Embed Size (px)
Citation preview

www.elsevier.com/locate/ynimg
NeuroImage 30 (2006) 570 – 579
Cortical activation in the processing of passive sentences
in L1 and L2: An fMRI study
Satoru Yokoyama,a,b,* Hideyuki Okamoto,b,d Tadao Miyamoto,a Kei Yoshimoto,a Jungho Kim,f
Kazuki Iwata, b,c Hyeonjeong Jeong, a,b Shinya Uchida, b,e Naho Ikuta, b,c Yuko Sassa, b,c
Wataru Nakamura,a Kaoru Horie,a Shigeru Sato,a and Ryuta Kawashimab
aGraduate School of International Cultural Studies (GSICS), Tohoku University, Kawauchi, Aoba-ku, Sendai-shi, Miyagi-ken, 980-8576, JapanbNew Industry Creation Hatchery Center, Tohoku University, 6-6-10, Aoba, Aza, Aramaki, Aoba-ku, Sendai-shi, Miyagi-ken, 980-8579, JapancLBC Research Center, Tohoku University 21st Century Center of Excellence Program in Humanities, Kawauchi, Aoba-ku, Sendai-shi,
Miyagi-ken, 980-8576, JapandGraduate School of Medicine, Tohoku University, Seiryo-cho, Aoba-ku, Sendai-shi, Miyagi-ken, 980-8574, JapaneDepartment of Radiology and Nuclear Medicine, IDAC, Tohoku University, Seiryo-cho, Aoba-ku, Sendai-shi, Miyagi-ken, 980-8574, JapanfGraduate School of Arts and Letters, Tohoku University, Kawauchi, Aoba-ku, Sendai-shi, Miyagi-ken, 980-8576, Japan
Received 19 May 2005; revised 29 September 2005; accepted 29 September 2005
Available online 21 November 2005
The question of whether the bilingual brain processes a first and second
language (L1 and L2, respectively) differently is a central issue in many
psycholinguistic and neurolinguistic studies. This study used functional
magnetic resonance imaging (fMRI) to investigate whether late
bilinguals process structurally complex sentences in L1 and L2 in
different cortical networks. For this purpose, we directly compared
brain activity during the processing of active and passive sentences in
both L1 and L2. We asked 36 healthy subjects to judge whether or not a
presented sentence was semantically plausible. Both L1 andL2 activated
the left hemispheric language-related regions such as the left inferior
frontal, superior/middle temporal, and parietal cortices. However, we
found different activation patterns between L1 and L2 in the processing
of passive sentences. Passive sentences elicited greater activation than
their active counterparts in the left pars triangularis, the premotor area,
and the superior parietal lobule in Japanese, but not in English.
Furthermore, there was a significant interaction between sentence type
(active versus passive) and language (Japanese versus English) in the left
pars orbitalis. The results of this study indicate that late bilinguals use
similar cortical regions to comprehend both L1 and L2. However, when
late bilinguals are presented with structurally complex sentences, the
involvement of these regions differs between L1 and L2. These results
suggest that, in addition to age of L2 acquisition and L2 proficiency,
differences in grammatical construction affect cortical representation
during the comprehension of L1 and L2.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Bilingual; Sentence comprehension; Passive sentence; fMRI
1053-8119/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.neuroimage.2005.09.066
* Corresponding author. Graduate School of International Cultural
Studies (GSICS), Tohoku University, Kawauchi, Aoba-ku, Sendai-shi,
Miyagi-ken, 980-8576, Japan. Fax: +81 22 795 4088.
E-mail address: [email protected] (S. Yokoyama).
Available online on ScienceDirect (www.sciencedirect.com).
Introduction
Many psycholinguistic and neurolinguistic studies have inves-
tigated the question of how the bilingual brain processes a first and
second language (L1 and L2, respectively). For example, based on
their study of aphasics, Albert and Obler (1978, p. 254) have argued
that L2 is represented more globally than L1 and that the right
hemisphere plays an important role in L2 representation. Other
researchers have presented evidence that following brain injury,
multilinguals exhibit different degrees of damage (Fabbro and
Paradis, 1995) and recovery (Junque et al., 1995). Furthermore, in an
experiment using electrical cortical stimulation during brain surgery,
Ojemann and Whitaker (1978) showed that L2 is more broadly
represented in epileptic bilingual patients than in their monolingual
counterparts. Although these studies did not lead to the same
conclusions, the combined evidence suggests that different cortical
areas are involved in the comprehension of L1 and L2. Neuro-
imaging studies, however, have yielded inconsistent results on this
question. Some imaging studies have found different activation
patterns for L1 and L2 (e.g., Klein et al., 1994; Perani et al., 1996;
Dehaene et al., 1997), while others have reported no difference in
activation between L1 and L2 (e.g., Klein et al., 1995; Illes et al.,
1999). In this paper, we report on a functional magnetic resonance
imaging (fMRI) study that investigates how the bilingual brain
processes L1 and L2.
Previous studies of bilinguals have considered various facets of
language processing, including phonological, lexical, and sentential
processing. Recent neuroimaging studies on bilingual sentence
comprehension have yielded relatively consistent results. In
previous studies on bilinguals, age of acquisition and proficiency
level have been considered to be the main factors that cause different
brain activation patterns between L1 and L2. In an fMRI study of

(3) John-ga Mary-wo home-ta.
John-NOM Mary-ACC praise-PAST
‘‘John praised Mary.’’
(4) Mary-ga John-ni home-rare-ta.
Mary-NOM John-DAT (by) praise-PASS-PAST
‘‘Mary was praised by John.’’
S. Yokoyama et al. / NeuroImage 30 (2006) 570–579 571
early Mandarin-to-English bilinguals acquiring the two languages
simultaneously, Chee et al. (1999) investigated whether the
bilinguals showed identical activation patterns for L1 and L2 in a
visually presented sentence comprehension task. The study revealed
no significant difference in activation patterns for L1 and L2. By
contrast, in a study of late Mandarin-to-English bilinguals acquiring
L2 later than L1, Luke et al. (2002) found that compared to
Mandarin, English produced greater activation in the left frontal
region during a syntactic error detection task; and greater activation
in the superior temporal region during a semantic error detection
task. Wartenburger et al. (2003) compared Italian-to-German
bilinguals on syntactic and semantic error detection tasks. The
subjects varied in age of acquisition and proficiency; the three
groups were high-proficiency early bilinguals, high-proficiency late
bilinguals, and low-proficiency late bilinguals. Early bilinguals
showed no significant difference between L1 and L2 in either the
syntactic or the semantic error detection task. By contrast, among
late bilinguals, L2 elicited greater activation in the left frontal and
parietal areas than L1, regardless of proficiency level. Furthermore,
Wartenburger et al. (2003) found that low- versus high-proficiency
late bilinguals showed different activation patterns. Based on these
observations, wemay assume that age of acquisition and proficiency
are the main factors that determine the pattern of brain activation in
L1 and L2 processing among bilinguals.
