Embed Size (px)
Citation preview

Molecular Plant • Volume 6 • Number 6 • Pages 1945–1960 • November 2013 RESEARCH ARTICLE
Control of Rice Embryo Development, Shoot Apical Meristem Maintenance, and Grain Yield by a Novel Cytochrome P450Weibing Yanga, Mingjun Gaoa, Xin Yina, Jiyun Liub, Yonghan Xuc, Longjun Zenga, Qun Lia, Shubiao Zhangd, Junmin Wangb, Xiaoming Zhangb, and Zuhua Hea,1
a National Key Laboratory of Plant Molecular Genetics and National Center for Plant Gene Research (Shanghai), Shanghai Institute of Plant Physiology and Ecology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, 300 Fenglin Road, Shanghai 200032, Chinab State Key Laboratory Breeding Base for Zhejiang Sustainable Pest and Disease Control, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, Chinac Zhejiang Province Key Laboratory of Technology for Improving Agricultural Product Quality, School of Agricultural and Food Science, Zhejiang Agriculture and Forest University, Hangzhou 311300, Chinad Institute of Genetics and Crop Breeding, Fujian Agriculture and Forestry University, Fuzhou 350002, China
ABSTRACT Angiosperm seeds usually consist of two major parts: the embryo and the endosperm. However, the molec-ular mechanism(s) underlying embryo and endosperm development remains largely unknown, particularly in rice, the model cereal. Here, we report the identification and functional characterization of the rice GIANT EMBRYO (GE) gene. Mutation of GE resulted in a large embryo in the seed, which was caused by excessive expansion of scutellum cells. Post-embryonic growth of ge seedling was severely inhibited due to defective shoot apical meristem (SAM) mainte-nance. Map-based cloning revealed that GE encodes a CYP78A subfamily P450 monooxygenase that is localized to the endoplasmic reticulum. GE is expressed predominantly in the scutellar epithelium, the interface region between embryo and endosperm. Overexpression of GE promoted cell proliferation and enhanced rice plant growth and grain yield, but reduced embryo size, suggesting that GE is critical for coordinating rice embryo and endosperm development. Moreover, transgenic Arabidopsis plants overexpressing AtCYP78A10, a GE homolog, also produced bigger seeds, implying a con-served role for the CYP78A subfamily of P450s in regulating seed development. Taken together, our results indicate that GE plays critical roles in regulating embryo development and SAM maintenance.
Key words: embryo; endosperm; shoot apical meristem; cytochrome P450; grain yield; rice.
InTROduCTIOnPlant seeds are the most important food resources for human beings (Tanksley and McCouch, 1997). A typical plant seed is mainly composed of two parts: the embryo and the endosperm. Seed development is initiated by double fertilization, during which one sperm fertilizes the egg cell to form a diploid zygote. The other sperm fuses with the two polar nuclei, giving rise to a triploid endosperm (Dresselhaus, 2006). In Arabidopsis thaliana, the zygote divides asymmetrically into a small, cytoplasmic apical cell and a large vacuolated basal cell. The apical cell divides and differentiates into shoot meristem, cotyledon, hypocotyl, and embryonic root primordial. The basal cell differentiates into a filamentous suspensor. The endosperm is a transient tissue that is finally consumed by the embryo (Jurgens et al., 2001; Lau et al., 2012). Compared with Arabidopsis, rice seed growth differs in several aspects. In rice, the division directions of early embryo cells are unfixed
and positional information is critical for embryo patterning. Grass-specific organs including coleoptile, scutellum, and epiblast are produced during rice embryo development (Itoh et al., 2005). Additionally, rice endosperm persists in the mature seed and accumulates most of the nutrients (Zhou et al., 2013).
Molecular genetic studies have revealed a complex frame-work involved in Arabidopsis embryogenesis, which depends on the distinct expression domains and coordinative functions of the MAPKK kinase gene YODA (YDA) (Lukowitz et al.,
1To whom correspondence should be addressed. E-mail [email protected], tel. 86-21-54924121, fax 86-21-54924123.
© The Author 2013. Published by the Molecular Plant Shanghai Editorial Office in association with Oxford University Press on behalf of CSPB and IPPE, SIBS, CAS.
doi:10.1093/mp/sst107, Advance Access publication 17 June 2013
Received 24 May 2013; accepted 9 June 2013

1946 Yang et al. • GE Controls Rice Embryo Development and Grain Yield
2004), WRKY DNA-BINDING PROTEIN 2 (WRKY2) transcript factor as well as WUSCHEL-RELATED HOMEOBOX (WOX) genes (Breuninger et al., 2008; Ueda et al., 2011). The auxin signaling pathway comprising directional auxin transport and local auxin response is also indispensable for proper embryo development (Hamann et al., 2002; Benkova et al., 2003; Friml et al., 2003; Geldner et al., 2003). Although a number of rice embryo mutants that exhibited defects in embryonic organ formation and embryo size have been isolated (Hong et al., 1995), none of the corresponding genes has been iden-tified so far, leaving the molecular mechanisms governing rice embryo growth and pattern formation poorly understood.
Plant seed size is determined by the coordinative growth of embryo, endosperm, and maternal tissue. In Arabidopsis, several positive regulators of seed size have been char-acterized, including HAIKU1, HAIKU2, and MINISEED3, which promote endosperm growth (Luo et al., 2005), and TRANSPARENT TESTA GLABRA2 (TTG2), which activates integument cell expansion (Garcia et al., 2005). On the other hand, APETALA2 (AP2), AUXIN RESPONSE FACTOR2 (ARF2), and DA1 were shown to be negative regulators that restrict seed growth (Jofuku et al., 2005; Schruff et al., 2006; Li et al., 2008). Recent studies also revealed the involvement of a novel CYP78A class of cytochrome P450 monooxygenases in con-trolling Arabidopsis seed development. Mutations in either KLUH/CYP78A5, EOD3/CYP78A6, or CYP78A9 all resulted in small seeds (Anastasiou et al., 2007; Fang et al., 2012). In rice, the seed size is an important agronomic trait controlled by multiple quantitative trait loci (QTLs), including GW2, GL3.1, GS3, GS5, GW5, and GW8, which regulate spikelet hull expan-sion, and GIF1, which regulates grain filling (Fan et al., 2006; Song et al., 2007; Wang et al., 2008a; Weng et al., 2008; Li et al., 2011; Qi et al., 2012; Wang et al., 2012).
The shoot apical meristem (SAM) is formed during early embryogenesis and gives rise to the entire aboveground plant body (Aichinger et al., 2012). Establishment and maintenance of SAM depends on an auto-feedback signaling loop compris-ing the WUSCHEL (WUS) and CLAVATA (CLV) in Arabidopsis (Brand et al., 2000; Schoof et al., 2000). Rice FON2-LIKE CLE PROTEIN1 (FCP1) and WUSCHEL-RELATED HOMEOBOX4 (WOX4) also constitute a feedback loop to promote shoot apical meristem maintenance (Ohmori et al., 2013). In addi-tion, small RNA-mediated gene regulation also affects rice SAM development. Mutation in SHOOTLESS2 (SHL2), SHOOT ORGANIZATION2 (SHO2, also known as SHL4), and SHO1, which encode homologs of Arabidopsis RNA-dependent RNA polymerase 6, AGO7, and DICER-like 4, respectively, caused complete loss or severe inhibition of SAM formation (Nagasaki et al., 2007).
Rice has been adopted as the model for cereal seed devel-opment. The giant embryo (ge) mutant has long been rec-ognized and used in rice breeding for its high nutritional contents (Zhang et al., 2007). However, the molecular mecha-nism underlying ge embryo development keeps unclear. Here, we report the identification and molecular characterization
of GIANT EMBRYO (GE) gene. GE encodes a cytochrome P450 belonging to the CYP78A subfamily. GE expressed predomi-nantly in the interface region between the embryo and the endosperm. GE controls embryo development and SAM main-tenance in a non-cell-autonomous manner. Increased expres-sion of GE could significantly promote rice growth and grain yield but restricted embryo development, suggesting that GE controls the coordination of rice embryo and endosperm growth.
RESuLTSCharacterization of Rice Embryo development Mutants
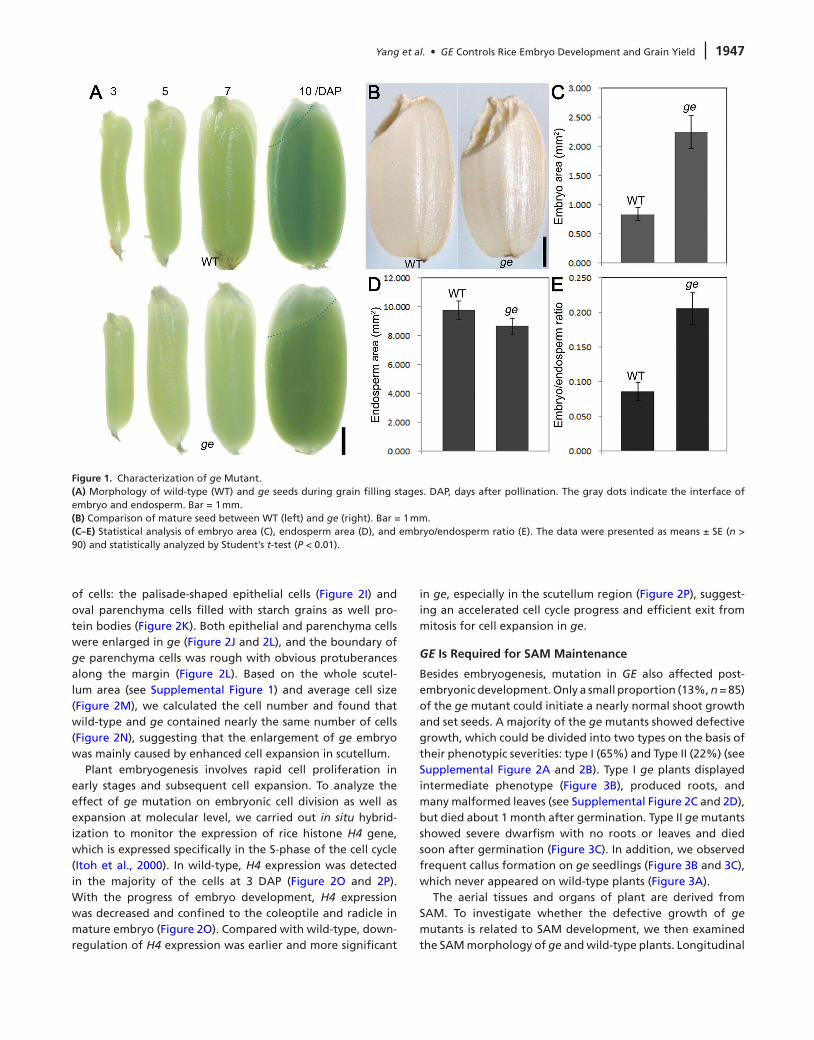
To understand the cellular and molecular mechanisms under-lying rice seed and embryo development, we looked for mutants with visible growth defects in seed development but displaying no obvious agronomic phenotypes from large populations of rice mutants generated by γ-ray irradia-tion and ethyl methanesulphonate (EMS) treatment (Wang et al., 2008a; Yang et al., 2011). One of the mutants showing large embryo, designated as ge, was identified and used in this study. The overall seed morphology of wild-type and ge mutant was comparable during earlier developmental stages. However, at 10 d after pollination (DAP), when the embryo development is nearly completed, a large embryo was devel-oped in ge seed (Figure 1A). There was no significant dif-ference in seed size between wild-type (10.62 mm2) and ge (10.93 mm2) (Figure 1B). The mature embryo of ge was two to three-fold bigger than that of wild-type (Figure 1C), whereas the ge endosperm size was reduced (Figure 1D). As a result, compared with the wild-type, the embryo/endosperm ratio was higher in ge (Figure 1E).
