Embed Size (px)
Citation preview

CURRENT STATUS OF WHEAT STEM RUST (Puccinia
graminis f. sp. tritici) AND REACTIONS OF GENOTYPES TO
PREDOMINANT RACES IN ARSI AND BALE ZONES OF
OROMIA, ETHIOPIA
M.Sc THESIS
LIDIYA TILAHUN HADIS
FEBRUARY 2019
JIMMA UNIVERSITY

i
CURRENT STATUS OF WHEAT STEM RUST (Puccinia graminis f. sp.
tritici) AND REACTIONS OF GENOTYPES TO PREDOMINANT
RACES IN ARSI AND BALE ZONES OF OROMIA, ETHIOPIA
A Thesis Submitted to the School of Graduate Studies
Jimma University, College of Agriculture and Veterinary Medicine
In Partial Fulfillment of the Requirements for the Degree of
Master of Science in CROP PROTECTION
Lidiya Tilahun Hadis
February 2019

ii
Jimma University College of Agriculture and Veterinary Medicine
Thesis Submission Request Form (F-08)
Name of student Lidiya Tilahun Hadis ID. No. RM1379/09
Program of study Plant Protection
Title: Current status of wheat stem rust (Puccinia graminis f. sp. tritici) and reactions of
genotypes to predominant races in Arsi and Bale zones of Oromia, Ethiopia
I have completed my thesis research work as per the approved proposal and it has been
evaluated and accepted by my advisors. Hence, I hereby kindly request the Department to
allow me to present the findings of my work and submit the thesis.
Lidiya Tilahun Hadis
Name & signature of student
We, the thesis advisors have evaluated the contents of this thesis and found to be satisfactory,
executed according to the approved proposal, written according to the standards and format of
the University and is ready to be submitted. Hence, we recommend the thesis to be submitted.
Major Adviser: Girma Adugna (PhD, Assoc. Professor) ____________ ___________
Name Signature Date
Co-advisor: Bekele Hundie (PhD) _______________ ______________
Name Signature Date
Internal Examiner:
Name: Fikre Lemessa (Professor) Signature ___________ Date _________
Decision/ suggestion of Department Graduate Council (DGC)
___________________________________________________________________________
___________________________________________________________________________
Chairperson, DGC Signature Date
_______________________ ____________ ___________
Chairperson, CGS Signature Date
_______________________ _____________ ____________

iii
DEDICATION
I dedicate this piece of work to The Almighty God, from him, through him, and
to him are all things to him be Glory forever.
Amen!

iv
STATEMENT OF THE AUTHOR
I declare that this piece of research work is my own and all sources of materials used for this
thesis work have been duly acknowledged. The thesis has been submitted in partial fulfillment
of the requirements for the degree of Master of Science at Jimma University and is reserved at
the University library to be made available to users. I solemnly declare that this thesis work is
not submitted to any other institution anywhere for the award of any academic degree,
diploma, or certificate.
With accurate acknowledgment of the source, users are free to use this thesis without special
permission. Permission for extended quotation or duplication of the manuscript in whole or in
part may be granted by the Horticulture and Plant Science Department Head, or Dean of
School of Graduate Studies of Jimma University. In all other instances, however, permission
should be obtained from the author.
Name: Lidiya Tilahun Hadis
Place: Jimma, Ethiopia
Signature: _____________________________
Date of Submission: _____________________

v
BIOGRAPHICAL SKETCH
The author, Lidiya Tilahun Hadis, was born on 07th
January 1992 in Mieso town West
Harerge Zone, Oromia Regional State. She attended her elementary school at Mieso
Missionary School in Mieso and secondary school at Nafiyad Primary and Secondary School
in Adama. She pursued her preparatory school education at Hawas Preparatory School in
Adama, Oromia Regional State. Following the completion of her preparatory education, she
joined Wollo University College of Agriculture and graduated with BSc Degree in Plant
Science on 08th
July, 2013. After graduation, she joined the Ethiopian Institute of Agricultural
Research (EIAR), at Kulumsa Agricultural Research Center (KARC) under Crop Protection
Research Division as a junior researcher. After two years‟ service, she has joined the graduate
studies program of Jimma University College of Agriculture and Veterinary Medicine to
pursue a graduate study leading to a Master of Science degree in Crop Protection.

vi
ACKNOWLEDGEMENTS
First and for most, I would like to praise the Almighty God for his endless opportunity and
help that enabled me to continue my M.Sc study and complete this research work
successfully.
First of all, I would like to express my sincere gratitude and deepest appreciation to Dr. Girma
Adugna (major advisor) and Dr. Bekele Hundie (Co-advisor). Without their keen interest in
the subject, invaluable academic guidance and continuous encouragement, it would have been
difficult to bring this study to its successful completion. I am greatly indebted to thank
Ethiopian Institute of Agricultural Research (EIAR), for the study leave and financial support
for my study. I gratefully acknowledge the immense contribution made by Jimma University
College of Agriculture and Veterinary Medicine for its facilitation and fruitful guidance
during class and research work. I am also acknowledging USAID/CIMMYT/EIAR project
‘Seed Multiplication and Delivery of High-yielding Rust Resistant Bread and Durum Wheat
Varieties to Ethiopian farmers’ for financial support for my research activities.
My special appreciation and thanks goes to Dr. Netsanet Bacha, Dr Getaneh Woldeab, Mr
Tsegaab Tesfaye, Mr Kitesa Gutu and the whole Ambo Plant Pathology team for their keen
cooperation by providing different stem rust races for field evaluation and skillful technical
support during seedling management and evaluation under greenhouse condition at Ambo. I
am also indebted to thank Dr Erena Edae for his unreserved assistance for data analysis.
Many peoples at Kulumsa Agricultural Research Center provided support during the entire
course of my study. Especially, I would like to thank all plant protection staff members for
their valuable support. Mr. Tamrat Negash, Mr Habtemariam Zegeye and Mrs. Getenesh
Demise deserve my special thanks for their genuine and unreserved support during my field
work and keen facilitation during my study.
Finally, all my heartfelt appreciation goes to my father Mr. Tilahun Hadis and my mother Mrs
Letebirhan Bogale whose staunch support and unreserved love was a source of inspiration to
complete my MSc study. I would like to acknowledge my brother Mikael Tilahun for his
moral support and encouragement during my study. Last but not least, I would like to thank
my husband Mr Zerihun Tadesse for his strong support in the entire period of my MSc study.

vii
LIST OF ABBREVIATIONS AND ACRONYMS
AARC - Ambo Agricultural Research Center
ANOVA - Analysis of Variance
APR - Adult Plant Resistance
AUDPC - Area under Disease Progress Curve
CSA - Central Statistics Agency
CV - Coefficient of Variation
DF - Degrees of Freedom
EIAR - Ethiopian Institute of Agricultural Research
FAO - Food and Agriculture Organization
IR - Infection Rate
IT - Infection Types
KARC - Kulumsa Agricultural Research Center
m.a.s.l - Meter Above Sea Level
MS - Mean Square
RH - Relative Humidity
SAS - Statistical Analysis System
SV - Source of Variation

viii
TABLE OF CONTENTS
Page
DEDICATION ........................................................................................................................................... iii
STATEMENT OF THE AUTHOR .......................................................................................................... iv
BIOGRAPHICAL SKETCH ..................................................................................................................... v
ACKNOWLEDGEMENTS ...................................................................................................................... vi
LIST OF ABBREVIATIONS AND ACRONYMS ................................................................................ vii
TABLE OF CONTENTS ........................................................................................................................ viii
LIST OF TABLES ...................................................................................................................................... x
LIST OF FIGURES ................................................................................................................................... xi
LIST OF TABLES IN APPENDIX ......................................................................................................... xii
ABSTRACT.............................................................................................................................................. xiii
1. INTRODUCTION ................................................................................................................................... 1
2. LITERATURE REVIEW ....................................................................................................................... 5
2.1. Origin and distribution of wheat ....................................................................................................... 5
2.2. Wheat production and importance in Ethiopia .................................................................................. 5
2.3. Wheat production constraints in Ethiopia ......................................................................................... 7
2.4. Wheat stem rust (Puccinia graminis f. sp. tritici) .............................................................................. 8
2.4.1. Taxonomy and historical perspectives ........................................................................................ 8
2.4.2. Rust biology, life cycle and geographic distribution .................................................................. 9
2.4.3. Epidemiology of wheat stem rust ............................................................................................. 10
2.4.3.1. Sources of inoculum ........................................................................................................... 10
2.4.3.2. Disease development .......................................................................................................... 11
2.4.3.3. Disease symptoms .............................................................................................................. 12
2.5. Effect of stem rust on wheat growth ............................................................................................... 12
2.6. Yield loss and economic importance ............................................................................................... 13
2.7. Distribution of wheat stem rust in Ethiopia ..................................................................................... 14
2.8. Management of stem rust ................................................................................................................ 15
2.8.1. Cultural practices ...................................................................................................................... 15
2.8.1.1. Elimination of green bridge between wheat crops ............................................................. 15
2.8.1.2. Eradication of alternate host ............................................................................................... 15
2.8.1.3. Pathogen monitoring .......................................................................................................... 16
2.8.1.4. Escape/Avoidance .............................................................................................................. 16

ix
2.8.2. Host plant resistance ................................................................................................................. 16
2.8.2.1. Seedling or all-stage resistance .......................................................................................... 17
2.8.2.2. Adult plant or race non-specific resistance ........................................................................ 18
2.8.3. Chemical control ....................................................................................................................... 18
2.8.4. Integrated wheat rust management ........................................................................................... 19
3. MATERIALS AND METHODS .......................................................................................................... 20
3.1. Description of the study areas ......................................................................................................... 20
3.2. Field survey ..................................................................................................................................... 21
3.2.1. Field assessment for stem rust .................................................................................................. 21
3.2.2. Collection of stem rust samples ................................................................................................ 22
3.2.3. Stem rust race analysis .............................................................................................................. 23
3.3. Seedling stage evaluation of wheat genotypes ................................................................................ 27
3.4. Adult plant stage evaluation of wheat genotypes ............................................................................ 29
3.5. Data analysis ................................................................................................................................... 31
4. RESULTS AND DISCUSSION ............................................................................................................ 33
4.1. Status and distribution of wheat stem rust ....................................................................................... 33
4.2. Virulence and physiological race composition of P. graminis f.sp. tritici ...................................... 38
4.3. Wheat genotypes response for seedling test .................................................................................... 42
4.4. Wheat genotypes response for adult plant stage evaluation ............................................................ 44
4.4.1. Final rust severity (FRS) ........................................................................................................... 44
4.4.2. Coefficient of infection (Cl) ..................................................................................................... 47
4.4.3. Area under disease progress curve (AUDPC) .......................................................................... 48
4.5. Correlation between slow rusting parameters of wheat stem rust ................................................... 52
4.6. Correlation between thousand kernel weight and disease parameters ............................................ 52
5. SUMMARY AND CONCLUSION ...................................................................................................... 54
6. REFERENCES ...................................................................................................................................... 57
7. APPENDIX ............................................................................................................................................ 68

x
LIST OF TABLES
Page
Table 1. Description of the study areas ................................................................................................. 21
Table 2. List of wheat stem rust differential lines used for race analysis and nomiclture .................... 25
Table 3. Wheat P. graminis f. sp. tritici gene differential sets and infection phenotype coding .......... 26
Table 4. Description of wheat genotypes used for seedling and adult plant stage evaluation .............. 27
Table 5. Description of single stem rust races used for seedling and adult plant evaluation ................ 28
Table 6. Description of reference genotypes used for field evaluation ................................................. 30
Table 7. Description of single stem rust race nurseries......................................................................... 30
Table 8. Wheat stem rust incidence and severity status in Arsi and Bale zones, 2017 ......................... 33
Table 9. Incidence and severity of wheat varieties assessed in Arsi and Bale zones, 2017 .................. 36
Table 10. The intensity of wheat stem rust across altitude ranges in Arsi and Bale zones, 2017 ......... 37
Table 11. Stem rust races identified in Arsi and Bale zones in 2017 .................................................... 38
Table 12. Virulence/Avirulence spectrum of races identified from Arsi and Bale zones, 2017 ........... 40
Table 13. Summary of disease parameters and TKW for predominate stem rust races ........................ 48
Table 14. Values of different disease parameters and TKW for reference varieties and differential
lines ....................................................................................................................................... 51
Table 15. Correlation coefficient for disease parameters and TKW among the wheat genotypes for
predominant stem rust races .................................................................................................. 53

xi
LIST OF FIGURES
Page
Figure 1. Life cycle of wheat stem rust (Puccinia graminis f.sp. tritici) ................................................ 9
Figure 2. Geographical locations for stem rust survey zones of Oromia region ................................... 20
Figure 3. Infection types of P. graminis f. sp. tritici stem rust and host response ................................ 25
Figure 4. Schematic overview of the protocols for seedling evaluation of genotypes in the greenhouse
at AARC, Ethiopia; (A) Seven days old seedling, (B) Inoculation of stem rust race, (C)
Seedling in the dew chamber for rust infection establishment and (D) Susceptible seedling
for stem rust ........................................................................................................................... 29
Figure 5. Stem rust prevalence across growth stage in Arsi and Bale zones during 2017 .................... 34
Figure 6. Disease prevalence across varieties in Arsi and Bale zones during 2017 ............................. 35
Figure 7. Distribution of stem rust races in Arsi and Bale zones in 2017 ............................................ 39
Figure 8. Percentage of stem rust races identified in Arsi and Bale zones, 2017 ................................. 41
Figure 9. Response of wheat genotypes at seedling stage for the four predominant races ................... 42
Figure 10. Response of wheat genotypes to stem rust race combination at seedling stage .................. 43
Figure 11. Response of wheat genotypes for individual stem rust races at Kulumsa in 2017 .............. 45
Figure 12. Stem rust severity of wheat genotypes for individual races at Kulumsa in 2017 ................ 45
Figure 13. Coefficient of infection (%) of wheat stem rust races on 202 genotypes at Kulumsa in 2017
............................................................................................................................................. 47
Figure 14. AUDPC values for reference varieties for predominant Sr races at Kulumsa in 2017 ....... 49
Figure 15. AUDPC values for 202 genotypes for predominant stem rust races at Kulumsa in 2017 ... 50

xii
LIST OF TABLES IN APPENDIX
Page
Appendix Table 1. Data sheet for wheat stem rust assessment in Arsi and Bale zones of Ethiopia
(2017) ...................................................................................................................... 69
Appendix Table 2. Stem rust races identified from Arsi and Bale zones during 2017 ........................ 70
Appendix Table 3. List of genotypes with different disease parameters scored for the four
predominant stem rust races ................................................................................... 71
Appendix Table 4. ANOVA mean square for TKW, AUDPC and ACI of the four stem rust races ... 78

xiii
CURRENT STATUS OF WHEAT STEM RUST (Puccinia graminis f. sp. tritici) AND
REACTIONS OF GENOTYPES TO PREDOMINANT RACES IN ARSI AND BALE
ZONES OF OROMIA, ETHIOPIA
Advisors: Girma Adugna (PhD, Assoc. Professor) and Bekele Hundie (PhD)
ABSTRACT
Wheat stem rust caused by Puccinia graminis f. sp. tritici is one of the most destructive
diseases of wheat which causes considerable yield losses in wheat growing areas worldwide.
Disease resistant cultivars provide one of the best means of controlling wheat stem rust. The
current study aimed at assessing the distribution of wheat stem rust in Arsi and Bale zones in
2017 cropping season and evaluating wheat genotypes resistance at seedling and adult plant
growth stage against the four predominant wheat stem rust races; TTKSK, TKTTF, TRTTF
and JRCQC. Wheat stem rust distribution was conducted in the main wheat belt areas of the
country namely in Arsi and Bale zones. Both seedling and adult plant stage evaluation
conducted on 202 wheat genotypes under greenhouse and field conditions, respectively. The
seedling resistance was evaluated by inoculating the wheat genotypes with four races
separately using completely randomized design with two replications at Ambo Agricultural
Research Center. The same wheat genotypes were evaluated for adult-plant stage resistance
at Kulumsa Agricultural Research Center using augmented design with five repeated checks
per blocks. Field survey result revealed that the distribution of stem rust varied depending on
agro-ecologies and varieties grown. Overall the average prevalence of stem rust was 63.0%
with individual zone prevalence of 52.2% in Arsi and 73.9% in Bale. From surveyed wheat
stem rust samples, five races (TKTTF, TTTTF, TTRTF, TTKSK and TRTTF) were identified
from Arsi and Bale zones. The most predominant races were TKTTF (53.0%), TTTTF (31.0%)
and TTRTF (10.0%). These races are virulent for 17-18 Sr genes. Seedling evaluation results
showed high level of variability in responses of genotypes for the four stem rust races.
Phenotypic variation was observed for infection types and level of stem rust severity for wheat
genotypes for field evaluation. There was highly significant difference among reference
variety and new genotypes for different disease parameters for each single race nursery.
From this study, five genotypes (ETBW 9045, ETBW 9464, ETBW 9569, K6295-4A and
Alidoro) showed seedling susceptible and field resistant reaction for the three major bread
wheat stem rust races (TKTTF, TRTTF and TTKSK). Therefore, this genotypes will be
potential source of resistance for further wheat breeding and pathological activities.
Key Words: Wheat, stem rust, race, resistance

1
1. INTRODUCTION
Wheat (Triticum aestivum L.) is one of the most important food security crops in the world,
accounting for a sizeable share of the global calories and protein consumed (Shiferaw et al.,
2013). It is one of the most widely grown cereal crops globally around 218 million hectares
with an average total production of 771 million metric tons worldwide. Global average
productivity is around 3.5 t ha-1
with high variability among countries and regions. It is the
most important food grain source for humans supplying 40% of the world‟s food and 25% of
calories consumed in developing countries (FAO 2017).
Wheat is an important staple food in the diets of several Ethiopian, providing about 15% of
the caloric intake for the country‟s over 90 million populations (FAO 2017), placing it second
after maize and slightly ahead of teff, sorghum, and enset, which contribute 10-12 percent
each (Minot et al., 2015). Bread wheat (Triticum aestivum L. Thell) and durum wheat
(Triticum. turgidum L. var. durum) are the two wheat species cultivated in Ethiopia.
Production and productivity of wheat shows increasing trend from time to time. On average,
for the last two decades, wheat area production and productivity increased by 38,762.98
hectare per year, 0.16 million tons per year and 0.07 tons per hectar per year, respectively. It
shows increasing scenario of area (92.29%), production (321.61%) and productivity
(119.26%). However, the productivity of wheat per unit area is low as compare to world
average (3.41 t ha-1
) (CSA, 1997-2017; FAO, 2017).
According to CSA (2017/18) report, the land cultivated to wheat is increased to 1.69 million
hectares and production has boosted to 46.42 million quintals with productivity of 2.73 t ha-1
,
which is still below the world‟s average of 3.5 t ha-1
(FAO, 2017). About 4.21 million
smallholder farmers (SHFs) engaged in wheat production in the country. Most wheat
production in Ethiopia comes from smallholder farmers and it is mainly grown in the central
and southeastern highlands. In Oromia region, wheat is one of the most important cereal crops
cultivated on 0.89 million hectares with production of 26.69 million quintals (CSA, 2017/18).
Arsi and Bale zones are included among the highest potential agro-ecologies in Eastern Africa
for wheat production with 0.47 million ha under wheat (Jobie, 2007; CSA, 2017/18). In Arsi
zone, wheat is growing on 0.2 million hectares with production and productivity of 6.54

2
million quintals and 3.20 tones per hectares, respectively. Similarly, in Bale zone, 0.16
million hectares of land covered by wheat with production and productivity of 5.55 million
quintals and 3.33 tones per hectares, respectively (CSA, 2017/18). This two zones
contributing 21.84% and 26.06% of the total area and production of wheat in Ethiopia,
respectively. This two zones has also a total share of nearly half of the area and production of
wheat from Oromia region. In addtion, 348,271 and 163,513 small scale wheat farmers
engaged on wheat production in Arsi and Bale zones, respectively (CSA, 2017/18).
Despite enormous economic and dietary values of the crop, the average yield has remained
low as compared to other wheat producing countries. This has been attributed to multifaceted
biotic and abiotic factors including insufficient/excess and erratic rainfall, poor agronomic
practices, poor soil fertility, diseases and insect pests. Wheat is susceptible to many diseases
including the highly destructive ones like rusts (Puccinia spp.), Septoria leaf blotches
(Septoria tritici), Fusarium head blight (Fusarium graminearum), Tan spot (Pyrenophora
tritici repentis), Smut (Ustilago tritici) and Powdery mildew (Erysiphe graminis f.sp.tritici)
(Prescott et al., 1986). Over 30 diseases have been reported on wheat in Ethiopia (Bekele
1985). Of these, fungal diseases like rusts (stem, stripe and leaf rust), Fussarium head blight
(FHB), Septoria blotch, Helmenthosporium spp., and tan spot are the dominant ones that were
reported over time (Badebo 2002; CIMMYT 2005).
Rusts, mainly stem and yellow rusts, have been considered the most important wheat diseases,
and thus, have been one of the most important priority research agenda since the inception of
wheat research on wheat diseases. The disease causing wheat rust fungi spread in the form of
clonally produced dikaryotic uredio spores, which can be dispersed by wind for thousands of
kilometers from initial infection sites across different areas from continent to continent.
Epidemics of wheat rusts can occur on a continental scale due to the widespread dispersal of
uredio spores (Khan et al., 2013). Wheat rust fungi are highly host specific obligate parasites
and changing to virulent races through mutation and sexual recombination. Consequently, the
three wheat rusts; leaf, stripe and stem rusts have played a devastating role in crop yield
reduction and significantly affected the socio-economic instability in the world (Rehman et
al., 2013).

3
Stem rust or black rust caused by Puccinia graminis f. sp. tritici is amongst the rust diseases
that can cause up to 100% yield loss if susceptible cultivars are grown and epidemic occurs
(Park, 2007). In Ethiopia, stem rust epidemics has knocked out major cultivars such as Enkoy
with Sr36 gene in 1994; and Digalu with SrTmp gene in 2013 and 2014, causing 100 percent
yield loss (Badebo and Hundie, 2016). The Digalu race (TKTTF) which is different from the
Ug99 race (TTKSK) is dominant across the major wheat growing regions of Ethiopia (Badebo
and Hundie, 2016) and becomes a major threat to wheat production in the country. Stem rust
isolates with virulence to Sr9e and Sr13 were first reported in Ethiopia in 1988 and 1989,
respectively (Hulluka et al., 1991).
According to Singh et al., 2006, the highland of Ethiopia is considered as a “hot spot” for the
development of stem rust diversity and nine races were identified, which include TTKSK,
TTKTF, TTKTK, JRCQC, TKTTF, TTKSC, TRTTF, SRKSC and RRKSF (Endale et al.,
2015). The high virulence diversity and evolution rate of the pathogen makes a considerable
proportion of wheat germplasm at risk in the country (Admassu et al., 2009). The persistence
of stem rust as a significant disease in wheat can be attributed to specific characteristics which
have a capacity to produce a large number of spores, disseminated by wind over long
distances and the ability to change genetically, thereby producing new races with increased
aggressiveness on resistant wheat cultivars (Roelfs et al., 1992; Singh et al., 2015).
Host resistance and chemical option are the two major rust managements means in Ethiopia
and worldwide. Host resistance (genetic control) has advantages for environmental and
economic reasons, particularly for farmers in the developing world because of the possibility
that rust pathogens develop resistance to fungicides (Oliver, 2014). Resistance conditioned by
major genes has been most widely emphasized strategy for mitigating rust threats and
reducing losses incurred. Pyramiding of several major genes into a single cultivar is also an
attractive breeding strategy for increasing resistance durability by reducing stepwise
accumulation of virulence by the pathogen against each gene. The alternative is the
development and employment of cultivars carrying durable or slow rusting resistance based
on quantitatively inherited, multiple genes referred to as adult plant resistance (Oliver, 2014).

4
Several wheat stem rust epidemics have been recorded in different parts of Ethiopia in recent
history that have caused great losses (Olivera et al., 2015 and Olivera et al., 2017). In 2016,
yield losses of 70.70% and 60.00% were reported in Arsi and Bale zones of Oromia region,
respectively (Hei et al., 2017). Currently, most of the released commercial bread wheat
varieties by the national wheat research program frequently defeated by new races of stem
rust. There is a need for farmers to use improved wheat genotypes possessing high resistance
to emerging new physiological races of Puccinia graminis f.sp. tritici. This necessitates, new
strategy to screen and identify resistant wheat genotypes by screening both at seedling and
adult plant stage to confirm adult plant resistant. Therefore, this study was designed based on
the following objectives.
General objective
To assess the current distributions of wheat stem rust and evaluate the reactions of
genotypes to major races of Puccinia graminis f. sp. tritici in Arsi and Bale zones of
Oromia, Ethiopia
Specific objectives
To assess the prevalence, incidence and severity of wheat stem rust in the Arsi and
Bale zones of Oromia in 2017 main cropping season
To evaluate released and promising wheat genotypes for their seedling and adult
plant reaction to predominant stem rust races

5
2. LITERATURE REVIEW
2.1. Origin and distribution of wheat
Wheat (Triticum Spp) is believed to have originated in South-western Asia. Some of the
earliest remains of the crop have been found in Syria, Jordan, and Turkey (Heun et al., 1997).
Primitive relatives of present day wheat have been discovered in some of the oldest
excavations of the world in eastern Iraq, which date back to 9,000 years and other
archeological findings show that bread wheat was grown in the Nile Valley about 5,000 B.C.
As well as in India, China, and even England at about the same time (Anonymous, 2015c).
Bread wheat (Triticum aestivum, 2n = 6x = 42, AABBDD) is an allohexaploid species that
consists of three genomes formed through two separate amphidiploidization events. The first
event involved hybridization of Triticumurartu (2n=2x=14, AA) with an unknown source of
the B genome, but speculated to be Aegilops speltoides (2n=2x=14, BB), that led to the
formation of wild T. dicoccoides (2n = 4x = 28, AABB). Subsequent hybridization of a
domesticated T. turgidum spp. Dicoccon with Ae. Tauschii (2n=2x=14, DD) formed T. spelta
(2n=6x= 42, AABBDD), the nascent hexaploid wheat from which free-threshing forms were
derived by mutation (Devos and Gale 1997; Dvorak et al., 1998).
Ethiopia has a wealth of variability in its genetic resource. Vavilov recognized Ethiopia as a
center of origin for tetraploid wheat types. Later on, however, due to the absence of ancestral
forms and wild relatives, Harlan indicated that Ethiopia is the center of diversity for tetraploid
wheat. Six species including Triticumdicoccum, T. durum, T. polonicum, T. compactum, T.
turgidumand T. aestivumwere identified. However, this valuable wealth is threatened by
different causes of genetic erosion including displacement by improved varieties and other
crops, drought and change in land use. The Portuguese and the Italians, probably, introduced
the hexaploid wheat (T. aestivum) although the exact period of introduction is unknown.
2.2. Wheat production and importance in Ethiopia
In Ethiopia, wheat is grown between 6o and 14
o N latitudes; and between 35
o and 42
o E
longitudes ranging in altitude from 1500 m to 3200 m. However, the most suitable regions fall
between 1900 and 2700 m. Hence, altitude plays an important role in the distribution of wheat

6
production through its influence on rainfall, temperature, diseases and pests. Soil types and
moisture regimes of the wheat producing regions vary from area to area.
Wheat is one of the most important cereal crops cultivated in a wide range of agro-ecologies
in Ethiopia. Bread wheat (Triticum aestivum L. em.Thell) and durum wheat (T. turgidum L.
var. durum) are the two species which are mainly cultivated in Ethiopia mainly by
smallholder farmers. According to Central Statistical Agency report (CSA, 2013/14), out of
the total grain crop area, 79.38% hectares was under cereals and wheat took up 12.94%
hectares of the grain crop area.
Currently national yield average obtained from wheat crop is 2.73 t ha-1
(CSA, 2017/18). The
productivity of wheat crop was by far lower than the potential yields obtained on research
stations and on farm verification trials that was 5 - 7 t ha-1
(Hailu et al,. 2002, Desalegn et al.,
2006). According to annual CSA figures, between 2006/7 and 2017/18 the area devoted to
wheat production largely remained constant fluctuating between 1.42 - 1.68 million hectares,
with an annual average growth rate of 1.24% and annual average area of 1.5 million hectares
over these years (CSA, 1999-2018). Ethiopia needs to increase its wheat production, and it
has the agro-climatic potential to do this in order to meet its national wheat requirements.
Research results and the recently conducted country-wide technological package
demonstrations have revealed that use of improved packages can double, triple or even
quadruple the present yield levels.
Wheat is used for the manufacture of flour for different purposes. Bread, biscuits and pasta
products such as macaroni, spaghetti and noodles are some of the industrial products. Wheat
is known to be a major source of energy and protein. Traditionally wheat is used for making
"Dabo", "Dabo kolo", "Ganfo", "Kinche" and other types of food. The straw is good source
for animal feed and is also used for thatching roofs. In terms of quality, wheat provides an
optimum amount of energy, protein, calcium and iron (Abera, 1991). The protein to energy
ratio of wheat is 12% as compared to 10.8% of barley, 10.7% of teff, 10.5% of sorghum and
10.1% of maize.
Compared to other cereals, farmers sell higher proportion of wheat that they produce and
generate income (CSA 2008-2015). The remaining is used for a combination of seed, in kind

7
payments for labor and animal feed. Wheat is thus one of the most important crops for food
security and farmer incomes in Ethiopia. The marketed volume of wheat covers only about
30% of the annual demand of the more than 200 mills found dispersed in the country, whose
combined annual capacity exceeds 2.3 million tons (EGTE Reports); some studies put this
figure as high as 3.2 million tons (Abu, 2012).
2.3. Wheat production constraints in Ethiopia
Wheat production in Ethiopia is constrained by a number of abiotic and biotic stresses at
different levels of intensity. This is further accentuated by the increasing incidence of climate
change, characterized by rising temperature (heat), less and erratic rainfall (drought) or
sometimes excessive rainfall (flooding), and when combined with virulent pests and diseases,
it makes agricultural productivity less predictable. Among abiotic factors, soil fertility and
moisture stress are the principal wheat production limiting factors in Ethiopia (Bogale et al.,
2011a; Bogale et al., 2011b; Haile et al., 2012).
The major biotic factors that limit wheat production in the country include diseases, pests and
weeds (Abebe et al., 2012). Among the diseases, rusts (stem rust (P. graminis f.sp. tritici), leaf
rust (P. triticina Eriks) and stripe rust (P. striiformis Westend. f.sp. tritici) are the most
important diseases reducing wheat production in Ethiopia. From the three rusts, stem rust has
been the most important disease of wheat in main wheat growing regions of Ethiopia
(Admassu et al., 2009; Denbel et al., 2013).
Stem rust (Puccinia graminis f.sp. tritici) is an obligate parasite, requiring living host tissue
for growth and reproduction (Kolmer et al., 2013). In epidemic situations, it causes yield
losses reaching up to 100% on susceptible cultivars (Park, 2007; Hodson, 2014). Stem rust
causes yield losses in several ways. The fungus absorbs nutrients from the plant tissues that
would be used for grain development in a healthy plant. By the time rust pustules break
through the epidermal tissue, the rust will have affected the plant transpiration, making the
metabolism less efficient. Interference with the vascular tissues results in shriveled grains.
Stem rust also can weaken wheat stems, so plants lodge, or fall over, in heavy winds and rain
(Roelfs, 1985a).

