not clear, but may be related to the specifically poised fluidity of the membrane in which it is embedded [11] or the specificity of the CoQ side chain interaction with integral mem- brane proteins [8].
Although homologues of CoQ represent major cellular components with functions intrinsic to the metabolism of the cell, the role and specificities of this molecule have been largely unstudied in the protozoa. The type of ubiquinone present in pro- tozoan species has been addressed in a few organ- isms [12,13], including the parasitic Plasmodium spp. (CoQs) [14] and Trypanosoma brucei (CoQ9) [15,16]. This is different from the host ubiquinone
in which the polyprenyl side chain consists of 10 isoprene units (CoQl0) [8]. The transfer of elec- trons by CoQ between electron transport chain components during respiratory processes repre- sents an important chemotherapeutic target [14,17,18], and the specificity of inhibitors for the parasite respiratory chain has been attributed to differences in the Q-binding sites of the integral membrane proteins in parasite and host respira- tory chains [19].
In this study we identified and quantified CoQ homologues from 11 species of protozoa and car- ried out a comparison of parasitic and free-living organisms. We present evidence that CoQ is com- mon amongst eukaryotic cells, including organ- isms known to lack mitochondria, and may func- tion in alternate roles to that of mitochondrial electron transport chain component.
2. Materials and methods
Organisms. The following organisms were ana- lyzed in this study: Leishmania donovani (promas- tigotes), Leishmania major (promastigotes), Leishmania pifanoi (amastigotes and promasti- gotes), Leishmania amazonensis (amastigotes and promastiogotes), Entamoeba histolytica (tropho- zoites), Giardia lamblia (trophozoites), Tritricho- monas foetus, Tetrahymena thermophila, Paramecium tetraurelia, Paramecium octaurelia, Parauronema acutum (with or without xeno- somes), Candida utilis and Escherichia coli. In ad- dition, the isolated cilia from P. tetraurelia were also examined for the presence of CoQ. All of the organisms (except for C. utilis) were cultured to late exponential phase of growth. Details of cul- ture media and washing solutions used for each species are given in Table 1. Solutions used for washing harvested pellets were as follows: P. tetra- urelia, P. octaurelia, T. thermophila, double-dis- tilled water; P. acutum, artificial sea water [20]; L. amazonensis, E. histolytica, T. foetus and G. lam- blia, phosphate buffered saline (200 mM, pH 7.4). All other harvested organisms were used as un- washed pellets. C. utilis was obtained freeze-dried from Sigma Chemical Co. (St. Louis, MO). Cilia were isolated from P. tetraurelia (51s) as pre-

,I.E. Ellis et al./Molecular and Biochemical Parasitology 65 (1994) 213-224 215
viously described [21]. The medium used for the growth of G. lamblia was bile-free. Yeast extract was from Difco Laboratories (Detriot, MI). Fetal bovine serum was obtained from Gibco Labora- tories (Grand Island, NY). Cells provided by other investigators were shipped frozen with dry ice, or lyophilized (P. tetraurelia cilia) and imme- diately extracted upon receipt.
1.0 ml min-1 using a Beckman 100A solvent deliv- ery system with a 421 system controller. Authentic standards (10 /~g m1-1) and samples were dis- solved in chloroform/methanol (1:2, v/v). Detec- tion of CoQ was carried out at Abs290 nm using a Hitachi variable wavelength UV detector (Hitachi, Tokyo, Japan). Data was plotted on a 3390A inte- grator (Hewlett Packard, Avondale, PA).
Lipid extraction and isolation of ubiquinone. Due to the labile nature of CoQ, precautions to prevent degradation were taken throughout the procedure to minimize degradation (e.g. inclusion of the anti- oxidant butylated hydroxytoluene (BHT) in all solvents, light exclusion from the samples, and storage under a N2 atmosphere). Total lipids were extracted from whole cells by the method of Bligh and Dyer [22] and purified by the method of Folch et al. [23]. Silicic acid adsorption column chroma- tography (Unisil, Clarkson Chemical, William- sport, PA) was used to separate total lipids into neutral (eluted with 50 ml chloroform) and polar (eluted with 50 ml methanol) lipid fractions [24]. The neutral lipid fraction was separated into dif- ferent classes by thin-layer chromatography (TLC) on glass-backed 500-#m thick Silica Gel G plates (Analtech, Newark, DE) employing the sol- vent system, petroleum ether/diethyl ether/acetic acid (80:20:1, v/v/v). Plates were visualized by staining with iodine vapor. Bands with migrations (RE) corresponding to those of authentic stan- dards of CoQ6 , COQT, CoQ9 and CoQ10 (Sigma) and CoQ8 purified from E. coli were scraped off the plate and eluted from the silica gel with chloroform/methanol (1:2, v/v). The isolated qui- none material was concentrated under a stream of N2 gas, redissolved in 200 #1 of chloroform/metha- nol (1:2, v/v) and subjected to high performance liquid chromatography (HPLC) analysis.
HPLC analysis of ubiquinone. Homologues of CoQ in TLC-purified material were separated by reverse phase HPLC by the method previously de- scribed [15] using an Ultrasphere ODS C18 column (5/~m particle size) (Beckman Instruments, Berke- ley, CA). Samples (20 #1) were injected onto the column and subjected to isocratic elution with hexane/methanol (1:9, v/v) at a flow rate of 0.8 or
Mass spectrometry. Fast atom bombardment io- nization-mass spectrometry (FAB-MS) was car- ried out on pooled fractions of CoQ collected after HPLC purification of samples. Positive and negative ion FAB-MS spectra were obtained after placing the equivalent of 10-50 ng of each com- pound dissolved in methanol onto a small drop of 3-nitrobenzyl alcohol spotted onto a copper target of an insertion probe. The probe was in- serted into the ion source of a VG Autospec Q mass spectrometer (VG Gas Analysis, Cheshire, UK) and the sample bombarded with a beam of cesium atoms (35 keV). Low resolution (1000) mass spectra were continually recorded (2 s scans) over the mass range m/z 150-1000.