Morphogenesis of ge Embryo
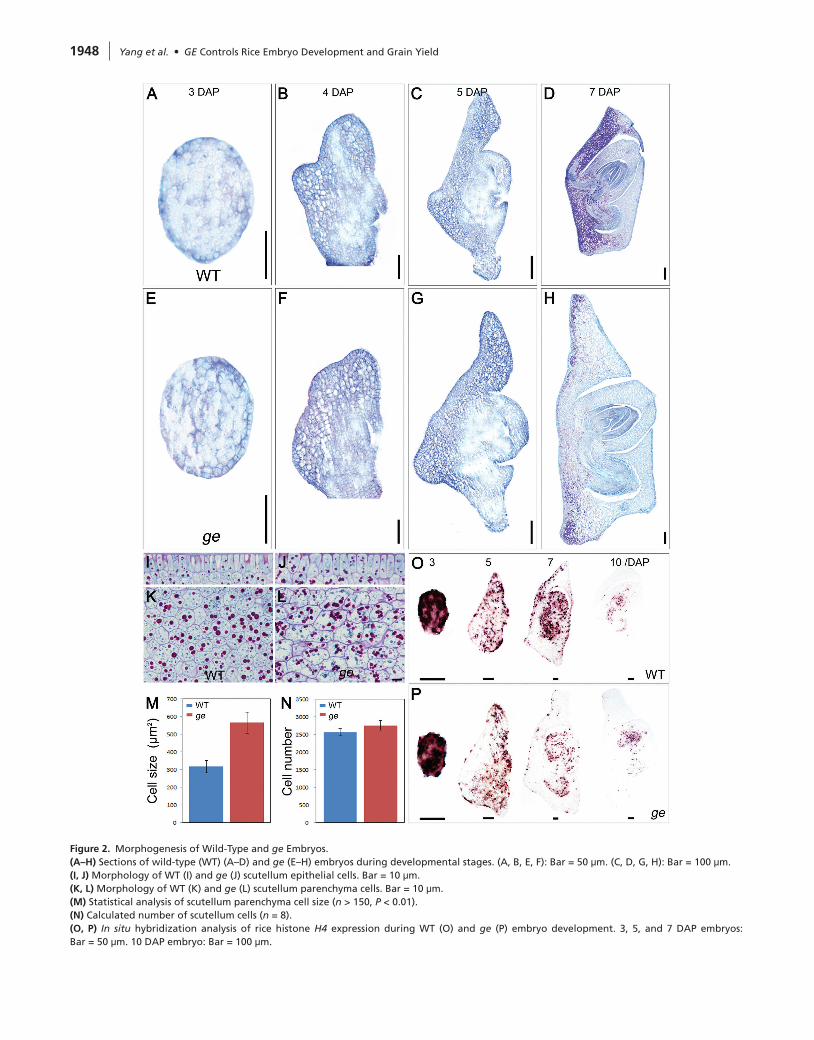
To dissect how ge mutation affects embryo development, we analyzed the whole embryogenesis processes of the wild-type and ge. At 3 DAP, the zygote divides quickly to form a globular embryo, in which only the epidermal cells differentiate (Figure 2A). The globular embryo of ge was indistinguishable from the wild-type, suggesting that mutation of GE does not interfere with early embryo patterning (Figure 2E). The coleoptile primordium and scutellum began to differentiate at 4 DAP (Figure 2B). Compared with wild-type, the scutellum of ge was larger at this stage (Figure 2F). Enlargement of ge scutellum was more evident from 5 to 7 DAP when all embryonic organs eventually formed and embryogenesis completed (Figure 2C, 2D, 2G, and 2H).
Rice embryo is composed of coleoptile, radicle, scutellum, epiblast, and SAM (Itoh et al., 2005). Each part of the ge embryo was relatively bigger than the wild-type but scutel-lum enlargement played the dominant role in promoting ge embryo growth (see Supplemental Figure 1). We next ana-lyzed the cell morphology to understand the cellular basis for scutellum enlargement. The scutellum contains two types
Yang et al. • GE Controls Rice Embryo Development and Grain Yield 1947
of cells: the palisade-shaped epithelial cells (Figure 2I) and oval parenchyma cells filled with starch grains as well pro-tein bodies (Figure 2K). Both epithelial and parenchyma cells were enlarged in ge (Figure 2J and 2L), and the boundary of ge parenchyma cells was rough with obvious protuberances along the margin (Figure 2L). Based on the whole scutel-lum area (see Supplemental Figure 1) and average cell size (Figure 2M), we calculated the cell number and found that wild-type and ge contained nearly the same number of cells (Figure 2N), suggesting that the enlargement of ge embryo was mainly caused by enhanced cell expansion in scutellum.
Plant embryogenesis involves rapid cell proliferation in early stages and subsequent cell expansion. To analyze the effect of ge mutation on embryonic cell division as well as expansion at molecular level, we carried out in situ hybrid-ization to monitor the expression of rice histone H4 gene, which is expressed specifically in the S-phase of the cell cycle (Itoh et al., 2000). In wild-type, H4 expression was detected in the majority of the cells at 3 DAP (Figure 2O and 2P). With the progress of embryo development, H4 expression was decreased and confined to the coleoptile and radicle in mature embryo (Figure 2O). Compared with wild-type, down-regulation of H4 expression was earlier and more significant
in ge, especially in the scutellum region (Figure 2P), suggest-ing an accelerated cell cycle progress and efficient exit from mitosis for cell expansion in ge.
GE Is Required for SAM Maintenance
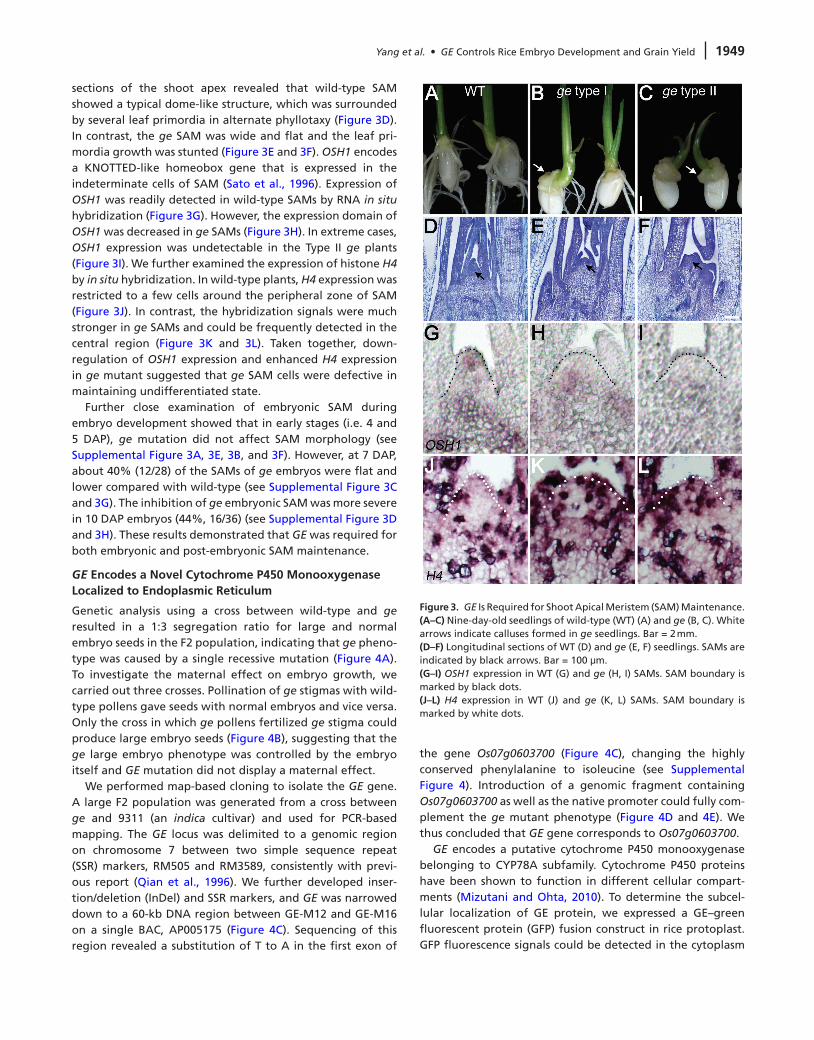
Besides embryogenesis, mutation in GE also affected post-embryonic development. Only a small proportion (13%, n = 85) of the ge mutant could initiate a nearly normal shoot growth and set seeds. A majority of the ge mutants showed defective growth, which could be divided into two types on the basis of their phenotypic severities: type I (65%) and Type II (22%) (see Supplemental Figure 2A and 2B). Type I ge plants displayed intermediate phenotype (Figure 3B), produced roots, and many malformed leaves (see Supplemental Figure 2C and 2D), but died about 1 month after germination. Type II ge mutants showed severe dwarfism with no roots or leaves and died soon after germination (Figure 3C). In addition, we observed frequent callus formation on ge seedlings (Figure 3B and 3C), which never appeared on wild-type plants (Figure 3A).
The aerial tissues and organs of plant are derived from SAM. To investigate whether the defective growth of ge mutants is related to SAM development, we then examined the SAM morphology of ge and wild-type plants. Longitudinal
Figure 1. Characterization of ge Mutant.(A) Morphology of wild-type (WT) and ge seeds during grain filling stages. DAP, days after pollination. The gray dots indicate the interface of embryo and endosperm. Bar = 1 mm.(B) Comparison of mature seed between WT (left) and ge (right). Bar = 1 mm.(C–E) Statistical analysis of embryo area (C), endosperm area (D), and embryo/endosperm ratio (E). The data were presented as means ± SE (n > 90) and statistically analyzed by Student’s t-test (P < 0.01).
1948 Yang et al. • GE Controls Rice Embryo Development and Grain Yield
Figure 2. Morphogenesis of Wild-Type and ge Embryos.(A–H) Sections of wild-type (WT) (A–D) and ge (E–H) embryos during developmental stages. (A, B, E, F): Bar = 50 μm. (C, D, G, H): Bar = 100 μm.(I, J) Morphology of WT (I) and ge (J) scutellum epithelial cells. Bar = 10 μm.(K, L) Morphology of WT (K) and ge (L) scutellum parenchyma cells. Bar = 10 μm.(M) Statistical analysis of scutellum parenchyma cell size (n > 150, P < 0.01).(n) Calculated number of scutellum cells (n = 8).(O, P) In situ hybridization analysis of rice histone H4 expression during WT (O) and ge (P) embryo development. 3, 5, and 7 DAP embryos: Bar = 50 μm. 10 DAP embryo: Bar = 100 μm.