8
In Ethiopia, stem rust epidemics has knocked out major cultivars such as Enkoy with Sr36
gene in 1994; and Digalu with SrTmp gene in 2013 and 2014, causing 100 per cent yield loss
(Badebo and Hundie, 2016). The Digalu race (TKTTF) which is different from the Ug99 race
(TKTTSK) is dominant across the major wheat growing regions of Ethiopia (Badebo and
Hundie, 2016) and becomes a major threat to wheat production in the country.
2.4. Wheat stem rust (Puccinia graminis f. sp. tritici)
2.4.1. Taxonomy and historical perspectives
The wheat stem rust pathogen belongs to the genus Puccinia, family Pucciniaceae, order
Uredinales, class Urediomycetes, phylum Basidiomycota and kingdom Fungi. Several
variants of the species P. graminis have been recognized and described as subspecies (subsp.),
varieties (var.) or formaespeciales (f.sp.). According to the nomenclature of Savile (1984),
there are three distinct morphological forms of P. graminis. These include P.graminissubsp.
graminis var. graminis, P. graminis subsp. graminis var. stakmanii, and P. graminis subsp.
graminicola. This classification of subspecies is justified based on morphological features,
host ranges and distinct evolutionary histories.
Despite the complexities of host specialization, most literature finds the f.sp. nomenclature
more workable; the var. designation rarely appears while the f.sp. is commonly used
worldwide (Anikster, 1984; Savile, 1984). Races or pathotypes constitute a taxon below the f.
sp. level. These are characterized by differences in physiological reactions, resulting in
different virulence/avirulence patterns on a selected set of differential genotypes within a host
genus. Pathogenic specialization has been most thoroughly studied for P. graminis f.sp. tritici
and f.sp. avenae.
Stem rust is the most destructive disease of wheat in several wheat-growing countries around
the world (Roelfs et al., 1992). It is mainly favored by a humid and warm temperature
conditions, and in the presence of susceptible wheat crop (Leonard and Szabo 2005). Several
major wheat production areas in the world have favorable environments for stem rust
development. Historically, stem rust has caused major devastation to wheat production in
areas including Africa, the Middle East, Asia (except Central Asia), Australia, New Zealand,
Europe, North Americas and South Americas (Saari and Prescott, 1985; Roelfs et al., 1992).

9
2.4.2. Rust biology, life cycle and geographic distribution
The life cycle of P. graminis f. sp. tritici involves both sexual and asexual stages. Barberries
as well as wheat are required for the complete life cycle of the pathogen. The sexual cycle
occurs on the alternate hosts barberry species or less commonly Mahonia (Singh, 1998) and
the asexual stage on wheat or other grassy hosts. The pathogen is a polymorphic species
producing a succession of different types of spores. P. graminis is thus heteroceous and
heterothalic in its life cycle and there are five distinct spore stages. These are uredial, telial,
basidial, pycnidial and acial states. Basidiospores and pycniospores are haploid while the
others are dikaryotic (Singh, 1998). Telial stage, the only true diploid stage of the fungus,
enables the pathogen to survive cold or dry conditions. Late in its development on wheat,
stem rust infection is converted from the phase of producing urediospores to teliospores.
Teliospores remain dormant in the field until the following season when they germinate and
immediately enter into the uni-nucleate and haploid basidiospore stage (Alexopoulos et al.,
1996).
Source: Roelfs et al., 1992
Figure 1. Life cycle of wheat stem rust (Puccinia graminis f.sp. tritici)

10
The life cycle of P. graminis commonly consists of continual uredinial generations. The
repeating uredinial stage is the major means through which the fungus initiates and
perpetuates epidemics. The pathogen develops successful asexual reproduction strategy that
apparently allows it to maintain necessary genes in blocks that are occasionally modified by
mutation and selection (Singh et al., 2002). It survives during the non-crop season on
volunteer cereal crops or on other gramineous hosts as urediospores, which can directly infect
wheat during the crop season. Survival is generally difficult for the pathogen outside the
cereal growing season. However, a few surviving local uredia can produce local inoculum and
thereby cause more infections than spores transported from severely rusted fields 100 kms
away (Roelfs, 1985b).
Stem rust (P. graminis) occurs in most areas of the world on wild grasses or cultivated
cereals. Most of the geographic information concerns distribution on cultivated cereals mainly
on wheat (Kurt, 2001). The geographic distribution of the pathogen and its epidemics appear
to be influenced by the prevailing climatic conditions, the movement of global air masses,
geographic features, availability of alternative grassy hosts or alternate sexual hosts and
cropping practices (Roelfs et al., 1992). Urediospores are long-lived and are resistant to
atmospheric conditions if their moisture content is moderate (20 to 30%) and can be carried
long distances (Singh et al., 2002). Due to such long distance transport of urediospores, the
occurrence of stem rust is limited only by the deposition pattern of the spores, suitable
weather conditions and the availability of susceptible hosts.
2.4.3. Epidemiology of wheat stem rust
2.4.3.1. Sources of inoculum
The source of primary inoculum of stem rust that infects the new wheat crop in the season
differs depending on the region in which the crop is grown. (Roelfs, 1985a). There are
generally two basic sources of inoculum for the cereal hosts, the urediospores and the
aeciospores. Urediospores originates from infected volunteer plants grown in the field or
roadside and/or from long distance transport within or outside the epidemiological zones. It
serves as a consistent and early source of aeciospres and a breeding ground for new
pathogenic races of the fungus (Kurt, 2001). The existence of barberry plants in Ethiopia was

11
reported from around Debresina and Menagesha areas but rust inoculum from this plant did
not infect wheat and other hosts of the pathogen (Sorokina et al., 1980). In Ethiopia
urediospores apparently over-season on volunteer plants and long distance transport of the
pathogen from other regions within or between epidemiological zones in East Africa is
probably an important source of inoculum.
Urediospores are the most principal inoculum for annual infection of wheat by stem rust in
most wheat producing regions of the world. In Bale zone, the bimodal rainfall pattern
provides green bridges between two seasons that could help the pathogen to recycle its
uredinial stages and assist the movement of the primary inoculum from previous crop to the
next crop (Bekele, 2003). Once the disease is established uniformly in a field, even at
extremely low levels, inoculum from nearby plots, other fields, or more distant regions does
not significantly affect the rate of rust development, eventual disease severity, or yield
(Peterson, 2001).
2.4.3.2. Disease development
The uredial stage of P. graminis is favored by hot days (25-30ºC) and mild nights (15-20ºC)
and wet leaves from rain or dew. The minimum, optimum and maximum temperatures for
spore germination are 2, 15 to 24 and 30ºC, respectively (Roelfs et al., 1992) and for
sporulation 5, 30 and 40ºC, respectively (Hogg et al., 1969; Rowell, 1984). Both aeciospores
and urediospores require free water for germination. Light is an important factor for the
development of penetration pegs from the appressorium, but it is seldom a limiting factor in
the field as dew often occurs in the morning. However, little infection results when evening
dew and/or rain are followed by winds causing a dry off prior to sunrise (Rowell, 1984). The
most important factor affecting the rate of stem rust development, ultimate severity, and
ultimate damage caused by the disease is the frequency of days with six or more hours of
moisture at 10ºC or higher, favorable period for infection, prior to dough stage of plant
growth (Peterson, 2001).
The uredial stage is initiated by germination of urediospores on its hosts, penetration,
development of an intercellular mycelium with intercellular haustoria and subsequent
sporulation. New urediospores are released approximately 7 to 15 days after infections

12
causing local buildup of disease (Hogg et al., 1969). Hence, there can be multiple generation
of inoculum produced during a single growing season. One uredinum can produce at least
100,000 urediospores leading to explosive epidemics during favorable environment. Disease
severity can increase extremely rapidly once the crop is uniformly infected (Beard et al.,
2004). Like all cereal rusts, the spores of P. graminis spread by plant-to-plant contact and by
air dispersal (Peterson, 2001). Most spores are spread only over short distances and contribute
to local epidemics within the crop. A very small proportion of spores gets into the atmosphere
and is capable of transport to long distances causing new infections.
2.4.3.3. Disease symptoms
The onset of stem rust is marked by an outbreak of elongated brown pustules on the leaves,
stems, leaf sheaths, spikes, glumes, awns and occasionally on grains of hosts; however, stem
and leaf sheaths are the main tissues affected (Singh, 1998). These pustules (uredia) may be
about 6mm or more in length and frequently run into one another. They burst early exposing a
brown powder (consisting of urediospores) and are surrounded by prominent epidermal
fringes (Singh, 1998). The telial stage occurs in the same tissue as the uredinial stage, but
becomes shiny-black. The teliospores are sessile, and the telial tissue is, therefore, firmer than
the uredinial tissue and hence no spores are released (Singh, 1998).
The pycnial stage occurs on the young leaves of the alternate host, mainly Berberis vulgaris.
Pycnial infections initially appear as light, chlorotic areas on the adaxial leaf surface, and then
become light orange-brown lesions, consisting of individual small cone-shaped eruptions (the
pycnia), often occurring in clusters (Singh et al., 2002). The aecia develop on the abaxial
surfaces of the leaves of the alternate host. When mature, they appear as bright orange,
closely packed, raised clusters of individual aecia. The aecia are cylindrical in shape and flare
out at their apices, appearing as a grouping of rings within the aecial cluster.
2.5. Effect of stem rust on wheat growth
Infection by stem rust under favorable environmental conditions can result in death of tillers
or entire plant. Heavy epidemic can halt plant growth or even kill the plant by reducing the
photosynthetic area, causing a loss of nutrient and water and disturbing the plant transport
system (Rowell and Roelfs, 1976). Stem rust attacks all above ground parts of the plant (Roelf

13
et al., 1992). Lesions of rust can occupy a significant portion of the host tissue, which are the
sources of most of the nutrients that are translocated to the developing grain (Roelfs, 1985b).
The rupture of the plant epidermal cells by the fungus results in a loss of water from the plant.
Because the pathogen also uses both water and nutrients from the plant to produce the large
volume of urediospores daily, the plant suffers added stress. An infected plant also has less
root growth, aggravating the imbalance in normal water requirements. Uredium of stem rust
develops on leaf sheaths and peduncle tissue. The fungus often penetrates through the tissue
of true stem. The rupture of the plant tissue by the fungus can disrupt transport of nutrients to
the roots and cause premature death of the roots (Bushnell and Rowell, 1968).
2.6. Yield loss and economic importance
Historically, stem rust has caused major devastation to wheat crops in most of the wheat
growing areas of the world (Roelfs et al., 1992). Damage due to stem rust is usually greatest
when the disease becomes severe before the grain is completely formed. In areas where
environmental conditions are favorable for disease development, susceptible varieties cannot
be grown (Kurt, 2001), but yield loss is generally influenced by resistance level, weather
conditions and the onset of the disease (Luig, 1985).
Widespread epidemics have been occurred regularly in Africa, China, and Asia (Saarii and
Prescott, 1985). Furthermore, highlands of Kenya, Ethiopia, Panama States, Brazil and South
India are reported to be the hot spot areas for stem rust (CIMMYT, 2005). According to
CIMMYT (2005), the losses due to stem rust of wheat alone in these countries ranged from
40% to 100% on susceptible varieties. In Ethiopia, losses in wheat yield due to stem rust have
been estimated to be 61% (Eshetu, 1985) whereas other reports indicated losses ranging from
67 to 100% on commercial wheat cultivars (Shank, 1994). Apart from losses in grain yield,
stem rust can significantly affect the test weight, grain quality and cause shriveled kernels
(Roelfs, 1985b). An increase of total nitrogen, protein nitrogen, soluble nitrogen and the ratio
of soluble to insoluble nitrogen occur as the rust develops (Singh, 1998).

14
2.7. Distribution of wheat stem rust in Ethiopia
Stem rust is one of the most important diseases of wheat in Ethiopia. The damage caused by
wheat stem rust can be more serious than any other cereal disease. It can destroy thousands of
hectares of a healthy crop with a high yield potential in less than a month (Hailu et al., 2015).
The source of primary inoculum of stem rust that infects the new wheat crop in the season
differs depending on the region in which the crop is grown (Roelfs, 1985a). The two basic
sources of inoculum for cereal hosts are the uredio spores and aecio spores. The uredospore
originates from infected volunteer plants grown in the fields or roadside and or from long
distance transport within or outsides the epidemiological zones.
Yield losses due to stem rust are estimated to reach up to 100% on susceptible cultivars in the
country (Hodson, 2013). In the past 20 years, severe epidemics of stem rust have been
reported in major wheat growing regions of Ethiopia. According to Shank (1994), the
continuous mono cropping of varieties like Enkoy and Lakech in 1993/94 in Oromiya region
(Arsi and Bale) resulted in yield losses of 65-100% and a 1998 epidemic attacked the high
yielding variety Kubsa and Shina, released in 1999 for north western Ethiopia. These varieties
were forced out of production because of stem rust epidemics in 2001. Since then there have
been no severe stem rust epidemics. But during the 2013/14 cropping season, the disease
caused 100% yield losses in cultivar Digalu (a reportedly resistant variety to stem rust) in
SNNPR and on the highlands of Oromiya region, (Olivera et al., 2015). The outbreak was
recorded in high altitude areas (>2200 m.a.s.l), although stem rust mainly occurred in the low
altitude areas of 1800 m.a.s.l, extending rust incidence to low, medium and high altitudes
production areas. Therefore, the highlands of Oromiya, SNNP, Tigray, and Amhara regions
are potential areas where wheat is predominantly cultivated and areas with frequent epidemic
occurrence in the country (Hailu et al., 2015).
Stem rust is particularly a major production constraint in most wheat-growing areas of
Ethiopia and the neighboring countries in east Africa, where yield losses of up to 100% have
been reported (Olivera et al., 2015). Major stem rust epidemics occurred in Ethiopia in 1974
and 1993, when popular wheat varieties, such as „Lacketch‟ and „Enkoy‟ suffered major yield
losses. Ug99 was first detected in Ethiopia in 2003 presenting a real threat to wheat
production in the country. In 2013/14, a new stem rust race designated as TKTTF has caused

15
severe epidemic on variety „Digalu‟ carrying SrTmp in Ethiopia (Olivera et al., 2015). The
continued evolution of new virulent races of the stem rust pathogen elucidates the dynamic
challenges of breeding for stem rust resistance.
2.8. Management of stem rust
2.8.1. Cultural practices
Several cultural methods can be used to reduce the intensity of an epidemic or provide long-
term partial control. Some of the strategies listed below;
2.8.1.1. Elimination of green bridge between wheat crops
The term “green bridge” refers to weeds and other crop volunteers that promote growth and
development of prevented pests and diseases in crop fields (Leonard and Szabo, 2005).
Through these green bridges diseases like rusts survive between seasons. The importance of
the green-bridge in carrying the disease from one crop to the next was emphasized by Zadoks
and Bouwman (Zadoks and Bouwman, 1985). In the case of stem rust, triticale, barley and
some grasses act as a green bridge. The green-bridge can effectively remove by tillage or
herbicides. In addition, in Ethiopian condition, two cropping cycle (main and short season) is
the main source of rust inoculum. This green bridge effect is mainly observed in Arsi and
Bale zones of wheat growing areas.
2.8.1.2. Eradication of alternate host
Common barberry (Berberis vulgaris L.) is an alternate host for wheat stem rust and also can
generate new races of stem rust (Jin et al., 2008). Eradication of barberry leads to delayed
onset of stem rust disease in the fields, reduced initial inoculum level in fields, and fewer
races due low frequency of evolution (Roelfs, 1985). In addition, successful eradication of
barberry has reduced the influence of the sexual cycle on the disease (Dubin and John, 2009).
Historically played an important role in sexual variability in Ethiopia and other countries
including United States. The first barberry eradication program was started in Rounen, France
(Roelfs, 1985a). The campaign to eradicate barberry in the U.S. started in 1918 (Campbell
and Long, 2001) and eliminated from most of wheat growing areas in U.S, thereby reducing
wheat rust epidemics in U.S. In Ethiopia, barberry is common especially in the northern part

16
of the country but less attention has been given to eradicate the plant (Personal
communication).
2.8.1.3. Pathogen monitoring
Population monitoring through trap nurseries was recommended by a CIMMYT expert panel
(CIMMYT, 2005). Trap nurseries are used for evaluating disease resistance sources
thoroughly by monitoring changes in the pathogen population. It can also be a source of
pathogen (stem rust races) cultures and for evaluating new germplasm (Ferrara, 2008). The
USDA‟s cereal rust laboratory in Minnesota is helping in race characterization by providing
assistance in developing stem rust nurseries at various locations in Africa and Asia to
determine presence and migration of Ug99 and other races in these areas (CIMMYT, 2005).
Ethiopia is one of target county benefited a lot from this initiative. In 1968, the first
international congress of Plant Pathology in London recommended global surveys for diseases
such as Pgt in order to determine regional differences in virulence (Park et al., 2010). For
global monitoring of cereal rust pathogens, they provided a broad outline for the global cereal
rust monitoring system (GCRMS) (Hodson, 2014). The success of this GCRMS is dependent
on the mutual cooperation at the international level by providing quality information on the
incidence of rusts and the pathotypes present (Park et al., 2010).
2.8.1.4. Escape/Avoidance
The timing of an epidemic is critical to the amount of damage that is sustained. The date of
disease onset is directly related to the development of an epidemic (Hamilton and Stakman,
1967) and is probably the single most important factor in determining the severity of the
epidemic (Roelfs, 1985a). Planting as early as possible and planting early-maturing cultivars
help to reduce the time of exposure of the crop to the pathogen and hence reduce yield loss.
2.8.2. Host plant resistance
Genetic resistance offers the most environmentally sustainable approach to control rust
diseases in wheat (Chen, 2005; Boyd, 2005). Developing wheat rust-resistant varieties has
been a priority of wheat breeding programs since the first description of the Mendelian nature
of resistance to stripe rust in wheat by Biffen (1905). Rust resistance genes in wheat are

17
generally categorized into two types; seedling (or all-stage) resistance and adult-plant
resistance (APR) that includes the high temperature adult plant (HTAP) resistance (Chen,
2005; Chen et al., 2014).
Since the occurrence of Ug99, CIMMYT breeders have been working to develop high
yielding wheat with high levels of stem rust resistance based on APR and indeed have
reported success and about 60% of CIMMYT lines and breeding germplasm carries Sr2
(Singh et al., 2011). Ethiopia is one of the country benefits from this germplasm source to
develop rust resistant varieties to the resource poor farmers.
2.8.2.1. Seedling or all-stage resistance
Seedling resistance, typically conferred by a single gene or simple combinations of single
genes, is a type of resistance that can be detected at the seedling stage and remains effective
throughout all stages of plant growth (Boyd, 2005; Chen et al., 2014). They are effective only
against isolates of the pathogen carrying the corresponding avirulence genes (race-specific
resistance), conferring protection in a gene-for-gene manner. Race-specific genes normally
provide highly effective resistance and are relatively easy to incorporate into commercial
cultivars due to simple inheritance. As a result, race specific resistance genes have been more
attractive to breeding programs and have provided significant economic benefits in
controlling rust disease in wheat. Despite these advantages, however, resistances conferred by
seedling resistance genes are often short lived, lacking the ability to provide an economic
level of protection over an extended period of time (Johnson, 1984). The lack of durability of
race-specific resistance is mainly due to their vulnerability to the rapid evolution of the
corresponding virulence in the pathogen that can overcome the effectiveness of such type of
resistance genes (McIntosh et al., 1995; Chen, 2005).
With the prospects of the current and historical circumstances in evolution of wheat rust
pathogen, notably the recent trend in evolution for more aggressive and highly virulent races,
it is a matter of time for race-specific resistance to succumb to the newly evolving virulent
races. This dictates the need for alternative strategies to deploy resistance genes in
commercial cultivars that provide a more lasting or durable protection.

18
2.8.2.2. Adult plant or race non-specific resistance
Adult plant resistance (APR) is a type of host defense mechanism that confers quantitative
and partial protection against the invading pathogen in a race non-specific manner (Chen,
2005). It is also known as polygenic, horizontal, minor, partial or slow rusting resistance, all
depicting its complex inheritance. Seedlings of wheat plants carrying adult plant resistance are
susceptible to the disease, but become effective and provide varying levels of resistance to
stem rust at various stages of post-seedling crop growth (Chen et al., 2014). Adult plant
resistance is often conditioned by quantitative trait loci that may have individually small
effects in reducing disease development, but operate collectively in an additive manner to
confer a high level of resistance.
APR is often characterized by its effectiveness against a wide range of pathogen races and has
been considered more durable, providing resistance without being readily overcome by the
change in pathogen virulence when the cultivar is widely grown in an area where the disease
is prevalent (Parlevliet, 1975; McIntosh et al., 1995). The deployment of cultivars carrying
APR based on multiple genes is particularly preferred to delay infection, growth and
reproduction of the pathogen in adult plants and circumvent “boom-and-bust” cycles (Chen, 2
005; Singh et al., 2005).
Despite the advantages of the quantitatively inherited APR genes in resistance, and the fact
that such genes are common within the primary gene pool and further, only limited number of
APR genes has been deliberately deployed by breeding programs. The complex inheritance of
highly effective APR resistance presents a relative difficulty in identification and routine
deployment of this type resistance in wheat breeding (Milus and Line, 1986; Chen et al.,
2007).
2.8.3. Chemical control
Currently, the large majority of wheat varieties are susceptible to either stem rust and/or
yellow rust. In the absence of an option to grow resistant varieties, the use of fungicides
becomes necessary. Adequate rust control could be attained by application of fungicides
before the rust starts and frequent application thereafter throughout the growing season
(Peterson, 2001). A number of fungicides are highly effective against stem rust and have been

19
used to successfully control the disease. These include triadimefon (Baylaton and Noble) 25%
WP at 0.5 l/ha, Rex Duoand propiconazole (Tilt and Bumper) 250 EC at 0.5 l/ha (Bekele,
2003).
Fungicides have not, however, been widely used for the control of cereal rust by subsistence
farmers. There are several reasons for this. The cost of fungicides is high and is a direct cost
to the producer. Chemical control is usually considered only where losses are expected to be
very high and where grain prices are highly subsidized and yields are high. Repeated
applications of fungicides are necessary under heavy epidemic conditions, increasing costs
further. Lack of knowledge and awareness about appropriate fungicides and unavailability of
the chemicals is also the main limitation particularly to the small-scale farmers. Early disease
detection and immediate application of fungicides should be considered in the control of stem
rust with fungicides. It has been reported that stem rust becomes more difficult to control as it
progresses. This is because fungicides reduce subsequent rust severity on plant parts that were
slightly infected at the time of fungicide application, but they are not effective on plant parts
that were heavily infected (Beard et al., 2004).
2.8.4. Integrated wheat rust management
Farmers have several options to manage wheat rusts of plant disease is most successful and
economical when all available pertinent information regarding the crop, its pathogens, the
history of disease in previous years, varietal resistance to diseases, the environmental
conditions expected to prevail, locality, availability of material, land, labor, and costs is taken
in to account in developing the control program.
Thus, in an integrated control program, several control methods are employed, including
regulatory inspections for nursery crop production, cultural practices (crop rotation,
sanitation, destruction of crop residues), biological control (resistant varieties, biocontrol
agents), physical control (temperature and moisture), and chemical control (fungicides). Each
one of these measures must be used for best results, and the routine use of each of them makes
all of them that much more effective (Agrios, 2005).

20
3. MATERIALS AND METHODS
3.1. Description of the study areas
This study consists of three broad activities; field survey, seedling and adult plant stage
evaluation of wheat genotypes. The detail description of each activities are discribed below.
Field survey: The survey of wheat stem rust (P.graminis f.sp. tritici) was carried out in the
major wheat growing areas of Arsi and Bale zones of Oromia during 2017 main cropping
season. Bale and Arsi highlands in southeastern Ethiopia are the major wheat producing areas
of Ethiopia and considered the wheat belt of East Africa. The region is also described as a
“hot spot” for wheat rusts ((Dereje Hailu and Chemeda Fininsa, 2009). The survey covered a
total of ten districts from Arsi and five districts from Bale zones. Detail discription of
districts, geographical location and weather conditions of the two zones listed in figure 2 and
table 1.
Figure 2. Geographical locations for stem rust survey zones of Oromia region
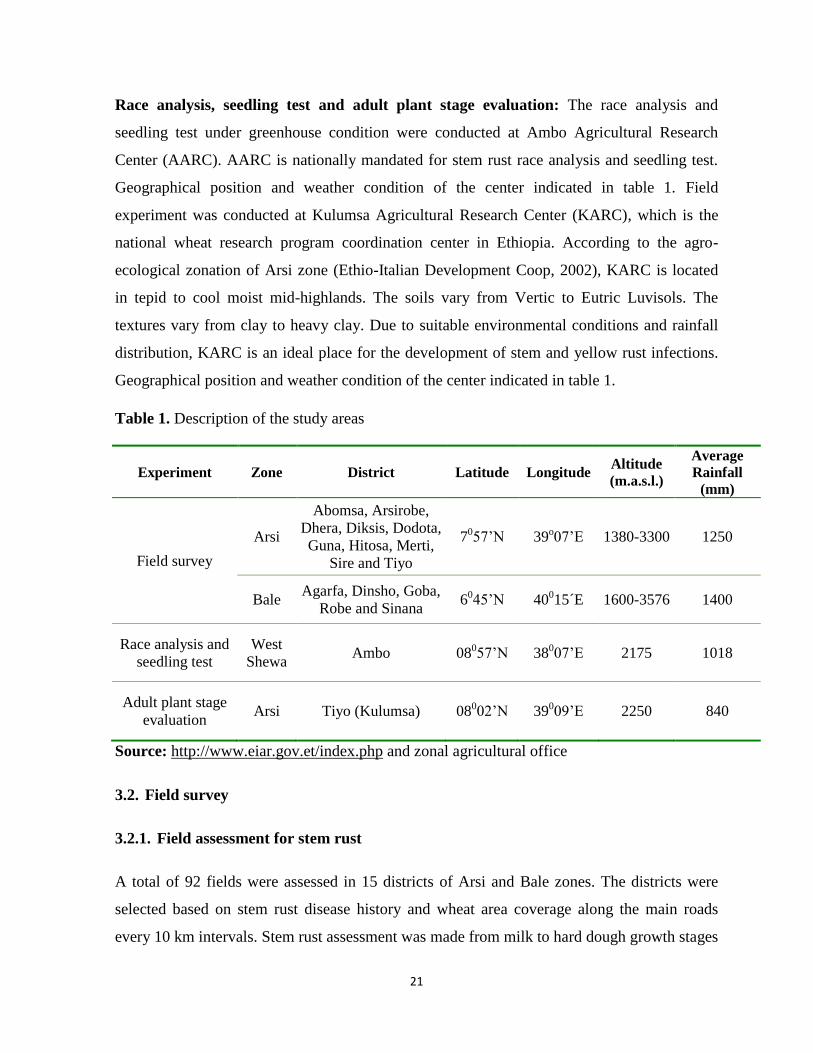
21
Race analysis, seedling test and adult plant stage evaluation: The race analysis and
seedling test under greenhouse condition were conducted at Ambo Agricultural Research
Center (AARC). AARC is nationally mandated for stem rust race analysis and seedling test.
Geographical position and weather condition of the center indicated in table 1. Field
experiment was conducted at Kulumsa Agricultural Research Center (KARC), which is the
national wheat research program coordination center in Ethiopia. According to the agro-
ecological zonation of Arsi zone (Ethio-Italian Development Coop, 2002), KARC is located
in tepid to cool moist mid-highlands. The soils vary from Vertic to Eutric Luvisols. The
textures vary from clay to heavy clay. Due to suitable environmental conditions and rainfall
distribution, KARC is an ideal place for the development of stem and yellow rust infections.
Geographical position and weather condition of the center indicated in table 1.
Table 1. Description of the study areas
Experiment Zone District Latitude Longitude Altitude
(m.a.s.l.)
Average
Rainfall
(mm)
Field survey
Arsi
Abomsa, Arsirobe,
Dhera, Diksis, Dodota,
Guna, Hitosa, Merti,
Sire and Tiyo
7057‟N 39o07‟E 1380-3300 1250
Bale Agarfa, Dinsho, Goba,
Robe and Sinana 6045‟N 40015´E 1600-3576 1400
Race analysis and
seedling test
West
Shewa Ambo 08057‟N 38007‟E 2175 1018
Adult plant stage
evaluation Arsi Tiyo (Kulumsa) 08002‟N 39009‟E 2250 840
Source: http://www.eiar.gov.et/index.php and zonal agricultural office
3.2. Field survey
3.2.1. Field assessment for stem rust
A total of 92 fields were assessed in 15 districts of Arsi and Bale zones. The districts were
selected based on stem rust disease history and wheat area coverage along the main roads
every 10 km intervals. Stem rust assessment was made from milk to hard dough growth stages

