Embed Size (px)
Citation preview

www.elsevier.com/locate/jembe
Journal of Experimental Marine Biolo
Contrasting contributions to inorganic nutrient recycling by the
co-occurring jellyfishes, Catostylus mosaicus and Phyllorhiza
punctata (Scyphozoa, Rhizostomeae)
K.A. Pitta,*, K. Koopb, D. Rissikc
aCentre for Aquatic Processes and Pollution and School of Environmental and Applied Sciences, Griffith University, PMB 50,
Gold Coast Mail Centre QLD 9726, AustraliabEnvironment and Conservation Science, NSW Department of Environment and Conservation, PO Box A290,
Sydney South NSW 1232, AustraliacDepartment of Infrastructure, Planning and Natural Resources, 33 Bridge Street, Sydney NSW 2000, Australia
Received 14 May 2004; received in revised form 1 August 2004; accepted 1 September 2004
Abstract
The rhizostome jellyfishes, Catostylus mosaicus and Phyllorhiza punctata abound in estuaries in New South Wales,
Australia. P. punctata contains symbiotic zooxanthellae but C. mosaicus contains few or no zooxanthellae. Our experiment
measured the rates at which NH3, PO4 and NOx were taken up or excreted by each species and in two controls: a bwater onlyQcontrol and a bmucusQ control. Rates of uptake or excretion were measured as changes in the nutrient concentration of the water
in the containers housing the animals over periods of 6 h. Experiments were repeated twice during the day and twice at night,
under both ambient and enriched nutrient conditions. Under ambient nutrient conditions, the flux of NH3 in the P. punctata
treatment did not differ from the controls but under enriched conditions P. punctata excreted NH3 during the night (49 Ag kg�1
WW (wet weight) h�1) and took up NH3 during the day (123 Ag kg�1 WW h�1). In contrast, C. mosaicus excreted NH3 at a rate
of 1555 Ag kg�1 WW h�1 during the day and 1004 Ag kg�1 WW h�1 during the night under both enriched and ambient nutrient
conditions. P. punctata neither took up nor excreted PO4 but C. mosaicus excreted PO4 at a faster rate during the day than night
(173 Ag kg�1 WW h�1 cf. 104 Ag kg�1 WW h�1). Both C. mosaicus and P. punctata excreted NOx and, although the rate of
excretion for P. punctata varied between the two experiments conducted during the day, the rate of excretion was consistently
greater than for C. mosaicus (52 and 80 Ag kg�1 WW h�1 cf. 26 Ag kg�1 WW h�1). Tubs containing P. punctata had a much
greater concentration of dissolved oxygen at the end of the experiments conducted during the day (152% saturation) than night
(60% saturation) but tubs containing C. mosaicus had a greater dissolved oxygen concentration during the night (47%
saturation) than day (39%). Overall, C. mosaicus appears to recycle more inorganic nutrients to estuaries than P. punctata.
Calculations of the importance of inorganic nitrogen excreted by this species during times of peak biomass in Lake Illawarra
suggest that it can meet about 8% of the phytoplankton primary production requirements of N and that its inorganic N excretion
0022-0981/$ - s
doi:10.1016/j.jem
* Correspon
E-mail addr
gy and Ecology 315 (2005) 71–86
ee front matter D 2004 Elsevier B.V. All rights reserved.
be.2004.09.007
ding author. Tel.: +61 7 5552 8324; fax: +61 7 5552 8064.
ess: [email protected] (K.A. Pitt).

K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–8672
rate is about 11% of measured inorganic ammonia fluxes from sediments in that system. Since the biomass of C. mosaicus often
exceeds several thousand tonnes, the contribution of inorganic nutrients by this species is substantial.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Gelatinous zooplankton; Excretion; Symbioses; Zooxanthellae
1. Introduction
Excretion of inorganic nutrients by consumers is a
major source of recycled nutrients for primary
producers (e.g., Uthicke, 2001; Priddle et al., 2003;
Reissig et al., 2003). Gelatinous zooplankters are
important consumers in pelagic systems where they
frequently represent a large fraction of the biomass
(e.g., Pages et al., 1996, 2001). Because these
consumers occur in such large biomasses, gelatinous
zooplankters are likely to be one of the largest
contributors of recycled nutrients in these areas.
Indeed, in the North Atlantic, excretion by gelatinous
zooplankters has been estimated to supply 39–63% of
the nitrogen required to sustain phytoplankton pro-
duction (Biggs, 1977).
Scyphozoan jellyfish are the most abundant gelat-
inous zooplankters in estuaries in eastern Australia.
Population blooms occur periodically, with abundan-
ces increasing up to 30-fold over periods of just weeks
to months (Pitt and Kingsford, 2000). During such
blooms, the biomass of jellyfish can measure thou-
sands of tonnes (Pitt and Kingsford, 2003) and the
medusae are likely to recycle enormous quantities of
inorganic nutrients to the water column. Scyphozoan
jellyfish differ from most gelatinous zooplankters in
that some species contain symbiotic dinoflagellates
(zooxanthellae) within their tissues (Balderston and
Claus, 1969). Jellyfish with symbiotic zooxanthellae
are likely to recycle fewer nutrients to the water
column since their symbionts may utilise the excre-
tory products of their host (Wilkerson and Kremer,
1992). Jellyfish with and without zooxanthellae (or
with different densities of zooxanthellae) are, there-
fore, likely to have contrasting roles in nutrient
recycling (Cates and McLaughlin, 1976; Muscatine
and Marian, 1982).
Catostylus mosaicus and Phyllorhiza punctata are
large, rhizostome medusae that occur abundantly in
estuaries and coastal lagoons in eastern Australia (Pitt
and Kingsford, 2000). Recent molecular studies show
that both species contain zooxanthellae (Moore,
personal communication), but a recent flurometric
study of the two species failed to detect any photo-
synthesis by C. mosaicus (Hill, personal communica-
tion). Hence, these species are likely to make
contrasting contributions to nutrient recycling in
estuaries.
The aim of our study was to compare rates of
uptake or excretion of the inorganic nutrients ammo-
nia (NH3), nitrate and nitrite (NOx) and phosphate
(PO4) by C. mosaicus and P. punctata. Specifically
we predicted that C. mosaicus would excrete inor-
ganic nutrients but P. punctata would actively deplete
inorganic nutrients from the surrounding water. Since
photosynthesis occurs only during daylight, rates of
nutrient uptake may vary diurnally. Hence we also
tested the hypothesis that the rate of nutrient uptake by
P. punctata would be faster during the day than during
the night while the rate of nutrient excretion by C.
mosaicus would be similar during both day and night.
2. Methods
The study was conducted during February 2003 at
Smiths Lake, an intermittently open coastal lagoon in
New South Wales, Australia. Like most coastal
lagoons in the region, Smiths Lake often contains
dense populations of C. mosaicus and P. punctata
(Pitt, personal observation).
2.1. Experimental treatments
Experiments were repeated twice during the day
(19 and 22/2/2003) and twice at night (18 and 20/2/
2003) to compare rates of nutrient flux when medusae
were and were not photosynthesising. Rates of
nutrient flux were measured for C. mosaicus, P.
punctata and two controls. The first control consisted

K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–86 73
of filtered estuarine water and was used to control for
possible variation in nutrient concentrations in the
absence of jellyfish. The second control was termed
the bmucusQ control. Observations showed that, unlikeP. punctata, C. mosaicus produces strings of mucus.
Decomposition of the mucus could potentially con-
tribute inorganic nutrients to the treatment, independ-
ent of excretion directly from the animal. To control
for the differing rates of mucus production between
species, mucus was collected by placing individual C.
mosiacus in tubs of filtered estuarine water. C.
mosaicus were confined for 1 h, after which the
medusae were discarded but the strings of mucus
retained.
2.2. Preparation of estuarine water
Estuarine water was collected approximately 500
m offshore and stored in clean, plastic tubs prior to
filtering. Water was pre-filtered through 10 and 1 Amnose-bag filters before being vacuum filtered through
glass fibre filters and 0.2 Am membrane filters. Forty
litres of filtered estuarine water was transferred to
each of sixteen 60-L rectangular tubs made of clear
plastic. Since it took approximately 16 h to filter all
the water required for each experiment, a lid was
placed on each tub after it was filled to prevent
evaporation or contamination by rain or other
sources.