Yang et al. • GE Controls Rice Embryo Development and Grain Yield 1949
sections of the shoot apex revealed that wild-type SAM showed a typical dome-like structure, which was surrounded by several leaf primordia in alternate phyllotaxy (Figure 3D). In contrast, the ge SAM was wide and flat and the leaf pri-mordia growth was stunted (Figure 3E and 3F). OSH1 encodes a KNOTTED-like homeobox gene that is expressed in the indeterminate cells of SAM (Sato et al., 1996). Expression of OSH1 was readily detected in wild-type SAMs by RNA in situ hybridization (Figure 3G). However, the expression domain of OSH1 was decreased in ge SAMs (Figure 3H). In extreme cases, OSH1 expression was undetectable in the Type II ge plants (Figure 3I). We further examined the expression of histone H4 by in situ hybridization. In wild-type plants, H4 expression was restricted to a few cells around the peripheral zone of SAM (Figure 3J). In contrast, the hybridization signals were much stronger in ge SAMs and could be frequently detected in the central region (Figure 3K and 3L). Taken together, down-regulation of OSH1 expression and enhanced H4 expression in ge mutant suggested that ge SAM cells were defective in maintaining undifferentiated state.
Further close examination of embryonic SAM during embryo development showed that in early stages (i.e. 4 and 5 DAP), ge mutation did not affect SAM morphology (see Supplemental Figure 3A, 3E, 3B, and 3F). However, at 7 DAP, about 40% (12/28) of the SAMs of ge embryos were flat and lower compared with wild-type (see Supplemental Figure 3C and 3G). The inhibition of ge embryonic SAM was more severe in 10 DAP embryos (44%, 16/36) (see Supplemental Figure 3D and 3H). These results demonstrated that GE was required for both embryonic and post-embryonic SAM maintenance.
GE Encodes a novel Cytochrome P450 Monooxygenase Localized to Endoplasmic Reticulum
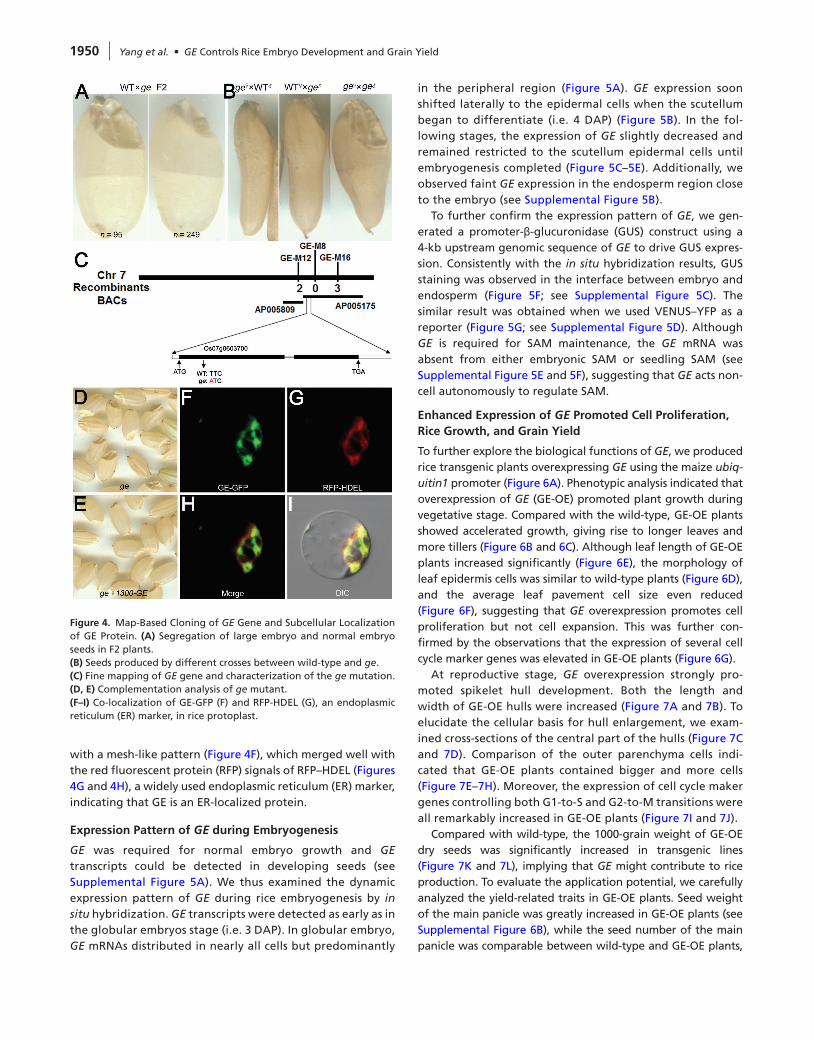
Genetic analysis using a cross between wild-type and ge resulted in a 1:3 segregation ratio for large and normal embryo seeds in the F2 population, indicating that ge pheno-type was caused by a single recessive mutation (Figure 4A). To investigate the maternal effect on embryo growth, we carried out three crosses. Pollination of ge stigmas with wild-type pollens gave seeds with normal embryos and vice versa. Only the cross in which ge pollens fertilized ge stigma could produce large embryo seeds (Figure 4B), suggesting that the ge large embryo phenotype was controlled by the embryo itself and GE mutation did not display a maternal effect.
We performed map-based cloning to isolate the GE gene. A large F2 population was generated from a cross between ge and 9311 (an indica cultivar) and used for PCR-based mapping. The GE locus was delimited to a genomic region on chromosome 7 between two simple sequence repeat (SSR) markers, RM505 and RM3589, consistently with previ-ous report (Qian et al., 1996). We further developed inser-tion/deletion (InDel) and SSR markers, and GE was narrowed down to a 60-kb DNA region between GE-M12 and GE-M16 on a single BAC, AP005175 (Figure 4C). Sequencing of this region revealed a substitution of T to A in the first exon of
the gene Os07g0603700 (Figure 4C), changing the highly conserved phenylalanine to isoleucine (see Supplemental Figure 4). Introduction of a genomic fragment containing Os07g0603700 as well as the native promoter could fully com-plement the ge mutant phenotype (Figure 4D and 4E). We thus concluded that GE gene corresponds to Os07g0603700.
GE encodes a putative cytochrome P450 monooxygenase belonging to CYP78A subfamily. Cytochrome P450 proteins have been shown to function in different cellular compart-ments (Mizutani and Ohta, 2010). To determine the subcel-lular localization of GE protein, we expressed a GE–green fluorescent protein (GFP) fusion construct in rice protoplast. GFP fluorescence signals could be detected in the cytoplasm
Figure 3. GE Is Required for Shoot Apical Meristem (SAM) Maintenance.(A–C) Nine-day-old seedlings of wild-type (WT) (A) and ge (B, C). White arrows indicate calluses formed in ge seedlings. Bar = 2 mm.(d–F) Longitudinal sections of WT (D) and ge (E, F) seedlings. SAMs are indicated by black arrows. Bar = 100 μm.(G–I) OSH1 expression in WT (G) and ge (H, I) SAMs. SAM boundary is marked by black dots.(J–L) H4 expression in WT (J) and ge (K, L) SAMs. SAM boundary is marked by white dots.
1950 Yang et al. • GE Controls Rice Embryo Development and Grain Yield
with a mesh-like pattern (Figure 4F), which merged well with the red fluorescent protein (RFP) signals of RFP–HDEL (Figures 4G and 4H), a widely used endoplasmic reticulum (ER) marker, indicating that GE is an ER-localized protein.
Expression Pattern of GE during Embryogenesis
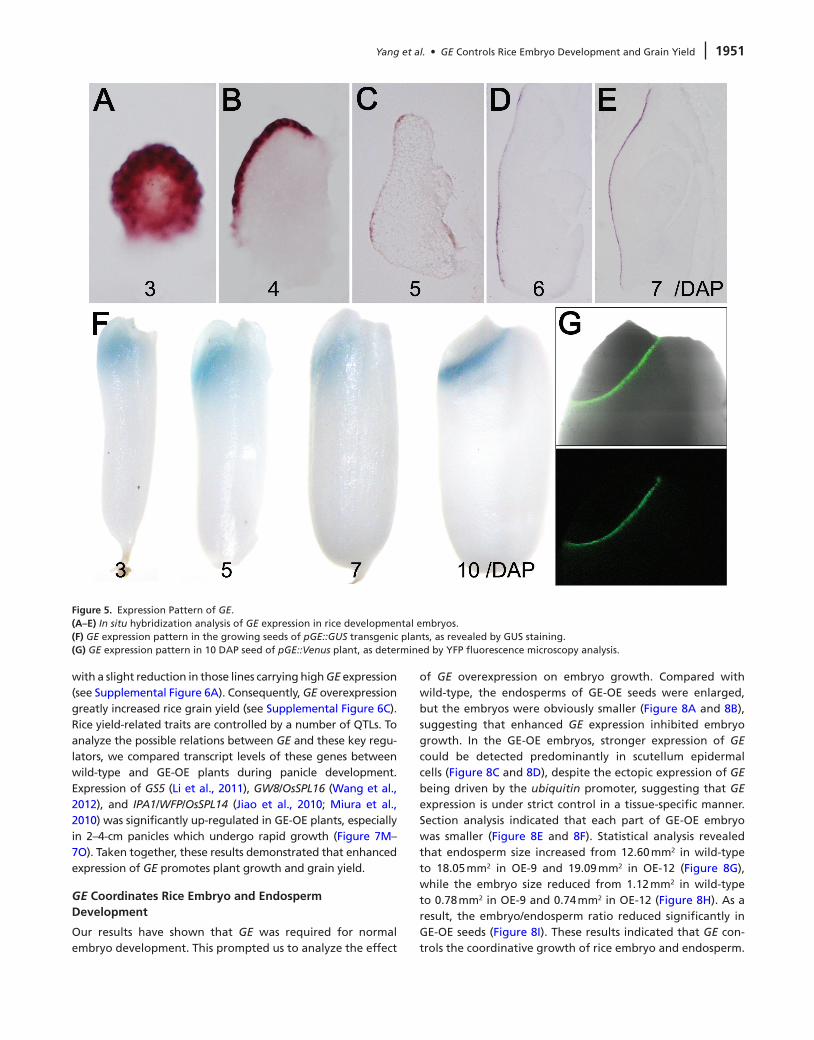
GE was required for normal embryo growth and GE transcripts could be detected in developing seeds (see Supplemental Figure 5A). We thus examined the dynamic expression pattern of GE during rice embryogenesis by in situ hybridization. GE transcripts were detected as early as in the globular embryos stage (i.e. 3 DAP). In globular embryo, GE mRNAs distributed in nearly all cells but predominantly
in the peripheral region (Figure 5A). GE expression soon shifted laterally to the epidermal cells when the scutellum began to differentiate (i.e. 4 DAP) (Figure 5B). In the fol-lowing stages, the expression of GE slightly decreased and remained restricted to the scutellum epidermal cells until embryogenesis completed (Figure 5C–5E). Additionally, we observed faint GE expression in the endosperm region close to the embryo (see Supplemental Figure 5B).