22
of the crop per field, along the two diagonals (in an “X” pattern) using 0.5m x 0.5m (0.25 m2)
quadrant and used to calculate average values of the field at five points.
The disease prevalance of stem rust under field condition was recorded percentage of leaf or
stem area covered by rust disease following the Modified Cobb‟s scale as developed by
Peterson et al., 1948. The prevalence of stem rust disease was calculated as:
Disease prevalence (%) = Number of infected fields x 100
Total number of field assessed
In each field, wheat plants within the quadrate were counted and recorded as diseased/infected
and healthy/non-infected and disease incidence was calculated. The incidence of stem rust
was calculated using the following formula;
Disease incidence (%) = Number of diseased plants x 100
Total number of plants in quadrants
The disease severity was measured as a percentage of stem/leaf area covered by rust disease
according to Modified Cobb‟s scale as developed by Peterson et al., (1948). The severity of
the disease was examined randomly by selecting 15 plants from a single quadrante and five
quadrante were used for the estimation of disease severity from a single wheat field.
Disease severity (%) = Area of plant tissue affected x 100
Total plant tissue area
In addition, data on geographical information (latitude, longitude and elevation) of each field
was recorded using GPS (e Trex LegendPS system, Garmin). Crop growth stage was
determind based on the decimalized key developed by Zadoks et al., 1974.
3.2.2. Collection of stem rust samples
A total of 40 stem rust samples were collected from randomly selected farmers' fields and
experimental plots at KARC. Stems of wheat plants infected with stem rust pustules were cut
into small pieces of 5-10 cm using scissors and placed in paper bags after the leaf sheath was
separated from the stem in order to keep stem and/or leaf sheath dry. The samples collected in
the paper bags were tagged with the name of the zone, district, variety, collector names and

23
date of collection. The samples within the paper bags were air dried and kept in refrigerator at
4 0
C until the survey in all districts were completed.
3.2.3. Stem rust race analysis
During field serveys, 40 wheat stem rust samples were collected from infested fields for race
analyisis. Detail procedures for race analysis startng from single pustule isolation upto race
designation described below.
Isolation and multiplication of single-pustules: The inoculum was multiplied and
maintained on standard rust susceptible variety “McNair” which does not carry any stem rust
resistant genes (Roelfs et al., 1992). Five seedlings of McNair for each sample were raised in
suitable 8 cm diameter clay pots that were filled with a mixture of steam sterilized soil, sand
and manure in the ratio of 2:1:1, respectively. Seven day old seedlings or when the primary
leaves were fully expanded and the second leaves beginning to grow, the leaves were rubbed
gently with clean (disinfected with 97% alcohol) moistened (with distilled water) fingers.
Greenhouse inoculations were carried out following procedures developed by Stakman et al.,
1962. The uredio spores of the stem rust were collected from 40 diseased wheat plant sample
by using motorized spore collector in a capsule container and diluted by using lightweight
mineral oil (SolTrol 130) chemicals. Then the spore mix were sprayed in to “McNair”
seedlings from a distance with clean motorized stem rust inoculator. For incubation,
inoculated plants were moistened with fine droplets of distilled water using atomizer after 20
minutes of inoculation and placed in dew chamber for 18 hrs dark period at 18-22 0C followed
by exposure to light for 4 hrs to provide favorable condition for stem rust infection. Seedlings
were allowed to dry/remove their dew/moisture for about 3-4 hrs. Following this, the
seedlings were transferred from dew chamber to glass compartments in the greenhouse where
conditions were regulated at 12 hrs photoperiod, at temperature range of 18-25 0C and relative
humidity (RH) of 60-70%.
After seven days of inoculation (when the flecks/symptoms was clearly visible) leaves
containing single fleck that produce single pustule was selected from the base of the leaves
and the remaining seedlings within the pots were eliminated using hand scissors. Only 2-3
leaves which contain single pustule were left and each of them was covered with cellophane

24
bag (145 × 235 mm) and tied up at the base with a rubber band to avoid cross contamination
(Fetch and Dunsmore, 2004). After two weeks of inoculation (when the monopustule was
well developed) each monopustule was sucked using electric power operated machine
(vacuum pump) and collected in capsule container separately. A suspension, prepared by
mixing urediospores of the monopustule in light weight mineral oil, was inoculated on seven-
day-old seedlings of the susceptible variety “McNair” for multiplication purpose on separate
pots. Soon after inoculation, the seedlings were placed in a humid chamber in dark condition
and transferred to a green house following the earlier procedure mentioned above. After
inoculation of 15 days, the spores of each monopustule/isolate were collected in separate test
tubes and stored at 4 0C until they were inoculated on the standard differential lines. This
procedure was repeated till sufficient amount of spores are produced in order to inoculate the
stem rust differential lines. By following this procedure a total of 32 monopustules/isolates
were developed from 40 wheat stem rust samples.
Inoculation of isolates with host differential lines: Five seeds from each of the 20 stem rust
wheat differential lines including the susceptible variety (McNair) were grown in 4 cm
diameter pots separately in the growth chamber (Table 2). The susceptible variety was used to
determine the viability of spores inoculated on the differential hosts and as a reference
suceptable variety. The single pustule spores/ isolate/ mixed with lightweight mineral oil
(approximately 4 mg of spores per 1 ml) was sprayed/inoculated on seven-day-old seedlings
of each line. Inoculation, incubation and greenhouse condition were applied similar to the
procedure mentioned above.
The natural day light was supplemented with additional 4 hrs/day that emitted by cool white
fluorescent tubes arranged directly above the plants in the greenhouse. Stem rust infection
types were scored 14 days after inoculation using the 0-4 scale of Stakman et al., 1962.
Infection types were grouped in to two, where, Low (resistance) = incompatibility (infection
phenotype 0, 0; (fleck), 1, 2, and 2+) and High (susceptible) = compatibility (infection
phenotype, 3-, 3+ and 4) (Fig 3).

25
Source: Stakman et al., 1962
Figure 3. Infection types of P. graminis f. sp. tritici stem rust and host response
Table 2. List of wheat stem rust differential lines used for race analysis and nomiclture
No Differential host lines Stem rust
genes Pedigree
1 LcSr24Ag 24 Little Club/Agent (Cl 13523)
2 W2691SrTt-1 36 Cl12632 T. timopheevii
3 ISr7b-Ra 7b Hope/Chinese Spring
4 ISr8a-Ra 8a Rieti/Wilhelmina//Akagomughi
5 CnSSrTmp Tmp Triumph 64(Cl 13679)/ Chinese Spring
6 Sr31(Benno)/6*LMPG 31 Kavkaz
7 CnS-T-.mono-deriv 21 Einkorn Cl 2433
8 Trident 38 Spear*4/VPM (Pl519303)
9 ISr9a-Ra 9a Red Egyptian/Chinese Spring
10 ISr9d-Ra 9d Hope/Chinese Spring
11 Combination VII 17 Esp 518/9
12 ISr5-Ra 5 Thatcher/Chinese Spring
13 ISr6-Ra 6 Red Egyptian/Chinese Spring
14 W2691Sr9b 9b Kenya 117A
15 Vernsteine 9e Little Club//3*Gabo/2*
16 W2691Sr10 10 Marquis*4/Egypt NA95/2/2*W2691
17 BtSr30Wst 30 Festival/Uruguay C10837
18 CnsSr9g 9g Selection from Kubanka (Cl1516)
19 ISr11-Ra 11 Kenya C6402/Pusa4/Dundee
20 McNair 701 McN Cl 15288
Note: Except Sr38 and SrMcN (Winter type) all lines are spring type
Source: Roelfs and Martens (1988) and Jin et al. (2008)
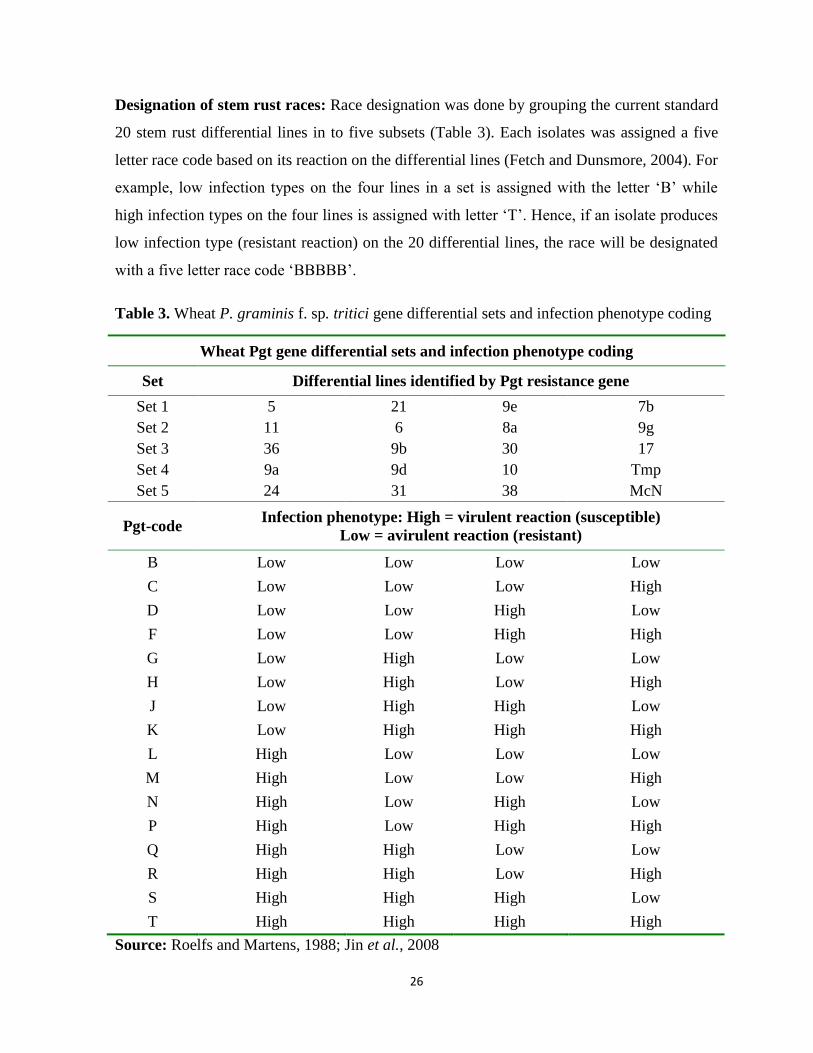
26
Designation of stem rust races: Race designation was done by grouping the current standard
20 stem rust differential lines in to five subsets (Table 3). Each isolates was assigned a five
letter race code based on its reaction on the differential lines (Fetch and Dunsmore, 2004). For
example, low infection types on the four lines in a set is assigned with the letter „B‟ while
high infection types on the four lines is assigned with letter „T‟. Hence, if an isolate produces
low infection type (resistant reaction) on the 20 differential lines, the race will be designated
with a five letter race code „BBBBB‟.
Table 3. Wheat P. graminis f. sp. tritici gene differential sets and infection phenotype coding
Wheat Pgt gene differential sets and infection phenotype coding
Set Differential lines identified by Pgt resistance gene
Set 1 5 21 9e 7b
Set 2 11 6 8a 9g
Set 3 36 9b 30 17
Set 4 9a 9d 10 Tmp
Set 5 24 31 38 McN
Pgt-code Infection phenotype: High = virulent reaction (susceptible)
Low = avirulent reaction (resistant)
B Low Low Low Low
C Low Low Low High
D Low Low High Low
F Low Low High High
G Low High Low Low
H Low High Low High
J Low High High Low
K Low High High High
L High Low Low Low
M High Low Low High
N High Low High Low
P High Low High High
Q High High Low Low
R High High Low High
S High High High Low
T High High High High
Source: Roelfs and Martens, 1988; Jin et al., 2008
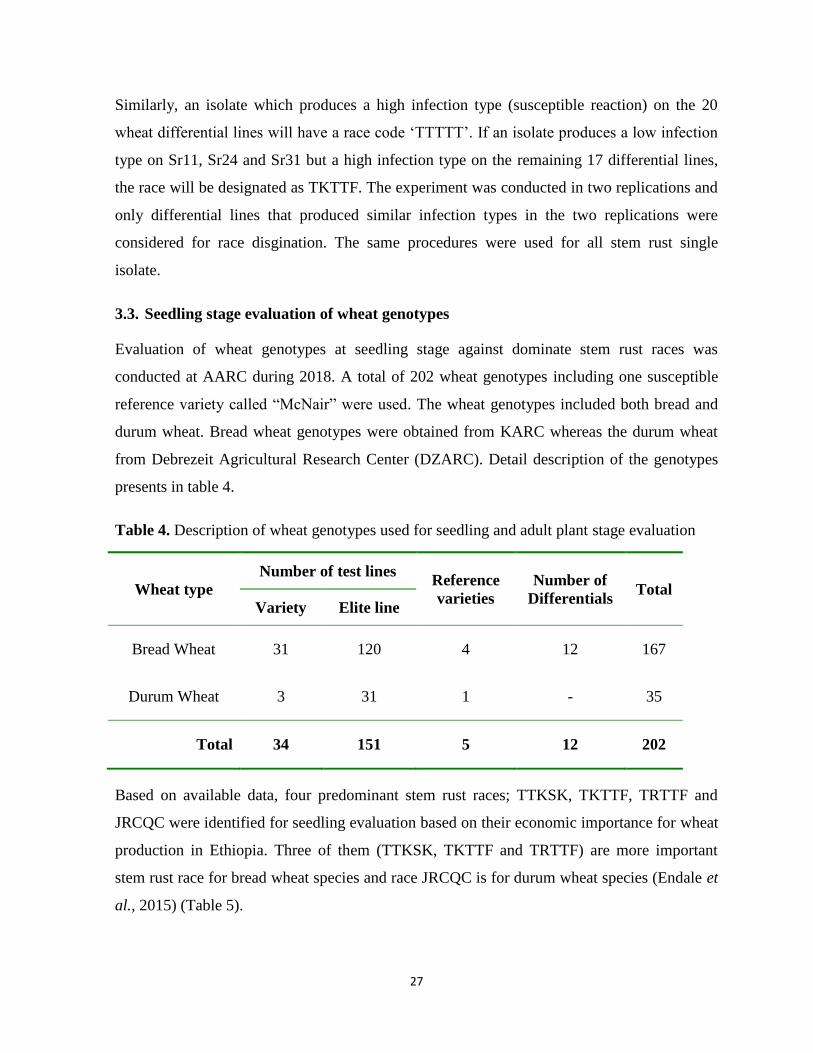
27
Similarly, an isolate which produces a high infection type (susceptible reaction) on the 20
wheat differential lines will have a race code „TTTTT‟. If an isolate produces a low infection
type on Sr11, Sr24 and Sr31 but a high infection type on the remaining 17 differential lines,
the race will be designated as TKTTF. The experiment was conducted in two replications and
only differential lines that produced similar infection types in the two replications were
considered for race disgination. The same procedures were used for all stem rust single
isolate.
3.3. Seedling stage evaluation of wheat genotypes
Evaluation of wheat genotypes at seedling stage against dominate stem rust races was
conducted at AARC during 2018. A total of 202 wheat genotypes including one susceptible
reference variety called “McNair” were used. The wheat genotypes included both bread and
durum wheat. Bread wheat genotypes were obtained from KARC whereas the durum wheat
from Debrezeit Agricultural Research Center (DZARC). Detail description of the genotypes
presents in table 4.
Table 4. Description of wheat genotypes used for seedling and adult plant stage evaluation
Wheat type
Number of test lines Reference
varieties
Number of
Differentials Total
Variety Elite line
Bread Wheat 31 120 4 12 167
Durum Wheat 3 31 1 - 35
Total 34 151 5 12 202
Based on available data, four predominant stem rust races; TTKSK, TKTTF, TRTTF and
JRCQC were identified for seedling evaluation based on their economic importance for wheat
production in Ethiopia. Three of them (TTKSK, TKTTF and TRTTF) are more important
stem rust race for bread wheat species and race JRCQC is for durum wheat species (Endale et
al., 2015) (Table 5).

28
Table 5. Description of single stem rust races used for seedling and adult plant evaluation
Race Origin Avirulence Virulence
TTKSK Uganda Sr24, 36, Tmp Sr5, 6, 7b, 8a, 9a, 9b, 9d, 9e, 9g, 10, 11, 17, 21, 30, 31,
38, McN
TKTTF Ethiopia Sr 11, 24, 31 Sr5, 6, 7b, 8a, 9a, 9b, 9d, 9e, 9g, 10, 17, 21, 30, 36, 38,,
Tmp and McN
TRTTF Yemen Sr8a, 24, 31 Sr5, 6, 7b, 9a, 9b, 9d, 9e, 9g, 10, 11, 17, 21, 30, 36, 38,
McN
JRCQC Ethiopia Sr5, 7b, 8a, 36, 9b,
30, Tmp, 24, 31, 38 Sr21, 9e, 11, 6, 9g, 17, 9a, 9d, McN
Source: Tesfaye, 2018
Race TTKSK (Ug99) has a wide virulence spectrum and is rapidly evolving in East Africa
(Singh et al., 2006). This race was dominant and widely distributed in most wheat growing
agroecologies of Ethiopia. The most virulent and new race, TKTTF (known as the “Digalu”
race in Ethiopia) has been identified as the causal race for the 2013/14 and 2014/15 outbreaks
(Olivera et al., 2015). Races TRTTF and JRCQC, both are present in Ethiopia, possess a
virulence combination that overcomes both the resistance genes Sr13 and Sr9e, two genes
present at high frequency in durum wheat (Klindworth et al., 2007).
Five seeds of each genotype were planted in a pot (4 cm diameter) separately using CRD
(Completely Randomized Design) with two replications. The seeds were pre-germinated on
filter paper in petri dish, and after the third day, the germinated seeds were planted in pots
filled by compost, sterilized soil and sand at a ratio of 1:2:1. Then seven-days-old seedlings
with the first leaf fully expanded and the second leaf just emerging, were inoculated with
spores of virulent races of TKTTF, TTKSK, TRTTF and JRCQC, separately. For incubation,
inoculated plants were moistened with fine droplets of distilled water by using atomizer and
after 20 minutes of inoculation they were placed in dew chamber for 18 hrs in a dark at 18-22
0C. Upon removal from chamber, seedlings were exposed to 3 hrs of fluorescent light to dry
dew on the leaves. Following this, the seedlings were transferred to the greenhouse where
conditions were regulated at 12 hrs photoperiod, at temperature range of 18-25 0
C and RH of
60-70% (Woldeab et al., 2017).

29
(A) (B)
(C) (D)
Figure 4: Schematic overview of the protocols for seedling evaluation of genotypes in the
greenhouse at AARC, Ethiopia; (A) Seven days old seedling, (B) Inoculation of
stem rust race, (C) Seedling in the dew chamber for rust infection establishment
and (D) Susceptible seedling for stem rust
Data on infection types (ITs) were recorded 14 days after inoculation using 0-4 scale based on
figure 3 (Stakman, et al., 1962). ITs readings of 3 (medium-size uredia with/without
chlorosis) and 4 (large uredia without chlorosis or necrosis) were regarded as compatible
reactions. Other readings, that are 0 (immune or fleck), 1 (small uredia with necrosis) and 2
(small to medium uredia with chlorosis or necrosis) were considered as incompatible. The
infection types were defined by modifying characters as follows: -, uredinia somewhat smaller
than normal; +, uredinia somewhat larger than normal for the infection type (Fig 3).
3.4. Adult plant stage evaluation of wheat genotypes
Field evaluation was conducted at KARC and the same genotypes and stem rust races (Table
4 and Table 5) were used. Four field nurseries, namely TTKSK, TKTTF, TRTTF and JRCQC
were established with isolation distances of 300 - 500 m (Table 6). The experiment was laid
out in augmented design with repeated susceptible, moderate and resistant reference varieties.
Each nursery has a total of eight blocks with 31 entries per block except the last block with 32

30
entries. Five wheat varieties „Kubsa‟, „Digalu‟, „Danda‟a‟, „Kingbird‟ and „Arendeto‟ were
used as a reference checks and repeated in all blocks (Table 6).
Table 6. Description of reference genotypes used for field evaluation
Nursery Name Type of Species Year of released Stem rust reaction
Kubsa Bread wheat 1995 Susceptible
Digalu Bread wheat 2005 Susceptible
Danda‟a Bread wheat 2010 Moderately resistance
Kingbird‟ Bread wheat 2015 Resistance
Arendeto‟ Durum wheat 1966 Susceptible
Each nursery was artificially inoculated for the specific stem rust race. Two rows of disease
spreading variety, specific variety susceptible to each specific stem rust race namely,
PBW343, Digalu, Lakech and ST464 was planted perpendicular (between blocks) to test
entries for each nursery (Table 7). Spreader rows were planted two weeks before entries were
planted in each nursery. The distance between spreader rows and spacing between spreader
rows and entries was kept at 0.5m and 0.2m, respectively. Oat and yellow dodder were drilled
around the nurseries to serve as an experimental guard rows border against incoming rust
spores. Testing entries were planted in double rows of 0.5-meter-long and 0.2m spacing
between rows. Weeding and other agronomic practices were applied according to
recommended practices for wheat production in Kulumsa area.
Table 7. Description of single stem rust race nurseries
Nursery Name Race Number of Plot Spreader
Variety
TTKSK-nursery TTKSK (Ug99) 249 PBW343
TKTTF-nursery TKTTF (Digalu Race) 249 Digalu
TRTTF-nursery TRTTF (Lakech Race) 249 Lakech
JRCQC-nursery JRCQC (Durum Race) 249 ST464

31
Single stem rust race inoculation: The spreader rows were inoculated with spores of
particular races; TTKSK, TKTTF, TRTTF and JRCQC. The inoculation was done six times
starting from stem elongation using injection and spraying methods (Roelfs et al., 1992) on
the spreader rows. One injection and five times spary were made at different growth stage of
the nurseries. The first inoculation was made by injection/needle inoculation at tillering
(before stem elongation) stage. Injection mix were prepared by mixing three gram of stem rust
spore, 500 ml of distilled water and two droplete of Tween20. Tween20 was used to make
even distrubution of the spors in the mix. The spray was done after wheat stem elongation or
flowering stage and it was a mix of one gram of specific stem rust race spores for each single
race with four litters of mineral oil by ULV pump sprayer. The sprays were made five times at
seven days interval. Specific sprayer was assigned to each specific race to aviod any
contamination from other races during innoculation. In addition, frequent personnel
movement among single-race nurseries without proper disinfection with alcohol was highly
restricted.
Disease severity and infection response were visually assessed four times starting at the onset
of disease on the entries with 10 days interval. Disease severity was assessed on the 0-100
modified Cobb scale (Peterson et al., 1948) whereas infection reaction was classified into
categories of resistant (R), moderately resistant (MR), moderately susceptible (MS), and
susceptible (S) based on pustule size and chlorosis (Roelfs et al., 1992). When more than one
infection response was observed on the same plot, all infection responses were recorded with
the most frequent infection response listed first.
3.5. Data analysis
Field survey: Survey data (prevalence, incidence and severity) were analyzed using the
descriptive statistical analysis over districts, varieties and crop growth stages.
Seedling stage evaluation: The original ratings, made using the scale from Stakman et al.,
(1962), were converted to a 0-9 linear disease scale for analysis: 0, 1-, 1, 1+, 2-, 2, 2+, 3-, 3,
3+ were coded as 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, respectively (Zhang et al,, 2014). Simple
descriptive statistics and correlation coefficient among stem rust races were analyzed using
SPSS software 2005.

32
Adult plant stage evaluation: Final Rust Severity (FRS), Final Coefficient of Infection
(FCI), Average Coefficient of Infection (ACI) and Area Under Disease Progress Curve
(AUDPC) were used as criterion to identify any possible source of partial resistance to stem
rust. Analysis of variance (ANOVA) was conducted on AUDPC, ACI and TKW in order to
determine resistance and differences among the wheat genotypes. A Pearson correlation
coefficient on FRS, AUDPC, ACI and TKW was done to establish the relationship between
different disease parameters and stem rust races.
The last stem rust severity score (4th
score) was considered as a FRS. CI was calculated by
multiplying the final percentage severity times a constant for host response: where immune =
0.0, R= 0.2, MR= 0.4, MRMS=0.6, MS = 0.8, and S= 1.0. (Stubbs et al., 1986). ACI for each
variety was computed for four observations recorded at an interval of ten days. The
coefficient of infection of each entry per each score was then averaged to give the average
coefficient of infection, ACI (Afzal, et al., 2009).
AUDPC: Stem rust disease severity scores taken at different times were used to calculate the
area under disease progress curve (AUPDC) of each line following Wilcoxson et al., 1975
method.
AUDPC=∑n-1
0.5 (Xi+ Xi+1) (ti+1 - ti)
Where, Xi is the cumulative disease severity expressed as a proportion at the ith
observation; ti
is the time (days after planting) at the ith
observation and n is total number of observations.
Since duration of assessment for each epidemic was different, AUDPC values will be
standardized through dividing the values by the total duration (tn -t1) of the epidemic

33
4. RESULTS AND DISCUSSION
4.1. Status and distribution of wheat stem rust
Wheat stem rust surveys were carried out in Arsi and Bale zones of Oromia Regional State,
Ethiopia, during early October to late November 2017 main cropping season in 92 wheat
fields from Arsi zone (ten districts) and Bale zone (five districts). The distribution or
prevalence of stem rust varied from 0.0 (rust free) to 100% (complete infection) of surveyed
fields depending on the varieties grown and agroecologies (Table 8). Stem rust was observed
in 63% wheat fields assessed, of which Goba and Agarfa from Bale zone and Tiyo from Arsi
zone were the major districts in which the disease was prevalent and its intensity was
relatively higher (Table 8). In contrast, the disease was not occurred in Dinsho (Bale zone)
and Abomsa, Diksis, Dodota, Guna, Merti and Sire district from Arsi zone.
Table 8. Wheat stem rust incidence and severity status in Arsi and Bale zones, 2017
Zone District
Altitude
range
(m.a.s.l.)
Assesse
d field
Infeste
d fields
Prevalence
(%)
Incidence
(%)
Severity
(%) Reaction
Arsi
Abomsa 1766-1831 2 0 0 0 0 0
Arsirobe 2425-2469 4 2 50 0-5 5 MR-MSS
Dhera 1631 1 1 100 10 5 MR
Diksis 2685-2750 3 0 0 0 0 0
Dodota 16321688 3 0 0 0 0 0
Guna 2387 1 0 0 0 0 0
Hitosa 1851-2141 4 3 75 0-15 5 MR - MRMS
Merti 1957-2019 2 0 0 0 0 0
Sire 1695-2356 7 0 0 0 0 0
Tiyo 2150-2250 19 18 94 0-100 5 - 60 MSS - S
Sub total 46 24 52
Bale
Agarfa 2346-2471 10 8 80 0-100 0-40 0-S
Dinsho 2731-2849 2 0 0 0 0 0
Goba 2448-2555 9 9 100 5-100 1-30 S
Robe 2450-2497 3 2 66 0-5 0-1 S
Sinana 2349-2462 22 15 68 0-50 0-40 S
Sub total 46 34 73
Over all Total 92 58 63.0
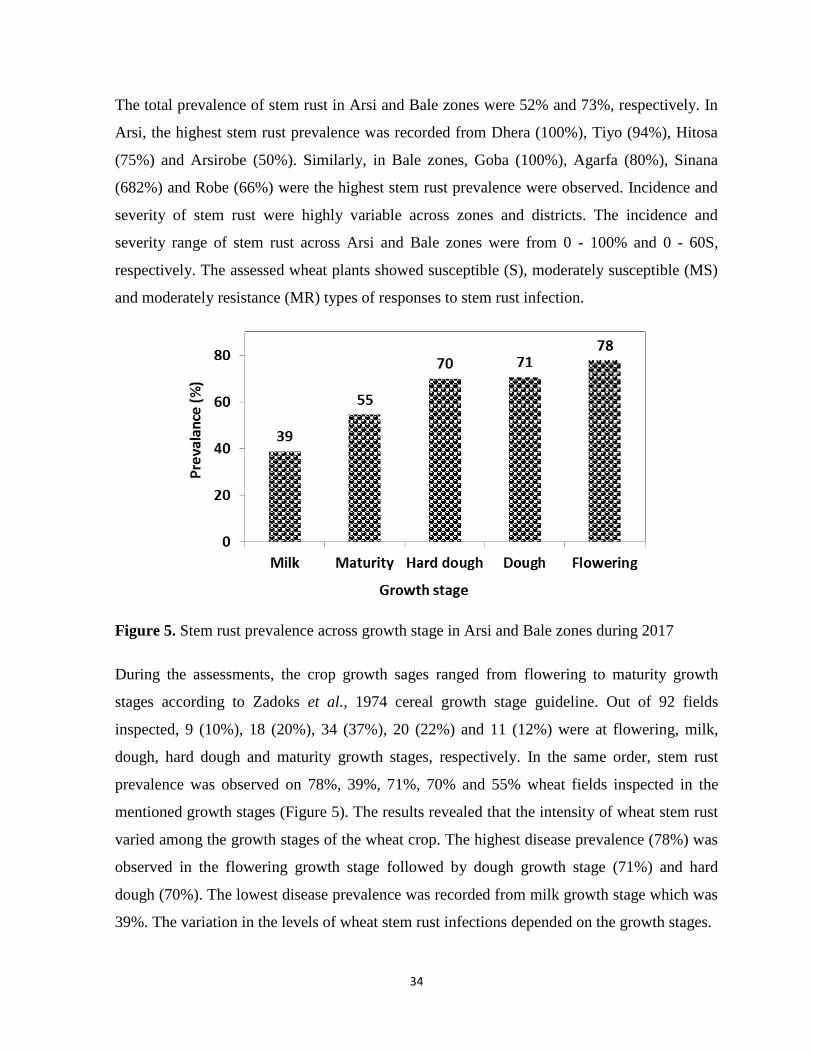
34
The total prevalence of stem rust in Arsi and Bale zones were 52% and 73%, respectively. In
Arsi, the highest stem rust prevalence was recorded from Dhera (100%), Tiyo (94%), Hitosa
(75%) and Arsirobe (50%). Similarly, in Bale zones, Goba (100%), Agarfa (80%), Sinana
(682%) and Robe (66%) were the highest stem rust prevalence were observed. Incidence and
severity of stem rust were highly variable across zones and districts. The incidence and
severity range of stem rust across Arsi and Bale zones were from 0 - 100% and 0 - 60S,
respectively. The assessed wheat plants showed susceptible (S), moderately susceptible (MS)
and moderately resistance (MR) types of responses to stem rust infection.
Figure 5. Stem rust prevalence across growth stage in Arsi and Bale zones during 2017
During the assessments, the crop growth sages ranged from flowering to maturity growth
stages according to Zadoks et al., 1974 cereal growth stage guideline. Out of 92 fields
inspected, 9 (10%), 18 (20%), 34 (37%), 20 (22%) and 11 (12%) were at flowering, milk,
dough, hard dough and maturity growth stages, respectively. In the same order, stem rust
prevalence was observed on 78%, 39%, 71%, 70% and 55% wheat fields inspected in the
mentioned growth stages (Figure 5). The results revealed that the intensity of wheat stem rust
varied among the growth stages of the wheat crop. The highest disease prevalence (78%) was
observed in the flowering growth stage followed by dough growth stage (71%) and hard
dough (70%). The lowest disease prevalence was recorded from milk growth stage which was
39%. The variation in the levels of wheat stem rust infections depended on the growth stages.