2.3. Sampling of water for nutrient analyses
Samples of water for nutrient analyses were
extracted from experimental tubs using a syringe.
Duplicate samples of 25 ml were filtered (0.45 Am)
and stored in clean plastic vials. Samples were
initially stored on ice and then transferred to a clean
freezer at the completion of each experiment. Nutrient
concentrations were determined by flow injection
analysis, ammonia by the automated phenate method,
phosphate by the automated ascorbic acid reduction
method and NOx (nitrite and nitrate) by the automated
cadmium reduction method at the New South Wales
Department of Environment and Conservation Water
Science Laboratories. Nutrient concentrations were
measured within two weeks of the experiment being
completed.
2.4. Experimental procedure
Duplicate samples of water were extracted from
each tub prior to starting each experiment to ensure
that no tubs were contaminated (referred to as Time
0). To increase the ability to detect the active uptake of
nutrients, ammonia (NH3), nitrate and nitrite (NOx)
and phosphate (PO4) were added at random to 8 of the
16 tubs to elevate them to concentrations of approx-
imately 58, 30 and 35 Ag L�1, respectively. These
concentrations reflected the upper limit of the nutrient
concentrations found naturally in the lake (Coade,
personal communication). Duplicate samples of water
were again extracted from all tubs immediately after
the addition of nutrients (Time 1). Two replicates of
each of the four treatments (i.e., C. mosaicus, P.
punctata and the Mucus and Water controls) were
assigned randomly to two tubs containing nutrients
and two tubs to which no nutrients had been added.
Jellyfish were collected from the field using a bucket
immediately prior to each experiment and care was
taken to select animals that were undamaged, were
actively swimming and were of similar sizes. The
average bell diameter (BD) of medusae used in the
experiment was identical for both C. mosaicus and P.
punctata and was 209 mm (S.E.=3.8 C. mosaicus;
S.E.=3.5 P. punctata). The mean weight, however,
differed between species: 2161 g (F103 S.E.) for C.
mosaicus and 1623 g (F162 S.E.) for P. punctata.
One C. mosaicus or one P. punctata was added to
each tub, as appropriate, immediately after the
addition of the nutrients. The mucus controls were
set-up slightly differently because the mucus was
collected by confining one C. mosaicus in each of
four tubs containing filtered seawater for 1 h prior to
the start of the experiment. After 1 h, the jellyfish
were removed and nutrients were added to two of the
tubs at the same time that nutrients were added to the
other treatments. Duplicate samples of water were
extracted from each tub every hour for 6 h (Times 2–
7). The bell diameter and wet weight of each jellyfish
was measured at the end of the experiment to
minimise stress to the animals caused by handling.
During the first day and first night experiments, the
tubs were placed in the water parallel to the shore at a
depth of approximately 20 cm so that the temperature
of the water in the tubs was the same as that in the
lake. During the second night experiment, tubs were
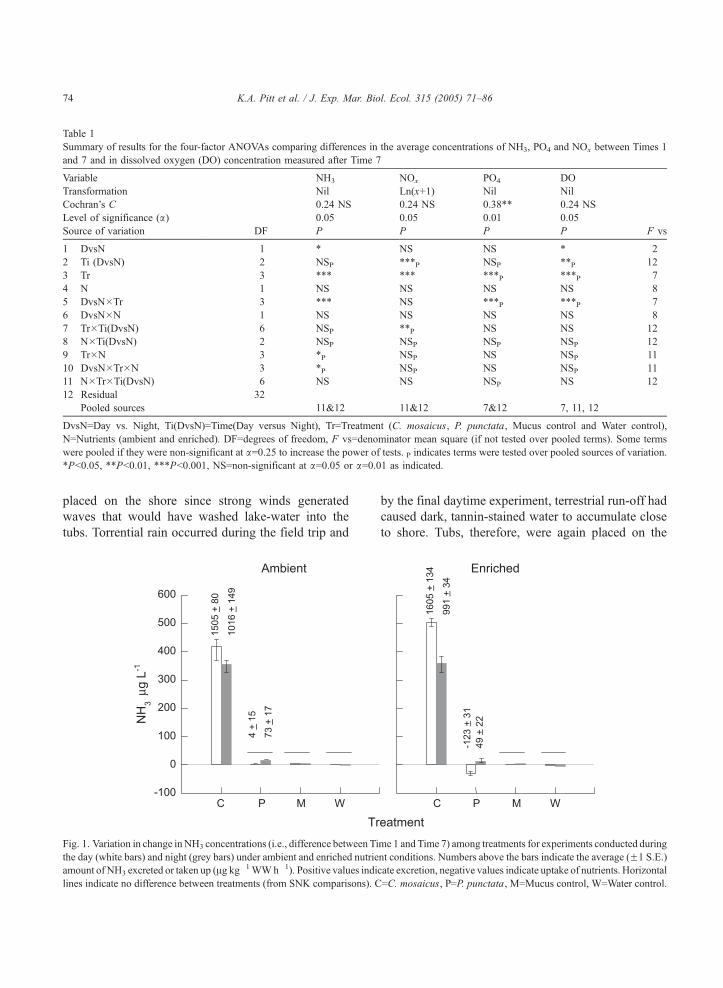
Table 1
Summary of results for the four-factor ANOVAs comparing differences in the average concentrations of NH3, PO4 and NOx between Times 1
and 7 and in dissolved oxygen (DO) concentration measured after Time 7
Variable NH3 NOx PO4 DO
Transformation Nil Ln(x+1) Nil Nil
Cochran’s C 0.24 NS 0.24 NS 0.38** 0.24 NS
Level of significance (a) 0.05 0.05 0.01 0.05
Source of variation DF P P P P F vs
1 DvsN 1 * NS NS * 2
2 Ti (DvsN) 2 NSP ***P NSP **P 12
3 Tr 3 *** *** ***P ***P 7
4 N 1 NS NS NS NS 8
5 DvsN�Tr 3 *** NS ***P ***P 7
6 DvsN�N 1 NS NS NS NS 8
7 Tr�Ti(DvsN) 6 NSP **P NS NS 12
8 N�Ti(DvsN) 2 NSP NSP NSP NSP 12
9 Tr�N 3 *P NSP NS NSP 11
10 DvsN�Tr�N 3 *P NSP NS NSP 11
11 N�Tr�Ti(DvsN) 6 NS NS NSP NS 12
12 Residual 32
Pooled sources 11&12 11&12 7&12 7, 11, 12
DvsN=Day vs. Night, Ti(DvsN)=Time(Day versus Night), Tr=Treatment (C. mosaicus, P. punctata, Mucus control and Water control),
N=Nutrients (ambient and enriched). DF=degrees of freedom, F vs=denominator mean square (if not tested over pooled terms). Some terms
were pooled if they were non-significant at a=0.25 to increase the power of tests. P indicates terms were tested over pooled sources of variation.
*Pb0.05, **Pb0.01, ***Pb0.001, NS=non-significant at a=0.05 or a=0.01 as indicated.
K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–8674
placed on the shore since strong winds generated
waves that would have washed lake-water into the
tubs. Torrential rain occurred during the field trip and
Fig. 1. Variation in change in NH3 concentrations (i.e., difference between Ti
the day (white bars) and night (grey bars) under ambient and enriched nutrie
amount of NH3 excreted or taken up (Ag kg�1WWh�1). Positive values indi
lines indicate no difference between treatments (from SNK comparisons). C
by the final daytime experiment, terrestrial run-off had
caused dark, tannin-stained water to accumulate close
to shore. Tubs, therefore, were again placed on the
me 1 and Time 7) among treatments for experiments conducted during
nt conditions. Numbers above the bars indicate the average (F1 S.E.)
cate excretion, negative values indicate uptake of nutrients. Horizontal
=C. mosaicus, P=P. punctata, M=Mucus control, W=Water control.