Protein and dry weight determination. Samples were analyzed for protein by the method of Low- ry [25] using bovine serum albumin (BSA) as stan- dard. Dry weights were measured after drying samples under a continuous N2 stream at 30°C.
3. Results
Identification of ubiquinone homologues. Evi- dence for the identity of CoQ homologues in sam- ples of protozoan neutral lipid fractions was determined by cochromatography with authentic standards by two chromatographic procedures: TLC and HPLC. Where large amounts of iso- lated CoQ homologues could be collected by elu- tion from HPLC, the compound was analyzed by FAB-MS for molecular weight determination.
The homologues of CoQ separated by TLC had RE values in the range of 0.35-0.41, for chain lengths from 6 to 10 isoprene units, respectively. Resolution of individual CoQ homologues was in- sufficient in itself to identify and quantify each

216 J.E. Ellis et al./Molecular and Biochemical Parasitology 65 (1994) 213-224
E
o a
O~ 0 E
0 ffi
.~_ N
A Q6
lli' 20 30 40 50 60 70 0 10
B
C 16 26 3~ 4d sd 8d 7d
\
0.001
0 10 20 30 40 50 60 70
Time (min)
Fig. 1. Separation, detection and quantitation of coenzyme Q by HPLC. (A) Chromatogram of authentic s tandards of COQ6,7,9,10 (200 ng) and a trace of CoQs, separated by HPLC and detected by absorbance at 290 nm. (B) CoQ homologues resolved by HPLC of material eluted from the CoQ TLC band of L, donovani neutral lipids. (C) CoQs separated by HPLC of the CoQ TLC band of the neutral lipids from P. octaurelia.
individual component by this method, therefore the whole CoQ band was eluted for each organ- ism under study and subjected to HPLC. Separa- tion of individual ubiquinones was achieved by reverse phase HPLC. A mixture of authentic stan- dards of CoQ separated by HPLC resulted in clear baseline separations with approximate retention times of 12, 17, 26, 38 and 58 min for C o Q 6 ,
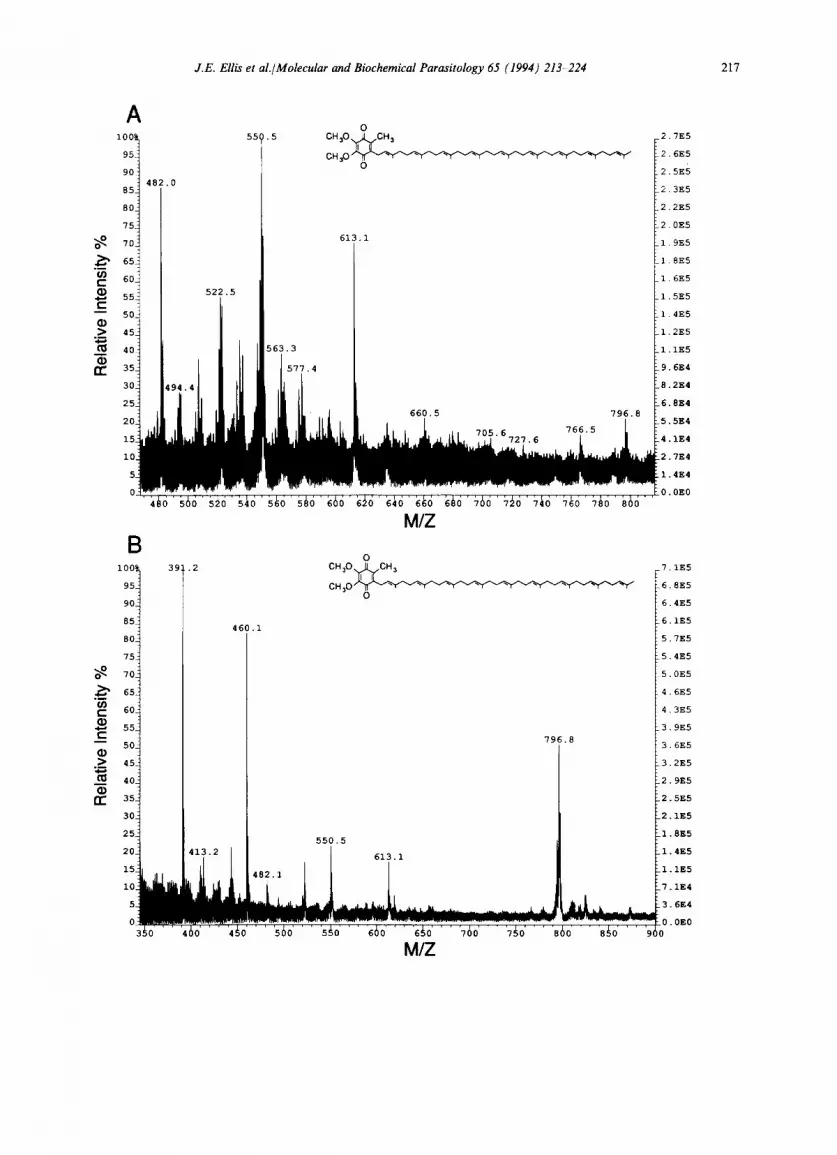
CoQ7, CoQs, CoQ9 and CoQ10, respectively (Fig. 1A). Identification of protozoan components was performed by comparison of retention times and cochromatography by spiking of samples with authentic standards. Quantification of CoQ (Abs290 nm) was obtained by measurement of HPLC peak areas compared with known amounts of authentic standard. This was com- pared for each organism and calculated on the basis of dry weight and protein. The limit of detec- tion for this technique was found to be ca. 2 ng. HPLC separation of CoQ homologues in neutral lipids of L. donovani and P. octaurelia are pre- sented (Fig. 1 B, C). Collection of HPLC fractions and subsequent FAB-MS analysis confirmed the identity of CoQ9 isolated from L. donovani and C. utilis (Fig. 2A, B). The positive ion FAB-MS spectra of these compounds revealed intense [M +H] ÷ ions with negligible fragmentation of the parent ion. Other ions in the spectra are de- rived from the matrix. Under negative ion record- ing no significant ions were found. The limit of detection for positive ion FAB-MS of CoQ was found to be in the order of 2 ng.