35
From this study, the prevalence of stem rust was not higher as compared to the previous
reports. According to Mideksa et al., (2018), temperature (minimum, maximum and average);
rainfall and relative humidity interacted simultaneously in nature and contributed collectively
to make the environment favorable for the stem rust development, which showed the
existence of multicollinearity among them. If one of these weather factors missed, disease will
not be developed. However, if disease develops, its severity fluctuates with changing
environmental conditions. Eventually, environmental conditions, amount of inoculum, host
susceptibility, host physiological growth stage and timing of the epidemic are all factor that
affects the degree of damage significantly (Duveiller, 2007). This would be one of the reasons
for low stem rust prevalence in the surveyed areas of Arsi and Bale zones in 2017 main
cropping season.
Figure 6. Disease prevalence across varieties in Arsi and Bale zones during 2017
Wide range of wheat varieties were recorded during the survey. Farmers in the assessed areas
were engaged in both bread and durum wheat production. However, eighty-nine of the
assessed fields were bread wheat and only three fields were durum wheat (Alemtena, Tesfaye
and Ude) (Table 9). This indicated that bread wheat is widely cultivated in the wheat belt
areas of Ethiopia in Arsi and Bale zones. The diffusion of durum wheat varieties was small
mainly due to problems associated with marketing rather than rust disease incidences (Sherif
et al., 2016).
36
Table 9. Incidence and severity of wheat varieties assessed in Arsi and Bale zones, 2017
Variety name Total number of
field assessed
Variety
response at fields
Range
Incidence
(%)
Severity
(%)
Alemtene 1 S 100 40
Batu 2 0 0 0
Danda'a 9 MSS 30-40 15-20
Dashen 1 MR 15 5
Digalu 4 S 50-70 20-40
Galama 1 0 0 0
Hidasse 12 MR-S 5-100 0-30
Hulluka 3 S 0-5 0-1
K6295-4A 3 0 0 0
Kakaba 1 S 5 1
Kubsa 5 S 0-40 0-30
Medewolabu 5 S 0-50 0-10
Ogolcho 21 MR-S 0-50 0-30
Pavon-76 1 0 0 0
Sanete 1 S 25 40
Tesfaye 1 S 100 20
Ude 1 S 5 1
Breeding Lines (Bread wheat) 12 MR-S 30-100 15-60
Unknown (Bread wheat) 8 S 0-5 0-1
Since the inception of wheat improvement in Ethiopia, close to 100 bread wheat varieties
have been released for commercial production. However, the majority was abandoned mainly
due to their susceptibility to new races of yellow and/or stem rusts and very few varieties are
currently under production (Source: Ministry of Ariculture variety registration book).
From this study, fourteen bread wheat varieties were recorded from the assessed fields of Arsi
and Bale zones (Dashen, Galama, Pavon-76, Kakaba, Sanete, Batu, K6295-4A, Hulluka,
Digalu, Kubsa, Medewolabu, Danda'a, Hidasse, Ogolcho) (Table9). In some fields, it was
difficult to differentiate the name of the varieties and the data is present as unknown variety.
But, all those unknown varieties were bread wheat based on their morphological
characteristics. In addition, some promising lines were surveyed from KARC breeding plots.
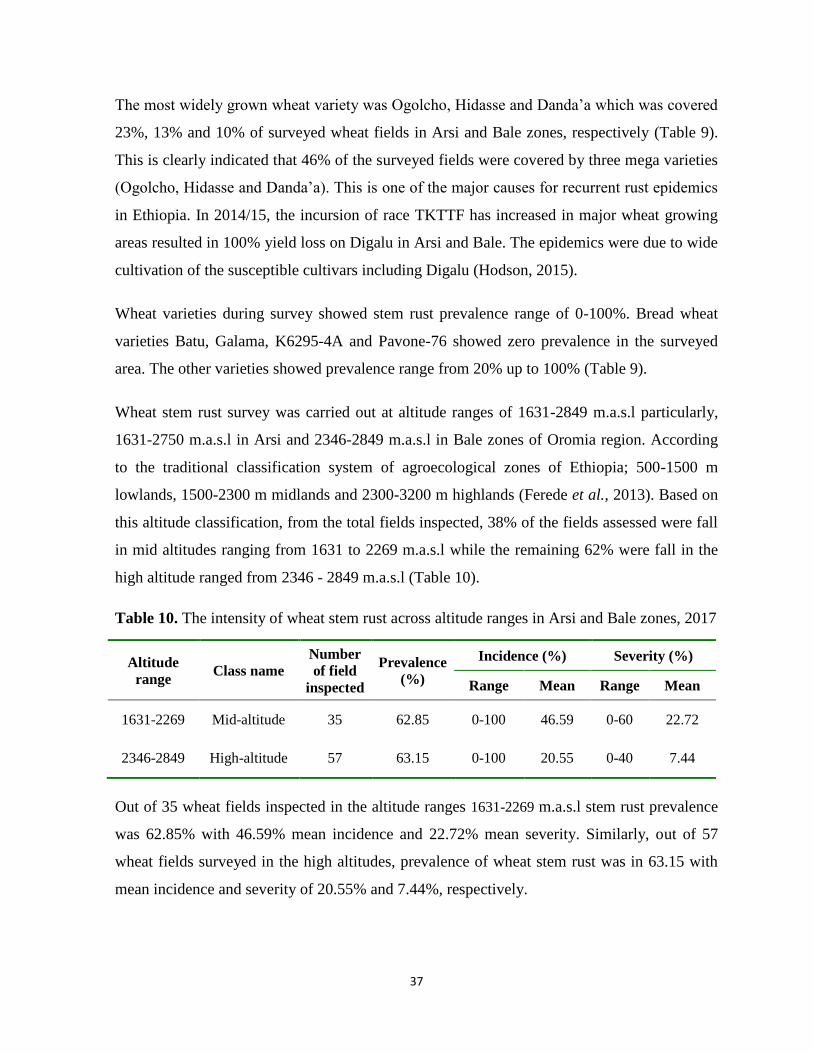
37
The most widely grown wheat variety was Ogolcho, Hidasse and Danda‟a which was covered
23%, 13% and 10% of surveyed wheat fields in Arsi and Bale zones, respectively (Table 9).
This is clearly indicated that 46% of the surveyed fields were covered by three mega varieties
(Ogolcho, Hidasse and Danda‟a). This is one of the major causes for recurrent rust epidemics
in Ethiopia. In 2014/15, the incursion of race TKTTF has increased in major wheat growing
areas resulted in 100% yield loss on Digalu in Arsi and Bale. The epidemics were due to wide
cultivation of the susceptible cultivars including Digalu (Hodson, 2015).
Wheat varieties during survey showed stem rust prevalence range of 0-100%. Bread wheat
varieties Batu, Galama, K6295-4A and Pavone-76 showed zero prevalence in the surveyed
area. The other varieties showed prevalence range from 20% up to 100% (Table 9).
Wheat stem rust survey was carried out at altitude ranges of 1631-2849 m.a.s.l particularly,
1631-2750 m.a.s.l in Arsi and 2346-2849 m.a.s.l in Bale zones of Oromia region. According
to the traditional classification system of agroecological zones of Ethiopia; 500-1500 m
lowlands, 1500-2300 m midlands and 2300-3200 m highlands (Ferede et al., 2013). Based on
this altitude classification, from the total fields inspected, 38% of the fields assessed were fall
in mid altitudes ranging from 1631 to 2269 m.a.s.l while the remaining 62% were fall in the
high altitude ranged from 2346 - 2849 m.a.s.l (Table 10).
Table 10. The intensity of wheat stem rust across altitude ranges in Arsi and Bale zones, 2017
Altitude
range Class name
Number
of field
inspected
Prevalence
(%)
Incidence (%) Severity (%)
Range Mean Range Mean
1631-2269 Mid-altitude 35 62.85 0-100 46.59 0-60 22.72
2346-2849 High-altitude 57 63.15 0-100 20.55 0-40 7.44
Out of 35 wheat fields inspected in the altitude ranges 1631-2269 m.a.s.l stem rust prevalence
was 62.85% with 46.59% mean incidence and 22.72% mean severity. Similarly, out of 57
wheat fields surveyed in the high altitudes, prevalence of wheat stem rust was in 63.15 with
mean incidence and severity of 20.55% and 7.44%, respectively.
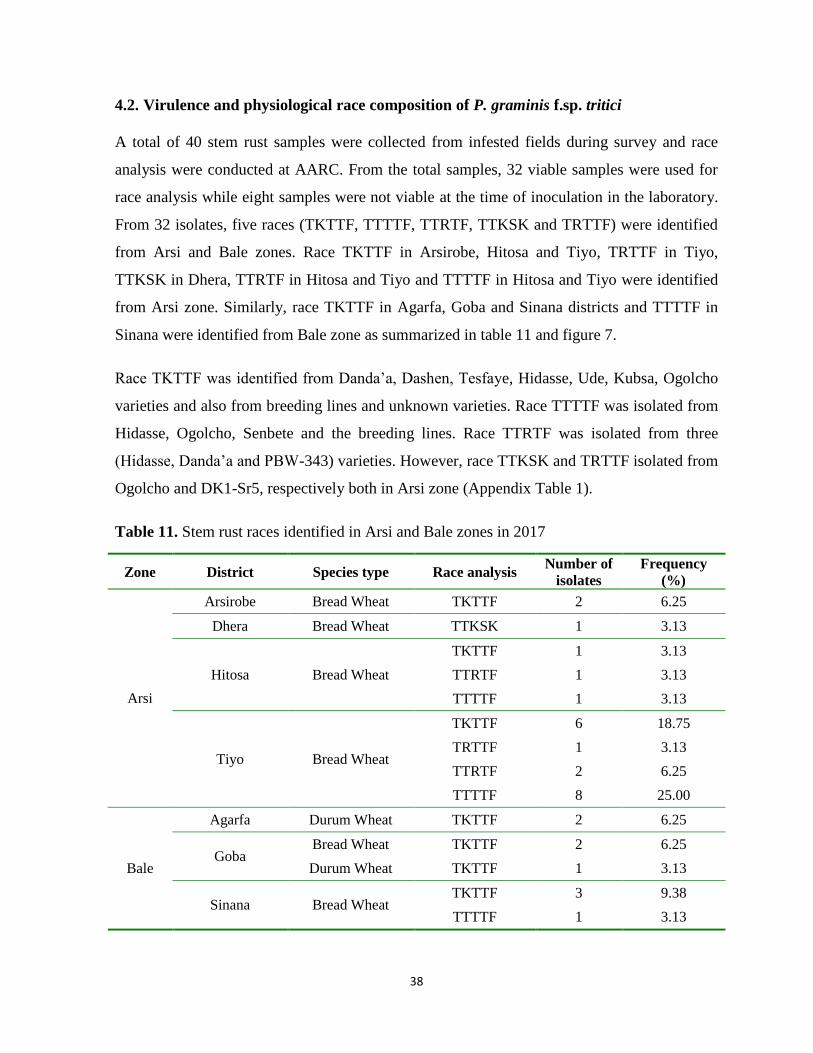
38
4.2. Virulence and physiological race composition of P. graminis f.sp. tritici
A total of 40 stem rust samples were collected from infested fields during survey and race
analysis were conducted at AARC. From the total samples, 32 viable samples were used for
race analysis while eight samples were not viable at the time of inoculation in the laboratory.
From 32 isolates, five races (TKTTF, TTTTF, TTRTF, TTKSK and TRTTF) were identified
from Arsi and Bale zones. Race TKTTF in Arsirobe, Hitosa and Tiyo, TRTTF in Tiyo,
TTKSK in Dhera, TTRTF in Hitosa and Tiyo and TTTTF in Hitosa and Tiyo were identified
from Arsi zone. Similarly, race TKTTF in Agarfa, Goba and Sinana districts and TTTTF in
Sinana were identified from Bale zone as summarized in table 11 and figure 7.
Race TKTTF was identified from Danda‟a, Dashen, Tesfaye, Hidasse, Ude, Kubsa, Ogolcho
varieties and also from breeding lines and unknown varieties. Race TTTTF was isolated from
Hidasse, Ogolcho, Senbete and the breeding lines. Race TTRTF was isolated from three
(Hidasse, Danda‟a and PBW-343) varieties. However, race TTKSK and TRTTF isolated from
Ogolcho and DK1-Sr5, respectively both in Arsi zone (Appendix Table 1).
Table 11. Stem rust races identified in Arsi and Bale zones in 2017
Zone District Species type Race analysis Number of
isolates
Frequency
(%)
Arsi
Arsirobe Bread Wheat TKTTF 2 6.25
Dhera Bread Wheat TTKSK 1 3.13
Hitosa Bread Wheat
TKTTF 1 3.13
TTRTF 1 3.13
TTTTF 1 3.13
Tiyo Bread Wheat
TKTTF 6 18.75
TRTTF 1 3.13
TTRTF 2 6.25
TTTTF 8 25.00
Bale
Agarfa Durum Wheat TKTTF 2 6.25
Goba Bread Wheat TKTTF 2 6.25
Durum Wheat TKTTF 1 3.13
Sinana Bread Wheat TKTTF 3 9.38
TTTTF 1 3.13
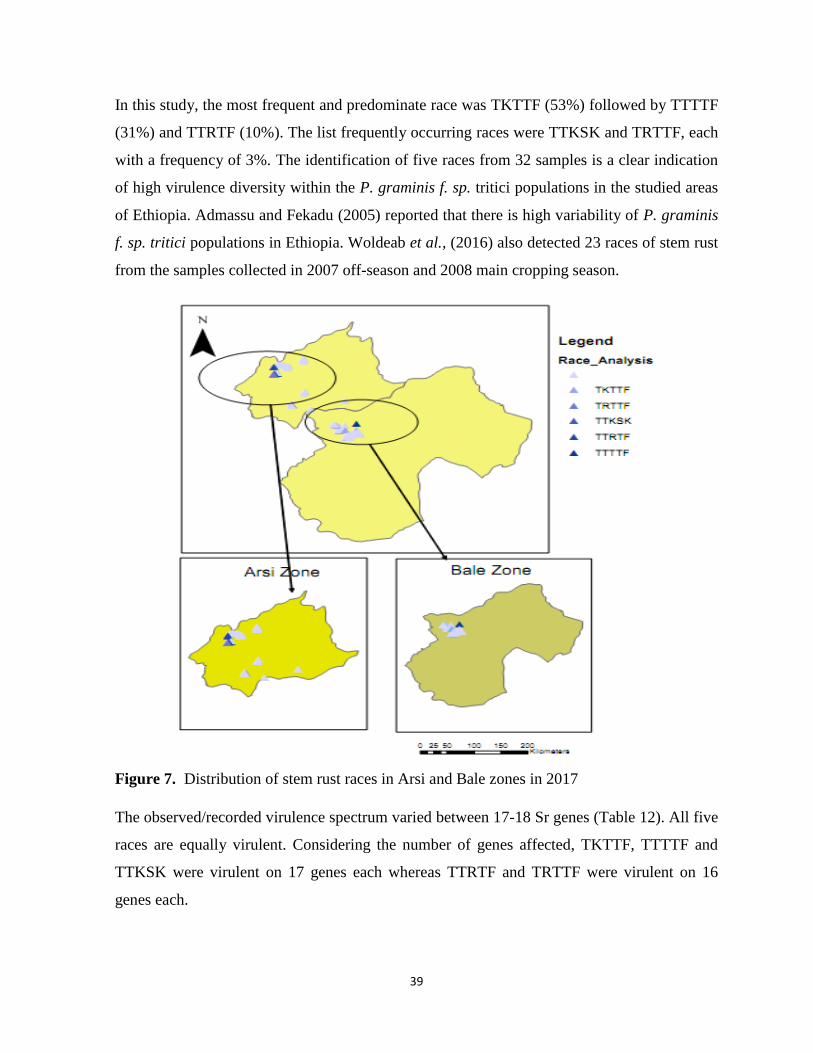
39
In this study, the most frequent and predominate race was TKTTF (53%) followed by TTTTF
(31%) and TTRTF (10%). The list frequently occurring races were TTKSK and TRTTF, each
with a frequency of 3%. The identification of five races from 32 samples is a clear indication
of high virulence diversity within the P. graminis f. sp. tritici populations in the studied areas
of Ethiopia. Admassu and Fekadu (2005) reported that there is high variability of P. graminis
f. sp. tritici populations in Ethiopia. Woldeab et al., (2016) also detected 23 races of stem rust
from the samples collected in 2007 off-season and 2008 main cropping season.
Figure 7. Distribution of stem rust races in Arsi and Bale zones in 2017
The observed/recorded virulence spectrum varied between 17-18 Sr genes (Table 12). All five
races are equally virulent. Considering the number of genes affected, TKTTF, TTTTF and
TTKSK were virulent on 17 genes each whereas TTRTF and TRTTF were virulent on 16
genes each.

40
Table 12. Virulence/Avirulence spectrum of races identified from Arsi and Bale zones, 2017
Identified
Race
Sr
Avirulence Sr Virulence
TKTTF Sr11,24,31 Sr5,6,7b,8a,9a,9b,9d,9e,9g,10,17,21,30,36,38,Tmp, McN
TTTTF Sr24,31 Sr5,6,7b,8a,9a,9b,9d,9e,9g,10,11,17,21,30,36,38, Tmp, McN
TTRTF Sr30,24,31 Sr5,6,7b,8a,9a,9b,9d,9e,9g,10,11,17,21,36,38, Tmp, McN
TTKSK Sr24,36,Tmp Sr5,6,7b,8a,9a,9b,9d,9e,9g,10,11,17,21,30,31,38,McN
TRTTF Sr8a,24,31 Sr5,6,7b,9a,9b,9d,9e,9g,10,11,17,21,30,36,38, Tmp, McN
Race TKTTF, also known as the “Digalu race”, caused destruction of the popular bread wheat
variety “Digalu” in Ethiopia and led to yield loss up to 100% in 2013-2015 (Olivera et al.,
2015). This epidemic is believed to be due to the narrow genetic background of stem rust
resistance in widely planted Digalu, conferred by the single gene SrTmp, coupled with
environmental conditions favoring stem rust outbreak. After its severe outbreak in 2013, race
TKTTF has become the predominant race in Ethiopia (Olivera et al., 2017). This study also
confirmed that high frequency of TKTTF in the surveyed area. Similar result was reported by
Endale et al., 2015 from West Shewa and South West Shewa zones, Oromia region. This
showed that, TKTTF is still one of the most virulent race on wheat varieties and it has rapidly
spread to a wide range of wheat growing areas. This might be due to favorable environmental
conditions as well as continued cultivation of Digalu and other susceptible wheat varieties in
those areas. TKTTF is genetically different from members of the Ug99 race group and it is
avirulent to stem rust resistance genes Sr11, Sr24 and Sr31 (Olivera et al., 2015). Therefore,
these genes could be deployed in combination with other genes to combat race TKTTF.
Race TTTTF is the second spreading stem rust race from the surveyed areas. Similar study in
East Shewa zone in central Ethiopia, race TTTTF were identified as the dominant races in
major durum wheat producing districts in the zone (Lemma et al., 2014). Race TTTTF is the
most virulent race known in the United States, producing high infection types (ITs) on the
majority of stem rust differential lines (Jin et al., 2007).
Race TTKSK (also known as Ug99) was first identified in Uganda in 1999, to which most
commercial wheat cultivars and breeding lines worldwide are susceptible (Pretorius et al.,
2000; Singh, et al., 2006; 2011). Since 1999, the migration of the Ug99 race has taken place

41
from Uganda, to Kenya in 2001, to Ethiopia in 2003 and has been shown in most of the wheat
production in those areas and shown virulence for the gene Sr31 (Singh et al., 2006). In 2005,
Ethiopian reports confirmed its presence in six dispersed locations and was spread to most of
wheat growing areas in the country and becoming the main threat for wheat production. In
2010 and 2011 seasons, wheat stem rust virulence survey conducted in Ethiopia yielded race
TTKSK as the dominant races (Woldeab et al., 2016). Race TTKSK was the only dominant
race detected from samples collected in the off-season and main seasons of 2012 and 2013 in
various wheat growing agro-ecologies of Ethiopia (Woldeab et al., 2016). The East African
highlands are a known „„hot-spot‟‟ for the evolution and survival of new rust races (Saari and
Prescott, 1985). The favorable environmental conditions and the presence of host plants year-
round favor the survival and buildup of pathogen populations. This study also clearly shows a
dramatic shift of the race statues from TTKSK to other aggressive stem rust race such as
TKTTF and TTTTF (Fig. 8).
Figure 8. Percentage of stem rust races identified in Arsi and Bale zones, 2017
Race TRTTF was also one of stem rust races detected from this survey although at the lowest
frequency. Stem rust samples collected during 2009 from stem rust screening nursery in
Debre Zeit yielded three important races viz., TTKSK, TRTTF and JRCQC (Olivera et al.,
2012) and races TRTTF and JRCQC have combined virulence for stem rust resistance genes
Sr9e and Sr13 (Olivera et al., 2012).
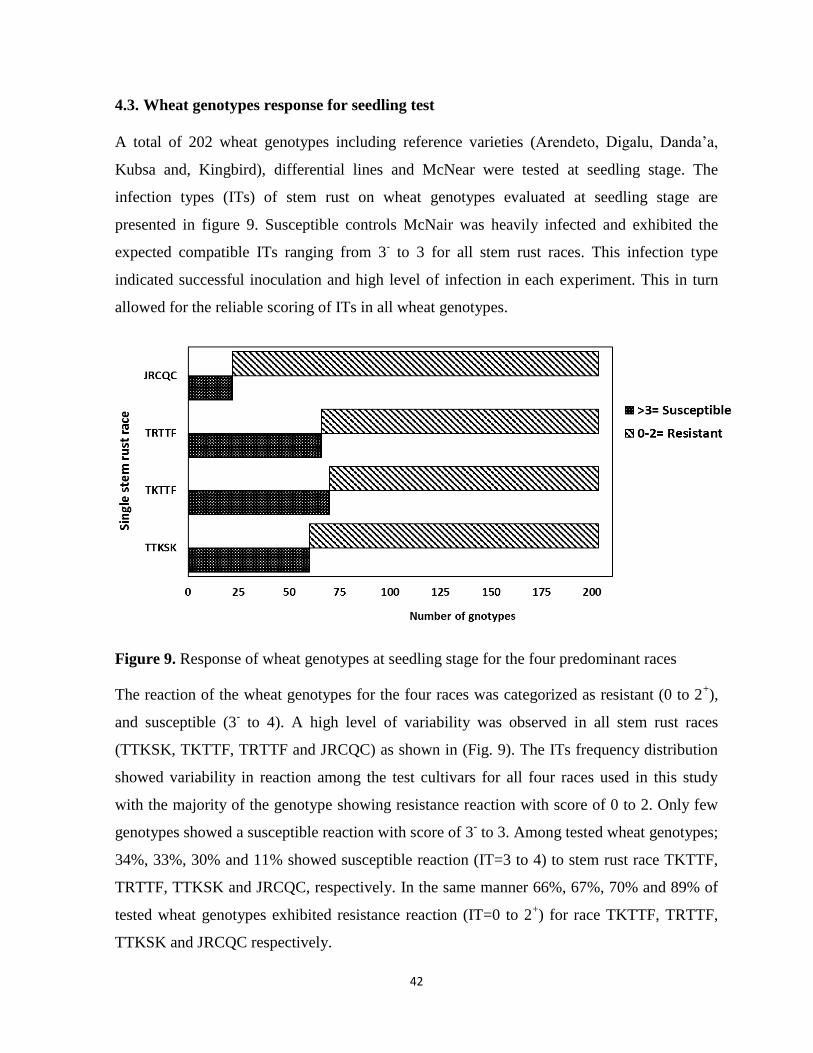
42
4.3. Wheat genotypes response for seedling test
A total of 202 wheat genotypes including reference varieties (Arendeto, Digalu, Danda‟a,
Kubsa and, Kingbird), differential lines and McNear were tested at seedling stage. The
infection types (ITs) of stem rust on wheat genotypes evaluated at seedling stage are
presented in figure 9. Susceptible controls McNair was heavily infected and exhibited the
expected compatible ITs ranging from 3- to 3 for all stem rust races. This infection type
indicated successful inoculation and high level of infection in each experiment. This in turn
allowed for the reliable scoring of ITs in all wheat genotypes.
Figure 9. Response of wheat genotypes at seedling stage for the four predominant races
The reaction of the wheat genotypes for the four races was categorized as resistant (0 to 2+),
and susceptible (3- to 4). A high level of variability was observed in all stem rust races
(TTKSK, TKTTF, TRTTF and JRCQC) as shown in (Fig. 9). The ITs frequency distribution
showed variability in reaction among the test cultivars for all four races used in this study
with the majority of the genotype showing resistance reaction with score of 0 to 2. Only few
genotypes showed a susceptible reaction with score of 3- to 3. Among tested wheat genotypes;
34%, 33%, 30% and 11% showed susceptible reaction (IT=3 to 4) to stem rust race TKTTF,
TRTTF, TTKSK and JRCQC, respectively. In the same manner 66%, 67%, 70% and 89% of
tested wheat genotypes exhibited resistance reaction (IT=0 to 2+) for race TKTTF, TRTTF,
TTKSK and JRCQC respectively.
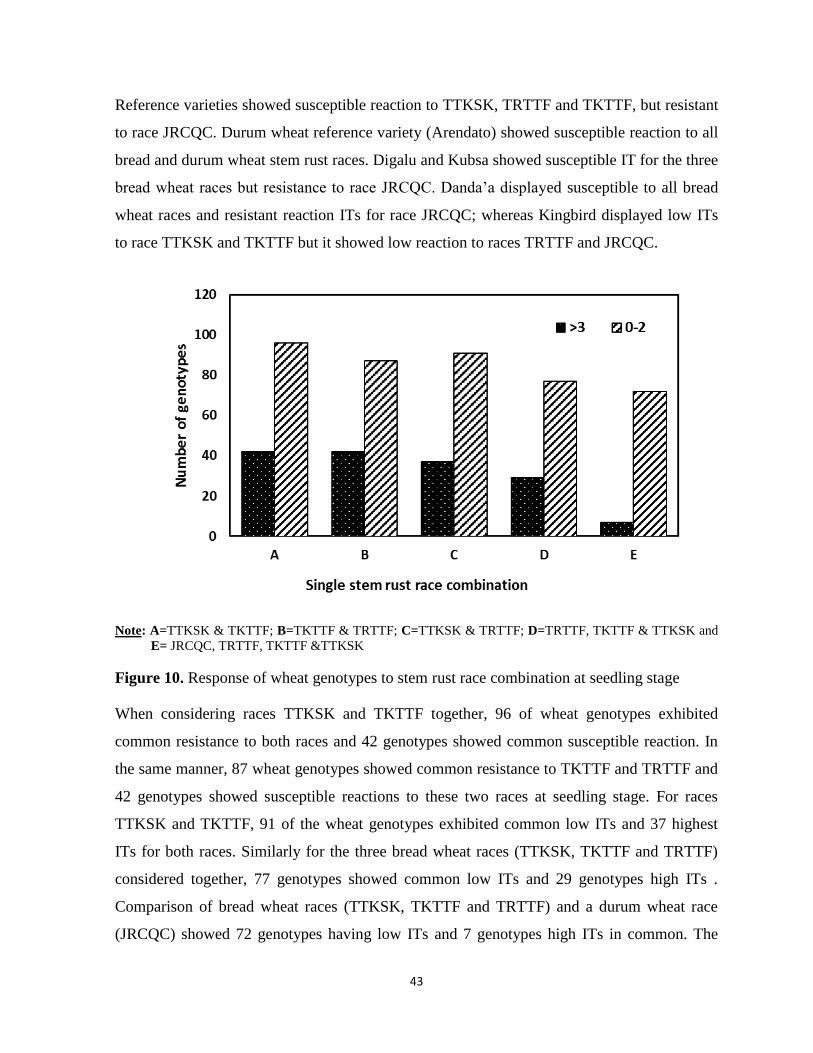
43
Reference varieties showed susceptible reaction to TTKSK, TRTTF and TKTTF, but resistant
to race JRCQC. Durum wheat reference variety (Arendato) showed susceptible reaction to all
bread and durum wheat stem rust races. Digalu and Kubsa showed susceptible IT for the three
bread wheat races but resistance to race JRCQC. Danda‟a displayed susceptible to all bread
wheat races and resistant reaction ITs for race JRCQC; whereas Kingbird displayed low ITs
to race TTKSK and TKTTF but it showed low reaction to races TRTTF and JRCQC.
Note: A=TTKSK & TKTTF; B=TKTTF & TRTTF; C=TTKSK & TRTTF; D=TRTTF, TKTTF & TTKSK and
E= JRCQC, TRTTF, TKTTF &TTKSK
Figure 10. Response of wheat genotypes to stem rust race combination at seedling stage
When considering races TTKSK and TKTTF together, 96 of wheat genotypes exhibited
common resistance to both races and 42 genotypes showed common susceptible reaction. In
the same manner, 87 wheat genotypes showed common resistance to TKTTF and TRTTF and
42 genotypes showed susceptible reactions to these two races at seedling stage. For races
TTKSK and TKTTF, 91 of the wheat genotypes exhibited common low ITs and 37 highest
ITs for both races. Similarly for the three bread wheat races (TTKSK, TKTTF and TRTTF)
considered together, 77 genotypes showed common low ITs and 29 genotypes high ITs .
Comparison of bread wheat races (TTKSK, TKTTF and TRTTF) and a durum wheat race
(JRCQC) showed 72 genotypes having low ITs and 7 genotypes high ITs in common. The