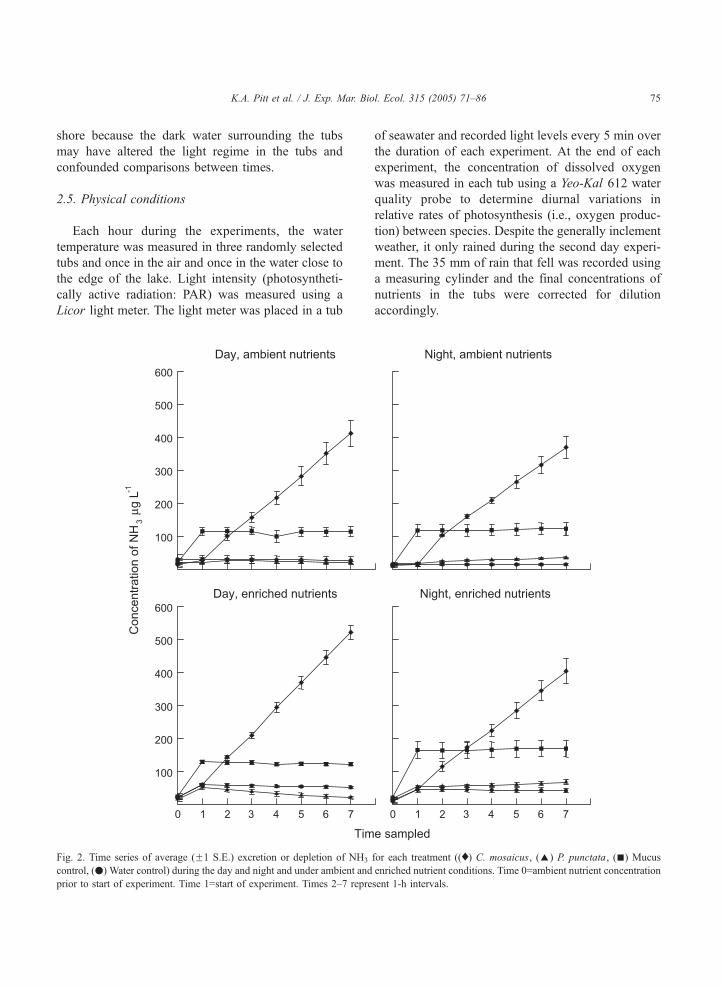
K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–86 75
shore because the dark water surrounding the tubs
may have altered the light regime in the tubs and
confounded comparisons between times.
2.5. Physical conditions
Each hour during the experiments, the water
temperature was measured in three randomly selected
tubs and once in the air and once in the water close to
the edge of the lake. Light intensity (photosyntheti-
cally active radiation: PAR) was measured using a
Licor light meter. The light meter was placed in a tub
Fig. 2. Time series of average (F1 S.E.) excretion or depletion of NH3
control, (.) Water control) during the day and night and under ambient and
prior to start of experiment. Time 1=start of experiment. Times 2–7 repre
of seawater and recorded light levels every 5 min over
the duration of each experiment. At the end of each
experiment, the concentration of dissolved oxygen
was measured in each tub using a Yeo-Kal 612 water
quality probe to determine diurnal variations in
relative rates of photosynthesis (i.e., oxygen produc-
tion) between species. Despite the generally inclement
weather, it only rained during the second day experi-
ment. The 35 mm of rain that fell was recorded using
a measuring cylinder and the final concentrations of
nutrients in the tubs were corrected for dilution
accordingly.
for each treatment ((x) C. mosaicus, (E) P. punctata, (n) Mucus
enriched nutrient conditions. Time 0=ambient nutrient concentration
sent 1-h intervals.
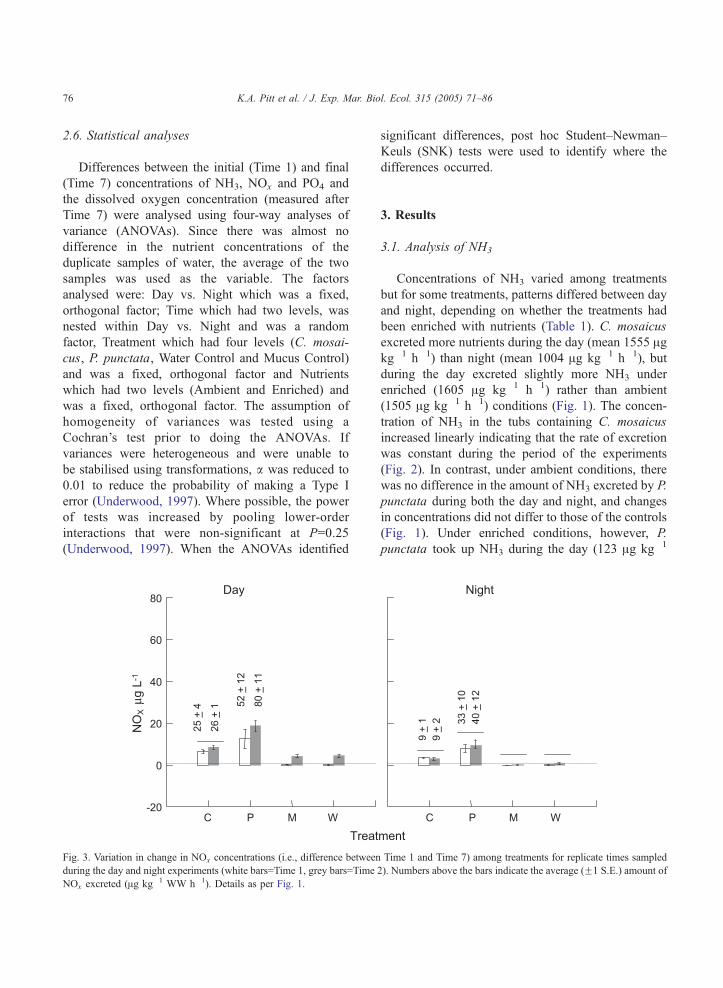
K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–8676
2.6. Statistical analyses
Differences between the initial (Time 1) and final
(Time 7) concentrations of NH3, NOx and PO4 and
the dissolved oxygen concentration (measured after
Time 7) were analysed using four-way analyses of
variance (ANOVAs). Since there was almost no
difference in the nutrient concentrations of the
duplicate samples of water, the average of the two
samples was used as the variable. The factors
analysed were: Day vs. Night which was a fixed,
orthogonal factor; Time which had two levels, was
nested within Day vs. Night and was a random
factor, Treatment which had four levels (C. mosai-
cus, P. punctata, Water Control and Mucus Control)
and was a fixed, orthogonal factor and Nutrients
which had two levels (Ambient and Enriched) and
was a fixed, orthogonal factor. The assumption of
homogeneity of variances was tested using a
Cochran’s test prior to doing the ANOVAs. If
variances were heterogeneous and were unable to
be stabilised using transformations, a was reduced to
0.01 to reduce the probability of making a Type I
error (Underwood, 1997). Where possible, the power
of tests was increased by pooling lower-order
interactions that were non-significant at P=0.25
(Underwood, 1997). When the ANOVAs identified
Fig. 3. Variation in change in NOx concentrations (i.e., difference between
during the day and night experiments (white bars=Time 1, grey bars=Time
NOx excreted (Ag kg�1 WW h�1). Details as per Fig. 1.
significant differences, post hoc Student–Newman–
Keuls (SNK) tests were used to identify where the
differences occurred.
3. Results
3.1. Analysis of NH3
Concentrations of NH3 varied among treatments
but for some treatments, patterns differed between day
and night, depending on whether the treatments had
been enriched with nutrients (Table 1). C. mosaicus
excreted more nutrients during the day (mean 1555 Agkg�1 h�1) than night (mean 1004 Ag kg�1 h�1), but
during the day excreted slightly more NH3 under
enriched (1605 Ag kg�1 h�1) rather than ambient
(1505 Ag kg�1 h�1) conditions (Fig. 1). The concen-
tration of NH3 in the tubs containing C. mosaicus
increased linearly indicating that the rate of excretion
was constant during the period of the experiments
(Fig. 2). In contrast, under ambient conditions, there
was no difference in the amount of NH3 excreted by P.
punctata during both the day and night, and changes
in concentrations did not differ to those of the controls
(Fig. 1). Under enriched conditions, however, P.
punctata took up NH3 during the day (123 Ag kg�1
Time 1 and Time 7) among treatments for replicate times sampled
2). Numbers above the bars indicate the average (F1 S.E.) amount of
K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–86 77
h�1) and excreted it during the night (49 Ag kg�1 h�1)
(Fig. 1). Although P. punctata did influence the flux
of nutrients under enriched conditions, the amount of
NH3 taken up or excreted by P. punctata was
substantially less than that excreted by C. mosaicus.