Leishmania spp. The predominant homologue of L. donovani promastigotes w a s C o Q 9 (Table 1). Lower amounts of CoQ8 and CoQ10 were also detectable in this organism. Similarly, promasti- gotes of L. major contained C o Q 9 as the major homologue, with minor contribution from CoQ8. The total CoQ concentration in L. donovani pro- mastigotes, and promastigote and amastigote
Fig. 2. Fast atom bombardment ionization mass spectra (35 keV) of CoQ9 purified by HPLC of neutral lipids from parasitic protists. In the upper panel (A) CoQ9 (3 ng) was isolated by TLC and purified by HPLC from L. donovani neutral lipids. The pseudo-molecular ion [M + H] + for CoQ9 was present at m/z of 796.8 and corresponds to the molecular weight for CoQ9. In the lower panel (B) CoQ9 (45 ng) was purified by HPLC from C. utilis neutral lipids. Ions detected at m/z 460.1 and 613.1 correspond to adducts from the matrix solution, 3-nitrobenzyl alcohol.
Rela
tive
Inte
nsity
%
I,
~o
~o
o~o -r
~1,~,
, .....
,,,,,,,,
,,,,,,,,
,,,, ..
....
....
,i,
, ...
......
......
......
..
N
Rela
tive
Inte
nsity
%
co
o~
==
~a
L~
"o
=o
T
r~
t~
218 J.E. Ellis et al./Molecular and Biochemical Parasitology 65 (1994) 213-224
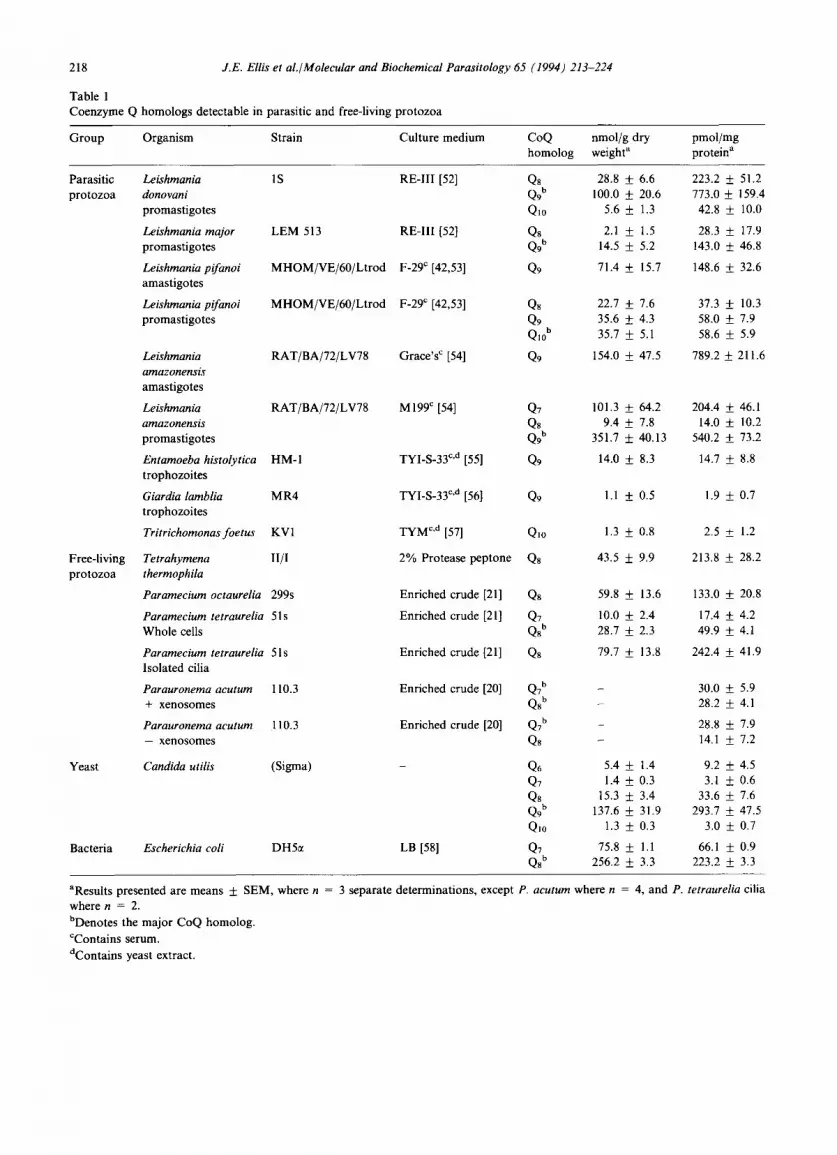
Table 1 Coenzyme Q homologs detectable in parasitic and free-living protozoa
Group Organism Strain Culture medium CoQ nmol/g dry pmol/mg homolog weight a protein a
Parasitic Leishmania 1S RE-Ill [52] Q8 28.8 + 6.6 223.2 protozoa donovani Q9 b 100.0 _+ 20.6 773.0
promastigotes Qlo 5.6 + 1.3 42.8
Leishmania major LEM 513 RE-III [52] Q8 2.1 + 1.5 28.3 promastigotes Q9 b 14.5 + 5.2 143.0
Leishmania pifanoi MHOM/VE/60/Ltrod F-29 c [42,53] Q9 71.4 + 15.7 148.6 amastigotes
Leishmaniapifanoi MHOM/VE/60/Ltrod F-29 ¢ [42,53] Q8 22.7 + 7.6 37.3 + 10.3 promastigotes Q9 35.6 + 4.3 58.0 + 7.9
Q10 b 35.7 + 5.1 58.6 + 5.9
Leishmania RAT/BA/72/LV78 Grace's ~ [54] Q9 154.0 + 47.5 789.2 + 211.6 amazonensis amastigotes
Leishmania RAT/BA/72/LV78 M199 ¢ [54] Q7 101.3 ___ 64.2 204.4 _+ 46.1 amazonensis Qs 9.4 + 7.8 14.0 _+ 10.2 promastigotes Q9 b 351.7 ___ 40.13 540.2 + 73.2
Entamoeba histolytica HM-I TYI-S-33 c'd [55] Q9 14.0 ___ 8.3 14.7 + 8.8 trophozoites
Giardia lamblia MR4 TYI-S-33 c'd [56] Q9 1.1 + 0.5 1.9 _+ 0.7 trophozoites
Tritrichomonasfoetus KV1 TYM c'd [57] Qlo 1.3 + 0.8 2.5 + 1.2
Tetrahymena II/I 2% Protease peptone Qs 43.5 __+ 9.9 213.8 + 28.2 thermophila
Paramecium octaurelia 299s Enriched crude [21] Qs 59.8 _+ 13.6 133.0 _+ 20.8
Paramecium tetraurelia 51s Enriched crude [21] Q7 10.0 _+ 2.4 17.4 + 4.2 Whole cells Qs b 28.7 + 2.3 49.9 + 4.1
Paramecium tetraurelia 51s Enriched crude [21] Qs 79.7 _+ 13.8 242.4 _+ 41.9 Isolated cilia
Parauronema acutum 110.3 Enriched crude [20] Q7 b - 30.0 ± 5.9 + xenosomes Q8 b - 28.2 + 4.1
Parauronema acutum 110.3 Enriched crude [20] Q7 b - 28.8 + 7.9 - xenosomes Q8 - 14.1 + 7.2
Candida utilis (Sigma) - Q6 5.4 + 1.4 9.2 + 4.5 Q7 1.4 + 0.3 3.1 ___ 0.6 Qs 15.3 +_ 3.4 33.6 + 7.6 Q9 b 137.6 + 31.9 293.7 + 47.5 Q10 1.3 + 0.3 3.0 + 0.7
Escherichia coli DH5ct LB [58] Q7 75.8 ___ 1.1 66.1 + 0.9 Q8 t' 256.2 ___ 3.3 223.2 __+ 3.3
Free-living protozoa
Yeast
Bacteria
___ 51.2 + 159.4 + 10.0
_+ 17.9 + 46.8
+ 32.6
aResults presented are means + SEM, where n = 3 separate determinations, except P. acutum where n = 4, and P. tetraurelia cilia where n = 2.
bDenotes the major CoQ homolog.
CContains serum.
dContains yeast extract.

J.E. Ellis et al./Molecular and Biochemical Parasitology 65 (1994) 213-224 219