44
current results discovered that the majority of the tested wheat genotypes infection types score
of 0 to 2 (Fig. 10) that is resistance to tested four races at seedling test.
4.4. Wheat genotypes response for adult plant stage evaluation
Field experiments to evaluate wheat genotypes for prevailing stem rust races were carried out
at KARC and showed high disease severity in all stem rust races. Phenotypic variation was
observed for infection types and level of stem rust severity for wheat genotypes. Terminal
score ranged from 0 (immune) to 80% S (highly susceptible) for all stem rust races. The
maximum rust severity was recorded 80%, 70%, 60% and 50% from TRTTF, TTKSK,
JRCQC and TKTTF, respectively. The response of genotypes for specific wheat stem rust
race varied from highly susceptible (S) to zero (no infection) as shown in appendix table 2.
Mean square values of AUDPC, ACI and TKW for the four stem rust races are shown in
appendix table 3. According to mean square value, there was highly significant difference
(P<0.001) between check genotypes for different disease parameters and TKW for each single
race nursery. This indicates that there was a clear difference between susceptible and resistant
check varieties response for all stem rust races. Similarly, highly significant difference
(P<0.001) observed for all disease parameters and TKW of the new genotypes except for
TKW (TKTTF and TRTTF) (Appendix Table 3).
4.4.1. Final rust severity (FRS)
The final rust severity score showed more number of genotypes was grouped under MR and S
types of reaction for all stem rust races (Fig. 11). From field observations 38, 36, 7 and 13
genotypes showed 0-R types of reaction for race TKTTF, TRTTF, TTKSK and JRCQC,
respectively. Similarly 105, 99, 87 and 78 genotypes showed MR-MS type of reaction in the
same order. More number of genotypes (111) was showed susceptible type of reaction (MSS-
S) for durum wheat stem rust race called JRCQC. In the same way, 59 (TKTTF), 67 (TRTTF)
and 108 (TTKSK) genotypes showed susceptible reaction for the races.
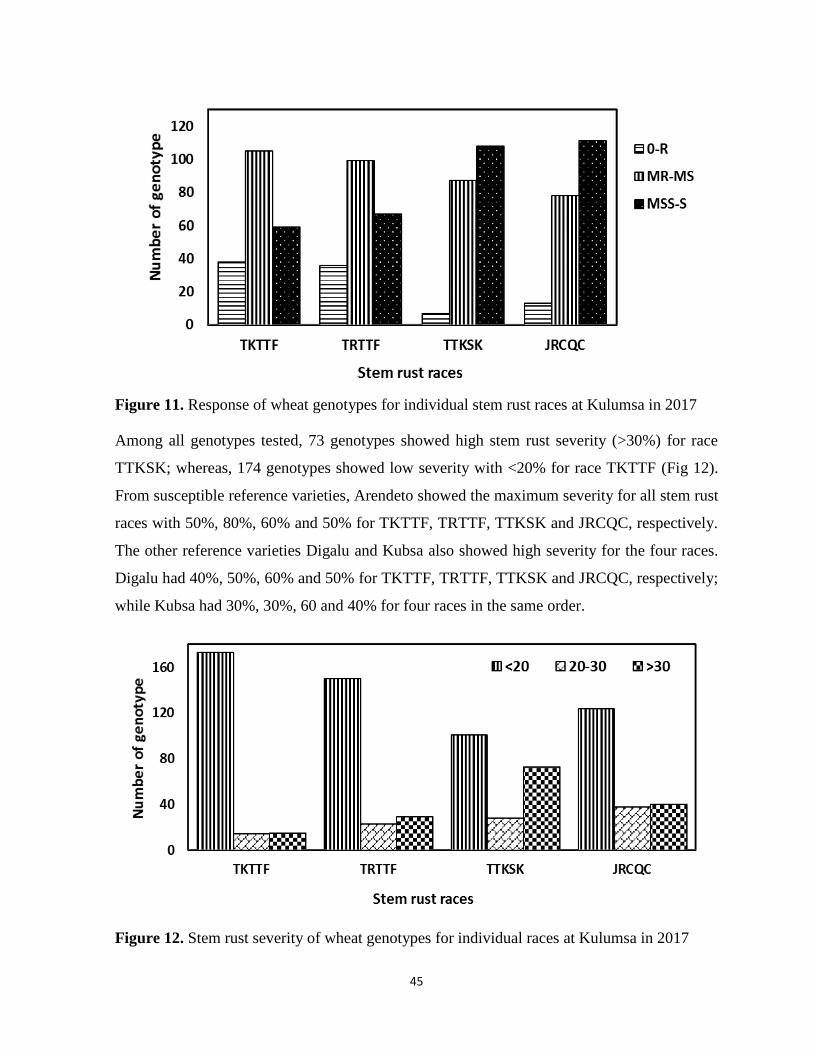
45
Figure 11. Response of wheat genotypes for individual stem rust races at Kulumsa in 2017
Among all genotypes tested, 73 genotypes showed high stem rust severity (>30%) for race
TTKSK; whereas, 174 genotypes showed low severity with <20% for race TKTTF (Fig 12).
From susceptible reference varieties, Arendeto showed the maximum severity for all stem rust
races with 50%, 80%, 60% and 50% for TKTTF, TRTTF, TTKSK and JRCQC, respectively.
The other reference varieties Digalu and Kubsa also showed high severity for the four races.
Digalu had 40%, 50%, 60% and 50% for TKTTF, TRTTF, TTKSK and JRCQC, respectively;
while Kubsa had 30%, 30%, 60 and 40% for four races in the same order.
Figure 12. Stem rust severity of wheat genotypes for individual races at Kulumsa in 2017

46
APR type check Danda‟a showed low severity for race TKTTF (20%) and TRTTF (25%) but
slightly higher for race TTKSK (50%) and JRCQC (40%). The resistant check Kingbird
showed low stem rust severity of 10%, 5%, 10% and 5% for TKTTF, TRTTF, TTKSK and
JRCQC, respectively (Table 14). The results clearly indicated that the expected rust severity
status from the check genotypes vis-a-vis susceptible, APR type and resistant genotypes.
From this study, most of the genotypes showed low to moderate severity (<20) for all stem
rust races and this will give a chance to identify valuable lines for future breeding and
pathological research.
From the seedling and field response studies of genotypes against the four stem rust races, 31
genotypes showed resistance for both seedling and adult-plant stages (Appendix Table 2).
Park (2007) suggested that when genotypes show rust resistance at both seedling and adult-
plant stages, it can be referred to as all-stage resistance. Possibly, these genotypes might have
all-stage resistance to those races. The seedling resistance is not growth stage-dependent
(Park, 2007 and Singh, 2005). However, seedling resistance does not always provide
protection against rust at adult-plant stages. In this study also suggested that genotypes
possessed seedling resistance but failed to protect at adult-plant stage (Appendix Table 2).
According to some works, a genotype with stem rust resistance at seedling stage alone is not
sustainable and effective for a long-term deployment (Park, 2007 and Singh, 2005). Often,
seedling resistance is governed by major gene(s) and frequent mutations in corresponding
avirulence genes in the rust pathogen may lead to catastrophic failure of the crop (Park, 2007).
APR is often characterized by its effectiveness against a wide range of pathogen races and has
been considered more durable, providing resistance without being readily overcome by the
change in pathogen virulence when the cultivar is widely grown in an area where the disease
is prevalent (Parlevliet 1975; McIntosh et al., 1995). From this study, only genotype G42
(ETBW 9464) showed seedling susceptible but adult plant resistant for all bread and durum
wheat stem rust races used in this study. However, four genotypes (G40=ETBW 9045,
G42=ETBW 9464, G175=ETBW 9569, and G196=Alidoro) showed seedling susceptible and
field resistant reaction for the three major bread wheat stem rust races (TKTTF, TRTTF and
TTKSK). The deployment of cultivars carrying APR based on multiple genes is particularly

47
preferred to delay infection, growth and reproduction of the pathogen in adult plants and
circumvent “boom-and-bust” cycles (Chen, 2005; Singh et al., 2005).
4.4.2. Coefficient of infection (Cl)
The data on disease severity and host reaction were combined to calculate CI. According to
Ali et al., (2009), varieties with CI values of 0-20, 21-40, 41-60 were regarded as possessing
high, moderate and low levels of slow rusting resistance, respectively. In this study 174
(86%), 150 (74%), 110 (54%) and 125 (62%) genotypes showed CI values between 0-20 for
races TKTTF, TRTTF, TTKSK, TRTTF and JRCQC, respectively (Fig 13). These are
designated as having a high level of slow rusting. Under moderate levels of slow rusting
resistance, which means CI value between 21 to 40 categories, were 23 (TKTTF), 36
(TRTTF), 43 (TTKSK) and 57 (JRCQC) genotypes. A total of 5, 10, 48 and 20 genotypes
were grouped under low levels of slow rusting resistance categories (CI value 40 - 60) for
race TKTTF, TRTTF, TTKSK and JRCQC, respectively (Appendix Table 2). Different
studies appraised slow rusting resistance to wheat stem rust using coefficient of infection and
reported the presence of different partial resistance conferring genes in wheat lines (Pathan
and Park 2006, and Draz et al., 2015).
Figure 13. Coefficient of infection (%) of wheat stem rust races on 202 genotypes at Kulumsa
in 2017
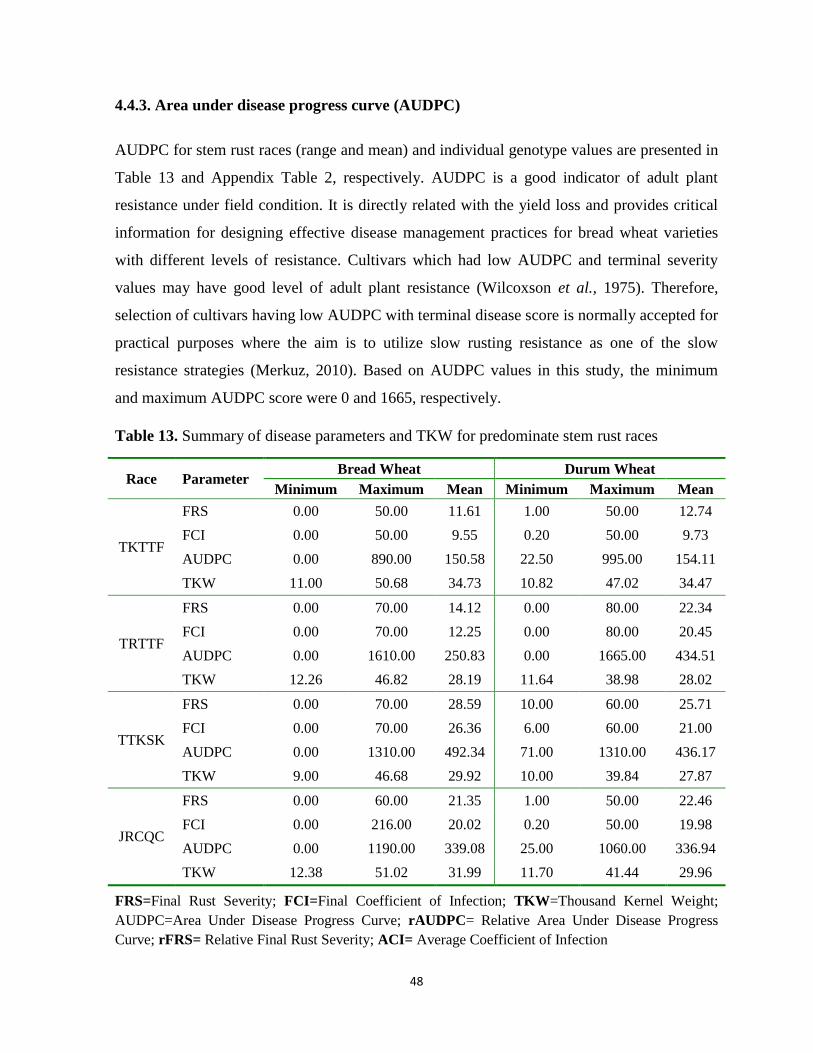
48
4.4.3. Area under disease progress curve (AUDPC)
AUDPC for stem rust races (range and mean) and individual genotype values are presented in
Table 13 and Appendix Table 2, respectively. AUDPC is a good indicator of adult plant
resistance under field condition. It is directly related with the yield loss and provides critical
information for designing effective disease management practices for bread wheat varieties
with different levels of resistance. Cultivars which had low AUDPC and terminal severity
values may have good level of adult plant resistance (Wilcoxson et al., 1975). Therefore,
selection of cultivars having low AUDPC with terminal disease score is normally accepted for
practical purposes where the aim is to utilize slow rusting resistance as one of the slow
resistance strategies (Merkuz, 2010). Based on AUDPC values in this study, the minimum
and maximum AUDPC score were 0 and 1665, respectively.
Table 13. Summary of disease parameters and TKW for predominate stem rust races
Race Parameter Bread Wheat Durum Wheat
Minimum Maximum Mean Minimum Maximum Mean
TKTTF
FRS 0.00 50.00 11.61 1.00 50.00 12.74
FCI 0.00 50.00 9.55 0.20 50.00 9.73
AUDPC 0.00 890.00 150.58 22.50 995.00 154.11
TKW 11.00 50.68 34.73 10.82 47.02 34.47
TRTTF
FRS 0.00 70.00 14.12 0.00 80.00 22.34
FCI 0.00 70.00 12.25 0.00 80.00 20.45
AUDPC 0.00 1610.00 250.83 0.00 1665.00 434.51
TKW 12.26 46.82 28.19 11.64 38.98 28.02
TTKSK
FRS 0.00 70.00 28.59 10.00 60.00 25.71
FCI 0.00 70.00 26.36 6.00 60.00 21.00
AUDPC 0.00 1310.00 492.34 71.00 1310.00 436.17
TKW 9.00 46.68 29.92 10.00 39.84 27.87
JRCQC
FRS 0.00 60.00 21.35 1.00 50.00 22.46
FCI 0.00 216.00 20.02 0.20 50.00 19.98
AUDPC 0.00 1190.00 339.08 25.00 1060.00 336.94
TKW 12.38 51.02 31.99 11.70 41.44 29.96
FRS=Final Rust Severity; FCI=Final Coefficient of Infection; TKW=Thousand Kernel Weight;
AUDPC=Area Under Disease Progress Curve; rAUDPC= Relative Area Under Disease Progress
Curve; rFRS= Relative Final Rust Severity; ACI= Average Coefficient of Infection
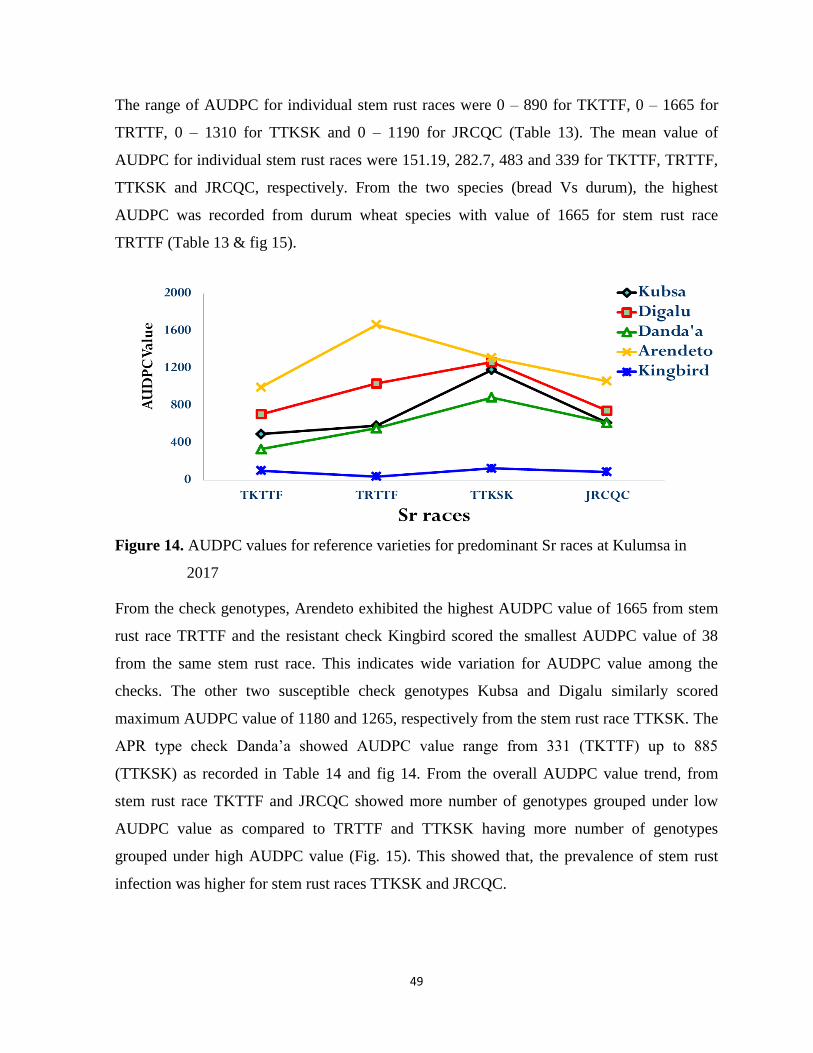
49
The range of AUDPC for individual stem rust races were 0 – 890 for TKTTF, 0 – 1665 for
TRTTF, 0 – 1310 for TTKSK and 0 – 1190 for JRCQC (Table 13). The mean value of
AUDPC for individual stem rust races were 151.19, 282.7, 483 and 339 for TKTTF, TRTTF,
TTKSK and JRCQC, respectively. From the two species (bread Vs durum), the highest
AUDPC was recorded from durum wheat species with value of 1665 for stem rust race
TRTTF (Table 13 & fig 15).
Figure 14. AUDPC values for reference varieties for predominant Sr races at Kulumsa in
2017
From the check genotypes, Arendeto exhibited the highest AUDPC value of 1665 from stem
rust race TRTTF and the resistant check Kingbird scored the smallest AUDPC value of 38
from the same stem rust race. This indicates wide variation for AUDPC value among the
checks. The other two susceptible check genotypes Kubsa and Digalu similarly scored
maximum AUDPC value of 1180 and 1265, respectively from the stem rust race TTKSK. The
APR type check Danda‟a showed AUDPC value range from 331 (TKTTF) up to 885
(TTKSK) as recorded in Table 14 and fig 14. From the overall AUDPC value trend, from
stem rust race TKTTF and JRCQC showed more number of genotypes grouped under low
AUDPC value as compared to TRTTF and TTKSK having more number of genotypes
grouped under high AUDPC value (Fig. 15). This showed that, the prevalence of stem rust
infection was higher for stem rust races TTKSK and JRCQC.

50
Figure 15. AUDPC values for 202 genotypes for predominant stem rust races at Kulumsa in
2017
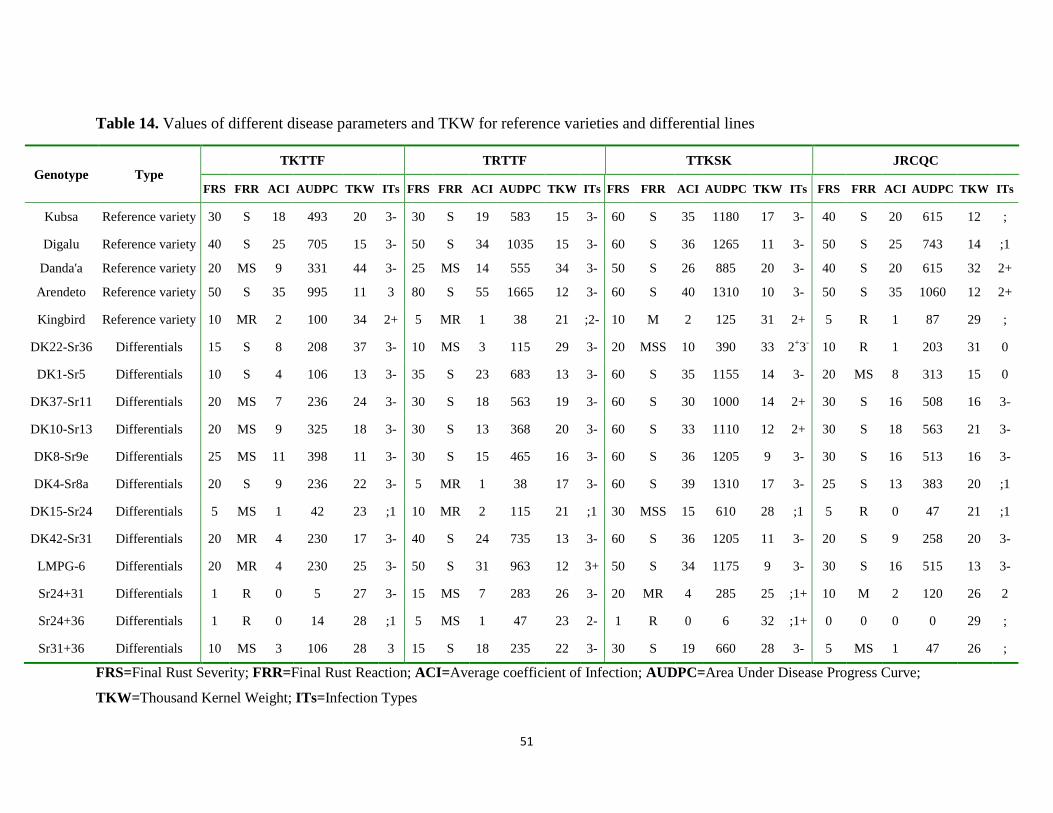
51
Table 14. Values of different disease parameters and TKW for reference varieties and differential lines
Genotype Type TKTTF TRTTF TTKSK JRCQC
FRS FRR ACI AUDPC TKW ITs FRS FRR ACI AUDPC TKW ITs FRS FRR ACI AUDPC TKW ITs FRS FRR ACI AUDPC TKW ITs
Kubsa Reference variety 30 S 18 493 20 3- 30 S 19 583 15 3- 60 S 35 1180 17 3- 40 S 20 615 12 ;
Digalu Reference variety 40 S 25 705 15 3- 50 S 34 1035 15 3- 60 S 36 1265 11 3- 50 S 25 743 14 ;1
Danda'a Reference variety 20 MS 9 331 44 3- 25 MS 14 555 34 3- 50 S 26 885 20 3- 40 S 20 615 32 2+
Arendeto Reference variety 50 S 35 995 11 3 80 S 55 1665 12 3- 60 S 40 1310 10 3- 50 S 35 1060 12 2+
Kingbird Reference variety 10 MR 2 100 34 2+ 5 MR 1 38 21 ;2- 10 M 2 125 31 2+ 5 R 1 87 29 ;
DK22-Sr36 Differentials 15 S 8 208 37 3- 10 MS 3 115 29 3- 20 MSS 10 390 33 2+3
- 10 R 1 203 31 0
DK1-Sr5 Differentials 10 S 4 106 13 3- 35 S 23 683 13 3- 60 S 35 1155 14 3- 20 MS 8 313 15 0
DK37-Sr11 Differentials 20 MS 7 236 24 3- 30 S 18 563 19 3- 60 S 30 1000 14 2+ 30 S 16 508 16 3-
DK10-Sr13 Differentials 20 MS 9 325 18 3- 30 S 13 368 20 3- 60 S 33 1110 12 2+ 30 S 18 563 21 3-
DK8-Sr9e Differentials 25 MS 11 398 11 3- 30 S 15 465 16 3- 60 S 36 1205 9 3- 30 S 16 513 16 3-
DK4-Sr8a Differentials 20 S 9 236 22 3- 5 MR 1 38 17 3- 60 S 39 1310 17 3- 25 S 13 383 20 ;1
DK15-Sr24 Differentials 5 MS 1 42 23 ;1 10 MR 2 115 21 ;1 30 MSS 15 610 28 ;1 5 R 0 47 21 ;1
DK42-Sr31 Differentials 20 MR 4 230 17 3- 40 S 24 735 13 3- 60 S 36 1205 11 3- 20 S 9 258 20 3-
LMPG-6 Differentials 20 MR 4 230 25 3- 50 S 31 963 12 3+ 50 S 34 1175 9 3- 30 S 16 515 13 3-
Sr24+31 Differentials 1 R 0 5 27 3- 15 MS 7 283 26 3- 20 MR 4 285 25 ;1+ 10 M 2 120 26 2
Sr24+36 Differentials 1 R 0 14 28 ;1 5 MS 1 47 23 2- 1 R 0 6 32 ;1+ 0 0 0 0 29 ;
Sr31+36 Differentials 10 MS 3 106 28 3 15 S 18 235 22 3- 30 S 19 660 28 3- 5 MS 1 47 26 ;
FRS=Final Rust Severity; FRR=Final Rust Reaction; ACI=Average coefficient of Infection; AUDPC=Area Under Disease Progress Curve;
TKW=Thousand Kernel Weight; ITs=Infection Types

52
4.5. Correlation between slow rusting parameters of wheat stem rust
Correlation coefficients of genotypes for disease parameters (FRS, AUDPC and ACI) showed
in Table 15. The correlations among the field based slow rusting parameters were positive and
highly significantly correlated within each single race nursery with correlation coefficients
ranging from 0.58 to 0.99. The highest correlation observed from stem rust race TRTTF
between AUDPC vs FRS (r=0.99), ACI vs AUDPC (r=0.99) and from stem rust race TTKSK
between AUDPC vs FRS (r=0.99). A high correlation would mean that the genotypes ranked
fairly similar in all stem rust races and the effects due to race change were minimal. The
lowest correlation value perceived from stem rust race of JRCQC and TRTTF between
AUDPC vs FRS (r=0.58).
In this study, a high and strong positive correlation was noted among all the epidemiological
parameters, AUDPC, FRS and ACI which were used to assess partial resistance at p < 0.001.
These epidemiological parameters give a dependable rate of disease increase and are related
with components of partial resistance like low receptivity, longer latent period and smaller
pustules (McNeil et al., 2008). Similar findings have been reported in previous studies
(Safavi, 2012; Safavi and Afshari, 2012; Ali et al., 2009).
4.6. Correlation between thousand kernel weight and disease parameters
The correlation coefficients considered between pairs of the respective disease parameters
(AUDPC, FRS and ACI) with TKW were highly and negatively correlated for the entire stem
rusts race except for JRCQC (Table 15). The negative relationship between TKW and disease
parameters showed harmful effects of stem rust on the yield component (TKW). However,
TKW correlations among TKW of single races (TKTTF, TRTTF, TTKSK and JRCQC) were
positive and highly and significantly correlated.
The large negative correlation between TKW and stem rust parameters (FRS, AUDPC and
ACI) can be attributed to the fact that the fungus damages the vascular system of the
susceptible host plant extensively limiting transportation of water and nutrients from the soil
to the developing kernel and other organs as well as interfering with translocation of
photosynthesis, which leads to shriveled grains (Singh et al., 2006; Merkuz, and Getachew,
2012b).
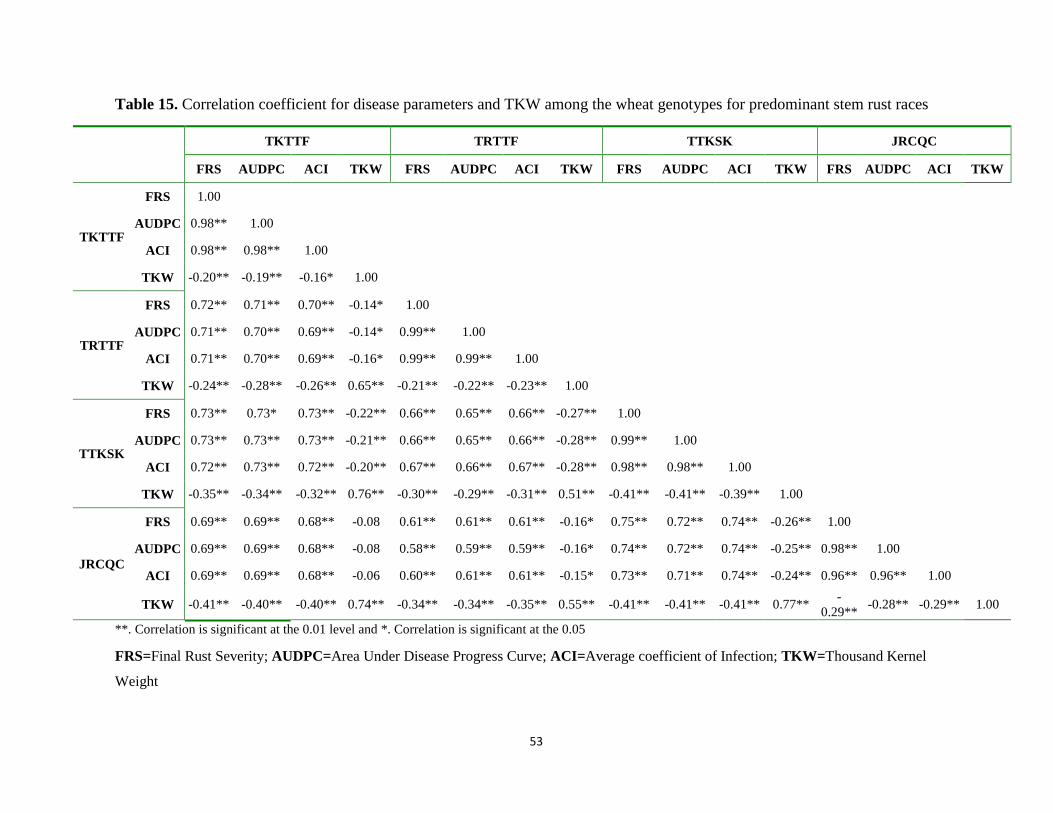
53
Table 15. Correlation coefficient for disease parameters and TKW among the wheat genotypes for predominant stem rust races
TKTTF TRTTF TTKSK JRCQC
FRS AUDPC ACI TKW FRS AUDPC ACI TKW FRS AUDPC ACI TKW FRS AUDPC ACI TKW
TKTTF
FRS 1.00
AUDPC 0.98** 1.00
ACI 0.98** 0.98** 1.00
TKW -0.20** -0.19** -0.16* 1.00
TRTTF
FRS 0.72** 0.71** 0.70** -0.14* 1.00
AUDPC 0.71** 0.70** 0.69** -0.14* 0.99** 1.00
ACI 0.71** 0.70** 0.69** -0.16* 0.99** 0.99** 1.00
TKW -0.24** -0.28** -0.26** 0.65** -0.21** -0.22** -0.23** 1.00
TTKSK
FRS 0.73** 0.73* 0.73** -0.22** 0.66** 0.65** 0.66** -0.27** 1.00
AUDPC 0.73** 0.73** 0.73** -0.21** 0.66** 0.65** 0.66** -0.28** 0.99** 1.00
ACI 0.72** 0.73** 0.72** -0.20** 0.67** 0.66** 0.67** -0.28** 0.98** 0.98** 1.00
TKW -0.35** -0.34** -0.32** 0.76** -0.30** -0.29** -0.31** 0.51** -0.41** -0.41** -0.39** 1.00
JRCQC
FRS 0.69** 0.69** 0.68** -0.08 0.61** 0.61** 0.61** -0.16* 0.75** 0.72** 0.74** -0.26** 1.00
AUDPC 0.69** 0.69** 0.68** -0.08 0.58** 0.59** 0.59** -0.16* 0.74** 0.72** 0.74** -0.25** 0.98** 1.00
ACI 0.69** 0.69** 0.68** -0.06 0.60** 0.61** 0.61** -0.15* 0.73** 0.71** 0.74** -0.24** 0.96** 0.96** 1.00
TKW -0.41** -0.40** -0.40** 0.74** -0.34** -0.34** -0.35** 0.55** -0.41** -0.41** -0.41** 0.77** -
0.29** -0.28** -0.29** 1.00
**. Correlation is significant at the 0.01 level and *. Correlation is significant at the 0.05
FRS=Final Rust Severity; AUDPC=Area Under Disease Progress Curve; ACI=Average coefficient of Infection; TKW=Thousand Kernel
Weight