Concentrations of NH3 did not vary diurnally or under
ambient or enriched conditions for either the Mucus or
Water controls (Fig. 1).
3.2. NOx
The flux of NOx was much smaller than that of
either NH3 or PO4 (Fig. 3). Concentrations of NOx
Fig. 4. Time series of average (F1 S.E.) excretion or depletion of NOx f
enriched nutrient conditions. Details as per Fig. 2.
varied among treatments but for P. punctata and the
two controls, the concentration of NOx also varied
between the two daytime experiments (Table 1), with
concentrations being greater during the second day-
time experiment (Fig. 3). Rates of excretion of NOx
by C. mosaicus, however, were consistent between
times. During both daytime experiments, the rate of
excretion by C. mosaicus (average 25.5 Ag kg�1 h�1)
was less than that of P. punctata (52 and 80 Ag kg�1
h�1; Fig. 3). Rates of excretion of NOx by P. punctata
were mostly linear under ambient nutrient conditions
indicating a constant rate of excretion, but under
enriched conditions, the rate of uptake appeared to
or each treatment during the day and night and under ambient and
K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–8678
slow or stop after Time 5 during the day, but
seemed to accelerate between Times 6 and 7 at night
(Fig. 4).
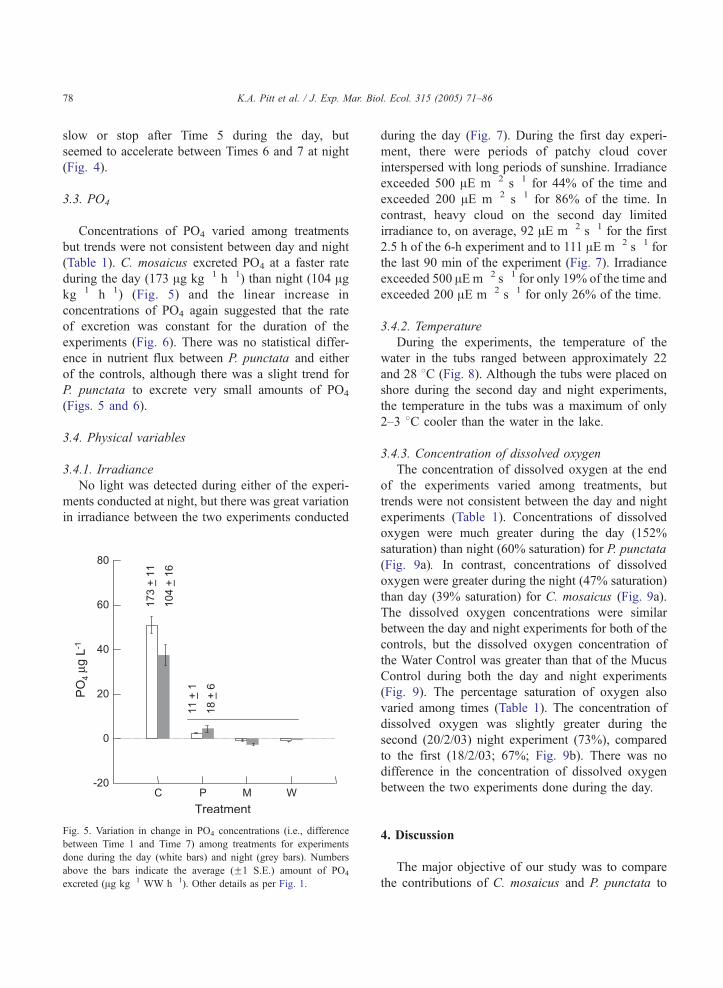
3.3. PO4
Concentrations of PO4 varied among treatments
but trends were not consistent between day and night
(Table 1). C. mosaicus excreted PO4 at a faster rate
during the day (173 Ag kg�1 h�1) than night (104 Agkg�1 h�1) (Fig. 5) and the linear increase in
concentrations of PO4 again suggested that the rate
of excretion was constant for the duration of the
experiments (Fig. 6). There was no statistical differ-
ence in nutrient flux between P. punctata and either
of the controls, although there was a slight trend for
P. punctata to excrete very small amounts of PO4
(Figs. 5 and 6).
3.4. Physical variables
3.4.1. Irradiance
No light was detected during either of the experi-
ments conducted at night, but there was great variation
in irradiance between the two experiments conducted
Fig. 5. Variation in change in PO4 concentrations (i.e., difference
between Time 1 and Time 7) among treatments for experiments
done during the day (white bars) and night (grey bars). Numbers
above the bars indicate the average (F1 S.E.) amount of PO4
excreted (Ag kg�1 WW h�1). Other details as per Fig. 1.
during the day (Fig. 7). During the first day experi-
ment, there were periods of patchy cloud cover
interspersed with long periods of sunshine. Irradiance
exceeded 500 AE m�2 s�1 for 44% of the time and
exceeded 200 AE m�2 s�1 for 86% of the time. In
contrast, heavy cloud on the second day limited
irradiance to, on average, 92 AE m�2 s�1 for the first
2.5 h of the 6-h experiment and to 111 AE m�2 s�1 for
the last 90 min of the experiment (Fig. 7). Irradiance
exceeded 500 AEm�2 s�1 for only 19% of the time and
exceeded 200 AE m�2 s�1 for only 26% of the time.
3.4.2. Temperature
During the experiments, the temperature of the
water in the tubs ranged between approximately 22
and 28 8C (Fig. 8). Although the tubs were placed on
shore during the second day and night experiments,
the temperature in the tubs was a maximum of only
2–3 8C cooler than the water in the lake.
3.4.3. Concentration of dissolved oxygen
The concentration of dissolved oxygen at the end
of the experiments varied among treatments, but
trends were not consistent between the day and night
experiments (Table 1). Concentrations of dissolved
oxygen were much greater during the day (152%
saturation) than night (60% saturation) for P. punctata
(Fig. 9a). In contrast, concentrations of dissolved
oxygen were greater during the night (47% saturation)
than day (39% saturation) for C. mosaicus (Fig. 9a).
The dissolved oxygen concentrations were similar
between the day and night experiments for both of the
controls, but the dissolved oxygen concentration of
the Water Control was greater than that of the Mucus
Control during both the day and night experiments
(Fig. 9). The percentage saturation of oxygen also
varied among times (Table 1). The concentration of
dissolved oxygen was slightly greater during the
second (20/2/03) night experiment (73%), compared
to the first (18/2/03; 67%; Fig. 9b). There was no
difference in the concentration of dissolved oxygen
between the two experiments done during the day.
4. Discussion
The major objective of our study was to compare
the contributions of C. mosaicus and P. punctata to
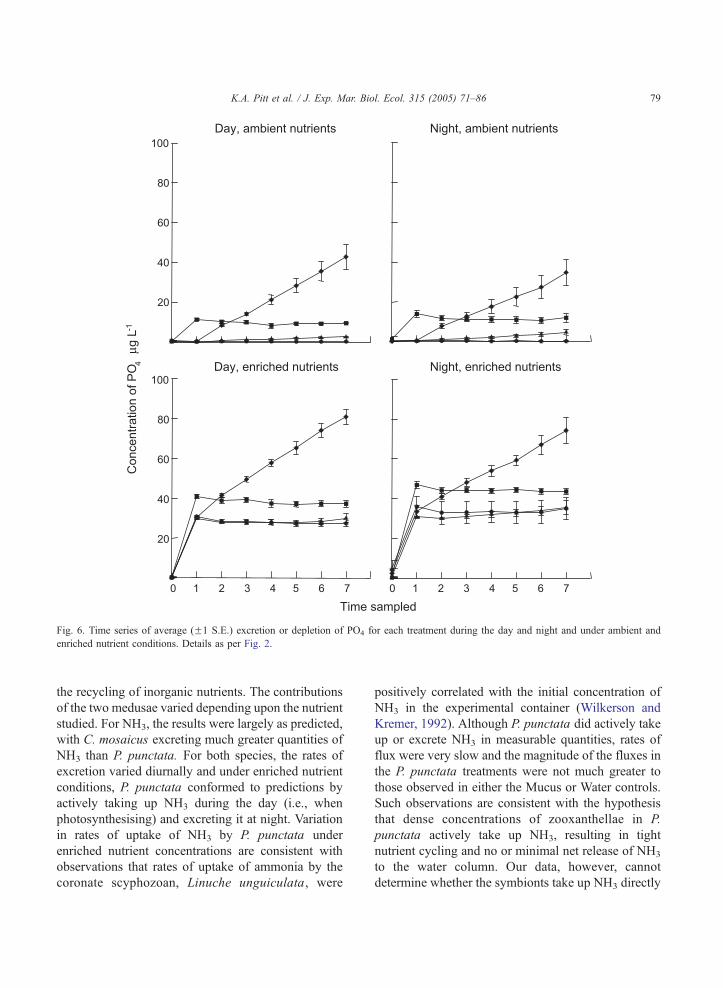
Fig. 6. Time series of average (F1 S.E.) excretion or depletion of PO4 for each treatment during the day and night and under ambient and
enriched nutrient conditions. Details as per Fig. 2.