forms of L. amazonensis, were substantially higher than any other organism analyzed in this study, even higher than that for aerobically grown yeast. Unlike the promastigotes of L. donovani and L. major, the promastigotes of L. pifanoi contained equivalent concentrations of CoQ9 and CoQlo and substantial amounts of CoQ8. Coenzyme Q7 and CoQ8 were detected along with CoQ9 in the promastigotes of L. amazonensis. Comparison with in vitro cultured amastigote s of L. pifanoi and L. amazonensis revealed that CoQ9 was the sole homologue in this life cycle form in both spe- cies. The total amount of CoQ was comparable in both life cycle stages.
'Anaerobic' protozoa. The protozoa which lack mitochondria were found to contain detectable CoQ, but the levels present were much lower than those found in the aerobic protozoa. The amount of CoQ was 5- to 50-fold lower in E. histolytica (CoQ9) and 15- to 350-fold lower for G. lambria (CoQ9) and T. foetus (COQlo) compared to the organisms that utilize mitochondrial respiration (Table 1).
Free-riving ciliates. Coenzyme Q8 was the predo- minant homologue among the free-living ciliates. Coenzyme Q7 was also detectable in some organ- isms (Table 1). Of the two species of Paramecium studied here, CoQ8 was the sole detectable homo- logue in P. octaurelia (Fig. 1C). However, in addi- tion to CoQs, CoQ7 was also present in P. tetraureria revealing differences between these two species grown under identical culture conditions. The isolated cilia from P. tetraureria (51s) con- tained significant amounts of CoQ8, the major ubiquinone homologue from this organism. CoQ7 which was present in whole cells was not detected in the cilia. Whole cells of P. acutum were ana- lyzed both with and without endosymbiotic bac- teria termed xenosomes. In the presence of the symbiont, CoQ8 was present at levels twice those found in organisms grown in its absence.
Ubiquinone in yeast and E. coli. The CoQ homo- logues present in C. utilis and E. cori were studied in order to compare the data obtained by this study with that from previous work. The presence
of CoQ9 and CoQ8 as the respective predominant CoQ of these organisms, confirmed previous stu- dies and the amounts detected were also compar- able to values in the literature [8].
Analyses of growth media. To examine whether CoQ present in the organisms might originate in the media used for their cultivation, growth med- ia were analyzed. CoQ was not detected in: 2% protease peptone (T. thermophila); a modified en- riched crude medium (Paramecium spp.) [20]; and a modified RE-III lacking BSA (L. donovani and L. major) [22]. However, CoQ8 was detected at low levels in the medium used for P. acutum culti- vation (0.2 nmol g-1 dry weight; 0.65 pmol mg-1 protein) [21], which may account for some of the CoQ8 in P. acutum, but it is unlikely that the ma- jor component, CoQ7 was simply scavenged by the ciliate. Analysis of growth media used to culture the 'anaerobic' protozoa failed to detect CoQ, therefore direct analysis of the media components was performed. Surprisingly, ubiquinone was not detectable in yeast extract (30 g extract = 30 1 of growth medium). Thus, CoQ is probably lost or destroyed during the extraction procedure applied for the commercial preparation of yeast extract from our supplier. Fetal bovine serum (100 ml = 1 1 growth medium), by contrast, contained detect- able CoQ9 (0.1 nmol g - i dry weight; 0.3 pmol mg -1 protein) and CoQl0 (0.4 nmol g i dry weight; 0.8 pmol mg-1 protein).