54
5. SUMMARY AND CONCLUSION
Wheat is one of the most important cereal crops cultivated in a wide range of agro-ecologies.
Most wheat production in Ethiopia comes from smallholder farmers and it is mainly grown in
the central and southeastern highlands. Despite enormous economic and dietary values of the
crop, the average yield has remained extremely low as compared to other wheat producing
countries. This has been attributed to multifaceted biotic and abiotic factors. Rusts, especially
stem rusts, have been considered the most important wheat diseases, and thus, have been one
of the most important priority research agenda since the inception of wheat research in
Ethiopia. It can cause up to 100% yield loss if susceptible cultivars are grown and epidemic
occurs. The use of resistant cultivars has been the major strategy to control stem rust of wheat
worldwide. This study was conducted with the major objectives of managing wheat stem rust
disease through durable resistance.
The study was conducted under three broad categories; field survey, seedling and field
evaluation for slow rusting resistance to stem rust among wheat genotypes. To achieve these
objectives, 10 districts in two zones were assessed by field surveys and 202 genotypes,
including checks and differential lines, were tested for their seedling and adult plant reaction
to major stem rust races during 2017 main cropping season. To test the slow rusting ability of
these varieties, greenhouse experiment was conducted at Ambo Agricultural Research Center
(AARC) while field experiment was conducted at Kulumsa Agricultural Research Center
(KARC). Four nurseries were established for TKTTF, TRTTF, TTKSK and JRCQC stem rust
races. Each nursery was planted in an augmented design with five repeated checks in each
blocks. In order to provide an artificial epidemic condition, particular varieties for each stem
rust race were planted as infector rows perpendicular to the main blocks for each nurseries
and inoculated particular stem rust races for each nurseries. Rust severity and response were
recorded at four stages of ten days‟ interval in each experimental plot, beginning with the
establishment of the rust in infector rows. TKW recorded for each nursery to assess the effect
of each stem rust race.
Field survey results showed that distribution or prevalence of stem rust varied from no
infection to heavy infection of the surveyed fields depending on agro-ecologies and varieties

55
grown. Stem rust was observed on 58 (63%) of the 92 wheat fields inspected. The overall
prevalence of stem ruse was 63.0%. Total prevalence‟s of stem rust in Arsi and Bale zones
were 52.2% and 73.9%, respectively. Major bread wheat varieties grown included Ogolcho,
Hidasse, Danda'a, Hulluka, Medawalabu, Kubsa, K-6295-4A, Batu, Kakaba, Senbete, Pavon-
76, Galama, Dashen and Digalu while few durum wheat varieties Alemtena, Tesfaye and Ude
were recorded in the surveyed areas.
A total of 40 stem rust samples were collected from infested fields during the survey and 32
viable samples were used for race analysis. A total of five races namley TKTTF, TTTTF,
TTRTF, TTKSK and TRTTF were identified from Arsi and Bale zones during the 2017 main
cropping season. The most predominant races were TKTTF (53.0%), TTTTF (31.0%) and
TTRTF (10.0%). The least frequent races were TTKSK and TRTTF with a frequency of 3.0%
each. Identification of five races from 32 samples is a clear indication of high virulence
diversity within the P. graminis f. sp. tritici populations in Ethiopia. The observed/recorded
virulence spectrum varied from 17-18 Sr genes. The widest virulence spectrum was recorded
on the race TTTTF which exhibited virulence on 18 Sr genes. The other virulence spectrum
was recorded on TKTTF, TTKSK, TTRTF and TRTTF races and showed virulence on 17 Sr
genes.
Seedling evaluation results showed high level of variability in responses of genotypes to stem
rust races TTKSK, TKTTF, TRTTF and JRCQC. The ITs frequency distribution shows a
variability in reaction among the test cultivars for all four races used in this study with the
majority of the genotype showing resistance reaction with score of 0 to 2. Only few genotypes
showed a susceptible reaction with score of 3- to 3. Correlation coefficients among stem rust
races result revealed that all races were positive and highly correlated each other‟s. The
highest correlation was observed between stem rust race TTKSK and TRTTF.
Field experiments showed high disease severity in all stem rust single races nurseries.
Phenotypic variation was observed for infection types and level of stem rust severity for
wheat genotypes. Terminal score ranged for all stem rust races from 0 (immune) to 80% S
(highly susceptible). From ANOVA mean square values, highly significant difference
(P<0.001) were observed among check and new genotypes for different disease parameters
(AUDPC and ACI) and TKW for each single race nursery. The final rust severity score

56
showed more number of genotypes were grouped under MR and S types of reaction for all
stem rust races. From seedling and field responses of genotypes for all stem rust races, 31
genotypes showed resistance to all four stem rust races at both seedling and adult-plant stages.
This suggested that when genotypes show resistance at both seedling and adult-plant stages, it
can be referred to as all-stage resistance. APR is often characterized by its effectiveness
against a wide range of pathogen races. From this study, only genotype G42 (ETBW 9464)
showed seedling susceptible but adult plant resistance for all wheat stem rust races used in
this study. However, four genotypes (G40, G42, G175, G195 and G196) showed seedling
susceptible and field resistant reaction for the three major bread wheat stem rust races
(TKTTF, TRTTF and TTKSK).
AUDPC is a good indicator of adult plant resistance under field condition and based on
AUDPC in this study, the values range from 0 - 1665. From check genotypes, Arendeto
exhibited the highest AUDPC value of 1665 from stem rust race TRTTF while the resistant
check Kingbird scored the smallest AUDPC value of 38 from the same stem rust race.
Correlation coefficients for slow rusting parameters (FRS, AUDPC and ACI) were positive
and highly significant within each single race nursery. Correlation coefficients ranged from
0.58 to 0.99. However, slow rusting parameters and TKW were highly and negatively
correlated in all stem rust races except in JRCQC.
In conclusion, from this study the current stem rust race distribution in Arsi and Bale zones
were identified and some genotypes having adult plant resistance selected based on seedling
and field evaluation. Those identified races will be helpful for future pathological activities to
screen wheat genotypes for those predominant races. The selected genotypes (G40, G42,
G175, G195 and G196) from this study also important input for wheat breeding crossing
programs and further pathological activities.

57
6. REFERENCES
Abebe, T., Getaneh, W., and Woubit, D., 2012. Analysis of pathogen virulence of wheat stem
rust and cultivar reaction to virulent races in Tigray, Ethiopia. African Journal of Plant
Science. 6(9): 244-250.
Abera, B., 1991. Biochemical aspects of wheat in human nutrition. In Hailu Gebre-Mariam,
Tanner, D.G., and Mengistu Hulluka (ed.) Wheat Research in Ethiopia: A Historical
Perspective. pp. 341-352. Addis Ababa: IAR/CIMMYT.
Abu, T., 2012. Ethiopia Grain and Feed Annual Report. GRAIN Report Number: ET 1201
(http://gain.fas.usda.gov/Recent%20GAIN%20Publications/Grain%20and%20Feed%2
0Annual_Addis%20Ababa_Ethiopia_4-17-2012.pdf.
Admassu, B., and Fekadu, E., 2005. Physiological races and virulence diversity of Puccinia
graminis f.sp. tritici on wheat in Ethiopi. Phytopathologia Mediterranea 44: 313-318.
Admassu, B., Friedt, W., and Ordon, F., 2009. Genetic characterization of graminis f. sp.
tritici populations from Ethiopia by SSRs. Journal of Phytopathol. 158: 806-812.
Afzal, S. N., I. Ahmedani, H. M.S., Munir, M., Firdous, S. S., Rauf, A., Ahmad, I., Rattu . R.
and Fayyaz, M., 2009. Resistance potential of wheat germplasm Triticum aestivum l.
against stripe rust disease under rain fed climate of Pakistan. Pakistan Journal of
Botany. 41 (3):1463-1475.
Agrios, G., 2005. Plant pathology, 5th
ed. Boston, Mass., U.S.A.: Elsevier Academic Press.
Alexopoulos, C.J., Mims, C.W., and Blackwell, M., 1996. Introductory Mycology. 4th
ed.
John Wiley and Sons. Inc., New York.
Ali, S., Shah, S.J.A., Khalil, I.H., Raman H, Maqbool K, Ullah W., 2009. Partial resistance to
yellow rust in introduced winter wheat germplasm at the north of Pakistan. Australia
Journal of Crop Science, 3: 37-43.
Anikster,Y., 1984. The formaespeciales. In: Bushnell, W.R., and Roelfs, A.P. (eds). The
Cereal Rusts. vol I. Origins, Specificity, Structure and Physiology. Academic Press,
inc., London.

58
Badebo, A., 2002. Breeding bread wheat with multiple disease resistance and high the
yielding for the Ethiopian highlands: broadening genetic basis of yellow rust and tan
spot resistance. Gottingen, Germany: Gottingen University, PhD thesis.
Badebo, A., and Hundie, B., 2016. Incidence and challenges of rusts in wheat production, in
Bishaw et al. (Ed.), containing the Menace of Wheat Rusts: Institutionalized
Interventions and Impacts, ISBN: 9789994466344, EIAR, Addis Ababa.
Beard, C., Jayasena K., Thomas, G., and Loughman, R., 2004. Managing Stem Rust of
Wheat. Plant Pathology, Department of Agriculture, Western Australia.
Bekele, E., 1985. A review of research on diseases of barley, tef and wheat in Ethiopia. In:
Tsedeke Abate (ed.), A review of crop protection research in Ethiopia. Institute of
Agricultural Research (IAR), Ethiopia. pp. 79-107.
Bekele, H., 2003. Short report on yellow rust and stem rust. In: Bedada Girma (ed.).
Proceedings of the Agronomy Workshop organized by Bale Agricultural Development
Enterprise. Nazereth, Ethiopia. pp. 67-78.
Biffen, R.H., 1905. Mendel‟s laws of inheritance and wheat breeding. Journal of Agricultural
Science 1: 4-48.
Bogale, A., N. Kedir, and H. Seboka, 2011b. Selection of some morphological traits of bread
wheat that enhance the competitiveness against wild oat (Avena fatua L.). World
Journal of Agricultural Sciences 7: 128-135.
Bogale, A., T. Kindie, and T. Geleto, 2011a. Morphological and physiological attributes
associated to drought tolerance of Ethiopian durum wheat genotypes under water
deficit condition. Journal of Biodiversity and Environmental Sciences 1: 22-36.
Boyd, L.A., 2005. Can Robigus defeat an old enemy? Yellow rust of wheat. Journal of
Agricultural Sciences 143: 233–243.
Bushnell, W. R., and Rowell, J.B., 1968. Premature death of adult rusted wheat plants in
relation to carbon dioxide evolution by root system. Phytopathology 58: 651-658

59
Campbell, CL., and Long, DL., 2001. The campaign to eradicate the common barberry in the
United States. Peterson PD, editor. In: Stem rust of wheat: From ancient enemy to
modern foe. St. Paul, Minn.: APS Press: pp.16-43.
Chen, W., Wellings, C., Chen, X., Kang, Z., Liu, T., 2014. Wheat stripe (yellow) rust caused
by Puccinia striiformis f. sp. tritici. Molecular Plant Pathology.15: 433–446.
Chen, W.Q., Xu, S.C., and Wu, L.R., 2007. Epidemiology and sustainable management of
wheat stripe rust caused by Puccinia striiformis West in China: a historical retrospect
and prospect. Sci. Agric. Sin. 40: 177–183.
Chen, X.M., 2005. Epidemiology and control of stripe rust Puccinia striiformisf. sp tritici on
wheat. Can. J. Plant Pathol. 27: 314-337.
CIMMYT., 2005. Sounding the alarm on global stem rust: an assessment of race Ug99 in
Kenya and Ethiopia and the potential for impact in neighboring countries and beyond.
Mexico city, Mexico.
CSA., 1995-2018. Agricultural Sample Survey 1995-2018. Report on area and production of
major crops. Addis Ababa, Ethiopia.
Denbel, W., Badebo, A., and Alemu, T., 2013. Evaluation of Ethiopian commercial wheat
cultivars for resistance to stem rust of wheat race „UG99‟. International Journal of
Agronomy and Plant Production, 4: 15-24.
Dereje,H.,& Chemeda. F., 2009. Relationship between stripe rust (Puccinia striiformis) and
common wheat (Triticum aestivum) yield loss in the highlands of Bale, southeastern
Ethiopia.
Devos, K.M., and Gale, M.D., 1997. Comparative genetics in the grasses. Plant Molecular
Biology 35: 3-15.
Draz, I.S., Abou-Elseoud, M.S., Kamara, A.M., Alaa-Eldein, O.A., El-Bebany, A.F., 2015.
Screening of wheat genotypes for leaf rust resistance along with grain yield. Ann.
Agricultural Science 60: 29-39.
Dubin, H.J., and B.P. Jone.P., 2009. Combating Stem and Leaf Rust of Wheat Historical
Perspective, Impacts, and Lessons Learned. IFPRI Discussion Paper 00910.

60
Duveiller, E., Singh, R.P., and Nicol, J.M., 2007. The challenges of maintaining wheat
productivity: pests, diseases, and potential epidemics. Euphytica, 157(3): 417-430.
Dvorak, J., Luo, M.C., Yang, Z.L., Zhang, H.B., 1998. The structure of the Aegilopstauschii
gene pool and the evolution of hexaploid wheat. Theoretical Applied Genetics 97: 657-
670.
Endale, H., Getaneh, W., Worku, D., Wubeshet, A., Teklay, A., 2015. Distribution of
Stem Rust (Puccinia graminis f. sp. tritici) Races in Ethiopia. Advances in Crop
Science and Technology 3: 173.
Eshetu B., 1985. A Review of Research on Diseases of Barley, Tef and Wheat in Ethiopia. In:
Tsedeke Abate (eds). A Review of Crop Protection Research in Ethiopia. Proceeding of
the First Ethiopian Crop Protection Symposium. 4-7 February, 1985. Addis Ababa,
Ethiopia: pp. 79-148.
FAO., 2017. Food and Agricultural Organization of the United Nations (FAO): FAO Global
Statistical Year book. http://www.fao.org/faostat/en/#data/QC. Metadata last certified
Nov. 2017.
Ferede, T., Ayenew, A., Hanjra, M., and Hanjra, M., 2013. Agro ecology matters: impacts of
climate change on agriculture and its implications for food security in Ethiopia. Global
food security: Emerging issues and economic implications: pp. 71-112.
Ferrara, G.O., 2008. Development of Trap Nurseries: Cooperative Screening, Evaluation,
Virulence Monitoring, Variants Detection, in: G. P. Singh, et al. (Eds.), International
Conference on Wheat Stem Rust Ug 99- A Threat to Food Security, New Delhi: pp.
56-61.
Fetch, TG., Dunsmore, KM., 2004. Physiological specialization of P. graminis on wheat,
barley, and oat in Canada in 2001. Canadian Journal of Plant Pathology 26: 148-55
Haile, D., Nigussie, D., and Ayana, A., 2012. Nitrogen use efficiency of bread wheat: Effects
of nitrogen rate and time of application. Journal of Soil Science and Plant Nutrition 12:
389-410.
Hailu, G., Tanner, DG., and Mengistu, H., (eds). 2002. Wheat research in Ethiopia: A
Historical perspective, IARI and CIMMYT, Addis Ababa.

61
Hailu, A., Woldeab, G., Dawit, W., and Hailu, E., 2015. Distribution of Wheat Stem Rust
(Puccinia graminis f.sp. tritici) in West and Southwest Shewa Zones and Identification
of its Phsiological Races. Advances in Crop Science and Technology: pp. 1-10.
Hamilton, L.M., and Stakman, E.C., 1967. Time of stem rust appearance on wheat in the
Western Mississippi basin in relation to the development of epidemics from 1921 to
1962. Phytopathology 57: 609-614.
Hei, N., Shimelis, A., and Laing, M., 2017. Appraisal of farmers‟ wheat production
constraints and breeding priorities in rust prone agro-ecologies of Ethiopia. African
journal of agricultural research. 12: 944-952.
Hodson, D., 2015. Summary of Ethiopia 2014/15 rust situation. Re-current, localized stem
rust epidemics caused by race TKTTF (“Digalu” race) in Ethiopia. Extreme caution &
vigilance needed in East Africa. Rust tracker.org, Global wheat rust monitoring system.
Hodson, D., 2013. Localized stem rust epidemic in southern Ethiopia extreme caution and
vigilance needed in East Africa and Middle East region. A global wheat rust
monitoring system.
Hodson, D., 2014. Keeping track of rust. Available at: http://rusttracker.cimmyt.org/?
page_id=40 Accessed 4 May 2014.
Hogg, W.H., Hounam, C.E., Mallik, A.K., and Zadoks, J.C., 1969. Meteorological factors
affecting the epidemiology of wheat rust. WMO. Tech. Note. 99: 143.
Hulluka, M., Woldeab, G., Adnew, Y., Desta, R., and Badebo, A., 1991. Wheat pathology
research in Ethiopia: 173-218 in: Wheat Research in Ethiopia: A Historical Perspective.
H. Gebre-Mariam, D. G. Tanner, and M. Hulluka, eds. IAR/CIMMYT, Addis Ababa,
Ethiopia.
Jin, Y., Szabo LJ., Pretorius, ZA., Singh, RP., Ward, R., 2008. Detection of virulence to
resistance gene Sr 24 within race TTKS of Puccinia graminis f. sp. tritici. Plant Dis 92:
923-926
Jin, Y., Singh, RP., Ward, RW., Wanyera, R., Kinyua, M., 2007. Characterization of seedling
infection types and adult plant infection responses of monogenic Sr gene lines to race
TTKS of Puccinia graminis f.sp. tritici. Plant Dis 91: 1096 - 1099.

62
Jobie, T., 2007. Delia flavibasis stein (Diptera: Anthomyiidae) as a major pest of wheat in the
major cereal belt, Southeast Ethiopia. African crop science conference proceedings 8:
965-969.
Johnson, R., 1984. A critical analysis of durable resistance. Annu Rev Phytopathol 22: 309-
330.
Khan, M.H., Bukhari, A., Dar, Z.A., and Rizvi, S.M., 2013. Status and strategies in breeding
for rust resistance in wheat. Agricultural Sciences, 4(06): 292.
Klindworth, D.L., Miller, J.D., Jin, Y., and Xu, S.S., 2007. Chromosomal locations of genes
for stem rust resistance in monogenic lines derived from tetraploid wheat accession
ST464. Crop science, 47(4): 1441-1450.
Kolmer, A., Ordonez, E., and Groth, V., 2013. The Rust Fungi. In: Encyclopedia of Life
Sciences (ELS). John Wiley and Sons, Ltd: Chichester. DOI:
10.1002/9780470015902.a0021264: 1-8.
Kurt, J.L., 2001. Black Stem Rust Biology and Treats to the Grower. U.S. Department of
Agriculture, Agricultural Research Service, Cereal Disease Laboratory. University of
Minnesota, st. Poul 55108.
Lemma, A., Woldeab, G., Semahegn, Y., and Dilnesaw, Z., 2014. Survey and virulence
distribution of wheat stem rust (Puccinia graminis f.sp. tritici) in the major wheat
growing areas of central Ethiopia. Sci-Afric Journal of Scientific Issues, Research and
Essays 2:474-78.
Leonard, J., and Szabo, J., 2005. Stem Rust of Small Grains and Grasses caused by Puccinia
graminis f.sp. tritici. doi:10.1111/j.1364-3703.2004.00273.x.
Luig, N.H., 1985. Epidemiology in Australia and New Zealand. In: A.P. Roelfs and W.R.
Bushnell (eds.) Cereal Rusts Vol. II Diseases, Distribution, Epidemiology and Control.
Academic Press, Orlando, USA: pp. 301-328.
McIntosh, R.A., Wellings C.R., Park R.F., 1995. Wheat Rusts: an Atlas of Resistance Genes.
CSIRO Publishing, Melbourne.

63
McNeil, MD., Kota, R., Paux, E., Dunn, D., McLean, R., Feuillet, C., Li, D., Kong, X.,
Lagudah, E., Zhang, JC., Jia, JZ., Spielmeyer, W., Bellgard, M., and Appels, R., 2008.
BAC-derived markers for assaying the stem rust resistance gene, Sr2, in wheat
breeding programs. Molecular Breeding 22: 15-24.
Mideksa, T., Fininsa, C., and Hundie, B., 2018. Analysis of Climate Variability Effects on
Wheat Stem Rust (Puccinia graminis f.sp tritici) Epidemics in Bale and Arsi Zones of
Oromia Regional State, Ethiopia. American Journal of Biological and Environmental
Statistics. Vol. 4, No. 2, doi: 10.11648/j.ajbes.20180402.12: 49-65.
Milus, E.A., and Line, R.F., 1986. Number of genes controlling high-temperature adult-plant
resistance to stripe rust in wheat. Phytopathology, 76: 93 - 96.
Minot, N., Warner, J., Lemma, S., Kassa, L., Gashaw, A., and Rashid, S., 2015. The Wheat
Supply Chain in Ethiopia: Patterns, Trends, and Policy Options. International Food
Policy Research Institute (IFPRI) Washington, DC.
Oliver, R.P., 2014. A reassessment of the risk of rust fungi developing resistance to
fungicides. Pest Manag. Sci. 70:1641–1645.
Olivera, P. D., Jin, Y., Rouse, M., Badebo, A., Fetch, T., Jr., Singh, R. P., and Yahyaoui, A.,
2012. Races of Puccinia graminis f. sp. tritici with combined virulence to Sr13 and
Sr9e in a field stem rust screening nursery in Ethiopia. Plant Dis. 96: 623-628.
Olivera, P., Newcomb, M., Flath, K., Sommerfeldt-Impe, N., Szabo, L., Carter, M., Luster,
D., and Jin, Y. 2017. Characterization of Puccinia graminis f. sp. tritici isolates derived
from an unusual wheat stem rust outbreak in Germany in 2013. Plant Pathology. 66,
no. 8: 1258–1266.
Olivera, P., Newcomb, M., Szabo, L.J., Rouse, M., Johnson, J., Gale, S., Luster, DG.,
Hodson, D., Cox, JA., Burgin, L., Hort, M., Gilligan, CA., Patpour, M.,Justesen, AF.,
Hovmøller, MS., Woldeab, G., Hailu, E., Hundie, B., Tadesse, K., Pumphrey, M.,
Singh, RP., and Jin, Y. 2015. Phenotypic and genotypic characterization of race
TKTTF of Puccinia graminis f. sp. tritici that caused a wheat stem rust epidemic in
southern Ethiopia in 2013–14. Phytopathology 105: 917-928.

64
Park, RF., Fetch, T., Hodson, DP., Jin, Y., and Nazari, K., 2010. International surveillance of
wheat rust pathogens-progress and challenges. BGRI 2010 Technical workshop; 30-31
May; St Petersburg, Russia.
Park, R. L., 2007. Stem rust of wheat in Australia. Aust. J. Agric. Res. 58: 558-566.
Parlevliet, J.E., 1975. Partial resistance of barley to leaf rust, Puccinia hordei. Effect of
cultivar and development stage on latent period. Euphytica 24: 21–27.
Pathan, AK., Park, RF., 2006. Evaluation of seedling and adult plant resistance to leaf rust in
European wheat cultivars. Euphytica 149: 327-342.
Peterson,, RF., Campbell, AR., Hannah, AE., 1948. A diagrammatic scale for estimating rust
intensity on leaves and stem of cereals. Canadian Journal Research 26: 490-500
Peterson, DP., 2001. Stem Rust of Wheat from Ancient Enemy to the Modern Foe. The
American Phytopathological Society. APS Press. St. Paul, Minnesota.
Prescott, JM., Burnett, PA., LeSaari, EE., Ranson, J., Bowman, J., Milliano, W., Singh, RP.,
and Bekele G., 1986. Wheat diseases and pests: a guide for field identification.
CIMMYT, Mixico city. DF. Mexico: pp. 135.
Pretorius, Z., Singh, R., Wagoire, W., and Payne, T., 2000. Detection of virulence to wheat
stem rust resistance gene Sr31 in Puccinia graminis f.sp. tritici in Uganda.
Phytopathology. 84: 203.
Rehman, A.U., Sajjad, M., Khan, S.H. and Ahmad, N., 2013. Prospects of wheat breeding for
durable resistance against brown, yellow and black rust fungi. International Journal of
Agricultural Biology 15: 1209-1220.
Roelfs, AP, Singh, RP, Saari, EE., 1992. Rust Diseases of Wheat: Concept and Methods of
Disease Management. CIMMYT: pp. 81.
Roelfs, A., and Martens, J., 1988. An international system of nomenclature for Puccinia
graminis f.sp. tritici. Phytopathology, 78: 526-533.
Roelfs, A.P., 1985a. Epidemiology in North America. In A.P. Roelfs and W.R. Bushnell.
(eds.) The Cereal Rusts Vol. II; Diseases, Distribution, Epidemiology and Control.
Academic Press, Orlando: pp. 403-434.

65
Roelfs, A.P., 1985b. Wheat and Rye Stem Rust. In A.P. Roelfs and W.R. Bushnell, (eds.)
Cereal Rusts Vol. II; Diseases, Distribution, Epidemiology, and Control, Academic
Press, Orlando: pp. 301-328.
Rowell, J. B., 1984. Controlled infection by Puccinia graminis f. sp. tritici under artificial
condition. In: Bushnel, W. R. and Roelfs, A. P. (eds). The Cereal Rusts. Academic
Press, Inc., London: pp. 291-331.
Rowell, J. B., and Roelfs, A.P., 1976. Wheat Stem Rust. In: Tummala, R.L., Haynes, D.R.
and Croft B.A. (eds.). Modeling for Pest Management, Concepts, Techniques and
Applications in USA/US. Second US/US. Symp., Michigan State University, East
Lansing: pp. 69-79.
Saari, E.E., and Prescott, J.M., 1985. World distribution in relation to economic losses. In:
Roelfs A.P. and Bushnell W.R. (eds.). The Cereal Rusts Vol. II; Diseases, Distribution,
Epidemiology, and Control. Academic Press, Orlando: pp. 259-298.
Safavi, S.A., and Afshari, F., 2012. Identification of resistance to Puccinia striiformis f.sp.
tritici in some elite wheat lines. Journal of Crop Protection 1: 293 - 302.
Safavi, SA., 2012. Evaluation of slow rusting parameters in thirty seven promising wheat
lines to yellow rust. Tech. J. Eng. Appl. Sci. 2: 324-329.
Savile, D.B.O., 1984. Taxonomy of the cereal rust fungi. In: Bushnell, W.R. and Roelfs, A.P.
(eds.). The cereal rusts, vol. 1, Origin, Specificity, Structures and physiology. Orlando,
F.L., USA Academic Press: pp. 79-112.
Shank, R., 1994. Wheat stem rust and drought effect on Bale agricultural production and
future prespects. Summerized report. 17-19 February. UNDP emergency Unit in
Ethiopia. Addis Ababa.
Sherif, A., Berhanu, B., Abebe, A., Zewdie, B., Dawit, A., and Asnake F., 2016. “Promoting
Rust-resistant Wheat Varieties and Technologies through Partnership Incidence and
challenges of rusts in wheat production”, in Bishaw et al. (Ed.), containing the Menace
of Wheat Rusts: Institutionalized Interventions and Impacts, ISBN: 9789994466344,
EIAR, Addis Ababa.