K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–86 79
the recycling of inorganic nutrients. The contributions
of the two medusae varied depending upon the nutrient
studied. For NH3, the results were largely as predicted,
with C. mosaicus excreting much greater quantities of
NH3 than P. punctata. For both species, the rates of
excretion varied diurnally and under enriched nutrient
conditions, P. punctata conformed to predictions by
actively taking up NH3 during the day (i.e., when
photosynthesising) and excreting it at night. Variation
in rates of uptake of NH3 by P. punctata under
enriched nutrient concentrations are consistent with
observations that rates of uptake of ammonia by the
coronate scyphozoan, Linuche unguiculata, were
positively correlated with the initial concentration of
NH3 in the experimental container (Wilkerson and
Kremer, 1992). Although P. punctata did actively take
up or excrete NH3 in measurable quantities, rates of
flux were very slow and the magnitude of the fluxes in
the P. punctata treatments were not much greater to
those observed in either the Mucus or Water controls.
Such observations are consistent with the hypothesis
that dense concentrations of zooxanthellae in P.
punctata actively take up NH3, resulting in tight
nutrient cycling and no or minimal net release of NH3
to the water column. Our data, however, cannot
determine whether the symbionts take up NH3 directly
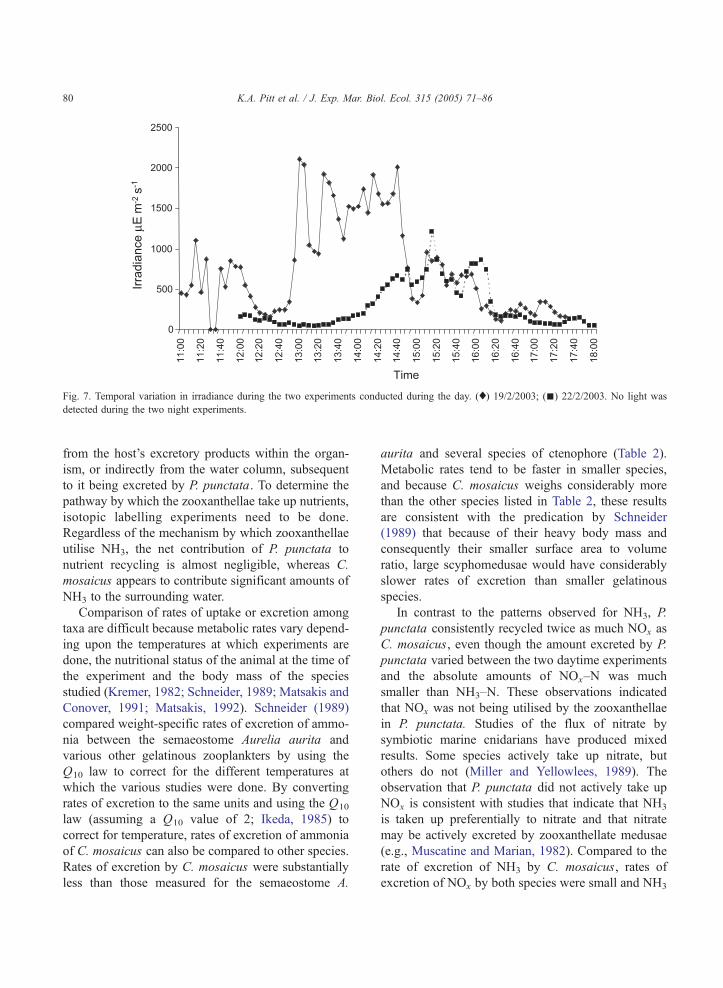
Fig. 7. Temporal variation in irradiance during the two experiments conducted during the day. (x) 19/2/2003; (n) 22/2/2003. No light was
detected during the two night experiments.
K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–8680
from the host’s excretory products within the organ-
ism, or indirectly from the water column, subsequent
to it being excreted by P. punctata. To determine the
pathway by which the zooxanthellae take up nutrients,
isotopic labelling experiments need to be done.
Regardless of the mechanism by which zooxanthellae
utilise NH3, the net contribution of P. punctata to
nutrient recycling is almost negligible, whereas C.
mosaicus appears to contribute significant amounts of
NH3 to the surrounding water.
Comparison of rates of uptake or excretion among
taxa are difficult because metabolic rates vary depend-
ing upon the temperatures at which experiments are
done, the nutritional status of the animal at the time of
the experiment and the body mass of the species
studied (Kremer, 1982; Schneider, 1989; Matsakis and
Conover, 1991; Matsakis, 1992). Schneider (1989)
compared weight-specific rates of excretion of ammo-
nia between the semaeostome Aurelia aurita and
various other gelatinous zooplankters by using the
Q10 law to correct for the different temperatures at
which the various studies were done. By converting
rates of excretion to the same units and using the Q10
law (assuming a Q10 value of 2; Ikeda, 1985) to
correct for temperature, rates of excretion of ammonia
of C. mosaicus can also be compared to other species.
Rates of excretion by C. mosaicus were substantially
less than those measured for the semaeostome A.
aurita and several species of ctenophore (Table 2).
Metabolic rates tend to be faster in smaller species,
and because C. mosaicus weighs considerably more
than the other species listed in Table 2, these results
are consistent with the predication by Schneider
(1989) that because of their heavy body mass and
consequently their smaller surface area to volume
ratio, large scyphomedusae would have considerably
slower rates of excretion than smaller gelatinous
species.
In contrast to the patterns observed for NH3, P.
punctata consistently recycled twice as much NOx as
C. mosaicus, even though the amount excreted by P.
punctata varied between the two daytime experiments
and the absolute amounts of NOx–N was much
smaller than NH3–N. These observations indicated
that NOx was not being utilised by the zooxanthellae
in P. punctata. Studies of the flux of nitrate by
symbiotic marine cnidarians have produced mixed
results. Some species actively take up nitrate, but
others do not (Miller and Yellowlees, 1989). The
observation that P. punctata did not actively take up
NOx is consistent with studies that indicate that NH3
is taken up preferentially to nitrate and that nitrate
may be actively excreted by zooxanthellate medusae
(e.g., Muscatine and Marian, 1982). Compared to the
rate of excretion of NH3 by C. mosaicus, rates of
excretion of NOx by both species were small and NH3
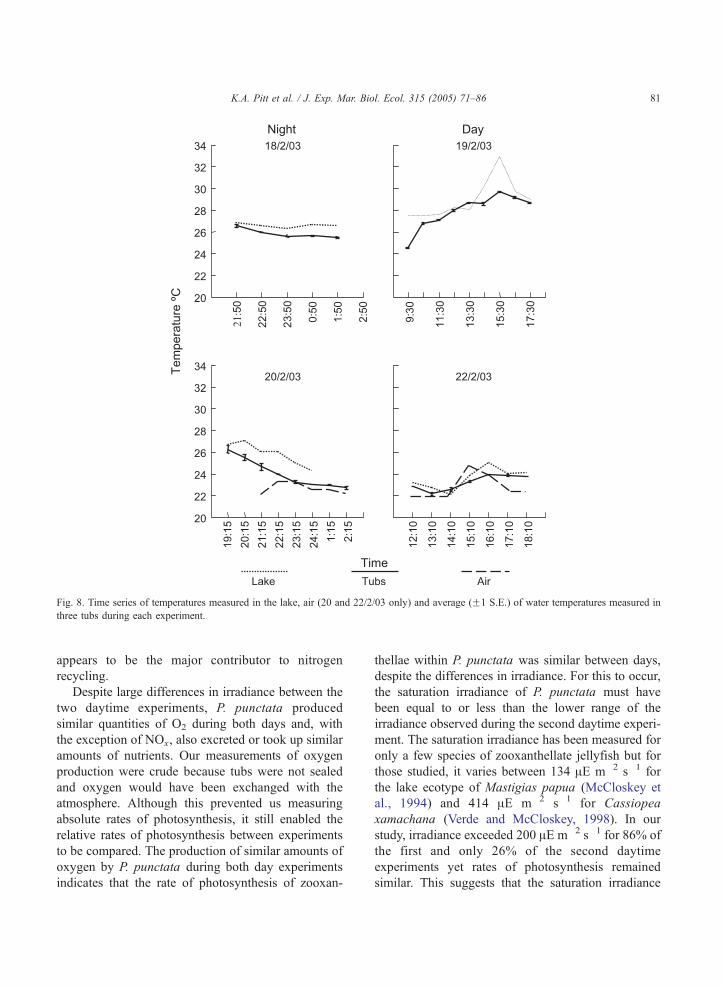
Fig. 8. Time series of temperatures measured in the lake, air (20 and 22/2/03 only) and average (F1 S.E.) of water temperatures measured in
three tubs during each experiment.