However, a recent neuroimaging study on bilingual sentence
comprehension (Hasegawa et al., 2002) lets us speculate that
sentence complexity may be an additional factor affecting differ-
ences in activation between L1 and L2. Hasegawa et al. (2002) based
their study on the findings of Carpenter and Just (1975) and
Carpenter et al. (1999), who reported that a negative sentence
requires more processing than its affirmative counterpart. Hasegawa
et al. (2002) compared the comprehension of negative versus
affirmative sentences among Japanese-to-English late bilinguals.
They reported that English (L2) negative sentences showed greater
activation than their affirmative counterparts in the left inferior
frontal, superior/middle temporal, and parietal cortices. Interesting-
ly, however, Japanese (L1) negative sentences produced no more
activation than Japanese affirmative sentences, suggesting that
structurally complex sentences may be processed differently in L1
and L2.
In the present study, we compared late bilinguals’ processing of
structurally simple versus complex sentences in Japanese (L1) and
English (L2). For this purpose, we compared brain activity during
the processing of active and passive sentences within each language.
We chose to compare active and passive sentences because the
Japanese direct passive semantically parallels the English passive
(Shibatani, 1990). Furthermore, as explained in detail below, the
syntactic differences between active and passive sentences in both
English and Japanese are similar.
Most languages mark the passive by a change in both the verb
form and the relative order of the agent and the theme (cf. Shibatani,
1990; O’Grady and Dobrovolsky, 1996). As shown in (1) and (2)
below, the English passive sentence has three distinct properties
(‘‘AUX’’, ‘‘PAST’’, and ‘‘PASS’’ are abbreviations for auxiliary, past
tense, and passive, respectively).
(1) John praised Mary.
praise-PAST
(2) Mary was praised by John.
AUX-PAST praise-PASS
First, a passive sentence contains some form of the auxiliary
verb ‘‘be’’, together with a verb in the past participle form, which is
normally marked by the suffix -ed or -en. Second, the order of the
agent and the theme is reversed in passive sentences relative to
active sentences. While the agent precedes the theme in active
sentences, the theme precedes the agent in passive sentences. The
final property of the English passive is that the agent occurs as an
oblique in a by-phrase.
The analogous situation occurs with the Japanese direct passive
sentence. As shown in (3) and (4) below, the Japanese direct
passive has three defining properties that are similar to those of the
English passive (‘‘NOM’’, ‘‘ACC’’, and ‘‘DAT’’ are abbreviations
for nominative, accusative, and dative, respectively).
First, a direct passive sentence encodes the passive voice with a
morpheme (ra)re on the verb. Second, the relative order of
thematic roles is reversed. Third, the agent occurs as part of a noun
phrase with an oblique case, which is marked by -ni in direct
passive sentences (cf. Shibatani, 1985). Thus, a Japanese direct
passive sentence is very similar to an English passive sentence.
It is important to note, however, that English and Japanese
passives differ in a number of ways. For example, Japanese has
three passive constructions: the direct passive, the indirect passive,
and the possessive passive; while English has only one passive
construction. Also, English uses psychological verbs in the passive
form (e.g., I am frightened), while Japanese does not. Keeping
these differences in mind, in our study, we restricted our stimuli to
Japanese direct passive sentences and English passive sentences
without psychological verbs.
In the present study, we compared brain activity during the
processing of active and passive sentences within each language to
determine whether the comprehension of structurally complex
sentences differs in L1 and L2. If we find different activation
patterns for Japanese and English, our hypothesis that structurally
complex sentences are processed differently in L1 and L2 will be
supported.
In addition, we statistically excluded the effect of task difficulty
as a confounding covariate in the analysis of covariance
(ANCOVA) when we directly compared the comprehension of
L1 and L2. We took this measure to address the points raised by
Hasegawa et al. (2002), who suggested that task difficulty or
workload generates different cortical representations during the
comprehension of L1 versus L2. Based on Hasegawa et al. (2002),
we speculate that when the effect of task difficulty is statistically
excluded, there will be no difference in the activation pattern of the
left hemispheric language-related regions in the comprehension of
L1 and L2.
Materials and methods
Participants
Thirty-six university students (30 males and 6 females; aged
between 18 and 29 years; the mean age of the participants was 21.1

S. Yokoyama et al. / NeuroImage 30 (2006) 570–579572
years) participated in this study. All participants were native
Japanese speakers, and all were right-handed, as assessed by the
Edinburgh Handedness Inventory (Oldfield, 1971). None of the
participants displayed any signs, or had any previous history of,
medical or neurological diseases. We obtained written informed
consent from each participant in accordance with the guidelines
approved by Tohoku University and the Helsinki Declaration of
Human Rights, 1975. The mean age and length of acquisition of
English (L2) for these participants were 11.8 (range = 8–13, SD =
1.4) and 9.2 years (range = 6–17, SD = 2.6), respectively. Thus, all
participants were late bilinguals. We assessed the participants on the
basis of the pre-level 2 English Language Proficiency test, which
was prepared by the Society for Testing English Proficiency. This
test, which includes listening and reading components, is generally
used to assess the proficiency of English as a foreign language in
Japan. The test has seven grades: one, pre-one, two, pre-two, three,
four, and five; five refers to the lowest level of proficiency, and one
to the highest. At pre-level 2 proficiency, the level selected for the
participants in this study, speakers are assumed to have a sufficient
level of comprehension and production to participate in daily life in
English. The mean scores in the listening and reading sections of the
proficiency test were 66.9% (range = 60.0–92.5, SD = 13.4) and
90.1% (range = 80–100, SD = 6.9), respectively. The mean total
score for the test was 78.6% (range = 66–92, SD = 8.4).
Materials
The experimental stimuli were 72 short sentences, half of which
were Japanese, and half English. For each language, 12 sentences
were semantically plausible, 12 sentences were semantically
implausible, and the remaining 12 were control stimuli. Of the
plausible and implausible sentences, half were active and half were
passive. All sentences consisted of three phrases: two noun phrases
and one verb phrase. We used translation equivalents in both
languages to ensure a comparable lexical level. Table 1 contains
some examples.
We composed the stimulus sentences as follows. For the
English sentences, we chose nouns and verbs generally used at
the junior high school level to ensure the participants’ familiarity
with them. We then translated the selected words into Japanese
and checked and controlled for frequency using the database
developed by Amano and Kondo (1999). Prior to the fMRI
experiment, we performed a lexical decision screening test to
eliminate any lexical items that would be difficult for learners of
English as a second language to process. Ten additional healthy
right-handed subjects (aged between 20 and 26 years; the mean
age of the participants was 22.3 years) participated in this test.
The mean reaction time (SD) for selected words was 1248 (258)
ms. The control stimuli consisted of three nouns or verbs that
were included in the stimulus sentences. We did not present the
Table 1
Examples of stimuli used in the present experiment
Sentence type Example
Plausible active sentence The hunter/shot/the deer
Plausible passive sentence The deer/was shot/by the hunter
Implausible active sentence The deer/shot/the hunter
Implausible passive sentence The hunter/was shot/by the deer
Control stimulus The deer/the hunter/the deer
control stimuli in a sentential context. While half of the stimuli
were identical, the remaining half were not.