To further confirm the expression pattern of GE, we gen-erated a promoter-β-glucuronidase (GUS) construct using a 4-kb upstream genomic sequence of GE to drive GUS expres-sion. Consistently with the in situ hybridization results, GUS staining was observed in the interface between embryo and endosperm (Figure 5F; see Supplemental Figure 5C). The similar result was obtained when we used VENUS–YFP as a reporter (Figure 5G; see Supplemental Figure 5D). Although GE is required for SAM maintenance, the GE mRNA was absent from either embryonic SAM or seedling SAM (see Supplemental Figure 5E and 5F), suggesting that GE acts non-cell autonomously to regulate SAM.
Enhanced Expression of GE Promoted Cell Proliferation, Rice Growth, and Grain Yield
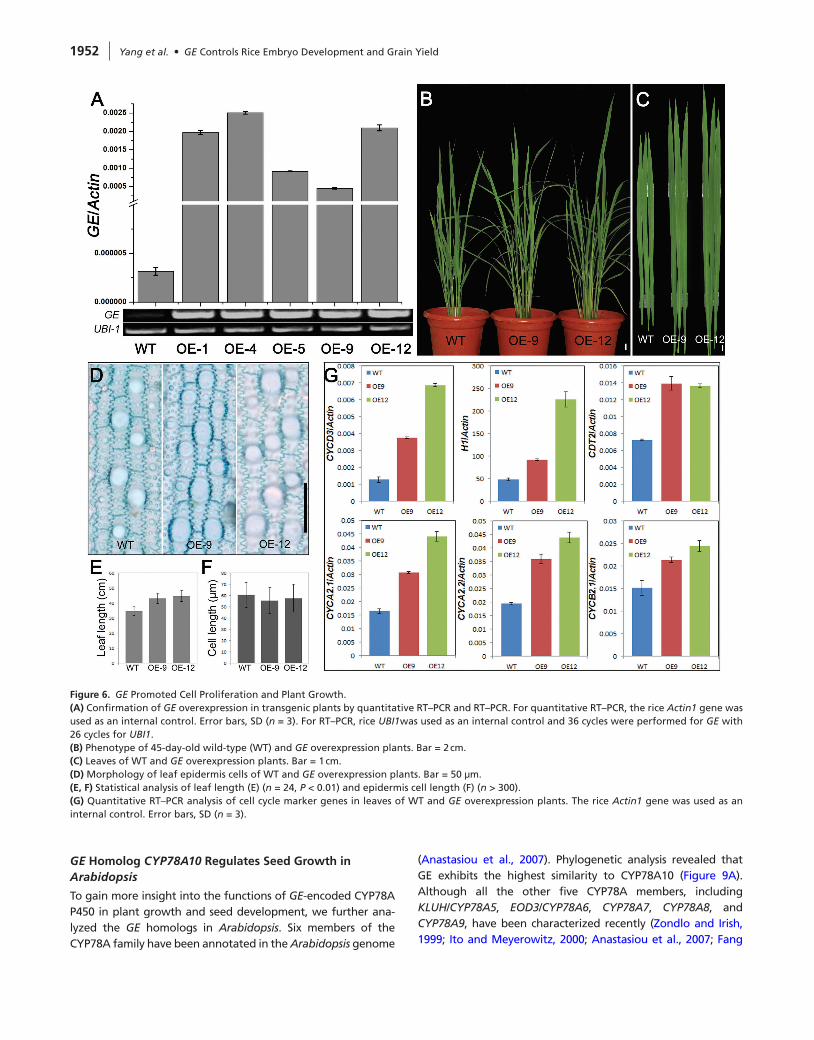
To further explore the biological functions of GE, we produced rice transgenic plants overexpressing GE using the maize ubiq-uitin1 promoter (Figure 6A). Phenotypic analysis indicated that overexpression of GE (GE-OE) promoted plant growth during vegetative stage. Compared with the wild-type, GE-OE plants showed accelerated growth, giving rise to longer leaves and more tillers (Figure 6B and 6C). Although leaf length of GE-OE plants increased significantly (Figure 6E), the morphology of leaf epidermis cells was similar to wild-type plants (Figure 6D), and the average leaf pavement cell size even reduced (Figure 6F), suggesting that GE overexpression promotes cell proliferation but not cell expansion. This was further con-firmed by the observations that the expression of several cell cycle marker genes was elevated in GE-OE plants (Figure 6G).
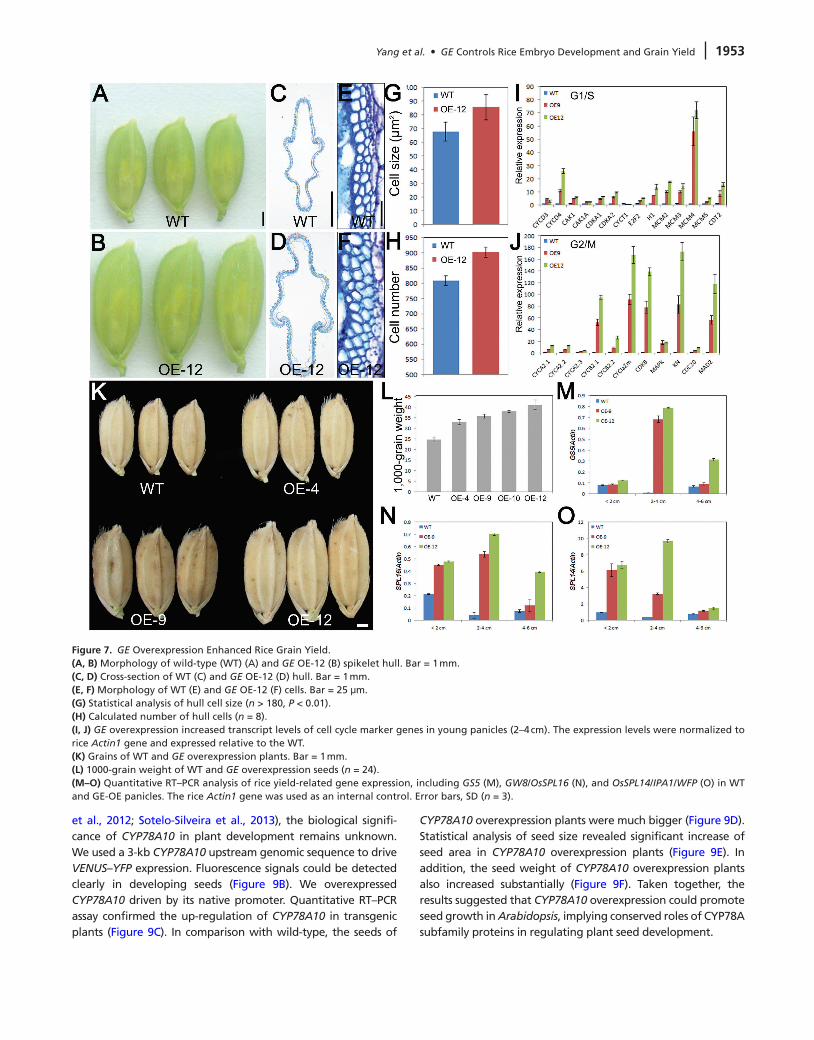
At reproductive stage, GE overexpression strongly pro-moted spikelet hull development. Both the length and width of GE-OE hulls were increased (Figure 7A and 7B). To elucidate the cellular basis for hull enlargement, we exam-ined cross-sections of the central part of the hulls (Figure 7C and 7D). Comparison of the outer parenchyma cells indi-cated that GE-OE plants contained bigger and more cells (Figure 7E–7H). Moreover, the expression of cell cycle maker genes controlling both G1-to-S and G2-to-M transitions were all remarkably increased in GE-OE plants (Figure 7I and 7J).
Compared with wild-type, the 1000-grain weight of GE-OE dry seeds was significantly increased in transgenic lines (Figure 7K and 7L), implying that GE might contribute to rice production. To evaluate the application potential, we carefully analyzed the yield-related traits in GE-OE plants. Seed weight of the main panicle was greatly increased in GE-OE plants (see Supplemental Figure 6B), while the seed number of the main panicle was comparable between wild-type and GE-OE plants,
Figure 4. Map-Based Cloning of GE Gene and Subcellular Localization of GE Protein. (A) Segregation of large embryo and normal embryo seeds in F2 plants.(B) Seeds produced by different crosses between wild-type and ge.(C) Fine mapping of GE gene and characterization of the ge mutation.(d, E) Complementation analysis of ge mutant.(F–I) Co-localization of GE-GFP (F) and RFP-HDEL (G), an endoplasmic reticulum (ER) marker, in rice protoplast.
Yang et al. • GE Controls Rice Embryo Development and Grain Yield 1951
with a slight reduction in those lines carrying high GE expression (see Supplemental Figure 6A). Consequently, GE overexpression greatly increased rice grain yield (see Supplemental Figure 6C). Rice yield-related traits are controlled by a number of QTLs. To analyze the possible relations between GE and these key regu-lators, we compared transcript levels of these genes between wild-type and GE-OE plants during panicle development. Expression of GS5 (Li et al., 2011), GW8/OsSPL16 (Wang et al., 2012), and IPA1/WFP/OsSPL14 (Jiao et al., 2010; Miura et al., 2010) was significantly up-regulated in GE-OE plants, especially in 2–4-cm panicles which undergo rapid growth (Figure 7M–7O). Taken together, these results demonstrated that enhanced expression of GE promotes plant growth and grain yield.