66
Shiferaw, B., Smale, M., Braun, H.J., Duveiller, E., Reynolds, M., and Muricho, G., 2013.
Crops that feed the world 10. Past successes and future challenges to the role played by
wheat in global food security. Food Security, 5(3): 291-317.
Singh, R. P., Huerta-Espino, J. and William, H.M., 2005. Genetics and breeding for durable
resistance to leaf and stripe rusts in wheat. Turk. Journal of Agriculture for Forestry.
29: 121-127.
Singh, R., Hodson, D., Huerta-Espino, J., Jin, Y., Bhavani, S., Njau, P., Herrera-Foessel, S.,
Singh, P., Singh, S., Govindan, V. 2011. The emergence of Ug99 races of the stem rust
fungus is a threat to world wheat production. Annu. Rev. Phytopathol. 49: 465–481.
Singh, R., Hodson, D., Jin, Y., Huerta-Espino, J., Kinyua, M., Wanyera, R., Njau, P., Ward,
R., 2006. Current status, likely migration, and strategies to mitigate the threat to wheat
production from race Ug99 (TTKS) of stem rust pathogen. CAB Review: Perspectives
in Agriculture, Veterinary Science, Nutrition and Natural Resources, 1: 1- 13.
Singh, R., Hodson, D., Jin, Y., Lagudah, E., Ayliffe, M., Bhavani, S., Rouse, M., Pretorius,
Z., Szabo, L., Huerta-espino, J., Basnet, B., Lan, C., and Hovmøller, M. 2015.
„Emergence and Spread of New Races of Wheat Stem Rust Fungus : Continued Threat
to Food Security and Prospects of Genetic Control‟: pp. 99.
Singh, R.P., Hodson, D.P., Jin, Y., Huerta-Espino, J., Kinyua, M.G., Wanyera, R., Njau, P.
and Ward, R.W., 2006. Current status, likely migration and strategies to mitigate the
threat to wheat production from race Ug99 (TTKS) of stem rust pathogen. CAB
reviews: perspectives in agriculture, veterinary science, nutrition and natural
resources, 1(54): 1-13.
Singh, R.P., Huerta-Espino, J., Roelfs, A.P., 2002. The Wheat Rusts. In: B.C. Curtis, S.
Rajaram, H. Gomez Macpherson (eds.). Bread Wheat Improvement and Production.
Food and Agriculture Organization (FAO) of the United Nations. Rome, Italy. pp. 554.
Singh, R.S., 1998. Plant Disease. Seventh ed. Oxford and IBH Publishing Co.Pv. LTD, New
Delhi.
Sorokina, G., Gorshkov, A. and Madumarov, T., 1980. Working out methods to enable an
improvement in resistance of cereals to the major fungal disease. Scientific

67
phytopathological laboratory (SPL), Progress Report for the Period 1978. Ambo,
Ethiopia.
Stakman, EC., Steward, DM., Loegering, WQ., 1962. Identification of physiologic races of
Puccinia graminis f.sp. tritici. Agric Res Serv E-617: 1-53
Stubbs, R. W., Prescott, J. M., Saari, E. E. and Dubin, H. J. 1986. Cereal Disease
Methodology Manual. Centro Intemacional de Mejoramiento de Maiz y Trigo
(CIMMYT), Mexicco. pp. 46.
Tesfaye, L., Seedling Resistance to Stem Rust (Puccinia graminis f.sp. tritici) and Molecular
Marker Analysis of Resistance Genes in Some Wheat Cultivars. Plant. Vol. 6, No. 1,
2018: 16-23. Wilcoxson, R.D., Skovmand, B. and Atif, A.H., 1975. Evaluation of
wheat cultivars ability to retard development of stem rust. Annals of Applied Biology
80: 275-2181
Woldeab, G., Hailu, E., and Bacha, N. 2017. Protocols for Race Analysis of Wheat Stem Rust
(Puccinia graminis f.sp. tritici). EIAR, Ambo, Ethiopia. Available online at
www.globalrust.org/race-manual/Ambo. pp. 1-26.
Woldeab, G., Hailu, E., and Negash, T., 2016. Detection of Barberry plants (Berberis holstii)
as an alternate host of stem rust (Puccinia graminis f.sp. tritici) of wheat in Ethiopia.
Pest Management Journal of Ethiopia 19. 17-26.
Zadoks, JC., Chang, TT., Kanzak, CF., 1974. A decimal code for the growth stage of cereals.
Weed Research 14: 415-421.
Zadoks, JC., and Bouwman, JJ., 1985. Epidemiology in Europe. In A.P. Roelfs and W.R.
Bushnell (eds). The Cereal Rusts Vol. II; Diseases, Distribution, Epidemiology, and
Control, Academic Press, Orlando. pp 329-369.

68
7. APPENDIX

69
Appendix Table 1. Data sheet for wheat stem rust assessment in Arsi and Bale zones of
Ethiopia (2017)
Part-I: Data collector information
Surveyor name: _________ Institution: ___________ Email: __________ Field code: ___________
Date (dd/mm/yy): _______ Region ________ Zone _______ District; _____ Location _________
Part-II: Field and crop management information
1. Field information:
1.1 Size of the wheat field (in ha): ____________
1.2 GPS data: Latitude (N)
1.3 Longitude (E)
1.4 Altitude (m) __________
1.5 Variety_____________
1.6 Growth stage Milky Flowering
Dough Hard dough
2. Crop management:
2.1 Frequency of tillage/ plough before sowing: ________________
2.2 Sowing date (dd/mm/yy): __________________
2.3 Panting/sowing pattern: Row Broadcast
2.4 Field history (previous crop): ____________________________
2.5 Field sanitation: Weedy Good Moderate
2.6 Crop stand/performance: Good Fair Bad
3. %SR Prevalence: ________________
4. %SR Incidence: _________________
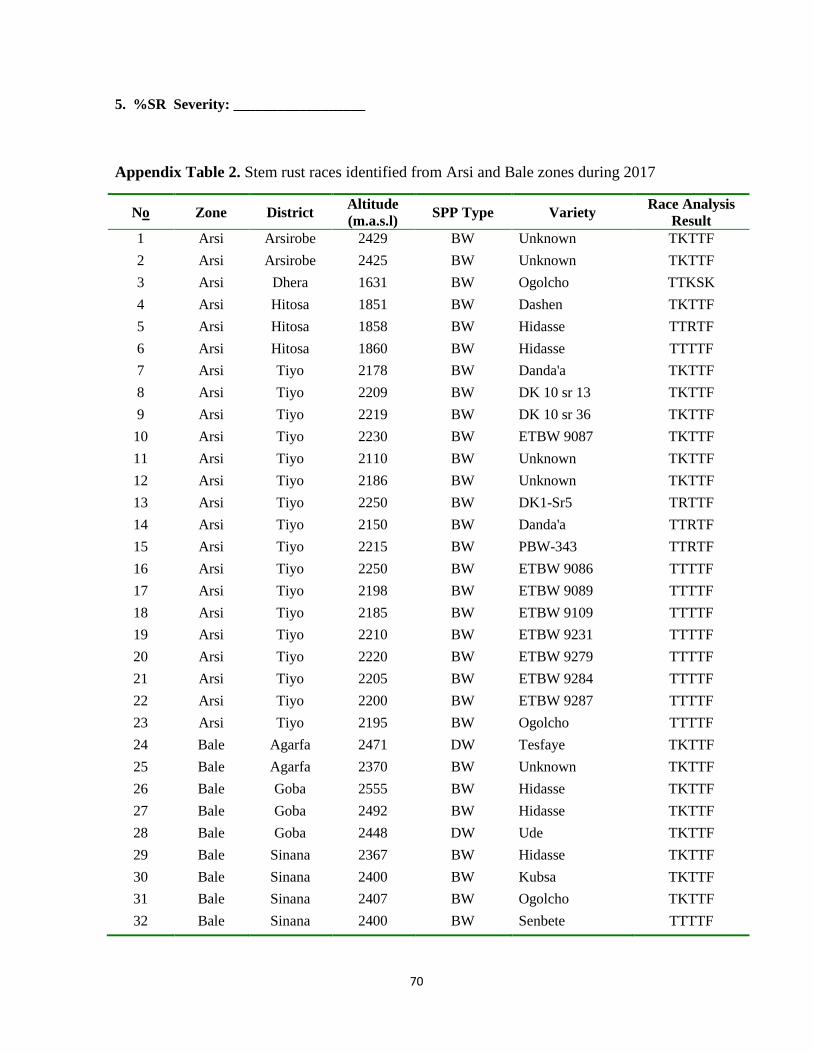
70
5. %SR Severity: __________________
Appendix Table 2. Stem rust races identified from Arsi and Bale zones during 2017
No Zone District Altitude
(m.a.s.l) SPP Type Variety
Race Analysis
Result
1 Arsi Arsirobe 2429 BW Unknown TKTTF
2 Arsi Arsirobe 2425 BW Unknown TKTTF
3 Arsi Dhera 1631 BW Ogolcho TTKSK
4 Arsi Hitosa 1851 BW Dashen TKTTF
5 Arsi Hitosa 1858 BW Hidasse TTRTF
6 Arsi Hitosa 1860 BW Hidasse TTTTF
7 Arsi Tiyo 2178 BW Danda'a TKTTF
8 Arsi Tiyo 2209 BW DK 10 sr 13 TKTTF
9 Arsi Tiyo 2219 BW DK 10 sr 36 TKTTF
10 Arsi Tiyo 2230 BW ETBW 9087 TKTTF
11 Arsi Tiyo 2110 BW Unknown TKTTF
12 Arsi Tiyo 2186 BW Unknown TKTTF
13 Arsi Tiyo 2250 BW DK1-Sr5 TRTTF
14 Arsi Tiyo 2150 BW Danda'a TTRTF
15 Arsi Tiyo 2215 BW PBW-343 TTRTF
16 Arsi Tiyo 2250 BW ETBW 9086 TTTTF
17 Arsi Tiyo 2198 BW ETBW 9089 TTTTF
18 Arsi Tiyo 2185 BW ETBW 9109 TTTTF
19 Arsi Tiyo 2210 BW ETBW 9231 TTTTF
20 Arsi Tiyo 2220 BW ETBW 9279 TTTTF
21 Arsi Tiyo 2205 BW ETBW 9284 TTTTF
22 Arsi Tiyo 2200 BW ETBW 9287 TTTTF
23 Arsi Tiyo 2195 BW Ogolcho TTTTF
24 Bale Agarfa 2471 DW Tesfaye TKTTF
25 Bale Agarfa 2370 BW Unknown TKTTF
26 Bale Goba 2555 BW Hidasse TKTTF
27 Bale Goba 2492 BW Hidasse TKTTF
28 Bale Goba 2448 DW Ude TKTTF
29 Bale Sinana 2367 BW Hidasse TKTTF
30 Bale Sinana 2400 BW Kubsa TKTTF
31 Bale Sinana 2407 BW Ogolcho TKTTF
32 Bale Sinana 2400 BW Senbete TTTTF
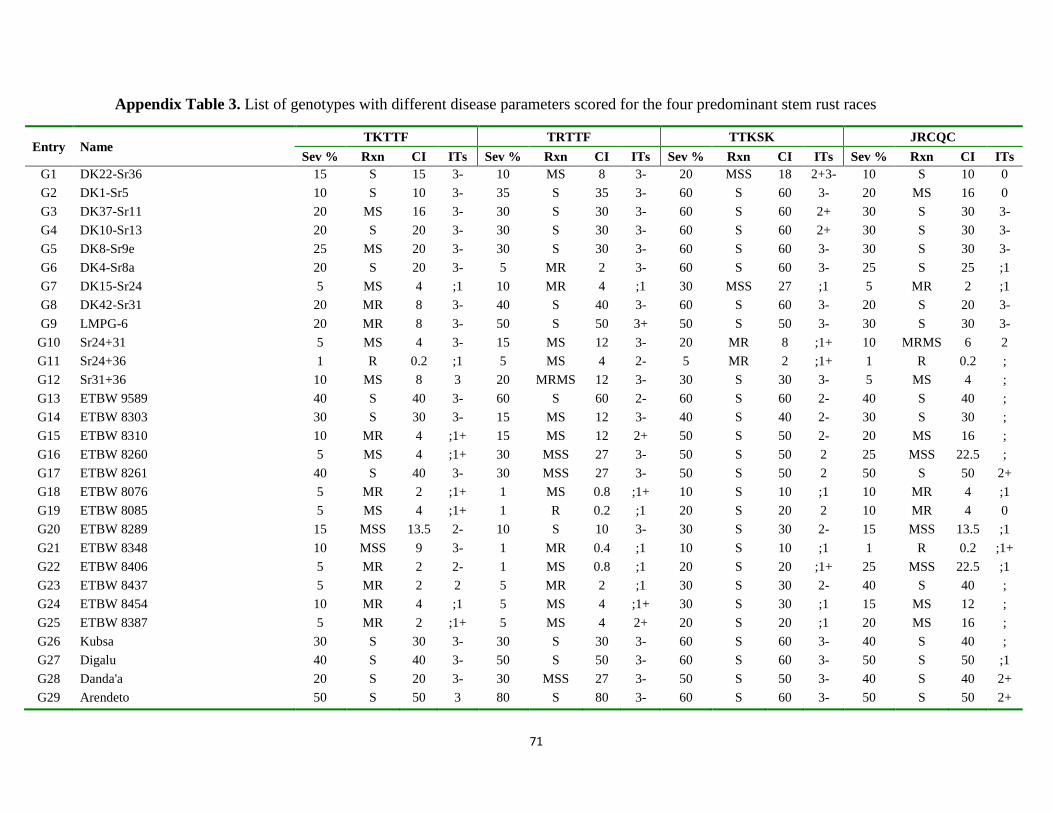
71
Appendix Table 3. List of genotypes with different disease parameters scored for the four predominant stem rust races
Entry Name TKTTF TRTTF TTKSK JRCQC
Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs
G1 DK22-Sr36 15 S 15 3- 10 MS 8 3- 20 MSS 18 2+3- 10 S 10 0
G2 DK1-Sr5 10 S 10 3- 35 S 35 3- 60 S 60 3- 20 MS 16 0
G3 DK37-Sr11 20 MS 16 3- 30 S 30 3- 60 S 60 2+ 30 S 30 3-
G4 DK10-Sr13 20 S 20 3- 30 S 30 3- 60 S 60 2+ 30 S 30 3-
G5 DK8-Sr9e 25 MS 20 3- 30 S 30 3- 60 S 60 3- 30 S 30 3-
G6 DK4-Sr8a 20 S 20 3- 5 MR 2 3- 60 S 60 3- 25 S 25 ;1
G7 DK15-Sr24 5 MS 4 ;1 10 MR 4 ;1 30 MSS 27 ;1 5 MR 2 ;1
G8 DK42-Sr31 20 MR 8 3- 40 S 40 3- 60 S 60 3- 20 S 20 3-
G9 LMPG-6 20 MR 8 3- 50 S 50 3+ 50 S 50 3- 30 S 30 3-
G10 Sr24+31 5 MS 4 3- 15 MS 12 3- 20 MR 8 ;1+ 10 MRMS 6 2
G11 Sr24+36 1 R 0.2 ;1 5 MS 4 2- 5 MR 2 ;1+ 1 R 0.2 ;
G12 Sr31+36 10 MS 8 3 20 MRMS 12 3- 30 S 30 3- 5 MS 4 ;
G13 ETBW 9589 40 S 40 3- 60 S 60 2- 60 S 60 2- 40 S 40 ;
G14 ETBW 8303 30 S 30 3- 15 MS 12 3- 40 S 40 2- 30 S 30 ;
G15 ETBW 8310 10 MR 4 ;1+ 15 MS 12 2+ 50 S 50 2- 20 MS 16 ;
G16 ETBW 8260 5 MS 4 ;1+ 30 MSS 27 3- 50 S 50 2 25 MSS 22.5 ;
G17 ETBW 8261 40 S 40 3- 30 MSS 27 3- 50 S 50 2 50 S 50 2+
G18 ETBW 8076 5 MR 2 ;1+ 1 MS 0.8 ;1+ 10 S 10 ;1 10 MR 4 ;1
G19 ETBW 8085 5 MS 4 ;1+ 1 R 0.2 ;1 20 S 20 2 10 MR 4 0
G20 ETBW 8289 15 MSS 13.5 2- 10 S 10 3- 30 S 30 2- 15 MSS 13.5 ;1
G21 ETBW 8348 10 MSS 9 3- 1 MR 0.4 ;1 10 S 10 ;1 1 R 0.2 ;1+
G22 ETBW 8406 5 MR 2 2- 1 MS 0.8 ;1 20 S 20 ;1+ 25 MSS 22.5 ;1
G23 ETBW 8437 5 MR 2 2 5 MR 2 ;1 30 S 30 2- 40 S 40 ;
G24 ETBW 8454 10 MR 4 ;1 5 MS 4 ;1+ 30 S 30 ;1 15 MS 12 ;
G25 ETBW 8387 5 MR 2 ;1+ 5 MS 4 2+ 20 S 20 ;1 20 MS 16 ;
G26 Kubsa 30 S 30 3- 30 S 30 3- 60 S 60 3- 40 S 40 ;
G27 Digalu 40 S 40 3- 50 S 50 3- 60 S 60 3- 50 S 50 ;1
G28 Danda'a 20 S 20 3- 30 MSS 27 3- 50 S 50 3- 40 S 40 2+
G29 Arendeto 50 S 50 3 80 S 80 3- 60 S 60 3- 50 S 50 2+
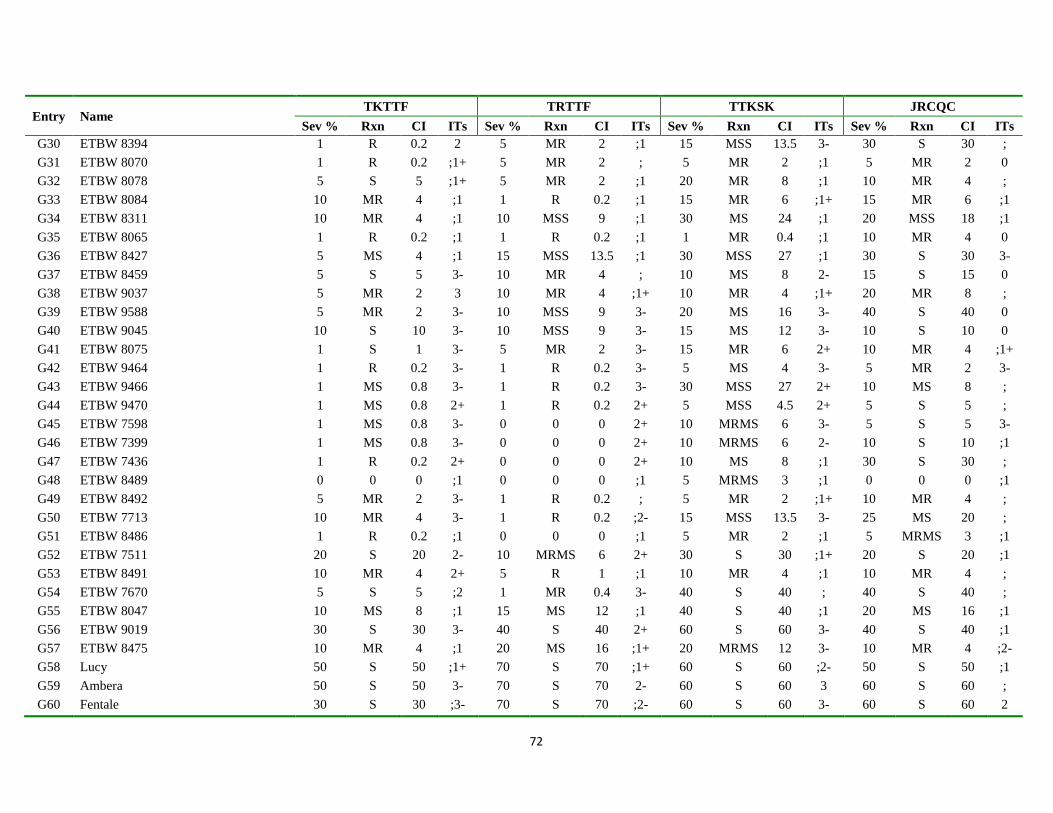
72
Entry Name TKTTF TRTTF TTKSK JRCQC
Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs
G30 ETBW 8394 1 R 0.2 2 5 MR 2 ;1 15 MSS 13.5 3- 30 S 30 ;
G31 ETBW 8070 1 R 0.2 ;1+ 5 MR 2 ; 5 MR 2 ;1 5 MR 2 0
G32 ETBW 8078 5 S 5 ;1+ 5 MR 2 ;1 20 MR 8 ;1 10 MR 4 ;
G33 ETBW 8084 10 MR 4 ;1 1 R 0.2 ;1 15 MR 6 ;1+ 15 MR 6 ;1
G34 ETBW 8311 10 MR 4 ;1 10 MSS 9 ;1 30 MS 24 ;1 20 MSS 18 ;1
G35 ETBW 8065 1 R 0.2 ;1 1 R 0.2 ;1 1 MR 0.4 ;1 10 MR 4 0
G36 ETBW 8427 5 MS 4 ;1 15 MSS 13.5 ;1 30 MSS 27 ;1 30 S 30 3-
G37 ETBW 8459 5 S 5 3- 10 MR 4 ; 10 MS 8 2- 15 S 15 0
G38 ETBW 9037 5 MR 2 3 10 MR 4 ;1+ 10 MR 4 ;1+ 20 MR 8 ;
G39 ETBW 9588 5 MR 2 3- 10 MSS 9 3- 20 MS 16 3- 40 S 40 0
G40 ETBW 9045 10 S 10 3- 10 MSS 9 3- 15 MS 12 3- 10 S 10 0
G41 ETBW 8075 1 S 1 3- 5 MR 2 3- 15 MR 6 2+ 10 MR 4 ;1+
G42 ETBW 9464 1 R 0.2 3- 1 R 0.2 3- 5 MS 4 3- 5 MR 2 3-
G43 ETBW 9466 1 MS 0.8 3- 1 R 0.2 3- 30 MSS 27 2+ 10 MS 8 ;
G44 ETBW 9470 1 MS 0.8 2+ 1 R 0.2 2+ 5 MSS 4.5 2+ 5 S 5 ;
G45 ETBW 7598 1 MS 0.8 3- 0 0 0 2+ 10 MRMS 6 3- 5 S 5 3-
G46 ETBW 7399 1 MS 0.8 3- 0 0 0 2+ 10 MRMS 6 2- 10 S 10 ;1
G47 ETBW 7436 1 R 0.2 2+ 0 0 0 2+ 10 MS 8 ;1 30 S 30 ;
G48 ETBW 8489 0 0 0 ;1 0 0 0 ;1 5 MRMS 3 ;1 0 0 0 ;1
G49 ETBW 8492 5 MR 2 3- 1 R 0.2 ; 5 MR 2 ;1+ 10 MR 4 ;
G50 ETBW 7713 10 MR 4 3- 1 R 0.2 ;2- 15 MSS 13.5 3- 25 MS 20 ;
G51 ETBW 8486 1 R 0.2 ;1 0 0 0 ;1 5 MR 2 ;1 5 MRMS 3 ;1
G52 ETBW 7511 20 S 20 2- 10 MRMS 6 2+ 30 S 30 ;1+ 20 S 20 ;1
G53 ETBW 8491 10 MR 4 2+ 5 R 1 ;1 10 MR 4 ;1 10 MR 4 ;
G54 ETBW 7670 5 S 5 ;2 1 MR 0.4 3- 40 S 40 ; 40 S 40 ;
G55 ETBW 8047 10 MS 8 ;1 15 MS 12 ;1 40 S 40 ;1 20 MS 16 ;1
G56 ETBW 9019 30 S 30 3- 40 S 40 2+ 60 S 60 3- 40 S 40 ;1
G57 ETBW 8475 10 MR 4 ;1 20 MS 16 ;1+ 20 MRMS 12 3- 10 MR 4 ;2-
G58 Lucy 50 S 50 ;1+ 70 S 70 ;1+ 60 S 60 ;2- 50 S 50 ;1
G59 Ambera 50 S 50 3- 70 S 70 2- 60 S 60 3 60 S 60 ;
G60 Fentale 30 S 30 ;3- 70 S 70 ;2- 60 S 60 3- 60 S 60 2