K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–86 81
appears to be the major contributor to nitrogen
recycling.
Despite large differences in irradiance between the
two daytime experiments, P. punctata produced
similar quantities of O2 during both days and, with
the exception of NOx, also excreted or took up similar
amounts of nutrients. Our measurements of oxygen
production were crude because tubs were not sealed
and oxygen would have been exchanged with the
atmosphere. Although this prevented us measuring
absolute rates of photosynthesis, it still enabled the
relative rates of photosynthesis between experiments
to be compared. The production of similar amounts of
oxygen by P. punctata during both day experiments
indicates that the rate of photosynthesis of zooxan-
thellae within P. punctata was similar between days,
despite the differences in irradiance. For this to occur,
the saturation irradiance of P. punctata must have
been equal to or less than the lower range of the
irradiance observed during the second daytime experi-
ment. The saturation irradiance has been measured for
only a few species of zooxanthellate jellyfish but for
those studied, it varies between 134 AE m�2 s�1 for
the lake ecotype of Mastigias papua (McCloskey et
al., 1994) and 414 AE m�2 s�1 for Cassiopea
xamachana (Verde and McCloskey, 1998). In our
study, irradiance exceeded 200 AE m�2 s�1 for 86% of
the first and only 26% of the second daytime
experiments yet rates of photosynthesis remained
similar. This suggests that the saturation irradiance
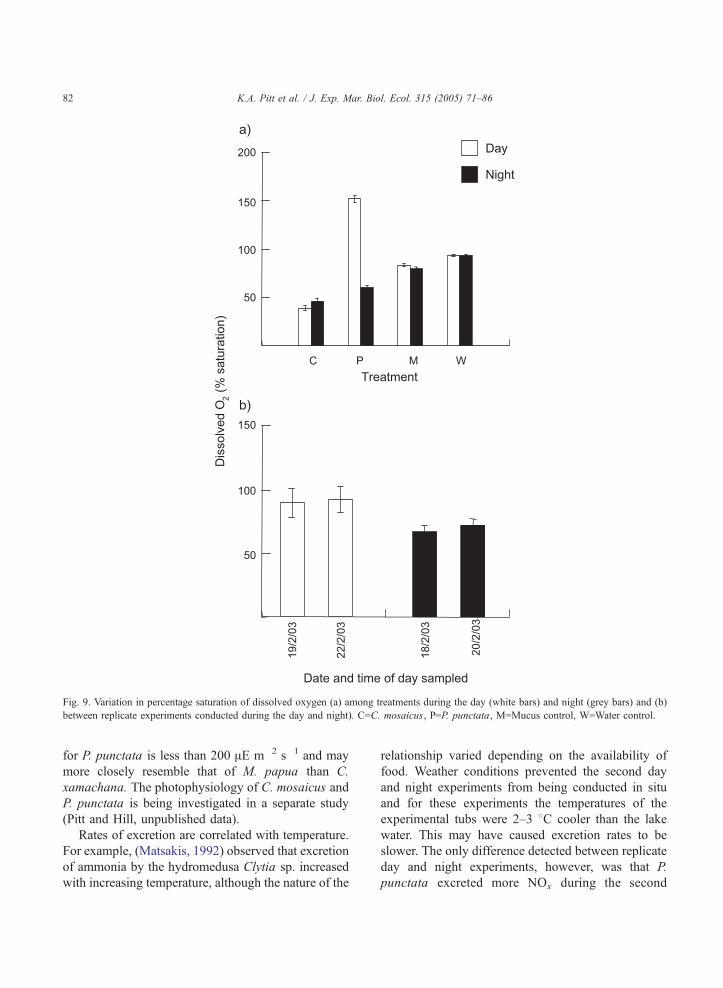
Fig. 9. Variation in percentage saturation of dissolved oxygen (a) among treatments during the day (white bars) and night (grey bars) and (b)
between replicate experiments conducted during the day and night). C=C. mosaicus, P=P. punctata, M=Mucus control, W=Water control.
K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–8682
for P. punctata is less than 200 AE m�2 s�1 and may
more closely resemble that of M. papua than C.
xamachana. The photophysiology of C. mosaicus and
P. punctata is being investigated in a separate study
(Pitt and Hill, unpublished data).
Rates of excretion are correlated with temperature.
For example, (Matsakis, 1992) observed that excretion
of ammonia by the hydromedusa Clytia sp. increased
with increasing temperature, although the nature of the
relationship varied depending on the availability of
food. Weather conditions prevented the second day
and night experiments from being conducted in situ
and for these experiments the temperatures of the
experimental tubs were 2–3 8C cooler than the lake
water. This may have caused excretion rates to be
slower. The only difference detected between replicate
day and night experiments, however, was that P.
punctata excreted more NOx during the second
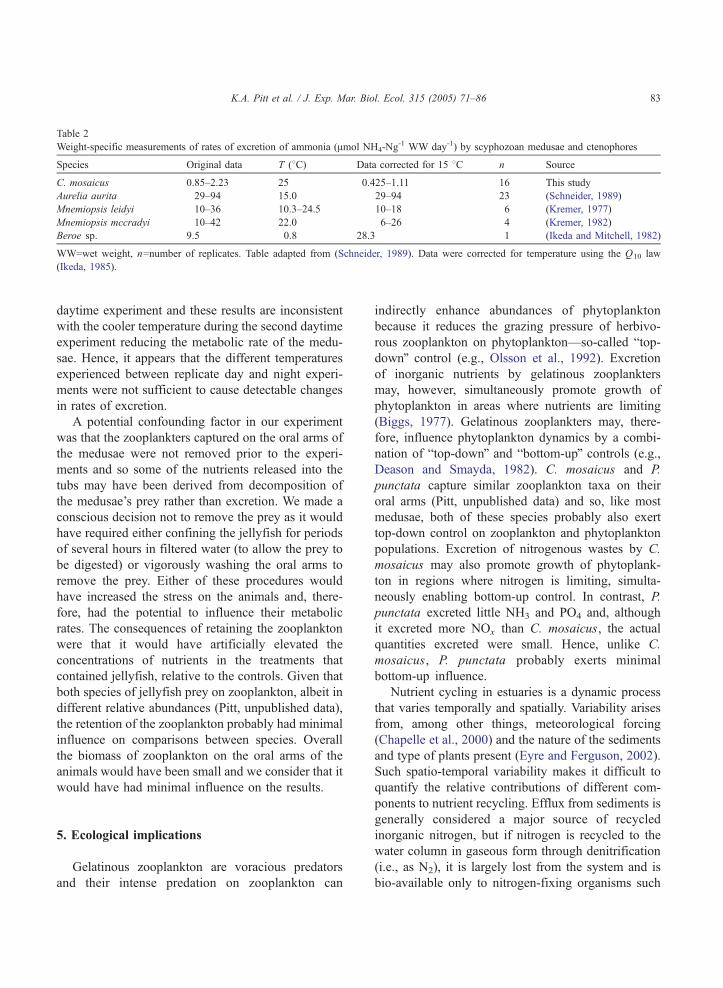
Table 2
Weight-specific measurements of rates of excretion of ammonia (Amol NH4-Ng-1 WW day-1) by scyphozoan medusae and ctenophores
Species Original data T (8C) Data corrected for 15 8C n Source
C. mosaicus 0.85–2.23 25 0.425–1.11 16 This study
Aurelia aurita 29–94 15.0 29–94 23 (Schneider, 1989)
Mnemiopsis leidyi 10–36 10.3–24.5 10–18 6 (Kremer, 1977)
Mnemiopsis mccradyi 10–42 22.0 6–26 4 (Kremer, 1982)
Beroe sp. 9.5 �0.8 28.3 1 (Ikeda and Mitchell, 1982)
WW=wet weight, n=number of replicates. Table adapted from (Schneider, 1989). Data were corrected for temperature using the Q10 law
(Ikeda, 1985).