GE Coordinates Rice Embryo and Endosperm development
Our results have shown that GE was required for normal embryo development. This prompted us to analyze the effect
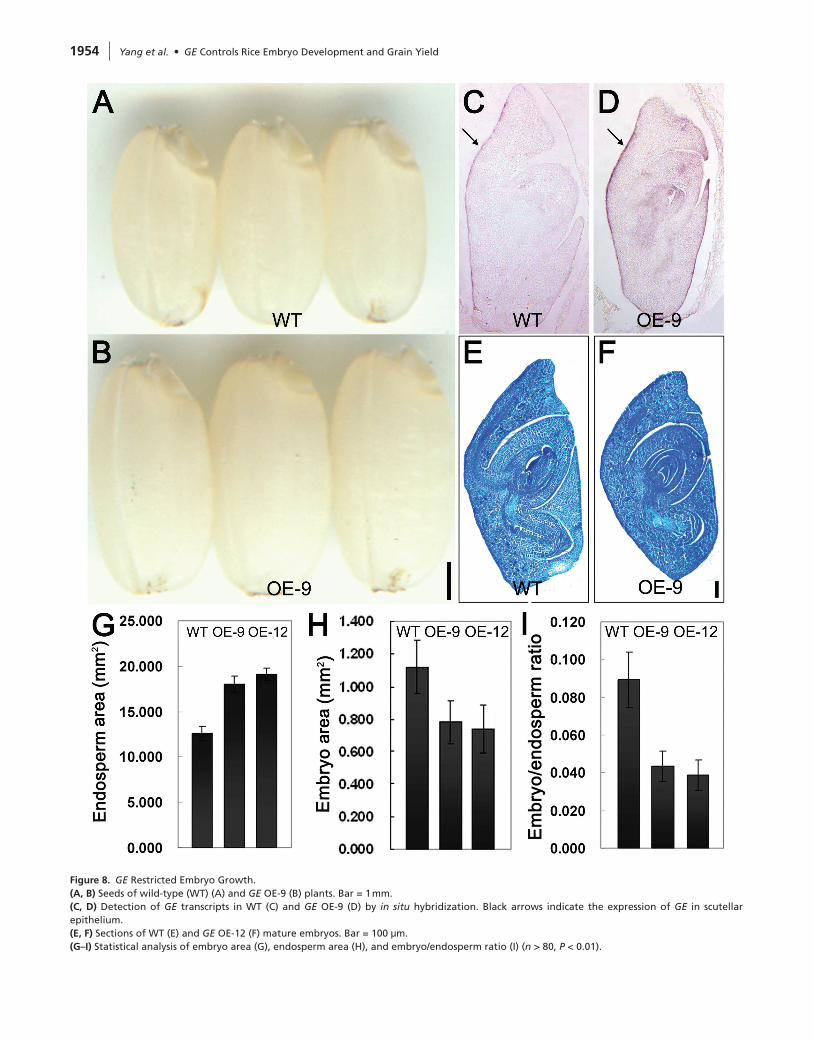
of GE overexpression on embryo growth. Compared with wild-type, the endosperms of GE-OE seeds were enlarged, but the embryos were obviously smaller (Figure 8A and 8B), suggesting that enhanced GE expression inhibited embryo growth. In the GE-OE embryos, stronger expression of GE could be detected predominantly in scutellum epidermal cells (Figure 8C and 8D), despite the ectopic expression of GE being driven by the ubiquitin promoter, suggesting that GE expression is under strict control in a tissue-specific manner. Section analysis indicated that each part of GE-OE embryo was smaller (Figure 8E and 8F). Statistical analysis revealed that endosperm size increased from 12.60 mm2 in wild-type to 18.05 mm2 in OE-9 and 19.09 mm2 in OE-12 (Figure 8G), while the embryo size reduced from 1.12 mm2 in wild-type to 0.78 mm2 in OE-9 and 0.74 mm2 in OE-12 (Figure 8H). As a result, the embryo/endosperm ratio reduced significantly in GE-OE seeds (Figure 8I). These results indicated that GE con-trols the coordinative growth of rice embryo and endosperm.
Figure 5. Expression Pattern of GE.(A–E) In situ hybridization analysis of GE expression in rice developmental embryos.(F) GE expression pattern in the growing seeds of pGE::GUS transgenic plants, as revealed by GUS staining.(G) GE expression pattern in 10 DAP seed of pGE::Venus plant, as determined by YFP fluorescence microscopy analysis.
1952 Yang et al. • GE Controls Rice Embryo Development and Grain Yield
GE Homolog CYP78A10 Regulates Seed Growth in Arabidopsis
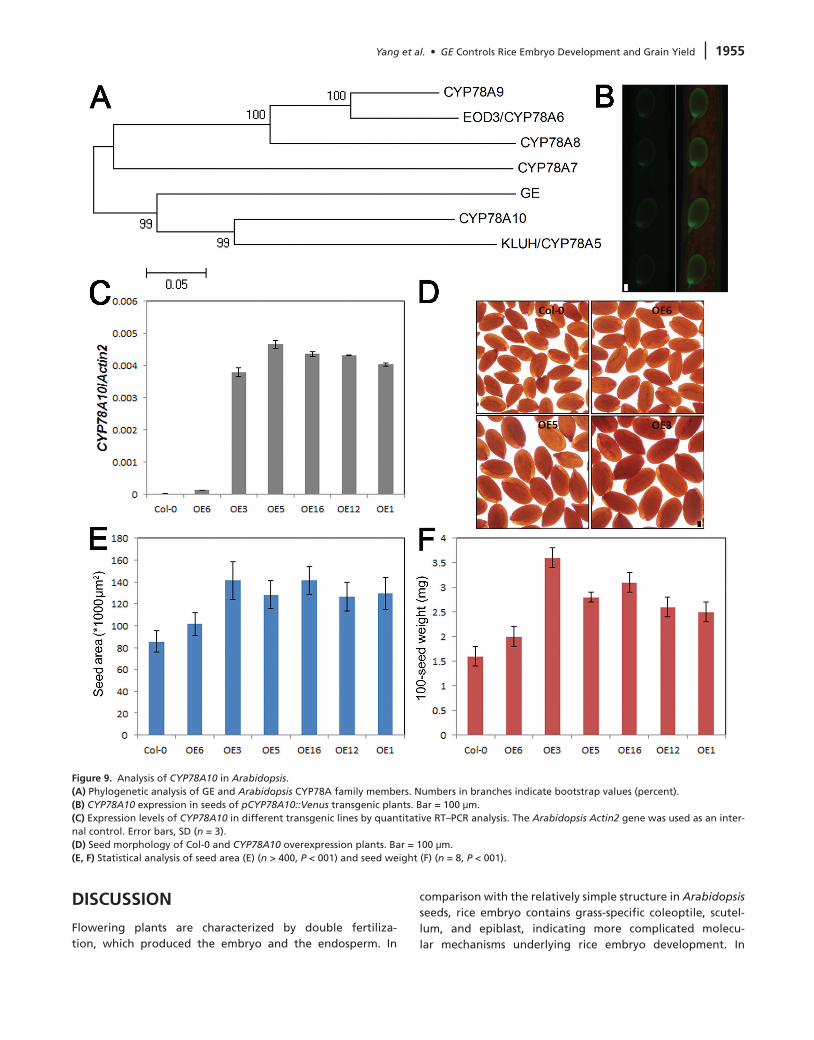
To gain more insight into the functions of GE-encoded CYP78A P450 in plant growth and seed development, we further ana-lyzed the GE homologs in Arabidopsis. Six members of the CYP78A family have been annotated in the Arabidopsis genome
(Anastasiou et al., 2007). Phylogenetic analysis revealed that GE exhibits the highest similarity to CYP78A10 (Figure 9A). Although all the other five CYP78A members, including KLUH/CYP78A5, EOD3/CYP78A6, CYP78A7, CYP78A8, and CYP78A9, have been characterized recently (Zondlo and Irish, 1999; Ito and Meyerowitz, 2000; Anastasiou et al., 2007; Fang
Figure 6. GE Promoted Cell Proliferation and Plant Growth.(A) Confirmation of GE overexpression in transgenic plants by quantitative RT–PCR and RT–PCR. For quantitative RT–PCR, the rice Actin1 gene was used as an internal control. Error bars, SD (n = 3). For RT–PCR, rice UBI1was used as an internal control and 36 cycles were performed for GE with 26 cycles for UBI1.(B) Phenotype of 45-day-old wild-type (WT) and GE overexpression plants. Bar = 2 cm.(C) Leaves of WT and GE overexpression plants. Bar = 1 cm.(d) Morphology of leaf epidermis cells of WT and GE overexpression plants. Bar = 50 μm.(E, F) Statistical analysis of leaf length (E) (n = 24, P < 0.01) and epidermis cell length (F) (n > 300).(G) Quantitative RT–PCR analysis of cell cycle marker genes in leaves of WT and GE overexpression plants. The rice Actin1 gene was used as an internal control. Error bars, SD (n = 3).
Yang et al. • GE Controls Rice Embryo Development and Grain Yield 1953
et al., 2012; Sotelo-Silveira et al., 2013), the biological signifi-cance of CYP78A10 in plant development remains unknown. We used a 3-kb CYP78A10 upstream genomic sequence to drive VENUS–YFP expression. Fluorescence signals could be detected clearly in developing seeds (Figure 9B). We overexpressed CYP78A10 driven by its native promoter. Quantitative RT–PCR assay confirmed the up-regulation of CYP78A10 in transgenic plants (Figure 9C). In comparison with wild-type, the seeds of
CYP78A10 overexpression plants were much bigger (Figure 9D). Statistical analysis of seed size revealed significant increase of seed area in CYP78A10 overexpression plants (Figure 9E). In addition, the seed weight of CYP78A10 overexpression plants also increased substantially (Figure 9F). Taken together, the results suggested that CYP78A10 overexpression could promote seed growth in Arabidopsis, implying conserved roles of CYP78A subfamily proteins in regulating plant seed development.
Figure 7. GE Overexpression Enhanced Rice Grain Yield.(A, B) Morphology of wild-type (WT) (A) and GE OE-12 (B) spikelet hull. Bar = 1 mm.(C, d) Cross-section of WT (C) and GE OE-12 (D) hull. Bar = 1 mm.(E, F) Morphology of WT (E) and GE OE-12 (F) cells. Bar = 25 μm.(G) Statistical analysis of hull cell size (n > 180, P < 0.01).(H) Calculated number of hull cells (n = 8).(I, J) GE overexpression increased transcript levels of cell cycle marker genes in young panicles (2–4 cm). The expression levels were normalized to rice Actin1 gene and expressed relative to the WT.(K) Grains of WT and GE overexpression plants. Bar = 1 mm.(L) 1000-grain weight of WT and GE overexpression seeds (n = 24).(M–O) Quantitative RT–PCR analysis of rice yield-related gene expression, including GS5 (M), GW8/OsSPL16 (N), and OsSPL14/IPA1/WFP (O) in WT and GE-OE panicles. The rice Actin1 gene was used as an internal control. Error bars, SD (n = 3).
1954 Yang et al. • GE Controls Rice Embryo Development and Grain Yield
Figure 8. GE Restricted Embryo Growth.(A, B) Seeds of wild-type (WT) (A) and GE OE-9 (B) plants. Bar = 1 mm.(C, d) Detection of GE transcripts in WT (C) and GE OE-9 (D) by in situ hybridization. Black arrows indicate the expression of GE in scutellar epithelium.(E, F) Sections of WT (E) and GE OE-12 (F) mature embryos. Bar = 100 μm.(G–I) Statistical analysis of embryo area (G), endosperm area (H), and embryo/endosperm ratio (I) (n > 80, P < 0.01).
Yang et al. • GE Controls Rice Embryo Development and Grain Yield 1955
dISCuSSIOn
Flowering plants are characterized by double fertiliza-tion, which produced the embryo and the endosperm. In
comparison with the relatively simple structure in Arabidopsis seeds, rice embryo contains grass-specific coleoptile, scutel-lum, and epiblast, indicating more complicated molecu-lar mechanisms underlying rice embryo development. In
Figure 9. Analysis of CYP78A10 in Arabidopsis.(A) Phylogenetic analysis of GE and Arabidopsis CYP78A family members. Numbers in branches indicate bootstrap values (percent).(B) CYP78A10 expression in seeds of pCYP78A10::Venus transgenic plants. Bar = 100 μm.(C) Expression levels of CYP78A10 in different transgenic lines by quantitative RT–PCR analysis. The Arabidopsis Actin2 gene was used as an inter-nal control. Error bars, SD (n = 3).(d) Seed morphology of Col-0 and CYP78A10 overexpression plants. Bar = 100 μm.(E, F) Statistical analysis of seed area (E) (n > 400, P < 001) and seed weight (F) (n = 8, P < 001).