4. Discussion
Data presented here is the first comprehensive study of the ubiquinones in protozoan species. The ubiquitous nature of CoQ in eukaryotic cells was confirmed as all of the organisms examined were found to possess this important cellular com- ponent, regardless of the presence or absence of identifiable mitochondria. A clear correlation ex- ists however, between the relative amounts of CoQ and the possession of this respiratory organelle as significantly lower levels of CoQ were present in organisms lacking mitochondria.
The central role of CoQ in the mitochondrial metabolism of protozoa is well established

220 J.E. Ellis et al./Molecular and Biochemical Parasitology 65 (1994) 213-224
[15,26,27]. However, details concerning the precise mechanism of electron transport in these organ- isms is largely based on studies with mammalian, bacterial and yeast systems. A large endogenous CoQ pool is a prerequisite for bioenergetic activ- ity, thus a large proportion of cellular ubiquinone is probably associated with the inner mitochon- drial membrane. In mammalian cells it is now well established that CoQ is present at a multiple sites within the cell and carries out a variety of cellular functions. Significant ubiquinone levels are associated with the Golgi, lysosomal and plas- ma membranes [1]. This is particularly interesting in Leishmania promastigotes, where CoQ levels three times higher than those found in aerobically grown yeast were detected. These organisms in- completely oxidize substrates to organic acids and CO2, and rely on glycolysis rather than respiration for their major source of ATP [28]. The subcellular location of CoQ in protozoa has not been specifi- cally addressed and the question of other cellular locations and functions of this molecule remains intriguing. The detection of ubiquinone in the ci- lia of P. tetraurelia represents the first non-mito- chondrial location for CoQ in a protozoan cell [29], and the first detection of ubiquinone in this organelle. Ubiquinone was significantly enriched in this organelle by comparison with whole cells. Possible role(s) for CoQ in ciliary membranes in- clude, participation in redox pathways, membrane antioxidant or a role in membrane fluidity.
Cell surface membrane electron transport has been characterized in Tetrahymena employing transmembrane ferricyanide reduction [30]. In this study it was observed that ciliary beat fre- quency was stimulated by ferricyanide and that the ferricyanide reduction was especially sensitive to inhibition by chloroquine [30]. Chloroquine is highly toxic to Tetrahymena [30] and Paramecium (Joachim Schultz, personal communication) and thus its site of action may include the ciliary mem- brane, which constitutes about half the area of the cell surface membrane of these holotrichous cili- ates.
Studies on the effects of respiratory inhibitors on E. histolytica [31,32] and G. lamblia [33] have suggested that CoQ may be a component of a transport chain in which O2 can serve as an elec-
tron acceptor. The transport chains of these or- ganisms, however, are unlike those present in eu- karyotes which contain mitochondria, in that cy- tochromes are absent and FeS proteins and flavin are predominant components [31,34]. However, the presence of relatively high concentrations of ubiquinone is a requirement for bioenergetic activ- ity and even a partial extraction of ubiquinone from the mitochondrial membrane results in an immediate decrease in oxidase activity [35]. There- fore, the possible role(s) that CoQ may play at such low levels is intriguing. Although the subcel- lular location of CoQ in these organisms is not known, it is possible that CoQ may be located in the plasma membrane or endoplasmic reticulum and have a role similar to that proposed for CoQ in purified plasma membranes from human and rat cells [4]. Indeed, it has been proposed that plas- ma membrane electron transport may play an es- sential role as an oxidase in these organisms [30]. A functional role for CoQ in this and other ami- tochondrial eukaryotes awaits elucidation.
Coenzyme Q9 predominates in the ubiquinones of Leishmania spp. This finding is in agreement with previous studies carried out with L. donova- hi, L. tropica and L. tarentolae (Rangnathan, G. (1984), PhD thesis, University of Cincinnati). Data presented here is the first indication that polyprenyl side chain length specificity exists in different life cycle forms of an organism (L. pifa- noi and L. amazonensis). Differences in the re- spiratory metabolism of promastigotes and amas- tigotes have been implicated both from ultrastruc- tural studies where the promastigote (insect) stage was found to contain well developed mitochondria by comparison with the amastigote (macrophage) stage where they become degenerate and the amas- tigotes proposed to be anaerobic [36]. Those stu- dies were carried out with in vitro cultured pro- mastigotes and amastigotes isolated from fresh tis- sue. Respiratory studies with both life cycle forms of L. mexicana showed that metabolic differences existed: amastigotes preferred to utilize fatty acids, whereas promastigotes preferentially respired su- gars [37]. More recently, comparative ultrastruc- tural studies with in vitro cultured amastigotes as well as promastigotes of L. pifanoi indicated simi- lar mitochondrial elements are present in both

J.E. Ellis et al./Molecular and Biochemical Parasitology 65 (1994) 21~224 221
forms [38]. The high levels of ubiquinone detected in both life cycle stages in this study is consistent with those ultrastructural data. The precise roles and importance of mitochondrial metabolism in cultured vs naturally occurring leishmanias re- mains unclear. Significant differences exist be- tween promastigotes and amastigotes of Leishma- nia spp. with respect to ubiquinone homologue content which may reflect differences in the meta- bolism of these life cycle stages. For example, CoQ homologues play different roles in cellular meta- bolism in rabbit liver cell preparations. In this sys- tem, CoQ9 accumulates in the cytosol, and may function as an antioxidant. CoQi0 however, was localized in the mitochondria, and functions in the respiratory chain [39]. It is' interesting to spec- ulate that CoQ may have antioxidant properties in this and other protozoa, as toxicity to 02 or pro- ducts of its reduction (superoxide, peroxide, hy- droxyl radical, etc.) are manifest, most notably in the 'anaerobic' protozoa and in Leishmania spp. [40,41]. However, conditions for axenic growth of amastigotes and promastigotes differ in pH and temperature [42] and therefore it cannot be ruled out that changes in membrane fluidity at different growth temperatures could bring about differences in ubiquinone homologues.