Task procedure
We presented the stimuli visually in a phrase-by-phrase
manner on a black projection screen. Fig. 1 illustrates this
procedure. Following an initial resting period (12 s), the stimuli
were presented in epochs lasting 28 s (18 epochs, pseudo-
randomized), followed by a 28-s resting period, during which a
fixation cross was presented. Prior to each epoch, an instruction
word was presented for 1 s (e.g., ‘‘bun’’, which means ‘‘sentence’’
in Japanese, for the sentence epoch; or ‘‘tan’’, which means
‘‘word’’ in Japanese, for the control epoch). Each epoch contained
four randomized trials. Each phrase was presented for 1 s.
Intervals for switching screens were inserted between phrases.
Plausible and implausible sentences were presented in the
sentence epoch. Immediately after the presentation of the third
phrase of a sentence, a fixation cross was displayed for a period
of 3.8 s. During this period, subjects were asked to press the left
button with their index finger if the sentence was plausible, and
the right button with their middle finger if the sentence was
implausible. Therefore, the intervals between the subject’s
response and the start of the next trial varied in relation to the
subject’s reaction time. In the control epoch, three noun phrases
or verb phrases were sequentially presented in a randomized
order. These phrases did not comprise a sentence. After the
presentation of the third phrase of these stimuli, a fixation cross
lasting 3.8 s was displayed. Subjects were then asked to press the
right button if the presented phrases were identical, and the left
button if the phrases were not identical. This control condition
was intended to control for the effects of visual input, working
memory, word recognition, and motor output. The order of the
conditions, such as the Japanese active, passive, and control, and
the English active, passive, and control conditions, was pseudo-
randomized and counterbalanced across subjects. We conducted a
brief practice session before performing the fMRI in order to
familiarize the participants with the task involved in the study.
This practice session consisted of 10 trials that were similar to the
task performed in this study.
fMRI data acquisition
We performed the fMRI on a 1.5 T Siemens Symphony
scanner (Siemens, Erlangen, Germany) at Tohoku University.
Head motion was minimized by placing pillows and cushions
around the head. Forty-two axial slices (thickness, 3 mm; FOV,
192 mm; data matrix, 64 � 64 voxels; in-plane resolution, 2 �2 mm) were acquired every 4 s during functional measurements
(BOLD-sensitive gradient EPI sequence; TR = 4000 ms; TE =
50 ms; flip angle = 90-). The seven initial scans were dummy
scans that were used to equilibrate the state of magnetization
and were excluded from the analysis. After the functional
imaging, anatomical images of T1-weighted MDEFT images
(thickness, 1 mm; FOV, 256 mm; data matrix, 192 � 224; TR =
1900 ms; TE = 3.93 ms) were also acquired from all
participants.
The accuracy rate and response time for all tasks used as
behavioral data were collected using a Windows-based com-
puter, which was used in the presentation of the stimuli for the
tasks.

Table 2
Behavioral data
Accuracy
rate (%)
SD Reaction
time (ms)
SD
Japanese active 95.5 21 1435 631
Japanese passive 94.9 17 1659 709
English active 91.6 26 1492 761
English passive 88.8 32 1692 902
Fig. 1. Procedure followed in the present experiment.
S. Yokoyama et al. / NeuroImage 30 (2006) 570–579 573
Data analysis
The fMRI time-series data were analyzed using SPM99
implemented on MATLAB (The Mathworks Inc., Natick, MA).
The data were realigned, spatially normalized to the standard brain
space, and smoothed with an isotropic Gaussian kernel of 9 mm
full width at half maximum using a standard SPM method (Friston
et al., 1995), and a high-pass frequency filter (128 s) was applied.
The time series were modeled using box-car regressors for each
epoch and convolved with the hemodynamic response function.
For each subject, we computed contrasts among (a) Japanese active
sentences; (b) Japanese passive sentences; (c) English active
sentences; (d) English passive sentences; (e) the Japanese control
task; and (f) the English control task.
We computed group effects using these contrast images with
the random effect model. Since the reaction time and the
accuracy rate exhibited significant statistical differences among
conditions (see Results), we included these values as a
confounding covariate in the ANCOVA in order to compensate
for the effect of task difficulty. Although there is a confound
involved in the different time intervals between the presentation
of active and passive sentences in both English and Japanese, we
assume that this confound corresponds to reaction time. An
anatomical location was identified using a template created by
Talairach and Tournoux (1988). In the contrasts of L1 plus L2
versus their controls and in the comparisons between L1 and L2,
a threshold was set at P < 0.05, corrected. In the comparisons of
task versus control conditions and task versus task conditions, a
threshold of P < 0.001, uncorrected, was established. Although
we did not perform correction for multiple comparisons in these
task versus task comparisons, we believe that the statistical results
were nevertheless conservative since these analyses employed the
random effect model.
However, we also performed ROI analyses to confirm statistical
significance and to determine the characteristics of each activated
area. In order to compare the intensity of activation, we defined
ROIs as the significantly activated clusters when contrasting
‘‘Japanese passive’’ versus ‘‘Japanese active’’ and ‘‘English
passive’’ versus ‘‘English active’’ (cf. Miura et al., 2005). Mean
percent signal changes were calculated in each ROI for each
subject in each condition. ANOVAs were conducted for all
conditions, and a post hoc multiple comparison (Bonferroni
method) was performed.
The data regarding accuracy rate and response time for all tasks
were analyzed by the analysis of variance (ANOVA). The data
were analyzed by ANOVA, and a post hoc multiple comparison
(Bonferroni method) was performed. When there was a significant
difference among conditions in the behavioral data, we performed
correlation analysis between the values of percent signal changes in
each ROI and the behavioral data, in order to confirm whether
there was a significant correlation between behavioral data and
brain activity for each ROI.