1956 Yang et al. • GE Controls Rice Embryo Development and Grain Yield
addition, Arabidopsis endosperm is a transient tissue that is rapidly consumed by the developing embryo. In sharp con-trast, rice endosperm grows continuously and occupies most of the space in the mature seed. Therefore, coordination of embryo and endosperm development is of special importance for proper rice seed development, although the molecular signaling pathways remain poorly understood.
The primary function of endosperm is to support embryo growth by delivering nutrients. The sucrose transporters AtSUC5 and OsSUT1 both expressed in the endosperm and were suspected to deliver sucrose to the embryo (Hirose et al., 1997; Baud et al., 2005). Disruption of endosperm develop-ment such as suppressing cellularization resulted in failure of glucose and fructose translocation to embryo and subsequent embryo defects (Hehenberger et al., 2012). Besides serving as a nutritive tissue for the embryo, the endosperm also gener-ates signal molecules to modulate embryo growth, including small secreted peptides (Fiume and Fletcher, 2012). On the other hand, the developing embryo also sends positive sig-nals to stimulate endosperm growth, since the cdc2a mutant could only produce one sperm to fertilize the egg cell, but the unfertilized endosperm still developed (Nowack et al., 2006). In rice, it was suggested that growth of the endosperm and embryo restricts each other due to space limitation (Hong et al., 1996). Indeed, the temperature-sensitive embryoless 1 produced embryoless seeds with large endosperm at high temperature but large embryo seeds containing small or even no endosperm at low temperature (Honga et al., 1995). Down-regulation of Orysa;CycB1;1 expression caused severe endosperm defects but produced very large embryos (Guo et al., 2010). We found that mutation in GE resulted in seeds with large embryos and small endosperms and GE overex-pression produced big endosperm but small embryo seeds. Therefore, GE might function as a master regulator that coor-dinates rice embryo and endosperm growth.
GE encodes a cytochromes P450 belonging to the CYP78A subfamily and expressed predominantly in the epidermal cells of globular embryo and scutellum. Epidermal cells are the first differentiated cells during embryogenesis (Itoh et al., 2005; Lau et al., 2012). These palisade-shaped cells often contain cutin depositions on the cell wall surface, which are supposed to protect the embryo from changing environment and organ fusion during embryo development (Tanaka et al., 2001). A specific set of genes involved in fatty acid metabo-lism such as FDH (Yephremov et al., 1999) and signal trans-duction such as ATML1, PDF2, ACR4, and ALE2 (Tanaka et al., 2002, 2007) are expressed in these cells to control cutin for-mation and normal embryo development. The cutin layer has also been predicted to exist in the scutellum epithelial cells of monocots such as rice, wheat, and barley (Fincher, 1989). The cutin layer constitutes a physical barrier to prevent pas-sive nutrient transfer from endosperm to embryo (Schreiber, 2010). Members of the CYP78A family have been shown to exhibit fatty acid ω-hydroxylation activity in vitro (Imaishi et al., 2000; Kai et al., 2009), while C16 and C18 ω-hydroxy fatty acids are the basic components of cutin (Pollard et al.,
2008). Thus, we propose that loss of function of GE P450 might change the physical properties of the scutellar epithe-lium cell walls or disrupt the cuticle structure, leading to more nutrient flux into the embryo, giving rise to enhanced embryo growth. However, another possibility could not be excluded that GE might be involved in the production of novel sign-aling molecules, given that P450s have been implicated in the biosynthesis or catabolism of plant hormones (Mizutani and Ohta, 2010). In addition, several pieces of evidence indi-cated that some plant hormones are produced specifically in rice scutellar epithelium cells. For example, gibberellin (GA) biosynthetic genes are expressed in the epithelium (Kaneko et al., 2003). And expression of auxin-responsive DR5–GUS and auxin biosynthesis gene TDD1 can also be detected in this layer of cells, suggesting that scutellar epithelium is the auxin maximum region (Sazuka et al., 2009). Therefore, GE may alternatively generate a novel growth regulator that controls embryo growth and SAM maintenance.
CYP78A family contains six members in Arabidopsis, five of which have been shown to regulate flower and seed develop-ment (Mizutani and Ohta, 2010). Mutation of KLUH/CYP78A5 resulted in small flowers and seeds due to reduced cell pro-liferation (Anastasiou et al., 2007); CYP78A9 act redundantly with EOD3/CYP78A6 and CYP78A8 to control seed size and outer integument development (Fang et al., 2012; Sotelo-Silveira et al., 2013). Overexpression of KLUH/CYP78A5, EOD3/CYP78A6, and CYP78A9 displayed similar growth phe-notypes, including delayed flower bud opening, longevity, big flowers with short stamens and reduced fertility (Ito and Meyerowitz, 2000; Anastasiou et al., 2007; Fang et al., 2012; Sotelo-Silveira et al., 2013). So far, the only identified CYP78A member in rice is PLASTOCHRON1 (PLA1), which encodes the CYP78A11 to control leaf initiation (Miyoshi et al., 2004). Similarly to rice pla1mutant, the cyp78a5/klu mutant also dis-played a faster leaf initiation phenotype (Wang et al., 2008b). Our current study identified GE P450, adding new insight into the biological roles of CYP78A in controlling rice embryo development and SAM maintenance. Interestingly, another rice plastochron mutant pla3 also contained enlarged embryos similar to ge (Kawakatsu et al., 2009). PLA3 encodes a putative glutamate carboxypeptidase, whose Arabidopsis homolog AMP1 has been shown to be required for miRNA inhibition of target mRNA translation most recently (Helliwell et al., 2001; Li et al., 2013). These observations indicated a genetic relationship between CYP78A P450s, AMP1, and PLA3-encoded glutamate carboxypeptidases, which probably involves miRNA-mediated translation inhibition. Despite the biological significance, the catalytic functions of plant CYP78A P450s remain largely unknown. Given that the growth phe-notypes of ge and other CYP78A mutants are unrelated to classical phytohormones (Anastasiou et al., 2007), and their probable fatty acid ω-hydroxylation activities (Imaishi et al., 2000; Kai et al., 2009), it is tempting to speculate that GE would produce a novel fatty acid-related plant growth regu-lator or signal molecule that inhibits scutellum cell expansion in rice embryo.

Yang et al. • GE Controls Rice Embryo Development and Grain Yield 1957
Seed size is an important agronomic trait that controls rice grain yield. We found that increasing the expression of a single gene, GE, could sufficiently promote rice seed growth and subsequent yield improvement. Rice grain yield is con-trolled by a number of QTLs derived from natural variations (Yano, 2001). In GE-OE plants, the expression of several QTLs increased significantly, including GS5, GW8, and IPA1. GS5 encodes a putative serine carboxypeptidase, whereas GW8 and IPA1 both encode SQUAMOSA PROMOTER BINDING PROTEIN-LIKE transcription factors, all of which are positive regulators of grain size (Jiao et al., 2010; Miura et al., 2010; Li et al., 2011; Wang et al., 2012). GE functions together with these key regulators to promote cell proliferation and expansion, leading to enhanced hull growth and grain fill-ing. In addition, overexpression of GE homolog CYP78A10 in Arabidopsis also produced bigger and heavier seeds, sug-gesting a conserved role for this class of P450 proteins in facilitating seed growth. Very recently, Nagasawa et al. (2013) reported the characterization of the same locus responsible for rice GE phenotype. They showed that GE functions in the embryo to control cell size and in the endosperm to regulate cell death via ROS signaling, and GE acts downstream of the REDUCED EMBRYO genes. Consistently with our study, they also found that GE predominantly expressed in the interface of the embryo and the endosperm, and increased GE expres-sion caused small embryo, emphasizing a pivotal role of GE in rice embryo development as well as embryo and endosperm coordination. The absence of GE mRNA in the SAM indicated that GE might generate a mobile signal to regulate SAM development in a non-cell autonomous manner, as suggested for KLUH/CYP78A5 (Anastasiou et al., 2007). In supporting of this hypothesis, Nagasawa et al. (2013) demonstrated that GE expression in either embryo or endosperm could control embryo/endosperm size. Furthermore, based on compre-hensive cellular, genetic, and molecular biology analysis, our current study revealed additional functions of GE in SAM main-tenance and rice grain yield improvement. Characterization of the catalytic functions of GE and other CYP78As and the possible signal molecule(s) they produced may reveal a new mechanism underlying plant growth regulation and provide new approaches for improving rice grain yield.
METHOdSPlant Materials and Growth Conditions
The rice (Oryza sativa L.) ge mutant was isolated from EMS-induced mutation of a japonica cultivar (cv. Dongjin). Plants were cultivated in an experimental field during natural grow-ing seasons. For seedling analysis, sterile rice seeds were grown on ½ Murashige and Skoog (MS) medium in a growth chamber under conditions of 12-h day, 28°C, 80% RH followed by 12-h night, 26°C, 60% RH. For Arabidopsis growing, seeds were sur-face-sterilized and sowed on ½ MS medium. After 3 d at 4°C in the dark, plates were transferred to a growth chamber at 20–22°C under a 16-h light/8-h dark photoperiod. One-week-old seedlings were transferred to soil for further growth.
For statistical analysis of seed, embryo, and endosperm area, seeds of rice and Arabidopsis were photographed. The digital images were used for measurement in Image J soft-ware (version 1.38; http://rsbweb.nih.gov/ij/) and analyzed by Student’s t-test in the R programming language (version 2.11.0; www.R-project.org).
Histological Analysis
Tissues were fixed overnight at 4°C in FAA (50% ethanol, 5% acetic acid, and 3.7% formaldehyde) and dehydrated in a graded ethanol series. Following substitution with xylene, the samples were embedded in Paraplast (Leica) and sectioned at 8-μm thickness using a rotary microtome (Leica). Sections were stained with 0.05% toluidine blue and observed with a light microscope (Leica). For resin sections, samples were fixed in FAA and dehydrated in an ethanol series. The sam-ples were embedded in Technovit 7100 resin (Hereaus Kulzer) and polymerized at 45°C. Transverse sections of 2 μm were cut using an Ultratome III ultramicrotome (Leica) and then stained by periodic acid-Schiff reaction for starches and amido black for proteins.
Map-Based Cloning of GE and Complementation
The ge mutant was crossed with 9311 (an indica cultivar) to generate an F2 mapping population. GE was mapped to a 60-kb region on chromosome 7. Genomic DNA fragments of this region were amplified from ge and wild-type plants, sequenced, and compared using MegAlign (DNASTAR). For complementation, a 7.8-kb genomic DNA fragment contain-ing the entire GE coding region, the 3.5-kb promoter region, and the 2.6-kb 3’ region was digested from BAC AP005175. The fragment was inserted into the binary vector pCambia1300 (GenBank accession number AF234296). The resulting con-struct, 1300-GE, was then introduced into the ge background via Agrobacterium tumefaciens-mediated transformation. More than 15 independent transgenic lines were produced that could successfully complement the mutant phenotypes.