K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–86 83
daytime experiment and these results are inconsistent
with the cooler temperature during the second daytime
experiment reducing the metabolic rate of the medu-
sae. Hence, it appears that the different temperatures
experienced between replicate day and night experi-
ments were not sufficient to cause detectable changes
in rates of excretion.
A potential confounding factor in our experiment
was that the zooplankters captured on the oral arms of
the medusae were not removed prior to the experi-
ments and so some of the nutrients released into the
tubs may have been derived from decomposition of
the medusae’s prey rather than excretion. We made a
conscious decision not to remove the prey as it would
have required either confining the jellyfish for periods
of several hours in filtered water (to allow the prey to
be digested) or vigorously washing the oral arms to
remove the prey. Either of these procedures would
have increased the stress on the animals and, there-
fore, had the potential to influence their metabolic
rates. The consequences of retaining the zooplankton
were that it would have artificially elevated the
concentrations of nutrients in the treatments that
contained jellyfish, relative to the controls. Given that
both species of jellyfish prey on zooplankton, albeit in
different relative abundances (Pitt, unpublished data),
the retention of the zooplankton probably had minimal
influence on comparisons between species. Overall
the biomass of zooplankton on the oral arms of the
animals would have been small and we consider that it
would have had minimal influence on the results.
5. Ecological implications
Gelatinous zooplankton are voracious predators
and their intense predation on zooplankton can
indirectly enhance abundances of phytoplankton
because it reduces the grazing pressure of herbivo-
rous zooplankton on phytoplankton—so-called btop-downQ control (e.g., Olsson et al., 1992). Excretion
of inorganic nutrients by gelatinous zooplankters
may, however, simultaneously promote growth of
phytoplankton in areas where nutrients are limiting
(Biggs, 1977). Gelatinous zooplankters may, there-
fore, influence phytoplankton dynamics by a combi-
nation of btop-downQ and bbottom-upQ controls (e.g.,Deason and Smayda, 1982). C. mosaicus and P.
punctata capture similar zooplankton taxa on their
oral arms (Pitt, unpublished data) and so, like most
medusae, both of these species probably also exert
top-down control on zooplankton and phytoplankton
populations. Excretion of nitrogenous wastes by C.
mosaicus may also promote growth of phytoplank-
ton in regions where nitrogen is limiting, simulta-
neously enabling bottom-up control. In contrast, P.
punctata excreted little NH3 and PO4 and, although
it excreted more NOx than C. mosaicus, the actual
quantities excreted were small. Hence, unlike C.
mosaicus, P. punctata probably exerts minimal
bottom-up influence.
Nutrient cycling in estuaries is a dynamic process
that varies temporally and spatially. Variability arises
from, among other things, meteorological forcing
(Chapelle et al., 2000) and the nature of the sediments
and type of plants present (Eyre and Ferguson, 2002).
Such spatio-temporal variability makes it difficult to
quantify the relative contributions of different com-
ponents to nutrient recycling. Efflux from sediments is
generally considered a major source of recycled
inorganic nitrogen, but if nitrogen is recycled to the
water column in gaseous form through denitrification
(i.e., as N2), it is largely lost from the system and is
bio-available only to nitrogen-fixing organisms such

K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–8684
as cyanobacteria (Eyre and Ferguson, 2002). The
nitrogen excreted by jellyfish (predominantly NH3–N)
is in a form that is readily utilised by primary
producers and, therefore, when jellyfish are abundant,
they are likely to be an important source of bio-
available forms of nitrogen for primary producers.
The influence of medusae on nutrient recycling will
be proportional to the size of their populations. Large
and rapid fluctuations in abundances are characteristic
of jellyfish populations. For example, in 1998 in Lake
Illawarra, a coastal lagoon 80 km south of Sydney that
has a surface area of 35 km2, there was a 30-fold
increase in the abundance of C. mosaicus over a period
of just 6 weeks (Pitt and Kingsford, 2000), with the
biomass of the population bloom conservatively
estimated at 18,000 tonnes (Pitt and Kingsford,
2003). If we assume that rates of excretion of NH3
measured in Smiths Lake were similar to those of
medusae in Lake Illawarra at the time of the bloom,
then our measurements of inorganic nutrient cycling
by C. mosaicus allow us to estimate the importance of
these organisms for nutrient regeneration in Lake
Illawarra. The measured ammonia regeneration rates
of 1505 Ag NH3 kg�1 h�1 (light) and 1016 Ag NH3
kg�1 h�1 (dark) give a daily nutrient regeneration rate
of 30 mg NH3 kg�1 day�1 assuming equal day/night
length of 12 h. At times of peak jellyfish biomass
measured as around 18,000 tonnes, this gives an
excretion rate of 540 kg day�1 for the estuary as a
whole or a rate of 15 mg NH3 m�2 day�1, equivalent to
13 mg NH3–N m�2 day�1. Detailed studies by S.
Wellman (University of Newcastle, Australia; personal
communication) of primary production in the estuary
suggest that a typical primary production rate for the
estuary is about 1000 mg C m�2 day�1. Using the
Redfield ratio of 6.6 for C/N of phytoplankton, this
suggests that phytoplankton produces 152 mg N m�2
day�1. Comparison with the measured ammonia–N
excretion rate of C. mosaicus shows that NH3–N
excretion can account for about 8% of the inorganic
nitrogen requirements of phytoplankton consumers in
this system.
Further studies by the NSWDepartment of Environ-
ment and Conservation (Potts, personal communica-
tion) have shown that sediment regeneration of NH3–N
averages 4 mmol NH3–N m�2 day�1 during the day
and 13 mmol NH3–N m�2 day�1 at night giving an
average daily flux rate of 9 mmol NH3–N m�2 day�1.
Again, comparing these measured rates with the
ammonia regeneration rates of C. mosaicus shows that
ammonia regeneration by the jellyfish amounts to
about 11% of the measured ammonia regeneration
rates from sediments, generally believed to be the
primary source of inorganic nutrients to this system.
The figures calculated above are likely to be
conservative because the average size of medusae
during the population bloom upon which the figures
were calculated was very small (77 mm BD; Pitt and
Kingsford, 2003). Given that rates of excretion are
usually negatively correlated to size (Matsakis, 1992)
and that excretion rates were determined for much
larger animals (mean 209 mm BD), excretion by the
medusae during the bloom was probably greater than
estimated for the large adult medusae used in the
current study. Measurements of how rates of excretion
vary with size are required.
The degree of flushing of an estuary will also affect
the relative contribution of excretion by jellyfish to
nutrient recycling. Diverse types of estuaries occur
along the east coast of Australia, ranging from marine-
dominated, open embayments that are well flushed, to
shallow coastal lagoons that have minimal riverine
input and only open to the sea after periods of heavy
rain cause the entrances of the lagoons to breach (Roy
et al., 2002). C. mosaicus and P. punctata occur in
many different types of estuaries, but their influence
on nutrient recycling will vary, depending on the
degree of flushing of the estuary. Excretion of
nitrogenous compounds by C. mosaicus, in particular,
will have a far greater effect in lagoons that are
isolated from the sea and poorly flushed.
There is growing evidence that populations of
gelatinous zooplankton are increasing globally (Mills,
2001), either due to the introduction of exotic species
into new regions (Shiganova, 1998) or due to
increases in the biomass of populations in their native
environments (Brodeur et al., 2002). In Australia,
populations of C. mosaicus and P. punctata some-
times form blooms (Pitt and Kingsford, 2003),
although due to the lack of long-term data, it is not
possible to determine whether this represents a generic
increase in biomass. If, indeed, populations of
medusae are increasing, then, in addition to their
intense grazing, azooxanthellate species, or those with
relatively few symbionts, are also likely to have a
major influence on nutrient recycling.