Results
Behavioral data
Table 2 summarizes the accuracy rate and reaction time. The
difference in the accuracy rates between the active and passive
sentence conditions was not statistically significant (P > 0.1), but
there was a significant difference between the Japanese and
English conditions (P < 0.001). In the post hoc analysis, there
was a significant difference between Japanese and English in both
the active and passive sentence conditions (P < 0.01 and P < 0.001
for active and passive sentences, respectively). The difference in
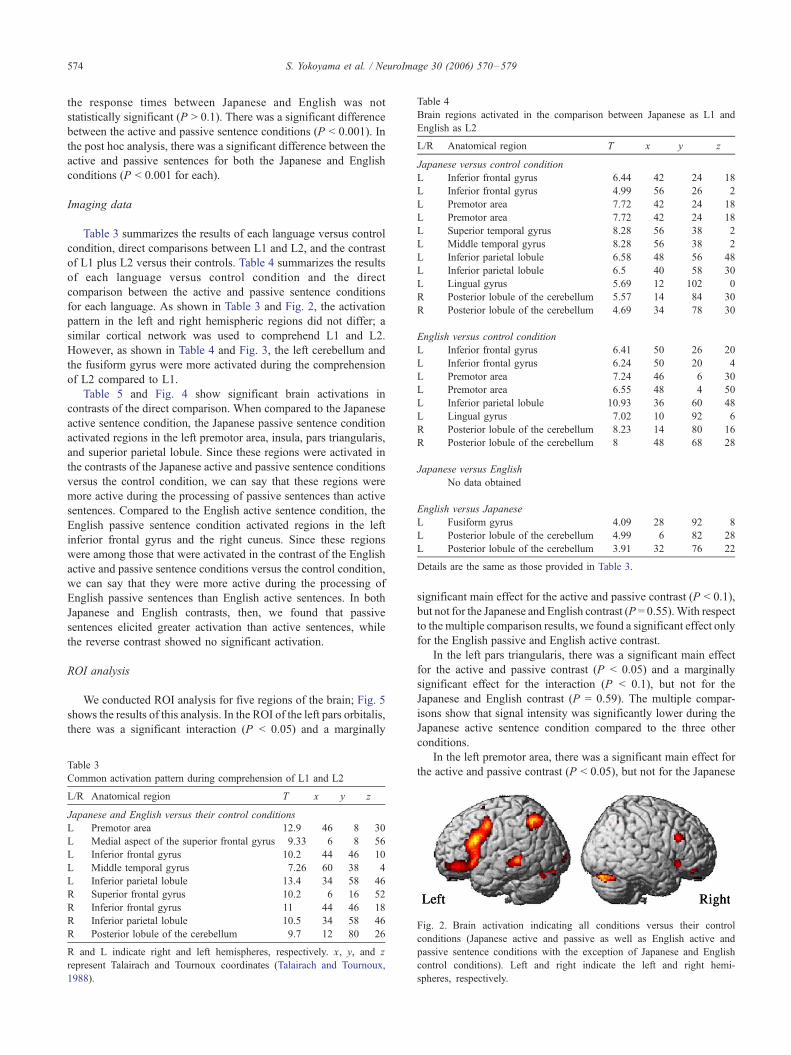
Table 4
Brain regions activated in the comparison between Japanese as L1 and
English as L2
L/R Anatomical region T x y z
Japanese versus control condition
L Inferior frontal gyrus 6.44 �42 24 �18L Inferior frontal gyrus 4.99 �56 26 �2L Premotor area 7.72 �42 24 18
L Premotor area 7.72 �42 24 18
L Superior temporal gyrus 8.28 �56 �38 �2L Middle temporal gyrus 8.28 �56 �38 �2L Inferior parietal lobule 6.58 �48 �56 48
L Inferior parietal lobule 6.5 �40 �58 30
L Lingual gyrus 5.69 �12 �102 0
R Posterior lobule of the cerebellum 5.57 14 �84 �30R Posterior lobule of the cerebellum 4.69 34 �78 �30
English versus control condition
L Inferior frontal gyrus 6.41 �50 26 20
L Inferior frontal gyrus 6.24 �50 20 �4L Premotor area 7.24 �46 6 30
L Premotor area 6.55 �48 4 50
L Inferior parietal lobule 10.93 �36 �60 48
L Lingual gyrus 7.02 �10 �92 �6R Posterior lobule of the cerebellum 8.23 14 �80 �16R Posterior lobule of the cerebellum 8 48 �68 �28
Japanese versus English
No data obtained
English versus Japanese
L Fusiform gyrus 4.09 �28 �92 �8L Posterior lobule of the cerebellum 4.99 �6 �82 �28L Posterior lobule of the cerebellum 3.91 �32 �76 �22Details are the same as those provided in Table 3.
S. Yokoyama et al. / NeuroImage 30 (2006) 570–579574
the response times between Japanese and English was not
statistically significant (P > 0.1). There was a significant difference
between the active and passive sentence conditions (P < 0.001). In
the post hoc analysis, there was a significant difference between the
active and passive sentences for both the Japanese and English
conditions (P < 0.001 for each).
Imaging data
Table 3 summarizes the results of each language versus control
condition, direct comparisons between L1 and L2, and the contrast
of L1 plus L2 versus their controls. Table 4 summarizes the results
of each language versus control condition and the direct
comparison between the active and passive sentence conditions
for each language. As shown in Table 3 and Fig. 2, the activation
pattern in the left and right hemispheric regions did not differ; a
similar cortical network was used to comprehend L1 and L2.
However, as shown in Table 4 and Fig. 3, the left cerebellum and
the fusiform gyrus were more activated during the comprehension
of L2 compared to L1.
Table 5 and Fig. 4 show significant brain activations in
contrasts of the direct comparison. When compared to the Japanese
active sentence condition, the Japanese passive sentence condition
activated regions in the left premotor area, insula, pars triangularis,
and superior parietal lobule. Since these regions were activated in
the contrasts of the Japanese active and passive sentence conditions
versus the control condition, we can say that these regions were
more active during the processing of passive sentences than active
sentences. Compared to the English active sentence condition, the
English passive sentence condition activated regions in the left
inferior frontal gyrus and the right cuneus. Since these regions
were among those that were activated in the contrast of the English
active and passive sentence conditions versus the control condition,
we can say that they were more active during the processing of
English passive sentences than English active sentences. In both
Japanese and English contrasts, then, we found that passive
sentences elicited greater activation than active sentences, while
the reverse contrast showed no significant activation.
ROI analysis
We conducted ROI analysis for five regions of the brain; Fig. 5
shows the results of this analysis. In the ROI of the left pars orbitalis,
there was a significant interaction (P < 0.05) and a marginally
Table 3
Common activation pattern during comprehension of L1 and L2
L/R Anatomical region T x y z
Japanese and English versus their control conditions
L Premotor area 12.9 �46 8 30
L Medial aspect of the superior frontal gyrus 9.33 �6 8 56
L Inferior frontal gyrus 10.2 �44 46 �10L Middle temporal gyrus 7.26 �60 �38 �4L Inferior parietal lobule 13.4 �34 �58 46
R Superior frontal gyrus 10.2 6 16 52
R Inferior frontal gyrus 11 44 46 �18R Inferior parietal lobule 10.5 34 �58 46
R Posterior lobule of the cerebellum 9.7 12 �80 �26R and L indicate right and left hemispheres, respectively. x, y, and z
represent Talairach and Tournoux coordinates (Talairach and Tournoux,
1988).
significant main effect for the active and passive contrast (P < 0.1),
but not for the Japanese and English contrast (P = 0.55).With respect
to the multiple comparison results, we found a significant effect only
for the English passive and English active contrast.
In the left pars triangularis, there was a significant main effect
for the active and passive contrast (P < 0.05) and a marginally
significant effect for the interaction (P < 0.1), but not for the
Japanese and English contrast (P = 0.59). The multiple compar-
isons show that signal intensity was significantly lower during the
Japanese active sentence condition compared to the three other
conditions.
In the left premotor area, there was a significant main effect for
the active and passive contrast (P < 0.05), but not for the Japanese
Fig. 2. Brain activation indicating all conditions versus their control
conditions (Japanese active and passive as well as English active and
passive sentence conditions with the exception of Japanese and English
control conditions). Left and right indicate the left and right hemi-
spheres, respectively.

Fig. 3. Brain activation indicating the difference between Japanese and
English (the English active and passive sentence conditions with the
exception of Japanese active and passive sentence conditions). The figure
on the left side indicates the left hemisphere. In the figure on the right side,
the left side indicates the left hemisphere.