Elevated levels of CoQ8 in the xenosome-bear- ing ciliate P. acutum supports previous studies suggesting that the presence of the endosymbiotic bacterium stimulated respiratory rate and host glycogen metabolism [43]. However, whether the increased amount of CoQ is due to the symbiont itself or the increased metabolic activity of the host is not clear. The xenosome cannot be cul- tured outside the protozoan host but it is known that the genome of this gram-negative bacterium is about one-tenth that of E. coli. Immunological studies suggest that 30% of the symbiont protein molecules are of host origin [44]. It would be inter- esting to determine if xenosomal CoQ is also of host origin.
The biosynthesis of ubiquinone by protozoa is poorly understood. The lack of detectable CoQ in the media used for the axenic cultivation of L. donovani, L. major and the free-living ciliates sug- gests that these organisms can synthesize ubiqui- none in order to meet their nutritional require-
ments during in vitro growth. The polyprenyl side chain of CoQ is probably synthesized by the me- valonic acid pathway from which certain protozoa (e.g. Leishmania, Trypanosoma, Tetrahymena) also synthesize sterols [16,45,46]. The origin of the ben- zoquinone ring, however, is less clear. Tetrahyme- na possess a functional shikimic acid pathway [47], which is common among the bacteria, fungi and plants. This pathway is absent in mammalian cells that utilize aromatic amino acids (e.g. pheny- lalanine or tyrosine) for CoQ ring biosynthesis. The presence of the shikimic acid pathway in parasitic protozoa awaits investigation, but may represent an important chemotherapeutic target. Lipid biosynthetic deficiencies have been pro- posed for the anaerobic protozoa [48-50]. These organisms utilize preformed lipids to meet their nutritional requirements. Accumulation of CoQ during scavenging of lipids from the culture med- ia may account for the presence of CoQ in these parasites as serum was found to contain both CoQ9 and CoQ10. However, some species specifi- city was evident in that only CoQ9 was detectable in G. lamblia and E. histolytica and only CoQl0 was detected in T. foetus. This indicates that non- discriminatory uptake of CoQ from the medium into these organisms grown with fetal bovine ser- um probably does not explain the presence of CoQ in the organism.
Quinones have been used increasingly as a taxonomic tool for bacteria and yeast. Data ob- tained here and from previous investigations indi- cates that there may be a correlation between pro- tozoan phyla and the predominant CoQ detected in the organism; thus CoQ8 was common to the free-living ciliates and CoQ9 predominated in the parasitic flagellates, with the exception of T. foe- tus. By contrast, the sporozoan malarial parasite, Plasmodium, contains CoQ8 as the predominant homologue of ubiquinone [14]. The data regard- ing other protozoan phyla are lacking at this time. These observation have implications in the use of CoQ homologues as taxonomic criteria in protozoa. A variety of quinones, other than CoQ, which differ by ring and side chain modifications can be found to occur not only in bacterial species (e.g. menaquinone [10]), but also in some eukaryo- tic organisms such as the parasitic helminths (e.g.

222 J.E. Ellis et al./Molecular and Biochemical Parasitology 65 (1994) 213-224
rhodoqu inone [51]). The part icipat ion o f other quinones in electron t ranspor t or other processes in p r o t o z o a remains to be established. The identi- fication o f CoQ homologues in several p ro tozoan species, including those known to lack mi tochon- dria suggest that in addi t ion to the well described role in respiratory processes, C o Q m a y play alter- nate roles in p ro tozoan metabolism.
Ubiquinone-l ike inhibitors o f the parasite re- spiratory chain have been studied extensively in the past few years, part icularly as potential anti- malarials [17]. It has become apparent that cer- tain drugs, such as the hydroxynaph thoqu inones , exhibit specificity for the parasite electron trans- por t chain, due to differences in the quinone-bind- ing sites o f the bCl complex o f P. falciparum, com- pared with that o f the host [19]. Major differences exist between parasite and host respiratory chains in a number o f organisms, but finding inhibitors which are no t toxic to the host remains a difficult task which is hampered by the lack o f informat ion regarding detailed character izat ion o f the respira- tory chain o f many p ro tozoan pathogens. Im- proved unders tanding o f the roles o f C o Q in para- sitic species may enable identification o f novel drug targets in organisms which include some of the major pathogens o f man.
Acknowledgements
The authors would like to thank the following colleagues who contr ibuted organisms to this study: Dr. E.L. Jarroll, Dr. J.M. Wingfield, Dr. D. Beach, Dr. D.G. Lindmark , Dr. D. M c M a - hon-Prat t , Dr. L.S. Diamond , Dr. A.T. Soldo, Dr. K-P. Chang. Isolated cilia were kindly do- nated by Dr. J. Schulz and S. Klumpp. Thanks also to Mike Wyder for prepara t ion o f L. donova-
ni neutral lipids and for other technical assistance. We are grateful to Jinan Cai for technical support in the mass spectrometer studies and to Dr. Ha r ry Rudney for his encouragement and critical reading of the manuscript . This work was suppor ted by a grant f rom the Nat iona l Institutes o f Heal th (RO 1 A I 29316).
References
[1] Kal+n, A., Norling, B., Appelkvist, E.L. and Dallner, G. (1987) Ubiquinone synthesis in the microsomal fraction of rat liver. Biochim. Biophys. Acta 926, 70-80.
[2] Lenaz, G. (1985) A biochemical rationale for the therapeutic effects of coenzyme Q. In: Coenzyme Q. Biochemistry, Bioenergetics and Clinical Applications of Ubiquinone (Lenaz, G., ed.), pp. 435-440, Wiley Inter- science, Chichester.
[3] Sun, I.L., Sun, E.E., Crane, F.L. and Morrr, D.J. (1990) Evidence for Coenzyme Q function in transplasma membrane electron transport. Biochim. Biophys. Res. Commun. 172, 979-984.
[4] Sun, I.L., Sun, E.E., Crane, F.L., MorrO, D.J., Lindgren, A. and Lrw, H. (1992) Requirement for coenzyme Q in plasma membrane electron transport. Proc. Natl. Acad. Sci. USA 89, 11126-11130.
[5] Crane, F.L., Sun, I.L., Barr, R. and Morrr, D.J. (1984) Coenzyme Q in Golgi apparatus membrane redox activity and proton uptake. In: Biomedical and Clinical Aspects of Coenzyme Q (Folkers, K. and Yamamura, Y., eds.), Vol. 4, pp. 77-86, Elsevier, Amsterdam.