K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–86 85
Acknowledgments
We are very grateful to J. Grayson, J. Browne, B.
Clynick and P. Barnes for their tireless efforts in the
field. Thanks also to G. Coade for preparing nutrients
and useful discussion, S. Wellman for analysing the
nutrient samples, A. Redden for use of her filtering
equipment and T. Glasby for helpful discussion
regarding experimental design. We also wish to thank
The University of Sydney for use of their facilities.
The project was conducted at the University of New
South Wales’ Smiths Lake Field Station and was
funded by an Australian Research Council Strategic
Partnership-Industry Training and Research grant
awarded to M. Kingsford, K. Pitt, K. Koop and D.
Rissik. [SS]
References
Balderston, W.L., Claus, G., 1969. A study of the symbiotic
relationship between Symbiodinium microadriaticum Freuden-
thal, a zooxanthella, and the upside down jellyfish Cassiopea
spec. Nova Hedwig. 17, 373–382.
Biggs, D., 1977. Respiration and ammonium excretion by open
ocean gelatinous zooplankton. Limnol. Oceanogr. 22, 108–117.
Brodeur, R., Sugisaki, H., Hunt, G.J., 2002. Increases in jellyfish
biomass in the Bering Sea: implications for the ecosystem. Mar.
Ecol., Prog. Ser. 233, 89–103.
Cates, N., McLaughlin, J.A.J., 1976. Differences of ammonia
metabolism in symbiotic and aposymbiotic Condylactus and
Cassiopea spp. J. Exp. Mar. Biol. Ecol. 21, 1–5.
Chapelle, A., Menesguen, A., Deslous-Paoli, J.-M., Souchu, P.,
Mazouni, N., Vaquer, A., Millet, B., 2000. Modelling nitrogen,
primary production and oxygen in a Mediterranean lagoon.
Impact of oyster farming and inputs from the watershed. Ecol.
Model. 127, 161–181.
Deason, E., Smayda, T., 1982. Ctenophore–zooplankton–phyto-
plankton interactions in Narragansett Bay, Rhode Island, USA,
during 1972–1977. J. Plankton Res. 4, 203–217.
Eyre, B.D., Ferguson, A.J.P., 2002. Comparison of carbon
production and decomosition, benthic nutrient fluxes and
denitrification in seagrass, phytoplankton, benthic microalgae-
and macroalgae-dominated warm–temperate Australian lagoons.
Mar. Ecol., Prog. Ser. 229, 43–59.
Ikeda, T., 1985. Metabolic rates of epipelagic marine zooplankton as
a function of body mass and temperature. Mar. Biol. 85, 1–11.
Ikeda, T., Mitchell, A.W., 1982. Oxygen uptake, ammonia excretion
and phosphate excretion by krill and other Antarctic zooplank-
ton in relation to their body size and chemical composition. Mar.
Biol. 71, 283–298.
Kremer, P., 1977. Respiration and excretion by the ctenophore
Mnemiopsis leidyi. Mar. Biol. 44, 43–50.
Kremer, P., 1982. Effect of food availability on the metabolism of
the ctenophore Mnemiopsis mccradyi. Mar. Biol. 71, 149–156.
Matsakis, S., 1992. Ammonia excretion rate of Clytia spp.
hydromedusae (Cnidaria Thecata): effects of individual dry
weight, temperature and food availability. Mar. Ecol., Prog. Ser.
84, 55–63.
Matsakis, S., Conover, R.J., 1991. Abundance and feeding of
medusae and their potential impact as predators on other
zooplankton in Bedford Basin (Nova Scotia, Canada) during
spring. Can. J. Fish. Aquat. Sci. 48, 1419–1430.
McCloskey, L.R., Muscatine, L., Wilkerson, F.P., 1994. Daily
photosynthesis, respiration, and carbon budgets in a tropical
marine jellyfish (Mastigias sp.). Mar. Biol. 119, 13–22.
Miller, D., Yellowlees, D., 1989. Inorganic nitrogen uptake by
symbiotic marine cnidarians: a critical review. Proc. R. Soc.
London, Ser. B 237, 109–125.
Mills, C.E., 2001. Jellyfish blooms: are populations increasing
globally in response to changing ocean conditions? Hydro-
biologia 451, 55–68.
Muscatine, L., Marian, R.E., 1982. Dissolved inorganic nitrogen
flux in symbiotic and nonsymbiotic medusae. Limnol. Oceanogr.
27, 910–917.
Olsson, P., Graneli, E., Carlsson, P., Abreu, P., 1992. Structuring of
a postspring phytoplankton community by manipulation of
trophic interactions. J. Exp. Mar. Biol. Ecol. 158, 249–266.
Pages, F., White, M.G., Rodhouse, P.G., 1996. Abundance of
gelatinous carnivores in the nekton community of the Antarctic
Polar Frontal Zone in summer 1994. Mar. Ecol., Prog. Ser. 141,
139–147.
Pages, F., Gonzalez, H.E., Ramon, M., Sobarzo, M., Gili, J.-M.,
2001. Gelatinous zooplankton assemblages associated with
water masses in the Humboldt Current System, and potential
predatory impact by Bassia bassensis (Siphonophora: Calyco-
phorae). Mar. Ecol., Prog. Ser. 210, 13–24.
Pitt, K.A., Kingsford, M.J., 2000. Geographic separation of stocks of
the edible jellyfish Catostylus mosaicus (Rhizostomeae) in New
South Wales, Australia. Mar. Ecol., Prog. Ser. 136, 143–155.
Pitt, K.A., Kingsford, M.J., 2003. Temporal variation in the virgin
biomass of the edible jellyfish, Catostylus mosaicus (Scypho-
zoa, Rhizostomeae). Fish. Res. 63, 303–313.
Priddle, J., Whitehouse, M.J., Ward, P., Shreeve, R.S., Brierley,
A.S., Atkinson, A., Watkins, J.L., Brandon, M.A., Cripps, G.C.,
2003. Biogeochemistry of a Southern Ocean plankton ecosys-
tem: using natural variability in community composition to
study the role of metazooplankton in carbon and nitrogen
cycles. J. Geophys. Res. 108 (C4), 8082.
Reissig, M., Queimalinos, P., Balseiro, E.G., 2003. Effects of
Galaxias maculatus on nutrient dynamics and phytoplankton
biomass in a North Patagonian oligotrophic lake. Environ. Biol.
Fish. 68, 15–24.
Roy, P.S., Williams, R.J., Jones, A.R., Yassini, I., Gibbs, P.J.,
Coates, B., West, R.J., Scanes, P.R., Hudson, J.P., Nichol, S.,
2002. Structure and function of south-east Australian estuaries.
Estuar. Coast. Shelf Sci. 53, 351–384.
Schneider, G., 1989. The common jellyfish Aurelia aurita: standing
stock, excretion and nutrient regeneration in the Kiel Bight,
Western Baltic. Mar. Biol. 100, 507–514.

K.A. Pitt et al. / J. Exp. Mar. Biol. Ecol. 315 (2005) 71–8686
Shiganova, T.A., 1998. Invasion of the Black Sea by the ctenophore
Mnemiopsis leidyi and recent changes in pelagic community
structure. Fish. Oceanogr. 7, 305–310.
Underwood, A.J., 1997. Experiments in Ecology: Their Logical
Design and Interpretation Using Analysis of Variance. Cam-
bridge University Press, Cambridge.
Uthicke, S., 2001. Nutrient regeneration by abundant coral reef
holothurians. J. Exp. Mar. Biol. Ecol. 265, 153–170.
Verde, E.A., McCloskey, L.R., 1998. Production, respiration, and
photophysiology of the mangrove jellyfish Cassiopea xama-
chana symbiotic with zooxanthellae: effect of jellyfish size and
season. Mar. Ecol., Prog. Ser. 168, 147–162.
Wilkerson, F.P., Kremer, P., 1992. DIN, DON and PO4 flux by a
medusa with algal symbionts. Mar. Ecol., Prog. Ser. 90,
237–250.






