Table 5
Brain regions activated in the comparison among Japanese active, Japanese
passive, English active, and English passive sentence conditions
L/R Anatomical region T x y z
Japanese active versus control condition
L Inferior frontal gyrus 6.44 �42 24 �18L Inferior frontal gyrus 5.85 �44 22 �32L Inferior frontal gyrus 4.99 �56 26 �2L Premotor area 4.45 �40 22 22
L Premotor area 3.74 �48 4 50
L Middle temporal gyrus 5.75 �54 �38 �2L Superior temporal sulcus 3.94 �58 �50 12
L Inferior parietal lobule 5.85 �38 �56 28
L Intraparietal sulcus 4.4 �40 �64 38
R Posterior lobule of the cerebellum 5.57 14 �84 �30R Posterior lobule of the cerebellum 4.69 34 �78 30
Japanese passive versus control condition
L Inferior frontal gyrus 6.87 �44 28 �18L Middle frontal gyrus 5.85 �42 22 28
L Middle frontal gyrus 5.88 �54 30 10
L Premotor area 4.56 �44 8 42
L Premotor area 3.98 �38 12 52
L Premotor area 3.89 �40 18 42
L Superior frontal gyrus 3.86 �6 6 68
L Medial aspect of the superior frontal gyrus 5.55 �2 46 36
R Medial aspect of the superior frontal gyrus 3.99 4 32 48
L Middle temporal gyrus 5.8 �62 �42 0
L Middle temporal gyrus 4.06 �56 �28 �10L Insula 4.37 �32 28 �12L Fusiform gyrus 3.99 �40 �56 �12L Inferior parietal lobule 7.57 �42 �58 30
L Superior parietal lobule 6.39 �38 �70 48
L Superior parietal lobule 4.7 �48 �60 48
R Posterior lobule of the cerebellum 9.39 14 �82 �32R Posterior lobule of the cerebellum 4.58 34 �82 �26
S. Yokoyama et al. / NeuroImage 30 (2006) 570–579 575
and English contrast (P = 0.15) and the interaction (P = 0.33). The
multiple comparisons show a significant difference only for the
Japanese passive and Japanese active contrast.
In the left insula, there was a significant main effect for the
active and passive contrast (P < 0.01), but not for the Japanese and
English contrast (P = 0.92) and the interaction (P = 0.47). The
multiple comparisons show a significant difference only for the
Japanese passive and Japanese active contrast.
In the left superior parietal lobule, there was a significant main
effect for both the active and passive contrast (P < 0.05) and the
Japanese and English contrast (P < 0.05), but not for the interaction
(P = 0.18). The multiple comparisons show that signal intensity is
significantly lower during the Japanese active sentence condition
compared to the three other conditions.
In the analysis of the correlation between the behavioral data
and the % signal change for each ROI, there was no significant
correlation for any of the ROIs in terms of accuracy rates or
response time. Table 6 summarizes the results of the correlation
analysis.
English active versus control condition
L Inferior frontal gyrus 4.45 �50 22 �2L Premotor area 4.21 �50 8 46
L Medial aspect of the superior frontal gyrus 3.89 �8 26 44
L Precentral gyrus 4.12 �36 0 40
L Middle temporal gyrus 3.86 �52 �40 �2R Posterior lobule of the cerebellum 4.02 32 �74 �32R Posterior lobule of the cerebellum 3.87 8 �82 �26L Posterior lobule of the cerebellum 3.68 �4 �82 �24
English passive versus control condition
L Inferior frontal gyrus 4.63 �44 24 �6L Inferior frontal gyrus 4.41 �46 22 �18L Premotor area 3.61 �48 6 38
L Superior frontal gyrus 4.4 �4 6 68
L Cuneus 3.91 �8 �88 �2R Posterior lobule of the cerebellum 5.5 12 �84 �22
Japanese passive versus active
L Superior parietal lobule 3.95 �28 �74 46
L Inferior frontal gyrus: pars triangularis 3.9 �52 34 14
L Premotor area 3.79 �52 20 30
L Insula 3.62 �32 18 �4
English passive versus active
L Inferior frontal gyrus: pars orbitalis 3.91 �50 20 �10R Cuneus 4.48 �6 �88 0
Details are the same as those provided in Table 3.
Discussion
In this study, we directly compared late bilinguals’ processing
of Japanese (L1) and English (L2). We found that the language-
related regions were commonly activated during the processing of
both L1 and L2; and that when the effect of task difficulty was
statistically excluded, there was no difference in the activation
pattern of the hemispheric regions related to language function.
Thus, our fMRI experiment indicates that late bilinguals use a
similar neural basis to comprehend both L1 and L2. In contrast, we
compared the processing of Japanese direct passive sentences with
English passive sentences and found different activation patterns
between L1 and L2. These results support our hypothesis that late
bilinguals use language-related regions of the brain differently
when processing structurally complex sentences in L1 versus L2.
We discuss these results in more detail below.
Difference between L1 and L2
In contrast to previous neuroimaging studies, we found no
difference between L1 and L2 in the activation pattern of the
hemispheric regions related to language function, such as the left
inferior frontal, superior/middle temporal, and the parietal regions
(see Table 4 and Fig. 3). As described in the Introduction, previous
studies have reported greater activation in these areas for L2
compared to L1. Our study suggests that such results may be
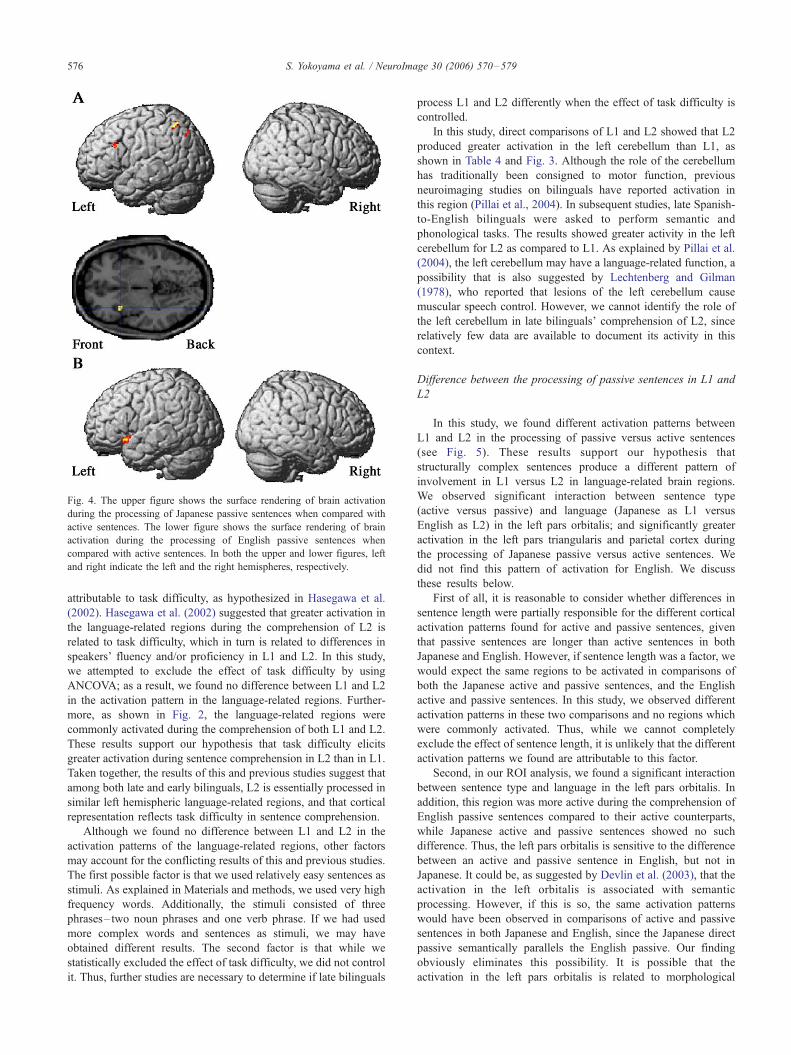
Fig. 4. The upper figure shows the surface rendering of brain activation
during the processing of Japanese passive sentences when compared with
active sentences. The lower figure shows the surface rendering of brain
activation during the processing of English passive sentences when
compared with active sentences. In both the upper and lower figures, left
and right indicate the left and the right hemispheres, respectively.