[6] Crane, F.L., Sun, I.L. and Sun, E.E. (1993) The essential functions of Coenzyme Q. Clin. Invest. 71, $55-$59.
[7] Ernster, L. and Forsmark-Andrre, P. (1993) Ubiquinol: an endogenous antioxidant in aerobic organisms. Clin. Invest. 71, $60-$65.
[8] Ramasarma, T. (1985) Natural occurrence and distribu- tion of coenzyme Q. In: Coenzyme Q. Biochemistry, Bioenergetics and Clinical Applications of Ubiquinone (Lenaz, G., ed.), pp. 67-81, Wiley Interscience, Chichester, UK.
[9] Yamada, Y., Sugihara, K., Van Eijw, G.W., Roeijmans, H.J. and De Hoog, G.S. (1989) Coenzyrne Q systems in ascomycetous black yeasts. Ant. Van Leeuwenhoek. 56, 349-356.
[10] Collins, M.D. and Jones, D. (1981) Distribution of isoprenoid structural types in bacteria and their taxo- nomic implications. Microbiol. Rev. 45, 316,354.
[11] Lenaz, G., Esposti, M.D., Fato, R. and Cabrini, L. (1981) Studies on coenzyme Q enzymes: role of the isoprenoid chain in the function of ubiquinone. In: Biomedical and Clinical Aspects of Coenzyme Q (Folkers, K. and Yamamura, Y., eds.), Vol. 3, pp. 169-182, Elsevier, Amsterdam.
[12] Vakirtzi-Lemonais, C., Kidder, G.W. and Dewey, V.C. (1963) Ubiquinone in four genera of protozoa. Comp. Biochem. Physiol. 8, 331-334.
[13] Long, B.H. and Coe, E.L. (1973) Characterization of ubiquinone from vegetative amoebae and mature soro- carps of the cellular slime mold, Dictyostelium discoideum. Comp. Biochem. Physiol. 45B, 933 943.
[14] Porter, T.H. and Folkers, K. (1974) Antimetabolites of Coenzyme Q. Their potential application as antimalarials. Angewandte Chemic 13, 559-569.
[15] Clarkson, A.B., Bienen, E.J., Pollakis, G. and Grady,

J.E. Ellis et al./Molecular and Biochemical Parasitology 65 (1994) 213~24 223
R.W. (1989) Respiration of bloodstream forms of the parasite Trypanosoma brucei brucei is dependent on a plant-like alternative oxidase. J. Biol. Chem. 264, 17770- 17776.
[16] L6w, P., Dallner, G., Mayor, S., Cohen, S., Chait, B.T. and Menon, A.K. (1991) The mevalonate pathway in the bloodstream form of Trypanosoma brucei. J. Biol. Chem. 266, 19250-19257.
[17] Hudson, A.T., Randall, A.W., Fry, M., Ginger, G.D., Hill, B., Latter, V.S., McHardy, N. and Williams, R.B. (1985) Novel anti-malarial hydroxynaphthoquinones with potent broad spectrum anti-protozoal activity. Parasitology 90, 45 55.
[18] Clarkson, A.B., Bienen, E.J., Pollakis, G. and Grady, R.W. (1989) Trypanocidal CoQ analogs: their effect on other mitochondrial systems. Comp. Biochem. Physiol. 94B, 245-251.
[19] Vaidya, A.B., Lashgari, M.S., Pologe, L.G. and Morrisey, J. (1993) Structural features of Plasmodium cytochrome b that may underlie susceptibility to 8-aminoquinolines and hydroxynaphthoquinones. Mol. Biochem. Parasitol. 58, 33-42.
[20] Soldo, A.T. and Merlin, E.J. (1972) The cultivation of symbiote-free marine ciliates in axenic medium. J. Protozool. 19, 519-524.
[21] Kaneshiro, E.S., Beischel, L.S., Merkel, S.J. and Rhoads, D.E. (1979) The fatty acid composition of Paramecium aurelia cells and cilia: changes with culture age. J. Protozool. 26, 147-158.
[22] Bligh, E.G. and Dyer, W.J. (1959) A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911-917.
[23] Folch, J. Lees, M. and Sloane-Stanley, G.H. (1957) A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 226, 497-509.
[24] Kates, M. (1986) Techniques in lipidology, 2nd edn. pp. 232-240, Elsevier, Amsterdam.
[25] Lowry, D.H., Rosebrough, N.J., Fare, A.L. and Randall, R.J. (1951) Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193, 265-275.
[26] Fry, M. (1991) Mitochondria of Plasmodium. In: Biochemical Protozoology (Coombs, G.H. and North, M.J., eds.), pp. 154-167, Taylor and Francis, London.
[27] Martin, E. and Mukkada, A.J. (1979) Identification of the terminal respiratory chain in kinetoplast mitochondrial complexes of Leishmania tropica promastigotes. J. Biol. Chem. 254, 12192 12198.
[28] Jeronimo, S.M.B. and Pearson, R.D. (1993) The Leishma- nia: protozoan adapted for extracellular and intracellular survival. In: Subcellular Biochemistry (Avila, J.L. and Harris, J.R., eds.), Vol. 18, pp. 1-37, Plenum Press, London.
[29] Kaneshiro, E.S., Meyer, K.B. and Reese, M.L. (1983) The neutral lipids of Paramecium tetraurelia: changes with culture age and detection of steryl esters in ciliary membranes. J. Protozool. 30, 392-396.
[30] Barr, R., Bransletter, B.A., Rajicek, A., Crane, F.L. and
L6w, H. (1991) Chloroquine-sensitive transplasmalemma electron transport in Tetrahymena pyriformis: a hypothesis for control of parasite protozoa through transmembrane redox. Biochim. Biophys. Acta 1058, 261-268.
[31] Weinbach, E.C. (1981) Biochemistry of enteric parasitic protozoa. Trends Biochem. Sci. 6, 254-257.