73
Entry Name TKTTF TRTTF TTKSK JRCQC
Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs
G61 Gambo 40 S 40 3- 50 S 50 ;1+ 60 S 60 2+3- 50 S 50 ;2-
G62 Werer-1 10 MR 4 3- 5 MS 4 2+ 40 S 40 3- 40 S 40 ;1+
G63 Werer-2 20 S 20 3- 30 S 30 2+ 30 S 30 2- 30 S 30 2+
G64 Wane 10 MR 4 ;1+ 20 MR 8 ;1+ 15 MR 6 3- 10 MR 4 ;1+
G65 Lemu 1 MS 0.8 3- 1 MR 0.4 ;2- 5 MR 2 ;1 1 R 0.2 0
G66 ETBW 9587 10 MS 8 2- 1 MR 0.4 2- 10 MR 4 ;2- 10 S 10 0
G67 Kingbird 10 MR 4 2+ 5 MR 2 ;2- 10 MRMS 6 2+ 10 MR 4 ;
G68 Liben 10 MR 4 ;2 10 MR 4 ;1 50 S 50 ;1 30 S 30 ;1
G69 Bulluk 1 R 0.2 ;1 1 MR 0.4 ;1+ 5 MR 2 ;1 1 R 0.2 ;
G70 Hobora 5 MR 2 ;1 1 R 0.2 ;1 5 MR 2 ;1 1 R 0.2 ;
G71 Dembel 5 MR 2 ;1 1 R 0.2 ;1 10 MR 4 ;1 5 MR 2 ;
G72 Honkolo 5 MR 2 ;1 1 R 0.2 ;1 15 MR 6 ;1 5 MR 2 ;
G73 Biqa 10 MRMS 6 3- 20 MSS 18 ;1+ 35 S 35 2+3- 10 S 10 ;1
G74 Sanate 10 MRMS 6 ;1 5 MR 2 ;1 20 MS 16 ;1 1 R 0.2 ;
G75 Mandoyu 5 MR 2 ;1 5 MR 2 ;1 20 MS 16 ;1 5 MR 2 ;1
G76 Hidasse 5 MR 2 3- 30 MSS 27 3- 40 S 40 2+ 10 MR 4 ;
G77 Ogolcho 1 R 0.2 ;1+ 1 MR 0.4 ;1+ 15 MR 6 ;1 10 MR 4 ;
G78 Hulluka 1 R 0.2 ;1 1 R 0.2 ;1 5 MR 2 ;1 1 R 0.2 ;1
G79 Shorima 1 R 0.2 2- 1 MRMS 0.6 ; 10 MR 4 ;1 5 MR 2 ;
G80 ETBW 6809 25 S 25 2+3- 30 S 30 3- 60 S 60 ;1 50 S 50 ;2+
G81 ETBW 6825 40 S 40 3- 20 S 20 3- 60 S 60 3- 60 S 60 ;1
G82 ETBW 7687 30 S 30 3- 15 MSS 13.5 2- 60 S 60 3- 50 S 50 ;1
G83 ETBW 7690 30 S 30 3- 15 MSS 13.5 3- 45 S 45 3- 40 S 40 ;
G84 BW candidate-2 1 R 0.2 ;1 1 R 0.2 ;1 1 R 0.2 ;1+ 5 MR 2 ;
G85 ETBW 6785 0 0 0 ;1 1 MR 0.4 ;1 1 R 0.2 ;1 1 MR 0.4 ;
G86 DW candidat-2 5 MR 2 2+ 30 MSS 27 3- 10 MS 8 2+ 10 MSS 9 2+
G87 DW candidat-1 10 MRMS 6 23- 40 S 40 3- 25 MS 20 2+ 20 MSS 18 ;1+
G88 BW candidate-1 10 MRMS 6 ;2- 30 MS 24 ;1+ 20 MRMS 12 2+ 15 MR 6 ;1
G89 F2-D/off/173/2010-81-2-3H 5 MR 2 2- 15 MRMS 9 ;2- 20 MSS 18 2 30 S 30 ;1
G90 CD13DZOS F6SR 2013 MS DZLS-1 10 MS 8 ;2+ 15 MSS 13.5 2 10 MRMS 6 2+3- 1 R 0.2 ;2+
G91 F2-D/off/149/2010-80-1-1H 20 S 20 ;1+ 30 MSS 27 2- 30 MS 24 2+ 20 MSS 18 ;1
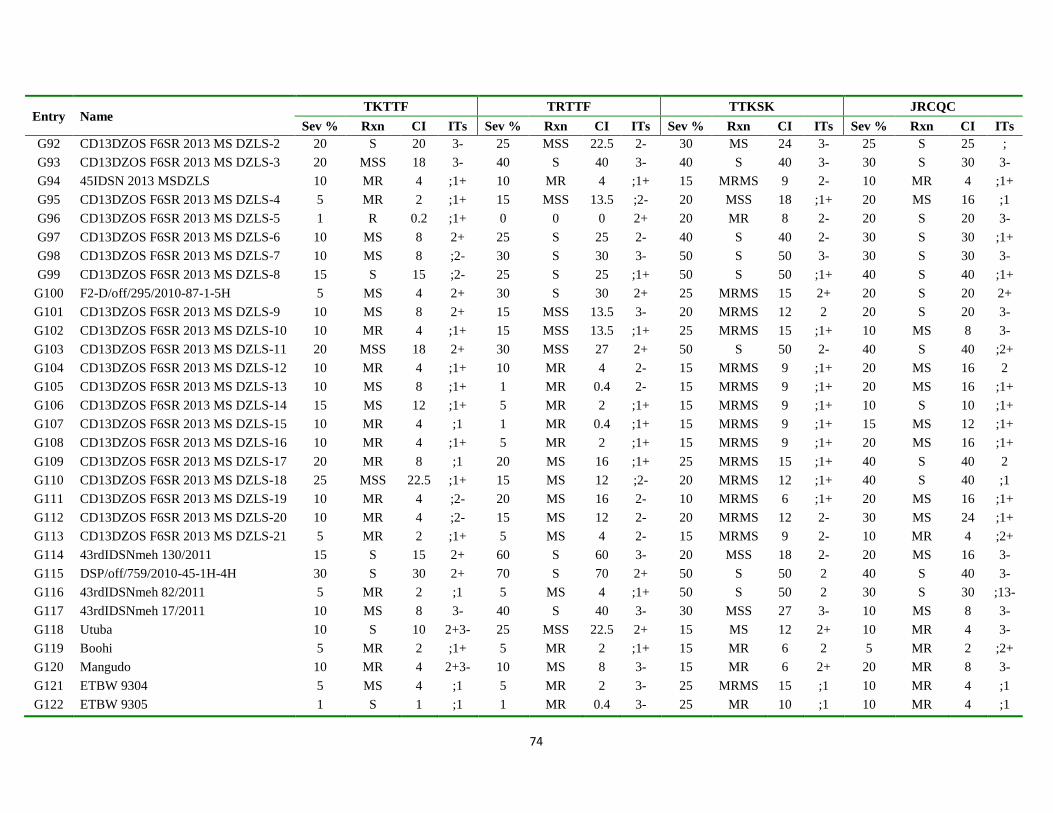
74
Entry Name TKTTF TRTTF TTKSK JRCQC
Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs
G92 CD13DZOS F6SR 2013 MS DZLS-2 20 S 20 3- 25 MSS 22.5 2- 30 MS 24 3- 25 S 25 ;
G93 CD13DZOS F6SR 2013 MS DZLS-3 20 MSS 18 3- 40 S 40 3- 40 S 40 3- 30 S 30 3-
G94 45IDSN 2013 MSDZLS 10 MR 4 ;1+ 10 MR 4 ;1+ 15 MRMS 9 2- 10 MR 4 ;1+
G95 CD13DZOS F6SR 2013 MS DZLS-4 5 MR 2 ;1+ 15 MSS 13.5 ;2- 20 MSS 18 ;1+ 20 MS 16 ;1
G96 CD13DZOS F6SR 2013 MS DZLS-5 1 R 0.2 ;1+ 0 0 0 2+ 20 MR 8 2- 20 S 20 3-
G97 CD13DZOS F6SR 2013 MS DZLS-6 10 MS 8 2+ 25 S 25 2- 40 S 40 2- 30 S 30 ;1+
G98 CD13DZOS F6SR 2013 MS DZLS-7 10 MS 8 ;2- 30 S 30 3- 50 S 50 3- 30 S 30 3-
G99 CD13DZOS F6SR 2013 MS DZLS-8 15 S 15 ;2- 25 S 25 ;1+ 50 S 50 ;1+ 40 S 40 ;1+
G100 F2-D/off/295/2010-87-1-5H 5 MS 4 2+ 30 S 30 2+ 25 MRMS 15 2+ 20 S 20 2+
G101 CD13DZOS F6SR 2013 MS DZLS-9 10 MS 8 2+ 15 MSS 13.5 3- 20 MRMS 12 2 20 S 20 3-
G102 CD13DZOS F6SR 2013 MS DZLS-10 10 MR 4 ;1+ 15 MSS 13.5 ;1+ 25 MRMS 15 ;1+ 10 MS 8 3-
G103 CD13DZOS F6SR 2013 MS DZLS-11 20 MSS 18 2+ 30 MSS 27 2+ 50 S 50 2- 40 S 40 ;2+
G104 CD13DZOS F6SR 2013 MS DZLS-12 10 MR 4 ;1+ 10 MR 4 2- 15 MRMS 9 ;1+ 20 MS 16 2
G105 CD13DZOS F6SR 2013 MS DZLS-13 10 MS 8 ;1+ 1 MR 0.4 2- 15 MRMS 9 ;1+ 20 MS 16 ;1+
G106 CD13DZOS F6SR 2013 MS DZLS-14 15 MS 12 ;1+ 5 MR 2 ;1+ 15 MRMS 9 ;1+ 10 S 10 ;1+
G107 CD13DZOS F6SR 2013 MS DZLS-15 10 MR 4 ;1 1 MR 0.4 ;1+ 15 MRMS 9 ;1+ 15 MS 12 ;1+
G108 CD13DZOS F6SR 2013 MS DZLS-16 10 MR 4 ;1+ 5 MR 2 ;1+ 15 MRMS 9 ;1+ 20 MS 16 ;1+
G109 CD13DZOS F6SR 2013 MS DZLS-17 20 MR 8 ;1 20 MS 16 ;1+ 25 MRMS 15 ;1+ 40 S 40 2
G110 CD13DZOS F6SR 2013 MS DZLS-18 25 MSS 22.5 ;1+ 15 MS 12 ;2- 20 MRMS 12 ;1+ 40 S 40 ;1
G111 CD13DZOS F6SR 2013 MS DZLS-19 10 MR 4 ;2- 20 MS 16 2- 10 MRMS 6 ;1+ 20 MS 16 ;1+
G112 CD13DZOS F6SR 2013 MS DZLS-20 10 MR 4 ;2- 15 MS 12 2- 20 MRMS 12 2- 30 MS 24 ;1+
G113 CD13DZOS F6SR 2013 MS DZLS-21 5 MR 2 ;1+ 5 MS 4 2- 15 MRMS 9 2- 10 MR 4 ;2+
G114 43rdIDSNmeh 130/2011 15 S 15 2+ 60 S 60 3- 20 MSS 18 2- 20 MS 16 3-
G115 DSP/off/759/2010-45-1H-4H 30 S 30 2+ 70 S 70 2+ 50 S 50 2 40 S 40 3-
G116 43rdIDSNmeh 82/2011 5 MR 2 ;1 5 MS 4 ;1+ 50 S 50 2 30 S 30 ;13-
G117 43rdIDSNmeh 17/2011 10 MS 8 3- 40 S 40 3- 30 MSS 27 3- 10 MS 8 3-
G118 Utuba 10 S 10 2+3- 25 MSS 22.5 2+ 15 MS 12 2+ 10 MR 4 3-
G119 Boohi 5 MR 2 ;1+ 5 MR 2 ;1+ 15 MR 6 2 5 MR 2 ;2+
G120 Mangudo 10 MR 4 2+3- 10 MS 8 3- 15 MR 6 2+ 20 MR 8 3-
G121 ETBW 9304 5 MS 4 ;1 5 MR 2 3- 25 MRMS 15 ;1 10 MR 4 ;1
G122 ETBW 9305 1 S 1 ;1 1 MR 0.4 3- 25 MR 10 ;1 10 MR 4 ;1
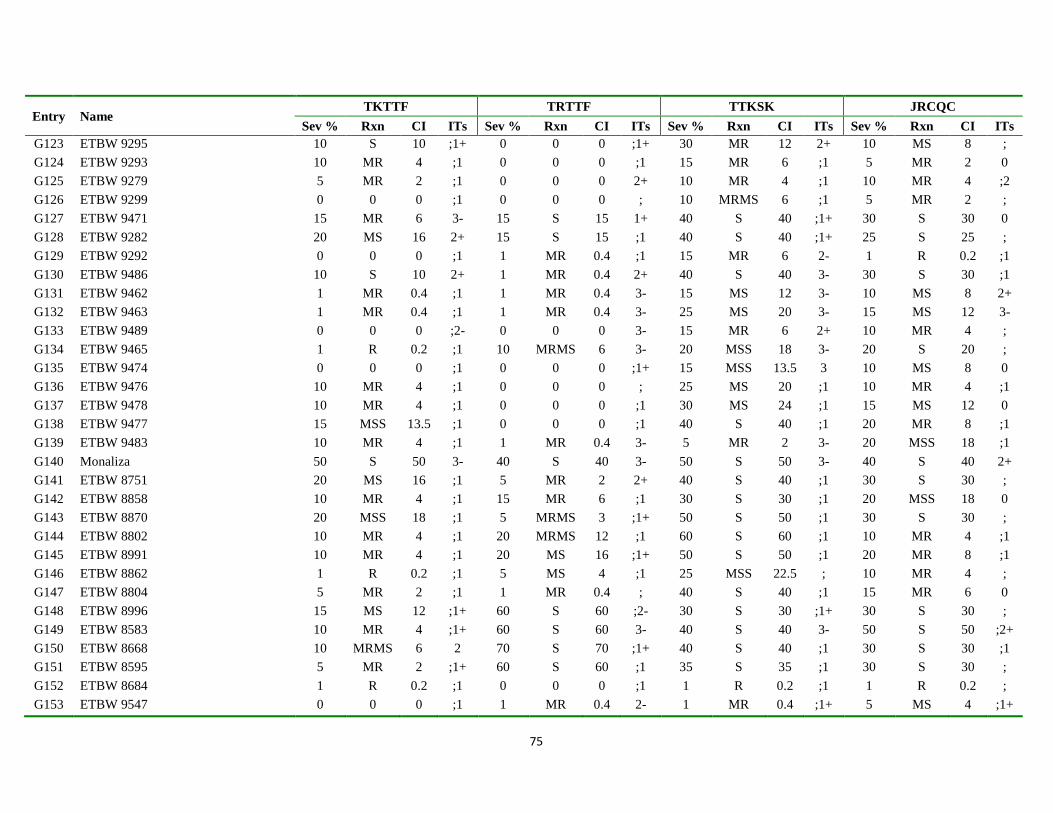
75
Entry Name TKTTF TRTTF TTKSK JRCQC
Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs
G123 ETBW 9295 10 S 10 ;1+ 0 0 0 ;1+ 30 MR 12 2+ 10 MS 8 ;
G124 ETBW 9293 10 MR 4 ;1 0 0 0 ;1 15 MR 6 ;1 5 MR 2 0
G125 ETBW 9279 5 MR 2 ;1 0 0 0 2+ 10 MR 4 ;1 10 MR 4 ;2
G126 ETBW 9299 0 0 0 ;1 0 0 0 ; 10 MRMS 6 ;1 5 MR 2 ;
G127 ETBW 9471 15 MR 6 3- 15 S 15 1+ 40 S 40 ;1+ 30 S 30 0
G128 ETBW 9282 20 MS 16 2+ 15 S 15 ;1 40 S 40 ;1+ 25 S 25 ;
G129 ETBW 9292 0 0 0 ;1 1 MR 0.4 ;1 15 MR 6 2- 1 R 0.2 ;1
G130 ETBW 9486 10 S 10 2+ 1 MR 0.4 2+ 40 S 40 3- 30 S 30 ;1
G131 ETBW 9462 1 MR 0.4 ;1 1 MR 0.4 3- 15 MS 12 3- 10 MS 8 2+
G132 ETBW 9463 1 MR 0.4 ;1 1 MR 0.4 3- 25 MS 20 3- 15 MS 12 3-
G133 ETBW 9489 0 0 0 ;2- 0 0 0 3- 15 MR 6 2+ 10 MR 4 ;
G134 ETBW 9465 1 R 0.2 ;1 10 MRMS 6 3- 20 MSS 18 3- 20 S 20 ;
G135 ETBW 9474 0 0 0 ;1 0 0 0 ;1+ 15 MSS 13.5 3 10 MS 8 0
G136 ETBW 9476 10 MR 4 ;1 0 0 0 ; 25 MS 20 ;1 10 MR 4 ;1
G137 ETBW 9478 10 MR 4 ;1 0 0 0 ;1 30 MS 24 ;1 15 MS 12 0
G138 ETBW 9477 15 MSS 13.5 ;1 0 0 0 ;1 40 S 40 ;1 20 MR 8 ;1
G139 ETBW 9483 10 MR 4 ;1 1 MR 0.4 3- 5 MR 2 3- 20 MSS 18 ;1
G140 Monaliza 50 S 50 3- 40 S 40 3- 50 S 50 3- 40 S 40 2+
G141 ETBW 8751 20 MS 16 ;1 5 MR 2 2+ 40 S 40 ;1 30 S 30 ;
G142 ETBW 8858 10 MR 4 ;1 15 MR 6 ;1 30 S 30 ;1 20 MSS 18 0
G143 ETBW 8870 20 MSS 18 ;1 5 MRMS 3 ;1+ 50 S 50 ;1 30 S 30 ;
G144 ETBW 8802 10 MR 4 ;1 20 MRMS 12 ;1 60 S 60 ;1 10 MR 4 ;1
G145 ETBW 8991 10 MR 4 ;1 20 MS 16 ;1+ 50 S 50 ;1 20 MR 8 ;1
G146 ETBW 8862 1 R 0.2 ;1 5 MS 4 ;1 25 MSS 22.5 ; 10 MR 4 ;
G147 ETBW 8804 5 MR 2 ;1 1 MR 0.4 ; 40 S 40 ;1 15 MR 6 0
G148 ETBW 8996 15 MS 12 ;1+ 60 S 60 ;2- 30 S 30 ;1+ 30 S 30 ;
G149 ETBW 8583 10 MR 4 ;1+ 60 S 60 3- 40 S 40 3- 50 S 50 ;2+
G150 ETBW 8668 10 MRMS 6 2 70 S 70 ;1+ 40 S 40 ;1 30 S 30 ;1
G151 ETBW 8595 5 MR 2 ;1+ 60 S 60 ;1 35 S 35 ;1 30 S 30 ;
G152 ETBW 8684 1 R 0.2 ;1 0 0 0 ;1 1 R 0.2 ;1 1 R 0.2 ;
G153 ETBW 9547 0 0 0 ;1 1 MR 0.4 2- 1 MR 0.4 ;1+ 5 MS 4 ;1+
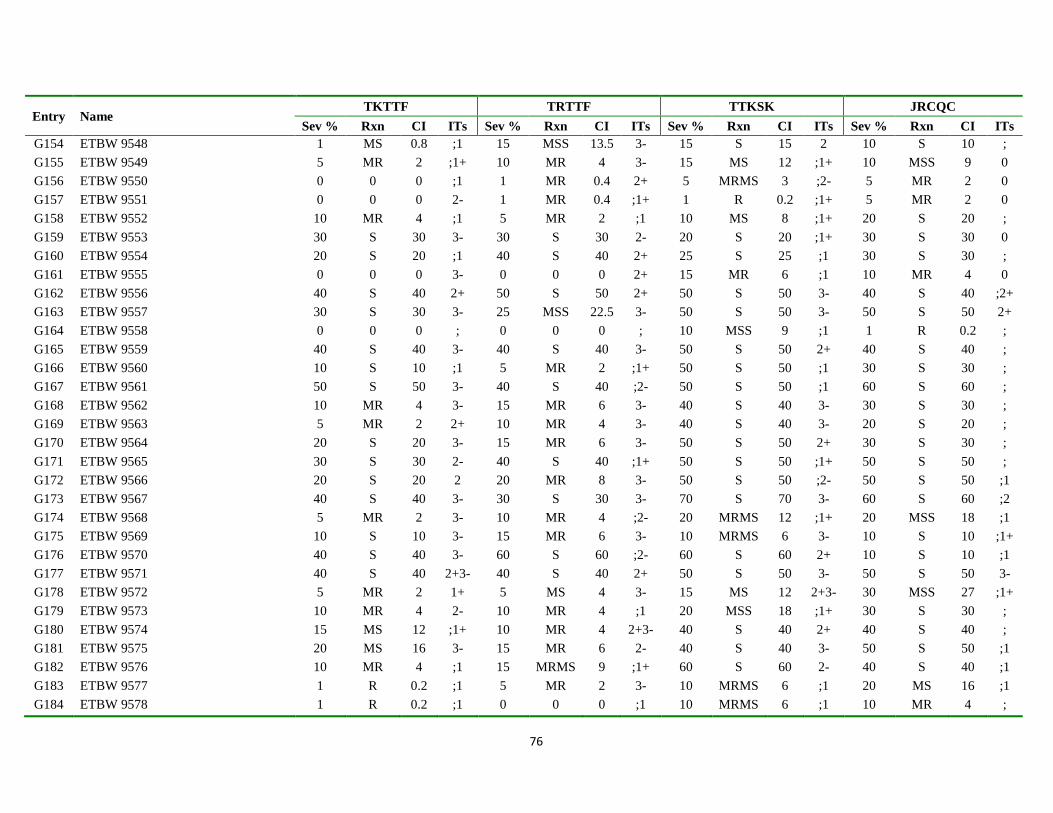
76
Entry Name TKTTF TRTTF TTKSK JRCQC
Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs
G154 ETBW 9548 1 MS 0.8 ;1 15 MSS 13.5 3- 15 S 15 2 10 S 10 ;
G155 ETBW 9549 5 MR 2 ;1+ 10 MR 4 3- 15 MS 12 ;1+ 10 MSS 9 0
G156 ETBW 9550 0 0 0 ;1 1 MR 0.4 2+ 5 MRMS 3 ;2- 5 MR 2 0
G157 ETBW 9551 0 0 0 2- 1 MR 0.4 ;1+ 1 R 0.2 ;1+ 5 MR 2 0
G158 ETBW 9552 10 MR 4 ;1 5 MR 2 ;1 10 MS 8 ;1+ 20 S 20 ;
G159 ETBW 9553 30 S 30 3- 30 S 30 2- 20 S 20 ;1+ 30 S 30 0
G160 ETBW 9554 20 S 20 ;1 40 S 40 2+ 25 S 25 ;1 30 S 30 ;
G161 ETBW 9555 0 0 0 3- 0 0 0 2+ 15 MR 6 ;1 10 MR 4 0
G162 ETBW 9556 40 S 40 2+ 50 S 50 2+ 50 S 50 3- 40 S 40 ;2+
G163 ETBW 9557 30 S 30 3- 25 MSS 22.5 3- 50 S 50 3- 50 S 50 2+
G164 ETBW 9558 0 0 0 ; 0 0 0 ; 10 MSS 9 ;1 1 R 0.2 ;
G165 ETBW 9559 40 S 40 3- 40 S 40 3- 50 S 50 2+ 40 S 40 ;
G166 ETBW 9560 10 S 10 ;1 5 MR 2 ;1+ 50 S 50 ;1 30 S 30 ;
G167 ETBW 9561 50 S 50 3- 40 S 40 ;2- 50 S 50 ;1 60 S 60 ;
G168 ETBW 9562 10 MR 4 3- 15 MR 6 3- 40 S 40 3- 30 S 30 ;
G169 ETBW 9563 5 MR 2 2+ 10 MR 4 3- 40 S 40 3- 20 S 20 ;
G170 ETBW 9564 20 S 20 3- 15 MR 6 3- 50 S 50 2+ 30 S 30 ;
G171 ETBW 9565 30 S 30 2- 40 S 40 ;1+ 50 S 50 ;1+ 50 S 50 ;
G172 ETBW 9566 20 S 20 2 20 MR 8 3- 50 S 50 ;2- 50 S 50 ;1
G173 ETBW 9567 40 S 40 3- 30 S 30 3- 70 S 70 3- 60 S 60 ;2
G174 ETBW 9568 5 MR 2 3- 10 MR 4 ;2- 20 MRMS 12 ;1+ 20 MSS 18 ;1
G175 ETBW 9569 10 S 10 3- 15 MR 6 3- 10 MRMS 6 3- 10 S 10 ;1+
G176 ETBW 9570 40 S 40 3- 60 S 60 ;2- 60 S 60 2+ 10 S 10 ;1
G177 ETBW 9571 40 S 40 2+3- 40 S 40 2+ 50 S 50 3- 50 S 50 3-
G178 ETBW 9572 5 MR 2 1+ 5 MS 4 3- 15 MS 12 2+3- 30 MSS 27 ;1+
G179 ETBW 9573 10 MR 4 2- 10 MR 4 ;1 20 MSS 18 ;1+ 30 S 30 ;
G180 ETBW 9574 15 MS 12 ;1+ 10 MR 4 2+3- 40 S 40 2+ 40 S 40 ;
G181 ETBW 9575 20 MS 16 3- 15 MR 6 2- 40 S 40 3- 50 S 50 ;1
G182 ETBW 9576 10 MR 4 ;1 15 MRMS 9 ;1+ 60 S 60 2- 40 S 40 ;1
G183 ETBW 9577 1 R 0.2 ;1 5 MR 2 3- 10 MRMS 6 ;1 20 MS 16 ;1
G184 ETBW 9578 1 R 0.2 ;1 0 0 0 ;1 10 MRMS 6 ;1 10 MR 4 ;
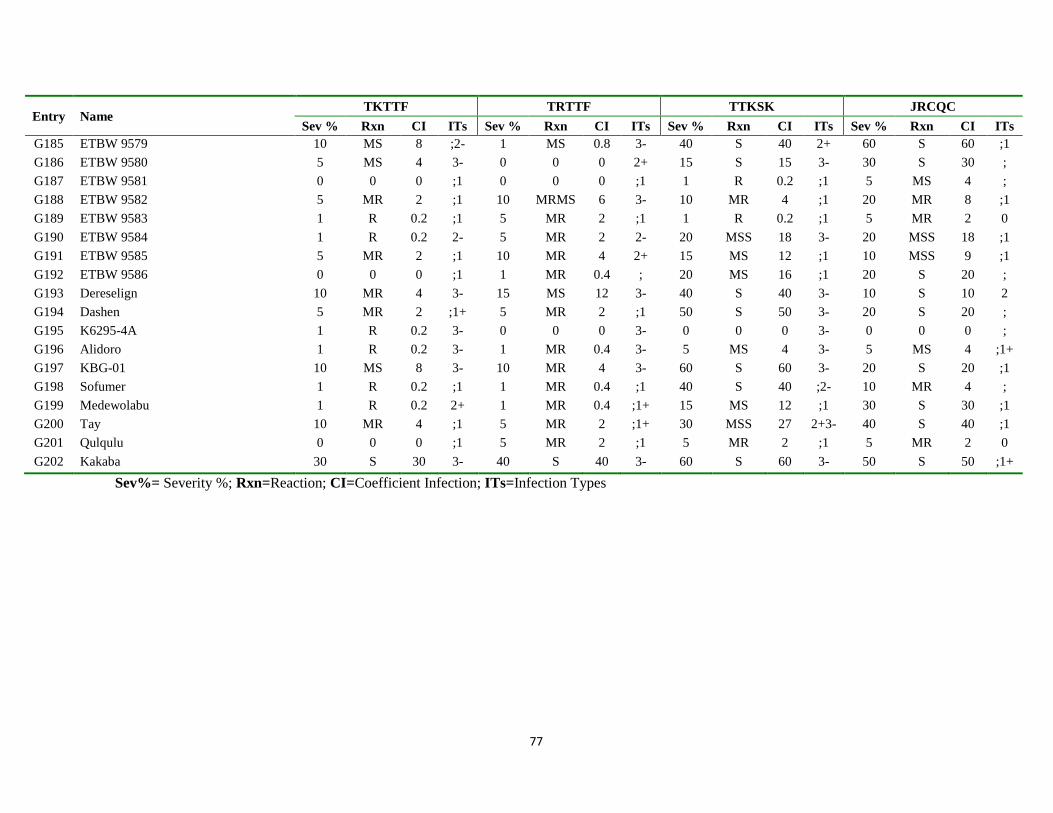
77
Entry Name TKTTF TRTTF TTKSK JRCQC
Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs Sev % Rxn CI ITs
G185 ETBW 9579 10 MS 8 ;2- 1 MS 0.8 3- 40 S 40 2+ 60 S 60 ;1
G186 ETBW 9580 5 MS 4 3- 0 0 0 2+ 15 S 15 3- 30 S 30 ;
G187 ETBW 9581 0 0 0 ;1 0 0 0 ;1 1 R 0.2 ;1 5 MS 4 ;
G188 ETBW 9582 5 MR 2 ;1 10 MRMS 6 3- 10 MR 4 ;1 20 MR 8 ;1
G189 ETBW 9583 1 R 0.2 ;1 5 MR 2 ;1 1 R 0.2 ;1 5 MR 2 0
G190 ETBW 9584 1 R 0.2 2- 5 MR 2 2- 20 MSS 18 3- 20 MSS 18 ;1
G191 ETBW 9585 5 MR 2 ;1 10 MR 4 2+ 15 MS 12 ;1 10 MSS 9 ;1
G192 ETBW 9586 0 0 0 ;1 1 MR 0.4 ; 20 MS 16 ;1 20 S 20 ;
G193 Dereselign 10 MR 4 3- 15 MS 12 3- 40 S 40 3- 10 S 10 2
G194 Dashen 5 MR 2 ;1+ 5 MR 2 ;1 50 S 50 3- 20 S 20 ;
G195 K6295-4A 1 R 0.2 3- 0 0 0 3- 0 0 0 3- 0 0 0 ;
G196 Alidoro 1 R 0.2 3- 1 MR 0.4 3- 5 MS 4 3- 5 MS 4 ;1+
G197 KBG-01 10 MS 8 3- 10 MR 4 3- 60 S 60 3- 20 S 20 ;1
G198 Sofumer 1 R 0.2 ;1 1 MR 0.4 ;1 40 S 40 ;2- 10 MR 4 ;
G199 Medewolabu 1 R 0.2 2+ 1 MR 0.4 ;1+ 15 MS 12 ;1 30 S 30 ;1
G200 Tay 10 MR 4 ;1 5 MR 2 ;1+ 30 MSS 27 2+3- 40 S 40 ;1
G201 Qulqulu 0 0 0 ;1 5 MR 2 ;1 5 MR 2 ;1 5 MR 2 0
G202 Kakaba 30 S 30 3- 40 S 40 3- 60 S 60 3- 50 S 50 ;1+
Sev%= Severity %; Rxn=Reaction; CI=Coefficient Infection; ITs=Infection Types
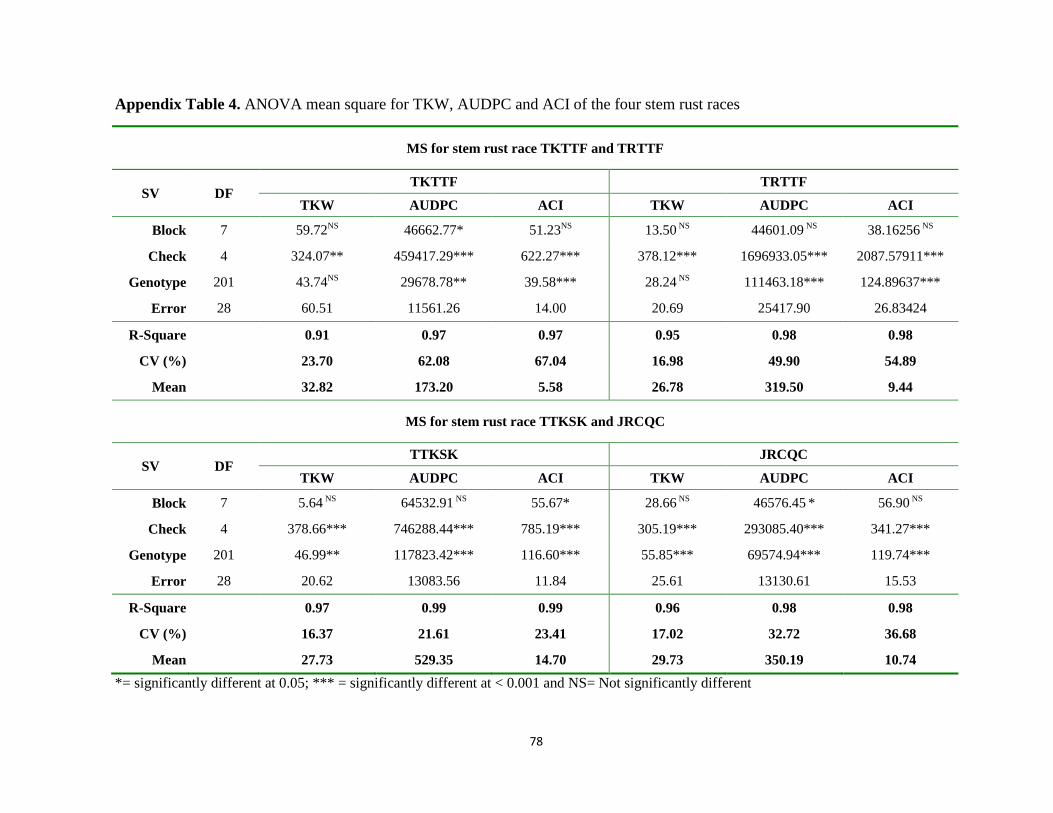
78
Appendix Table 4. ANOVA mean square for TKW, AUDPC and ACI of the four stem rust races
MS for stem rust race TKTTF and TRTTF
SV DF TKTTF TRTTF
TKW AUDPC ACI TKW AUDPC ACI
Block 7 59.72NS 46662.77* 51.23NS 13.50 NS 44601.09 NS 38.16256 NS
Check 4 324.07** 459417.29*** 622.27*** 378.12*** 1696933.05*** 2087.57911***
Genotype 201 43.74NS 29678.78** 39.58*** 28.24 NS 111463.18*** 124.89637***
Error 28 60.51 11561.26 14.00 20.69 25417.90 26.83424
R-Square
0.91 0.97 0.97 0.95 0.98 0.98
CV (%)
23.70 62.08 67.04 16.98 49.90 54.89
Mean
32.82 173.20 5.58 26.78 319.50 9.44
MS for stem rust race TTKSK and JRCQC
SV DF TTKSK JRCQC
TKW AUDPC ACI TKW AUDPC ACI
Block 7 5.64 NS 64532.91 NS 55.67* 28.66 NS 46576.45 * 56.90 NS
Check 4 378.66*** 746288.44*** 785.19*** 305.19*** 293085.40*** 341.27***
Genotype 201 46.99** 117823.42*** 116.60*** 55.85*** 69574.94*** 119.74***
Error 28 20.62 13083.56 11.84 25.61 13130.61 15.53
R-Square
0.97 0.99 0.99 0.96 0.98 0.98
CV (%)
16.37 21.61 23.41 17.02 32.72 36.68
Mean
27.73 529.35 14.70 29.73 350.19 10.74
*= significantly different at 0.05; *** = significantly different at < 0.001 and NS= Not significantly different





