S. Yokoyama et al. / NeuroImage 30 (2006) 570–579576
attributable to task difficulty, as hypothesized in Hasegawa et al.
(2002). Hasegawa et al. (2002) suggested that greater activation in
the language-related regions during the comprehension of L2 is
related to task difficulty, which in turn is related to differences in
speakers’ fluency and/or proficiency in L1 and L2. In this study,
we attempted to exclude the effect of task difficulty by using
ANCOVA; as a result, we found no difference between L1 and L2
in the activation pattern in the language-related regions. Further-
more, as shown in Fig. 2, the language-related regions were
commonly activated during the comprehension of both L1 and L2.
These results support our hypothesis that task difficulty elicits
greater activation during sentence comprehension in L2 than in L1.
Taken together, the results of this and previous studies suggest that
among both late and early bilinguals, L2 is essentially processed in
similar left hemispheric language-related regions, and that cortical
representation reflects task difficulty in sentence comprehension.
Although we found no difference between L1 and L2 in the
activation patterns of the language-related regions, other factors
may account for the conflicting results of this and previous studies.
The first possible factor is that we used relatively easy sentences as
stimuli. As explained in Materials and methods, we used very high
frequency words. Additionally, the stimuli consisted of three
phrases– two noun phrases and one verb phrase. If we had used
more complex words and sentences as stimuli, we may have
obtained different results. The second factor is that while we
statistically excluded the effect of task difficulty, we did not control
it. Thus, further studies are necessary to determine if late bilinguals
process L1 and L2 differently when the effect of task difficulty is
controlled.
In this study, direct comparisons of L1 and L2 showed that L2
produced greater activation in the left cerebellum than L1, as
shown in Table 4 and Fig. 3. Although the role of the cerebellum
has traditionally been consigned to motor function, previous
neuroimaging studies on bilinguals have reported activation in
this region (Pillai et al., 2004). In subsequent studies, late Spanish-
to-English bilinguals were asked to perform semantic and
phonological tasks. The results showed greater activity in the left
cerebellum for L2 as compared to L1. As explained by Pillai et al.
(2004), the left cerebellum may have a language-related function, a
possibility that is also suggested by Lechtenberg and Gilman
(1978), who reported that lesions of the left cerebellum cause
muscular speech control. However, we cannot identify the role of
the left cerebellum in late bilinguals’ comprehension of L2, since
relatively few data are available to document its activity in this
context.
Difference between the processing of passive sentences in L1 and
L2
In this study, we found different activation patterns between
L1 and L2 in the processing of passive versus active sentences
(see Fig. 5). These results support our hypothesis that
structurally complex sentences produce a different pattern of
involvement in L1 versus L2 in language-related brain regions.
We observed significant interaction between sentence type
(active versus passive) and language (Japanese as L1 versus
English as L2) in the left pars orbitalis; and significantly greater
activation in the left pars triangularis and parietal cortex during
the processing of Japanese passive versus active sentences. We
did not find this pattern of activation for English. We discuss
these results below.
First of all, it is reasonable to consider whether differences in
sentence length were partially responsible for the different cortical
activation patterns found for active and passive sentences, given
that passive sentences are longer than active sentences in both
Japanese and English. However, if sentence length was a factor, we
would expect the same regions to be activated in comparisons of
both the Japanese active and passive sentences, and the English
active and passive sentences. In this study, we observed different
activation patterns in these two comparisons and no regions which
were commonly activated. Thus, while we cannot completely
exclude the effect of sentence length, it is unlikely that the different
activation patterns we found are attributable to this factor.
Second, in our ROI analysis, we found a significant interaction
between sentence type and language in the left pars orbitalis. In
addition, this region was more active during the comprehension of
English passive sentences compared to their active counterparts,
while Japanese active and passive sentences showed no such
difference. Thus, the left pars orbitalis is sensitive to the difference
between an active and passive sentence in English, but not in
Japanese. It could be, as suggested by Devlin et al. (2003), that the
activation in the left orbitalis is associated with semantic
processing. However, if this is so, the same activation patterns
would have been observed in comparisons of active and passive
sentences in both Japanese and English, since the Japanese direct
passive semantically parallels the English passive. Our finding
obviously eliminates this possibility. It is possible that the
activation in the left pars orbitalis is related to morphological
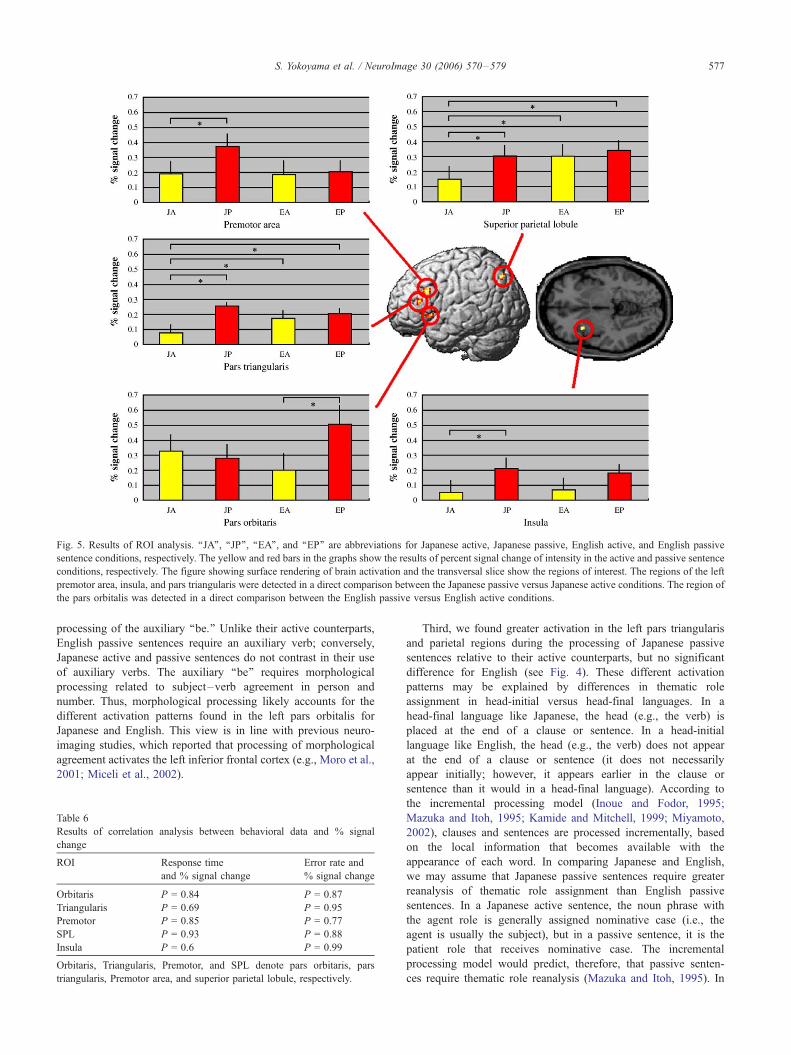
Fig. 5. Results of ROI analysis. ‘‘JA’’, ‘‘JP’’, ‘‘EA’’, and ‘‘EP’’ are abbreviations for Japanese active, Japanese passive, English active, and English passive
sentence conditions, respectively. The yellow and red bars in the graphs show the results of percent signal change of intensity in the active and passive sentence
conditions, respectively. The figure showing surface rendering of brain activation and the transversal slice show the regions of interest. The regions of the left
premotor area, insula, and pars triangularis were detected in a direct comparison between the Japanese passive versus Japanese active conditions. The region of
the pars orbitalis was detected in a direct comparison between the English passive versus English active conditions.