[32] Weinbach, E.C., Harlow, D.R., Clagett, C.E. and Diamond, L.S. (1977) Entamoeba histolytica: diaphorase activities. Exp. Parasitol. 41, 186-197.
[33] Paget, T.A., Manning, P. and Jarroll, E.L. (1993) Oxygen uptake in cysts and trophozoites of Giardia lamblia. J. Euk. Microbiol. 40, 246-250.
[34] Ellis, J.E., Williams, R., Cole, D., Cammack, R. and Lloyd, D. (1993) Electron transport components of the parasitic protozoon Giardia lamblia. FEBS Lett. 325, 196- 200.
[35] Schneider, H., Lemasters, J.J. and Hackenbrock, C.R. (1985) Membrane fluidity and mobility of ubiquinone. In: Coenzyme Q. Biochemistry, Bioenergetics and Clinical Applications of Ubiquinone (Lenaz, G., ed.), pp. 201-214, Wiley Interscience, Chichester, UK.
[36] Rudzinska, M.A., D'Alesandro, P.A. and Trager, W. (1964) The fine structure of Leishmania donovani and the role of the kinetoplast in the leishmania to leptomonad transformation. J. Protozool. 11, 166-191.
[37] Hart, D.T. and Coombs, G.H. (1982) Leishmania mexicana: energy metabolism of amastigotes and promas- tigotes. Exp. Parasitol. 54, 397-409.
[38] Pan, A.A. and Pan, S.C. (1986) Leishmania mexicana: comparative fine structure of amastigotes and promasti- gotes in vitro and in vivo. Exp. Parasitol. 62, 254-265.
[39] Matsura, T., Yamada, K. and Kawasaki, T. (1991) Changes in the content and intracellular distribution of CoQ homologs in rabbit liver during growth. Biochem. Biophys. Acta 1083, 277-282.
[40] Lloyd, D. and Coombs, G.H. (1989) Aerotolerantly anaerobic protozoa: some unsolved problems. In: Bio- chemistry and Molecular Biology of 'Anaerobic' Protozoa (Lloyd, D., Coombs, G.H. and Paget, T.A., eds.), pp. 267- 286, Harwood Academic, Chur, Switzerland.
[41] Haidaris, C.G. and Bonventre, P.F. (1982) A role for oxygen-dependent mechanisms in killing of Leishmania donovani tissue forms in activated macrophages. J. Immunol. 129, 850-855.
[42] Pan, A.A., Duboise, S.M., Eperon, S.M., Rivas, L., Hodgkinson, V., Traub-Cseko, Y. and McMahon-Pratt, D. (1993) Developmental life cycle of Leishmania cultiva- tion and characterization of cultured extracellular amasti- gotes. J. Euk. Microbiol. 40, 213 223.
[43] Soldo, A.T. (1983) The biology of the xenosome, an intracellular symbiont. Int. Rev. Cytol. Suppl. 14, 79-109.
[44] Soldo, A.T., Brickson, S.A. and Vazquez, D. (1992) The molecular biology of a bacterial endosymbiont. J. Proto- zool. 39, 196-198.
[45] Holz, G.G., Jr. (1985) The lipids of Leishmanias. In: Leishmaniasis (Chang, K.-P. and Bray, R.S., eds.), pp. 81 92, Elsevier, Amsterdam.

224 J.E. Ellis et al./Molecular and Biochemical Parasitology 65 (1994) 213-224
[46] Conner, R.L., Landrey, J.R., Burns, C.H. and Mallory, F.B. (1968) Cholesterol inhibition of pentacyclic triterpe- noid biosynthesis in Tetrahymena pyriformis. J. Protozool. 15, 600q505.
[47] Miller, J.E. (1965) Biosynthesis of the benzoquinone ring of ubiquinone in Tetrahymena pyriformis. Biochem. Biophys. Res. Commun. 19, 335 339.
[48] Beach, D.H., Holz, G.G. Jr., Singh, B.N. and Lindmark, D.G. (1990) Fatty acid and sterol metabolism of cultured Trichomonas vaginalis and Tritrichomonas foetus. Mol. Biochem. Parasitol. 38, 175-190.
[49] Jarroll, E.L., Manning, P., Berrada, A., Hare, D. and Lindmark, D.G. (1989) Biochemistry and metabolism of Giardia. J. Protozool. 36, 190-197.
[50] Van Vliet, H.H.D.M., Op den Kamp, J.A.F. and Van Deenen, L.L.M. (1975) Lipid auxotrophy and the effect on lipid composition of Entamoeba invadens. J. Protozool. 22, 428-432.
[51] K6hler, P. (1985) New strategies of energy conservation in helminths. Mol. Biochem. Parasitol. 17, 1-18.
[52] Kaneshiro, E.S., Gottlieb, M. and Dwyer, D.M. (1982) Cell surface origin of antigens shed by Leishmania
donovani during growth in axenic culture. Infect. Immun. 37, 558-567.
[53] Pan, A.A. (1984) Leishmania mexicana: serial cultivation of intracellular stages in a cell-free medium. Exp. Parasitol. 58, 7~80.
[54] Lui, X. and Chang, K-P. (1992) Extrachromosomal genetic complementation of surface metalloproteinase (gp63)- deficient Leishmania increases their binding to macro- phages. Proc. Natl. Acad. Sci. USA 89, 4991-4995.
[55] Diamond, L.S., Harlow, D.R. and Cunnick, C.C. (1978) A new medium for axenic cultivation of Entamoeba histoly- tica and other Entamoeba. Trans. R. Soc. Trop. Med. Hyg. 72, 431-432.
[56] Keister, D.B. (1983) Axenic culture of Giardia lamblia in TYI-S-33 medium supplemented with bile. Trans. R. Soc. Trop. Med. Hyg. 77, 487-488.
[57] Diamond, L.S. (1957) The establishment of various trichomonads of animals and man in axenic cultures. J. Parasitol. 43, 488-490.
[58] Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) In: Molecular Cloning: a Laboratory Manual, Vol. 3, 2nd edn., Cold Spring Harbor, New York.





