Subcellular Localization of GE
The coding sequencing of GE was amplified using primers GE-GFP-F (XhoI) and GE-GFP-R (SpeI) and inserted into pA7–GFP. The resulting GE-GFP, along with RFP-HDEL, was co-transformed into rice protoplasts prepared from leaf sheaths of 11-day-old seedlings. Rice protoplast transformation was performed according to previously reported methods (Bart et al., 2006).
Overexpression and Tissue-Specific Expression of GE and CYP79A10
For GE overexpression, the full-length genomic DNA sequence was amplified using primers GE-OE-F (BamHI) and GE-OE-R (KpnI). The resulting fragment was subsequently cloned into the binary vector pUN1301 and transformed into wild-type plants (japonica cv. Nipponbare). More than 15 independent transgenic lines were produced and used for further analysis. For reporter gene construction, the GUS coding sequence as

1958 Yang et al. • GE Controls Rice Embryo Development and Grain Yield
well as Nos terminator was digested from pBI101 using HindIII and EcoRI, and then inserted into pCambia1300, resulting in 1300-GUS-Nos. The VENUS–YFP coding sequencing was amplified using primers Venus-F (BamHI) and Venus-R (KpnI) and then cloned into 1300-GUS-Nos, resulting in 1300-Venus-Nos. A 4-kb promoter of GE was amplified using primers GE-promoter-F (SalI) and GE-promoter-R (BamHI) and intro-duced into 1300-GUS-Nos and 1300-Venus-Nos. The resulting constructs, pGE::GUS and pGE::Venus were transformed into wild-type Nipponbare.
A 3-kb promoter of CYP78A10 was amplified using primers A10-promoter-F (SalI) and A10-promoter-R (BamHI), and then introduced into 1300-Venus-Nos, giving rise to pA10::Venus. The genomic sequencing of CYP78A10 was amplified using primers A10-OE-F (SmaI) and A10-OE-R (SacI), and then intro-duced into pA10::Venus to produce the construct pA10::gDNA for overexpression. pA10::Venus and pA10::gDNA were trans-formed into Arabidopsis Col-0. More than 30 positive trans-genic lines were generated and used for analysis.
For GUS staining, various tissues of pGE::GUS transgenic plants were incubated in a solution containing 50 mM NaPO4 buffer (pH 7.0), 5 mM K3Fe(CN)6, 5 mM K4Fe(CN)6, 0.1% Triton X-100, and 1 mM X-Gluc and stained at 37°C.
RnA Isolation and Quantitative RT–PCR
Total RNAs were extracted from different tissues of rice and Arabidopsis plants using TRIzol reagent (Invitrogen) according to the manufacturer’s instructions. For RT–PCR analysis, total RNA (2 μg) was reverse transcribed into cDNA using oligo(dT) primer and SuperScript III reverse transcriptase (Invitrogen) and then used as templates for PCR with gene-specific primers. Quantitative RT–PCR analysis was performed using SYBR Premix Ex Taq (TaKaRa) and gene-specific primers. The rice Actin1 and Arabidopsis Actin2 genes were used as referent genes.
Phylogenetic Analysis
Protein sequences of CYP78A family members in rice, Arabidopsis, maize, medicago, and moss were obtained from the CYP450 Engineering Database v2 (www.cyped.uni-stuttgart.de/cgi-bin/CYPED/index_org). Multiple sequence alignments of protein were done in Clustal X (1.83). A phy-logenetic tree of the aligned sequence was constructed by MEGA (version 4.0.2) using the neighbor-joining method with the following parameters: Poisson correction, complete dele-tion, and bootstrap (1000 replicates, random seed).
In Situ Hybridization
For in situ hybridization, digoxigenin-labeled RNAs were pro-duced from the coding sequences of GE, OSH1, and H4 accord-ing to the manufacturer’s instructions (Roche). Tissues were fixed in 0.1 M sodium phosphate buffer containing 4% para-formaldehyde, 0.25% glutaraldehyde, 0.1% Tween-20, and 0.1% Triton X-100. Fixed samples were embedded in Paraplast Plus and sectioned at 8–10-μm thickness. The sections were mounted on Poly-L-lysine-treated slides (Sigma). Hybridization
and immunological detection of the hybridized signals were performed as previously described (Sato et al., 1996).
SuPPLEMEnTARY dATASupplementary Data are available at Molecular Plant Online.
FUNDING
This work was supported by Natural Science Foundation of China grants (91117018) and by the CAS International Partnership Program for Creative Research Teams.
ACKNOWLEDGMENTS
We are grateful to Dr. Jiawei Wang and Dr. Ertao Wang (Shanghai Institute of Plant Physiology and Ecology) for criti-cal reading of the manuscript. We thank Xiaoyan Gao, Xiaoshu Gao, Jiexin Li, Zhiping Zhang, and Guoxin Xu for sample prep-aration, electron microscopy, and confocal microscopy help. No conflict of interest declared.
REFERENCES
Aichinger, E., Kornet, n., Friedrich, T., and Laux, T. (2012). Plant stem cell niches. Annu. Rev. Plant Biol. 63, 615–636.
Anastasiou, E., Kenz, S., Gerstung, M., MacLean, d., Timmer, J., Fleck, C., and Lenhard, M. (2007). Control of plant organ size by KLUH/CYP78A5-dependent intercellular signaling. Dev. Cell. 13, 843–856.
Bart, R., Chern, M., Park, C.J., Bartley, L., and Ronald, P.C. (2006). A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts. Plant Methods. 2, 13.
Baud, S., Wuilleme, S., Lemoine, R., Kronenberger, J., Caboche, M., Lepiniec, L., and Rochat, C. (2005). The AtSUC5 sucrose trans-porter specifically expressed in the endosperm is involved in early seed development in Arabidopsis. Plant J. 43, 824–836.
Benkova, E., Michniewicz, M., Sauer, M., Teichmann, T., Seifertova, d., Jurgens, G., and Friml J. (2003). Local, efflux-dependent auxin gradients as a common module for plant organ forma-tion. Cell. 115, 591–602.
Brand, u., Fletcher, J.C., Hobe, M., Meyerowitz, E.M., and Simon, R. (2000). Dependence of stem cell fate in Arabidopsis on a feed-back loop regulated by CLV3 activity. Science. 289, 617–619.
Breuninger, H., Rikirsch, E., Hermann, M., ueda, M., and Laux, T. (2008). Differential expression of WOX genes mediates apical–basal axis formation in the Arabidopsis embryo. Dev. Cell. 14, 867–876.
dresselhaus, T. (2006). Cell–cell communication during double fer-tilization. Curr. Opin. Plant Biol. 9, 41–47.
Fan, C., Xing, Y., Mao, H., Lu, T., Han, B., Xu, C., Li, X., and Zhang, Q. (2006). GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet. 112, 1164–1171.
Fang, W., Wang, Z., Cui, R., Li, J., and Li, Y. (2012). Maternal control of seed size by EOD3/CYP78A6 in Arabidopsis thaliana. Plant J. 70, 929–939.

Yang et al. • GE Controls Rice Embryo Development and Grain Yield 1959
Fincher, G.B. (1989). Molecular and cellular biology associated with endosperm mobilization in germinating cereal grains. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40, 305–346.
Fiume, E., and Fletcher, J.C. (2012). Regulation of Arabidopsis embryo and endosperm development by the polypeptide sign-aling molecule CLE8. Plant Cell. 24, 1000–1012.
Friml, J., Vieten, A., Sauer, M., Weijers, d., Schwarz, H., Hamann, T., Offringa, R., and Jurgens, G. (2003). Efflux-dependent auxin gradients establish the apical–basal axis of Arabidopsis. Nature. 426, 147–153.
Garcia, d., Fitz Gerald, J.n., and Berger, F. (2005). Maternal con-trol of integument cell elongation and zygotic control of endosperm growth are coordinated to determine seed size in Arabidopsis. Plant Cell. 17, 52–60.
Geldner, n., Anders, n., Wolters, H., Keicher, J., Kornberger, W., Muller, P., delbarre, A., ueda, T., nakano, A., and Jurgens, G. (2003). The Arabidopsis GNOM ARF-GEF mediates endosomal recycling, auxin transport, and auxin-dependent plant growth. Cell. 112, 219–230.
Guo, J., Wang, F., Song, J., Sun, W., and Zhang, X.S. (2010). The expression of Orysa;CycB1;1 is essential for endosperm formation and causes embryo enlargement in rice. Planta. 231, 293–303.
Hamann, T., Benkova, E., Baurle, I., Kientz, M., and Jurgens, G. (2002). The Arabidopsis BODENLOS gene encodes an auxin response protein inhibiting MONOPTEROS-mediated embryo patterning. Genes Dev. 16, 1610–1615.
Hehenberger, E., Kradolfer, d., and Köhler, C. (2012). Endosperm cellularization defines an important developmental transition for embryo development. Development. 139, 2031–2039.
Helliwell, C.A., Chin-Atkins, A.n., Wilson, I.W., Chapple, R., dennis, E.S., and Chaudhury, A. (2001). The Arabidopsis AMP1 gene encodes a putative glutamate carboxypeptidase. Plant Cell. 13, 2115–2125.
Hirose, T., Imaizumi, n., Scofield, G.n., Furbank, R.T., and Ohsugi, R. (1997). cDNA cloning and tissue specific expression of a gene for sucrose transporter from rice (Oryza sativa L.). Plant Cell Physiol. 38, 1389–1396.
Hong, S.K., Aoki, T., Kitano, H., Satoh, H., and nagato, Y. (1995). Phenotypic diversity of 188 rice embryo mutants. Dev. Genet. 16, 298–310.
Hong, S.K., Kitano, H., Satoh, H., and nagato, Y. (1996). How is embryo size genetically regulated in rice? Development. 122, 2051–2058.
Honga, S.K., Aokia, T., Kitanob, H., Satohc, H., and nagato, Y. (1995). Temperature-sensitive mutation, embryoless 1, affects both embryo and endosperm development in rice. Plant Sci. 108, 165–172.
Imaishi, H., Matsuo, S., Swai, E., and Ohkawa, H. (2000). CYP78A1 preferentially expressed in developing inflorescences of Zea mays encoded acytochrome P450-dependent lauric acid 12-monooxy-genase. Biosci. Biotechnol. Biochem. 64, 1696–1701.
Ito, T., and Meyerowitz, E.M. (2000). Overexpression of a gene encoding a cytochrome P450, CYP78A9, induces large and seedless fruit in Arabidopsis. Plant Cell. 12, 1541–1550.
Itoh, J.I., Kitano, H., Matsuoka, M., and nagato, Y. (2000). SHOOT ORGANIZATION genes regulate shoot apical meristem organi-zation and the pattern of leaf primordium initiation in rice. Plant Cell. 12, 2161–2174.
Itoh, J.I., nonomura, K., Ikeda, K., Yamaki, S., Inukai, Y., Yamagishi, H., Kitano, H., and nagato, Y. (2005). Rice plant development: from zygote to spikelet. Plant Cell Physiol. 46, 23–47.
Jiao, Y., Wang, Y., Xue, d., Wang, J., Yan, M., Liu, G., dong, G., Zeng, d., Lu, Z., Zhu, X., et al. (2010). Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 42, 541–544.