S. Yokoyama et al. / NeuroImage 30 (2006) 570–579 577
processing of the auxiliary ‘‘be.’’ Unlike their active counterparts,
English passive sentences require an auxiliary verb; conversely,
Japanese active and passive sentences do not contrast in their use
of auxiliary verbs. The auxiliary ‘‘be’’ requires morphological
processing related to subject–verb agreement in person and
number. Thus, morphological processing likely accounts for the
different activation patterns found in the left pars orbitalis for
Japanese and English. This view is in line with previous neuro-
imaging studies, which reported that processing of morphological
agreement activates the left inferior frontal cortex (e.g., Moro et al.,
2001; Miceli et al., 2002).
Table 6
Results of correlation analysis between behavioral data and % signal
change
ROI Response time
and % signal change
Error rate and
% signal change
Orbitaris P = 0.84 P = 0.87
Triangularis P = 0.69 P = 0.95
Premotor P = 0.85 P = 0.77
SPL P = 0.93 P = 0.88
Insula P = 0.6 P = 0.99
Orbitaris, Triangularis, Premotor, and SPL denote pars orbitaris, pars
triangularis, Premotor area, and superior parietal lobule, respectively.
Third, we found greater activation in the left pars triangularis
and parietal regions during the processing of Japanese passive
sentences relative to their active counterparts, but no significant
difference for English (see Fig. 4). These different activation
patterns may be explained by differences in thematic role
assignment in head-initial versus head-final languages. In a
head-final language like Japanese, the head (e.g., the verb) is
placed at the end of a clause or sentence. In a head-initial
language like English, the head (e.g., the verb) does not appear
at the end of a clause or sentence (it does not necessarily
appear initially; however, it appears earlier in the clause or
sentence than it would in a head-final language). According to
the incremental processing model (Inoue and Fodor, 1995;
Mazuka and Itoh, 1995; Kamide and Mitchell, 1999; Miyamoto,
2002), clauses and sentences are processed incrementally, based
on the local information that becomes available with the
appearance of each word. In comparing Japanese and English,
we may assume that Japanese passive sentences require greater
reanalysis of thematic role assignment than English passive
sentences. In a Japanese active sentence, the noun phrase with
the agent role is generally assigned nominative case (i.e., the
agent is usually the subject), but in a passive sentence, it is the
patient role that receives nominative case. The incremental
processing model would predict, therefore, that passive senten-
ces require thematic role reanalysis (Mazuka and Itoh, 1995). In

S. Yokoyama et al. / NeuroImage 30 (2006) 570–579578
addition, Japanese passive sentences require thematic role
reanalysis for all the components that appear before the head,
since the head verb appears at the end of the clause or sentence.
In contrast, in English passive sentences, only the subject
requires thematic role reanalysis, since the verb is preceded by
only one noun phrase. This difference between Japanese and
English is reflected in our data. We found that the left
triangularis and the parietal region were more active during
the comprehension of Japanese passive sentences compared to
their active counterparts, while the comparison of English
passive and active sentences showed no difference in activation
in these regions. If we assume that the left pars triangularis and
parietal cortex are both involved in thematic role assignment,
then our results are compatible with the incremental processing
model’s predictions with respect to thematic processing.
Accordingly, our results are consistent with the findings that
the left pars triangularis (Newman et al., 2003) and the parietal
region (Inui et al., 1998) are involved in thematic processing.
Fourth, the different activation patterns observed in our study
may be attributable not only to the differences in grammatical
construction discussed above, but also to participants’ L2
proficiency. The participants in our study were late bilinguals,
with greater proficiency in L1 than L2. They may have found it
difficult to learn L2 grammatical properties that were not present in
their L1. Thus, the different activation patterns we found may stem
from the interaction between differences in grammatical construc-
tion between L1 and L2 and the participants’ L2 proficiency.
Fifth, it must be noted that between-language interference may
occur in a study design like ours, where sentences in the same
language continue in a block. Although we counter-balanced the
order of blocks across subjects, we cannot completely exclude the
effects of between-language interference. To address this issue, in
future studies of bilingual sentence comprehension, an event-
related or ‘‘odd ball’’ paradigm may be preferable.
Conclusion
In this study, we used fMRI to investigate whether the
comprehension of structurally complex sentences generates differ-
ent activation patterns in L1 and L2. First, our results indicate that
L1 and L2 are processed in similar language-related regions, such
as the left inferior frontal, superior/middle temporal, and parietal
regions, and that there is no difference in the activation pattern in
these regions between L1 and L2 when the effect of task difficulty
is statistically excluded. These results suggest that late bilinguals
use a similar neural basis to comprehend both L1 and L2.
However, we also found different activation patterns for the
processing of passive sentences in L1 versus L2. Passive sentences
elicited greater activation than their active counterparts in the left
pars triangularis, the premotor area, and the superior parietal lobule
in Japanese, but not in English. Furthermore, we found a
significant interaction between sentence type (active versus
passive) and language (Japanese versus English) in the left pars
orbitalis, indicating that structurally complex sentences generate a
different pattern of involvement in these regions for L1 and L2. We
attribute this contrast in cortical representation to an interaction
between differences in grammatical construction between L1 and
L2, and L2 proficiency. Therefore, further neuroimaging studies on
late bilinguals’ comprehension of L1 and L2 should take into
account not only age of acquisition and L2 proficiency, but also
task difficulty and differences in grammatical construction between
L1 and L2.
Acknowledgments
We thank Professor Noriaki Yusa of Miyagi Gakuin
Women’s University and Associate Professor Masatoshi Koi-
zumi of Tohoku University for their helpful comments. We also
thank two anonymous reviewers for their helpful comments.
This study was supported by JST/RISTEX, R&D Promotion
Scheme for Regional Proposals Promoted by TAO, and the 21st
Century Center of Excellence (COE) Program (Ministry of
Education, Culture, Sports, Science and Technology) entitled ‘‘A
Strategic Research and Education Center for an Integrated
Approach to Language and Cognition’’ (Tohoku University).