Jofuku, K.d., Omidyar, P.K., Gee, Z., and Okamuro, J.K. (2005). Control of seed mass and seed yield by the floral homeotic gene APETALA2. Proc. Natl Acad. Sci. U S A. 102, 3117–3122.
Jurgens, G. (2001). Apical–basal pattern formation in Arabidopsis embryogenesis. EMBO J. 20, 3609–3616.
Kai, K., Hashidzume, H., Yoshimura, K., Suzuki, H., Sakurai, n, Shibata, d., and Ohta, d. (2009). Metabolomics for the charac-terization of cytochromes P450-dependent fatty acid hydroxy-lation reactions in Arabidopsis. Plant Biotechnol. 26, 175–182.
Kaneko, M., Itoh, H., Inukai, Y., Sakamoto, T., ueguchi-Tanaka, M., Ashikari, M., and Matsuoka, M. (2003). Where do gibberel-lin biosynthesis and gibberellin signaling occur in rice plants? Plant J. 35, 104–115.
Kawakatsu, T., Taramino, G., Itoh, J., Allen, J., Sato, Y., Hong, S.K., Yule, R., nagasawa, n., Kojima, M., Kusaba, M., et al. (2009). PLASTOCHRON3/GOLIATH encodes a glutamate carboxypepti-dase required for proper development in rice. Plant J. 58, 1028–1040.
Lau, S., Slane, d., Herud, O., Kong, J., and Jürgens, G. (2012). Early embryogenesis in flowering plants: setting up the basic body pattern. Annu. Rev. Plant Biol. 63, 483–506.
Li, S., Liu, L., Zhuang, X., Yu, Y., Liu, X., Cui, X., Ji, L., Pan, Z., Cao, X., Mo, B., et al. (2013). MicroRNAs inhibit the translation of target mRNAs on the endoplasmic reticulum in Arabidopsis. Cell. 153, 562–574.
Li, Y., Fan, C., Xing, Y., Jiang, Y., Luo, L., Sun, L., Shao, d., Xu, C., Li, X., Xiao, J., et al. (2011). Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat. Genet. 43, 1266–1269.
Li, Y., Zheng, L., Corke, F., Smith, C., and Bevan, M.W. (2008). Control of final seed and organ size by the DA1 gene family in Arabidopsis thaliana. Genes Dev. 22, 1331–1336.
Lukowitz, W., Roeder, A., Parmenter, d., and Somerville, C. (2004). A MAPKK kinase gene regulates extra-embryonic cell fate in Arabidopsis. Cell. 116, 109–119.
Luo, M., dennis, E.S., Berger, F., Peacock, W.J., and Chaudhury, A. (2005). MINISEED3 (MINI3), a WRKY family gene, and HAIKU2 (IKU2), a leucine-rich repeat (LRR) KINASE gene, are regulators of seed size in Arabidopsis. Proc. Natl Acad. Sci. U S A. 102, 17531–17536.
Miura, K., Ikeda, M., Matsubara, A., Song, X.J., Ito, M., Asano, K., Matsuoka, M., Kitano, H., and Ashikari, M. (2010). OsSPL14 pro-motes panicle branching and higher grain productivity in rice. Nat. Genet. 42, 545–549.
Miyoshi, K., Ahn, B.O., Kawakatsu, T., Ito, Y., Itoh, J., nagato, Y., and Kurata, n. (2004). PLASTOCHRON1, a timekeeper of leaf initiation in rice, encodes cytochrome P450. Proc. Natl Acad. Sci. U S A. 101, 875–880.
Mizutani, M., and Ohta, d. (2010). Diversification of P450 genes during land plant evolution. Annu. Rev. Plant Biol. 61, 291–315.

1960 Yang et al. • GE Controls Rice Embryo Development and Grain Yield
nagasaki, H., Itoh, J., Hayashi, K., Hibara, K., Satoh-nagasawa, n., nosaka, M., Mukouhata, M., Ashikari, M., Kitano, H., Matsuoka, M., et al. (2007). The small interfering RNA produc-tion pathway is required for shoot meristem initiation in rice. Proc. Natl Acad. Sci. U S A. 104, 14867–14871.
nagasawa, n., Hibara, K.I., Heppard, E.P., Vander Velden, K.A., Luck, S., Beatty, M., nagato, Y., and Sakai, H. (2013). GIANT EMBRYO, encoding CYP78A13, affects size balance between embryo and endosperm in rice. Plant J., 29, 10.1111/tpj.12223.
nowack, M.K., Grini, P.E., Jakoby, M.J., Lafos, M., Koncz, C., and Schnittger, A. (2006). A positive signal from the fertilization of the egg cell sets off endosperm proliferation in angiosperm embryogenesis. Nat. Genet. 38, 63–67.
Ohmori, Y., Tanaka, W., Kojima, M., Sakakibara, H., and Hirano HY. (2013). WUSCHEL-RELATED HOMEOBOX4 is involved in meris-tem maintenance and is negatively regulated by the CLE gene FCP1 in rice. Plant Cell. 25, 229–241.
Pollard, M., Beisson, F., Li, Y., and Ohlrogge, J.B. (2008). Building lipid barriers: biosynthesis of cutin and suberin. Trends Plant Sci. 13, 236–246.
Qi, P., Lin, Y.S., Song, X.J., Shen, J.B., Huang, W., Shan, J.X., Zhu, M.Z., Jiang, L., Gao, J.P., and Lin, H.X. (2012). The novel quan-titative trait locus GL3.1 controls rice grain size and yield by regulating Cyclin-T1;3. Cell Res. 22, 1666–1680.
Qian, Q., Xiong, Z.M., Min, S.K., and Zhu, L.H. (1996). The RFLP of tagging of giant embryo gene. Chinese J. Rice Sci. 10, 65–70.
Sato, Y., Hong, S.K., Tagiri, A., Kitano, H., Yamamoto, n., nagato, Y., and Matsuoka, M. (1996). A rice homeobox gene, OSH1, is expressed before organ differentiation in a specific region during early embryogenesis. Proc. Natl Acad. Sci. U S A. 93, 8117–8122.
Sazuka, T., Kamiya, n., nishimura, T., Ohmae, K., Sato, Y., Imamura, K., nagato, Y., Koshiba, T., nagamura, Y., Ashikari, M., et al. (2009). A rice tryptophan deficient dwarf mutant, tdd1, con-tains a reduced level of indole acetic acid and develops abnor-mal flowers and organless embryos. Plant J. 60, 227–241.
Schoof, H., Lenhard, M., Haecker, A., Mayer, K.F.X., Jürgens, G., and Laux, T. (2000). The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell. 100, 635–644.
Schreiber, L. (2010). Transport barriers made of cutin, suberin and associated waxes. Trends Plant Sci. 15, 546–553.
Schruff, M.C., Spielman, M., Tiwari, S., Adams, S., Fenby, n., and Scott, R.J. (2006). The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development. 133, 251–261.
Song, X.J., Huang, W., Shi, M., Zhu, M.Z., and Lin, H.X. (2007). A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat. Genet. 39, 623–630.
Sotelo-Silveira, M., Cucinotta, M., Chauvin, A.L., Chávez Montes, R.A., Colombo, L., Marsch-Martínez, n., and de Folter, S. (2013). Cytochrome P450 CYP78A9 is involved in Arabidopsis reproduc-tive development. Plant Physiol. 162, 779–799.
Tanaka, H., Onouchi, H., Kondo, M., Hara-nishimura, I., nishimura, M., Machida, C., and Machida, Y. (2001). A subtilisin-like
serine protease is required for epidermal surface formation in Arabidopsis embryos and juvenile plants. Development. 128, 4681–4689.
Tanaka, H., Watanabe, M., Sasabe, M., Hiroe, T., Tanaka, T., Tsukaya, H., Ikezaki, M., Machida, C., and Machida, Y. (2007). Novel recep-tor-like kinase ALE2 controls shoot development by specifying epidermis in Arabidopsis. Development. 134, 1643–1652.
Tanaka, H., Watanabe, M., Watanabe, d., Tanaka, T., Machida, C., and Machida, Y. (2002). ACR4, a putative receptor kinase gene of Arabidopsis thaliana, that is expressed in the outer cell lay-ers of embryos and plants, is involved in proper embryogenesis. Plant Cell Physiol. 43, 419–428.
Tanksley, S.d., and McCouch, S.R. (1997). Seed banks and molecu-lar maps: unlocking genetic potential from the wild. Science. 277, 1063–1066.
ueda, M., Zhang, Z., and Laux, T. (2011). Transcriptional activa-tion of Arabidopsis axis patterning genes WOX8/9 links zygote polarity to embryo development. Dev. Cell. 20, 264–270.
Wang, E., Wang, J., Zhu, X., Hao, W., Wang, L., Li, Q., Zhang, L., He, W., Lu, B., Lin, H., et al. (2008a). Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat. Genet. 40, 1370–1374.
Wang, J.W., Schwab, R., Czech, B., Mica, E., and Weigel, d. (2008b). Dual effects of miR156-targeted SPL genes and CYP78A5/KLUH on plastochron length and organ size in Arabidopsis thaliana. Plant Cell. 20, 1231–1243.
Wang, S., Wu, K., Yuan, Q., Liu, X., Liu, Z., Lin, X., Zeng, R., Zhu, H., dong, G., Qian, Q., et al. (2012). Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet. 44, 950–954.
Weng, J., Gu, S., Wan, X., Gao, H., Guo, T., Su, n., Lei, C., Zhang, X., Cheng, Z., Guo, X., et al. (2008). Isolation and initial charac-terization of GW5, a major QTL associated with rice grain width and weight. Cell Res. 18, 1199–1209.
Yang, W., Ren, S., Zhang, X., Gao, M., Ye, S., Qi, Y., Zheng, Y., Wang, J., Zeng, L., Li, Q., et al. (2011). BENT UPPERMOST INTERNODE1 encodes the class II formin FH5 crucial for actin organization and rice development. Plant Cell. 23, 661–680.
Yano, M. (2001). Genetic and molecular dissection of naturally occurring variation. Curr. Opin. Plant Biol. 4, 130–135.
Yephremov, A., Wisman, E., Huijser, P., Huijser, C., Wellesen, K., and Saedler, H. (1999). Characterization of the FIDDLEHEAD gene of Arabidopsis reveals a link between adhesion response and cell differentiation in the epidermis. Plant Cell. 11, 2187–2201.
Zhang, L.L., Shu, X.L., Wang, X.Y., Lu, H.J., Shu, Q.Y., and Wu, d.X. (2007). Characterization of indica-type giant embryo mutant rice enriched with nutritional components. Cereal Res. Commun. 35, 1459–1468.
Zhou, S.R., Yin, L.L., and Xue, H.W. (2013). Functional genomics based understanding of rice endosperm. Curr. Opin. Plant Biol. 16, 236–246.
Zondlo, S.C., and Irish, V.F. (1999). CYP78A5 encodes a cytochrome P450 that marks the shoot apical meristem boundary in Arabidopsis. Plant J. 19, 259–268.




