References
Albert, M.L., Obler, L.K., 1978. The Bilingual Brain. Academic Press, New
York.
Amano, S., Kondo, T., 1999. Nihongo no Goi-Tokusei, Dai 2 kan, Hindo.
Lexical Properties of Japanese, vol. 2. NTT Communication Science
Laboratories, Kyoto. Frequency.
Carpenter, P.A., Just, M.A., 1975. Sentence comprehension: a psycho-
linguistic processing model of verification. Psychol. Rev. 82, 45–73.
Carpenter, P.A., Just, M.A., Keller, T.A., Eddy, W.F., Thulborn, K.R., 1999.
Time course of fMRI-activation in language and spatial networks during
sentence comprehension. Nueroimage 10, 216–224.
Chee, M.W., Caplan, D., Soon, C.S., Sriram, N., Tan, E.W., Thiel, T.,
Weekes, B., 1999. Processing of visually presented sentences in
Mandarin and English studied with fMRI. Neuron 23, 127–137.
Dehaene, S., Dupoux, E., Mehler, J., Cohen, L., Paulesu, E., Perani, D., van
de Moortele, P.F., Lehericy, S., Le Bihan, D., 1997. Anatomical
variability in the cortical representation of first and second language.
NeuroReport 8, 3809–3815.
Devlin, J.T., Matthews, P.M., Rushworth, M.F., 2003. Semantic processing
in the left inferior prefrontal cortex: a combined functional magnetic
resonance imaging and transcranial magnetic stimulation study. J. Cogn.
Neurosci. 15, 71–84.
Fabbro, F., Paradis, M., 1995. Differential impairments in four multilingual
patients with subcortical lesions. In: Paradis, M. (Ed.), Aspects of
Bilingual Aphasia. Pergamon Press, London, pp. 139–176.
Friston, K., Holmes, A.P., Worsley, K., Poline, J.B., Frith, C., Frackowiak,
R.S., 1995. Statistical parametric maps in functional imaging: a general
linear approach. Human Brain Mapping 2, 189–210.
Hasegawa, M., Carpenter, P., Just, M., 2002. An fMRI study of bilingual
sentence comprehension and workload. NeuroImage 15, 647–660.
Illes, J., Francis, W., Desmond, J., Gabrieli, J., Glover, G., Poldrack, R.,
Lee, C., Wagner, D., 1999. Convergent cortical representation of
semantic processing in bilinguals. Brain Lang. 70, 347–363.
Inoue, A., Fodor, J., 1995. Information-paced parsing of Japanese. In:
Mazuka, R., Nagai, N. (Eds.), Japanese Sentence Processing. Lawrence
Erlbaum Associates, Mahwah, pp. 9–63.
Inui, T., Otsu, Y., Tanaka, S., Okada, T., Nishizawa, S., Konishi, J., 1998. A
functional MRI analysis of comprehension processes of Japanese
sentences. NeuroReport 9, 3325–3328.
Junque, C., Vendrell, P., Vendrell, J., 1995. Differential impairments and
specific phenomena in 50 Catalan–Spanish bilingual aphasic patients.
In: Paradis, M. (Ed.), Aspects of Bilingual Aphasia. Pergamon Press,
London, pp. 139–176.
Kamide, Y., Mitchell, D., 1999. Incremental pre-head attachment in
Japanese parsing. Lang. Cogn. Processes 14, 631–662.
Klein, D., Zatorre, R.J., Milner, B., Meyer, E., 1994. Left putaminal

S. Yokoyama et al. / NeuroImage 30 (2006) 570–579 579
activation when speaking a second language: evidence from PET.
NeuroReport 5, 2295–2297.
Klein, D., Milner, B., Zatorre, R.J., Meyer, E., Evans, A.C., 1995. The
neural substrates underlying word generation: a bilingual functional-
imaging study. Proc. Natl. Acad. Sci. U. S. A. 92, 2899–2903.
Lechtenberg, R., Gilman, S., 1978. Speech disorders in cerebellar disease.
Ann. Neurol. 3, 285–290.
Luke, K., Liu, H., Wai, Y., Wan, Y., Tan, L.H., 2002. Functional anatomy of
syntactic and semantic processing in language comprehension. Hum.
Brain Mapp. 16, 133–145.
Mazuka, R., Itoh, K., 1995. Can Japanese speakers be led down the garden-
path? In: Mazuka, R., Nagai, N. (Eds.), Japanese Sentence Processing.
Lawrence Erlbaum Associates, Mahwah, pp. 295–329.
Miceli, G., Turriziani, P., Caltagirone, C., Capasso, R., Tomaiuolo, F.,
Caramazza, A., 2002. The neural correlates of grammatical gender: an
fMRI investigation. J. Cogn. Neurosci. 14, 618–628.
Miura, N., Watanabe, J., Iwata, K., Sassa, Y., Riera, J., Tsuchiya, H.,
Sato, S., Horie, K., Takahashi, M., Kitamura, M., Kawashima, R.,
2005. Cortical activation during reading of ancient versus modern
Japanese texts: fMRI study. NeuroImage 26, 426–431.
Miyamoto, E., 2002. Case markers as clause boundary inducers in
Japanese. J. Psycholinguist. Res. 31, 307–347.
Moro, A., Tettamanti, M., Perani, D., Donati, D., Cappa, C., Fazio, F., 2001.
Syntax and the brain: disentangling grammar by selective anomalies.
NeuroImage 13, 110–118.
Newman, S.D., Just, M.A., Keller, T.A., Roth, J., Carpenter, P.A., 2003.
Differential effects of syntactic and semantic processing on the
subregions of Broca’s area. Cogn. Brain Res. 16, 297–307.
O’Grady, W., Dobrovolsky, M., 1996. Contemporary Linguistic Analysis.
Copp Clark, Toronto.
Ojemann, G.A., Whitaker, H.A., 1978. The Bilingual Brain. Arch. Neurol.
35, 409–412.
Oldfield, R., 1971. The assessment and analysis of handedness: the
Edinburgh inventory. Neuropsychologia 9, 812–815.
Perani, D., Dehaene, S., Grassi, F., Cohen, L., Cappa, S.F., Dupoux, E.,
Fazio, F., Mehler, J., 1996. Brain processing of native and foreign
languages. NeuroReport 7, 2439–2444.
Pillai, J., Allison, J., Sethuraman, S., Araque, J., Thiruvaiyaru, D., Ison, C.,
Loring, D., Lavin, T., 2004. Functional MR imaging study of language-
related differences in bilingual cerebellar activation. Am. J. Neuro-
radiol. 25, 523–532.
Shibatani, M., 1985. Passives and related constructions: a prototype
analysis. Language 61, 821–848.
Shibatani, M., 1990. The Languages of Japan. Cambridge Univ. Press,
Cambridge.
Talairach, J., Tournoux, P., 1988. Co-Planar Stereotaxic Atlas of the Human
Brain. Thieme, Stuttgart.
Wartenburger, I., Heekeren, H., Abutalebi, J., Cappa, S., Villringer, A.,
Perani, D., 2003. Early setting of grammatical processing in the
bilingual brain. Neuron 37, 159–170.





























